Embed Size (px)
Citation preview

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/269098630
Thepresenceofneuropeptidesinthemammalianpinealandsubcommisuralorgan.
ARTICLEinNEUROSCIENCELETTERS·JANUARY1978
ImpactFactor:2.03
CITATIONS
2
READS
20
5AUTHORS,INCLUDING:
PaulPevet
UniversityofStrasbourg
814PUBLICATIONS17,963CITATIONS
SEEPROFILE
RuudBuijs
UniversidadNacionalAutóno…
431PUBLICATIONS20,614CITATIONS
SEEPROFILE
Availablefrom:PaulPevet
Retrievedon:14January2016

Cell Tiss. Res., 153, 277--292 (1974) �9 by Springer-Verlag 1974
The Pineal Gland of the Mole (Talpa europaea L.) I. The F ine S t ruc tu re of the P inea locy tes
Paul Pevet*
The Netherlands Central Institute for Brain Research, Amsterdam, The Netherlands (Dir. Prof. Dr. J. Arifins Kappers) - Laboratoire de Physiologic Compar6e, Poitiers, France (Dir. Prof. Dr. A. Peyre) - Service Gen6ral de Microscopie Electronique, appliqu6e k la Biologic,
Poitiers, France (Dir. Prof. Dr. J. Schrevel)
Received May 20, 1974
Summary. The ultrastructure of the mole pinealocytes, a mammal which lives practically in complete darkness, has been examined and compared with that of other mammals. Mite- chondria, ribosomes, smooth and granular endoplasmic reticulum, lysosomes and lipid inclu- clusions are present in the perikaryon. The presence of a paracrystalline structure of a possibly proteinaceous nature in some cisterns of the granular endoplasmic reticulum and between the two layers of the nuclear membrane, is characteristic of the mole pinealocyte. The Golgi complex produces clear vesicles of 500-1500 A in diameter. Occasionally, some dense core secretory vesicles were observed in the perikaryon and in the ending of cell processes. Their presumed origin from the Golgi complex could not yet be demonstrated. A large number of ciliary derivatives (9+0 pattern) are also present in the mole pinealocyte.
Key words: Ultrastructure - - Pineal Gland - - Mole - - Paracrystalline structures.
Rdsumd. Les pindalocytes (au sens strict: Wolfe, 1965) de l'6piphyse de la Taupe (animal vivant pratiquement toujours dans une complbte obscurit6) examines au microscope 61ectro- nique, ont dt6 eompar6s ~ ceux d'autres Mammifbres. Dans le pdrikaryon des mitochondries, des ribosomes, du r6ticulum endoplasmique lisse et granulaire et quelques lysosomes et in- clusions lipidiques sont prdsents. La pr6sence k l'intdrieur de certaines eavitds du rdticulum endoplasmique granulaire et parfois entre les deux feuillets de l'enveloppe nucldaire, de struc- tures paracristallines (de nature protdique .9) est caract6ristique des pin6alocytes de cet animal. L'appareil de Golgi s6cr6te des vdsicules claires de 500 ~ 1500/~ de diambtre. Quelques tr6s rares grains de s6cr6tion, dont l'origine golgienne n'a pas encore 6t6 ddmontr6, ont 6t6 observ6 dans le pdrikaryon et ~ l'extrdmit6 de certains prolongements. Un grand hombre de structures ciliaires (9+0 paires de tubules) ont 6galement 6t6 observds dans les pin6alocytes.
Introduction
There is much evidence t ha t the pineal gland exerts a significant influence upon reproduct ion and tha t this influence depends, at least part ly, on photo- periodic control (for a review of the l i terature, see W u r t m a n , Axelrod and Kelly, 1968). Most invest igat ions on m a m m a l i a n pineal u l t ras t ruc ture have been per-
Send o//print requests to: Dr. P. Pevet, Netherlands Central Institute for Brain Research, Ijdijk 28, Amsterdam-O, Holland. * Acknowledgements. In gratitude and with admiration this paper is dedicated to Professor Dr. J. Ari~ns Kappers and to Professor Dr. J.-P. Collin who, in permitting me to work without material cares, enabled me to perform this investigation. The author wishes to thank Dr. A. R. Smith, Dr. H. J. Romijn and Prof. Dr. J. Arifins Kappers for their help in the present work and for the revision of the paper, and Miss P. RSring for typing the manuscript.
19 Calc. Tiss. Res., Vol. 153

278 P. Pevet
formed in common laboratory animals which are usually reared under controlled conditions. A few studies of the pineal gland have dealt with species subjected to natural environmental conditions (hedgehog - Pevet, 1972, 1974; Pevet and Saboureau, 1973; pocket gopher - Sheridan and Reiter, 1973; macaque - Bererhi and Abbas-Terki, 1970; antarct ic seal - Cuello, 1973). I n view of the fact tha t the pineal gland inhibits the reproductive system during prolonged periods of darkness (Reiter, 1967, 1968, 1969) and controls the sexual cycle of mammals (Reiter, 1973), it seems also of interest to examine the gland of wild mammals living under natural lighting conditions and showing a characteristic sexual cycle. I n earlier investi- gations (Pevet, 1972, 1974; Pevet and Saboureau, 1972, 1973), the pineal gland of the hedgehog (Erinaceus europaeus L., Insectivora) was studied. The present report describes the fine structure of the mole pineal (Talpa europaea L., Insecti- vora), another lower mammal belonging to the same family. Like the hedgehog, the mole has a well-known sexual cycle (Courrier, 1927; Peyre, 1968). While, moreover, the mole lives practically always in complete darkness, it seemed of special importance to investigate its pineal structure, innervat ion and function. The present paper is concerned with the mole's pinealocyte structure only.
Material and Methods
All 20 moles, both females and males, used in this study were captured in Deux-S~vres (France) during January-June 1972. Immediately after capture the animals were transported to the laboratory 1. They were decapitated under ether anesthesia 3 hours after capture. The skull was opened and the region of the pineal flooded with cold fixative (glutaraldehyde 2.5 % - phosphate buffer 0.1 M, pH 7.25, at 4 ~ The time elapsed between decapitation and flooding of the gland was less than one minute. After its removal, the gland was placed in the same fixative at a temperature of 4 ~ for i to 2 hours. After washing the organs in phosphate buffer during 20 miniutes, they were placed in a postfixative consisting of OsO 4 1% and phosphate buffer 0.2 M (pH of 7.25), at room temperature during one hour. The glands were then dehyd- rated and embedded in araldite (Glauert and Glauert, 1958).
Thin sections were cut on a LKB ultramicrotome with a glass knife, made with a Knife Maker LKB. The sections were stained with uranyl acetate and lead citrate (Reynolds, 1963; Venable and Coggeschall, 1965). For the observations a Philips 200 electron microscope was used.
Results
Perikaryon. Mole pinealocytes are irregular in shape and processes emerge from their cell bodies (Fig. 2). Profiles of both longitudinal and cross sections of such processes are observed in the vicinity of the perivascular space.
The nucleus is large and lobulated (Fig. 1). Oval and polygonal nuclei were, however, also observed. Chromatine is widely dispersed in the nucleoplasma. Only few chromatine concentrat ions are located in a zone adjoining the nuclear envelope (Fig. 1). The nucleolus is large and composed of an electron-dense irregular nucleoma (Fig. 13). These features helped to distinguish the nuclei of pinealocytes from those of other pineal elements. The nuclear envelope, perforated at intervals
I The moles were sacrificed and their pineal glands fixed and embedded in the Laboratoire de Physiologic Compar6e et S.G.M.E.A.B., Universit6 de Poitiers, France. The sections were cut and examined in the Netherlands Central Institute for Brain Research, Amsterdam, The Netherlands
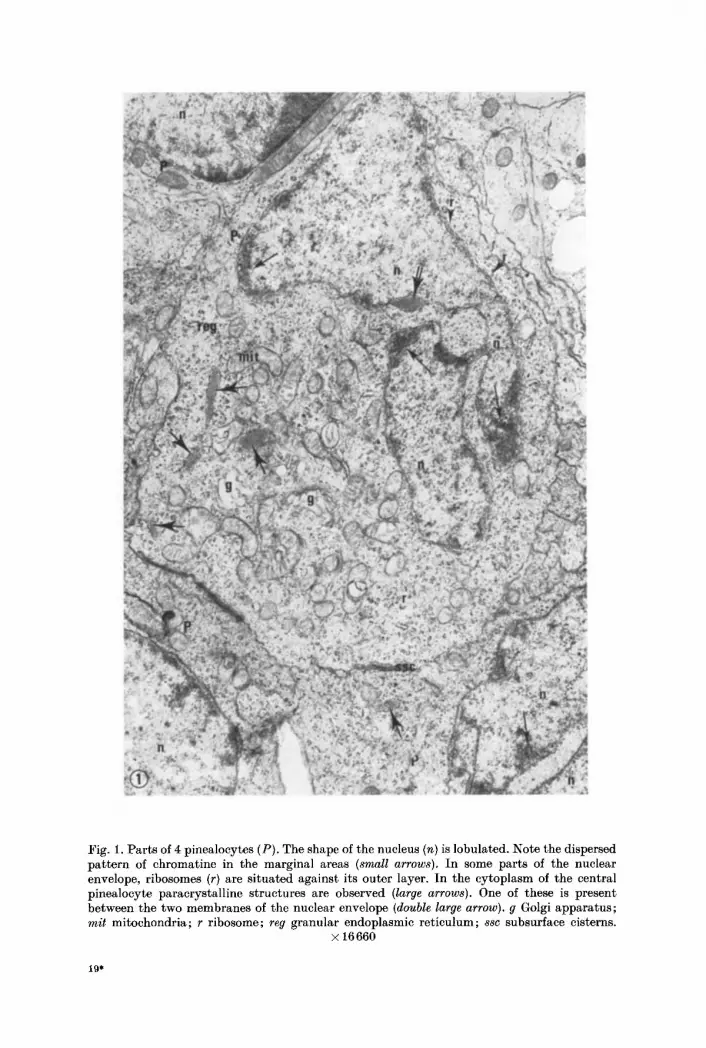
Fig. 1. Par ts of 4 pinealocytes (P). The shape of the nucleus (n) is lobulated. Note the dispersed pa t te rn of chromatine in the marginal areas (small arrows). In some parts of the nuclear envelope, ribosomes (r) are si tuated against its outer layer. In the cytoplasm of the central pinealoeyte paracrystall ine structures are observed (large arrows). One of these is present between the two membranes of the nuclear envelope (double large arrow), g Golgi apparatus; mit mitochondria; r ribosome; reg granular endoplasmic ret iculum; ssc subsurface cisterns.
• 16660
19"

280 P. Pevet
Fig. 2. Pa r t of the perikaryon of a pinealoeyte (P) and the proximal par t of a process which is characterized by microtubules (t) oriented parallel to its axis. Microtubules (arrows) are also seen in the cytoplasm of the cell body. mit mitochondria; r ribosomes; reg granular endoplasmic
reticulum. •
Fig. 3. Concentric laminae of membrane-bound cisterns observed in some pinealocytic processes •

Structure of Mole Pinealocytes. I 281
Fig. 4. Golgi apparatus (g) which appears to be very active. Many clear vesicles (arrows) are associated with the Golgi sacs. • 30000
Fig. 5. Transverse section of ciliary derivative in a pinealocyte, containing9~0pairsoftubules. • 56000
Fig. 6. Golgi apparatus (g) in the non-active phase. Some small clear vesicles are seen (arrows). • 30000
Fig. 7. Ciliary structure (c) in pinealocyte, mit mitochondria; n nucleus; pcs paracrystalline structure. X 13 000
Fig. 8. Smooth endoplasmic reticulum showing a peculiar shape (*). I t constitutes a system of twelve very long and flattened cisterns. This system is in direct topographical relationship
with the membrane of the nucleus (n). x 24125
Fig. 9. Smooth endoplasmic reticulum of the same shape as shown in Fig. 8 (*) in the cyto- plasm of a pinealocyte, mit mitochondria; pcs paracrystalline structure. • 47 060

282 P. Pevet

Structure of Mole Pinealocytes. I 283
by pores, consists of two electron-dense membranes. I n some parts of this nuclear envelope, m a n y ribosomes are lying against the outer membrane (Figs. 1, 15).
A prominent Golgi complex is present and usually widely dispersed around the nucleus of the pinealocyte. Each Golgi apparatus consists of a system of f lat tened sacs which is associated with a populat ion of vesicles of varying diameter (Figs. 4, 13). I n some pinealocytes the Golgi appara tus is very small (Fig. 6), but in most it is large. Here, the sacs are long and lucent vesicles are often observed in asso- ciation with them (Fig. 4). Golgi complexes associated with or free of these vesicles were practically never found together in one pinealocyte.
I n 3 of the 20 moles examined we observed some rare but very characteristic secretory (granular) vesicles measuring 800-2000 A in diameter (Figs. 10-12).
Numerous relatively large elongated or spherical mitochondria are found within the cytoplasm; they are bound by two unit membranes (Figs. 1, 2, 19). Membrane-bound dense bodies, presumably lysosomes (Fig. 13), show a single membrane and a granular matrix. Lipid droplets are sparse.
Smooth endoplasmic reticulum is very rare. If present, it often shows a peculiar shape (Figs. 8, 9) and is consti tuted by some very flat cisterns. Quite often this ret iculum appears to be connected with the nuclear envelope (Fig. 8).
Single ribosomes and polysomes are scattered throughout the cytoplasm (Figs. 1, 2, 10, 13). Par t of the ribosomes are associated with the double cyto- plasmic membrane (Figs. l, 2, 13) and with the outer layer of the nuclear envelope (Figs. 1, 15) to form the rough surfaced or granular endoplasmic reticulum which is abundan t in the pinealocytes.
I n the cytoplasm (Figs. l, 7, 15, 17-18, 20-21) and also between the two layers of the nuclear envelope (Figs. 1, 18-19, 22) of some pinealocytes a peculiar para- crystalline s tructure was observed, si tuated in cisterns of the granular endoplasmic reticulum. If present within the nuclear envelope, m a n y ribosomes were associated with the outer layer of this envelope.
These paracrystal l ine structures were never observed to lie freely in the cyto- plasm. Generally, they show the shape of a lozenge (Figs. 15, 18). Their length is 0.5-1.5 ~, their width 0.15-0.3 ~z. Format ions having this part icular shape show a
Fig. 10. Pinealocyte (P). In the cytoplasm there is one typical secretory granule (gs), and possibly another one (arrow). n nucleus; r ribosomes, x 9 500
Fig. 11. Ending of pinealocyte process. Many clear vesicles (v) and also one secretory granule (arrow) are present, mit mitochondria, x 19700
Fig. 12. Secretory granule. Between the dense matrix and the membrane there is always a clear halo (arrow). x 78000
Fig. 13. Part of epiphyseal tissue. In the contact area between the central pinealocyte (P) and the other pinealocytes (P) there are four subsurface cisterns (2 to 2) (arrows). g Golgi apparatus;
lys lysosomal structure; mit mitochandria; n nucleus; reg granular endoplasmic reticulum
Fig. 14. Subsurface cistern complex which consists of two cisterns (*) present in two pinealo- cytes and facing each other. Some ribosomes (arrows) are situated on their membranes facing the interior of the cell. Between the two cisterns, the plasma membranes of the two cells are
observed (big arrows), x 87 800

284 P. Pevet
Fig. 15. Paracrystalline structure (pcs) in a pinealocyte. The structure lies in a cistern of the granular endoplasmic reticulum. I t has a characteristic lozenge shape and its paracrystalline aspect is very obvious. Ribosomes (small arrows) are situated against the membrane of the reticular cistern and the outer layer of the nuclear envelope (large arrows), g Golgi apparatus;
n nucleus; r ribosomes. • 27 680
Fig. 16. Another paracrystalline structure (pcs) which has not a lozenge shape, n nucleus; r ribosomes. • 43 920

Structure of Mole Pinealocytes. I 285
clear paracrystalline structure (Figs. 15-18) consisting of many parallel striae which are situated perpendicularly to the long axis of the formation and associated in pairs (Fig. 17). The width between two associated striae is 120-140/~, while tha t between the pairs is 200-300 A. Many of these formations, whose paracrystalline structure was scarcely distinct, have also been observed (Figs. 19-22). In these cases they were generally spherical in shape, measuring 0.5-1.5 ~x in diameter.
So far, it has been impossible to determine whether the different types of paracrystalline formations present were either identical, but sectioned in different planes, or identical, but representing different developmental stages, or, indeed, quite different formations.
Ciliated structures (Fig. 7) were found in many pinealocytes. When seen in cross-sections they exhibit a 9~-0 arrangement of pairs of tubules (Fig. 5). The diameter of the cilia is 0.2-0.3 ~z while that of the tubules is 150-170 A.
In numerous pinealocytes subsurface cisterns are present. They are situated very close to the plasmalemma and found in the perikarya exclusively, never in the processes of pinealocytes. In contrast to subsurface cisterns present in rat pinealocytes (Wolfe, 1965), those in mole pinealocytes are rarely entirely devoid of ribosomes which are situated on that membrane of the cistern which faces the interior of the cell body. Ribosomes on the part of the cistern membrane facing the plasmalemma are, however, lacking. Rather often, two subsurface cisterns were seen facing each other, being present in adjacent pincalocytes.
Cell processes. The processes of mole pinealocytes are very long. Some organel- les, in particular mitochondria, were observed in their cytoplasm (Fig. 2). Further- more, they contain many microtubules with a diameter of approximately 200- 250 A which run parallel to the long axis of the processes (Fig. 2). These micro- tubules could also be observed in the cytoplasm of the cell body (Fig. 2), but there they did not show any special and constant distribution. Occasionally, concentric laminae of membrane-bound cisterns can be observed in the processes (Fig. 3).
The endings of the mole pinealocyte processes are club-shaped. They contain numerous clear vesicles varying from 300-350/~ in diameter. In a few rare cases endings of the processes show one or more secretory granular vesicles (Fig. 11).
Discussion
The pinealocyte of the mole, like that of other mammals (Kappers, 1969; Karasek, 1971), can be considered the characteristic cell element of the pineal gland. The problem of the presence of light and dark pinealocytes also exists in the mole. This difference has been earlier observed in the pineal gland of the rabbit (Wartenberg and Gusek, 1965; Romijn, 1972), rat (Gusek et al., 1965), hamster (Sheridan and Reiter, 1968, 1970), pocket gopher (Sheridan and Reiter, 1973) and
Fig. 17. Larger magnification of the paracrystalline structure illustrated in Fig. 16. Its crystalline organization (pcs) which is made up of parallel striae associated in pairs (*) is
clearly seen. • 79 800
Fig. 18. Paracrystalline structured (pcs) between the two layers of the nuclear envelope (arrows) In the cytoplasm another paracrystalline structure (pcs). g Golgi apparatus; n nucleus;
r ribosomes. • 31900

286 P. Pevet
Fig. 19. Paracrystall ine structures (pcs) whose crystalline aspect is not very apparent . They are si tuated between the two membranes of the nuclear envelope. Many ribosomes are present on the outer layer (arrows). g golgi apparatus mit mitochondria; r ribosomes; reg granular
endoplasmic reticulum. •

Structure of Mole Pinealocytes. I 287
hedgehog (Pevet, 1972, 1974; Pevet and Saboureau, 1973). In the pineal gland of the hedgehog, we could clearly demonstrate that the number of dark and light pinealocytes depends on the fixative used. In the mole, dark and light pinealocytes were also observed, but, having used a single fixative only, it has not been possible to determine whether the cytological picture of the mole pinealocyte varies also with different types of fixatives. The ultrastructure of the two cell types was not basically different. I t is likely that the light and dark pinealocytes in the mole and also in the hedgehog pineal gland reflect differences in functional stages. According to Romijn (1972), differences in content and location between dark and light rabbit pinealocytes may occur.
I t is now generally accepted that the mammalian pineal is a glandular organ and that the pinealocytes are involved in the synthesis of pineal secretory products (Kappers, 1969). Cytological proofs of a secretory process have been looked for in the mole pineal. In this respect, the presence of two different aspects of the Golgi apparatus is interesting. In fact, the well-developed, large Golgi apparatus pro- bably represents an active functional phase of this organelle while a small Golgi apparatus points to a state of lesser activity. Although a Golgi apparatus in its non-active phase could be possibly confused with a transversel section of a Golgi apparatus in an active phase, the actual existence of these two phases of the Golgi apparatus is most probable because they were practically never observed in the same pinealocyte. Moreover, the ratio between the number of active and non- active Golgi complexes is the same in the pineal of all moles killed during the same period of the year; this ratio seems to present a functional phase of the gland during that special part of the year, and it may change at different periods.
Clear vesicles, measuring 500-1000 A in diameter, have been observed in association with the Golgi apparatus in the cytoplasm of the cell body. They are evidently produced by this structure. Clear vesicles have also been found in the pinealocytes of several other mammals such as rat (Wolfe, 1965), cattle (Anderson, 1965), dog (Sano and Mashimo, 1966), hamster (Sheridan and Reiter, 1968, 1970), mouse (Pellegrino de Iraldi, 1969), rabbit (Romijn, 1972), pocket gopher (Sheridan and Reiter, 1973)and hedgehog (Pevet, 1972, 1974; Pevet and Saboureau, 1973). The functional role and the biochemical nature of these vesicles are not known.
Secretory vesicles containing a dense core, which were observed in the pinealo-
Fig. 20. In the cytoplasm of one pinealocyte, three paracrystalline structures (pcs) are seen. Their crystalline aspect is not very apparent. The shape of these structures is not that of a lozenge and they are rather large. Within one of these structures one can discern a crystalline
organization (arrows). • 24100
Fig. 21. In the cytoplasm of a pinealocyte, four paracrystalline structures (pcs), the crystalline aspect of which is not very apparent, are present. Within two of them it is possible to observe crystalline organization (arrows). In a noradrenergic ending (NA), many small vesicles (v), some of them having a dense core, are present. In others, also a large granular vesicle can be seen (small arrow), g Golgi apparatus; r ribosomes; reg granular endoplasmic reticulum.
• 25250
Fig. 22. In this pinealocyte a large number of paracrystalline structures (arrows) are observed between the two membranes of the nuclear envelope, n nucleus. • 22960

288 P. Pevet
cytes of all other mammalian species studied and also in all principal epiphyseal cells of lower vertebrates (Collin and Meiniel, 1972), have also been found in mole pinealocytes. In this animal, however, they are very scarce. We never observed a secretory granular vesicle in association with a Golgi apparatus; therefore, it is difficult to decide whether they are produced by the Golgi complex. Because secretory vesicles are so scarce, a secretory polarity (Collin, 1969) could not be demonstrated in the mole pinealocyte. I t is likely that such a secretory polarity exists, because some secretory granular vesicles were observed in the terminal buds of pinealocyte processes.
The scarcity of secretory granular vesicles is characteristic - in our study - of the pinealocytes of moles captured from January to June. I t is quite possible that during another part of the year the production of these granules is increased, perhaps in relation with the sexual cycle. An investigation, now in progress, of the mole pinealocyte during the course of the entire sexual cycle will perhaps permit us to verify this hypothesis.
The presence of paracrystalline inclusions has been revealed in many types of cells (Fawcett, 1967). They have been reported to occur in nearly all compart- ments of the cell, i.e., in the nucleus, mitochondria, Golgi complex, secretory granules, endoplasmic reticulum and free in the cytoplasm (Fawcett, 1967), but, so far, they were never observed in pinealocytes. These formations have usually been assumed to consist of protein, but in a few cases only the specific protein has been identified (for more details, see Fawcett, 1967). The chemical nature and the physiological significance of the majority of the crystals described are not known. In the mole, these paracrystalline formations are always located in the granular endoplasmic reticulum. Although their biochemical composition could not be determined, their presence may point to an important function, probably protein synthesis. Paracrystalline formations could be caused by an accumulation of compounds due to an excessive synthesis or by preventing the diffusion of com- pound(s) produced by the endoplasmic reticulum. Possibly, a relation exists between the large number of paracrystalline formations present in the mole pinealocytes and the extreme scarcity of secretory granular vesicles. Both features seem to be very characteristic of the mole pineal gland.
The peculiar packets of smooth endoplasmic reticulum somewhat resemble the annulate lamellae earlier observed by Kessel (1968). The structure seems to be closely associated with the nuclear envelope (Figs. 8), being rarely situated free in the cytoplasm (Fig. 9). Kessel (1968) mentioned that the annulate lamellae, situated close to the nuclear membrane, originate by differentiation of the nuclear envelope and that they migrate into other regions of the cytoplasm. So far, it is impossible to confirm that the structures described in the present paper are homologous with the annulate lamcllae of Kessel. In pinealocytes of other mammals a similar structure has never been described. At present the function of this peculiar formation of the smooth reticulum is unknown.
In the mole pineal gland, a large number of subsurface cisterns were observed in pinealocytes touching the inner layer of the plasmalemma. These structures have been described in many cells (Rosenbluth, 1962). They have also been found by Wolfe (1965) in the pinealocytes of the rat and by Cuello (1973) in those of the antarctic seal. In pinealocytes of the field mouse and of the hedgehog these struc-

Structure of Mole Pinealocytes. I 289
tures were also observed by the present author (unpublished). When facing each other in two adjacent pinealocytes, they may establish a pathway for the exchange of metabolites or of ions between these cells.
Like the pinealocytes of many other mammals, the pinealocytes of the mole are characterized by the piesence of one or several processes differing in length and mostly showing a club-shaped ending (Kappers, 1969). In the mole these endings contain a large number of small clear vesicles measuring 300400 A in diameter. Due to the scarcity of secretory granular vesicles in the mole pinealo- cyte, which are much more common in the pinealocytes of other mammals, the endings of the pinealocyte processes in the mole pineal gland are not easily identi- fied. When granular vesicles are totally absent in them, they may be confused with the terminals of cholinergic nerve fibres because both (the endings of the pinealocyte processes and the nerve terminals) contain lucent vesicles of the same diameter. The presence of pineal cholinergic nerve terminals in the mole cannot be excluded because their presence has been surmised by some authors in other mammals, while they were clearly demonstrated in the rabbit pineal organ by Romijn (1972, 1973). In some pinealocyte processes few secretory vesicles were present; their endings could be identified beyond doubt. In contrast to the hedgehog (Pevet, 1974), terminal buds of pinealocyte processes in the mole cannot be confused with pineal noradrenergic nerve endings because the latter contain many small vesicles showing a dense core (c]. Taxi and Droz, 1966), also in the mole pineal (:Fig. 21), while in the hedgehog lucent vesicles dominate.
Some concentric laminar formations, similar to the "systbme r@ticulaire, stade V" as observed in the male hedgehog pinealocyte processes during the period of sexual quiescence (Pevet, 1972, 1974; Pevet and Saboureau, 1972, 1973), were likewise found in processes of mole pinealocytes. Such formations have also been described in the pineal of several other mammalian species and of lower verte- brates (see Pevet and Saboureau, 1973, for references). In the mole we never observed the earlier stages of development of these concentric laminar formations, but their development has earlier been demonstrated in the hedgehog pinealocytes during the period of sexual quiescence (Pevet, 1972, 1974; Pevet and Saboureau, 1973). All moles investigated in the present study were captured during the first part of the year (1972, January-June) (for information on the mole sexual cycle, see Courrier, 1927, and Peyre, 1968). As yet, it has not been possible to establish any functional relationship between the concentric laminar system observed in the mole pineal and t h e " syst@me r@ticulaire stade V" observed in the hedgehog pineal.
Many mole pinealoeytes are characterized by the presence of ciliary derivatives. At the ultrastructural level all motile cilia so far known exhibit nine pairs of peripheral and two central tubules (9+2). The shafts or connecting pieces of retinal rods (De Robertis, 1956; Tokuyasu and Yamada, 1959) and cones (Ueno, 1961) and of pineal photoreceptor cell outer segments (Collin, 1969, 1971) belong to the 9 + 0 type. Several investigators (Barnes 1961; Sorokin, 1962) have sug- gested that these 9 + 0 cilia have a sensory function. The ciliary derivatives ob- served in the present study are characterized by a 9 + 0 tubular pattern. Ciliary structures have also been observed in pinealocytes of other mammals, such as rat (Wolfe, 1965; Karasek, 1970), cattle (Anderson, 1965), hedgehog (Pevet, 1972, 1974; Pevet and Saboureau, 1973) ane hamster (Clabough, 1971), but in these

290 P. Pevet
animals the i r in terna l s t ruc ture has not been described. F r o m the presence of c i l iary de r iva t ives in fe ta l r a t and hams te r Clabough (1973) concluded t h a t the p ineal g land conta ins cells der ived from the photoreceptor -ce l l line supposed to be presen t in the m a m m a l i a n pineal g land b y Collin (1969, 1971). I n cont ras t to the p ineal g land of o ther adu l t mammals , in the mole a large q u a n t i t y of ci l iary deriv- a t ives occurs. I t is l ike ly t h a t these ci l iary der iva t ives arc phylogenet ic relics. Dur ing the phylogenet ic deve lopmen t of ve r t eb ra t e s a g radua l reduc t ion of the d i rec t pho torecep t ive funct ion of the p inea l organ occurs, and the neurosensory photorecept ive e lements become r u d i m e n t a r y (Collin, 1969, 1971). I t should, however, be men t ioned t h a t c i l iary der iva t ives also occur in quite different cells of ec todermal origin, for instance, in nerve cells.
I n the p resen t inves t iga t ion i t has been shown t h a t the u l t r a s t ruc tu ra l aspect of the mole p inea locyte differs somewhat from t h a t observed in the p inea locytes of o ther mammal s so far examined . As the moles p rac t ica l ly live in pe rmanen t darkness , and as i t is well known tha t the q u a n t i t y of env i ronmenta l l ighting exer ts an impor t an t influence on the s t ruc ture and funct ion of the p ineal organ (see W u r t m a n , Axc l rod and Kel ly , 1968; Kappers , 1969), i t is t e m p t i n g to cor- re la te the par t i cu la r ecological s i tua t ion of these animals wi th the differences found in the u l t r a s t ruc tu re of the i r p inealocytes . The produc t ion of paracrys ta l l ine format ions was never observed before in any m a m m a l i a n pineal gland.
References Anderson, E. : The anatomy of bovine and ovine pineals. Light and electron microscopic studies.
J. Ultrastruct. Res., Suppl. 8, 1-80 (1965) Barnes, B. G. : Ciliated secretory cells in the pars distalis of the mouse hypophysis. J. Ultra-
struct. Res., Suppl. 5, 453-467 (1961) Bererhi, A., Abbas-Terki, M.: Structure fine de l'~piphyse du Magot d'Alg~rie (Macacus
sylvanus L.) Bull. Ass. Anat. 148, 285-294 (1970) Clabough, J. W.: Ultrastructural features of the pineal gland in normal and light deprived
golden hamsters. Z. Zellforseh. 114, 151-164 (1971) Clabough, J. W. : Cytological aspects of pineal development in rats and hamsters. Amer. J.
Anat. 135, 215-230 (1973) Collin, J. P. : Contribution ~ l'~tude de l'organe pineal. De l'~piphyse sensorielle ~ la glande
pin~ale: modalit~s de transformation et implications fonctionelles. Ann. Star. Biol. de Besse-en-Chandesse, Suppl. l , 1-359 (1969)
Collin, J. P.: Differentiation and regression of the cells of the sensory line in the epiphysis cerebri. In: The Pineal Gland. CIBA Foundation Symposium. London 1970. Eds. G.E.W. Wolstenholme and J. Knight, p. 79-125. Churchill Livingstone 1971
Collin, J. P., Meiniel, A. : L'organe pineal. Etudes combin~es ultrastructurales, cytochimiques (monoamines) et exp~rimentales, chez Testudo mauritanica: grains denses des cellules de la lign~e ,,sensorielle" chez les Vertebras. Arch. Anat. micr. Morph. exp. 60, 269-304 (1972)
Courrier, 1%. : Etude sur le d~terminisme des caract+res sexuels secondaires chez quelques Mammif+res k l'activit~ testiculaire p~riodique. Arch. Biol. (Liege) 37, 173-334 (1927)
Cuello, A.C.: Ultrastructural characteristics and innervation of the pineal organ of the antarctic seal (Leptonychotes weddelli). J. Morph. 141, 218-226 (1973)
De Robertis, E. : Morphogenesis of the retinal rods. An electron microscope study. J. biophys. biochem. Cytol., Suppl. 2, 209-218 (1956)
Fawcett, D. W. : An atlas of fine structure. The Cell. Philadelphia: W. B. Saunders & Co. 1967 Glauert, A. M., Glauert, R. H. : Araldite as an embedding medium for electron microscopy.
J. biophys, biochem. Cytol. 4, 191-194 (1958) Gusek, W., Buss, H., Wartenberg, H. : Weitere Untersuchungen zur Feinstruktur der Epiphysis
cerebri normaler und vorbehandelter Ratten. Progr. Brain Res. 10, 317-330 (1965)

Structure of Mole Pinealocytes. I 291
Kappers, J. Ari~ns: The mammalian pineal organ. J. neuro-visc. Rel., Suppl. 9, 140-184 (1969) Karasek, M. : The cilia in the white rat pineal gland. J. Microsc. 9, 1103-1104 (1970) Karasek, M. : The role of the pineal body in mammals. Polish Endocrinol. 22, 315-327 (1971) Kessel, R. G. : Annulate lamellae. J. Ultrastruct. Res., Suppl. 10, 1-83 (1968) Pevet, P. : Etude ultrastrueturale de l'~piphyse du H~risson male. Evolution en fonction du
cycle sexuel. Th~se I I Ie cycle. Universit~ de Poitiers (1972) Pevet, P. : Etude structurale et ultrastructurale de l'~piphyse du tt~risson male (Erinaceus
europaeus L.). Evolution en fonction du cycle sexuel. Vme entretiens de Chiz~. Probl~mes endocriniens chez les Mammif~res sauvages - Aspects M~taboliques et Ecophysiologiques, 11-12-13-octobre 1973 (in press) (1974)
Pevet, P., Saboureau, M. : Modifications ultrastructurales dans l'~piphyse du H~risson m~le au cours du repos sexuel. Congr~s de la Soc. Europ. d'endocr. Comp. Montpellier 1971. Gen. comp. Endocr. 18, 3 (1972)
Pevet, P., Saboureau, M. : L'~piphyse du H6risson (Erinaceus europaeus L.) male. 1) Les pin~a- locytes et leur variations ultrastructurales consid~r~es au cours du cycle sexuel. Z. Zell- forsch. 143, 367-385 (1973)
Pellegrino de Iraldi, A. : Granulated vesicles in the pineal gland of the mouse. Z. Zellforsch. 1Ol, 408-418 (1969)
Peyre, A. : Cycles g~nitaux et correlation hypophyso-g~nitales chez trois Insectivores Euro- p~ens. In : Cycles g6nitaux saisonniers de Mammif~res sauvages, p. 133-149. Paris: Masson& Cie. 1968
Reiter, R. J. : The effect of pineal grafts, pinealectomy and denervation of the pineal gland on the reproductive organs of male hamsters. Neuroendocrinol. 2, 138-146 (1967)
Reiter, R. J. : Morphological studies on the reproductive organs of blinded male hamsters and the effect of pinealectomy and superior cervical ganglionectomy. Anat. Rec. 160, 13-24 (1968)
Reiter, R. J. : Pineal function in long term blinded male and female golden hamsters. Gen. comp. Endocr. 12, 460468 (1969)
Reiter, R. J. : Pineal control of a seasonal reproductive rhythm in male golden hamsters exposed to natural daylight and temperature. Endocrinology 92, 2, 423430 (1973)
Reynolds, E. S. : The use of lead citrate at high p i t as an electron-opaque stain in electron microscopy. J. Cell Biol. 17, 208-212 (1963)
Romijn, H. J. : Structure and innervation of the pineal gland of the rabbit, Oryctolagus cuni- culus L., with some functional considerations. Th~se, Free University, Amsterdam (197 2)
Romijn, H. J. : Structure and innervation of the pineal gland of the rabbit, Oryctolagus cuni- culus L. I) A light microscopic investigation. Z. Zellforsch. 189, 473-485 (1973)
tr J. : Subsurface cisterns and their relationship to the neuronal plasma membrane, J. Cell Biol. 13, 405-421 (1962)
Sano, I., Mashimo, T. : Elektronenmikroskopische Untersuchungen an der Epiphysis cerebri beim Hund. Z. Zellforsch. 69, 129-139 (1966)
Sheridan, M. N., Reiter, R. J . : The fine structure of the hamster pineal gland. Amer. J. Anat. 122, 357-376 (1968)
Sheridan, M. N., Reiter, R. J. : Observations in the pineal system in the hamster. II) Fine structure of the deep pineal. J. Morph. 181, 163-177 (1970)
Sheridan, M. N., Reiter, R. J . : The fine structure of the pineal gland of the pocket gopher, Geomys bursarius. Amer. J. Anat. 136, 363-382 (1973)
Sorokin, S. : Centrioles and the formation of rudimentary cilia by fibroblast and smooth muscle cells. J. Cell Biol. 15, 363-377 (1962)
Taxi, J. , Droz, B.: Etude de l'incorporation de noradr~naline-3H (NA-aH) et de 5-hydroxy- tryptophane-aH (5-HTP-aH) darts les fibres nerveuses du canal @f6rent et de l'intcstin. C. R. Acad. Sci. (Paris) 263, 1237-1240 (1966)
Tokuyasu, K., Yamada, E. : The fine structure of the retina studied with the electron micro- scope. IV. Morphogenesis of outer segments of retinal rods. J. biophys, biochem. Cytol. 6, 225-230 (1959)
Ueno, K. : Morphogenesis of the retinal cone studied with the electron microscope. Jap. J. Ophthal. 5. 114 122 (1966)

292 P. Pevet
Venable, J. M., Coggeschall, W. : Simplified lead stain for use in electron microscopy. J. Cell Biol. 25, 407-408 (1965)
Wartenberg, It. : The mammalian pineal organ: electron microscopic studies on the fine structure of pinealocytes, glial cells and on the perivascular compartement. Z. Zellforsch. 86, 74-97 (1968)
Wartenberg, H., Gusek, W. : Licht- und elektronenmikroskopische Beobachtungen fiber die Struktur der Epiphysis cerebri des Kaninchens. Progr. Brain Res. 10, 296-316 (1965)
Wolfe, D. E. : The epiphyseal cell: an electron-microscopic study of its intercellular relationship an intracellular morphology in the pineal body of the albino rat. Progr. Brain Res. 10, 332-376 (1965)
Wurtmann, 1%. J. , Axelrod, J., Kelly, D. E. : The pineal. New-York-London: Academic Press 1968





























