Embed Size (px)
Citation preview

J Comp Physiol A (2009) 195:225–240
DOI 10.1007/s00359-008-0400-9ORIGINAL PAPER
The function of wall-following behaviors in the Mexican blind caveWsh and a sighted relative, the Mexican tetra (Astyanax)
Saurabh Sharma · Sheryl Coombs · Paul Patton · Theresa Burt de Perera
Received: 28 August 2008 / Revised: 21 November 2008 / Accepted: 23 November 2008 / Published online: 18 December 2008© Springer-Verlag 2008
Abstract Mexican blind caveWsh exhibit an uncondi-tioned wall-following behavior in response to novel envi-ronments. Similar behaviors have been observed in a widevariety of animals, but the biological signiWcance and evo-lutionary history of this behavior are largely unknown. Inthis study, the behaviors of Mexican blind caveWsh (Astya-nax sp.) and sighted Mexican tetra (Astyanax mexicanus)were videotaped after Wsh were introduced into a novelenvironment under dark (infrared) or well-lit conditions.Under dark conditions, both sighted and blind morphsexhibited wall-following behaviors with subtle but signiW-cant diVerences. Blind morphs swam more nearly parallelto the wall, exhibited greater wall-following continuity andreached higher levels of sustained swimming speeds morequickly than sighted morphs. In contrast, sighted morphs inthe light remained motionless near the wall for long periodsof time or moved slowly around the center of the tank with-out entraining to the walls. These results are consistent withthe idea that wall-following is a shared, primitive trait thatserves an exploratory function under dark conditions tocompensate for the absence of vision. This behavior hasbecome more honed in blind morphs for exploratory pur-poses—in large part due to the enhanced, active-Xow sens-ing abilities of the lateral line.
Keywords Spatial cognition · Exploration · Lateral line · Hydrodynamic · Wall-following
Introduction
Wall-following behaviors represent an unconditioned pref-erence for following and staying near or maintaining contact(thigmotaxis) with vertically-oriented walls or boundaries ofan environment. They have been observed under laboratoryconditions for a wide range of invertebrate and vertebratespecies, including cockroaches (Creed and Miller 1990;Jeanson et al. 2003; Besson and Martin 2004; Martin 2004),crayWsh (Basel and Sandemann 2000), rodents (Barnett1963; Treit and Fundytus 1988; Simon et al. 1994), blindhumans (Kallai et al. 2004, 2007) and blind caveWsh (Brederand Gresser 1941; Gertychowa 1970; Teyke 1985, 1989;Abdel-Latif et al. 1990; Burt de Perera 2008).
Wall-following behaviors are often elicited when ani-mals encounter novel environments (Weissert and vonCampenhausen 1981; Teyke 1985, 1989; Wolfer et al.1998; Basel and Sandemann 2000). As part of an overallresponse to novel and unknown circumstances, wall-fol-lowing behaviors could be stress-induced and thereby,serve a protective purpose for Wnding shelter and/or escaperoutes. Consistent with a protective function are experimen-tal Wndings indicating that wall-following behaviors arelinked to anxiety in rodents (Treit and Fundytus 1988;Simon et al. 1994) and humans (Kallai et al. 2007). In thesestudies, anti-anxiety drugs reduced the proclivity of animalsto stay near the walls and increased the amount of timespent in the center of the test arena. For this reason, wall-seeking behaviors in these studies have been described as‘centrophobic’, which implies a ‘protection-seeking’ or‘anxiety-relieving’ function.
S. Sharma · S. Coombs (&) · P. PattonDepartment of Biological Sciences and JP Scott Center for Neuroscience, Mind and Behavior, Bowling Green State University, Bowling Green, OH 43403, USAe-mail: [email protected]
T. B. de PereraAnimal Behavior Research Group, Department of Zoology, Oxford University, Oxford, UK
123

226 J Comp Physiol A (2009) 195:225–240
Wall-following behaviors might also function as anoverall exploratory strategy for discovering and locatingpotential resources in a new environment and in a moregeneral sense, for building spatial knowledge about the rela-tive locations of various resources. For example, it hasbeen suggested by Kallai et al. (2007) that blind humansuse wall-following behaviors to build spatial knowledge ofthe size and shape of the perimeter, which can then be usedas an external frame of reference or “home base” for map-ping out the spatial relationships of objects in the remaininginterior space. Indeed, Hill et al. (1993) report that blindhumans link objects in the interior space to the perimeter ofthe environment as a successful strategy for Wnding andnavigating around objects in a novel environment. Simi-larly, wall-following behaviors in non-human animals maybe part of a general class of behaviors involving theirability to orient to (and navigate around) stationary featuresof the terrain (e.g., trees, rocks, hills) that have fairly stablelocations over time.
Spatial orientation of this type may additionally involvelearning and remembering the relative locations or geomet-ric conWguration of several landmark features, as mightoccur for animals that maintain restricted territories andhome ranges. Whereas initial encounters of novel objects(e.g., landmark features) in the environment are likely toinvolve an egocentric (internal) frame of reference (i.e.,where is the landmark feature in relation to me?), spatialrelationships between features could be learned andmapped over time with respect to an allocentric (external),map-like frame of reference (i.e., where is the feature inrelation to a constellation of landmark features?).
In this respect, it is interesting to note that wall-follow-ing behaviors are often, but not always associated with spe-cies that are active under visually-deprived conditions and/or circumstances that limit or prevent the use of vision. Inthe absence of vision and other long-range senses (e.g.,echolocation), animals cannot sense distant landmark fea-tures from a single vantage point in order to learn their spa-tial relationships, but rather must move about in theenvironment to bring their short-range senses within closerange of each landmark feature. Examples include theactive head and whisker (macrovibrissae) movements ofnocturnal rodents (Fanselow and Nicolelis 1999; Mitchin-son et al. 2007) and rostral antennae manipulations of cray-Wsh (Patullo and Macmillan 2006) and cockroaches (Creedand Miller 1990) to sense and explore wall surfaces. Simi-larly, it has been proposed that blind caveWsh use their‘touch-at-a-distance’ lateral line system for sensing station-ary objects (Dijkgraaf 1963). When swimming through thewater, Wsh generate a hydrodynamic Xow Weld thatbecomes distorted by nearby objects, including planar sur-faces like walls (Hassan 1989, 1992a, b). The Xow-Welddistortions near the body surface can be detected by the
lateral line. Indeed, experiments with blind caveWsh haverevealed that despite the absence of vision, they can avoidstationary obstacles (John 1957; Gertychowa 1970; Teyke1985), discriminate between stationary objects with diVer-ent spatial characteristics (Weissert and von Campenhausen1981), and detect novel changes in familiar landmark con-Wgurations (Burt de Perera 2004b).
For these short-range senses, knowledge of the spatialconWguration of the environment can only be achievedthrough sequential samples of the environmental space andby temporally linking information from one sample at onemoment in time to that of another sample at the nextmoment in time. In this context, it is easy to imagine howorienting behaviors might be preferentially guided andentrained by continuous (e.g., walls) or closely-spaced fea-tures vs. those that are discontinuous and widely spaced.Indeed, Biro et al. (2004) show that pigeons often followlinear landmarks such as roads or railway tracks whenreturning to a familiar release site, a strategy that mayreduce the number of visual “waypoints” to be memorizedand thus, the overall ‘cognitive load’.
Despite the wide variety of animals for which wall-fol-lowing behaviors have been observed and reported, therehas been surprisingly little in the way of systematic andquantitative descriptions of the behavior in any species. Forexample, direct measurements of, e.g., sequential distancesfrom and orientations to the wall, direction of travel alongthe wall and the degree to which the wall is continuously‘followed’ are largely missing. Thus, operational criteriafor deWning the behavior have seldom been adopted orapplied. Furthermore, the true function or functions of thisbehavior are unknown for any species. Although it has beenhypothesized that wall-following behaviors could serve aprotective or exploratory function for Wnding resourcesand/or building spatial knowledge, neither of these hypo-theses have been directly tested. Likewise, it is oftenassumed that wall-following behaviors Wgure prominentlyin the lives of animals without vision or restricted use ofvision, but the prevalence of this behavior in sighted animals isunknown and the evolution of wall-following behaviors inresponse to the loss of vision has not been studied.
In this study, we document the behavioral responses ofthe cave-dwelling, eyeless Mexican blind caveWsh (Astya-nax sp.) and the eyed, stream-dwelling Mexican tetra(A. mexicanus) to a novel environment in the presence andabsence of visible light. A number of studies have shownthat Mexican blind caveWsh respond to novel environmentsby increasing their swimming speeds (Teyke 1985; Burt dePerera 2004a) and exhibiting a tendency to preferentiallyswim close to the outer boundary of the environment(Gertychowa 1970; Teyke 1985; Abdel-Latif et al. 1990).Both the wall-following preference (measured as propor-tion of time spent near the wall vs. in the center of the tank)
123

J Comp Physiol A (2009) 195:225–240 227
and swimming speed were reported to decline graduallyover approximately the same time period of up to 15 h(Abdel-Latif et al. 1990), presumably as Wsh become morefamiliar with the environment. Missing from this picture,however, is a detailed analysis of the initial, short-termdevelopment of wall-following behaviors in both blind andsighted morphs. Although it has been reported that sightedmorphs, when experimentally deprived of vision, behavesimilarly to naturally-blind cave morphs (Breder 1943;John 1957; Hassan et al. 1992), quantitative details on thedegree of similarity (or dissimilarity) are lacking.
In addition to providing a more detailed and systematicdescription of wall-following behaviors in both eyed andeyeless morphs, we examine the question of whether thewall-following behavior in blind morphs is a widely-sharedprimitive trait or newly-evolved specialized trait and if aprimitive trait, the extent to which the behavior may havebecome specialized for spatial exploration in the dark.Finally, we test the exploratory hypothesis of wall-follow-ing function—i.e., that wall-following behavior is a senso-rimotor strategy for acquiring spatial knowledge whenlong-range sensory cues are unavailable. We reason that ifthe exploratory hypothesis is true, then wall-followingbehaviors should be exhibited by both blind and sightedmorphs deprived of long-range visual cues, but not bysighted morphs with access to visual information.
Materials and methods
Overview of experimental design
The behaviors of blind and sighted (Astyanax) morphs werevideotaped during and after their initial introduction into anovel arena under dark (infrared) and visible light condi-tions. To eliminate chemosensory ‘novelty’ cues and tominimize stress factors associated with diVerences betweenwater quality in home and experimental tanks (e.g., watertemperature, conductivity, pH etc.), the novel test arenawas Wlled with water from the Wsh’s home tank. Possibledisturbances from substrate borne vibrations (e.g., thosearising from some one walking across the room) were mini-mized with a vibration–isolation table. Fish movementswere subsequently tracked with an automatic image-track-ing system to provide a post-hoc analysis of how the Wsh’sswimming speed and position (distance and orientation)with respect to the arena walls varied over time.
Experimental animals
Naïve blind caveWsh (Astyanax sp.) (3–6 cm in standardbody length, SL) and sighted Mexican tetra (A. mexicanus)(SL, 4–6 cm) were used for all experiments. Blind morphs
were obtained from commercial aquarium suppliers,whereas sighted morphs were provided by Dr. TimothyBonner at Texas State University, San Marcos, Texas. Fishwere maintained in 75.8 l aquaria at 20–25°C and 12:12light/dark cycles. Protocols for the maintenance, care andexperimental use of Wsh followed the guidelines for theCare and Use of Laboratory Animals and have beenapproved by Bowling Green State University InstitutionalAnimal Care and Use Committee.
Experimental setup
Fish were tested in a circular arena (30 cm or »6 SL indiameter) housed inside a larger rectangular tank resting ontop of a vibration–isolation table. A video camera (SonyHandicam DCR-HC 42) mounted »1 m above the testingarena recorded the swimming behavior of Wsh. The entireset-up was enclosed in a light-tight enclosure made withthick black curtains to prevent any visible light from enter-ing the testing arena. A 115 £ 77 cm matrix of infra red(IR) LED diodes (»10 Amp, 20 V, � < 970 nm) placedunderneath the test arena provided an upwelling source ofIR illumination for the dark conditions of these experi-ments. For visible light conditions, the IR matrix wasreplaced with two 150 W (Halco PureLite) incandescentneodymium bulbs to produce an upwelling broad spectrumlight. A white plastic sheet above the light sources servedas a diVuser to evenly distribute the light across the Xoor ofthe experimental arena. To minimize depth of Weld errorsdue to vertical excursions of the Wsh, water depth in theexperimental arena was kept shallow (5 cm).
Behavioral procedures
Fish were transferred from their home tank to a transportbucket and the experimental arena using a plastic-lined,water-Wlled net to prevent any mechanical damage to super-Wcial neuromasts of the lateral line system. Fish were intro-duced into a small holding ring (»15 cm in diameter) in thecenter of the rectangular tank. The circular testing arenawas then placed around the holding ring, the dark room cur-tains were sealed shut, the outside room lights were turnedoV, and the IR illumination was turned on. Fish were given5 min to acclimate and recover from the transfer process.After 5 min of acclimation, the holding ring was slowlyremoved to minimize water motions and Wsh were releasedinto the experimental arena where their swimming behaviorwas videotaped for 10–20 min. There was a gap of approxi-mately 1 min between the removal of the holding ring andthe onset of video capture. In an additional set of experi-ments on sighted individuals, the 5 min period of acclima-tion was extended to 1 h to give individuals more time toadapt to the dark conditions before testing began and to
123

228 J Comp Physiol A (2009) 195:225–240
control for the possibility that behavioral responses weredue to an abrupt loss of vision rather than exposure to anovel environment. One hour was based on the typical time(»40 to 60 min) that it takes for the vertebrate retina torecover light sensitivity when background lights areabruptly turned oV (Dowling 1967).
Data analysis
Video Capture Software (Winnov, Version 3.2.4185) wasused for online viewing and capturing of digital images ofthe swimming behavior of the Wsh at the rate of 5 framesper second. The frame-to-frame position of the Wsh wassubsequently tracked using Image Pro Software (Version6.0 Media Cybernetics). The software has an automaticimage detection and tracking feature that Wrst Wts an ellipseto the outline of the Wsh’s main body (from the tip of thesnout to the base of the caudal peduncle) and then deter-mines the centroid of the ellipse. For each video frame, thesoftware determines (a) the position of the centroid inCartesian co-ordinates relative to a Wxed reference point onthe video screen and (b) the major and minor axis of theellipse. Custom MATLAB subroutines subsequently com-pute the ellipse’s orientation to and distance from the wallsof the arena, as well as distance moved from one frame tothe next in order to derive a characteristic swimming veloc-ity for each frame pair.
Distance to the wall was computed as the shortest dis-tance from the centroid of the ellipse to the wall. Orienta-tion to the wall was deWned by the direction in which thesnout-end of the ellipse was pointing with respect to thewall. The ‘snout-end’ of the ellipse was distinguished fromthe ‘tail-end’ by determining the direction of movementfrom one frame to the next and by making the assumptionthat motion was head-Wrst (i.e., Wsh were moving forwardsrather than backwards or sidewards). By convention, anglesof 0° and 180° represent cases in which the long axis of theWsh was orthogonal to the wall surface and the Wsh washeading directly towards and away from the wall, respec-tively. In contrast, 90° and 270° represent cases in whichthe Wsh’s body was parallel to the nearest wall section andthe Wsh was moving in a clockwise (90) or counterclock-wise (270) direction around the circular arena.
Potential sources of measurement errors include pixelresolution, which was measured to be <1 mm, and thegoodness of Wt between the ellipse and the Wsh’s body.Errors in estimating centroid distance and Wsh orientationdepend largely on the bending angle of the Wsh. When theWsh’s body is straight, the ellipse Wtting procedure gives areliable estimate of both body orientation and centroid posi-tion. If the Wsh is involved in a wide-angle turn and thebending angle is large, the Wt is such that the centroid of theellipse is shifted a few mm relative to the true centroid of
the Wsh. The orientation of the ellipse will also be shiftedrelative to the true orientation of the Wsh. However, mea-surement errors due to large bending angles are assumed tobe infrequent, small and randomly distributed over timeand between populations and treatments.
In order to determine if Wsh exhibited preferred wall ori-entations, a modiWed version of the vector strength, r, of acircular distribution (Batschelet 1981), which varies from 0(Wsh headings uniformly distributed across 360°) to 1 (allWsh headings the same), was computed. Rather than com-puting the vector strength from a full 360° worth of data,however, clockwise (0°–180°) and counterclockwise(180°–360°) directions of movement along the wall werecollapsed into a single 180° distribution. This modiWcationpreserves the relative orientation of the Wsh’s body withrespect to the wall while removing direction of travel diVer-ences due to clockwise versus counterclockwise move-ments around the rim.
To determine if Wsh exhibited regional preferences orevidence of non-uniform travel around the perimeter of thetank, the angular position of the Wsh with respect to the tankperimeter was measured using a compass coordinate sys-tem (i.e., with respect to an arbitrary north (0°), south(180°), east (270°) or west (90°) side of the tank) for casesin which the distance to the wall was <0.5 SL. The vectorstrength of the full 360° circular distribution of angularpositions was also computed (Batschelet 1981).
Three additional indices (wall-preference, wall-follow-ing and long-bout indices) were used to characterize thestrength and nature of wall-following behaviors. The wall-preference-index (the fraction of video frames in whichWsh-to-wall distances were less than 0.5 SL) was used todetermine the degree to which Wsh were near the wall ver-sus other areas of the tank. The wall-following index pro-vided a measure of the degree to which Wsh continuouslytraveled along the wall while maintaining a narrow range ofbody orientations and distances with respect to the wall.For this index, a single bout of continuous wall-followingbehavior was deWned as when Wsh (1) traveled a minimumdistance of 2 SLs along the wall surface while maintaining(2) a distance of <0.5 SLs away from the wall and (3) abody orientation of §30° of parallel to the wall circumfer-ence. Brief, single frame (1/5th sec) departures from crite-ria 2 and 3 were ignored to allow for transient departuresfrom the criteria and to reduce the number of occasionalerrors due to the ellipse-Wtting procedure. The total distancetraveled during consecutive wall-following bouts was thendivided by the total path length (distance traveled near oraway from the wall during a speciWed time interval) to yielda wall-following index for 1- or 10-min time-intervals.Because high wall-following indices could result frommany, closely spaced short bouts or a few long bouts, anadditional metric, the long-bout index, was created to
123
J Comp Physiol A (2009) 195:225–240 229
distinguish between these two possibilities. This index wassimply the percent contribution of long bouts (those greaterin length than 10 SL) to the total distance traveled.
The 0.5 SL distance criteria used for various wall-fol-lowing indices was selected as a conservative criterion,given diVerences in body length/width ratios between thetwo populations. This distance represented between 3 and 5times the body width of sighted individuals (mean = 4.3 §0.33) and between 2 and 4 times the body width of blindindividuals (mean = 2.9 § 0.56). Thus, a criterion based onbody width, rather than length, would have required sightedmorphs to have been much closer to the wall (in relativebody-width terms) than blind morphs and would havelikely increased, rather than decreased the statistical signiW-cance of diVerences reported in this study.
Statistical analysis
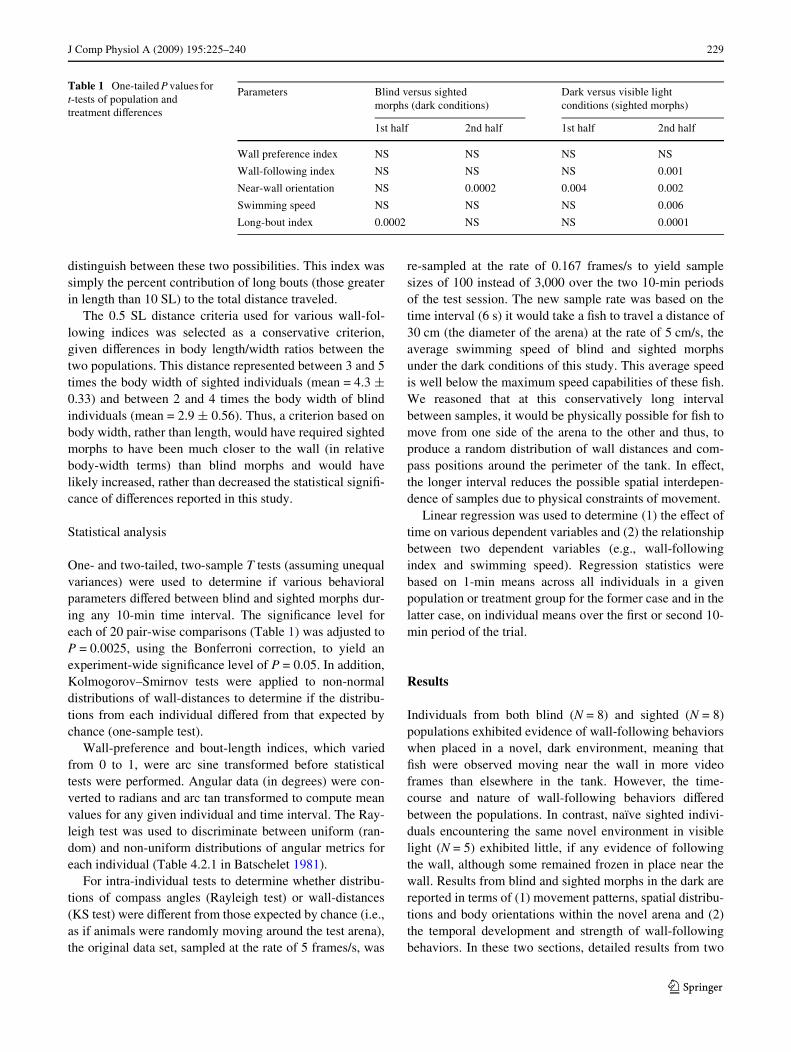
One- and two-tailed, two-sample T tests (assuming unequalvariances) were used to determine if various behavioralparameters diVered between blind and sighted morphs dur-ing any 10-min time interval. The signiWcance level foreach of 20 pair-wise comparisons (Table 1) was adjusted toP = 0.0025, using the Bonferroni correction, to yield anexperiment-wide signiWcance level of P = 0.05. In addition,Kolmogorov–Smirnov tests were applied to non-normaldistributions of wall-distances to determine if the distribu-tions from each individual diVered from that expected bychance (one-sample test).
Wall-preference and bout-length indices, which variedfrom 0 to 1, were arc sine transformed before statisticaltests were performed. Angular data (in degrees) were con-verted to radians and arc tan transformed to compute meanvalues for any given individual and time interval. The Ray-leigh test was used to discriminate between uniform (ran-dom) and non-uniform distributions of angular metrics foreach individual (Table 4.2.1 in Batschelet 1981).
For intra-individual tests to determine whether distribu-tions of compass angles (Rayleigh test) or wall-distances(KS test) were diVerent from those expected by chance (i.e.,as if animals were randomly moving around the test arena),the original data set, sampled at the rate of 5 frames/s, was
re-sampled at the rate of 0.167 frames/s to yield samplesizes of 100 instead of 3,000 over the two 10-min periodsof the test session. The new sample rate was based on thetime interval (6 s) it would take a Wsh to travel a distance of30 cm (the diameter of the arena) at the rate of 5 cm/s, theaverage swimming speed of blind and sighted morphsunder the dark conditions of this study. This average speedis well below the maximum speed capabilities of these Wsh.We reasoned that at this conservatively long intervalbetween samples, it would be physically possible for Wsh tomove from one side of the arena to the other and thus, toproduce a random distribution of wall distances and com-pass positions around the perimeter of the tank. In eVect,the longer interval reduces the possible spatial interdepen-dence of samples due to physical constraints of movement.
Linear regression was used to determine (1) the eVect oftime on various dependent variables and (2) the relationshipbetween two dependent variables (e.g., wall-followingindex and swimming speed). Regression statistics werebased on 1-min means across all individuals in a givenpopulation or treatment group for the former case and in thelatter case, on individual means over the Wrst or second 10-min period of the trial.
Results
Individuals from both blind (N = 8) and sighted (N = 8)populations exhibited evidence of wall-following behaviorswhen placed in a novel, dark environment, meaning thatWsh were observed moving near the wall in more videoframes than elsewhere in the tank. However, the time-course and nature of wall-following behaviors diVeredbetween the populations. In contrast, naïve sighted indivi-duals encountering the same novel environment in visiblelight (N = 5) exhibited little, if any evidence of followingthe wall, although some remained frozen in place near thewall. Results from blind and sighted morphs in the dark arereported in terms of (1) movement patterns, spatial distribu-tions and body orientations within the novel arena and (2)the temporal development and strength of wall-followingbehaviors. In these two sections, detailed results from two
Table 1 One-tailed P values for t-tests of population and treatment diVerences
Parameters Blind versus sighted morphs (dark conditions)
Dark versus visible light conditions (sighted morphs)
1st half 2nd half 1st half 2nd half
Wall preference index NS NS NS NS
Wall-following index NS NS NS 0.001
Near-wall orientation NS 0.0002 0.004 0.002
Swimming speed NS NS NS 0.006
Long-bout index 0.0002 NS NS 0.0001
123
230 J Comp Physiol A (2009) 195:225–240
individuals, one sighted (SF 5) and one blind (BC 30) arehighlighted as examples that bracket the range of variabilityobserved across the two populations in the dark. A thirdsection presents results from sighted morphs under visiblelight compared to sighted and blind morphs in the dark. Thelast (fourth) section examines the eVects of swimmingspeed and Wsh length on wall-following behaviors ofsighted and blind morphs in the dark.
Movement patterns, spatial distributions and body orientations of blind and sighted morphs in the dark
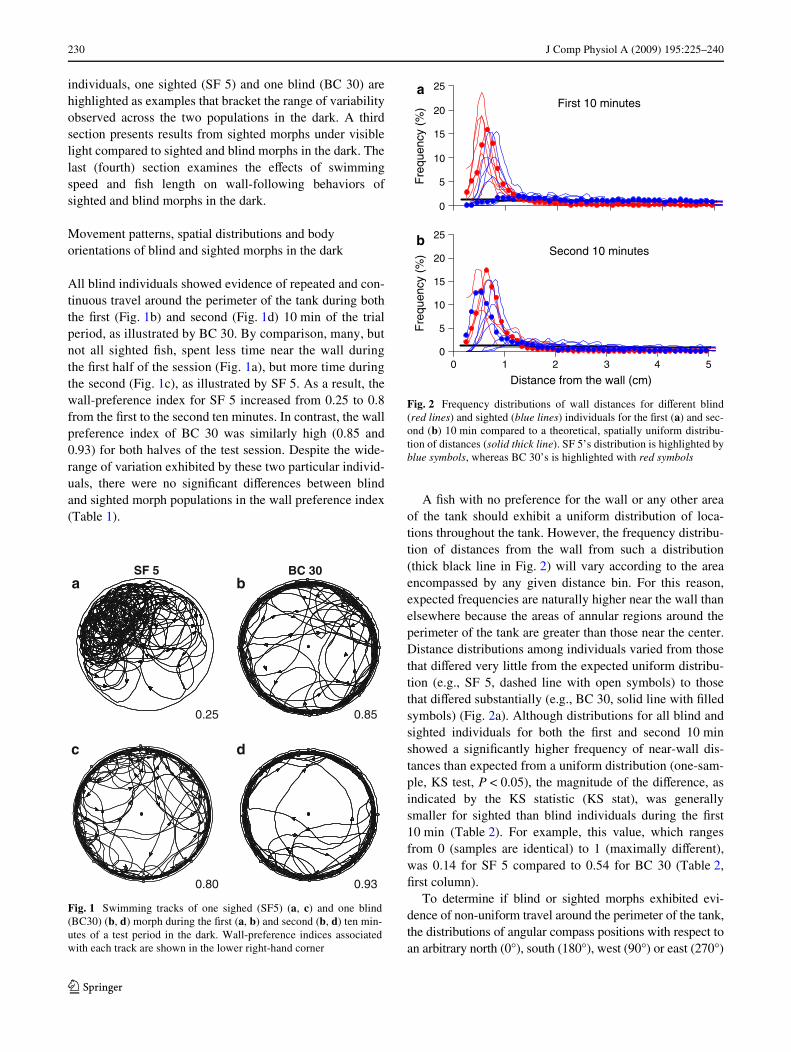
All blind individuals showed evidence of repeated and con-tinuous travel around the perimeter of the tank during boththe Wrst (Fig. 1b) and second (Fig. 1d) 10 min of the trialperiod, as illustrated by BC 30. By comparison, many, butnot all sighted Wsh, spent less time near the wall duringthe Wrst half of the session (Fig. 1a), but more time duringthe second (Fig. 1c), as illustrated by SF 5. As a result, thewall-preference index for SF 5 increased from 0.25 to 0.8from the Wrst to the second ten minutes. In contrast, the wallpreference index of BC 30 was similarly high (0.85 and0.93) for both halves of the test session. Despite the wide-range of variation exhibited by these two particular individ-uals, there were no signiWcant diVerences between blindand sighted morph populations in the wall preference index(Table 1).
A Wsh with no preference for the wall or any other areaof the tank should exhibit a uniform distribution of loca-tions throughout the tank. However, the frequency distribu-tion of distances from the wall from such a distribution(thick black line in Fig. 2) will vary according to the areaencompassed by any given distance bin. For this reason,expected frequencies are naturally higher near the wall thanelsewhere because the areas of annular regions around theperimeter of the tank are greater than those near the center.Distance distributions among individuals varied from thosethat diVered very little from the expected uniform distribu-tion (e.g., SF 5, dashed line with open symbols) to thosethat diVered substantially (e.g., BC 30, solid line with Wlledsymbols) (Fig. 2a). Although distributions for all blind andsighted individuals for both the Wrst and second 10 minshowed a signiWcantly higher frequency of near-wall dis-tances than expected from a uniform distribution (one-sam-ple, KS test, P < 0.05), the magnitude of the diVerence, asindicated by the KS statistic (KS stat), was generallysmaller for sighted than blind individuals during the Wrst10 min (Table 2). For example, this value, which rangesfrom 0 (samples are identical) to 1 (maximally diVerent),was 0.14 for SF 5 compared to 0.54 for BC 30 (Table 2,Wrst column).
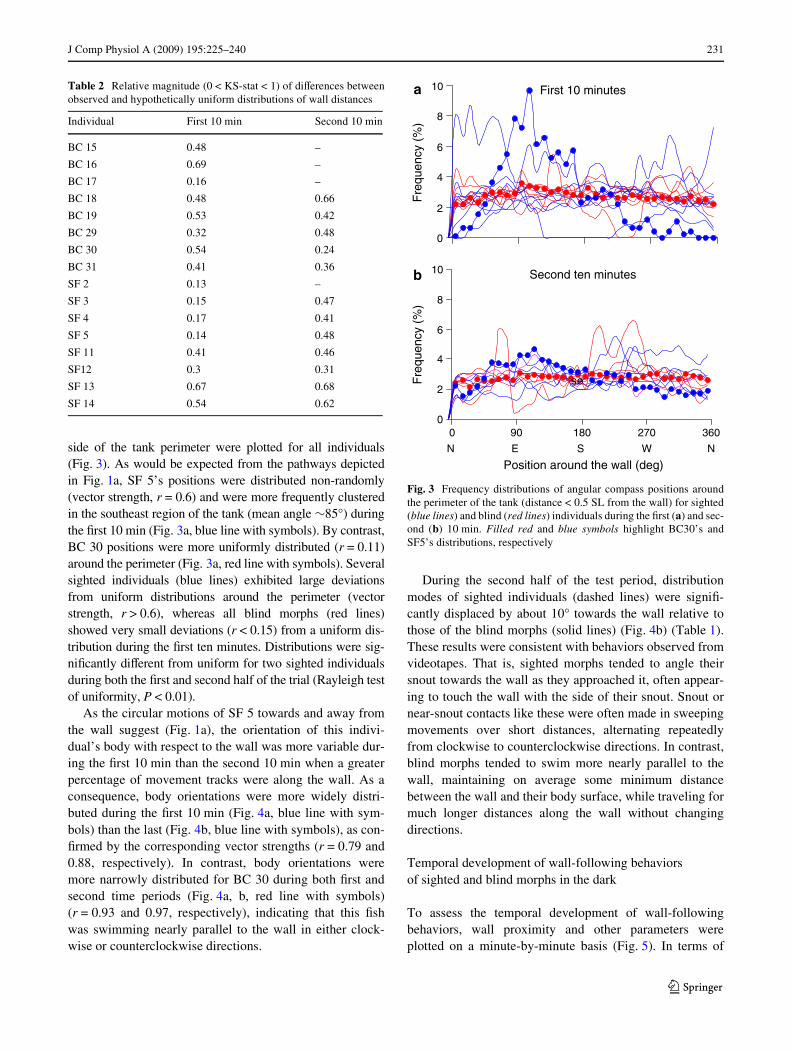
To determine if blind or sighted morphs exhibited evi-dence of non-uniform travel around the perimeter of the tank,the distributions of angular compass positions with respect toan arbitrary north (0°), south (180°), west (90°) or east (270°)
Fig. 1 Swimming tracks of one sighed (SF5) (a, c) and one blind(BC30) (b, d) morph during the Wrst (a, b) and second (b, d) ten min-utes of a test period in the dark. Wall-preference indices associatedwith each track are shown in the lower right-hand corner
a b
c d
0.25 0.85
0.80 0.93
BC 30SF 5
Fig. 2 Frequency distributions of wall distances for diVerent blind(red lines) and sighted (blue lines) individuals for the Wrst (a) and sec-ond (b) 10 min compared to a theoretical, spatially uniform distribu-tion of distances (solid thick line). SF 5’s distribution is highlighted byblue symbols, whereas BC 30’s is highlighted with red symbols
0
5
10
15
20
25
Fre
quen
cy (
%) First 10 minutes
0
5
10
15
20
25
0 1 2 3 4 5
Distance from the wall (cm)
Fre
quen
cy (
%)
b
a
Second 10 minutes
123
J Comp Physiol A (2009) 195:225–240 231
side of the tank perimeter were plotted for all individuals(Fig. 3). As would be expected from the pathways depictedin Fig. 1a, SF 5’s positions were distributed non-randomly(vector strength, r = 0.6) and were more frequently clusteredin the southeast region of the tank (mean angle »85°) duringthe Wrst 10 min (Fig. 3a, blue line with symbols). By contrast,BC 30 positions were more uniformly distributed (r = 0.11)around the perimeter (Fig. 3a, red line with symbols). Severalsighted individuals (blue lines) exhibited large deviationsfrom uniform distributions around the perimeter (vectorstrength, r > 0.6), whereas all blind morphs (red lines)showed very small deviations (r < 0.15) from a uniform dis-tribution during the Wrst ten minutes. Distributions were sig-niWcantly diVerent from uniform for two sighted individualsduring both the Wrst and second half of the trial (Rayleigh testof uniformity, P < 0.01).
As the circular motions of SF 5 towards and away fromthe wall suggest (Fig. 1a), the orientation of this indivi-dual’s body with respect to the wall was more variable dur-ing the Wrst 10 min than the second 10 min when a greaterpercentage of movement tracks were along the wall. As aconsequence, body orientations were more widely distri-buted during the Wrst 10 min (Fig. 4a, blue line with sym-bols) than the last (Fig. 4b, blue line with symbols), as con-Wrmed by the corresponding vector strengths (r = 0.79 and0.88, respectively). In contrast, body orientations weremore narrowly distributed for BC 30 during both Wrst andsecond time periods (Fig. 4a, b, red line with symbols)(r = 0.93 and 0.97, respectively), indicating that this Wshwas swimming nearly parallel to the wall in either clock-wise or counterclockwise directions.
During the second half of the test period, distributionmodes of sighted individuals (dashed lines) were signiW-cantly displaced by about 10° towards the wall relative tothose of the blind morphs (solid lines) (Fig. 4b) (Table 1).These results were consistent with behaviors observed fromvideotapes. That is, sighted morphs tended to angle theirsnout towards the wall as they approached it, often appear-ing to touch the wall with the side of their snout. Snout ornear-snout contacts like these were often made in sweepingmovements over short distances, alternating repeatedlyfrom clockwise to counterclockwise directions. In contrast,blind morphs tended to swim more nearly parallel to thewall, maintaining on average some minimum distancebetween the wall and their body surface, while traveling formuch longer distances along the wall without changingdirections.
Temporal development of wall-following behaviors of sighted and blind morphs in the dark
To assess the temporal development of wall-followingbehaviors, wall proximity and other parameters wereplotted on a minute-by-minute basis (Fig. 5). In terms of
Table 2 Relative magnitude (0 < KS-stat < 1) of diVerences betweenobserved and hypothetically uniform distributions of wall distances
Individual First 10 min Second 10 min
BC 15 0.48 –
BC 16 0.69 –
BC 17 0.16 –
BC 18 0.48 0.66
BC 19 0.53 0.42
BC 29 0.32 0.48
BC 30 0.54 0.24
BC 31 0.41 0.36
SF 2 0.13 –
SF 3 0.15 0.47
SF 4 0.17 0.41
SF 5 0.14 0.48
SF 11 0.41 0.46
SF12 0.3 0.31
SF 13 0.67 0.68
SF 14 0.54 0.62
Fig. 3 Frequency distributions of angular compass positions aroundthe perimeter of the tank (distance < 0.5 SL from the wall) for sighted(blue lines) and blind (red lines) individuals during the Wrst (a) and sec-ond (b) 10 min. Filled red and blue symbols highlight BC30’s andSF5’s distributions, respectively
First 10 minutes
0
2
4
6
8
10
Fre
quen
cy (
%)
Second ten minutes
0
2
4
6
8
10
0 90 180 270 360
Position around the wall (deg)
Fre
quen
cy (
%)
N E S N
se
a
b
W
123
232 J Comp Physiol A (2009) 195:225–240
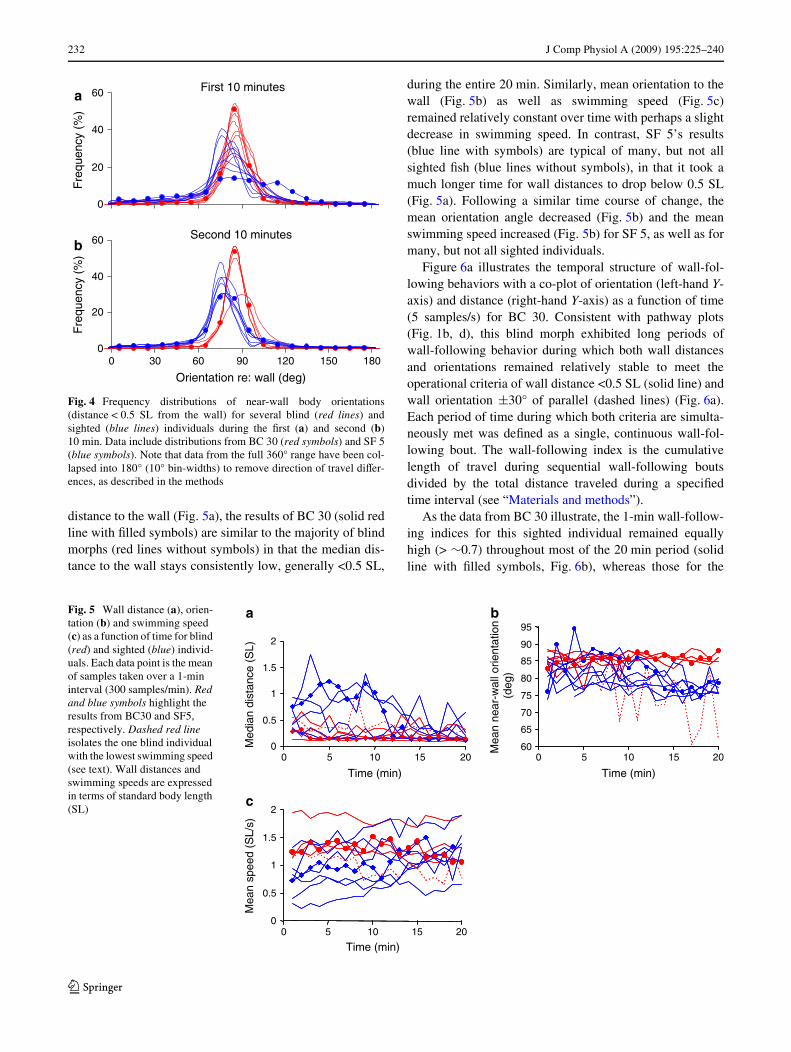
distance to the wall (Fig. 5a), the results of BC 30 (solid redline with Wlled symbols) are similar to the majority of blindmorphs (red lines without symbols) in that the median dis-tance to the wall stays consistently low, generally <0.5 SL,
during the entire 20 min. Similarly, mean orientation to thewall (Fig. 5b) as well as swimming speed (Fig. 5c)remained relatively constant over time with perhaps a slightdecrease in swimming speed. In contrast, SF 5’s results(blue line with symbols) are typical of many, but not allsighted Wsh (blue lines without symbols), in that it took amuch longer time for wall distances to drop below 0.5 SL(Fig. 5a). Following a similar time course of change, themean orientation angle decreased (Fig. 5b) and the meanswimming speed increased (Fig. 5b) for SF 5, as well as formany, but not all sighted individuals.
Figure 6a illustrates the temporal structure of wall-fol-lowing behaviors with a co-plot of orientation (left-hand Y-axis) and distance (right-hand Y-axis) as a function of time(5 samples/s) for BC 30. Consistent with pathway plots(Fig. 1b, d), this blind morph exhibited long periods ofwall-following behavior during which both wall distancesand orientations remained relatively stable to meet theoperational criteria of wall distance <0.5 SL (solid line) andwall orientation §30° of parallel (dashed lines) (Fig. 6a).Each period of time during which both criteria are simulta-neously met was deWned as a single, continuous wall-fol-lowing bout. The wall-following index is the cumulativelength of travel during sequential wall-following boutsdivided by the total distance traveled during a speciWedtime interval (see “Materials and methods”).
As the data from BC 30 illustrate, the 1-min wall-follow-ing indices for this sighted individual remained equallyhigh (> »0.7) throughout most of the 20 min period (solidline with Wlled symbols, Fig. 6b), whereas those for the
Fig. 4 Frequency distributions of near-wall body orientations(distance < 0.5 SL from the wall) for several blind (red lines) andsighted (blue lines) individuals during the Wrst (a) and second (b)10 min. Data include distributions from BC 30 (red symbols) and SF 5(blue symbols). Note that data from the full 360° range have been col-lapsed into 180° (10° bin-widths) to remove direction of travel diVer-ences, as described in the methods
0
20
40
60F
requ
ency
(%
)First 10 minutes
0
20
40
60
0 30 60 90 120 150 180
Orientation re: wall (deg)
Fre
quen
cy (
%)
a
bSecond 10 minutes
Fig. 5 Wall distance (a), orien-tation (b) and swimming speed (c) as a function of time for blind (red) and sighted (blue) individ-uals. Each data point is the mean of samples taken over a 1-min interval (300 samples/min). Red and blue symbols highlight the results from BC30 and SF5, respectively. Dashed red line isolates the one blind individual with the lowest swimming speed (see text). Wall distances and swimming speeds are expressed in terms of standard body length (SL)
0
0.5
1
1.5
2
0 5 10 15 20 0 5 10 15 20
Time (min)
Med
ian
dist
ance
(S
L)
0
0.5
1
1.5
2
0 5 10 15 20
Time (min)
Mea
n sp
eed
(SL/
s)
60
65
70
75
80
85
90
95
Time (min)
Mea
n ne
ar-w
all o
rient
atio
n (d
eg)
a
c
b
123
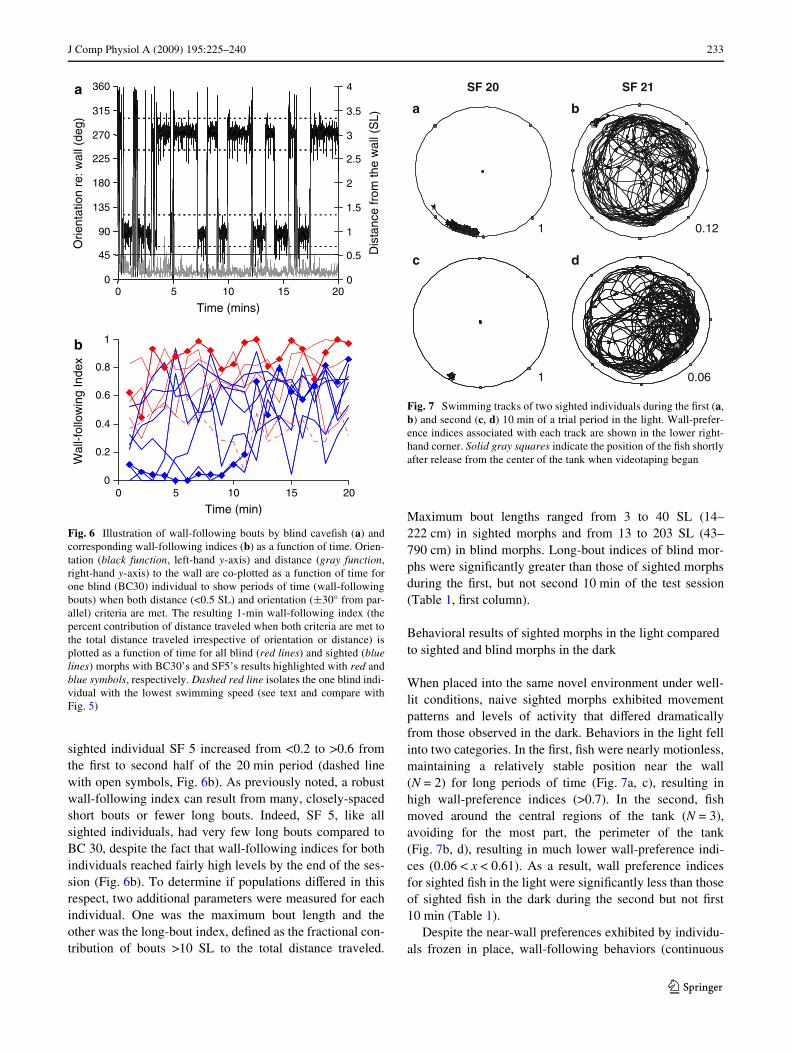
J Comp Physiol A (2009) 195:225–240 233
sighted individual SF 5 increased from <0.2 to >0.6 fromthe Wrst to second half of the 20 min period (dashed linewith open symbols, Fig. 6b). As previously noted, a robustwall-following index can result from many, closely-spacedshort bouts or fewer long bouts. Indeed, SF 5, like allsighted individuals, had very few long bouts compared toBC 30, despite the fact that wall-following indices for bothindividuals reached fairly high levels by the end of the ses-sion (Fig. 6b). To determine if populations diVered in thisrespect, two additional parameters were measured for eachindividual. One was the maximum bout length and theother was the long-bout index, deWned as the fractional con-tribution of bouts >10 SL to the total distance traveled.
Maximum bout lengths ranged from 3 to 40 SL (14–222 cm) in sighted morphs and from 13 to 203 SL (43–790 cm) in blind morphs. Long-bout indices of blind mor-phs were signiWcantly greater than those of sighted morphsduring the Wrst, but not second 10 min of the test session(Table 1, Wrst column).
Behavioral results of sighted morphs in the light compared to sighted and blind morphs in the dark
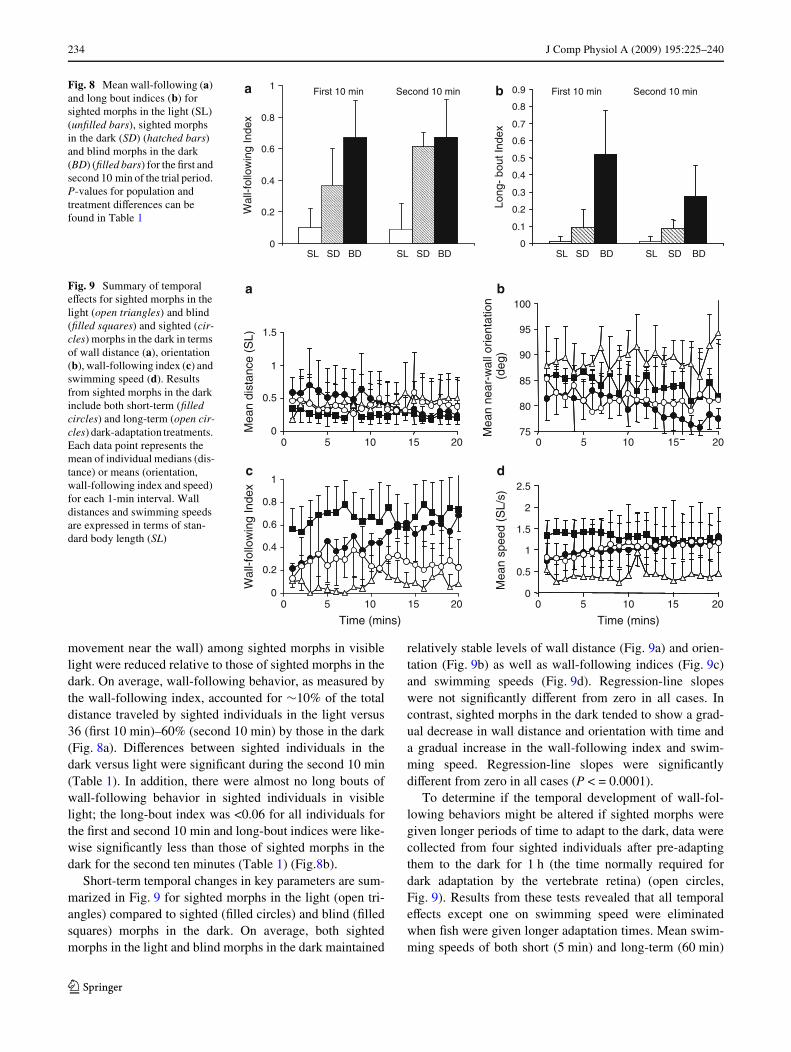
When placed into the same novel environment under well-lit conditions, naive sighted morphs exhibited movementpatterns and levels of activity that diVered dramaticallyfrom those observed in the dark. Behaviors in the light fellinto two categories. In the Wrst, Wsh were nearly motionless,maintaining a relatively stable position near the wall(N = 2) for long periods of time (Fig. 7a, c), resulting inhigh wall-preference indices (>0.7). In the second, Wshmoved around the central regions of the tank (N = 3),avoiding for the most part, the perimeter of the tank(Fig. 7b, d), resulting in much lower wall-preference indi-ces (0.06 < x < 0.61). As a result, wall preference indicesfor sighted Wsh in the light were signiWcantly less than thoseof sighted Wsh in the dark during the second but not Wrst10 min (Table 1).
Despite the near-wall preferences exhibited by individu-als frozen in place, wall-following behaviors (continuous
Fig. 6 Illustration of wall-following bouts by blind caveWsh (a) andcorresponding wall-following indices (b) as a function of time. Orien-tation (black function, left-hand y-axis) and distance (gray function,right-hand y-axis) to the wall are co-plotted as a function of time forone blind (BC30) individual to show periods of time (wall-followingbouts) when both distance (<0.5 SL) and orientation (§30° from par-allel) criteria are met. The resulting 1-min wall-following index (thepercent contribution of distance traveled when both criteria are met tothe total distance traveled irrespective of orientation or distance) isplotted as a function of time for all blind (red lines) and sighted (bluelines) morphs with BC30’s and SF5’s results highlighted with red andblue symbols, respectively. Dashed red line isolates the one blind indi-vidual with the lowest swimming speed (see text and compare withFig. 5)
a
0
0.2
0.4
0.6
0.8
1
0 5 10 15 20
0 5 10 15 20
Time (min)
b
Wal
l-fol
low
ing
Inde
x
0
45
90
135
180
225
270
315
360
Time (mins)
Orie
ntat
ion
re: w
all (
deg)
0
0.5
1
1.5
2
2.5
3
3.5
4
Dis
tanc
e fr
om th
e w
all (
SL)
Fig. 7 Swimming tracks of two sighted individuals during the Wrst (a,b) and second (c, d) 10 min of a trial period in the light. Wall-prefer-ence indices associated with each track are shown in the lower right-hand corner. Solid gray squares indicate the position of the Wsh shortlyafter release from the center of the tank when videotaping began
1
0.06
d
1
0.12
b
SF 21SF 20
a
c
123
234 J Comp Physiol A (2009) 195:225–240
movement near the wall) among sighted morphs in visiblelight were reduced relative to those of sighted morphs in thedark. On average, wall-following behavior, as measured bythe wall-following index, accounted for »10% of the totaldistance traveled by sighted individuals in the light versus36 (Wrst 10 min)–60% (second 10 min) by those in the dark(Fig. 8a). DiVerences between sighted individuals in thedark versus light were signiWcant during the second 10 min(Table 1). In addition, there were almost no long bouts ofwall-following behavior in sighted individuals in visiblelight; the long-bout index was <0.06 for all individuals forthe Wrst and second 10 min and long-bout indices were like-wise signiWcantly less than those of sighted morphs in thedark for the second ten minutes (Table 1) (Fig.8b).
Short-term temporal changes in key parameters are sum-marized in Fig. 9 for sighted morphs in the light (open tri-angles) compared to sighted (Wlled circles) and blind (Wlledsquares) morphs in the dark. On average, both sightedmorphs in the light and blind morphs in the dark maintained
relatively stable levels of wall distance (Fig. 9a) and orien-tation (Fig. 9b) as well as wall-following indices (Fig. 9c)and swimming speeds (Fig. 9d). Regression-line slopeswere not signiWcantly diVerent from zero in all cases. Incontrast, sighted morphs in the dark tended to show a grad-ual decrease in wall distance and orientation with time anda gradual increase in the wall-following index and swim-ming speed. Regression-line slopes were signiWcantlydiVerent from zero in all cases (P < = 0.0001).
To determine if the temporal development of wall-fol-lowing behaviors might be altered if sighted morphs weregiven longer periods of time to adapt to the dark, data werecollected from four sighted individuals after pre-adaptingthem to the dark for 1 h (the time normally required fordark adaptation by the vertebrate retina) (open circles,Fig. 9). Results from these tests revealed that all temporaleVects except one on swimming speed were eliminatedwhen Wsh were given longer adaptation times. Mean swim-ming speeds of both short (5 min) and long-term (60 min)
Fig. 8 Mean wall-following (a) and long bout indices (b) for sighted morphs in the light (SL) (unWlled bars), sighted morphs in the dark (SD) (hatched bars) and blind morphs in the dark (BD) (Wlled bars) for the Wrst and second 10 min of the trial period. P-values for population and treatment diVerences can be found in Table 1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Long
- bo
ut In
dex
0
0.2
0.4
0.6
0.8
1
Wal
l-fol
low
ing
Inde
xSL SD BD SL SD BD SL SD BD SL SD BD
First 10 min Second 10 minFirst 10 min Second 10 mina b
Fig. 9 Summary of temporal eVects for sighted morphs in the light (open triangles) and blind (Wlled squares) and sighted (cir-cles) morphs in the dark in terms of wall distance (a), orientation (b), wall-following index (c) and swimming speed (d). Results from sighted morphs in the dark include both short-term (Wlled circles) and long-term (open cir-cles) dark-adaptation treatments. Each data point represents the mean of individual medians (dis-tance) or means (orientation, wall-following index and speed) for each 1-min interval. Wall distances and swimming speeds are expressed in terms of stan-dard body length (SL)
75
80
85
90
95
100
0 5 10 15 20
0 5 10 15 20
Mea
n ne
ar-w
all o
rient
atio
n (d
eg)
0
0.5
1
1.5
0 5 10 15 20
0 5 10 15 20
Mea
n di
stan
ce (
SL)
0
0.5
1
1.5
2
2.5
Time (mins)
Mea
n sp
eed
(SL/
s)
0
0.2
0.4
0.6
0.8
1
Time (mins)
Wal
l-fol
low
ing
Inde
x
a b
c d
123

J Comp Physiol A (2009) 195:225–240 235
adapted groups increased at the same rate (slopes of regres-sion lines = 0.02 SL/min) and the regression-line slope wassigniWcantly diVerent from zero for the short-term adaptedgroup (P = 0.0001) and almost, but not quite signiWcantlydiVerent from zero for the long-term adapted group(P = 0.07).
The eVects of swimming speed and Wsh length
Because there was a consistent trend for sighted Wsh toincrease their swimming speed in the dark, a linear regres-sion analysis was performed to determine if swimmingspeed was correlated with key wall-following parameters.Mean swimming speed was not signiWcantly correlatedwith the wall-following index for either blind or sightedmorphs. However, the long-bout index, which was uncorre-lated with speed for sighted morphs (either short- or long-term adapted groups), was signiWcantly correlated withspeed in blind morphs (P = 0.004), accounting for 77% ofthe variance.
A Wnal regression analysis was performed to determineif wall-following abilities might have been constrained byWsh length, as might be expected if the degree of wall cur-vature prevented or constrained longer Wsh from swimmingparallel to the wall and thus, from meeting the criteria forcontinuous wall-following bouts. Neither maximum boutlengths nor long-bout indices were correlated with Wshlength in either blind or sighted populations.
Discussion
Functional signiWcance of wall-following behaviors
At least two biologically-relevant functions of wall-follow-ing behaviors—exploration and predator avoidance—areworth discussing in light of the current results. The explo-ratory hypothesis proposes that wall-following behaviorsrepresent a sensorimotor strategy for bringing short-rangesenses within range of environmental boundaries for thepurpose of (a) short-term localization of needed resources(e.g., food, shelter, escape routes) and/or (b) long-termacquisition of general knowledge about the spatial conWgu-ration of the environment and the relative location ofneeded resources within the environmental space. Experi-ments to test the proposed beneWts of wall-following func-tion (e.g., does wall-following behavior lead to enhancedabilities to Wnd spatial goals?) are needed before Wrm con-clusions about an exploratory function can be reached.However, reduced levels of locomotor activity and theabsence of wall-following behaviors in sighted morphsunder well-lit conditions are consistent with the exploratoryhypothesis. That is, when visual cues are available, locomotor
strategies to bring short-range senses within range of diVer-ent spatial features in the environment are unnecessary.Thus, it is theoretically possible for sighted individuals toacquire long-range spatial information about either thelocation of a needed resource or the general size and shapeof a well-lit, bounded environment without moving todiVerent locations, but impossible for either sighted or blindindividuals to do the same in the dark, assuming that otherlong-range cues (e.g., chemical or sonic) are unavailable aswell.
The possibility that Wsh may use other long-range sensesto acquire spatial information under these experimentalconditions is unlikely for several reasons. In general, thetemporal and spatial dispersion of chemical cues by wind orwater currents can be capricious, leading to temporal insta-bilities of spatial information (Koehl 2006). In stagnantwater, any chemical cues from the wall’s surface would dis-perse slowly and uniformly by diVusion, but in the presenceof a Wsh, wall chemicals could be dispersed more rapidlyand less uniformly by Wsh-generated water motions. Fur-thermore, the possibility of odor cue use by Wsh in thisstudy was reduced by the addition of familiar, home-tankwater into the novel arena. Long-range sonic (echolocation)cues are likewise unlikely, given that sounds have yet to berecorded from this species (Popper 1976, personal observa-tion). Similarly, it is doubtful that Wsh could make use ofany kind of long-range ‘acoustic scene analysis’ (Bregman1990), given the absence of ambient noise sources (e.g.,biotic sound producers, wind and wave action etc.) in thetest chamber.
Wall-proximity by blind or sighted morphs could alsoserve a protective function against predators. That is, Wshcould take up positions near the wall for purposes of shelterand/or to decrease the number of directions from which apredator might attack. This explanation is plausible forsighted Wsh, which were sometimes observed to take up sta-tionary positions near the wall in the light. Sighted morphshave natural predators in the wild and individuals in thisstudy were sometimes observed to exhibit possible frightresponses (rapid, erratic swimming) when Wrst placed intothe test arena. However, this explanation does not obvi-ously account for the wall-following behaviors observedfor blind morphs, which are believed to have few, if anynatural predators in the subterranean caves that theyinhabit, other than possibly conspeciWc adults preying uponthe young (Mitchell et al. 1977; Parzefall 1983; Wilkens1988).
One possibility worth considering is that the attraction towalls observed in both populations serves diVerent primaryfunctions under the two sensory conditions of these experi-ments. For sighted morphs in the light, wall proximity mayserve a protective function. When wall-proximity is cou-pled with the absence of movement, as was the case for
123

236 J Comp Physiol A (2009) 195:225–240
some of the sighted individuals in this study, this theoreti-cally increases the protective value of the behavior, sincevisual predators are strongly geared towards detectingvisual motion (Palmer 1999). For both sighted and blindmorphs in the dark, however, wall-following behaviorsmay serve an exploratory function. Depending on predatorypressures and levels of perceived threat (the ‘fear’ factor),exploration could be used to build general spatial knowl-edge of the environment over longer periods of time and/orto Wnd shelter or escape routes as quickly as possible.
There are other alternative explanations for wall-follow-ing and proximity behaviors. For example, it is possiblethat by staying motionless near the wall, sighted morphs invisible light minimized their exposure to bright light, sinceit is well-known that both sighted and blind morphs exhibitnegative phototaxis (Breder 1943; Breder and Rasquin1947; Gertychowa 1970; Langecker 1989; Yoshizawa andJeVery 2008). Arguing against this possibility is the factthat light in these experiments was delivered from anupwelling source to prevent shadows and through a diVuserto minimize spatial non-uniformities in light levels.
Another explanation for wall-following behavior is thatit is an incidental consequence of how the Wsh’s movementsare redirected every time the walls of a concave environ-ment are encountered as a barrier to forward motion (Creedand Miller 1990). A critical test of this hypothesis is howanimals respond when nearing the apex of a convex wallthat no longer impedes their forward motion. Recent stud-ies in our lab using a goggle-shaped arena with both con-cave and convex surfaces have revealed that blind morphscontinue to follow a convex wall after reaching the apex aslong as the radius of curvature is <»20 cm (S. Coombs,unpublished data). As the radius of curvature decreases,animals begin to show greater departures from the moresteeply inclined walls, but the degree of departure decreasesas swimming speed increases. This correlation is similar tothe correlation reported in this study between swimmingspeed and the persistence of wall-following behavior inblind (but not sighted) morphs, as measured by maximumbout lengths and long-bout indices.
The most parsimonious explanation of these results isthat wall-following behaviors by blind morphs cannot beexplained by a simple, reactive strategy to avoid barrierswhenever they are encountered. Rather, blind morphs seemto be actively attracted to the wall in a manner that dependson swimming speed and by inference, the amplitude of thehydrodynamic signal to the lateral line. In this light, depar-tures from steeply-curved convex surfaces are more likelydue to loss of sensory information about the wall’s presencethan the absence of an impending barrier. Although similarexperiments on sighted morphs in a goggle-shaped arenahave not been done, the absence of any correlation betweenswimming speed and measures of continuous wall-following
in this study suggests that hydrodynamic cues to the lateralline play a less signiWcant role in the wall-following behav-iors of sighted morphs compared to blind morphs.
Behavioral diVerences between blind and sighted morphs in the dark: the role of active sensing by the lateral line
Unlike sighted morphs in the light, both sighted and blindmorphs in the dark exhibited wall-following behaviors.However, their near-wall swimming behaviors diVered insubtle, but signiWcant ways, casting new light on claimsthat sighted morphs behave just like blind morphs whenvision is eliminated (Breder 1943; Breder and Rasquin1947). Blind morphs swam nearly parallel to the wall sur-face for long distances (up to 8 times the circumference ofthe arena) in the same direction, developing maximum andstable swimming speeds of »1.5 SL/s within 1–2 min afterWrst being introduced into the novel arena. In contrast,sighted morphs took much longer (»10 to 15 min) to attainsimilarly high swimming speeds and swam for shorter dis-tances along the wall with the head directed more inwardstowards the wall. Instead of covering the entire perimeter inone direction, they often switched from clockwise to coun-terclockwise directions to repeatedly cover the same, shortdistance. Alternatively, they swam in short loops, Wrsttowards and then away from a small region of the wall—apattern of behavior that was also observed away from thewall by sighted Wsh in the light.
There are several, perhaps inter-related explanations forthese diVerences. Behavioral diVerences may reXect diVer-ences in locomotor strategies (e.g., when to move, how farto move, where to move etc.) for acquiring information. Forsighted animals with access to vision, small left-right scan-ning movements of the head or eyes yield a relatively wideWeld of view at a distance and represent a relatively eVec-tive way of acquiring long-range information. This is truefor other long-range senses as well—e.g., the head scanningmovements of echolocating dolphins (Potter and Taylor2001). In light of these long-range motor strategies, the bi-directional sweeping movements of sighted individualsalong short stretches of the wall in the dark may reXect amotor strategy for long-range (visual) acquisition ofinformation. In contrast, the continuous, uni-directionalwall-following behavior of blind morphs may represent asensorimotor strategy that has evolved for optimum acqui-sition of information through short-range senses, presum-ably the lateral line, as will be discussed further in thefollowing sections.
Behavioral diVerences may also reXect diVerences in thesensorimotor abilities of the two morphs to sense and fol-low the wall. In particular, the non-visual sensing abilitiesof blind morphs are likely to be enhanced relative to sightedmorphs. For active Xow sensing by the lateral line, sensing
123

J Comp Physiol A (2009) 195:225–240 237
abilities could be enhanced in terms of both signal recep-tion (e.g., morphological and physiological adaptations forincreasing sensitivity and/or spatial resolution), but also interms of signal generation (e.g., locomotor adaptations forgenerating Xow signals).
There are several reasons to believe that active sensingby the lateral line plays a major role in the sensory guidanceof wall-following behaviors by blind morphs. First, the lat-eral line system is within detection range of the wall for themajority of near-wall distances reported in this study. Theactive sensing range of the lateral line for stationary sourceslike walls depends on the strength of the active signal (i.e.,on the swimming speed of the Wsh and the resulting ampli-tude of the self-generated Xow Weld) and the extent towhich the stationary object perturbs the active Xow Weld.For blind caveWsh, the detection range for actively-sensedstationary objects is predicted to be a small fraction of stan-dard body length (<0.2 SL), as estimated from obstacleavoidance tasks (Windsor 2008) and computational modelsof Wsh-shaped objects moving past planar surfaces (Hassan1985; Windsor 2008). The ‘damming pressure’ created by aWsh swimming towards a nearby wall could also theoreti-cally be detected by the inner ear of blind caveWsh, whichlike the majority of otophysan Wsh, have morphologicalspecializations (anterior swimbladder + Weberrian ossi-cles) for sensing pressure changes (Popper 1970).
Although the sensing range of the tactile sense wouldseem to be even shorter, blind caveWsh have been observedtouching walls with extended pectoral Wns while swimmingpast them (Windsor 2008). The pectoral Wn lengths of blindcaveWsh in this study ranged from »0.6 to 0.75 cm (»0.15SL) and were easily within reach of the wall for the major-ity of reported wall-distances. Some pectoral Wn contactwith the wall under such circumstances thus seems inevita-ble. Although a contribution of pectoral Wn touch orpressure detection by the inner ear cannot be ruled out forwall-following behaviors, recent studies on blind morphsindicate that wall-following indices are dramaticallydecreased when the lateral line system is pharmacologicallyblocked (S. Coombs, unpublished data). In addition, thecorrelations between swimming speed (and by inference,hydrodynamic signal strength) and various metrics for thestrength of wall-following behaviors are also consistentwith active sensing by the lateral line.
On theoretical grounds alone, the wall-following beha-viors of blind morphs appear to be better suited for acquiringspatial information via the lateral line than those of sightedmorphs. By swimming at relatively high, but stable speedsand maintaining nearly parallel orientations to the wall,blind morphs are likely to improve their ability to detectwall-generated distortions in their own self-generated XowWeld. A high swimming speed increases the probability thatthe signal distortions will be detected since the amplitude of
the Xow Weld is a function of both cross-sectional area andswimming speed (Hassan 1985, 1989, 1992a, b; Teyke1988). A stable swimming speed ensures that diVerent sig-nal strengths arising from, e.g., landmarks of diVerent sizesor at diVerent distances will not be confused with diVerentsignal strengths caused by variable swimming speeds.
Body orientation is also important for both signal recep-tion and generation. Because the Wsh’s body is laterallycompressed and lateral line sense organs are distributed allover the head and body (Schemmel 1973), the Weld of viewwill be much narrower in front of the head than along theXanks. This is particularly important for two reasons. Oneinvolves how distance of a stimulus source is encoded bythe lateral line (Hassan 1989; Coombs et al. 2002) and theother how information from sequentially encountered land-marks may be serially linked in time for the purpose oflearning spatial relationships. As distance between the Wshand any Xow-distorting obstacle increases, the pattern ofactivity along distributed lateral line sensors broadens andthe peak activity decreases (Hassan 1985). Distance encod-ing of far sources require longer arrays than does distanceencoding of near sources. Lateral Welds of view (i.e., Wsh’sbody oriented parallel to the wall) will thus encode a largerrange of distances than frontal views (Wsh pointing towardthe wall).
Lateral Welds of view also increase the body distanceover which diVerent spatial features might be simulta-neously sensed and registered as the Wsh swims past them.In this respect, the propensity for blind morphs to swimcontinuously along the wall (or in theory, other spatiallydistributed landmark features) for long distances in thesame direction might assist them in constructing a sequen-tial map of features encountered at diVerent points in timealong a single trajectory. Such an encoding might beregarded as a kind of one-dimensional, allocentric represen-tation of feature sequences. Indeed, there is some evidenceto suggest that blind caveWsh can learn both the spatial con-Wguration of a set of landmarks (Burt de Perera 2004b), aswell as the spatial order of landmarks within the conWgura-tion (Burt de Perera 2004a) by swimming around the con-Wguration. Whether or not sequentially acquiredinformation about spatial relationships could later be con-verted into a two or three dimensional allocentric map thatallows Wsh to take the shortest possible route to a goal (e.g.,a food source) remains to be seen.
If more lateral (=parallel orientation) Welds of view arebeneWcial to exploratory behaviors, why is it that the orien-tation of blind morphs is shifted by »5–10° away from par-allel and towards the wall? One simple explanation is thatthis is merely a consequence of Wsh being unable to main-tain a strictly parallel orientation while making frequentturns to follow the curved surface. If that were the case,however, one would expect that there would be more turns
123

238 J Comp Physiol A (2009) 195:225–240
away from the wall than towards the wall and thus, that thedistribution would be shifted in the opposite direction fromthat observed. Another possibility is that observed bodyorientations could be a compromise driven by the cost/ben-eWt ratio of lateral versus frontal views in terms of hydrody-namic signal strength, Weld of view, and spatial resolution.Lateral line sense organs on the head are more denselypacked than those on the rest of the body (Schemmel 1967,1973), so the sensitivity and spatial resolution of informa-tion in a frontal ‘view’ is likely to be greater than that of alateral view. Moreover, the strength of the hydrodynamicXow Weld in front of the Wsh for any given swimming speedwill be approximately twice that of the Xow amplitude tothe side, assuming that the Wsh behaves like a dipole (Kal-mijn 1988). Nevertheless, for a Wsh that is swimming in aforward direction, there is a stagnation point directly infront of the snout where Xow velocity plummets to zero.The stagnation point coupled with a complete absence oflateral line sensors in this region (personal observation),results in an eVective lateral line ‘blind’ spot. To summa-rize, a lateral ‘view’ (parallel orientation) is good for dis-tance encoding and sequential linking of encounteredlandmark features, whereas a frontal ‘view (orthogonal ori-entation) yields a higher signal strength and better spatialresolution, except directly in front of the snout. To avoidthe blind spot and to maximize sensing advantages thatcome with both frontal and lateral views, body orientationsto the wall would likely deviate somewhat from purely par-allel or orthogonal, depending on the sensory task.
In terms of active sensing by the lateral line, it wouldappear that the behavior of blind morphs gives themenhanced capabilities relative to sighted morphs for bothsignal generation and reception. In addition to the various(putative) beneWts gained from the blind morph’s wall-fol-lowing behaviors, as discussed above, there are likely to bemorphological advantages. For example, blind morphshave broader heads than sighted morphs (JeVrey 2001, per-sonal observation), so the amplitude of the Xow Weld infront of a blind morph will theoretically be greater than thatin front of a sighted morph swimming at the same speed.SuperWcial neuromasts on the head are also likely to bemore sensitive in blind morphs than sighted morphs owingto their larger circumference and corresponding increasedsurface area and lengths of overlying cupulae (Teyke1990).
Evolution of wall-following behaviors
Conclusions about the evolution of wall-following beha-viors of blind caveWsh are complicated by the fact that thereare many cave-dwelling populations of blind caveWsh in asmany as 30 diVerent caves, largely in northeast and centralMexico (Schemmel 1967). Indeed, the current evidence
suggests that eye loss has evolved multiple times in geo-graphically-isolated caves (Mitchell et al. 1977; Borowskyand Wilkens 2002; JeVery et al. 2003). Thus, the precisephylogenetic relationship between diVerent cave popula-tions and sighted morphs of Astyanax remains in question.Nevertheless, it is abundantly clear that all sighted andblind populations studied to date can successfully inter-breed (Sadoglu 1956, 1958; Borowsky 2008), indicatingthat all populations should be considered races of the samespecies (Sadoglu 1956) all descended from a commonsighted ancestor (JeVery et al. 2003). Given that bothsighted and blind morphs in this study exhibit wall-follow-ing behaviors in the dark, we tentatively conclude that blindmorphs have inherited a primitive form of wall-followingbehavior from the common sighted ancestor. Furthermore,the subtle, but signiWcant diVerences between wall-follow-ing behaviors of blind and sighted morphs suggest thatwall-following behaviors of blind morphs have become fur-ther honed for exploratory purposes, in large part due toenhanced lateral-line sensing capabilities. Additional stud-ies with outgroup comparisons are needed before it can bedetermined if wall-following behaviors observed in sightedAstyanax morphs under the conditions of this experimentrepresent a typical response of many, if not all sighted Wsh.
Sources of inter-individual variability and selection pressures in the evolution of wall-following behaviors
In general, behavioral responses of sighted morphs, underboth dark and light conditions, appeared to more variablethan those of blind morphs under dark conditions. Forexample, approximately half of sighted individuals in thelight moved very little, whereas the other half showed sub-stantial, if slow movements. Similarly, some sighted indi-viduals in the dark exhibited a preference for remainingnear the wall, whereas others did not. In contrast, all butone blind individual exhibited wall preferences. One obvi-ous source of wall-following variability among blind mor-phs is swimming speed, which accounted for >77% of thevariance in the long-bout index. In fact, the one blind indi-vidual that diVered from all the rest in having a wall-prefer-ence index <0.5 and low wall-following indices also hadthe lowest mean swimming speed (0.4 SL/s). This source ofinter-individual variability among blind morphs can, atleast in part, be understood in terms of the hydrodynamicsignal strength of the wall’s presence. Reduced swimmingspeeds will theoretically produce weaker signals of thewall’s presence and thus, lead to poorer wall-sensing andfollowing capabilities. The strong argument against swim-ming speed as a source of variability for sighted individualsin either light or dark conditions is the absence of any cor-relation between swimming speed and any wall-followingmetrics. Ironically, the few sighted individuals to show any
123

J Comp Physiol A (2009) 195:225–240 239
preference for being near the wall in the light were thosethat remained relatively motionless in one position for longperiods of time.
If swimming speed and by interference, ability toactively sense the wall with the lateral line system is not asource of inter-individual variability in sighted individuals,what accounts for the wide range of observed behaviors?One plausible explanation is that there is greater variabilityamong sighted individuals in terms of the shy-bold contin-uum of personality traits. It is now well-documented thatindividual diVerences in these traits can have profoundeVects on behavior in a wide variety of vertebrate groups,including Wsh (Sneddon 2003; Bell 2005; Frost et al. 2007)(Yoshida et al. 2005). Bold individuals tend to be moreactive, take more risks, learn more quickly and show higherlevels of aggression than shy individuals. Thus, in novelenvironments, it is expected that bold individuals will takemore risks in actively exploring the new territory. Further-more, risk of predation and food availability are thought toplay major, if conXicting roles in shaping the frequency ofbold versus shy traits in any given population. Given thecurrent state of our knowledge on the ecology of blind andsighted morphs, it is reasonable to assume that both foodavailability and risk of predation are lower for blind thansighted individuals in their natural habitat (Mitchell et al.1977; Parzefall 1983, 1993). As a result, these two, interac-tive selective pressures would theoretically favor a higherfrequency of bold than shy traits among blind morph popu-lations and perhaps more evenly balanced frequencies ofbold and shy traits in sighted populations.
In light of this scenario, the absence of movement andpreference for the wall exhibited by half of the sighted indi-viduals in the light might well be interpreted as risk-aver-sive, predator avoidance behaviors near the shy end of thecontinuum, whereas active movements away from the wallby the other half might be interpreted as less risk-aversive,exploratory behaviors nearer the bold end of the continuum.Similarly, the variability in the temporal onset and degreeof wall-following behaviors among sighted individuals inthe dark could be explained by variability in personalitytraits along the shy-bold continuum. In contrast, the rapiddevelopment of high swimming speeds and persistentlystrong wall-following behaviors by the majority of blindmorphs are consistent with a higher frequency of bold thanshy traits in these morphs, assuming an exploratory, ratherthan protective function for wall-following behaviors.
Acknowledgments This work was done in partial fulWllment for aMaster’s degree for SS. We thank the committee members, Drs. RobertHuber and Paul Moore for their insightful comments and helpful sug-gestions throughout. We thank Dr. Tim Bonner for providing Mexicantetra and Tristan Ula for her able assistance in the care and mainte-nance of experimental animals. TBdP is supported by a Royal SocietyDorothy Hodgkin fellowship and a L’Oreal UK Women in Science
Fellowship. Finally, SS, SC and PP would like to thank their col-leagues in a multi-university collaboration supported by the Bioin-spired Concepts program (funded by the AIR force OYce of ScientiWcResearch) and the BioSenSE program (funded by the Defense Ad-vanced Research Projects Agency). In particular, we thank the PI of theproject, Dr. Chang Liu for making the collaboration and support forthis work possible, and Drs. Friedrich Barth, Horst Bleckmann, PepeHumphrey and Doug Jones for their helpful insights and perspectives.These experiments comply with the “Principles of animal care”, publi-cation No. 86-23, revised 1985 of the National Institute of Health, andalso with the current laws of the United States.
References
Abdel-Latif H, Hassan E-S, Campenhausen C (1990) Sensory perfor-mance of blind Mexican cave Wsh after destruction of the canalneuromasts. Naturwiss 77:237–239
Barnett SA (1963) The rat: a study in behavior. Metheun, LondonBasel J, Sandemann D (2000) CrayWsh (Cherax destructor) use tactile
cues to detect and learn topographic changes in their environment.Ethol 106:247–259
Batschelet E (1981) Circular statistics in biology. Mathematics in biol-ogy. Academic Press, New York, pp 52–83
Bell AM (2005) Behavioural diVerences between individuals and twopopulations of stickelbacks (Gasterosteus aculeatus). J Evol Biol18:467–473
Besson M, Martin J-R (2004) Centrophobism/thigmotaxis, a new rolefor the mushroom bodies in Drosophila. J Neurobiol 62:386–396
Biro D, Meade J, Guilford T, Krebs JR (2004) Familiar route loyaltyimplies visual pilotage in the homing pigeon. Proc Nat Acad SciUSA 101(50):17440–17443
Borowsky R (2008) Restoring sight in blind caveWsh. Curr Biol18:R23–R24
Borowsky R, Wilkens H (2002) Mapping a cave Wsh genome: poly-genic systems and regressive evolution. J Hered 93:19–21
Breder CM (1943) Problems in the behavior and evolution of a speciesof blind cave Wsh. Trans NY Acad Sci 5:168–176
Breder CM, Gresser EB (1941) Correlation between structural eye de-fects and behavior in the Mexican blind characin. Zoologica26:123–131
Breder CM, Rasquin P (1947) Comparative studies in the light sensi-tivity of blind characins from a series of Mexican caves. Bull AmMus Nat Hist 89:324–351
Bregman AS (1990) Acoustic scene analysis: the perceptual organiza-tion of sound. MIT Press, Cambridge
Burt de Perera T (2004a) Fish can encode order in their spatial map.Proc R Soc Lond B 271:2131–2134
Burt de Perera T (2004b) Spatial parameters encoded in the spatial mapof the blind Mexican cave Wsh, Astyanax fasciatus. Anim Behav68:291–295
Burt de Perera T (2008) Strategic exploration of a novel 3D environ-ment by a Wsh. Curr Biol (in press)
Coombs S, New JG, Nelson ME (2002) Information-processing de-mands in electrosensory and mechanosensory lateral line systems.J Physiol 96:341–354
Creed RP, Miller JR (1990) Interperting animal wall-following behav-ior. Experientia 46:758–761
Dijkgraaf S (1963) The functioning and signiWcance of the lateral-lineorgans. Biol Rev 38:51–105
Dowling JE (1967) The site of visual adaptation. Science 155:273–279Fanselow EE, Nicolelis MAL (1999) Behavioral modulation of tactile
responses in the rat somatosensory system. J Neurosci 19:7603–7616
123

240 J Comp Physiol A (2009) 195:225–240
Frost AJ, Winrow-GiVen A, Ashley PJ, Sneddon LU (2007) Plasticityin animal personality traits: does prior experience alter the degreeof boldness? Proc R Soc Lond B 274:333–339
Gertychowa R (1970) Studies on the ethology and spacer orientation ofthe blind cave Wsh Anoptichthys jordani Hubbs et Innes 1936(Characidae). Folia Biol 18:9–69
Hassan E-S (1985) Mathematical analysis of the stimulus for the lateralline organ. Biol Cybern 52:23–36
Hassan E-S (1989) Hydrodynamic Imaging of the Surroundings by theLateral Line of the Blind Cave Fish Anoptichthys jordani. In:Coombs S, Görner P, Münz H (eds) The mechanosensory lateralline: neurobiology and evolution. Springer, New York, pp 217–227
Hassan E-S (1992a) Mathematical description of the stimuli to the lat-eral line system of Wsh derived from a three-dimensional XowWeld analysis. II. The case of gliding alongside or above a planesurface. Biol Cybern 66:453–461
Hassan E-S (1992b) Mathematical description of the stimuli to the lat-eral line system of Wsh derived from a three-dimensional XowWeld analysis. I. The case of moving in open water and of glidingtowards a plane surface. Biol Cybern 66:443–452
Hassan E-S, Abdel-Latif R, Biebricher R (1992) Studies on the eVectsof Ca++ and Co++ on the swimming behavior of the blind Mexi-can cave Wsh. J Comp Physiol A 171:413–419
Hill EW, Rieser JJ, Hill MM, Hill M, Halpin J, Halpin R (1993) Howpersons with visual impairments explore novel spaces: strategiesof good and poor performers. J Vis Impair Blind 87:295–301
Jeanson R, Blanco S, Fournier R, Deneubourg J-L, Fourcassie V, Ther-aulaz G (2003) A model of animal movements in a boundedspace. J Theor Biol 225:443–451
JeVery WR, Strickler AG, Yamamoto Y (2003) To see or not to see:evolution of eye degeneration in Mexican blind caveWsh. IntegComp Biol 43:531–541
John KR (1957) Observations on the behavior of blind and blindedWshes. Copeia 1957:123–132
Kallai J, Makany T, Karadi K, Jacobs WJ (2004) Spatial orientationstrategies in Morris-type virtual water task for humans. BehavBrain Res 159:187–196
Kallai J, Makany T, Nadel L, Jacobs JW, Csatho A, Karadi K, HorvathD, Kovacs-Labadi B, Jarai R (2007) Cognitive and aVective as-pects of thigmotaxis strategy in humans. Behav Neurosci 121:21–30
Kalmijn AJ (1988) Hydrodynamic and acoustic Weld detection. In:Atema J, Fay RR, Popper AN, Tavolga WN (eds) Sensory biologyof aquatic animals. Springer, New York, pp 83–130
Koehl M (2006) The Xuid mechanics of arthropod sniYng in turbulentodor plumes. Chem Senses 31:93–105
Langecker TG (1989) Studies on the light reaction of epigean and cavepopulations of Astyanax fasciatus (Characidae, Pisces). MemBiospeleol 16:169–176
Martin J-R (2004) A portrait of locomotor behavior in drosophiladetermined by a video-tracking paradigm. Behav Proc 67:207–219
Mitchell HC, Russell WH, Elliott WR (1977) Mexican Eyeless Chara-cin Wshes, Genus Astyanax: environment, distribution, and evolu-tion. Texas Tech Press, Lubbock
Mitchinson B, Martin CJ, Grant RA, Prescott TJ (2007) Feedback con-trol in active sensing: rat exploratory whisking is modulated byenvironmental contact. Proc R Soc Lond B 274:1035–1041
Palmer S (1999) Vision science: photons to phenomenology. The MITPress, Cambridge
Parzefall J (1983) Field observation in epigean and cave populations ofthe Mexican Characid Astyanax mexicanus (Pisces, Characidae).Mem Biospeleol 10:171–176
Parzefall J (1993) Behavioural ecology of cave-dwelling Wsh. In:Pitcher TJ (ed) Behavior of Teleost Fishes, 2nd edn. Chapmanand Hall, London, pp 573–608
Patullo BW, Macmillan DL (2006) Corners and bubble wrap: the struc-ture and texture of surfaces inXuence crayWsh exploratory behav-ior. J Exp Biol 209:567–575
Potter JR, Taylor E (2001) On novel reception models for Bottlenosedolphin echolocation. In: Proc of the Inst of Acoust, vol 24. Bios-onar, Loughborough
Popper AN (1970) Auditory capacities of the Mexican blind caveWsh(Astyanax jordani) and its eyed ancestor (Astyanax mexicanus).Anim Behav 18:552–562
Sadoglu P (1956) A preliminary report on the genetics of the Mexicancave characins. Copeia 1956:113–114
Sadoglu P (1958) Mendelian inheritance in the hybrids between theMexican blind cave Wshes and their overground ancestor. VerhDtsch Zool Ges Graz 1957 Zool Anz Suppl 21:432–439
Schemmel C (1967) Vergleichende Untersuchungen an dep Hantsin-nesorganen ober- und unterirdisch lebender Astyanax - men. ZMorph Ökol Tiere 61:255–319
Schemmel C (1973) Les organes sensoriels cutanes des genre Astyanax(Pisces, Characidae) chez les formes occupant des biotopes sout-errains. Ann Speleol 28:209–219
Simon P, Dupuis R, Costentin J (1994) Thigmotaxis as an index ofanxiety in mice. InXuence of dopaminergic transmissions. BehavBrain Res 61:59–64
Sneddon LU (2003) The evidence for pain in Wsh: the use of morphineas an analgesic. Appl Anim Behav Sci 83:153–162
Teyke T (1985) Collision with and avoidance of obstacles by blindcave Wsh Anoptichthys jordani (Characidae). J Comp Physiol A157:837–843
Teyke T (1988) Flow Weld, swimming velocity and boundary layer:parameters which aVect the stimulus for the lateral line organ inblind Wsh. J Comp Physiol A 163:53–61
Teyke T (1989) Learning and remembering the environment in theblind cave Wsh Anoptichthys jordani. J Comp Physiol A 164:655–662
Teyke T (1990) Morphological diVerences in neuromasts of the blindcave Wsh Astyanax hubbsi and the sighted river Wsh Astyanax mex-icanus. Brain Behav Evol 35:23–30
Treit D, Fundytus M (1988) Thigmotaxis as a test of anxiolytic activityin rats. Pharm Biochem Behav 31:959–962
Weissert R, von Campenhausen C (1981) Discrimination between sta-tionary objects by the blind cave Wsh Anoptichthys jordani.J Comp Physiol 143:375–381
Wilkens H (1988) Evolution and genetics of epigean and cave Astya-nax fasciatus (Characidae, Pices): support for the neutral muta-tion theory. Evol Biol 23:271–367
Windsor S (2008) Swimming kinematics and hydrodynamic imagingin the blind Mexican caveWsh (Astyanax fasciatus). J Exp Biol211:2950–2959
Wolfer DP, Stagljar-Bozicevic M, Errington ML, Lipp H-P (1998)Spatial memory and learning in transgenic mice: fact or artifact?News Physiol Sci 13:118–123
Yoshida M, Nagamine M, Uematsu K (2005) Comparison of behav-ioral responses to a novel environment between three teleosts,bluegill Lepomis macrochirus, crucian carp Carassius lang-sdorWi, and goldWsh Carassius auratus. Fish Sci 71(2):314–319
Yoshizawa M, JeVery WR (2008) Shadow response in the blind cave-Wsh Astyanax reveals conservation of the functional pineal eye.J Exp Biol 211:292–299
123





























