Embed Size (px)
Citation preview

Eur. J. Biochem. 243. 502-510 (1997) 0 FEBS 1997
The neuronal cell-adhesion molecule axonin- 1 is specifically released by an endogenous glycosylphosphatidylinositol-specific phospholipase Ruth LIERHEIMER', Beat KUNZ', Lorenz VOGT', Reto SAVOCA', Urs BRODBECK2 and Peter SONDEREGGER' ' Institute of Biochemistry, University of Zurich, Switzerland ' Institute of Biochemistry and Molecuiarbiology, University of Bern, Switzerland
(Received 21 October 1996) ~ EJB 96 1548/1
Axonin-I , a member of the immunoglobulidfibronectin type-I11 family of cell-adhesion molecules, occurs both as a glycosylphosphatidylinositol-(glycosylPtdIns)-anchored membrane-bound and a soluble form. In vivo observations show that the major part of axonin-1 is found in the soluble fraction and that soluble axonin-I perturbs neurite fasciculation and pathfinding in the developing chicken embryo. This has prompted further investigations into the mechanism of the axonin-I release. We demonstrate here that axonin-1 released from dorsal root ganglion neurons contains ethanolamine and inositol, components of the glycosylPtdIns anchor. Secreted axonin-1 does not exhibit the cross-reacting determinant epitope, an indication that the cleavage of the anchor is not mediated by a phosphatidylinositol-specific phospholi- pase C. Treatment of dorsal root ganglion neurons with 1,lO-phenanthroline, an inhibitor of glycosylPtd- Ins-specific phospholipase D, reduces the release of axonin-I by 56 %. Moreover, glycosylPtdIns-specific phospholipase D activity was detected in dorsal root ganglion neurons and brain. These results suggest that axonin-1 is released from the membrane by an endogenously expressed glycosylPtdIns-specific phos- pholipase D in viva With domain-swaping experiments between axonin-1 and its non-released relative F1 1, deletion mutants and monoclonal antibodies, we demonstrate that the fourth fibronectin type-111-like domain of axonin-I is required for the generation of the soluble form of axonin-1.
Keywords; axonin-1 ; domain localization; glycosylphosphatidylinositol ; glycosylphosphatidylinositol- specific phospholipase D ; neuronal cell-adhesion molecule.
During the development of neuronal networks, growth cones are guided to their targets by attractive and repulsive factors, and by guidance cues expressed on cellular surfaces or deposited in the extracellular matrix (Jessel, 1988; Hynes and Lander, 1992; Luo et al., 1993; Goodman and Shatz, 1993; Tessier-Lavi- gne, 1994 ; Keynes and Cook, 1995). Axon-associated cell-adhe- sion molecules (AxCAMs), concentrated primarily on axons and growth cones, are known to play a prominent role in pathfinding decisions influenced by the local environment. AxCAMs are di- vided in three major groups; cadherins (Takeichi et al., 1991 ; Ranscht et al., 1994), integrins (Reichardt and Tomaselli, 1991) and neural surface proteins of the immunoglobulin superfamily (Sonderegger and Rathjen, 1992; Brummendorf and Rathjen, 1993). One subgroup of the Ig superfamily consists of Fll(con- tactin)/F3, TAG-l/axonin-1 and PANGBIG-I (Briimmendorf et al., 1989; Ranscht, 1988; Gennarini et al., 1989; Furley et al., 1990; Zuellig et al., 1992; Connelly et al., 1994; Yoshihara et al., 1994). All members of this group are glycosylphosphatidyli- nositol(g1ycosylPtdIns)-anchored proteins with six amino-termi- nal Ig-like and four fibronectin type-111-(FNII1)-like domains
Correspondence to P. Sonderegger, Institute of Biochemistry, Uni-
Fax: +41 1 363 7947. Abbreviations. AxCAMs, axon-associated cell-adhesion molecules;
DRG, dorsal root ganglion, FNIII, fihronectin type 111; glycosylPtdIns- PLD, glycosylphosphatidylinositol-specific phospholipase D; mf-AchE, membrane form of acetylcholinesterase; NgCAM, neuron-glia cell-adhe- sion molecule; NrCAM, NgCAM-related cell-adhesion molecule; PtdTns-PLC, phosphatidylinositol phospholipase C, s-AchE, soluble ace- tylcholinesterase.
versity of Zurich, Winterthurerstr. 190, CH-8057 Zurich, Switzerland
(Brummendorf and Rathjen, 1993). Axonin-I was initially iden- tified as an axonally secreted protein from cultured chicken dor- sal root ganglia (DRG) neurons (Stoeckli et al., 1989) and puri- fied from vitreous fluid of the chicken embryo (Ruegg et al., 1989). Soluble axonin-1 adsorbed to cell-culture dishes pro- motes neuronal outgrowth and, when added to the medium of DRG neuron cultures, it perturbs the fasciculation of the neurites in vitro (Stoeckli et al., 1991). GlycosylPtdIns-anchored mem- brane-bound axonin-I is known to interact homophilically (Rader et al., 1993) and heterophilically with neuron-glia cell- adhesion molecule (NgCAM) and NgCAM-related cell adhesion molecule (NrCAM) (Kuhn et al., 1991; Suter et al., 1995). The axonin-l/NgCAM interaction is involved in fasciculation while the interaction of axonin-I with NrCAM is thought to be re- quired for accurate pathfinding in vivo. Injections of soluble axo- nin-I or anti-(axonin-1) serum into the spinal cord of the deve- loping chicken embryo result in defasciculation and pathfinding errors of the commissural axons. These axons fail to cross the midline and join instead the ipsilateral longitudinal axon tracts (Stoeckli and Landmesser, 1995).
The closest relative to axonin-I is F11, which is 53% iden- tical at the amino acid level. F11 also interacts heterophilically with NgCAM and NrCAM (Brummendorf et al., 1993; Morales et al., 1993). Despite these structural and functional similarities, the majority of axonin-1 is secreted, while most of F11 remains on the cell surface in vivo (Stoeckli et al., 1991). In the present study, we have characterized the molecular mechanism of axo- nin-1 release. The results suggest that an endogenous glyco- sylPtdIns-specific phospholipase D (glycosylPtdIns-PLD) is in- volved in this release process. Furthermore, we found that the

Lierheimer et al. (EM J. Biochem. 243) 503
fourth FNIII-like domain of axonin-I is necessary for the secre- tion of axonin-I.
MATERIALS AND METHODS
Materials. Axonin-I was purified from vitreous fluid of 14- day-old chicken embryos (Ruegg et al., 1989). Rabbit polyclonal antibodies against axonin-1 were raised as previously described (Ruegg et al., 1989). Mouse monoclonal antibodies (mAbs) against axonin-I were raised as described elsewhere (Kunz, B. and Sonderegger, P., unpublished results). IgG and Fab were obtained as previously described (Stoeckli et al., 1991). Rabbit polyclonal serum against F11 was kindly provided by Dr F. G. Rathjen (Berlin, Germany). The anti-CRD serum was obtained from Oxford GlycoSystems Ltd. l,l0-Phenanthroline was ob- tained from Sigma. ["SIMethionine and [l-'H]ethanl-ol-2- amine hydrochloride were from Amersham. Myo[l,2-'H]inositol was from New England Nuclear.
Methods. Cell cultures. Dorsal root ganglia were dissected from 10-day-old chicken embryos and dissociated as described previously (Sonderegger et al., 1985). For the immunoprecipita- tion assay, 500000 cells were plated in 2 ml medium on colla- gen-(Boehringer Mannheim)-coated culture dishes of 35 mm di- ameter. The serum-free medium was composed of Eagle's mini- mal essential medium (Gibco-BRL) in Eagle's salts, 5 mg/ml Albumax, 20ng/ml nerve growth factor and N, (100 pg/ml transferrin, 200 pM putrescine, 15.78 ng/ml sodium selenite, 20 ng/ml triiodothyronine, 12.6 ng/ml progesterone, 200 ng/ml corticosterone, and 10 pg/ml insulin). Glutamine was freshly added to a concentration of 2 mM. Serum-containing medium was supplemented with 5 % horse serum and 5 % chicken embry- onic extract instead of Albumax. To minimize proliferation of non-neuronal cells, 0.12 mM S-fluorodesoxyuridine/0.3 mM uri- dine (Sigma) was added. To inactivate the glycosylPtdIns-PLD activity in the supplemented DRG medium, the pH was adjusted to 11 with 10 M NaOH and incubated for 1 h at 37°C. Then, the pH was adjusted to pH 7-8 with 6 M HCL. After this treat- ment, no glycosylPtdIns-PLD activity could be measured. The experiments with the anti-CRD serum, 1 ,lo-phenanthroline and monoclonal antibodies against axonin-1 were performed using serum-containing, serum-free and serum-free pH-inactivated medium.
Metabolic radiolabeling of proteins in DRG neurons and in myeloma cell lines. DRG cultures containing 500000 cellddish were metabolically radiolabeled 3 days after plating by addition of 50 pCi/ml ["S]methionine (10 pCi/pl) in 800 p1 medium for 16 h. DRG cultures for double-labeling experiments were grown for 4 days; subsequently, the medium was replaced by 2 ml ra- diolabeling medium containing 10 pCi [35S]methionine and 200 pCi [3H]ethanolamine. After 4-5 days, proteins were har- vested as described. The recombinant proteins of the myeloma cell lines were radioactively marked by resuspending 500 000 cells in 1 ml Dulbecco's modified Eagle's medium (Gibco-BRL) containing 10% fetal calf serum (Gibco-BRL) or serum-free me- dium (Amimed) and 50 pCi ["S]methionine.
Determination of glycosylPtdlns-anchor components in solu- ble axonin-I. The determination of ethanolamine and inositol of axonin-I purified from vitreous fluid was performed according to the method of Ferguson (1993).
Immunoprecipitation and quant$cation of the proteins. Cell- culture supernatants were collected and the cells washed once with 400 pl fresh medium. The washing solutions were com- bined with the supernatants. To harvest the glycosylPtdIns-an- chored proteins, cells were incubated with 800 pl 5 mg/ml solu- tion of Albumax in 0.5 mM MgC12, 136.9 mM NaCl, 2.7 mM
KCl, 8.1 mM Na,PO,,l.S mM KH,PO, (buffer A) containing 200 mU phosphatidylinositol-specific phospholipase C (Boeh- ringer Mannheim). After incubation for 1 h at 37"C, the super- natants were removed. After this procedure, more than 90% of the cells were viable and, when plated on laminin, they formed neurites. The plates were washed once with 400 p1 buffer A/ Albumax and the wash solution was added to the supernatants. To all probes 10% (by vol.) vitreous fluid (as carrier) of 14- day-old chicken embryos and 10% (by vol.) 10 X stock solution 500 mM Tris, 1.5 mM NaC1, 10% Triton X 100, 50 mM EDTA, pH 7.5 (buffer B) were added, then centrifuged for 1 h at 100000 g. No protease inhibitors were used because axonin-I was found to be stable in cell-culture medium for days. Axonin- 1 and the Ax-F1 1 hybrid were immunoprecipitated by adding 2% (by vol.) rabbit axonin-l antiserum. F11 and the F11-Ax- 1 hybrid were immunoprecipitated with rabbit F11 antiserum. After gently shaking overnight at room temperature, 2 % (by vol.) fixed Staphylococcus aureus was added to the samples to immobilize the antigen-antibody complex. The probes were then shaked again for 1 h and, afterwards, the suspensions were centrifuged and the pellets were washed five times with fresh buffer B by resuspension and centrifugation. The bound antigen- antibody complexes were then released by boiling the pellets for 10 min in reduced Laemmli-buffer (Laemmli, 1970).
The samples were centrifuged, and the supernatants of the subsequent centrifugation were separated by SDSPAGE. The gel was fixed in 30% ethanol, 10% acidic acid, washed with water and exposed for one day on a Phosphorimager storage plate. Quantification of the protein bands was performed by vol- ume integration using the ImageQuant software (Molecular dy- namics). For each result, at least five independent experiments were averaged.
Assay for glycosylPtdlns-spec@ phospholipase D activity. GlycosylPtdIns-specific phospholipase D activity was assayed using the purified membrane form of acetylcholinesterase (mf- AChE) from bovine erythrocytes as substrate. The mf-AChE is covalently linked to a glycosylPtdIns anchor, and the conversion of amphiphilic mf-AChE to soluble enzyme (s-AChE) serves as a measure for anchor-degrading activity (Stieger and Brodbeck, 1988). Under standard assay conditions, the reaction mixtures contained 0.6 pmol mf-AChE in 50 mM Mes, pH 6.5, 0.5 mM CaCI, and variable amounts of Triton X-114 (0.02-0.1 %) in a final volume of 100 pl. The assay was started by incubation of the tubes at 37°C in a water bath. The product of the reaction (s-AChE) was separated from the substrate by phase separation in Triton X-I 14, according to the method of Bordier (1981), and AChE activity was measured according to the method of Ellman et a]. (1961) as detailed by Heller et al. (1992). One unit of glycosylPtdIns-specific phospholipase D activity equals 1 pmol s-AChE produced/min (1 pmol AChE corresponds to 0.43 IU AChE activity (Stieger et al., 1991). Whole DRG ganglions and brain of 14-day-old chicken embryos were washed three times with buffer A. The intact vitreous body was also washed with buffer A and subsequently centrifugated to obtain the pure vitre- ous fluid.
Construction of Ax-I - FII hybrids. The axonin-1 -expressing myeloma cell line was described recently (Rader et al., 1993). To construct the F1 I-expressing and the Ax-I -F11-hybrid-ex- pressing cell lines, the cDNA fragments RLFl IF-RLFl lB, RLFl 1 F-RLF11B2 and RLFllF2-RLF11B were amplified using the polymerase chain reaction (Saiki et al., 1988) on the full-length F11 clone (Briimmendorf et al., 1989). PCR amplifi- cation was performed as described (Rader et al., 1993). RLFIIF- RLFllB was digested with Sac1 and HindIII and cloned into the identically digested vector pMAX (Rader et al., 1993) to pro- duce pMFl1. RLFllF2-RLFI 1B was digested with HindIII and

504 Lierheimer et al. (Eul: J. Biochem. 243)
cloned into the HindIII-digested vector pMAX to obtain pMAX- 1 -F11. RLFl 1F-RLF1 1B2 was digested with Sac1 and HindIII- and cloned into the similarly digested vector pMAX. The result- ing clone was digested with HindIII and ligated with the HindIII fragment of axonin-1 to give the F11-Ax-1 hybrid. All con- structs were verified by DNA sequencing. The oligonucleotides used were as follows:
RLFl1 F, S'-CTCTGAATAGG AGCTCACCATGAGGTT- CTTC-3' (SucI);
RLFl 1 F2, S'CCAAGCTTACCATCACAGAGAATCAGA- ACTG-3 (HindIII);
RLFl 1 B, S-CGAAGCTTACTTACCTAGAATCCCGAG- TAGGCAAGGACACC-3' (HindIII) (antisense: splice consensus donor site is bold) ;
AAAGTG-3' (HindIII). RLFl 1 B2,S'-GATGGCAAGCTTGGTTCCCCAGTTCCC-
The amino acid sequences of the resulting hybrids were as fol- lowed :
Ax-1 -FI 1 hybrid, Ax-I (MI-L693)-F11(P686-F1009); F11 -Ax-1 hybrid, F11 (Ml-P686)-Ax-l (S695-L1036).
Construction of'uxonin-I-deletion mutunts. A 1 .l-kb HindIII fragment, isolated from pMAX, was cloned into the phagemid vector pTZ19U (Bio-Rad) generating pTZ19UlaxH-H. For in vi- tro mutagenesis, the Escherichia coli strain CJ236 and the helper phagemid VCSMl3 (Stratagene) were used to obtain uracil-con- taining single-strand DNA as template (Yuckenberg et al., 1991). The domain deletions were made by loop-out oligonucleotide- directed mutagenesis using the following oligonucleotides :
RLdFn2,S'-CGTACCAAAGAAGCAGCACCCAAGGTGG-
RLAFn3, S'-GTGTACTCTGCAGAAGAAGAACCAAGGA- GGCC ACCAGGCAACATCTCC-3' ;
RLAFn4, S-AACATCACGACCACAAAGCCACCAACCA- GTATGATGGTGGAAGACTCC-3'.
Oligonucleotide phosphorylation, annealing and second-strand synthesis were performed as described (Yuckenberg et al., 1991) using pTZ19UhxH-H for RLdFn2, RLdFn3 and RLAFn4. All constructs were verified by DNA sequencing. The amino acid sequence of the resulting deletion mutants were as follows:
CACCATTCAGAGTTACGGCCAAAGCC-3' ;
AFn2, des-(T706-P807)-axonin-l ; AFn3, des-(K808-P908)-axonin-l; AFn4, des-(P907 -GI OOI)-axonin-l.
RESULTS
Axonin-1 and F11 differ significantly in their secretion. To quantify the extent of secretion of axonin-I and F11 in DRG cultures, dissociated chicken dorsal root ganglion neurons were metabolically radiolabeled with ["Slmethionine for 16 h. After that, supernatants containing soluble forms of axonin-1 and F11 were collected and the cells were incubated with phosphatidyl- inositol phospholipase C (PtdIns-PLC) to harvest the glycosyl- PtdIns-anchored fraction of the proteins. In both fractions, axo- nin-1 and F11 were immunoprecipitated with the corresponding polyclonal antisera and the ratios between the secreted and the membrane-bound forms of axonin-I and F11 were determined. 68% of the total amount of axonin-I was released (Fig. 1). In contrast, only 13% F11 was found in solution. These experi- ments reveal that, despite the high sequence similarity between these two molecules, axonin-1 is secreted to a much greater ex-
A F11
S P
125 -
B
50 5
5
axonin-1
S P
F11 axonin-1
Fig. 1. Quantification of the secretion of F11 and axonin-1 in DRG cultures. (A) DRG cultures were metabolically radiolabeled with ["S]methionine (SO pCi/ml). After 16 h. supernatants (S) containing the secreted proteins were collected. To determine the membrane-bound fraction (P), the cells were incubated with PtdIns-PLC for 1 h. Collected proteins were immunoprecipitated and subjected to SDSlPAGE (7.5 %) under reducing conditions. Protein bands were quantified after exposing on a Phosphorimager screen using the ImageQuant software. The molec- ular mass is indicated in kilodaltons (kDa). (B) The ratios of the secreted and the membrane-bound fractions of F11 and axonin-I were determined and the release was calculated. Columns represent the mean of five inde- pendent experiments ; error bars indicate standard deviation.
tent than F1 I . Control experiments suggest that the difference in the amount released is rather underestimated in the measure- ments provided here, because axonin-I was more efficiently cleaved from the cells than F11 by PtdIns-PLC. When cells were tested for axonin-1 and F11 after PtdIns-PLC treatment, no axo- nin-1 was found (see also Zuellig et al., 1992), whereas approxi- mately SO% of F11 showed resisted PtdIns-PLC cleavage (see also Wolff et al., 1989).
Soluble axonin-1 contains glycosylPtdIns anchor compo- nents. Axonin-1 is attached to the membrane by a glycosylPtd- Ins anchor (Zuellig et al., 1992). If the released form of axonin-1 is generated by a glycosylPtdIns-specific phospholipase, soluble axonin-I should contain part of the glycosylPtdIns-anchor. Therefore, secreted axonin-I was examined for the presence of ethanolamine by simultaneously radiolabeling the DRG cultures with ['5S]methionine and ['Hlethanolamine. After immunopreci- pitation, the ratio "SPH was determined. The "S value corre- sponded to the axonin-1 protein while the 'H value corresponded to the glycosylPtdIns anchor. The ratio 35S/'H for secreted axo- nin-I was 0.44 whereas PtdIns-PLC-cleaved axonin-I exhibited a "SPH ratio of 0.35. Assuming that all PtdIns-PLC-cleaved axonin-1 contains ethanolamine, 80 % of secreted axonin-I car- ries ethanolamine and, therefore, part of the glycosylPtdIns an- chor. The analogous experiment was carried out with ['Hlinosi- tol, another component of the glycosylPtdlns anchor. The "SPH ratios of secreted (0.77) and PtdIns-PLC-cleaved (0.46) axonin- 1 lead to the conclusion that 55% of the soluble axonin-I in DRG cultures contain inositol. Probably due to the inherently
A F11 axonin-1
S P S P
Lierheimer et al. ( E m J. Biochern. 243)
' O 8o i A
505
125 - B S P S P
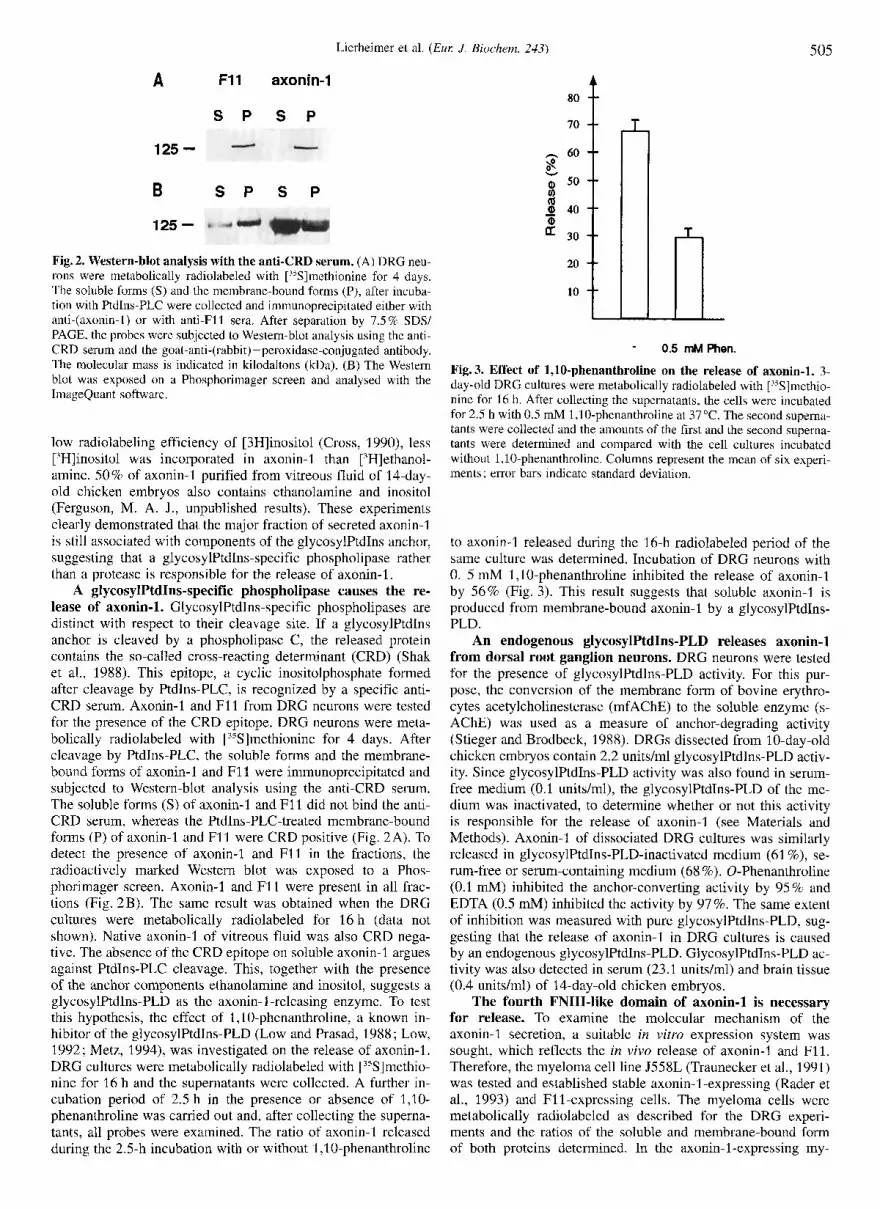
125 - Fig. 2. Western-blot analysis with the anti-CRD serum. (A) DRG neu- rons were metabolically radiolabeled with [35S]methionine for 4 days. The soluble forms (S) and the membrane-bound forms (P), after incuba- tion with Ptdlns-PLC were collected and immunoprecipitated either with anti-(axonin-I) or with anti-F11 sera. After separation by 7.5% SDS/ PAGE, the probes were subjected to Western-blot analysis using the anti- CRD serum and the goat-anti-(rabbit) -peroxidase-conjugated antibody. The molecular mass is indicated in kilodaltons (ma). (B) The Western blot was exposed on a Phosphorimager screen and analysed with the ImageQuant software.
low radiolabeling efficiency of [3H]inositol (Cross, 1990), less ['Hlinositol was incorporated in axonin-1 than ['Hlethanol- amine. 50% of axonin-1 purified from vitreous fluid of 14-day- old chicken embryos also contains ethanolamine and inositol (Ferguson, M. A. J., unpublished results). These experiments clearly demonstrated that the major fraction of secreted axonin-1 is still associated with components of the glycosylPtdIns anchor, suggesting that a glycosylPtdIns-specific phospholipase rather than a protease is responsible for the release of axonin-1.
A glycosylPtdIns-specific phospholipase causes the re- lease of axonin-1. GlycosylPtdIns-specific phospholipases are distinct with respect to their cleavage site. If a glycosylPtdIns anchor is cleaved by a phospholipase C, the released protein contains the so-called cross-reacting determinant (CRD) (Shak et al., 1988). This epitope, a cyclic inositolphosphate formed after cleavage by PtdIns-PLC, is recognized by a specific anti- CRD serum. Axonin-1 and F11 from DRG neurons were tested for the presence of the CRD epitope. DRG neurons were meta- bolically radiolabeled with [15S]methionine for 4 days. After cleavage by PtdIns-PLC, the soluble forms and the membrane- bound forms of axonin-1 and F11 were immunoprecipitated and subjected to Western-blot analysis using the anti-CRD serum. The soluble forms (S) of axonin-1 and F11 did not bind the anti- CRD serum, whereas the PtdIns-PLC-treated membrane-bound forms (P) of axonin-1 and F11 were CRD positive (Fig. 2A). To detect the presence of axonin-I and F11 in the fractions, the radioactively marked Western blot was exposed to a Phos- phorimager screen. Axonin-I and F11 were present in all frac- tions (Fig. 2B). The same result was obtained when the DRG cultures were metabolically radiolabeled for 16 h (data not shown). Native axonin-1 of vitreous fluid was also CRD nega- tive. The absence of the CRD epitope on soluble axonin-I argues against Ptdlns-PLC cleavage. This, together with the presence of the anchor components ethanolamine and inositol, suggests a glycosylPtdIns-PLD as the axonin-1 -releasing enzyme. To test this hypothesis, the effect of 1 ,lO-phenanthroline, a known in- hibitor of the glycosylPtd1ns-PLD (Low and Prasad, 1988; Low, 1992; Metz, 1994), was investigated on the release of axonin-1. DRG cultures were metabolically radiolabeled with ['5S]methio- nine for 16 h and the supernatants were collected. A further in- cubation period of 2.5 h in the presence or absence of 1,10- phenanthroline was carried out and, after collecting the superna- tants, all probes were examined. The ratio of axonin-1 released during the 2.5-h incubation with or without 1 ,lo-phenanthroline
- 0.5 mM Phen.
Fig. 3. Effect of lJ0-phenantbroline on the release of axonin-1. 3- day-old DRG cultures were metabolically radiolabeled with [-?S]methio- nine for 16 h. After collecting the supernatants, the cells were incubated for 2.5 h with 0.5 mM 1 ,lo-phenanthroline at 37OC. The second superna- tants were collected and the amounts of the first and the second superna- tants were determined and compared with the cell cultures incubated without 1,lO-phenanthroline. Columns represent the mean of six experi- ments ; error bars indicate standard deviation.
to axonin-1 released during the 16-h radiolabeled period of the same culture was determined. Incubation of DRG neurons with 0. 5 mM 1,10-phenanthroline inhibited the release of axonin-1 by 56% (Fig. 3). This result suggests that soluble axonin-1 is produced from membrane-bound axonin-1 by a glycosylPtdIns- PLD.
An endogenous glycosylPtdIns-PLD releases axonin-1 from dorsal root ganglion neurons. DRG neurons were tested for the presence of glycosylPtdIns-PLD activity. For this pur- pose, the conversion of the membrane form of bovine erythro- cytes acetylcholinesterase (mfAChE) to the soluble enzyme (s- AChE) was used as a measure of anchor-degrading activity (Stieger and Brodbeck, 1988). DRGs dissected from 10-day-old chicken embryos contain 2.2 unitdm1 glycosylPtdIns-PLD activ- ity. Since glycosylPtdIns-PLD activity was also found in serum- free medium (0.1 units/ml), the glycosylPtdIns-PLD of the me- dium was inactivated, to determine whether or not this activity is responsible for the release of axonin-1 (see Materials and Methods). Axonin-1 of dissociated DRG cultures was similarly released in glycosylPtdIns-PLD-inactivated medium (61 %), se- rum-free or serum-containing medium (68 %). 0-Phenanthroline (0.1 mM) inhibited the anchor-converting activity by 95% and EDTA (0.5 mM) inhibited the activity by 97%. The same extent of inhibition was measured with pure glycosylPtdIns-PLD, sug- gesting that the release of axonin-1 in DRG cultures is caused by an endogenous glycosylPtdIns-PLD. GlycosylPtdIns-PLD ac- tivity was also detected in serum (23.1 unitdml) and brain tissue (0.4 units/ml) of 14-day-old chicken embryos.
The fourth FNIII-like domain of axonin-1 is necessary for release. To examine the molecular mechanism of the axonin-1 secretion, a suitable in vitro expression system was sought, which reflects the in vivo release of axonin-1 and F11. Therefore, the myeloma cell line J558L (Traunecker et al., 1991) was tested and established stable axonin-1-expressing (Rader et al., 1993) and F11-expressing cells. The myeloma cells were metabolically radiolabeled as described for the DRG experi- ments and the ratios of the soluble and membrane-bound form of both proteins determined. In the axonin-1-expressing my-

506 Lierheimer et al. ( E m J. Biochem. 243)
Ax-1IF11 w *
t- \ *-, 8 - \
F11IAx-1 m> ?, & ?. $ ?, n n 1 1 1 1 1 1 -.4- -.0- .I
B 60
50
40
30
20
10
h
9 e 3 U
B
- AX-1 Ft t A X - t f f i i Fi l/AX-1
Fig.4. Quantification of the secretion of axonin-1 and F11 and the axonin-1IF11 hybrids expressed in myeloma cells. (A) Structures of the hybrids composed of axonin-1 and F11. The six Ig-like domains of axonin-1 are represented as loops and the four FNI11-like domains as open boxes, whereas corresponding domains of F11 are indicated as dashed loops and filled boxes. (B) Quantitative analysis of the secretion of axonin-I, F11, and the hybrids stably expressed in J558L myeloma cells are shown. The corresponding cell lines were metabolically radio- labeled with [35S]methionine and the quantification was done as outlined in Fig. 1 . The columns represent the mean of six independent experi- ments with three independently derived cell lines of each construct; error bars indicate standard deviation.
eloma cell line, 50% axonin-I was released, whereas in the F11- expressing cell line, only 23 % F11 was released (Fig. 4B). This indicates that axonin-1 and F11 also behave differently with re- spect to their release when expressed heterologically in a non- neuronal cell line, and it was concluded that the distinctive re- lease characteristics of axonin-I and F11 are determined by in- trinsic properties of the proteins.
As a first step towards domain localization of a putative secretion motif, the last three FNIII-like domains of axonin-I were exchanged with those of F11 and vice versa. The resulting axonin-1 -F11 hybrid molecule (Ax-1 -F11) contained all six Ig-like domains and the first FNIII-like domain of axonin-I (M1-L693) fused to the three C-terminal FNIII-like domains of F11 (P686-F1009). The complementary F11 -axonin-1 hybrid (F11 -Ax-1) was composed of all six Ig-like domains and the first FNIII-like domain of F11 (MI -P686) and the three C-termi- nal FNIII-like domains of axonin-1 (S695-Ll036) (Fig. 4A). The Ax-1 -F11-expressing cell line secreted 24% of the hybrid molecule, as did F11 cells. Conversely, the F11 -Ax-1-express- ing cell line exhibited 50% secretion, the same secretion be- haviour as Ax-1 cells (Fig. 4B). Thus, the substitution of the last three FNIII-like domains of axonin-1 with those of F11 re- duced the release of axonin-I to the level of F11. In contrast, the substitution of the last three FNIII-like domains of F11 with those of axonin-1 resulted in a secretion comparable with that
30
20
10
T
Ax-1 AFn2 AFn3 AFn4
Fig. 5. Quantification of the secretion of axonin-1 deletion mutants expressed in the myeloma cell system. (A) Structures of the axonin-l- deletion mutants. Domains of axonin-1 as defined by their similarity to the type-I11 domain of fibronectin and by exon-intron borders of the axonin-1 gene were deleted by oligonucleotide-directed mutagenesis. Lines indicate the deleted domain. The names of mutants indicate the domain that is deleted. (B) Quantification of the release of the axonin-I deletion mutants expressed in myeloma cells were carried out as outlined in Fig. 1. The columns represent the mean of seven independent experi- ments with three independently derived cell lines of each construct; error bars indicate standard deviation.
of axonin-1. These experiments indicate that the last three FNIII- like domains of axonin-I are necessary and sufficient for its secretion.
To localize the secretion motif more precisely, protein do- mains of axonin-1, as defined by their similarity to the type-111 domain of fibronectin and by exonlintron boundaries of the axo- nin-I gene (Giger et al., 1995), were deleted by oligonucleotide- directed mutagenesis (Fig. 5A). The mutants were expressed in myeloma cells and tested for their release properties. After meta- bolically radiolabeling, the ratios between the soluble and glyco- sylPtdIns-anchored forms were determined. The deletion mutant deficient for the second FNIII-like domain, denominated dFn2 [des-(T706-P807)-axonin-l], was released similarly to wild- type axonin-I (44% compared to 50%). In contrast, deletion of the fourth FNIII-like domain, dFn4 [des-(P907 - G1001)-axonin 11, reduced the release to the level of F11 (22 % ; Fig. 5 B). These experiments indicate that the fourth FNIII-like domain of axo- nin-1 is necessary for release. Surprisingly, dFn3 [des-(K808 - P908)-axonin-l], which lacks the third FNIII-like domain, was

507
B
80
70
60
50
2 40
30
20
lo
h
8 v
0 u) m
2
A
4
-- -- --
-- -- -- -- -- -
B 70 --
AX-1
Lierheimer et al. (Eul: J. Biochem. 243)
A
S P S P S P
125 - 125 -
no Fab X9C3 X7H1 Fab Fab
noFab X9C3 X7H 1 Fab Fab
Fig. 6. Analysis of release inhibition of axonin-1, expressed in the myeloma cell system, by Fabs, directed against the fourth FNIII-like domain of axonin-1. (A) Axonin-1-expressing myeloma cells (Ax-1 ) were metabolically radiolabeled with ["Slmethionine and incubated without or with 40 pg/ml Fab. After 16 h, supernatants (S) were col- lected and the cells were incubated with PtdIns-PLC for 1 h to harvest the membrane-bound form (P) of axonin-1. All fractions were collected, immunoprecipitated with anti-(axonin-1) serum, and analysed after exposing to a Phosphorimager screen. The molecular mass of axonin-I is indicated in kilodaltons (kDa). (B) The amounts of the secreted and the membrane-bound forms were determined and the amount released calculated. Columns represent the mean of eight experiments ; error bars indicate standard deviation.
released to a considerably higher proportion (65%) than wild- type axonin-1. The axonin-1, F1 1 - Ax hybrid and the axonin-I - deletion mutants expressed in myeloma cells were 100% sensi- tive to PtdIns-PLC-cleavage, in contrast to F11 and the Ax- F11 hybrid, where the PtdIns-PLC cleaves approximatly 50 % of these membrane-anchored forms (data not shown).
To confirm independently that the fourth FNIII-like domain of axonin-1 is required for secretion, a panel of mAbs directed against single Ig-like o r FNIII-like domains was tested for their capacity to inhibit axonin-1 release in myeloma cells. mAb X9C3, which recognizes an epitope localized on the fourth FNIII-like domain of axonin-1, was both as IgG (30 pg/ml final concentration) and as Fab (40 pg/ml final concentration) able to reduce the release of axonin-1 from 50% to 17%, comparable to F11 levels (23%; Fig. 6). The antibody X7H1, also directed against the fourth FNIII-like domain, did not reduce the release of axonin-I (52%). When these experiments were done in DRG cultures, X9C3 Fab decreased the release of axonin-1 from 68 % to 21 %. The Fab X7H1 did not significantly reduce the release of axonin-I (from 68% to 5 8 % , see Fig. 7). All other tested antibodies, directed against a single Tg-like or FNIII-like domain
axonin-1
S P S P S P
no Fab x9c3 X7H1 Fab Fab
T
noFab X9C3 X7H1 Fab Fab
Fig. 7. Analysis of release inhibition of axonin-1 of dorsal root gan- glion neurons by Fabs, directed against the fourth FNIII-like domain of axonin-1. (A) 3-day-old DRG cultures were metabolically rddio- labeled with [3SS]methionine and incubated without or with 40 pg/ml Fab. The quantitative analysis of the supernatants (S) and the membrane- bound form (P) of axonin-1 was donc as outlined in Fig. 1. The molecu- lar mass of axonin-1 is indicated in kilodaltons (kDa). (B) The amounts of the secreted and the membrane-bound forms were determined and the amount released calculated. Columns represent the mean of nine experiments ; error bars indicate standard deviation.
as determined by immunostaining and Western-blot analysis of the axonin-1-deletion mutants (data not shown; Rader et al., 1996), had no effect on axonin-1 secretion either in the myeloma cell system or in DRG cultures (data not shown). These results support the conclusion drawn from the deletion mutants that the fourth FNIII-like domain is involved in the release of axonin-1 from the membrane.
DISCUSSION
Many eucaryotic cell-surface proteins are linked to the mem- brane by a glycosylPtdIns anchor (Ferguson and Williams, 1988). This form of membrane anchorage can provide several functions including low turnover rates, sorting to the apical sur- face of polarised cells (Brown, 1992; Censullo and Davitz, 1994), and the possibility to produce soluble molecules by cleav- age of the glycosylPtdIns anchor by phospholipases (Ferguson and Williams, 1988; Low, 1989; Cross, 1990). Several glyco- sylPtdIns-anchored membrane-bound proteins have also been observed in biological fluids and cell-culture supernatants, One of these molecules is the chicken AxCAM axonin-1 which oc- curs both as a membrane-bound and as a secreted protein (Stoeckli et al., 1991). Based on in vitro and in vivo observa-

508 Lierheimer et al. (Eur: J. Biochem. 243)
tions, soluble axonin-1 has been implicated as a regulator of axonal fasciculation and pathfinding decisions. Here, we present evidence that the glycosylPtdIns anchor of axonin-1 is cleaved by an endogenous glycosylPtdIns-PLD activity and that the re- lease mechanism depends on the presence of the membrane- proxima1 fourth FNTII-like domain of axonin-I.
Evidence for glycosylPtdlns anchorage of membrane-bound axonin-1 has been presented previously. Membrane-bound axo- nin-1 of chicken DRG neurons is completely cleaved from the membrane by PtdIns-PLC (Zuellig et al., 1992). The membrane- bound and the soluble form of axonin-1 are immunologically identical (Ruegg et al., 1989) and no alternative splicing of the axonin-I mRNA was detected (Giger et al., 1995). These data indicated that the soluble form of axonin-1 is generated by post- translational cleavage either by a protease or a phospholipase activity. To examine whether the soluble form of axonin-I is generated by a protease that cleaves axonin-1 close to the C- terminus or a phospholipase that cleaves the glycosylPtdIns an- chor secreted axonin-I was tested for the presence of the glyco- sylPtdlns anchor components ethanolamine and inositol (Fergu- son and Williams, 1989; Cross, 1990). Approximately 80% of soluble axonin-I in DRG neuron cultures contained the etha- nolamine residue, indicating that a phospholipase rather than a protease is involved in cleavage. This result was confirmed by double-labeling experiments using ['H]inositol and ["S]methio- nine. Ethanolamine and inositol were also found in native axo- nin-I purified from vitreous fluid. Thus, the soluble form of axonin-I is derived from the glycosylPtdIns-anchored form, which is presumably cleaved by a phospholipase. The observa- tion that only 50% of axonin-1 purified from vitreous fluid con- tain glycosylPtdIns-anchor components could be explained by the presence of proteases which cleave axonin-1 after glyco- sylPtdIns-PLD cleavage. Two types of phospholipases are known, PtdIns-PLC and glycosylPtdIns-PLD. They differ in their cleavage site (Low, 1989). The conclusion that the release of glycosylPtdIns-anchored axonin-I is mediated by a glyco- sylPtdlns-PLD is supported by the following observations : (a) Secreted axonin-1 of DRG neurons and purified from vitreous fluid contains glycosylPtdIns-anchor components. (b) Soluble axonin-1 of DRG neurons and purified from vitreous fluid lack the CRD epitope, an epitope formed after cleavage of the glyco- sylPtdIns anchor by PtdIns-PLC but not by glycosylPtdIns-PLD (Ferguson and Williams, 1988; Shak et al., 1988). (c) Treatment of DRG neurons with 1 ,lo-phenanthroline, an inhibitor of glyco- sylPtdIns-PLD (Low and Prasad, 1988; Davitz et al., 1989), sig- nificantly reduces the release of axonin-1 .
GlycosylPtdIns-PLD activity was found in dorsal root gan- glion neurons, brain tissue and serum of the chicken embryo. To exclude the possibility that the detected glycosylPtdIns-PLD activity present in serum-free DRG medium is responsible for the release of axonin-1, this activity was inactivated. The release of axonin-1 in the inactivated medium was similar to the release of axonin-1 in serum-free and serum-containing media. This ob- servation is consistent with data showing that serum glycosyl- PtdIns-PLD activity does not release glycosylPtdIns-anchored proteins when added to cells (Davitz et al., 1989; Low and Hu- ang, 1991 ; Bergman and Carlson, 1994). These observations support the conclusion that an endogenous glycosylPtdIns-PLD activity expressed in DRG neurons is responsible for the release of axonin- 1. An endogenous glycosylPtdIns-PLD has been shown to release the basic fibroblast growth-factor-heparan- sulfate-proteoglycan complexes from human bone marrow cul- tures (Brunner et al., 1994). So far, the cellular location of the glycosylPtdlns-PLD that cleaves axonin-1 is unknown. The glycosylPtdIns-PLD could be secreted from neurons or it could be membrane-bound and located either at the surface or in-
ternally in post-Golgi vesicles. Recently, it has been shown that a cell-membrane-associated glycosylPtdIns-specific phospholi- pase releases the glycosylPtdIns-anchored complement regula- tory protein decay-acceleration factor from HeLa cells and the basic fibroblast growth-factor-binding heparan sulfate proteogly- can from bone marrow stromal cells (Metz et al., 1994).
In contrast to other axonal cell-adhesion molecules such as F11, NgCAM and NrCAM, axonin-1 exists predominantly as a secreted molecule in vivo (Stoeckli et al., 1991). Despite the structural and functional similarities between axonin-1 and F11, only axonin-I is cleaved to a great extent. The release of axonin- 1 and F11 observed in vivo has been quantified and axonin-I found to be secreted fivefold more than F11 in DRG neurons (Fig. 1). To characterize the molecular mechanism responsible for this difference, a suitable irz vitro system was sought which reflects this in vivo situation. Axonin-1 and F11 were expressed stably in the myeloma cell line J558L (Traunecker et al., 1991 ; Rader et al., 1993) and, as in the DRG neurons, a clear differ- ence was found in the release of axonin-1 and F11. The observa- tion that both proteins expressed in the same cell system still differ markedly in secretion strongly suggested that the proteins themselves contain the information for release. To support that the release of axonin-1 expressed in myeloma cells is also due to glycosylPtdIns-PLD as in DRG neurons, the soluble form of axonin-1 was tested for the CRD epitope. As in DRG neurons, soluble axonin-1 was CRD negative (data not shown). To inves- tigate which domains are necessary for release, hybrids of axo- nin-1 and F11 were constructed. The AxlF11 hybrid, which con- tains the last three FNIII-like domains of F11, is secreted as little as is F1 I and that the F11IAx hybrid is secreted as much as is axonin-1. In these experiments, the strong secretion prop- erty of axonin-1 was transferred to F11 and it could thereby be demonstrated that the last three FNIII-like domains of axonin-1 are necessary and sufficient for release. To investigate in more detail the localization of [he putative secretion motif, entire FNITI-like domains were deleted. The deletion of the second FNIII-like domain showed no effect on the secretion. Excision of the third FNIII-like domain of axonin-1 resulted in a signifi- cantly higher secretion, whereas deletion of the fourth FNIII- like domain of axonin-1 reduced the release level to that of F11 (Fig. 5) . Assuming that the fourth FNIII-like domain is neces- sary for release, the enhanced secretion of dFn3 could be ex- plained by better accessibility of the fourth FNIII-like domain for the glycosylPtdIns-phospholipase activity. With monoclonal antibodies directed against single Ig-like and FNIII-like do- mains, it was confirmed that the fourth FNIII-like domain is necessary for release. The Fab X9C3, directed against the fourth FNIII-like domain of axonin-I reduced the release in myeloma cells (Fig. 6) and in DRG cultures (Fig. 7) to the level of F11. This effect can be explained by the antibody being able to block the release-recognition motif on the fourth FNIII-like domain for the glycosylPtdIns-PLD. These data strongly suggest that the fourth FNIII-like domain of axonin-1 is necessary for release. One possible explanation for the low release level of F11 is the absence of the putative secretion motif. Alternatively, one can speculate that, despite sequence similarity between axonin-I and F11, the conformation of the fourth FNIII-like domain or of the whole molecule is different and, therefore, F11 is not accessible for glycosylPtdTns-PLD cleavage.
The biological significance of cleavage and release of mem- brane-associated glycosylPtdIns-anchored proteins by glyco- sylPtdTns-PLD is not yet known. It is conceivable that glyco- sylPtdIns-PLD activity regulates the surface concentration of membrane-bound axonin-1, which influences the interaction of axonin-1 with NgCAM andlor NrCAM. This could affect fascic- ulation or might influence the pathfinding decisions of axons in

Lierheimer et al. (Eur: J . Biochem. 243) 509
the developing chicken embryo (Stoeckli and Landmesser, 1995). Fasciculation and pathfinding can also be influenced by competition between membrane-bound axonin-1 and F11 for their heterologous binding partners. Alternatively, an increased local concentration of soluble axonin-1 might disturb the homo- philic axonin-I /axonin-1 interaction and/or the interaction be- tween axonin-I and NgCAM/NrCAM and might, therefore, have an anti-adhesive effect as shown recently (Stoeckli and Land- messer, 1995).
The authors thank Dr Michael A. J. Ferguson for the ethanolamine and inositol analyses of axonin-1 from vitreous fluid. We also thank Dr Fritz Rathjen for kindly providing polyclonal F11 antiserum, Dr Thomas Briimmendorf for the F11 cDNA and Dr Juan Riesgo-Escovar for read- ing the manuscript. This work was supported by Swiss National Science Foundation grants 3100-039209.93 (U. B.), 3100-033815.92/2 (P. S.), the Sandoz Stifzing, the Julius-Klaus-Sl~tung and the Fonds fur medi- zinische Forschung der Universitat Zurich.
REFERENCES Bergman, A. S. & Carlson, S. R. (1994) Saponin induced release of
cell-surface-anchored-Thy-l by serum glycosylphosphatidylinositol- specific phospholipase D, Biochem. J. 298, 661 -668.
Bordier, C. (1981) Phase separation of integral membrane proteins in Triton X114 solution, J. Biol. Chem. 256, 1604-1607.
Brown, D. A. (1992) Interactions between glycosylPtdlns-anchored pro- teins and membrane lipids, Trends Cell Biol. 2, 338-343.
Brummendorf, T., Wolff, J. M., Frank, R. & Rathjen, F. G. (1989) Neural cell recognition molecule F1 1 : homology with fibronectin type I11 and immunoglobulin type C domains, Neuron 2, 1351 - 1361,
Briimmendorf, T., Hubert, M., Treubert, U., Leuschner, R., T h o k , A. & Rathjen, F. G. (1993) The axonal recognition molecule F11 is a multifunctional protein: specific domains mediate interactions with NgCAM and restrictin, Neuron 10, 1-20.
Brummendorf, T. & Rathjen, F. G. (1993) Axonal glycoproteins with immunoglobulin- and fibronectin type 111-related domains in verte- brates: Structural features, binding activities, and signal transduction, J. Neurochem. 61, 1207-1219.
Brunner, G., Metz, C. N., Nguyen, H., Gabrilove, J., Patel, S. R., Davitz, M. A,, Rifkin, D. B. & Wilson, E. L. (1994) An endogenous glyco- sylphosphatidylinositol-specific phospholipase D releases basic fi- broblast growth factor-heparan sulfate proteoglycan complexes from human bone marrow cultures, Blood 83, 2115-2125.
Censullo, P. & Davitz, M. A. (1994) How glycosylPtdlns-anchored pro- teins turnover: or where do they go after arrival at the plasma mem- brane, Sem. Immunol. 6, 81 -88.
Connelly, M. A,, Grady, R. C., Mushinski, J. F. & Marcu, K. B. (1994) PANG, a gene encoding a neuronal glycoprotein, is ectopically acti- vated by intracisternal A-type particle long terminal repeats in mu- rine plasmacytomas, P roc. Nut1 Acud. Sci. USA 91, 1337 - 1341.
Cross, G. A. M. (1990) Glycolipid anchoring of plasma membrane pro- teins, Annu. Rev. Cell Biol. 6, 1-39.
Davitz, M. A,, Hom, J. & Scheikman, S. (1989) Purification of a glyco- syl-phosphatidylinositol-specific phospholipase D from human plasma, J. B id . Chem. 264, 13 760- 13 764.
Ellman, G. L., Courtney, D. K., Andres, V. & Featherstone, R. M. (1961) A new and rapid colorimetric determination of acetylcholinesterase activity, Biochem. Pharmacol. 7, 88-95.
Ferguson, M. A. J. & Williams, A. F. (1988) Cell-surface anchoring of proteins via Glycosyl-phosphatidylinositol structures, Annu. Rev. Biochem. 57, 285-320.
Ferguson, M. A. J. (1993) glycosylPtdIns membrane anchors: isolation and analysis, in Glycobiology, A Practical Approach (Fukuda, M. & Kobata, A,, eds) pp. 349-383, IRL Press, Oxford, UK.
Furley, A. J. , Morton, S. B., Manalo, D., Karagogeos, D., Dodd, J. & Jessell, T. M. (1990) The axonal glycoprotein TAG-1 is an immuno- globulin superfamily member with neurite outgrowth promoting ac- tivity, Cell 61, 157-170.
Gennarini, G., Cibelli, G., Rougon, G.. Mattei, M. G. & Goridis, C. (1989) The mouse neuronal cell surface protein F3: a phosphatidyli-
nositol-anchored member of the immunoglobulin superfamily related to chicken contactin, J. Cell Biol. IOY, 775-788.
Giger, R. J., Vogt, L., Zuellig, R. A,, Rader, C., Henehan-Beatty, A., Wolfer, D. P. & Sonderegger, P. (1 995) The gene of chicken axonin- 1, complete structure and analysis of the promoter, Eur: .I. Biochem.
Goodman, C. S. & Shatz, C. J. (1993) Developmental mechanisms that generate precise patterns of neuronal connectivity, Cell 10, 77- 98.
Heller, M., Bieri, S. & Brodbeck, U. (1992) A novel form of glycosyl- phosphatidylinositol-anchor converting activity with a specificity of a phospholipase D in mammalian liver membranes, Biochem. Bio- phys. Acta 1109, 109-116.
Hynes, R. 0. & Lander, L. A. D. (1992) Contact and adhesive specifi- cities in the associations, migrations and targeting of cells and axons, Cell 68, 303-322.
Jessel, T. M. (1988) Adhesion molecules and the hierarchy of neural development, Neuron I , 3-13.
Keynes, R. & Cook, G. M. W. (1995) Axon guidance molecules, Cell
Kuhn, T. B., Stoeckli, E. T., Condrau, M. A., Rathjen, F. G. & Sonderegger, P. (1991) Neurite outgrowth on immobilized axonin-I is mediated by a heterophilic interaction with L1(G4), J. Cell B id .
Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of the bacteriophage T4, Nature 227, 680- 685.
Low, M. G. (1989) The glycosyl-phosphatidylinositol anchor of mem- brane proteins, Biochim. Biophys. Acta 988, 427-454.
Low, M. G. & Huang, K. S. (1991) Factors affecting the ability of glyco- sylphosphatidylinositol-specific phospholipase D to degrade the membrane anchors of cell surface proteins, Biochem. J. 279, 483- 493.
Low, M. G. (1992) in Lipid modifications of proteins: a practical ap- proach (Hooper, N. M. & Turner, H. A,, eds) pp. 117-154, Oxford University Press, Oxford, UK.
Low, M. G. & Prasad, A. R. S. (1988) A phospholipase D specific for the phosphatidylinostitol anchor of cell-surface proteins is abundant in plasma, Proc. Nut1 Acad. Sci. USA 85, 980-984.
Luo, Y., Raible, D. & Raper, J. A. (1993) Collapsin: a protein in brain that induces the collapse and paralysis of neuronal growth cones, Cell 75, 211-221.
Metz, C. N., Brunner, G., Choi-Muira, N. H., Nguyen, H., Gabrilove, I., Caras, I. W., Altzuler, N., Rifkin, D. B., Wilson, E. L. & Davitz, M. A. (1 994) Release of glycosylPtdIns-anchored membrane proteins by a cell-associated glycosylPtdlns-specific phospholipase D, EMBO J.
Morales, G., Hubert, M., Bruemmendorf, T., Treubert, U., Tanok, A., Schwarz, U. & Rathjen, F. G. (1993) Induction of axonal growth by heterophilic interactions between the cell surface recognition pro- teins F11 and NrCAM/Bravo, Neuron 11, 1 113 - 1122.
Rader, C., Stoeckli, E. T., Ziegler, U., Osterwalder, T., Kunz, B. & Sonderegger, P. (1993) Cell-cell adhesion by homophilic interaction of the neuronal recognition molecule axonin-1, Eur: J. Biochem. 215,
Rader, C., Kunz, B., Lierheimer, R., Giger, R. J., Berger, P., Tittmann, P., Gross, H. & Sonderegger, P. (1996) Implications for the domain arrangement of axonin-1 derived from the mapping of its NgCAM binding site, EMBO J. 15, 2056-2068.
Ranscht, B. (1988) Sequence of contactin, a 130-kD glycoprotein con- centrated in areas of interneuronal contact, defines a new member of the immunoglobulin supergene family in the nervous system, J. Cell Biol. 107, 1561-1573.
Ranscht, B. (1 994) Cadherins and catenins : interactions and functions in embryonic development, Curr: Opin. Cell Biol. 6, 740-746.
Reichardt, L. F. & Tomaselli, K. J . (1991) Extracellular matrix molecules and their receptors: functions in neural development, Annu. Rev. Neurosci. 14, 531 -570.
Ruegg, M. A,, Stoeckli, E. T., Kuhn, T. B., Heller, M., Zuellig, R. & Sonderegger, P. (1989) Purification of axonin-1, a protein that is secreted from axons during neurogenesis, EMBO J. 8, 55-63.
Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S . J., Higuchi, P., Horn, G. T., Mullis, K. B. & Erlich, H. A. (1988) Primer-directed enzy-
227, 617-628.
83, 161-169.
5, 1113-1126.
13, 1741-1751.
133- 141.

510 Lierheimer et al. ( E m J. Biochem. 243)
matic amplification of DNA with a thermostable DNA polymerase, Science 239, 487-491.
Shak, S., Davitz, M. A, , Wolinsky, M. L., Nussenzweig, V., Turner, M. J. & Gurnett, A. (1 988) Partial characterization of the cross-reacting determinant, carbohydrate epitope shared by decay acceleration factor and the variant surface protein of the 31 african fiypanosoma buucei, J. lmrnuuol. 140. 2046-2050.
Sonderegger, P., Lemkin, P. E, Lipkin, L. E. & Nelson, P. G. (1985) Differential modulation of the expression of axonal proteins by non- neuronal cells of the peripheral and central nervous system, EMBO J, 4, 1395-1401.
Sonderegger, P. & Rathjen, F, G. (1992) Regulation of axonal growth in the vertebrate nervous system by interactions between glycoproteins belonging to two subgroups of the immunoglobulin superfamily, J. Cell Biol. 119, 1387-1394.
Stieger, S. & Brodbeck, U. (1988) Assay and purification of PI-specific phospholipase C from Bacillus cereus using commercially available phospholipase C. in Post-trurz.slation~11 modification of proteins by 1ipid.y (Brodbeck, U. & Bordier, C., eds) pp. 34-39, Springer-Verlag, Berlin.
Stieger, S., Diem, S., Jakob. A. & Brodbeck, U. (1991) Enzymatic prop- erties of phosphatidylinositol-glycan-specific phospholipase C from rat liver and phosphatidylinositol-glycan-specific phospholipase D from rat serum, Eur: J . Aiochem. 197, 67-73.
Stoeckli, E. T.. Lemkin, P. F., Kuhn, T. B., Ruegg, M. A,, Heller, M. M. & Sonderegger, P. ( 1 989) Identification of proteins secreted from axons of embryonic dorsal root ganglia neurons, E m J. Biochem.
Stoeckli, E. T., Kuhn, T. B., Duc, C. O., Ruegg, M. A. & Sonderegger, P. (1991) The axonally secreted protein axonin-1 is a potent substra- tum for ncurite growth, J. Cell Biol. 112, 449-455.
180, 249-258.
Stoeckli, E. T. & Landmesser, L. T. (1995) Axonin-I, Nr-CAM, and Ng- CAM play different roles in the in vivo guidance of chick commis- sural neurons, Neuron 14, 1165-1179.
Suter, D. M., Pollerberg, G. E., Buchstaller, A,, Giger, R. J., Dreyer. W. J. & Sonderegger, P. (1995) Binding between the neural cell adhesion molecules axonin-1 and NrCAM/Bravo is involved in neuron-glia interaction, J . Cell. Biol. 131, 1067-1081.
Takeichi. M. (1991) Cadherin cell adhesion receptors as a morphogenetic regulator, Science 251, 1451 -1455.
Tessier-Lavigne, M. (1994) Axon guidance by diffusible repellants and attractants, Curr: Opin. Gen. Dev. 4, 596-601.
Traunecker. A,, Oliveri, E & Karjalainen, K. (1991) Myeloma based expression system for production of large mammalian proteins, Trends Biotech. 9, 109-113.
Wolff, J. M, Briimmendorf, T. & Rathjen, F. G. (1989) Neural cell adhe- sion molecule F11 : membrane interaction by covalently attached phosphatidylinositol, Biochem. Biophys. Res. Commurz. 161, 931 - 938.
Yoshihara, Y., Kawasaki, M., Tani, A,, Tamada, A,, Nagata, S., Kagami- yama, H. & Mori, K. (1994) BIG-1 : a new TAG-I/F3-related mem- ber of the immunoglobulin superfamily with neurite outgrowth-pro- moting activity, Neuron 13, 415-426.
Yuckenberg, P. D., Witney, F., Geisselsoder. J. & McClary, J . (1991) Site-directed in vitro mutagenesis using uracil-containing DNA and phagemid vectors, in Directed nzutugerzesis: n pructicul upproach (McPherson, M. J., ed.) pp. 27-48, IRL Press, Oxford, UK.
Zuellig. R. A,, Rader, C., Schroeder, A., Kalousek, M. B., von Bohlen und Halbach. F., Osterwalder, T., Inan, C.. Stoeckli, E. T., Affolter, 1% U., Fritz, A,, Hafen, E. & Sonderegger, P. (1992) The axonally secreted cell adhesion molecule, axonin-1 : Primary structure, immu- noglobulin- and fibronectin-type-111-like domains, and glycosyl-pho- sphatidylinositol anchorage, Eur: 1. Biochem. 204. 453 -463.





























