Embed Size (px)
Citation preview

Mutation Research, 266 (1992) 117-134 © 1992 Elsevier Science Publishers B.V. All rights reserved 0027-5107/92/$05.00
117
MUT 05061
The induction of mitotic chromosome malsegregation in Aspergiilus nidulans. Quantitative structure activity relationship (QSAR) analysis
with chlorinated aliphatic hydrocarbons
R. Crebelli, C. Andreoli, A. Carere, G. Conti, L. Conti, M. Cotta Ramusino and R. Benigni
istituto Superiore di SanitY, Rome (Italy)
(Received 5 February 1991) (Revision received 27 September 1991)
(Accepted 8 October 1991)
Keywords: Aneuploidy; Aspergillus; Haloalkanes; QSAR analysis
Summary
The biological activity of 24 chlorinated aliphatic hydrocarbons has been studied in the mold Aspergillus nidulans. The ability to induce chromosome malsegregation, lethality and mitotic growth arrest has been experimentally determined for each chemical. These data, together with those of 11 related compounds previously investigated, generated a data base which was used for quantitative structure-activity relationship (QSAR) analysis. To this aim, both physico-chemical descriptors and electronic parameters of each compound have been calculated and included in the analysis.
The QSAR analysis indicated that toxic effects induced by chlorinated aliphatics in ,4. nidulans are mainly dependent on steric factors, as indicated by the correlation with molar ,'¢fractivity (MR). Conversely, the ease with which they accept electrons, parametrized by LUMO (energy of the lowest unoccupied molecular orbital), plays a prevailing role in determining the aneuploidizing properties. An involvement of free radicals, generated by the reductive metabolism of haloalkanes, is hypothesized as an explanation of the data.
Aneuploidy, i.e., the occurrence of fewer or more chromosomes than an exact multiple of the haploid set, has dramatic consequences on both somatic and germ cells, it is estimated that a sizeable fraction of all human conceptions (5- 10%) are chromosomally abnormal, thus resulting in early miscarriage (Hook, 1985; Hecht and
Correspondence: Dr. R. Crebelli, lstituto Superiore di Sanit[~, Viale Regina Elena 299, !-00161 Rome (Italy).
Hecht, 1987). Furthermore, aneuploidy is respon- sible for numerous human syndromes (Down, Turner, Klinefelter, etc.) which severely affect physical and mental health (Sankaranarayanan, 1979; Hook, 1985; Hecht and Hecht, 1987). Ge- nomic mutations are also supposed to play a role in the process of neoplastic transformation (Bar- rett et al., 1985; Cavenee et al., 1986; Evans, 1985).
Aneuploidy can be caused by chemical agents that disturb the processes of chromosome segre-

118
gation through multiple mechanisms: all the con- stituents of the mitotic (and meiotic) apparatus can be regarded as potential targets for aneu- ploidizing agents (aneugens): spindle fibers, kine- tochores, centrioles, membranes, etc. (Liang and Brinkley, 1985). Furthermore, interference with physiological processes such as energy production and Ca 2+ homeostasis can affect proper chromo- some segregation by indirect mechanisms leading to microtubule destabilization (Onfelt, 1986a).
Because of the multiplicity of cellular targets potentially affected by aneuploidizing chemicals, and the lack of a unique mechanism for the induction of chromosome malsegregation, it can be anticipated that the relationship between aneugenic activity and chemical structure will be a complex one. However, despite the difficulties inherent in studies on structure-activity relation- ships with aneugenic chemicals, it is likely that well-designed investigations, e.g.,, with series of congeneric chemicals, could provide useful infor- mation on 'structural alerts' for aneuploidy in- duction, and consequently help clarify the mecha- nism underlying the effect observed.
Previous work in this laboratory demonstrated the unusual activity of several chlorinated aliphati¢ hydrocarbons in genetic assays for chro- mosome maisegregation used as indicator of aneugenic potential in the mold Aspergillus nidu. lans (Crebelli et al., 1984, 1985) prompting fur- ther studies in this field. A first analysis of the interrelationships between biological properties of 11 chlorinated methanes and ethanes and their physico-chemical descriptors highlighted a disso- ciation between genetic and toxicity end-points, demonstrating that other parameters, in addition to lipophilicity and sterie factors, play a key role in aneuploidy induction in A. nidulans (Crebeili et al., 1988).
In this present work, the previous data base has been enlarged to 24 saturated and unsatu- rated chlorinated aliphatics which have been studied in detail in the same genetic system (i.e,, malsegregation in A. nidulans). Genetic (aneup- Ioidizing) and toxic (cytostatic and lethal) proper- ties have been experimentally determined for each chemical. In addition, physico-chemical descrip- tors have been calculated (or derived from the literature), together with quanto-mechanical pa-
rameters, and the existing interrelationships have been investigated by means of multivariate statis- tical methods.
Materials and methods
Chemicals Sources, chemical names, code numbers, pu-
rity and CAS numbers of the chlorinated aliphatic compounds investigated are shown in Table 1. Dimethyl sulfoxide (DMSO) was used as solvent. The amount of solvent applied did not exceed 2%. Historical control data demonstrate that, at these doses, DMSO does not affect spontaneous segregation rates of A. nidulans diploid strains nor significantly inhibits colony sporulation.
Strains and genetic systems The A. nidulans diploid strain P1, synthesized
in this laboratory (suAladE20, riboA1, fpaA1, anAl, proA1, pabaA1, yA2, adE20, b/A1; methA1, pyroA4; nicA2; lysB5; nicB8) and the haploid strain 35 (anAl, pabaA1, yA2; methA1, s12; nicA2; nicB8) were used throughout this work.
Euploid yellow segregants in the diploid strain Pl were detected as homozygous (yA2/yA2) or hemizygous (yA2/0) yellow sectors or patches in heterozygous (yA2/yA2 +) pale green colonies growing on complete medium. Mitotic segregants were isolated, often streaked for purification, and analyzed for their nutritional requirements in or- der to distinguish yellow whole chromosome seg- regants (which are p-fluorophenylalanine-re- sistant and require aneurine and p-aminobenzoic acid) from yellow cross-overs (which are p-fluoro- phenylalanine-sensitive and may require p- aminobenzoic acid) (Morpurgo et al., 1979).
Abnormal, presumptive aneuploid colonies in the haploid strain 35 were visually identified and analyzed by 'needle plating' (Killer, 1961) to dis- tinguish aneuploid, unstable types producing nor- mal vigorous sectors from stable abnormals, which are presumed to be mutant types.
Media Czapek-Dox minimal medium (Hopwood and
Sermonti, 1962) and a previously described com- plete medium (Bignami et aL, 1977) were used.

i19
TABLE I
HALOGENATED ALIPHATIC HYDROCARBONS INVESTIGATED IN MITOTIC SEGREGATION ASSAYS IN A. nidu- lans
Code Name CAS No. Purity Source number
1 Dichloromethane * 1665-00-5 > 99% Fluka 2 Chloroform * 865-49-6 > 99% C. Erba 3 Carbon tetrachloride * 56-23-5 > 99.5% C. Erba 4 l,l-Dichloroethane * 75-34-3 97% Janssen 5 1,2-Dichloroethane * 107-06-2 99.5% C. Erba 6 l,l ,l-Trichloroethane * 71-55-6 > 99% Aldrich 7 1,1,2-Trichloroet hane * 79-00-5 98% Aldrich 8 1,1,1,2-Tetrachloroethane * 630-20-6 99% Aldrich 9 1,1,2,2-Tetrachloroethane * 79-34-5 99% Aldrich
10 Pentachloroethane * 76-01-7 > 98% C. Erba 11 Hexachloroethane * 67-72-1 > 98% C. Erba 12 1,1,2-Trichloroethylene 79-01-6 > 99% BDH 13 Tetrachloroethylene 127-18-4 > 99.9% Aldrich 14 1,2-Dichloroethylene a 540-59-0 98% Aldrich 15 l,l-Dichloroethylene 75-35-4 99% Aldrich 16 1,2-Dichloropropane 78-78-5 99% Aldrich 17 2,2-Dichloropropane 594-20-7 98% Aldrich 18 1,3-Dichloropropane 142-28-9 99% Aldrich 19 1,2,3. Trichloropropane 96-18-4 99% Aldrich 20 l-Chlorobutane 109-69-3 99.5% Aldrich 21 2-Chlorobutane 78-86-4 99% Aldrich 22 1,3-Dichlorobutane 1190-22-3 99% Aldrich 23 2,3-Dichlorobutane 7581-97-7 98% Aldrich 24 l-Chloro.2-methylpropane 513-36-0 98% Aldrich 25 2-Chloro.2-methylpropane 507-20-0 99~ Aldrich 26 l.Chloropentane 543-59-9 99% Aldrich 27 I-Chlorohexane 544-10-5 99% Aldrich 28 I-Chlomoctane I '~ 1-85-3 99% Aldrich 29 l,I-Dichloropropene 563-58-6 97% Aldrich 30 2,3-Dichloro- l-propene 78-88-6 98% Aldrich 31 1,3-Dichloropropene ,L 542-75-6 95% Aldrich 32 I,l,3-Trichloropropene 2567-14-8 98% Aldrich 33 3-Chloro-2-chloromethylpropene 1871-57-4 > 99% Aldrich 34 I-Chloro-2-methylpropene 513-37-1 98% Aldrich 35 3-Chloro-2-methylpropene 563-47-3 98% Aldrich
* Raw data of mitotic segregation assays on these compounds have been repo~ted ia a previous paper (Crebelli et al., 1988). a Mixture of cis and trans isomers.
Treatments To obtain precise quantitative data, a liquid
exposure protocol was routinely used for treat- ments with test chemicals. This procedure is used for treatment of mononucleate conidia during early germination (i.e., during germ-tube emer- gence), in order to study the first mitotic division(s) (Bainbridge, 1971). Briefly, conidia (105/ml) were pre-incubated in a semi-liquid (0.15% agar) complete medium at 37°C with
gentle shaking. After 3.5 h, samples of swollen, pregerminating conidia were treated in sealed capped glass tubes with test chemicals until the emergence of germ tubes (usualgy 3-3.5 h). A range of concentrations was applied to obtain dose-effect relationships as well as to determine the lowest concentration arresting conidial germi- nation (ARR) or inducing one lethal hit per cell (37% of survivors, D37). Treatments were inter- rupted by serial dilutions with sterile water and

TA
BL
E 2
CH
LO
RIN
AT
ED
AL
IPH
AT
ICS
EV
AL
UA
TE
D A
S N
EG
AT
IVE
IN
AS
SA
YS
FO
R M
ITO
TIC
SE
GR
EG
AT
ION
IN
A.
nidu
lans
DIP
LO
ID S
TR
AIN
PI
Che
mic
al
conc
en-
Sur
viva
l S
core
d co
de a
tr
atio
n (%
) co
loni
es
(% v
:v) c
Yel
low
seg
rega
nts
b in
abn
orm
al c
olon
ies
Yel
low
seg
rega
nts
b in
nor
mal
col
onie
s
who
le c
hrom
osom
e cr
oss-
over
s w
hole
chr
omos
ome
cros
s-ov
ers
n (%
) n
(%)
n (%
) n
(%)
13
0.02
5 i0
0 43
0 0.
005
100
422
0.01
I0
0 41
4 0.
0125
74
46
5 0.
015
35
234
0.02
7
138
16
0.05
I0
0 28
8 0.
075
100
216
0.1
I00
431
0.15
86
38
9 0.
2 57
53
5 0.
25
17
285
17
0.05
!0
0 22
2 0.
1 89
45
3 0.
15
72
349
0.2
19
238
18
0.01
25
I00
307
0.02
5 lo
ft
750
0.05
89
71
4 0.
1 74
65
6 0.
15
23
312
10
0.02
5 !0
0 53
6 0.
05
74
493
0.07
5 20
40
6 0.
1 5
209
20
0.02
5 i0
0 28
4 0.
05
86
456
0.1
70
372
0.15
26
29
9
0 0
0 6
(1.4
0)
3 (0
.71)
0
1 (0
.24)
2
(0.4
7)
3 (O
.72)
0
0 2
(0.4
8)
2 (0
.43)
0
0 2
(0.4
3)
2 (0
.84)
1
(0.4
3)
0 3
(1.3
3)
0 0
0 1
(0.7
2)
0 0
0 1
(0.3
5)
! (0
.46)
1
(0.4
6)
0 2
(0.9
3)
I (0
.23)
0
0 1
(0.2
3)
! (0
.26)
0
0 1
(0.2
6)
0 0
0 2
(0.3
7)
0 0
0 3
(1.0
5)
0 0
0 3
(1.3
5)
0 0
0 1
(0.2
2)
1 (0
.29)
2
(0.5
7)
0 0
0 0
l (0
.42)
4
0.68
)
I (0
.33)
1
(0.3
3)
1 (0
.33)
4
(1.3
0)
! (0
.13)
0
1 (0
.13)
1
(0.1
3)
2 (0
.28)
0
0 2
(0.2
8)
4 (0
.61)
0
0 4
(0.6
1)
! (0
.32)
1
(0.3
2)
1 (0
.32)
2
(0.6
4)
2 (0
.37)
1
(0.1
9)
0 1
(0.1
9)
1 (0
.20)
1
(0.2
0)
0 1
(0.2
0)
0 0
1 (0
.25)
4
(0.9
9)
0 0
0 0
2 (O
.7O
) 0
0 0
0 1
(0.2
2)
0 3
(0.6
6)
! (0
.27)
0
0 4
(1.0
7)
1 (0
.33)
1
(0.3
3)
0 2
(0.6
7)

21
22
23
24
25
26
27
28
29
0.1
0.15
0.
2 0.
25
0.3
0.02
5 0.
05
0.07
5 0.
1
0.02
5 0.
05
0.07
5 0.
1
0.02
5 0.
05
0.1
0.15
0.
2
0.05
0.
1 0.
15
0.2
0.25
0.02
0.
04
0.06
0.
08
0.01
0.
O2
0.03
0.
04
0.00
625
0.01
25
0.02
5
0.01
25
0.02
5 0.
05
0.06
25
!00 87
73
58
18
100
100
61
23
95
86
69
47
100
100
100
37 7
i00
100
100
72
17
IO0
72
29 8
!00 71
27 5
100
30 8
!00 83
31
9
536
64O
77
7
733
292
545
525
418
235
549
496
6O8
713
247
452
48
t 26
6 13
7
371
226
648
40O
22
6
261
482
313
!17
46
3
512
266 89
6O9
286
295
270
379
286
131
0 2 3 2 I I 2 0 0 0 0 i 0 0 0 0 0 0 0 0 0 2 0 ! 2 I 0 0 i 0 0 2 0 1 I I 0 0
(0.3
1)
(0.3
9)
(0.2
7)
(0.3
4)
(0.1
8)
(0.3
8)
(0.1
6)
(0.5
0)
(0.3
8)
(0.4
1)
(O.3
2)
(0.1
9)
(0.3
3)
(0.3
4)
(I).
37)
(0.2
6)
0 0 0 0 1 0 0 0 0 3 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 I 0 0 0 0 0 0 0 0
(0.3
4)
(0.5
5)
(0.20)
(0.73)
(0.2
5)
(0.38}
0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 1 0 0 1 0 0 0 0 0 1 0 0 0 I 0 0 0 0
(o.1
6)
(o.3
4)
(0.54)
(0.1
5)
(0.3
8)
(0.38)
(0.34)
5 8 2 3 3 3 3 1 0 3 5 1 1 1 2 2 1 1 4 5 5 0 i 0 1 1 0 3 1 2 0 i 2 I 4 6 2 0
(0.93)
(1.25)
(0.26)
(0.41)
(1.03)
(0.55)
(0.57)
(0.24)
(0.55)
(1.01)
(0.1
6)
(0.1
4)
(0.4
0)
(0.4
4)
(0,4
2)
(0.38)
(0.73)
(1.08)
(2.21)
(0.77)
(0.44)
(0.21)
(0.32)
(0.6
5)
(0,1
9)
(0.7
5)
(0.1
6)
(0.7
0)
(0.3
4)
(1.4
8)
(1.58)
(0.7
0)
TA
BL
E 2
(co
ntin
ued)
Che
mic
al
CO
de
a
Con
cen-
S
urvi
val
Sco
red
trat
ion
(%)
colo
nies
(%
v:v
) ~
Yel
low
seg
rega
nts
b in
abn
orm
al c
olon
ies
Yel
low
seg
rega
nts
b in
nor
mal
col
onie
s
who
le c
hrom
osom
e cr
oss-
over
s w
hole
chr
omos
ome
cros
s-ov
ers
n (%
) n
(%)
n (%
) n
(%)
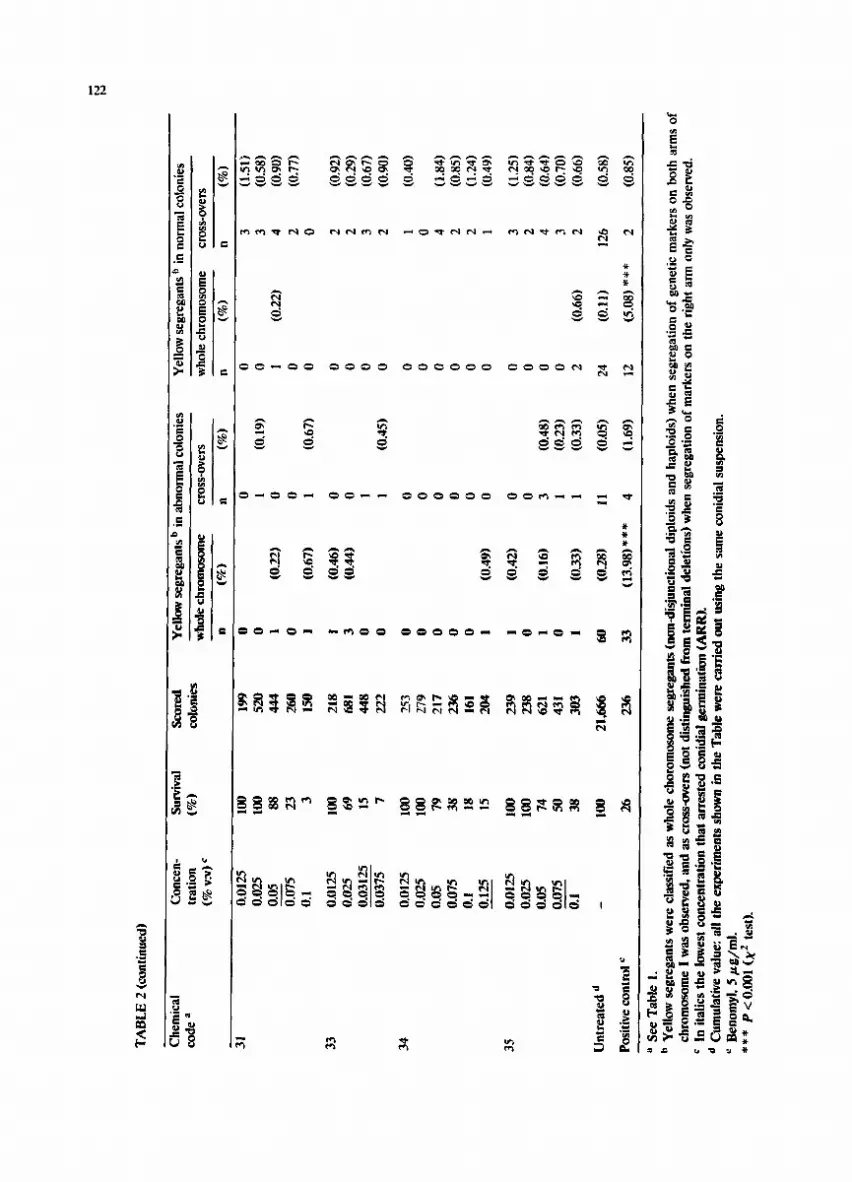
31
0.01
25
100
199
0 0
0 3
(1.5
1)
0.02
5 10
0 52
0 0
1 (0
.19)
0
3 (0
.58)
0.
05
88
444
1 (0
.22)
0
1 (0
.22)
4
(0.9
0)
0.07
5 23
26
0 0
0 0
2 (0
.77)
O
. 1
3 15
0 I
(0.6
7)
1 (0
.67)
0
0
33
0.01
25
100
218
! (0
.46)
0
0 2
(0.9
2)
0.02
5 69
68
1 3
{0.4
4)
0 0
2 (0
.29)
0.
0312
5 15
44
8 0
1 0
3 (0
.67)
0.
0375
7
222
0 1
(0.4
5)
0 2
(0.9
0)
34
0.01
25
100
!~3
0 0
0 1
(0.4
0)
0.02
5 10
0 27
9 0
0 0
0 0.
05
79
217
0 0
0 4
(1.8
4)
0.07
5 38
23
6 0
0 0
2 (0
.85)
0.
1 18
16
1 0
0 0
2 (1
.24)
0.
125
15
204
I (0
.49)
0
0 1
(0.4
9)
35
0.01
25
100
239
I (0
.42)
0
0 3
(1.2
5)
0.02
5 10
0 23
8 0
0 0
2 (0
.84)
0.
05
74
621
1 (0
.16)
3
(0.4
8)
0 4
(0.6
4)
0.07
5 50
43
1 0
1 (0
.23)
0
3 (0
.70)
0.
1 38
30
3 I
(0.3
3)
! (0
.33)
2
(0.6
6)
2 (0
.66)
Unt
reat
ed d
_
100
21,6
66
60
(0.2
8)
11
(0.0
5)
24
(0.1
1)
126
(0.5
8)
Pos
itiv
e co
ntro
l ¢
26
236
33
(13.
98)*
**
4 (1
.69)
12
(5
.08)
**
*
2 (0
.85)
a S
ee T
able
1.
b Y
ello
w s
egre
gant
s w
ere
clas
sifi
ed a
s w
hole
cho
rom
osom
e se
greg
ants
(no
n-di
sjun
ctio
nal
dipl
oids
and
hap
loid
s) w
hen
segr
egat
ion
of g
enet
ic m
arke
rs o
n bo
th a
rms
of
chro
mos
ome
1 w
as o
bser
ved,
and
as
cros
s-ov
ers
(not
dis
ting
uish
ed f
rom
ter
min
al d
elet
ions
) w
hen
segr
egat
ion
of m
arke
rs o
n th
e ri
ght
arm
onl
y w
as o
bser
ved.
c
In i
tali
cs t
he l
owes
t co
ncen
trat
ion
that
arr
este
d co
nidi
al g
erm
inat
ion
(AR
R).
°
Cum
ulat
ive
valu
e: a
ll t
he e
xper
imen
ts s
how
n in
the
Tab
le w
ere
carr
ied
out
usin
g th
e sa
me
coni
dial
sus
pens
ion.
B
enom
yl,
5 ~
g/m
l.
**
*
P<
0.0
01
(;
~2 t
est)
.

the conidia were plated on an agarized complete medium. Only plates with fewer than 20 colonies were used for the detection of mitotic segregants to prevent normal colonies from encroaching on abnormal, slow-growing aneuploids.
Quantitative structure-activity relationship (QS- AR) analysis
Physico-chemical and molecular orbital param- eters are presented in Table 5. The physico- chemical parameters ware: logarithm of the oc- tanol /water partition coefficient (Log P), molar refractivity (MR), estimated free energy of bind- ing to biological receptors (DG), boiling point (BP), and density (DENS). Log P was obtained from the literature or estimated according to Leo's fragment constant method (Lyman et al., 1982). MR was calculated according to Vogei's method (Lyman et al., 1982). DG was calculated as in Andrews et al. (1984). BP and DENS were found in the literature.
The molecular orbital parameters were: the energy of the highest occupied molecular orbital (HOMO), the energy of the lowest unoccupied molecular orbital (LUMO), the greatest bond length between a carbon and a halogen (DIST), the charge on the carbon bonded with the most distant halogen (CHARGE), the variance of the net charge of the atoms of the molecule (VARCH), and the difference L U M O - HOMO (DIF). Ab initio molecular orbital calculations were by a Gaussian 80 program, with fully opti- mized geometries (STO-3G, Murtangh-Sargent option).
Statistical methods The methods used for the exploration of
QSARs were: factor, canonical correlation, step- wise regression and stepwise linear discriminant analyses. The BMDP Statistical Software package was used (Dixon, 1981) (programs: BMDP4M, BMDP6M, BMDP2R, and BMDPTM).
Results
Biological activity of chlorinated aliphatics in As- pergillus
Twenty-four chlorinated aliphatic hydrocar- bons were assayed in the A. nidulans diploid
123
strain P1 to determine their effect on mitotic chromosome segregation, their ability to arrest conidial germination (and mitosis) and their cyto- toxic activity.
The chlorinated compounds investigated were: 4 chlorinated ethylenes, 4 propanes, 6 butanes, 4 propenes, 3 methyl propenes and 3 n-saturated aliphatics (l-chloropentane, 1-chlorohexane and l-chlorooctane) (see Table 1).
With each compound, pilot experiments with extensive microscope observation were carried out in order to determine the appropriate dose range for further testing. To this end, 4 doses in the range of concentrations which produced a delay or reversible arrest of conidial germination were selected. Usually the lowest concentration as- sayed slighly delayed the emergence of g~rm tubes, and the highest one induced severe delay or arrest of conidial germination. When the high- est dose tested induced less than one lethal hit per cell (i.e., > 37% survivors), 1 or 2 additional doses were assayed. The extensive work carried out with mitotic segregation systems in A. nidu- lans demonstrates that such a dose choice is appropriate for the detection of most aneugenic agents, which show up as positive in the dose range where mitosis of germinating conidia is impaired or reversibly arrested.
Nineteen out of the 24 compounds assayed were unable to induce reproducible, significant increases in segregation rates at any of the con- centrations assayed (Table 2). Therefore these chemicals were evaluated as negative, i.e., inef- fective in malsegregation induction in Aspergillus. Establishing evidence for a lack of activity in mitotic segregation assays may be more difficult than identifying a positive effect. Spontaneous segregation rates are usually low in A. nidulans and therefore, borderline positive effects may re- main undetected if a low number of colonies is scored. However, taking into account the sponta- neous segregation frequencies, the sample size of the experiments was sufficient in most cases to rule out the possibility of small (2-4-fold) in- creases in malsegregation rates with P = 95% (Mann et al., 1985).
The remaining 5 compounds, namely 1,1,2-tri- chloroethylene (code number 12), 1,2-dichloro- ethylene (14), 1,1-dichloroethylene (15), 2,3-di-

TA
BL
E 3
CH
LO
RIN
AT
ED
AL
IPH
AT
ICS
EV
AL
UA
TE
D A
S P
OS
ITIV
E 1
N A
SS
AY
S F
OR
MIT
OT
IC S
EG
RE
GA
TIO
N
IN A
. ni
dula
ns D
IPL
OID
ST
RA
IN P
I
Che
mic
al
Con
cen-
S
urvi
val
Sco
red
code
a
trat
ion
(%)
colo
nies
(%
v.~
) ~
Yel
low
scg
rcga
nts
b in
abn
orm
al c
olon
ies
Yel
low
seg
rega
nts
b in
nor
mal
col
onie
s
who
le c
hrom
osom
e cr
oss-
over
s w
hole
chr
omos
ome
cros
s-ov
ers
n (~
) n
(%)
n (%
) n
(%)
12
0.02
10
0 42
5 0
1 (0
.24)
0
2 (0
.47)
0.
04
100
310
4 (1
.29)
*
0 0
4 (1
.29)
0.
06
93
357
5 ( !
A0)
* *
0
0 3
(0.8
4)
0.07
90
38
4 8
(2.0
8) *
**
0
0 4
(1.0
4)
0.08
72
45
8 17
(3
.71)
**
*
0 0
2 (0
.44)
0.
09
52
588
14
(Z38
) *
**
0
0 1
(0.1
7)
0.1
10
385
12
(3.1
1) *
**
0
4 (1
.04)
0
0.11
5
214
5 (2
.33)
* *
* 0
0 0
0.12
3
195
21
(i.0
2)
0 1
(0.5
1)
0
14
0.05
94
24
8 3
(!.2
1)
0 0
1 (0
.40)
0.
075
80
186
5 (Z
68)
**
*
0 0
2 (1
.07)
0.
1 71
25
9 i0
(3
.86)
**
*
0 1
(0.3
9)
1 (0
.39)
0.
125
64
354
13
(3.6
7) *
**
0
1 (0
.28)
3
(0.8
5)
0.15
51
29
9 13
(4
.35)
**
*
0 0
1 (0
.33)
0.
162
40
331
3 (0
.90)
0
0 3
(0.9
1)
0.17
5 30
37
4 4
(1.0
7)
0 0
3 (1
.07)
15
0.02
5 10
0 26
0 2
(0.7
7)
0 0
4 (1
.54)
0.
05
93
815
9 (1
.10)
* *
0
0 10
(1
.23)
0.
1 85
39
2 8
(2.0
4) *
**
0
0 2
(0.5
1)
0.1 2
5 70
33
4 8
(2.3
9) *
* *
1
(0.3
0)
1 (0
.30)
2
(0.6
0)
0.15
50
24
9 5
(2.4
0) *
**
0
0 3
(1.2
0)
0.17
5 41
44
0 10
(2
.27)
**
*
1 (0
.23)
0
0 0.
2 20
23
3 2
(0.8
6)
0 1
(0.4
3)
1 (0
.43)
30
0.01
5 lO
0 22
5 0
1 (0
.44)
1
(0.4
4)
4 (1
.78)
0.
0175
10
0 21
0 0
1 (0
.48)
0
4 (1
.90)
0.
0187
83
35
0 6
(I.7
1)
0 0
2 (0
.57)
0.
02
73
395
13
(3.2
9)
1 (0
.25)
1
(0.2
5)
2 (0
.51)
0.
0225
59
54
0 I1
(2
.04)
5
(0.9
2) *
**
2
(0.3
7)
5 (0
.92)
0.
025
39
821
44
(5.3
6)
5 (0
.61)
**
*
0 3
(0.3
7)
0.02
6 29
23
1 9
(3.8
9)
1 (0
.43)
0
2 (0
.87)
0.
027
22
285
9 (3
.16)
3
(1.0
5)
0 0
0.02
75
< 0.
1 0
32
0.00
25
100
336
! (0
.30)
0
0 1
(0.3
0)
0.00
5 95
25
8 7
(2.7
1) *
* *
0
1 (0
.39)
2
(0.7
8)
0.00
75
65
204
29
(14.
20)
* *
* 2
(0.9
8)
2 (0
.98)
2
(0.9
8)

Un
trea
ted
d
Pos
itiv
e co
ntro
l c
0.01
40
40
3 30
(7
.44)
**
* 3
0.01
25
36
310
30
(9.6
7) *
**
3
0.01
5 35
37
0 26
(7
.02)
**
*
6 0.
0175
36
18
1 12
(6
.62)
**
*
1 0.
02
28
339
22
(6.4
9) *
**
5
0.02
25
19
154
7 (4
.54)
**
*
2 0.
025
19
153
5 (3
.27)
**
*
5 0.
03
13
197
3 (1
.52)
8
- 10
0 2,
553
6 (0
.24)
1
30
240
30
(12.
50)
* *
* 1
(0.7
4)
3 (0
.74)
3
(0.7
4)
(0.9
7)
3 (0
.97)
3
(0.9
7)
(1.6
2) *
**
3 (0
.81)
2
(0.5
4)
(0.5
5)
0 2
(1.1
0)
(1.4
7) *
**
1
(0.2
9)
1 (0
.29)
(1
.30)
2
(1.3
0)
3 (1
.95)
(3
.27)
**
*
0 1
(0.6
5)
(4.0
6) *
**
0
1 (0
.51)
(0.0
4)
2 (0
.08)
16
(0
.63)
(0.4
2)
3 (1
.25)
5
(2.0
8)
a-d
See
foo
tnot
es t
o T
able
2.
Thi
aben
dazo
le,
40
/zg
/ml.
*
p<
o.o
s;*
*p
<O
.Ol;
**
*p
<o
.OO
l(x
2te
st).
TA
BL
E 4
IND
UC
TIO
N O
F M
OR
PH
OL
OG
ICA
L
AB
NO
RM
AL
CO
LO
NIE
S
IN A
. ni
dula
ns H
AP
LO
ID S
TR
AIN
35
Tre
atm
ent
Con
cert
- S
urvi
val
Sco
red
Abn
orm
al c
olon
ies
trat
ion
(%)
colo
nies
al
l ty
pes
stab
le
unst
able
(%
v:v
) n
(%)
n (%
)
Un
trea
ted
-
100
2,64
6 6
(0.2
3)
1 (0
.04)
2,
3-D
ichl
oro-
pr
open
e 0.
025
33
747
31
(4.1
5) *
* *
0
!, 1
,3-T
rich
loro
- pr
open
e 0.
0125
92
40
9 22
(5
.38)
* *
* 1
(0.2
4)
Pos
itiv
e co
ntro
l b
33
103
25
(24.
27)
* **
0
n
(%)
5 (0.19)
28
(3.75) *
* *
18
(4.40) *
* *
22
(21.36)
* * *
unde
fine
d a
[]
(%)
0 3 (0.40)
3 (0.73)
3 (2.91)
a S
emi-
leth
al o
r ac
onid
ial
colo
nies
whi
ch c
ould
not
be
succ
es.~
fully
an
alyz
ed b
y re
plat
ing.
b
Ben
omyl
, 5
~tg
/ml.
*
**
P
<0
.00
1
(X 2
tes
t).
L~

TA
BL
E 5
CH
EM
ICA
L A
ND
BIO
LO
GIC
AL
DE
SC
RIP
TO
RS
OF
CH
LO
RIN
AT
ED
AL
IPH
AT
IC H
YD
RO
CA
RB
ON
S
Co
Che
mic
al
Che
mic
o-ph
ysic
al p
aram
eter
s
(cod
e)
LO
G P
D
G
MR
B
P
Mol
ecul
ar o
rbit
al p
aram
eter
s B
iolo
gica
l par
amet
ers
DE
NS
H
OM
O
LU
MO
D
IST
C
HA
RG
E
VA
RC
H
LE
C
AR
R
D37
INCR
I 1.
260
- 10
.600
16
.335
39
.900
1.
325
-0.4
00
0.
289
!.80
3 0.
117
0.01
6 62
.700
94
.000
14
1.00
0 11
.200
2
1.98
0 -9
.30
0
21.1
51
61.0
00
1.49
2__
-0.4
18
0.
231
!.81
1 0.
077
0.01
2 20
.000
25
.000
27
,500
4.
800
3 2.
830
- 8.
000
25.9
67
77.0
00
i..59
4 -0
_437
0.
174
!.81
8 0.
039
0.00
8 4.
100
10.4
00
7.30
0 11
.800
4
!.80
0 -9
.80
0
20.9
60
57.0
00
i.77
6 -0
.38
6
0-29
2 1.
820
0.14
1 0.
017
23.8
00
47.6
00
38.1
00
4.40
0 5
[.48
0 -
10.5
00
20.9
82
83.0
00
i.25t
~ --
0.38
0 0.
288
1.80
9 0.
150
0.01
4 25
.400
50
.800
44
.400
20
.100
(,
2.50
0 -8
.50
0
25.7
58
75.0
00
!.33
8 -0
.41
1
0.23
2 1.
821
0.09
6 0,
016
500.
000
10.0
00
11.0
00
1.00
0 "
2.07
0 -9
.20
0
25.7
98
!12.
500
1.43
5 -0
.39
3
0.25
9 !.
818
0.12
3 0.
014
8.10
0 10
.700
11
,800
9.
900
8 3.
050
- 7.
900
30.6
14
138.
000
1.59
8 -0
.40
7
0.21
7 !.
822
0.08
2 0.
013
1.20
0 2.
800
2.40
0 49
.600
t~
2.
660
- 7.
900
30.6
14
147.
000
1.58
6 -0
.4~
0.
235
!.81
6 0.
108
0.01
3 1.
900
2.80
0 3.
300
3,50
0 I0
5.
720
-6.6
00
35
.430
16
1.50
0 1.
680
-0.4
06
0.
__~6
1.
822
0.07
3 0.
010
500.
000
1.70
0 1.
200
1.00
0 11
4.
620
-5.3
00
40
.246
18
6.00
0 2.
091
-0.4
18
0-
189
!.82
0 0.
045
0.00
7 50
0.00
0 0.
800
16.9
00
1.00
0 12
0.
960
-8.7
00
25
.317
86
.900
1.
464
--0_
_344
0.
225
!.77
5 0.
070
0.00
8 4.
500
11.1
00
10.6
00
13.2
00
13
1.27
0 -
7.40
0 30
.133
12
1.00
0 1
.69
.3
--03
-18
0.20
1 1.
769
0.04
1 0.
004
500.
000
1.20
0 1.
500
1.00
0 14
0.
650
- 10
.000
20
.501
54
.000
! ~
.265
--
0.33
8 0-
251
1.77
2 0.
114
0.01
5 11
.200
29
.900
25
.500
15
.500
15
!.
210
- 10
.000
20
.501
31
.000
1.
213
--0.
334
0.25
6 1.
776
0.09
4 0.
012
6.30
0 25
.000
21
.900
8.
600
16
2.02
0 -9
.70
0
25.6
07
95.5
00
1.15
6 --
0.38
0 0.
291
1.82
6 0.
030
0.02
3 50
0.00
0 15
.400
17
.400
1.
000
17
2.3
40
-
9.00
0 25
.585
67
.000
1.
082
- 0-
381
0.29
1 1.
828
0.13
7 0.
021
500.
000
19.2
00
17.2
00
1.00
0 18
1.
740
- 10
.400
25
.629
12
1.00
0 !.1
92-
-0.3
79
0.
343
1.81
6 --
0.05
6 0.
021
500.
000
10.5
00
13.7
00
1.00
0 19
2.
010
-9.1
00
30
.445
15
6.00
0 i.
387
-0.3
89
0.
273
!.81
9 0.
030
0.02
2 50
0.00
0 9.
400
6.70
0 1.
000
20
2.39
0 -
10.9
00
25.4
38
77.5
00
0.88
6 -0
.36
8
0.35
8 1.
812
--0.
056
0.01
0 50
0.00
0 14
.400
12
.400
1.
000
21
2.57
0 -
10.2
00
25
Ai6
69
.000
0.
873
-03
63
0.
353
1.82
6 0.
030
0.01
2 50
0.00
0 18
.900
25
.500
1.
000
22
2.28
0 -
9.60
0 30
.254
13
4.00
0 1.
115
- 0.
373
0.33
8 1.
824
0.03
0 0.
014
500.
000
8.80
0 8.
300
1.00
0 23
2.
560
-8.9
00
30
.232
11
8.00
0 1.
107
--0.
373
0.31
7 1.
823
0.03
0 0.
014
500.
000
8.70
0 10
.500
1.
000
24
2.44
0 -
10.2
00
25-4
16
68.5
00
0.88
3 -0
.36
8
0.36
0 1.
814
-0.0
56
0.
012
500.
000
14.3
00
14.3
00
1,00
0 25
2.
440
- 9.
500
25.3
94
51.5
00
0.85
! -
0.36
2 0.
349
1.83
3 O
. 106
0.
015
500.
000
18.4
00
23.9
00
! .00
0 26
3.
110
- 10
.800
30
.085
10
7.50
0 0.
882
--0.
369
0.36
0 1.
814
-0.0
54
0.
009
500.
000
5.00
0 5.
000
1.00
0 27
3.
650
- 10
.700
34
.699
13
3.50
0 0.
879
-0.3
68
0.
360
1.81
3 -0
.05
4
0.00
8 50
0.00
0 2.
900
2.20
0 1.
000
28
4.85
0 -
6.30
0 43
.979
18
3.00
0 0.
875
- 0.
367
0.36
0 1.
814
- 0.
054
0.00
8 50
0.00
0 1.
500
1.00
0 1.
000
29
1.79
0 -9
.20
0
25.1
26
76.5
00
!.16
9 -0
22
7
0.26
2 1.
780
0.07
0 0.
012
500.
000
6.60
0 5.
000
1.00
0 30
1.
750
-9.9
00
25
.148
94
.000
1.
204
-0.3
39
0.
246
1.81
3 -0
.05
8
0.01
3 2.
000
2.70
0 2.
700
19.1
00
31
!.19
(I
-9.9
00
25
.148
11
2.00
0 1.
181
-0.3
37
0.
245
1.81
3 -0
.05
8
0.01
2 50
0.00
0 5.
300
7.80
0 !.
000
32
1.50
0 -
8.60
0 29
.964
13
1.50
0 1.
403
- 0.
345
O. ~
220
1.81
1 -
0.06
2 0.
011
0.50
0 2.
400
1.70
0 50
.700
33
2.
010
-9.8
00
29
.795
13
8.00
0 1.
080
-0.3
47
0.
245
1.81
8 -0
.06
3
0.01
3 50
0.00
0 2.
700
2.60
0 1,
000
34
1.89
0 -
9.70
0 24
.935
68
.0fl
0 0.
920
- 0.
305
0.24
4 !.
781
- 0.
040
0.01
3 50
0.00
0 12
. 700
9,
700
1.00
0 35
1.
890
- 10
.400
23
.929
75
.000
0.
917
-0.3
25
0.
3.80
1.
818
-0.0
61
0.
012
500.
000
7.60
0 10
,100
1.
000
Log
P,
part
itio
n co
effi
cien
t o
ctan
ol/
wat
er;
MR
, m
olar
ref
ract
ivit
y; D
G,
free
ene
rgy
of b
indi
ng t
o bi
olog
ical
rec
epto
rs (
see
text
); B
P,
boil
ing
poin
t; D
EN
S,
dens
ity;
H
OM
O,
ener
gy o
f th
e hi
ghes
t oc
cupi
ed m
olec
ular
orb
ital
; L
UM
O,
ener
gy o
f th
e lo
wes
t un
occu
pied
mol
ecul
ar o
rbit
al;
D1S
T,
high
est
bond
len
gth
carb
on-c
hlor
ine;
C
HA
RG
E,
char
ge o
n th
e ca
rbon
bo
nd
ed w
ith
the
mos
t di
stan
t ha
loge
n: L
EC
, lo
wes
t ef
fect
ive
conc
entr
atio
n (m
mol
ar)
in m
alse
greg
atio
n in
duct
ion.
The
val
ue 5
00 w
as
arbi
trar
ily
assi
gned
to
inef
fect
ive
com
poun
ds;
AR
R,
low
est
conc
entr
atio
n (m
mol
ar)
arre
stin
g co
nidi
al g
erm
inat
ion;
1)3
7, c
once
ntra
tion
(m
mol
ar)
asso
ciat
ed w
ith
37%
su
rviv
al.

chloropropene (30) and l,l,3-trichloropropene (32), produced reproducible increases in the fre- quency of euploid whole chromosome segregants (non-disjunctional diploids and haploids) and were further analyzed in order to obtain detailed dose-effect relationships (Table 3).
1,1,2-Trichloroethylene, 1,2-dichloroethylene, and 1,1-dichloroethylene displayed a pattern of activity typical of spindle poisons, inducing only unstable, morphological abnormal colonies with whole chromosome segregation (presumably hy- perhaploids and hyperdiploids), in a range of doses where mitotic growth was affected, but not arrested. Therefore, these chemicals were evalu- ated as positive for chromosome malsegregation induction.
In addition, 1,1,3-trichloropropene (and to a lesser extent 2,3-dichloropropene) also produced increased rates of cross-overs (not distinguished from terminal deletions), which are diagnostic for induced DNA damage (Crebelli and Carere, 1987). In this case, the nature of the primarily induced genetic evem cannot be established with- out further testing, due to possible secondary malsegregation of chromosomes in nuclei with unbalanced structural aberrations (Killer, 1986; Kiifer et al., 1986). Therefore, additional assays were carried out with a haploid strain, where secondary malsegregation is not expected to oc- cur (Kiifer et al., 1986).
The results obtained in assays with strain 35 are shown in Table 4. Exposure to l,l,3-trichloro- propene, 2,3-dichloropropene and to the positive control henomyl, significantly (P<0.001) in- creased the frequency of putative hyperploids, i.e., morphologically abnormal, unstable types producing vigorous wild-type sectors (g~ifer and Upshall, 1973). In conclusion, also l,l,3-trichloro- propene and 2,3-dichloropropene directly affect the segregation of chromosomes at mitosis in A. nidulans.
Interrelationships among cytotoxic, cytostatic and aneugenic properties of chlorinated aliphatics in Aspergillus
For each of the halogenated compounds stud- ied, the following parameters were experimen- tally determined or calculated by linear interpola- tion: LEC (lowest effective concentration in
127
TABLE 6
C O R R E L A T I O N M A T R I X OF B I O L O G I C A L PARAME- TERS
LEC A R R D37 INCR
LEC 1.000 A R R 0.969 1.000 D37 0.982 0.959 1.000 INCR - 0.305 - 0.324 - 0.261 1.000
maisegregation induction); INCR (the highest fold increase in malsegregant rates produced by chem- ical treatment in comparison with the historical spontaneous frequency); ARR (the lowest con-' centration producing a complete block of conidial germination); D37 (the dose producing 37% sur- vivors). These values are shown in Table 5, to- gether with those of a further 11 chlorinated methanes and ethanes (codes 1-11) previously studied in the same system (Crebelli et al., 1988). The correlation matrix of these parameters is shown in Table 6.
Among the positive compounds (13 chemicals), the efficiency of aneuploidy induction, i.e., the lowest active concentration required to exert a significant effect (LEC), was strongly correlated to the cytostatic activity of the chemicals (ARR), with Pearson's r value = 0.969. This value high- lights a general rule that holds for mitotic segre- gation induction in Aspergillus, i.e., that the range of effective doses is just below ARR, where mi- totic division is impaired but not completely blocked (Wood and Killer, 1969; Shanfield and K~ifer, 1971; Kiifer, 1986). Despite the strong correlation observed between LEC and ARR in the subset of positive compounds, the occurrence of a mechanistic link between chromosome malsegregation and induced block of mitotic de- velopment is not clearly seen. In fact, comparable arresting activities were found with most of the negative compounds investigated (Table 5).
A strong correlation was also found between ARR and D37, independently from the aneu- ploidizing properties of the chemicals, with r-- 0.959 (positives only) and r -- 0.950 (all data set).
Contrary to the effeciciency in aneuploidy in- duction (described by LEC), the effectiveness, i.e., the extent of the effect induced (described by

128
INCR), was not significantly related to any of the parameters considered (Table 6). These data con- firm and reinforce the conclusion of previous analyses carried out on a smaller data base (Crebelli et al., 1988), emphasizing the dissocia- tion between efficiency and effectiveness in aneu- ploidy induction in this biological system.
QSARs of aneugenic, cytotoxic and cytostatic ac- tion of aliphatic halogenated hydrocarbons
Quantitative structure-activity relationships (QSARs) have been formulated for the muta- genie action of several families of chemicals (Be- nigni et al., 1989; Frierson et al., 1986; Lopez de Compadre et al., 1990; Shusterman et al., 1989). The QSARs are useful for the identification of the structural features of a chemical influencing its mutagenicity, hence giving insights into the chemical mechanisms of mutagenesis. A crucial point of a QSAR analysis is the choice of the correct correlation when several alternative struc- ture-activity relationships are suggested by the data. This is because, in a given set of chemicals, 2 or more chemical properties can be intercorre. lated purely by chance. The identification of the chemical property that has a mechanistic correla- tion with biological activity becomes more diffi- cult. In fact, the parameter that better fits the data may not be the best parameter for under- standing the mechanism of the effect. To avoid this problem, we first performed a global expio. ration of the interrelationships among chemical properties (via factor analysis) and of the rela- tionships among chemical properties and biologi. cal activity (canonical correlation analysis), then we formalized the QSAR of each biological activ- ity under study (ARR, D37, LEC) within the framework of that general picture.
Factor analysis The chemical parameters were initially ana-
lyzed by factor analysis. Factor analysis is a multi- variate data analysis technique that indicates which variables (parameters, or descriptors) are intercorrelated. Correlated variables basically de- scribe the same effect or phenomenon. Moreover, the analysis generates factors, which are new artificial valiables (linear combinations of the original variables). Each factor summarized the
T A B L E 7
F A C T O R A N A L Y S I S
F A C T O R L O A D I N G S
O F C H E M I C A L P A R A M E T E R S :
Pa ramete r Fac tor 1 Factor 2 Fac tor 3
DG 0.888 - 0.282 - 0.013
MR 0.835 0.353 - 0.352
BP 0.820 0.218 - 0.261
Log P 0.785 0.442 - 0.023
D I F 0.076 - 0.880 - 0.329
L U M O - 0.388 0.862 - 0.055
DIST 0.314 0.662 0.449
DENS 0.574 - 0.635 0.385
C H A R G E - 0 . 0 0 2 - 0 . 4 1 7 0.747
H O M O - 0.558 - 0.115 - 0.726
V A R C H - 0.375 0.202 0.600
information represented by a group of intercorre- lated variables. Thus, factors summarize the phe- nomenon by representing the basic underlying effects, and can be used for every kind of analysis as an alternative to the original variables. The contributions of each original variable to the fac- tors are expressed by factor Ioadings (correlation coefficients of the original variables with the fac- tors) (Lebart et al., 1984).
In the present set of chemicals, factor analysis produced 3 factors (35, 27, and 19% of the vari- ance, respectively), thus pointing to 3 basic chem- ical properties of the compounds. Inspection of factor loadings (Table 7) permitted the identifica- tion of the factors as: (a) stericity (factor I), because of its correlation with MR; (b) elec- trophilicity (factor 2), because of its correlation with LUMO; and (c) electron-donating ability (factor 3), because of its correlation with HOMO.
Canonical correlation analysis Canonical correlation analysis of chemical ver-
sus biological parameters was also performed (Table 8). This technique points to the relation- ships between 2 sets of variables of different nature (here, biological and chemical variables). The analysis generates canonical variables (which are equivalent to the factors used in factor analy- sis) for each set of variables. This operation has the constraint that the first factor of one set is

T A B L E 8
C A N O N I C A L C O R R E L A T I O N A N A L Y S I S : C A N O N I C A L V A R I A B L E L O A D I N G S
Cor re la t ions o f canonical var iables (C NVR } with original vari- ab les
C N V R F I C N V R F 2
Biological parameters A R R 0.926 0.057 D37 0.768 - 0.012 L E C - 0.552 0.800
C N V R S I C NVR S 2
Chemical parameters M R - 0.853 0.043 BP - 0.762 - 0.169 C H A R G E 0.670 - 0.227 Log P - 0.552 0.143 L U M O 0.092 0.821 D E N S 0.093 - 0.746 D I F 0.144 0.607 D G - 0 . 5 1 3 - 0 . 4 4 2 V A R C H 0.385 0.404 H O M O - 0.109 0.332 D I S T - 0.214 0.216
maximally correlated with the first factor of the other set, and so on. The inspection of the corre- lations of the original variables with the canonical variables indicates what variables of one set are correlated with the variables of the other set.
For this analysis, compounds that did not in- duce aneuploidy were arbitrarily assigned a very high LEC value (500 mM). The first pair of canonical variables pointed to the correlation of cytotoxic and cytostatic effects (ARR and D37) with the chemical parameters MR and BP (re- lated to steric factors) and C H A R G E (related to electron-donating ability) (for the interpretation of variables, see Table 7). The second pair of canonical variables indicated a correlation be- tween LEC, and LUMO and DENS (both related to electrophilicity according to Table 7). Since LEC - with its arbitrary values for non-aneup- Ioidizing chemicals - here mainly represents the contrast between genetically positive and negative compounds, this result points to electrophilicity as an important component of aneuploidizing ac- tion in this experimental system.
129
QSAR analysis The QSARs for each biological activity were
studied in detail. To minimize the probability of chance correlations (Topliss and Edwards, 1979), the analyses were performed against small groups of the original variables (for example, MR, LUMO and HOMO, representing the 3 funda- mental properties highlighted by factor analysis), or against the factors.
Cytotoxic and cytostatic effects - highly corre- lated in this data set (see Table 6) - were mainly explained by the steric component, with minor electronic components (stepwise regression analy- sis):
Log(I /D37) = -0 .65 + 0.08 MR - 3,57 LUMO + 4.15 H O M O n = 34; r = 0.94; F ratio = 71.81; P=O.O000 (F ratios of individual variables at step 0: MR = 70.44; LUMG = 1.83; HOMO, = 4.15)
L O G ( I / D 3 7 ) = -0 .79 + 0.08 IVlP. - 3.68 DIF n =34; r=0.9,4; F ra t io= 110.21; P=0 .0000 ( F ratios of individual variables at step 0: MR = 70.44; DIF = 2.48)
Log( l /D37) = -0 .99 + 0.41 F I - 0.24 F3 - 0.13 F2 n = 34; r = 0,92; F ratio = 36.65; P = 0.0000 (F ratios of individual variables at step 0: FI = 34.98; F3 = 9,03; F2 = 3.98)
L o g ( I / A R R ) = -0 .61 + 0.08 MR - 3.10 LUMO + 4.03 H O M O n = 35; r = 0.92; F ratio = 60.47; P=O.O000 (F ratios of individual variables at step 0: M R = 7 6 . 3 5 ; L U M O = 3 . 0 5 ; H O M O = 0.00)
Log(1 /ARR) = -0 .83 + 0.07 MR - 3.26 DIF n - 35; r = 0.92; F ratio = 91.4; P -- 0.0000 (F ra- tios of individual variables at step 0: MR = 76.35; DIF = 2.83)
Log(1 /ARR) = -0 .92 + 0.36 F I - 0.22 F3 - 0.13 F2 n =35; r=0 .91 ; F ratio =38.82; P= 0.0000 ( F ratios of individual variables at step 0: F1 = 40.47; F3 = 8.64; F2 = 2.74)
The similarity of the sets of equations obtained for A R R and D37 is remarkable. This similarity

130
44
110
M R
10o
30- 19 o
3
16-
I )375
26 o
2? 0
6 e 9e 19o 230 220 260 32 e 33 °
2O
12e° 36 • 17~I 6 18Ooooo ~,~'30 o2, • l~s
35 21
2o 16e See4
11 16o
i I 0.262 0.350
l .umo FiB. 1, Planar representation of the distribution of MR and LUMO values in halogenated aliphatic hydrocarbons evalu- ated as positive (closed symbols) and negative (open symbols) in maisegregation assays in A. nidulans. For coda numbers of
tested chemicals, see Table I.
reflects their intercorrelations. The consistency of the equations based on the original variables and factors should also be noted; in both cases, steric effects (MR and F1) represent the chemical property most correlated with biological activity. The electronic component is expressed either by both LUMO and HOMO or by DIF (= LUMO -HOMO). DIF parametrizes the energy dis- tance between the highest occupied and lowest unoccupied molecular orbitais, it measures the ease of transfer of an electron from one energy level to the next, thus parametrizing chemical reactivity.
The repartition between the aneugenic and inactive chemicals was studied by stepwise linear discriminant analysis. This analysis confirmed the involvement of electrophilicity in the phe- nomenon: aneugens were more electrophilic than non-aneugens (classification 1) (see Fig. 1). Alter- native sets of variables were also able to discrimi-
nate between aneugens and non-aneugens; in ev- ery case, a term related to electrophilicity was present in the discriminant equations (DENS and F2) (see Table 7).
Classification 1: F-values, MR: 8.28, LUMO: 9.44; 80% correct Classification 2: F-values, DENS: 6.27, MR: 4.21; 86% correct Classification 3: F-values, F2: 13.8, F3: 6.0; 77% correct
The analysis of the modulation of LEC in the subset of aneugenic chemicals was also per- formed. Within the group of active compounds, LEC was strongly correlated with ARR and D37 (see Table 6). This correlation was confirmed by the QSAR of LEC, which had the same form of the QSARs of ARR and D37 (see above).
Log(1/LEC) = 1.77 + 0.08 MR - 7.12 DIF n = 13; r = 0.94; F ratio--40.31; P = 0.0000 (F ratios of individual variables at step 0: MR--21.55; DIF = 10.84)
Log(I/LEC) -- -0 .37 - 0.42 F2 + 0.44 F1 - 0.36 F3 n = 13; r -- 0.95; F ratio = 7.16; P < 0.02 (F ratios of individual variables at step 0 : F 2 = 7.57; F1 -- 6,26; F3 = 3.66)
Log( I /LEC) = 1.66 + 0.08 MR + 7.51 H O M O - 6.41 LUMO n- -13 ; r=0.94; F ratio--20.40; P=0.0000 (F ratios of individual variables at step 0: MR--21.55; HOMO = 0.50; L U M O = 8.91)
Discussion
The extensive investigation carried out on the physiology and ultrastructure of the mitotic appa- ratus has led to the identification of the struc- tures primarily involved in chromosome distribu- tion (including centrosomes, kinetochores and spindle microtubules) and has given insights into their functioning and regulation (see, e.g., Paweletz and Schroeter, 1987; Hyams and Brink. ley, 1989). This knowledge provides the scientific background to analyze the mechanism of action of those chemicals which affect chromosome dis-

tribution and induce aneuploidy in experimental systems. In particular, the interaction of chemical compounds with spindle microtubules has been extensively investigated, leading to a detailed de- scription of the mechanism of action of spindle poisons such as colchicine, vinblastine, taxoi, which directly interfere with microtubule assem- bly/ disassembly (Sakai et ai., 1982; Dustin, 1984).
Even though numerous chemicals have been identified which affect microtubule polymeriza- tion in vitro (Grrschel-Stewart et al., 1985; Zim- mermann et al., 1985, 1986; Albertini et al., 1988; Wallin et al., 1988; Brunner et al., 1991), and in spite of the great amount of data produced by in vitro short-term tests for chromosome malsegre- gation (Dellarco et al., 1986; Moustacchi et al., 1986), so far only limited structure-activity inves- tigation has been carried out. Apart from a more recent investigation performed on benzimidazole derivatives (Lacey and Watson, 1985), most of the information available on 'structural alerts' for aneuploidy derives from studies carried out in the 1940s (Ostergren and Levan, 1943; Ostergren, 1944), which associated the lypophilicity of chem- ically unrelated compounds wi'a the induction of mitotic spindle disturbance i'a Allium cepa root tips. This suggested the occurrence of a non- specific mechanism for the chemical induction of spindle disturbance, possibly related to physico- chemical interaction with hydrophobic cellular structures. The involvement of membrane dam- age was postulated (as reviewed in Onfelt, 1986), due to the strict association of spindle effects with the anesthetic properties of hydrophobic chemicals (Ostergren, 1944; Hansch and Dunn, 1972; Onfelt et al., 1986b). More recently, a lin- ear relationship between lipophilicity and spindle disturbance (c-mitosis induction) was also found in V79 Chinese hamster cells with a series of n-alcohols and chlorinated aliphatics (Onfelt, 1987). At present, the general relevance of such non-specific hydrophobic interactions within the mechanisms that lead to chromosome misdistri- bution is unclear. The mitotic arrest produced by lipophilic chemicals may be mainly related to the general toxicity induced, and it is conceivable that in actively metabolizing systems the extent of metabolic conversion of the parent compounds may play a pivotal role. Furthermore, the high
131
concentrations required to exert an effect related to lipophilicity in ~'itrc may be not attainable in vivo.
In order to provide insight into this matter, we have analyzed the relationship between toxic and aneugenic effects induced by chlorinated hydro- carbons, and their physico-chemical features. The mold A. nidulans was used as the test organism, because of its ability to carry out the metabolic conversion of several promutagenic and procar- cinogenic compounds (Bignami et al., 1981).
QSAR analyses carried out with 35 chiori- nated aliphatic hydrocarbons also indicate that toxicity (i.e., the induction of lethality) is mainly related to steric factors, best described by MR. This agrees with other analyses undertaken with aliphatic compounds (Hellberg et al. 1985; ()nfelt et al., 1986) which demonstrated that chemical bulk, rather than electronic parameters, is an important determinant of toxic effects. Moreover, the occurrence of the MR term in the correlation equation (without a corresponding hydrophobic- ity term) suggests that the molecules are not contacting hydrophobic space of cellular macro- molecules. In such a case, it is generally assumed that polar space (oxygen and nitrogen on the surface of the macromolecules) is involved (Bla- ney and Hansch, 1990).
The ability to arrest conidial germination, which is synchronized with first mitoses (Bain- bridge et al., 1971), was also mainly related to the same steric factor. This end-point (the arrest of mitotic growth of germinating conidia) is not comparable to a c-mitotic effect, where cells are blocked at a precise phase of mitosis (metaphase). Rather, it reflects the general toxicity of the compound, and it is not expected to be specifi- cally related to a spindle effect. Accordingly, closely related chemicals such as 1,1,2- and 1,1,1- trichloroethane displayed identical arresting ac- tivities, but opposite aneuploidizing properties (see Table 5).
As far as the aneugenic activity is concerned, the QSAR analysis carried out in this study points to the prevailing role of electronic parameters. In particular, the ease in nccepting electrons (parametrized by LUMO and DENS) enters into the discriminant equations, together with MR. This eleetrophilic character is probably inade-

132
quate for a significant direct interaction with DNA. In fact, none of the compounds studied were able to induce mitotic crossing-over, as evi- denced by the induction of twin spots in morpho- logically normal colonies (data not shown). How- ever, it is an important factor determining the relative propensity to undergo reductive metab- olism by an 'electron capture' mechanism medi- ated by cytochromes (Hanzlik, 1981). Even though the direct interference of these chlorinated com- pounds with microtubule polymerization (as sug- gested for aprotic polar solvents by Gr6schel- Stewart et al., 1985) cannot be ruled out, the induction of radical damages provides an attrac- tive hypothesis for the mechanism of action of haloalkanes. Spindle functioning could be af- fected by free radicals generated by the reductive metabolism of haloalkanes through multiple mechanisms: lipid peroxidation of membranes, reaction with critical sulfhydryl groups (e.g., on microtubules and C a 2+ ATPase), alteration of the redox state of the cell, depletion of reduced glutathione (Onfelt, 1986). In particular, damage to the complex system of membranous structures surrounding the spindle (Paweletz and Schroeter, 1987) will lead to impaired control of calcium homeostasis and alterations of the fine regulation of the polymerization of microtubules (Hepler et al., 1984; Paweletz and Fehst, 1984), resulting in abnormal chromosome segregation.
According to the hypothesized role of reduc- tire metabolism, preliminary experiments with te- trachloroethanes demonstrate an increased activ- ity in assays carried out in the presence of lower oxygen concentration, which can shift metabolic conversion of haloalkanes toward reductive path- ways (Hanzlik, 1981; Macdonald, 1982) and a protective effect of radical scavengers (data not shown). Furthermore, the involvement of free radicals in the mechanism of chromosome malsegregation induced by CCI 4 in A. nidulans was previously suggested on the basis of the pro- tective effect exerted by cysteamine (Gualandi, 1984).
Further investigation of the substituent effect in simple haloalkanes (Slater and Sawyer, 1971) and of the quantitative relationship between in- duced radical damage and chromosome maiseg- regation in A. nidulans is under way in order to
verify the indication provided by this first explo- ration of the QSARs in the (geno)toxicity of haloaikanes.
In conclusion, QSAR analysis indicates that the basic determinants of toxic effects and chro- mosome malsegregation induced by a series of chlorinated aliphatic hydrocarbons in A. nidulans are different, thus confirming that in our data set aneuploidy is a 'unique' genetic effect, other than general toxicity. The clues provided for a possible role of reductive metabolism, with the production of free radicals, are worth considering. Free radi- cal damage is known to affect the integrity of the cytoskeleton (Bellomo et al., 1990a,b), and it could conceivably lead to a wrong chromosome distri- bution through impaired spindle functioning.
Acknowledgements
The technical assistance of Mr. M.C. D'Ascoli, B. Crochi and L. Gargano is gratefully acknowl- edged. This work was partially supported by EEC Contract No. EV4V-0044-1(a).
References
Albertini, S., U. Friederieh, C. Holderegger and F.E. Wiirgler (1988) 3"he in vitro porcine brain tubulin assembly assay: effects of a genotoxie carcinogen (aflatoxin B,), eight [t~mor promoters and nine miscellaneous substances, Mu- tation Res., 201, 183-292.
Andrews, P.R., DJ. Craik and J.L. Martin (1984) Functional group contributions to drug-receptor interactions, J. Med. Chem., 27, 1684-1657.
Bainbridge, B.W. (1971) Macromolecular composition and nuclear division during spore germination in Aslre~illus nidulans, J. Gen. Microbiol., 66, 319-325.
Barrett, J.C, H. Osbimura, N. Tamaka and I". Tsutsui (1985) Role of aneuploidy in early and late stages of neoplastic progression of Syrian hamster embuo cells in culture, in: V.L. Dellarco, P.E. Woytek and A. Hollaender (Eds.), Aneupluidy: Etiology and Mechanisms, Plenum. New York, pp. 523-538.
Bellomo, G., F. Mirabelli, M. Vairetti, F. losi and W. Malorni (1990a) Cytoskeleton as a target of menadione induced oxidative stress in cultured mammalian cells. !. Biochemi- cal and immuno~tochemical features, J. Cell Physiol., 143, ! 18-128.
Bellomo, G., F. Mirabelli, P. Richelmi, W. Malorni, F. losi and S. Orrenius (1990b)Cytoskeleton as a target in quinone toxicity, Free Radical Res. Commun., 8, 391-399.

Benigni, R., C. Andreoli and A. Giuliani (1989) Quantitative structure-activity relationships: principles, and applica- tions to mutagenicity and carcinogenicity, Mutation Res., 221, 197-216.
Bignami, M., F. Aulicino, A. Velcich, A. Carere and G. Morpurgo (1977) Mutagenic and recombinogenic action of pesticides in Aspergillus nidulans, Mutation Res., 46, 395- 402.
Bignami, M., G. Conti, R. Crebelli and A. Carere (1981) Growth-mediated metabolic activation of promutagens in Aspergillus nidulans, Mutation Res., 80, 265-272.
Blaney, J.M., and C. Hansch (1990) Application of molecular graphics to the analysis of macromolecular structures, in: C. Hansch (Ed.), Comprehensive Medicinal Chemistry, Vol. IV, Pergamon, Oxford, pp. 459-496.
Brunner, M., S. Albertini and F.E. Wiirgler (1991) Effects of 10 known or suspected spindle poisons in the in vitro porcine brain tubulin assembly assay, Mutagenesis, 6, 65- 70.
Cavenee, W.B., A. Koufos and M.F. Hansen (1986) Recessive mutant genes predisposing to human cancer, Mutation Res., 168, 3-14.
Crebelli, R., and A. Carere (1987) Chemical and physical agents assayed in tests for mitotic intergenic and intra- genie recombination in Aspergillus nidulans diploid strains, Mutagenesis, 2, 469-475.
Crebelli, R., G. Conti, L. Conti and A, Carere (1984) Induc- tion of somatic segregation by halogenated aliphatic hy- drocarbons in Aspergillus nidulans, Mutation Res., 138, 33-38.
Crebelli, R., G. Conti, L. Conti and A. Carere (1985) Muta- genicity of trichloroethylene, trichloroethanol and chloral hydrate in Aspergillus nidulans, Mutation Res., 155, 105- II1.
Crebelli, R., R. Benigni, J. Franekic, G. Conti, L. Conti and A. Carere (1988) Induction of chromosome malsegregation by halogenated organic solvents in Asper#llus tlidulans: unspecific or specific mechanism?, Mutation Res., 201, 401-411.
Dellarco, V.L., K.H. Mavournin and M.D. Waters (1986) Aneuploidy Data Review Committee: summary compila- tion of chemical data base and evaluation of test method- ologies, Mutation Res., 167, 149-169.
Dixon, W.J. (Ed.), (1981) BMDP Statistical Software, Univer- sity of California Press, Berkeley, CA.
Dustin, P. (1984) Microtubules, Springer, Berlin. Evans, H.J. (1985) Neoplasia and cytogenetic abnormalities,
in: V.L. Dellarco, P.E. Woytek and A. Hollaender (Eds.), Aneuploidy: Etiology and Mechanisms, Plenum, New York, pp. 165-178.
Frierson, M.R., G. Klopman and H,S. Rosenkranz (1986) Structure-activity relationships (SARs) among mutagens and carcinogens: a review, Environ. Mutagen., 8, 283-327.
Gr6escheI-Stewart, U., V.W. Mayer, R.E. Taylor-Mayer and F.K. Zimmermann (1985) Aprotic polar solvents inducing chromosomal malsegregation in yeast interfere with the assembly of porcine brain tubulin in vitro, Mutation Res., 149, 333-338.
133
Gualandi, G. (1984) Genotoxicity of the free-radical produc- ers CCI 4 and lipoperoxide in Aspergillus nidulans, Muta- tion Res., 136, 109-114.
Hansch, C., and W.J. Dunn Ill (1972) Linear relationships between lipophilic character and biological activity of drugs, J. Pharm. Sci., 61, 1-19.
Hanzlik, R.P. (1981) Reactivity and toxicity among halo- genated methanes and related compounds. A physico- chemical correlate with predictive value, Biochem. Phar- macol., 30, 3027-3030.
Hecht, F., and B.K. Hecht (1987) Aneuploidy in humans: dimensions, demography, and danger of abnormal num- bers of chromosomes, in: B.IC Vig and A.A, Sandberg (Eds.), Aneuploidy, Part A: Incidence and Etiology, Liss, New York,
Hellberg, S., S. Wold, W.J. Dunn !I!, J. Gasteiger and M.G. Hutchings (1985) The anesthetic activity and toxicity of halogenated ethyl methyl ethers, a multivariate QSAR modelled by PLS, Quant. Struct. Act. Relat., 4, 1-11.
Hepler, P.K., and S.M. Wolniak (1984) Membranes in the mitotic apparatus: their structure and function, Int. Rev. Cytol., 90, 169-238.
Hook, E.B. (1985) The impact of aneuploidy upon public health: mortality and morbidity associated with human chromosome abnormalities, in: V.L. Dellarco, P.E. Woytek and A. Hollaender (Eds.), Aneuploidy: Etiology and Mechanisms, Plenum, New York, pp. 7-33.
Hopward, D.A., and G. Sermonti (1962) Genetics of Strepto- myces coelicolor, Adv. Genet., 11,273-342.
Hyams, J.S., and B.R. Brinkley (Eds.) (1989) Mitosis: Molecules and Mechanisms, Academic Press, london.
K~ifer, E. (1961)The processes of spontaneous recombination in vegetative nuclei of Aspergillus nidulans, Genetics, 46, 1581-1609.
Kiifer. E. (1986)Tests which distinguish induced crossing-over and aneuploidy from secondary segregation in Aspergillus treated with chloral hydrate and ~-rays, Mutation Res., 164, 145-166.
Kifer, E., and A. Upshall (1973) The phenotypes of the eight disomics and trisomics in Aspergillus ,idulans, J, Hcred., 64, 35-38.
K.:ifer, E, B,R. Scott and A. Kappas (1986) Systems and results of tests for chemical induction of mitotic segrega- tion in Aspergillus nidulans, Mutation Res., 167, 9-34.
Lacey, E., and T.R. Watson (1985) Structure-activity relation- ships of benzimidazole carbamates as inhibitors of mam- malian tubulin in vitro, Biochem. Pharmacol., 34, 1073- 1077.
Lebart, L., A. Morineau and K.M. Warwick (1984) Multivari- ate Descriptive Statistical Analysis, Wiley, New York.
Liang, J.C., and B.R. Brinkley (1985) Chemical probes and possible targets for the induction of aneuploidy, in: V.L. Dellarco, P.E. Woytek and A. Hollaender (Eds.), Aneu- ploidy: Etiology and Mechanisms, Plenum, New York, pp. 491-505.
Lopez de Compadre, R.L., A. Kumar Debnath, A.J. Shuster- man and C. Hansch (1990) LUMO energies and hydropho- bicity as determinants of mutagenicity by nitroaromatic

134
compounds in Salmonella o'phimurium, Environ. Mol. Mu- tagen., 15, 44-55.
Lyman, W.J., W.F. Reehl and Rosenblatt, D.H. 11982) Hand- book of Chemical Property Estimation Methods, McGraw- Hill, New York.
Macdonald, T.L. 11982) Chemical mechanism of halocarbon metabolism, CRC Crit. Rev. Toxicol, 11, 85-119.
Mann, R.C., D.M. Popp and RA. Popp 11985) Critical sample size for determining the statistical significance of mutation frequencies, Mutation Res., 143, 93-100.
Morpurgo, G., D. Bellincampi, G. Gualandi, L. Baldineili and O. Serlupi Crescenzi 11979) Analysis of mitotic non-dis- junction with Aspergillus nidulans, Environ. Health Per- spect., 31, 81-95.
Moustacchi, E., A. Carere, G. Morpurgo, C. Ramel and F.E. Wiirgler 11986) Assays for genetic changes in fungi, in: R. Montesano, H. Bartsch, H. Vainio, J. Wiibourn and H. Yamasaki (Eds.), Long-term and Short-term Assays for Carcinogens: A Critical Appraisal, IARC Sci. Publ. No. 83, International Agency for Research on Cancer, Lyon, pp. 303-349.
Onfelt, A. 11986) Mechanistic aspects on chemical induction of spindle disturbances and abnormal chromosome num- bers, Mutation Res., 168, 269-300.
Onfelt, A. (1987) Spindle disturbance in mammalian cells. !11. Toxocity, c-mitosis and aneuploidy with 22 different com- pounds. Specific and unspecific mechanisms, Mutation Res., 182, 135-154.
Onfelt, A., S. Hellber8 and S. Wold 11986) Relationships between induction of anesthesia and mitotic spindle dis- turbances studied by means of principal components anal- ysis, Mutation Res., 174, 109-113.
Ostergren, G. (1944) Colchicine mitosis, chromosome contrac- tion, narcosis and protein chain folding, Hereditas, 30, 429-46%
Ostergren, G,, and A. Levan (1943) The connection between c-mitotic activity and water solubility in some monocyclic compounds, Hereditas, 29, 496-498,
Paweletz, N., and M, Fehst (1984) The vesicular compartment of the mitotic apparatus in mammalian ceils, Cell Biol, lnt. Rep., 8, 675-688.
Paweletz, N., and D. Schroeter 11987) On the ultrastructure of the mitotic apparatus, in: B.K. Vig and A.A. Sandberg (Eds.), Aneuploidy, Part A: Incidence and Etiology, Liss, New York, pp. 341-393.
Sakai, H., H. Mohri and G.G. Borisy (Eds.) 11982) Biological Functions of Microtubules and Related Structures, Aca- demic Press, New York.
Sankaranarayanan, K. (1979) The role of non-disjunction in anenploidy in man: an overview, Mutation Res., 61, 1-28.
Shanfield, B., and E. K~ifer (1971) Chemical induction of mitotic recombination in Aspergillus nidulans, Genetics, 67, 209-219.
Shusterman, A.J., A. Kumar Debnath, C. Hansch, G.W. Horn, F.R. Fronczek, A.C. Greene and S.F. Watkins 11989) Mutagenicity of dimethyl heteroaromatic triazenes in the Ames test: the role of hydrophobicity and electronic ef- fects, Mol. Pharmacol., 36, 939-944.
Slater, T.F., and B.C. Sawyer 11971) The stimulatory effects of carbon tetrachloride and other halogeno-alkanes on perox- idative reactions in rat liver fractions in vitro, Biochem. J., 123, 805-814.
Topliss, J.G., and R.P. Edwards 11979) Change factors in studies of quantitative structure-activity relationships, J. Mud. Chem., 22, 1238-1244.
Wallin, M., B. Frid6n and M. Billger 11988) Studies of the interaction of chemicals with microtubule assembly in vitro can be used as an assay for detection of cytotoxic chemi- cals and possible inducers of aneuploidy, Mutation Res., 201,303-311.
Wood, S., and E. Kiifer 11969) Effects of ultraviolet irradia- tion on heterozygous diploids of Aspergillus nidulans, i. UV-induced mitotic crossing over, Genetics, 62, 507-518.
Zimmermann, F.K., U, Gr6escheI-Stewart, I. Scheel and M,A. Resnick (1985) Genetic change may be caused by interfer- ence with protein-protein interactions, Mutation Res., 150, 203-210.
Zimmermann, F,K., J.H, Henning, 1. Scheel and M. Oehler 11986) Genetic and anti-tubulin effects induced by pyri- dine derivatives, Mutation Res., 163, 23-31.





























