Embed Size (px)
Citation preview

TB
V
a
b
R
A
fTWfm(sg©
K
1
cHsoilcdt
C(
0d
Zoologischer Anzeiger 250 (2011) 246–257
he male genitalia in the subfamily Agathidinae (Hymenoptera:raconidae): Morphological information of species on generic level
ladimir Zikica,∗, Cees van Achterbergb, Sasa S. Stankovica, Marijana Ilica
Faculty of Sciences, Department of Biology and Ecology, University of Nis, ul. Visegradska 33, 18000 Nis, SerbiaDepartment of Terrestrial Zoology, Netherlands Centre for Biodiversity Naturalis, Postbus 9517, 2300 RA Leiden, Netherlands
eceived 21 January 2011; received in revised form 19 April 2011; accepted 21 April 2011
bstract
The morphology of the male genitalia of 46 Agathidinae species belonging to 20 genera has been investigated. Samplesrom various geographical regions were used to discover the evolution of the male genitalia within the subfamily Agathidinae.hose male genitalia may contain important phylogenetic information at least in certain groups of Braconidae. For the Neworld, three of four investigated species of Cremnops (C. haematodes, C. montrealensis and C. vulgaris) could suggest the
orm of the new genus with the other members of New World Cremnops, partly because of the deviating morphology of theale genitalia. Comparative morphology of the male genitalia of Bassus confirms that it is polyphyletic. The status of Lytopylus
L. erythrogaster) is again actualised. Once synonymised as Aerophilopsis, this species could be excluded from other Lytopylus
pecies. The statistical analysis showed that smaller specimens (<6.67 mm) give less details on the morphology of the maleenitalia comparing to larger.2011 Elsevier GmbH. All rights reserved.
Cremn
tmom(tmit
eywords: Braconidae; Agathidinae; Morphology; Male genitalia;
. Introduction
The Braconidae belong to the parasitoid group, whichomprises about 60% of the known 145,000 species ofymenoptera (Huber, 2009), but taking in account the unde-
cribed species found in surveys this will be at least 80%r more. Biology of Agathidinae is very diverse, depend-ng on the species. They are koinobiont endoparasitoids ofepidopteran larvae, attacking all larval instars exposed or
oncealed. Also, may be gregarious or solitary, nocturnal oriurnal, very variable in size (Sharkey et al., 2009). Withinhe parasitoid group the male genitalia are hardly used both at∗Corresponding author. Tel.: +381 63415426; fax: +381 18533015.E-mail addresses: [email protected] (V. Zikic),
[email protected], [email protected]. van Achterberg).
Rh(aBB(f
044-5231/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.jcz.2011.04.006
ops
he species level and above this level. Outside Hymenoptera,ale genitalia are widely used for the higher level taxonomy
f insects (e.g. Snodgrass, 1957) who revised the externalorphology of the male genitalia of insects or Kristensen
1998). Among the Hymenoptera to some degree male geni-alia are used for the higher phylogeny of some groups. The
ost important starting point in the study of the male gen-talia of Hymenoptera is given by Snodgrass (1941), afterhat several hymenopterologists investigated this group (e.g.asnitsyn, 1988) and mainly in the Aculeate families for theigher phylogeny (e.g. Brothers, 1975) or within familiese.g. Pompilidae (Evans, 1950), Thynnidae (van Achterbergnd van Harten, 2009) and Bethylidae (Evans, 1978)). Withinraconidae a few examples at species level are known, e.g.
elokobylskij (1987, 1992, 1993) and Belokobylskij et al.2004) give the information about male genitalia of two sub-amilies: Doryctinae and Exothecinae concentrating on the

er Anze
mto(ctLnSM(gdoAa(t(Mtipiohofi
tpta
etbtelttibt
2
ontoa
iMdalZ
oc1(iTmdtcvMw2(mltcm
3
itmH1at1pDegli(p111Z
V. Zikic et al. / Zoologisch
ain evolutionary transformations of their structures andheir phylogenetic meaning; Walker (1994) for two speciesf Apanteles Foerster, 1862 (Microgastrinae); Haeselbarth1973) and van Achterberg (1980) for a few species of Bla-us Nees, 1819 (Blacinae). At the genus or the subgenus levelhere have been some attempts, some with less result (e.g.oan (1974) for Peristenus and related genera in the Euphori-ae) than others (e.g. Mason (1981); males of members ofathon Mason, 1981 (a subgenus of the genus Apanteles;icrogastrinae) are supposed to have enlarged parameres
claspers) compared to other genera or subgenera of Micro-astrinae. Tobias (1967, 1968) was the first who indicatedifferences of male genitalia between several subfamiliesf Braconidae. van Achterberg (1984) and Quicke and vanchterberg (1990) used some male genitalia traits to char-
cterise some higher groups in the Braconidae and Quicke1988) gives some generalised differences of male geni-alia in genera groups of the subfamily Braconinae. Maetô1987) gives a comparative study of braconid male genitalia;
aetô (1996) he analyzes the inter-generic variation amonghe species of the subfamily Microgastrinae. An extensivelyllustrated summary of the male genitalia in Braconidae wasublished in Serbian by Brajkovic (1989), along with thellustrations of mouth parts and the female genitalia. It isbvious that small parasitoids (less than 5 mm length of body)ave reduced male genitalia in general what makes the studyf male genitalia less rewarding. This observation comesrom the personal experience of authors working on Aphidi-nae (Aphidius) and Alysiinae (Chorebus) (unpublished data).
For answering the question if male genitalia are of impor-ance for the phylogeny of genera in the Braconidae it is morerofitable (and also more practical) to look for the groups con-aining specimens exceeding 8–10 mm as e.g., Braconinaend Agathidinae.
The present account deals with the morphological differ-nces occurring among species of the subfamily Agathidinaehroughout the world given that the overall knowledge of theasic group of Braconidae (Helconoid group, together withhe Sigalphinae; Quicke and van Achterberg, 1990; Quicket al., 1999) is limited. In these two groups male genitalia areess reduced and/or specialised than more derived groups inhe Braconidae, as e.g. Euphorinae or Alysiinae. Important ishe presence of a phylogeny of the subfamily; for the Agath-dinae a recent cladogram (Sharkey et al., 2006) is availableased on molecular and morphological data (but no data onhe male genitalia).
. Material and methods
For this study we used original material of male genitaliaf 46 taxa belonging to 20 genera of the subfamily Agathidi-
ae from various countries of the world (Table 1). Especiallyropical taxa with few species are under sampled, despiteur efforts to include as much as possible. The specimensre collected by sweeping and by Malaise trap. The spec-AiHo
iger 250 (2011) 246–257 247
mens from the USA are deposited at the Natural Historyuseum, Washington, DC, USA; specimens from Serbia are
eposited at the Faculty of Sciences, Department of Biologynd Ecology, University of Nis, Serbia. The rest of the ana-yzed material is deposited at the Department of Terrestrialoology, Netherlands Centre for Biodiversity Naturalis.Genital apparatus were pulled out from the metasoma
f specimens using the fine forceps and needles. Afterleaning surrounding tissue, male genitalia were boiled in0% KOH. Dissection is conducted using stereo microscopeBresser Science ETD 101 7–45x Microscoperesser) and gen-tal structures were mounted in Canada balsam (Stary, 1970).he structures were studied using a DM LS phase-contrasticroscope (Leica, Wetzlar, Germany), and after that some
rawings were drawn. For the statistical analyses, we usedhe Statistica 6 software package (StatSoft, 2001) examinedharacters are given in the Table 2. We performed the multi-ariate analysis of variance (ANOVA) on selected characters.ean body lengths (mbl) and mean forewing lengths (mfwl)ere given from the Taxapad software package (Yu et al.,005). The rest of measurements such is aedeagus lengthmbl/mal), the mean number of setae on cuspis (mcs), theean teeth on digitus (mdt), the size ratio between body
ength and fore wings length (mbl/mfwl) are calculated forhis purpose. Mean value for characters mcs and mdt is cal-ulated according to both vollselas. Scales under each figureeasure 0.1 mm.
. Results and discussion
Morphologically, the subfamily Agathidinae can be splitnto two major groups on the basis of the shape of the forearsal claws. Either the fore tarsal claws are simple or with aore or less lamelliform, submedial lobe (the tribe Agathidinialiday, 1833, and including the subtribe Earinina Sharkey,992; Fig. 57) or the fore tarsal claws are bifurcate (or “bifid”)nd with its inner tooth usually large (the tribe Cremnop-ini Sharkey, 1992 s.l. and including Disophrini Sharkey,992) the recognition of the latter makes the Cremnoptiniaraphyletic as shown by Sharkey et al., 2006). The use ofisophrini Sharkey (also spelt as Disophryini by Sharkey
t al., 2006) has to be abandoned. There is a minor thirdroup with a deep medio-posterior depression of the scutel-um and vein 2-CU of hind wing at least basally tubular,ncluding the very derived tribe Mesocoelini Viereck, 1918Sharkey et al., 2006). The tribe Cremnoptini as far as sam-led comprises the Old World genera Euagathis Szépligeti,900, Disophrys Foerster, 1862, Coccygidium de Saussure,892 s.l., Biroia Szépligeti, 1900 and Gyrochus Enderlein,920 and the New World genera Labagathis Enderlein, 1920,acremnops Sharkey and Wharton, 1985 and Zelomorpha
shmead, 1900. The genitalia are fairly homogeneous, hav-ng the dorsal inner corner of the parameres beak-shaped.owever, Zelomorpha, is aberrant by having this cornerf the parameres differently shaped (Fig. 17), compared to

248 V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257
Table 1. List of Agathidinae taxa with examined male genitalia.
No. Species Locality
1. Aerophilopsis erythrogaster (Viereck 1913) U.S.A. Coll. Natural History Museum, Washington, DC2. Agathirsia nigricauda (Viereck 1905) U.S.A. Coll. Natural History Museum, Washington, DC3. Agathirsia testacea (Muesebeck 1927) U.S.A. Coll. Natural History Museum, Washington, DC4. Agathis fuscipennis (Zetterstedt 1838) Serbia, Sicevo gorge, 15.08.1997, leg. V. Zikic5. Agathis gibbosa (Say 1838) U.S.A. Coll. Natural History Museum, Washington, DC6. Agathis montana (Shestakov 1932) Montenegro, Mt. Durmitor: Pitomine (1450 m a.s.l.), 27.06.1988, leg. A. Cetkovic7. Agathis nigra (Nees 1812) Serbia, Sicevo gorge, 15.08.1997, leg. V. Zikic8. Agathis umbellatarum (Nees 1812) Turkey, Istanbul, H. R. Th. v. Oorschot and H. Wiering. Yakuplu, 30 km W of Istanbul,
06.06.1979. Det. C. v. Achterberg, 19989. Alabagrus albispina (Cameron 1887) Suriname, Kwatta, 28.01.1964. DC Geijskes; Seacoast in Avicennia forest. Det M.
Sharkey10. Alabagrus erythromelas (Brullé 1846) Suriname, Lelydorp, Sumatra road, Sand ridges in savannah forest. 24–28. IV 1984. DC
Geijskes. (Museum Leiden). Det M. Sharkey11. Alabagrus levipodeum (Sharkey 1988) Colombia, Amazonas, Amacayacu Park, W Leticia, 3◦48′21′′S, 70◦18′23′′W,
01–04.10.1997, sweeping, M. Sharkey, RMNH 199812. Alabagrus maculipes (Cameron 1887) Mexico, Veracruz, Cordoba, 850 m, 05.05.1963, C and O Epping. Det M. Sharkey13. Alabagrus stigma (Brullé 1846) U.S.A. Coll. Natural History Museum, Washington, DC14. Alabagrus texanus (Cresson 1872) U.S.A. Coll. Natural History Museum, Washington, DC15. Biroia taeniocauda (Enderlein 1920) Indonesia, Samarinda, 11.05.1937, leg. Walsh. Conspecific with type of Isoptronotum
End. Det. C. v. Achterberg, 197316. Braunsia sp. (Australia) Australia, N.T. Kakadu N.P. Border store, Manngarra walk, 29.03.1991. leg C. v.
Achterberg, RMNH 199117. Camptothlipsis armenicus (Telenga 1955) Italy, Frali, Ronta de Casena, VII 2000, ex Anarsia lineatella D. Davide, RMNH 200018. Camptothlipsis sp. (India) India, Kerala, 10.12.1988. P. K. Sumodan, RMNH 199119. Coccygidium sp. (Senegal) Senegal, Rd. Toll. 18.07.1989. H. v. d. Valk, RMNH 1990. no 2 m20. Crassomicrodus medius (Cresson 1865) U.S.A. Coll. Natural History Museum, Washington, DC21. Crassomicrodus pallens (Cresson 1873) U.S.A. Coll. Natural History Museum, Washington, DC22. Cremnops desertor (Linnaeus 1758) Netherlands, 52◦19′N, 5◦07′E, Muiderberg, 19.06.1969. J B. Wolsehrija. leg. C. v.
Achterberg 197323. Cremnops haematodes (Brullé 1846) U.S.A. Coll. Natural History Museum, Washington, DC24. Cremnops montrealensis (Morrison 1953) U.S.A. Coll. Natural History Museum, Washington, DC25. Cremnops vulgaris (Cresson 1865) U.S.A. Coll. Natural History Museum, Washington, DC26. Disophrys initiator (Fonscolombe 1846) Serbia, Nis, 20.07.2005, leg. V. Zikic27. Disophrys sp. (Indonesia) Indonesia, N. Sumatra, Bukit Lawang, nr Bohorok river, c 200 m, rainforest 3–5. III
1994. C. v. Achterberg, RMNH 1994. Det. C. v. Achterberg 199428. Disophrys sp. (Thailand) Thailand, 50 m, Phuket Isl. S.; 7◦47′N, 98◦19′E; 01.08.1986, leg.R. Hensen29. Earinus elator (Fabricius 1804) Serbia, Sicevo gorge, 04.03.1997, leg. V. Zikic30. Euagathis abbotti (Ashmead 1900) Indonesia, Java, Museum Leiden, Tapos, 1936. leg. J. v. Vecht31. Euagathis chinensis (Holmgren 1868) Indonesia, Sumatra, Medan, Deli, Det. C. v. Achterberg, 199432. Euagathis forticarinata (Cameron 1899) Indonesia, Sumatra, Medan, Deli, Det. C. v. Achterberg, 199433. Euagathis punctata (Szépligeti 1902) Indonesia, Sulawesi, nr Luwuk, Bunga, c 300 m, 21–31.10.1989, Mal Trap 15, leg. C.v.
Achterberg, RMNH 198934. Gyrochus sp. (Indonesia) Indonesia, Coll. Betrem35. Labagathis rufoatra (Enderlein 1920) Suriname, Museum Leiden; Brokopondo distr., Brownsberg. 07.07.1975. P. H. van
Doesburg, 20, 21 (compared and conspecific with holotype) det. C. v. Achterberg 198636. Lytopylus rufipes (Nees 1812) Bulgaria, Coll. Zaykov, RHNM Leiden 1991, 01. VI 1976, Plovdiv, leg Zaykov. Det C.
v. Achterberg 199437. Therophilus agilis (Cresson 1873) U.S.A. Coll. Natural History Museum, Washington, DC38. Therophilus calcaratus (Cresson 1873) U.S.A. Coll. Natural History Museum, Washington, DC39. Therophilus festivus (Muesebeck 1953) U.S.A. Coll. Natural History Museum, Washington, DC40. Therophilus marshi (Bhat and Gupta 1977) Malaysia, SE Sabah, nr Danum Valley Field C. c. 150 m Wono, Mal. Trap 5,
26.05.1987–20.06.1987, RMNH 1987. leg. C. v. Achterberg, and D. Kennedy41. Therophilus perforator (Provancher 1880) U.S.A. Coll. Natural History Museum, Washington, DC42. Therophilus simillimus (Cresson 1873) U.S.A. Coll. Natural History Museum, Washington, DC43. Therophilus sp. (Trinidad) Trinidad, San Ferdinando, 04.05.1997, leg. R Someyer, B. v. Aartsen44. Therophilus tumidulus (Nees 1812) Bulgaria, Coll. Zaykov, 28.06.1986, Rhodopi, leg J. Kolarev. RHNM Leiden 199145. Zacremnops cressoni (Cameron 1887) U.S.A. Coll. Natural History Museum, Washington, DC46. Zelomorpha arizonensis (Ashmead 1900) U.S.A. Coll. Natural History Museum, Washington, DC
V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257 249
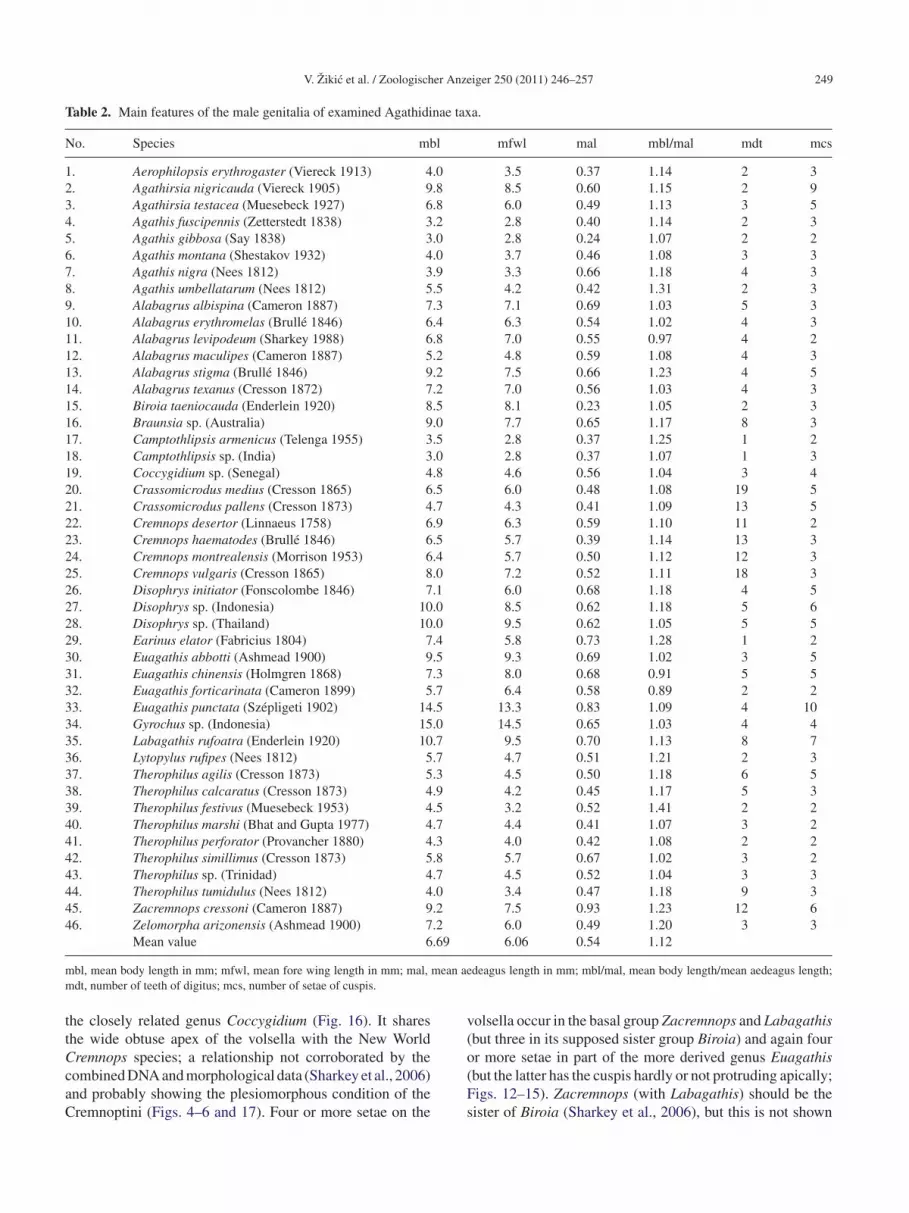
Table 2. Main features of the male genitalia of examined Agathidinae taxa.
No. Species mbl mfwl mal mbl/mal mdt mcs
1. Aerophilopsis erythrogaster (Viereck 1913) 4.0 3.5 0.37 1.14 2 32. Agathirsia nigricauda (Viereck 1905) 9.8 8.5 0.60 1.15 2 93. Agathirsia testacea (Muesebeck 1927) 6.8 6.0 0.49 1.13 3 54. Agathis fuscipennis (Zetterstedt 1838) 3.2 2.8 0.40 1.14 2 35. Agathis gibbosa (Say 1838) 3.0 2.8 0.24 1.07 2 26. Agathis montana (Shestakov 1932) 4.0 3.7 0.46 1.08 3 37. Agathis nigra (Nees 1812) 3.9 3.3 0.66 1.18 4 38. Agathis umbellatarum (Nees 1812) 5.5 4.2 0.42 1.31 2 39. Alabagrus albispina (Cameron 1887) 7.3 7.1 0.69 1.03 5 310. Alabagrus erythromelas (Brullé 1846) 6.4 6.3 0.54 1.02 4 311. Alabagrus levipodeum (Sharkey 1988) 6.8 7.0 0.55 0.97 4 212. Alabagrus maculipes (Cameron 1887) 5.2 4.8 0.59 1.08 4 313. Alabagrus stigma (Brullé 1846) 9.2 7.5 0.66 1.23 4 514. Alabagrus texanus (Cresson 1872) 7.2 7.0 0.56 1.03 4 315. Biroia taeniocauda (Enderlein 1920) 8.5 8.1 0.23 1.05 2 316. Braunsia sp. (Australia) 9.0 7.7 0.65 1.17 8 317. Camptothlipsis armenicus (Telenga 1955) 3.5 2.8 0.37 1.25 1 218. Camptothlipsis sp. (India) 3.0 2.8 0.37 1.07 1 319. Coccygidium sp. (Senegal) 4.8 4.6 0.56 1.04 3 420. Crassomicrodus medius (Cresson 1865) 6.5 6.0 0.48 1.08 19 521. Crassomicrodus pallens (Cresson 1873) 4.7 4.3 0.41 1.09 13 522. Cremnops desertor (Linnaeus 1758) 6.9 6.3 0.59 1.10 11 223. Cremnops haematodes (Brullé 1846) 6.5 5.7 0.39 1.14 13 324. Cremnops montrealensis (Morrison 1953) 6.4 5.7 0.50 1.12 12 325. Cremnops vulgaris (Cresson 1865) 8.0 7.2 0.52 1.11 18 326. Disophrys initiator (Fonscolombe 1846) 7.1 6.0 0.68 1.18 4 527. Disophrys sp. (Indonesia) 10.0 8.5 0.62 1.18 5 628. Disophrys sp. (Thailand) 10.0 9.5 0.62 1.05 5 529. Earinus elator (Fabricius 1804) 7.4 5.8 0.73 1.28 1 230. Euagathis abbotti (Ashmead 1900) 9.5 9.3 0.69 1.02 3 531. Euagathis chinensis (Holmgren 1868) 7.3 8.0 0.68 0.91 5 532. Euagathis forticarinata (Cameron 1899) 5.7 6.4 0.58 0.89 2 233. Euagathis punctata (Szépligeti 1902) 14.5 13.3 0.83 1.09 4 1034. Gyrochus sp. (Indonesia) 15.0 14.5 0.65 1.03 4 435. Labagathis rufoatra (Enderlein 1920) 10.7 9.5 0.70 1.13 8 736. Lytopylus rufipes (Nees 1812) 5.7 4.7 0.51 1.21 2 337. Therophilus agilis (Cresson 1873) 5.3 4.5 0.50 1.18 6 538. Therophilus calcaratus (Cresson 1873) 4.9 4.2 0.45 1.17 5 339. Therophilus festivus (Muesebeck 1953) 4.5 3.2 0.52 1.41 2 240. Therophilus marshi (Bhat and Gupta 1977) 4.7 4.4 0.41 1.07 3 241. Therophilus perforator (Provancher 1880) 4.3 4.0 0.42 1.08 2 242. Therophilus simillimus (Cresson 1873) 5.8 5.7 0.67 1.02 3 243. Therophilus sp. (Trinidad) 4.7 4.5 0.52 1.04 3 344. Therophilus tumidulus (Nees 1812) 4.0 3.4 0.47 1.18 9 345. Zacremnops cressoni (Cameron 1887) 9.2 7.5 0.93 1.23 12 646. Zelomorpha arizonensis (Ashmead 1900) 7.2 6.0 0.49 1.20 3 3
Mean value 6.69 6.06 0.54 1.12
m mean aem
ttCcaC
v(o
bl, mean body length in mm; mfwl, mean fore wing length in mm; mal,dt, number of teeth of digitus; mcs, number of setae of cuspis.
he closely related genus Coccygidium (Fig. 16). It shareshe wide obtuse apex of the volsella with the New Worldremnops species; a relationship not corroborated by the
ombined DNA and morphological data (Sharkey et al., 2006)nd probably showing the plesiomorphous condition of theremnoptini (Figs. 4–6 and 17). Four or more setae on the(Fs
deagus length in mm; mbl/mal, mean body length/mean aedeagus length;
olsella occur in the basal group Zacremnops and Labagathisbut three in its supposed sister group Biroia) and again fourr more setae in part of the more derived genus Euagathis
but the latter has the cuspis hardly or not protruding apically;igs. 12–15). Zacremnops (with Labagathis) should be theister of Biroia (Sharkey et al., 2006), but this is not shown
250 V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257
Figs. 1–9. Male genitalia of Agathidinae. 1, Aerophilopsis erythrogaster Viereck, 1913; 2, Agathirsia nigricauda (Viereck, 1905); 3, A.testacea Muesebeck, 1927; 4, Agathis fuscipennis (Zetterstedt, 1838); 5, A. gibbosa (Say, 1836); 6, A. montana Shestakov, 1932; 7, A. nigraN (Cam
bctogis
sTss
ees, 1812; 8, A. umbellatarum Nees, 1812; 9, Alabagrus albispina
y the morphology of the male genitalia (Figs. 2 and 3). Inonclusion, in the Cremnoptini the male genitalia supply lit-le additional support of the known all inclusive phylogenyf the group (Sharkey et al., 2006). Male genitalia obviously
ive support to the validity of the identification of the exam-ned species. Thus, analyses of morphology of the genitaltructures of deviant group such as the New World Cremnops,gCr
eron, 1887).
how additional differences among the rest of the species.hree species of Cremnops: C. haematodes, C. montrealen-is and C. vulgaris suggest to be separate as a new genus orubgenus versus Cremnops and Zelomorpha versus Coccy-
idium (Fig. 19). The latter were supposed to be synonyms byhou and Sharkey (1989), but they have been separated againecently by Sharkey et al. (2006). We suppose that further

V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257 251
Figs. 10–18. Male genitalia of Agathidinae. 10, Alabagrus erythromelas (Brullé, 1846); 11, A. levipodeum Sharkey, 1988; 12, A. maculipes( s (Cress Campt
msposgbfC
aCcCCm
Cameron, 1887); 13, A. stigma (Brullé, 1846); 14, Alabagrus texanup. (Australia); 17, Camptothlipsis armenicus (Telenga, 1955); 18,
orphological and molecular methods will give the definitivetatus of this very variable genus. In this analysis we pro-ose the new genus, Neocremnops for the New World speciesf Cremnops (Type species: Bracon montrealensis Morri-on, 1917. Etymology: from “neo” (Latin for “new”) and theeneric name Cremnops Foerster, 1862, because all its mem-
ers occur in the New World (gender: masculine), includingollowing species: Cremnops cameronii (Dalla Torre, 1898);. caribensis (Berta, 1998); C. apicalipennis Berta, 1998; C.1B1
son, 1872); 15, Biroia taeniocauda (Enderlein, 1920); 16, Braunsiaothlipsis sp. (India).
shmeadi Morrison, 1917; C. californicus Morrison, 1917;. caribensis Berta, 1998; C. comstocki Morrison, 1917; C.rassifemur Muesebeck, 1927; C. guanicanus Wolcott, 1924;. haematodes Brullé, 1846; C. kelloggii Morrison, 1917;. marshi Berta, 1998; C. melanoptera Ashmead, 1894; C.ontrealensis Morrison, 1917; C. nigrosternum (Morrison,
917); C. pectoralis (Ashmead, 1894); C. plesiopectoraliserta, 1998; C. sharkei Berta, 1998; C. shenefelti Marsh,961; C. slossonae Morrison, 1917; C. tibiomaculatus Berta,
252 V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257
Figs. 19–27. Male genitalia of Agathidinae. 19, Coccygidium sp. (Senegal); 20, Crassomicrodus medius (Cresson, 1865); 21, C. pallens( aematov 46); 2
1sBm(vnoo
oFlrrl
Cresson, 1873); 22, Cremnops desertor (Linnaeus, 1758); 23, C. hulgaris (Cresson, 1865); 26, Disophrys initiator (Fonscolombe, 18
998; C. virginiensis Morrison, 1917; C. vulgaris Cres-on, 1865; C. washingtonensis Shenefelt, 1937; C. willinkierta, 1998, and C. yucatanus Berta, 1998, because all itsembers occur in the New World, volsella with 3 setae
Figs. 4–6) vs. C. desertor (volsella with 2 setae (Fig. 7);olsella wide (Figs. 4–6) vs. C. desertor (volsella moderately
arrow (Fig. 7); lateral crests of frons reaching posterior ocellir nearly so (Fig. 48; remain distinctly removed: Fig. 53);uter hind tarsal claw similar to inner claw (Figs. 49 and 50;cww
des (Brullé, 1846); 24, C. montrealensis (Morrison, 1917); 25, C.7, Disophrys sp. (Indonesia).
uter hind claw distinctly differentiated from inner claw:igs. 54 and 55); scapus gradually bent (Fig. 51; angu-
arly bent: Fig. 56); pair of crests between antennal socketsemain separated posteriorly or reduced (Fig. 48; united oreduced: Fig. 53); wings largely darkened (banded, rarelyargely darkened); precoxal sulcus present posteriorly and
renulate, rarely with smooth pit or absent (absent, rarelyith smooth groove); subpronope elliptical (Fig. 52), butide and more or less triangular in N. cameronii (Dalla Torre)
V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257 253
F Thailan( rinatas lus ru
aEBccvo
i(
Fdrctop
igs. 28–36. Male genitalia of Agathidinae. 28, Disophrys sp. (Ashmead, 1900); 31, E. chinensis (Holmgren, 1868); 32, E. forticap. (Indonesia); 35, Labagathis rufoatra Enderlein 1920; 36, Lytopy
nd N. caribensis (Berta) (wide oval or triangular: Fig. 57).xcluded are C. ferrugineus Cameron, 1887; C. punctatuserta, 1998 and C. turrialbae Berta, 1998, because of thearinate hind trochantellus and C. boliviensis Berta, 1988; C.ubensis Cresson, 1865; C. misionensis Berta, 1988, and C.iolaceipennis Cameron, 1887, because of the aberrant shapef the pronotum and the protruding stemmaticum.
The variation of the male genitalia within the Agathidinis more extensive and the phylogeny of the tribe is less clearSharkey et al., 2006), mainly because of the genus Bassus
o(s
d); 29, Earinus elator (Fabricius, 1804). 30, Euagathis abbotti(Cameron, 1899); 33, E. punctata Szépligeti, 1902; 34, Gyrochus
fipes (Nees, 1812) [= sculptured Bassus p.p.].
abricius, 1804, s.l. wherein usually all groups are placedifficult to separate from each other. The subtribe Earinina isepresented in this study by the New World genera Crassomi-rodus Ashmead, 1900, and Agathirsia Westwood, 1882, andhe wide-spread genus Earinus Wesmael, 1837. Earinus hasnly two setae on the volsella, the dorsal inner corners ofarameres are comparatively narrow and the volsella with-
ut distinct cuspis (Fig. 18). Both other genera have similarand likely less derived) male genitalia with four or moreetae, wide and truncate inner corners of the parameres
254 V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257
Figs. 37–45. Male genitalia of Agathidinae. 37 Therophilus agilis (Cresson, 1873); 38, T. calcarata (Cresson, 1873); 39, T. festivus (Mue-s us (C( i (Cam
atssWa(t
BActw
ebeck, 1953); 40, T. perforator (Provancher, 1880); 41, T. simillimMalaysia); 44, Therophilus sp. (Trinidad); 45, Zacremnops cresson
nd a wide and truncate cuspis (Figs. 19–22). The sub-ribe Agathidina is represented by the polyphyletic and widepread Bassus-complex (including the genera Camptothlip-is Enderlein, 1920, Bassus Fabricius, 1804 s.s., Therophilusesmael, 1837 and Lytopylus Foerster, 1862), the large
nd varied New World genus Alabagrus Enderlein, 1920Figs. 23–28), and some smaller genera: the mainly Holarc-ic genus Agathis Latreille, 1805, and the wide-spread genus
w
t
resson, 1873); 42, T. tumidulus (Nees, 1812); 43, Therophilus sp.eron, 1887).
raunsia Kriechbaumer, 1894. At the base of the subtribegathidina (Sharkey et al., 2006) are the species of Bassus-
omplex with a largely smooth third and second metasomalergites (Bassus Fabricius s.s. and Therophilus Wesmael) orith the second tergite granulated combined with a reduced
ing venation (Camptothlipsis).The last attempts to solve the phylogenetic relations amonghe species of the genus Bassus (Sharkey et al., 2006) con-

V. Zikic et al. / Zoologischer Anzeiger 250 (2011) 246–257 255
Figs. 46 and 57. Male genitalia of Agathidinae. 46, Zelomorpha arizonensis Ashmead, 1900; 47, Sigalphus irrorator (Fabricius, 1775) (afterTobias, 1968; modified). 48–52, Cremnops montrealensis (Morrison, 1917), female, USA, Ann Arbor. 53–57, Cremnops desertor (Linnaeus,1 , 54, ol 1×); 4
dtdtpmc(l(iiAfn
co
iilit(o(fcmotal
758), female, Netherlands, Waarder. 48, 53, head, dorsal aspect; 49ateral aspect; 52, 57, pronotum, dorsal aspect. 48, 53: scale-line (=
ucted by phylogenetic analyses based on morphology andhe D2–3 regions of 28S rDNA and Sharkey et al. (2009)id not correct this problem. Our support to clarify its posi-ion comparing the male genitalia, unfortunately leave theolyphyletic nature of the genus Bassus, overlapping withore derived genera such as Alabagrus, Agathis, Bassus-
omplex with striate third and second metasomal tergitesLytopylus) and Braunsia. One species of Agathis (A. umbel-atarum Nees, 1812), the species of Lytopylus FoersterLytopylus rufipes (Nees, 1812)) and Braunsia have the dorsalnner corners of the parameres distinctly protruding, resultingn a distinct concavity (Figs. 29, 34 and 35). Other species ofgathis have no concavity (Figs. 30–32) and one species has
our setae on the volsella (Fig. 33), contrary to the normalumber of two or three setae within this group.
Remains the heterogeneous remnant of the Bassus-omplex with more or less beak-shaped inner dorsal cornersf the parameres (Figs. 37–43), but the corners are wider
FSe
uter hind tarsal claw; 50, 43, inner hind tarsal claw; 51, 56, scapus,9, 50, 54, 55: 2.3×; 51, 56: 1.9×; 52, 57: 1.4×.
n the Therophilus sp. from Malaysia (Fig. 44) and narrown the Therophilus sp. from Trinidad (Fig. 45)/and may beargely absent (Figs. 36 and 46). The shape of the volsellas very variable and the number of setae on it is normallywo, but sometimes may be three (Figs. 45 and 46) or fiveFig. 38). The included species with the granulated sec-nd and third metasomal tergites (Camptothlipsis armenicusTelenga, 1955) and Therophilus agilis (Cresson, 1873)) dif-ers both in venation (the former has no second submarginalell of fore wing; the latter has one) and in the shape of theale genitalia. As one could expect, the sculpture of the sec-
nd tergite is not a reliable character to assign species ofhe Bassus-complex a genus rank and the male genitalia givedditional support to this. The male genitalia of Camptoth-ipsis armenicus are more derived (e.g. no distinct digitus:
ig. 36) than other members of the group. According toharkey et al. (2009) Aerophilopsis Viereck, 1913 is consid-red as a synonym of Lytopylus Forster, 1913. However based
2 er Anze
ol(oi
radabgCaossapaTaitio(ssammadnbnP
A
fwa
R
v
v
B
B
B
B
B
B
C
E
E
H
K
L
M
M
M
Q
56 V. Zikic et al. / Zoologisch
n our results Aerophilopsis could be excluded from Lytopy-us as separate genus because of the aberrant male genitaliaFigs. 35 and 46); the shape of the digitus is similar to somef the Therophilus species (Figs. 40–42) but it has differentnner corners of the parameres (Fig. 46).
In conclusion, also in the Agathidini the male genitaliaender some clues about the relationships within the groupbove species level, but the large supra-specific variationoes not allow more than a limited contribution. There isn interesting similarity between the male genitalia of theasal groups of the Cremnoptini (Zacremnops and Laba-athis: Figs. 1 and 2) and of Agathidini (Agathirsia andrassomicrodus: Figs. 19–22), together with those of theberrant (because of granulated second tergite and numberf setae on the volsella) Therophilus agilis (Fig. 38). It mayhow the plesiomorphic condition of the male genitalia in theubfamily Agathidinae with four or more setae on the volselland with a wide obtuse cuspis next to the digitus. In the sup-osed sister group Sigalphinae (Fig. 47) the cuspis is smallnd separated from the volsella and the volsella is slender. Inable 2, the Agathidinae with body size less than 6.69 mmnd the aedeagus not exceeding 0.54 mm are listed as small;n general, the number of setae on the cuspis and the teeth onhe digitus are reduced, so we concluded that size does mattern analysis of the details on the male genitalia in this groupf braconids. The results of multivariate analysis of varianceANOVA) based on the mean body length (mbl) shows thetatistically significance (F = 8.45; df = 36; P < 0.001). Theame is shown for the number of teeth on the digitus by thenalysis of one way ANOVA for the following characters:ean forewing length (mfwl) (F = 80.02; df = 9; P < 0.001),ean aedeagus length (mbl/mal) (F = 6.42; df = 9; P < 0.001)
nd the mean number of setae on cuspis (mcs) (F = 6.75;f = 9; P < 0.001). The univariate tests of significance for theumber of mean teeth on digitus (mdt) and the size ratioetween body length and fore wings length (mbl/mfwl) doot show valuable differences: for (mdt) (F = 1.92; df = 9;< 0.08), for (mbl/mfwl) (F = 0.37; df = 1; P < 0.543).
cknowledgements
We thank to Dr. Zeljko Tomanovic and Andjeljko Petrovicor the cooperation and technical support. This investigationas supported by the Grant 43001 (The Ministry of Education
nd Science of the Republic of Serbia).
eferences
an Achterberg, C., 1984. Essay on the phylogeny of Braconidae(Hymenoptera: Ichneumonoidea). Entomolosk Tidskrift 105,
41–57, Figs. 1–17.an Achterberg, C., van Harten, A., 2009. Order Hymenoptera,family Thynnidae. Genera Lamprowara Boni Bartalucci, IswaraWestwood and Komarowia Radoszkowski (Hymenoptera: Thyn-
Q
iger 250 (2011) 246–257
nidae: Myzininae: Meriini). Arthropod Fauna of the UAE 2,298–334, Figs. 1–145.
elokobylskij, S.A., 1987. The male genitalia of parasitic waspsof the subfamily Doryctinae (Hymenoptera, Braconidae), theirevolution and maining for classification. Proceedings of the All-Union Entomological Society 69, 209–219 (in Russian).
elokobylskij, S.A., 1992. On the classification and phylogeny ofthe braconide wasps of subfamilies Doryctinae and Exotheci-nae (Hymenoptera, Braconidae). Part I. On the classification 1.Entomologicheskoe Obozrenie 71 (4), 900–928 (in Russian).
elokobylskij, S.A., 1993. On the classification and phylogeny ofthe braconide wasps of subfamilies Doryctinae and Exotheci-nae (Hymenoptera, Braconidae). Part I. On the classification 2.Entomologicheskoe Obozrenie 72 (N), 143–164 (in Russian).
elokobylskij, S.A., Zaldivare-Riveron, A., Quicke, D., 2004. Phy-logeny of the genera of the parasitic wasps subfamily Doryctinae(Hymenoptera: Braconidae) based on morphological evidence.Zoological Journal of the Linnean Society 142, 369–404.
rajkovic, M.M., 1989. Parazitske ose. Braconidae, Hymenoptera.Uporedno-morfoloska studija usnih i genitalnih struktura, sis-tematika i filogenija. Naucna knjiga, Beograd, pp. 1–99, Figs.1–673.
rothers, D.J., 1975. Phylogeny and classification of the aculeateHymenoptera, with special reference to Mutillidae. Kansas Uni-versity Science Bulletin 50, 483–648, Figs. 1–101, 7 tables.
hou, L.-Y., Sharkey, M.J., 1989. The Braconidae (Hymenoptera)of Taiwan. I. Agathidinae. Journal of the Taiwan Museum 42,147–233, Figs. 1–309.
vans, H.E., 1950. A taxonomic study of the Nearctic spiderwasps belonging to the tribe Pompilini (Hymenoptera: Pompil-idae). Transactions of the American Entomological Society 75,133–270.
vans, H.E., 1978. The Bethylidae of America north of Mexico.Memoirs of the American Entomological Institute 27, 1–332.
uber, J.T., 2009. Biodiversity of hymenoptera. In: Foottit, R.G.,Adler, P.H. (Eds.), Insect Biodiversity, Science and Society.Wiley-Blackwell, pp. 303–323, i–xxi + 1–632.
ristensen, N.P., 1998. Lepidoptera, moths and butterflies. 1. Evo-lution, systematics, and biogeography. Handbuch der Zoologie4 (35), 1–487, ill.
oan, C.C., 1974. The European species of Leiophron Neesand Peristenus Foerster (Hymenoptera: Braconidae, Euphori-nae). Transactions of the Royal Entomological Society 126,207–238.
aetô, K., 1996. Inter-generic variation in the external malegenitalia of the subfamily Microgastrinae (Hymenoptera, Bra-conidae), with a reassessment of Mason’s tribal system. Journalof Hymenoptera Research 5, 38–52.
aetô, K., 1987. A comparative morphology of the male internalreproductive organs of the family Braconidae (Hymenoptera,Ichneumonoidea). Kontyû 55, 3242.
ason, W.R.M., 1981. The polyphyletic nature of ApantelesFoerster (Hymenoptera: Braconidae): a phylogeny and reclassifi-cation of Microgastrinae. Memoirs of the Entomological Societyof Canada 115, 1–147, Figs. 1–110.
uicke, D.L.J., van Achterberg, C., 1990. Phylogeny of the subfam-ilies of the family Braconidae (Hymenoptera: Ichneumonoidea).
Zoologische Verhandelingen Leiden 258, 1–95, Figs. 1–180.uicke, D.L.J., 1988. Inter-generic variation in the male genitalia ofthe Braconinae (Insecta, Hymenoptera, Braconidae). ZoologicaScripta 17, 399–409.

er Anze
Q
R
S
S
S
S
S
S
T
T
W
V. Zikic et al. / Zoologisch
uicke, D.L.J., Basibuyuk, H.H., Fitton, M.G., Rasnitsyn, A.P.,1999. Morphological, palaeontological and molecular aspectsof ichneumonoid phylogeny (Hymenoptera, Insecta). ZoologicaScripta 28, 175–202.
asnitsyn, A.P., 1988. An outline of evolution of the hymenopterousinsects (Order Vespida). Oriental Insects 22, 115–145, Figs. 1–4.
harkey, M.J., Laurenne, N.M., Sharanowski, B., Quicke, D.L.J.,Murray, D., 2006. Revision of the Agathidinae (Hymenoptera:Braconidae) with comparisons of static and dynamic alignments.Cladistics 22 (6), 546–567.
harkey, M.J., Yu, D.S., Noort, S. van, Seltmann, K., Penev,L., 2009. Revision of the Oriental genera of Agathidinae(Hymenoptera, Braconidae) with an emphasis on Thailandincluding interactive keys to genera published in three differentformats. ZooKeys 21, 19–54.
nodgrass, R.E., 1941. The male genitalia of Hymenoptera. Smith-
sonian Miscellaneous Collections 99, 1–86.nodgrass, R.E., 1957. A revised interpretation of external repro-ductive organs of male insects. Smithsonian MiscellaneousCollections 135, 1–60.
Y
iger 250 (2011) 246–257 257
tary, P., 1970. Biology of Aphid Parasites (Hymenoptera: Aphidi-idae) with Respect to Integrated Control. Series Entomologica.6. Dr. W. Junk, The Hague, the Netherlands.
tatSoft, 2001. Statistica (data Analysis Software System), Version6. Statsoft, Inc., Tulsa, OK, http://www.statsoft.com.
obias, V.I., 1967. A review of the classification, phylogenyand evolution of the family Braconidae (Hymenoptera). Ento-mologicheskoye Obozreniye 46 (3), 645–669 (EntomologicalReview 46(3), 387–399).
obias, V.I., 1968. On the classification and phylogeny of the fam-ily Braconidae. Special issue dedicated to the memory of H.A.Kholodkovskogo, Academy of Sciences of USSR, pp. 3–43,Figs. 1–4 (in Russian).
alker, A.K., 1994. Species of Microgastrinae (Hymenoptera: Bra-conidae) parasitizing lepidopterous cereal stem borers in Africa.Bulletin of Entomological Research 84, 421–434.
u, D.S., Achterberg, C.v., Horstmann, K., 2005. WorldIchneumonoidea 2004 – Taxonomy, biology, morphologyand distribution. CD-Rom, Taxapad, Vancouver, Canada,www.taxapad.com.





























