Embed Size (px)
Citation preview

Life Sciences 87 (2010) 169–174
Contents lists available at ScienceDirect
Life Sciences
j ourna l homepage: www.e lsev ie r.com/ locate / l i fesc ie
Insulin temporal sensitivity and its signaling pathway in the rat pineal gland
Rodrigo A. Peliciari-Garcia a, Anderson C. Marçal a, Jéssica A. Silva a, Daniella Carmo-Buonfiglio a,Fernanda G. Amaral a, Solange Castro Afeche b, José Cipolla-Neto a, Carla R.O. Carvalho a,⁎a University of São Paulo, Institute of Biomedical Sciences-I, Department of Physiology and Biophysics, São Paulo, SP, Brazilb Laboratory of Pharmacology, Butantan Institute, São Paulo, SP, Brazil
⁎ Corresponding author. Tel.: +55 11 3091 7247; faxE-mail address: [email protected] (C.R.O. Carvalho).
0024-3205/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.lfs.2010.06.005
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 31 October 2009Accepted 9 June 2010Keywords:Pineal glandMelatonin synthesisInsulin actionArylalkylamine-N-acetyltransferase (AANAT)PI3K
Aims: In our previous work, we reported that the insulin potentiating effect on melatonin synthesis isregulated by a post-transcriptional mechanism. However, the major proteins of the insulin signalingpathway (ISP) and the possible pathway component recruited on the potentiating effect of insulin had notbeen characterized. A second question raised was whether windows of sensitivity to insulin exist in thepineal gland due to insulin rhythmic secretion pattern.Main methods: Melatonin content from norepinephrine(NE)-synchronized pineal gland cultures wasquantified by high performance liquid chromatography with electrochemical detection and arylalkyla-mine-N-acetyltransferase (AANAT) activity was assayed by radiometry. Immunoblotting and immunopre-cipitation techniques were performed to establish the ISP proteins expression and the formation of 14-3-3:
AANAT complex, respectively.Key findings: The temporal insulin susceptibility protocol revealed two periods of insulin potentiating effect,one at the beginning and another one at the end of the in vitro induced “night”. In some Timed-insulinStimulation (TSs), insulin also promoted a reduction on melatonin synthesis, showing its dual action incultured pineal glands. The major ISP components, such as IRβ, IGF-1R, IRS-1, IRS-2 and PI3K(p85), as welltyrosine phosphorylation of pp85 were characterized within pineal glands. Insulin is not involved in the 14-3-3:AANAT complex formation. The blockage of PI3K by LY 294002 reduced melatonin synthesis and AANATactivity.Significance: The present study demonstrated windows of differential insulin sensitivity, a functional ISP andthe PI3K-dependent insulin potentiating effect on NE-mediated melatonin synthesis, supporting thehypothesis of a crosstalk between noradrenergic and insulin pathways in the rat pineal gland.© 2010 Elsevier Inc. All rights reserved.
Introduction
The mammalian pineal gland synthesizes and releases thehormone melatonin during the dark period. Due to its physiologicalrhythmic production, melatonin is one of the most reliable internaltiming signals as far as circadian and seasonal rhythms are concerned(Afeche et al. 2008). The light/dark cycle, via retino-hypothalamicpathway entrains the master circadian clock, the suprachiasmaticnuclei (SCN), which control the nocturnal norepinephrine (NE)release from sympathetic nerve endings into pineal gland.
The NE interaction with β1 Gs-protein-coupled adrenoreceptorsactivates the cyclic AMP-dependent protein kinase (PKA), inducingthe phosphorylation of cAMP responsive element binding (CREB),which phosphorylated, drives the transcription and translation oftryptophan hydroxylase (TPOH) and arylalkylamine-N-acetyltrans-ferase (AANAT) (Simonneaux and Ribelayga 2003). AANAT controls
: +55 11 3091 7629.
l rights reserved.
the daily melatonin production and is considered the most importantenzyme on melatonin synthesis (Klein 2007). At the same time, NEinteracts with α1 Gq-protein-coupled adrenoreceptors and potenti-ates β1 signaling via protein kinase C (PKC), leading to an increase inintracellular PKA levels (Klein et al. 1983; Sugden et al. 1985). Theactivation of PKA also induces the formation of the 14-3-3:AANATcomplex, which protects AANAT against proteosomal proteolysis, thusassuring melatonin production (Klein 2007).
Among its actions, melatonin promotes the timing of central(Gillette andMitchell 2002) and peripheral metabolic functions (Limaet al. 1998; Picinato et al. 2002a; Alonso-Vale et al. 2006, 2008),playing an important role in energy metabolism, insulin action (Limaet al. 1994, Anhê et al. 2004) and secretion (Picinato et al. 2002a,b,2008), as well as in white adipose tissue function (Alonso-Vale et al.2004a,b, 2005).
As a counterpoint to these melatonin's actions on insulinsecretion/action, Garcia et al. (2008) reported that insulin acts inthe rat pineal gland potentiating NE-mediated melatonin synthesis,through the regulation of TPOH and AANAT activities. This pinealeffect of insulin seems to depend on post-transcriptional mechanisms

Fig. 1. Temporal insulin susceptibility protocol. Pineal gland cultures were norepi-nephrine (NE)-synchronized (12 h pattern of NE presence or absence — horizontalsuperior bar). On the final 12 h period of NE stimulation, considered as the in vitroinduced “night”, the glands were distributed into 12 groups. Each group receivedinsulin (Ins 10 nM) stimulation only for 1 h, called a Timed-insulin Stimulation (TS)(black squares). At the end of each TS, the glands were collected (black arrows) andfrozen on dry ice and kept at−80 °C until melatonin assay. The moment at each this TSoccurred varied along the 12 h period of NE stimulation, generating 12 windows ofpossible differential sensitivity to insulin presence.
170 R.A. Peliciari-Garcia et al. / Life Sciences 87 (2010) 169–174
since no differences in the enzymes gene expression were character-ized. The hypothesis that remained to be tested was whether insulinwould potentiate melatonin synthesis by inducing a further 14-3-3:AANAT complex formation leading to an increase on melatonincontent. Related to insulin signaling pathway (ISP) in the rat pinealgland, studies have demonstrated in cultured cells, under NEstimulation, the participation of p38mitogen-activated protein kinase(MAPK), p42/44 MAPK, as well as MAPKphosphatase-1 on themodulation of melatonin synthesis (Chik et al. 2004; Ho et al. 2006;Price et al. 2007).
However, as the major protein participants of the ISP, like insulinreceptor β subunit (IRβ), insulin growth factor-1 receptor (IGF-1R), IRsubstrate-1/2 (IRS-1/2) and the p85 subunit of phosphoinositide 3-kinase (PI3K) had not been characterized in the rat pineal gland, oneof the aims of this study was to investigate the ISP proteins expressionas well as the target protein responsible for the insulin potentiatingeffect on NE-stimulated melatonin synthesis observed in our previouswork. We also intended to investigate if any temporal susceptibilitywindows to insulin would exist in the pineal gland, determining inwhich moment of the NE-stimulated phase the potentiating insulineffect would happen.
Materials and methods
Animals
Two month-old male Wistar rats weighing 150–180 g wereobtained from the Institute of Biomedical Sciences-I, University ofSão Paulo, São Paulo, SP, Brazil. The animals were kept on a 12 h:12 hlight/dark cycle (lights on at 06:00 am), in a temperature controlledroom (21±2 °C), with food andwater ad libitum. Ethics approval wasgranted by the Committee of Ethics in Animal Experimentation of theInstitute of Biomedical Sciences, University of São Paulo, São Paulo, SP,Brazil.
Pineal gland cultures
The standard and NE-synchronized pineal gland cultures wereperformed as previously described (Garcia et al. 2008). Briefly, instandard cultures the glands did not receive any kind of treatment(only medium change every 24 h). After 48 h of culture, the glandswere collected and the total protein extractionwas performed. OnNE-synchronized cultures, the glands were extracted just at the beginningof the dark period of the light-dark cycle and immediately incubatedin a medium containing 1 μM of NE, maintaining the glands in arhythmic exposition to NE, similar to physiological stimulationnoticed in vivo. After 12 h of NE incubation, the mediumwas changedfor another one without NE and this alternation between media, 12 hwith or without NE, lasted for 48 h. After that, the glands were thendistributed into the following experimental groups: Control, NE[1 μM], NE+Ins [10 nM] and NE+Ins+LY 294002 (LY) [1 μM], aspecific inhibitor of PI3K activation (Wu et al. 2008). All drugs werepurchased from Sigma, St. Louis, MO, USA. At the end of theexperiments (5 h of stimulation) the glands were frozen on dry iceand kept at −80 °C or immediately processed according to eachexperimental protocol.
Temporal insulin susceptibility protocolIn the final 12 h period of NE stimulation (60 h total time),
considered as the in vitro induced “night”, the NE stimulated glandswere distributed into 12 groups (one-hour interval each). Thechallenged glands additionally received insulin [10 nM] stimulationduring only 1 h, called a Timed-insulin Stimulation (TS). The momentat each TS occurred varied along the 12 h period of NE stimulation,generating 12 windows where the temporal susceptibility to insulinpresence were investigated. At the end of each TS, the glands were
collected (Fig. 1— black arrows) and frozen on dry ice, kept at−80 °Cuntil melatonin assay.
AANAT activity
AANAT activity was measured by a radiometric assay (Garcia et al.2008; Deguchi and Axelrod, 1972; Parfitt et al. 1975). Briefly, 100 μL of0.1 M sodium phosphate buffer, pH 6.8, containing 40 mM tryptamineand [3H]- acetyl coenzyme A (2 mM, final specific activity=4 mCi/mmol) (Amersham Biosciences, Piscataway, NJ, USA) were added to amicrocentrifuge tube containing one gland kept at 4 °C. The glandswere sonicated and then incubated at 37 °C for 20 min. The reactionproduct 3[H] N-acetyltryptamine was extracted with chloroform(1 mL). Samples of 500 μL were evaporated in a scintillation vial andradioactivity was determined with a β scintillation fluid counter.

Fig. 2. Windows of sensitivities to insulin in the rat pineal gland. Melatonin contentfrom pineal glands submitted to the temporal insulin susceptibility protocol wasquantified by HPLC and the values were expressed in ng/gland, plotted asmeans±SEM.NE+Ins was represented as black bars and NE as diagonal black line bars. Two-wayANOVA, Bonferroni's post-hoc test, **pb0.01 vs NE TS 4 and NE TS 11, respectively. *p b
0.05 vs NE TS 5. @p b 0.001 vs NE TS 6 and 8. #pb0.01 vs NE TS 10. One-way ANOVA,Bonferroni's post-hoc test, ***pb0.05 vs all NE TSs, except 5 and 10. n=6 glands pergroup with 3 repetitions.
171R.A. Peliciari-Garcia et al. / Life Sciences 87 (2010) 169–174
Immunoblotting and immunoprecipitation
Reagents for sodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE) and immunoblotting (IB) were obtained fromBio-Rad (Hercules, CA, USA). Trizma, aprotinin, dithiothreitol, TritonX-100, glycerol, Tween 20, bovine serum albumin, fraction V wereobtained from Sigma (St. Louis, MO, USA). Immunoblot enhancedchemiluminescence detection kit was purchased from AmershamPharmacia Biotech (Buckinghamshire, UK) and nitrocellulose mem-brane (0.45 μm) was purchased from Bio-Rad (Hercules, CA, USA).Antibodies against IRβ, IGF-1R, IRS-1, IRS-2, PI3K p85 subunit,phosphotyrosine, and α-Tubulin were ordered from Santa CruzBiotechnology Inc. (Santa Cruz, CA, USA); anti-AANAT (pT31) andanti-14-3-3 phospho Serine 185 were purchased from Millipore(Billerica, MA, USA). Pineal glands were homogenized in solubiliza-tion buffer (100 mM Tris, 1% SDS, 10 mM EDTA, 100 mM Na2P2O7,100 mM NaF, 10 mM Na2VO4). The homogenates were centrifuged at17,530 g for 20 min at 4 °C to remove insoluble material. Proteinconcentration in the supernatants was determined by the Bradforddye method using Bio-Rad reagent. Proteins were treated withLaemmli sample buffer (Laemmli 1970) containing dithiothreitoland boiled for 5 min before being loaded into 6.5% or 10% or 14% SDS-PAGE in a Bio-Rad miniature slab gel apparatus. Same concentrationaliquots (50 μg) were subjected to SDS-PAGE. Protein transfer fromthe gel to the nitrocellulosemembranewas performed for 1 h at 120 Vin a Bio-Radminiature transfer apparatus. Nonspecific protein bindingto the nitrocellulose was reduced by preincubation for 1 h at 22 °C inblocking buffer (5% nonfat dry milk, 10 mM Tris, 150 mM NaCl, and0.02% Tween 20). Membranes were probed overnight at 4 °C withantibodies diluted in blocking buffer with 3% non fat dry milk, andthen washed for 30 min in blocking buffer without milk. After thisprocedure, membranes were incubated with peroxidase-conjugatedsecondary antibody for 1 h and processed for enhanced chemilumi-nescence to visualize the immunoreactive bands. The autoradiographswere scanned and band intensities were quantified using ScionImaging software (Scion Corp., Frederick, MD, USA). For immunopre-cipitation (IP), pineal extracts were centrifuged at 15,000 g for 20 minat 4 °C to remove insoluble material. The supernatant (3 mg ofprotein) was used for IP with anti-14-3-3 and protein A-Sepharose6 MB prior to Laemmli sample buffer containing 100 mM dithiothrei-tol. Same concentration samples (50 μg) with Laemmli sample bufferwere used for total extracts. Samples were heated in a boiling waterbath for 5 min and then subjected to SDS-PAGE (14% bis-acrylamide).Protein transfer from gel to nitrocellulose membrane was performedfor 90 min at 120 V (Towbin et al. 1979). Nonspecific protein bindingto the nitrocellulose was reduced by preincubating the membraneovernight at 4 °C in blocking buffer (5% nonfat dry milk, 10 mM Tris,150 mM NaCl and 0.02% Tween-20). The nitrocellulose blot wasincubated overnight at 4 °C with anti-AANAT (1:400) diluted inblocking buffer (3% BSA) and then washed for 30 min with blockingbuffer without milk. Bound antibodies were detected with horserad-ish peroxidase-conjugated anti IgG (1:10000) and visualized withenhanced chemiluminescence detection system. HRP-conjugatedanti-IgG bound to the antibodies was detected by autoradiographyusing preflashed Kodak XAR film (Eastman Kodak Co., Rochester, NY).Band intensities were quantitated from the developed autoradio-graphs using Scion Imaging software.
Chromatography
Pineal gland melatonin levels were measured by HPLC withelectrochemical detection running Empower software (Waters Sys-tem, Milford, MA, USA). Melatonin was separated on a Resolve C18column (5 μm, 150×3.9 mm). The chromatographic system wasisocratically operated with the following mobile phase: 0.1 M sodiumacetate, 0.1 M citric acid, 0.15 mM EDTA, 30% methanol, pH 3.7, at a
flow rate of 1 mL/min. The electrochemical detector potential wasadjusted to+900 mV. The elution time for melatonin was about6 min. Each gland was sonicated (Microson XL 2005, Heat System Inc.,Farmingdale, NY, USA) in a solution of 0.1 M perchloric acid (120 μL),containing 0.02% EDTA and 0.02% sodium bisulfate. After centrifuga-tion (2 min, 13,000 g, Eppendorf 5415C centrifuge, Brinkman Instru-ments Inc., Westbury, NY, USA), 40 μL of the supernatant was injectedinto the chromatographic system (Injector Mod. 7125, 20 μL loop,Rheodyne Inc., San Francisco, CA, USA).
Statistical analysis
Results were plotted as mean±SEM. For each experimental block(at least 3 replications) One-way or Two-way ANOVA followed byBonferroni's multiple comparisons post-hoc test was performed usingGraphPad Prism (GraphPad Software version 5.01, San Diego, CA,USA).
Results
According to the temporal insulin susceptibility protocol, periodsof differential sensitivities were observed. The presence of insulin onlyfor 1 h-period was able to further increase melatonin synthesis,induced by NE, exclusively in two specific moments: one at thebeginning (TS 4) and a second one at the end (TS 11) of the NE phase.In other TSs, insulin did not show its potentiating effect on melatoninsynthesis. However, insulin also evoked a reduction on melatoninsynthesis observed in the TSs 5, 6, 8 and 10. As expected, the NEstimulated groups, demonstrated the maximum induction of melato-nin synthesis around TSs 6–8 (Fig. 2).
In order to analyze the basal expression of proteins commonlyrecruited by the ISP, an immunoblotting was performed fromstandard cultured pineal glands without any possible treatmentand/or stimulation interference. The analysis revealed the expressionof IRβ, IGF-1R, IRS-1, IRS-2 and p85 subunit of the PI3K. Liver samplesare only demonstrated as positive controls and no comparisonsbetween liver and pineal gland protein amounts were performed(Fig. 3).
The state of the primary protein related to the signaling of insulinpathway, IRβ, was investigated within pineal gland according to thetyrosine pp85 phosphorylation. The synchronized pineal glandculture revealed increased tyrosine pp85 phosphorylation only in
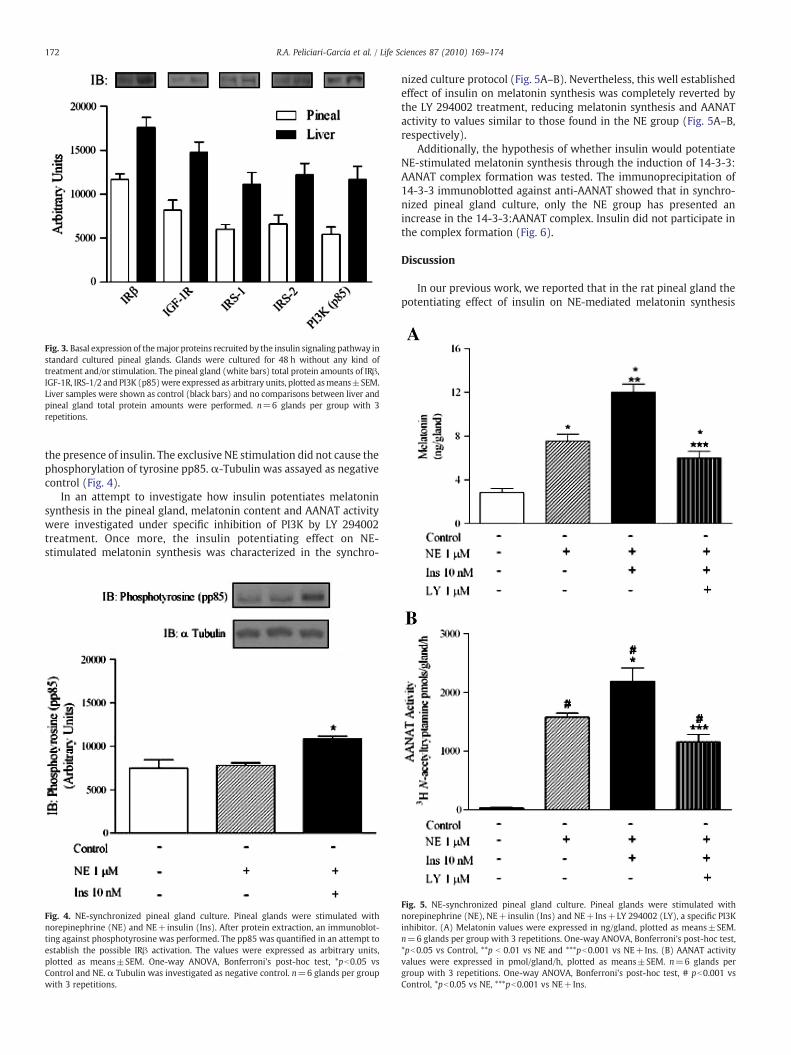
Fig. 3. Basal expression of themajor proteins recruited by the insulin signaling pathway instandard cultured pineal glands. Glands were cultured for 48 h without any kind oftreatment and/or stimulation. The pineal gland (white bars) total protein amounts of IRβ,IGF-1R, IRS-1/2 and PI3K (p85)were expressed as arbitrary units, plotted asmeans±SEM.Liver samples were shown as control (black bars) and no comparisons between liver andpineal gland total protein amounts were performed. n=6 glands per group with 3repetitions.
172 R.A. Peliciari-Garcia et al. / Life Sciences 87 (2010) 169–174
the presence of insulin. The exclusive NE stimulation did not cause thephosphorylation of tyrosine pp85. α-Tubulin was assayed as negativecontrol (Fig. 4).
In an attempt to investigate how insulin potentiates melatoninsynthesis in the pineal gland, melatonin content and AANAT activitywere investigated under specific inhibition of PI3K by LY 294002treatment. Once more, the insulin potentiating effect on NE-stimulated melatonin synthesis was characterized in the synchro-
Fig. 4. NE-synchronized pineal gland culture. Pineal glands were stimulated withnorepinephrine (NE) and NE+insulin (Ins). After protein extraction, an immunoblot-ting against phosphotyrosine was performed. The pp85 was quantified in an attempt toestablish the possible IRβ activation. The values were expressed as arbitrary units,plotted as means±SEM. One-way ANOVA, Bonferroni's post-hoc test, *pb0.05 vsControl and NE. α Tubulin was investigated as negative control. n=6 glands per groupwith 3 repetitions.
nized culture protocol (Fig. 5A–B). Nevertheless, this well establishedeffect of insulin on melatonin synthesis was completely reverted bythe LY 294002 treatment, reducing melatonin synthesis and AANATactivity to values similar to those found in the NE group (Fig. 5A–B,respectively).
Additionally, the hypothesis of whether insulin would potentiateNE-stimulated melatonin synthesis through the induction of 14-3-3:AANAT complex formation was tested. The immunoprecipitation of14-3-3 immunoblotted against anti-AANAT showed that in synchro-nized pineal gland culture, only the NE group has presented anincrease in the 14-3-3:AANAT complex. Insulin did not participate inthe complex formation (Fig. 6).
Discussion
In our previous work, we reported that in the rat pineal gland thepotentiating effect of insulin on NE-mediated melatonin synthesis
Fig. 5. NE-synchronized pineal gland culture. Pineal glands were stimulated withnorepinephrine (NE), NE+insulin (Ins) and NE+Ins+LY 294002 (LY), a specific PI3Kinhibitor. (A) Melatonin values were expressed in ng/gland, plotted as means±SEM.n=6 glands per group with 3 repetitions. One-way ANOVA, Bonferroni's post-hoc test,*pb0.05 vs Control, **p b 0.01 vs NE and ***pb0.001 vs NE+Ins. (B) AANAT activityvalues were expressed in pmol/gland/h, plotted as means±SEM. n=6 glands pergroup with 3 repetitions. One-way ANOVA, Bonferroni's post-hoc test, # pb0.001 vsControl, *pb0.05 vs NE, ***pb0.001 vs NE+Ins.
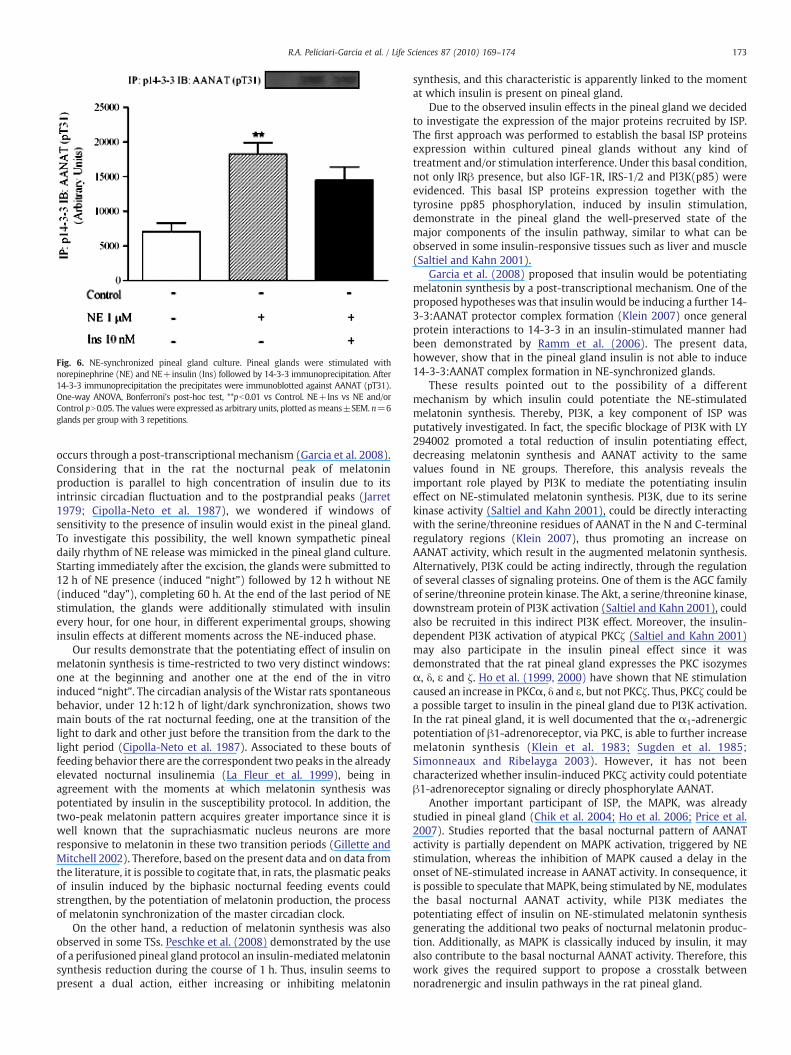
Fig. 6. NE-synchronized pineal gland culture. Pineal glands were stimulated withnorepinephrine (NE) and NE+insulin (Ins) followed by 14-3-3 immunoprecipitation. After14-3-3 immunoprecipitation the precipitates were immunoblotted against AANAT (pT31).One-way ANOVA, Bonferroni's post-hoc test, **pb0.01 vs Control. NE+Ins vs NE and/orControl pN0.05. The values were expressed as arbitrary units, plotted asmeans±SEM. n=6glands per group with 3 repetitions.
173R.A. Peliciari-Garcia et al. / Life Sciences 87 (2010) 169–174
occurs through a post-transcriptional mechanism (Garcia et al. 2008).Considering that in the rat the nocturnal peak of melatoninproduction is parallel to high concentration of insulin due to itsintrinsic circadian fluctuation and to the postprandial peaks (Jarret1979; Cipolla-Neto et al. 1987), we wondered if windows ofsensitivity to the presence of insulin would exist in the pineal gland.To investigate this possibility, the well known sympathetic pinealdaily rhythm of NE release was mimicked in the pineal gland culture.Starting immediately after the excision, the glands were submitted to12 h of NE presence (induced “night”) followed by 12 h without NE(induced “day”), completing 60 h. At the end of the last period of NEstimulation, the glands were additionally stimulated with insulinevery hour, for one hour, in different experimental groups, showinginsulin effects at different moments across the NE-induced phase.
Our results demonstrate that the potentiating effect of insulin onmelatonin synthesis is time-restricted to two very distinct windows:one at the beginning and another one at the end of the in vitroinduced “night”. The circadian analysis of theWistar rats spontaneousbehavior, under 12 h:12 h of light/dark synchronization, shows twomain bouts of the rat nocturnal feeding, one at the transition of thelight to dark and other just before the transition from the dark to thelight period (Cipolla-Neto et al. 1987). Associated to these bouts offeeding behavior there are the correspondent two peaks in the alreadyelevated nocturnal insulinemia (La Fleur et al. 1999), being inagreement with the moments at which melatonin synthesis waspotentiated by insulin in the susceptibility protocol. In addition, thetwo-peak melatonin pattern acquires greater importance since it iswell known that the suprachiasmatic nucleus neurons are moreresponsive to melatonin in these two transition periods (Gillette andMitchell 2002). Therefore, based on the present data and on data fromthe literature, it is possible to cogitate that, in rats, the plasmatic peaksof insulin induced by the biphasic nocturnal feeding events couldstrengthen, by the potentiation of melatonin production, the processof melatonin synchronization of the master circadian clock.
On the other hand, a reduction of melatonin synthesis was alsoobserved in some TSs. Peschke et al. (2008) demonstrated by the useof a perifusioned pineal gland protocol an insulin-mediatedmelatoninsynthesis reduction during the course of 1 h. Thus, insulin seems topresent a dual action, either increasing or inhibiting melatonin
synthesis, and this characteristic is apparently linked to the momentat which insulin is present on pineal gland.
Due to the observed insulin effects in the pineal gland we decidedto investigate the expression of the major proteins recruited by ISP.The first approach was performed to establish the basal ISP proteinsexpression within cultured pineal glands without any kind oftreatment and/or stimulation interference. Under this basal condition,not only IRβ presence, but also IGF-1R, IRS-1/2 and PI3K(p85) wereevidenced. This basal ISP proteins expression together with thetyrosine pp85 phosphorylation, induced by insulin stimulation,demonstrate in the pineal gland the well-preserved state of themajor components of the insulin pathway, similar to what can beobserved in some insulin-responsive tissues such as liver and muscle(Saltiel and Kahn 2001).
Garcia et al. (2008) proposed that insulin would be potentiatingmelatonin synthesis by a post-transcriptional mechanism. One of theproposed hypotheses was that insulin would be inducing a further 14-3-3:AANAT protector complex formation (Klein 2007) once generalprotein interactions to 14-3-3 in an insulin-stimulated manner hadbeen demonstrated by Ramm et al. (2006). The present data,however, show that in the pineal gland insulin is not able to induce14-3-3:AANAT complex formation in NE-synchronized glands.
These results pointed out to the possibility of a differentmechanism by which insulin could potentiate the NE-stimulatedmelatonin synthesis. Thereby, PI3K, a key component of ISP wasputatively investigated. In fact, the specific blockage of PI3K with LY294002 promoted a total reduction of insulin potentiating effect,decreasing melatonin synthesis and AANAT activity to the samevalues found in NE groups. Therefore, this analysis reveals theimportant role played by PI3K to mediate the potentiating insulineffect on NE-stimulated melatonin synthesis. PI3K, due to its serinekinase activity (Saltiel and Kahn 2001), could be directly interactingwith the serine/threonine residues of AANAT in the N and C-terminalregulatory regions (Klein 2007), thus promoting an increase onAANAT activity, which result in the augmented melatonin synthesis.Alternatively, PI3K could be acting indirectly, through the regulationof several classes of signaling proteins. One of them is the AGC familyof serine/threonine protein kinase. The Akt, a serine/threonine kinase,downstream protein of PI3K activation (Saltiel and Kahn 2001), couldalso be recruited in this indirect PI3K effect. Moreover, the insulin-dependent PI3K activation of atypical PKCζ (Saltiel and Kahn 2001)may also participate in the insulin pineal effect since it wasdemonstrated that the rat pineal gland expresses the PKC isozymesα, δ, ε and ζ. Ho et al. (1999, 2000) have shown that NE stimulationcaused an increase in PKCα, δ and ε, but not PKCζ. Thus, PKCζ could bea possible target to insulin in the pineal gland due to PI3K activation.In the rat pineal gland, it is well documented that the α1-adrenergicpotentiation of β1-adrenoreceptor, via PKC, is able to further increasemelatonin synthesis (Klein et al. 1983; Sugden et al. 1985;Simonneaux and Ribelayga 2003). However, it has not beencharacterized whether insulin-induced PKCζ activity could potentiateβ1-adrenoreceptor signaling or direcly phosphorylate AANAT.
Another important participant of ISP, the MAPK, was alreadystudied in pineal gland (Chik et al. 2004; Ho et al. 2006; Price et al.2007). Studies reported that the basal nocturnal pattern of AANATactivity is partially dependent on MAPK activation, triggered by NEstimulation, whereas the inhibition of MAPK caused a delay in theonset of NE-stimulated increase in AANAT activity. In consequence, itis possible to speculate that MAPK, being stimulated by NE, modulatesthe basal nocturnal AANAT activity, while PI3K mediates thepotentiating effect of insulin on NE-stimulated melatonin synthesisgenerating the additional two peaks of nocturnal melatonin produc-tion. Additionally, as MAPK is classically induced by insulin, it mayalso contribute to the basal nocturnal AANAT activity. Therefore, thiswork gives the required support to propose a crosstalk betweennoradrenergic and insulin pathways in the rat pineal gland.

174 R.A. Peliciari-Garcia et al. / Life Sciences 87 (2010) 169–174
Conclusions
In summary, the present results demonstrate in cultured rat pinealgland periods of differential insulin sensitivity, the expression ofmajor proteins commonly recruited by the insulin signaling pathwaylike IRβ, IGF-1R, IRS-1/2 and PI3K(p85) as well as the PI3K-dependentpotentiating effect of insulin on NE-mediated melatonin synthesis.
Conflict of interest
A conflicting interest exists when professional judgement concerning a primaryinterest (such as patient's welfare or the validity of research) may be influenced by asecondary interest (such as financial gain or personal rivalry). It may arise for theauthors when they have financial interest that may influence their interpretation oftheir results or those of others. Examples of potential conflicts of interest includeemployment, consultancies, stock ownership, honoraria, paid expert testimony, patentapplications/registrations, and grants or other funding.
Funding source
All sources of funding should also be acknowledged and youshould declare any involvement of study sponsors in the study design;collection, analysis and interpretation of data; the writing of themanuscript; the decision to submit the manuscript for publication. Ifthe study sponsors had no such involvement, this should be stated.
Acknowledgements
Supported by FAPESP, RAPG 2007/07790-6, JAS 2007/08733-06,ACM 2005/60000-8, JCN and CROC grant 04/06767-2.
References
Afeche SC, Amaral FG, Villela DCM, Abrahão MV, Peres R, Cipolla-Neto J. Melatonin andpineal gland (Chapter 10). In: Romano, E, De Luca, S (Eds.), New Research onNeurosecretory Systems, vol. 1. Nova Science Publishers, Hauppauge, pp. 155–157,2008.
Alonso-Vale MI, Adreotti S, Borges-Silva C, Mukai PY, Cipolla-Neto J, Lima FB.Intermittent and rhythmic exposure to melatonin in primary cultured adipocytesenhances the insulin and dexamethasone effects on leptin expression. Journal ofPineal Research 41 (1), 28–34, 2006.
Alonso-Vale MI, Adreotti S, Peres SB, Anhê GF, Borges-Silva C, Cipolla-Neto J, Lima FB.Melatonin enhances leptin expression by rat adipocytes in the presence of insulin.American Journal of Physiology Endocrinology and Metabolism 288 (4), 805–812,2005.
Alonso-Vale MI, Andreotti S, Mukai PY, Borges-Silva CN, Peres SB, Cipolla-Neto J, LimaFB. Melatonin and the circadian entrainment of metabolic and hormonal activitiesin primary isolated adipocytes. Journal of Pineal Research 45 (4), 422–429, 2008.
Alonso-Vale MI, Anhê GF, Borges-Silva CN, Andreotti S, Peres SB, Cipolla-Neto J, Lima FB.Pinealectomy alters adipose tissue adaptability to fasting in rats. Metabolism 53(4), 500–506, 2004a.
Alonso-Vale MI, Borges-Silva CN, Anhê GF, Andreotti S, Machado MA, Cipolla-Neto J,Lima FB. Light/dark cycle-dependent metabolic changes in adipose tissue ofpinealectomized rats. Hormone and Metabolism Research 36 (7), 474–479, 2004b.
Anhê GF, Caperuto LC, Pereira-da-Silva M, Souza LC, Hirata AE, Velloso LA, Cipolla-NetoJ, Carvalho CR. In vivo activation of insulin receptor tyrosine kinase by melatonin inthe rat hypothalamus. Journal of Neurochemistry 90 (3), 559–566, 2004.
Chik CL, Mackova M, Price D, Ho AK. Adrenergic regulation and diurnal rhythm of p38mitogen-activated protein kinase phosphorylation in the rat pineal gland.Endocrinology 145 (11), 5194–5201, 2004.
Cipolla-Neto J, Recine EG, Menna-Barreto LS, Marques N, Afeche SC, Schott C, FortunatoG, Sothern RB, Halberg F. Perinatal malnutrition, suprachiasmatic nuclear lesioningand circadian-ultradian aspects of spontaneous behavior of albino rats. Progress inClinical and Biological Research 227B, 473–489, 1987.
Deguchi T, Axelrod J. Sensitive assay for serotonin N-acetyltransferase activity in ratpineal. Analytical Biochemistry 50 (1), 174–179, 1972.
Garcia RAP, Afeche SC, Scialfa JH, Amaral FG, Santos SHJ, Lima FB, Young ME, Cipolla-Neto J. Insulin modulates norepinephrine-mediates melatonin synthesis incultured rat pineal gland. Life Sciences 82 (1–2), 108–114, 2008.
Gillette MU, Mitchell JW. Signaling in the suprachiasmatic nucleus: selectivelyresponsive and integrative. Cell and Tissue Research 309 (1), 99–107, 2002.
Ho AK, Hashimoto K, Matowe W, Chik CL. Adrenergic regulation of translocation ofprotein kinase C isozymes in rat pinealocytes. Molecular and Cellular Endocrinol-ogy 150 (1–2), 169–178, 1999.
Ho AK, Ling A, Chik CL. Intracellular pH on translocation of protein kinase C isozymes inrat pinealocytes. Journal of Neurochemistry 75 (5), 1845–1851, 2000.
Ho AK, Price DM, Terrif D, Chik CL. Timing of mitogen-activated protein kinase (MAPK)activation in the rat pineal gland. Molecular and Cellular Endocrinology 252 (1–2),34–39, 2006.
Jarret RJ. Rhythms in insulin and glucose. In: Krieger, DT (Ed.), Endocrine Rhythms, vol.1. Raven Press, New York, pp. 247–258, 1979.
Klein DC, Sugden D, Weller JL. Postsynaptic α-adrenergic receptors potentiate the β-adrenergic stimulation of pineal serotonin N-acetyltransferase. Proceedings of theNational Academy of Sciences of the United States of America 80 (2), 599–603,1983.
Klein DC. Arylalkylamine N-acetyltransferase: “the Timezyme”. The Journal ofBiological Chemistry 282 (7), 4233–4237, 2007.
La Fleur SE, Kalsbeek A, Wortel J, Buijis RM. A suprachiasmatic nucleus generatedrhythm in basal glucose concentrations. Journal of Neuroendocrinology 11 (8),643–652, 1999.
Laemmli UK. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227 (5259), 680–685, 1970.
Lima FB, Machado UF, Bartol I, Seraphim PM, Sumida DH, Moraes SM, Hell NS, OkamotoMM, Saad MJ, Carvalho CR, Cipolla-Neto J. Pinealectomy causes glucose intoleranceand decreases adipose cell responsiveness to insulin in rats. The American Journalof Physiology 275 (6 Pt 1), 934–941, 1998.
Lima FB, Matsushita DH, Hell NS, Dolnikoff MS, Okamoto MM, Cipolla-Neto J. Theregulation of insulin action in isolated adipocytes. Role of the periodicity of foodintake, time of day and melatonin. Brazilian Journal of Medical and BiologicalResearch 27 (4), 995–1000, 1994.
Parfitt A, Weller JL, Klein DC, Sakai KK, Marks BH. Blockade by ouabain or elevatedpotassium ion concentration of the adrenergic and adenosine cyclic 3′, 5′-monophosphate-induced stimulation of pineal serotonin N-acetyltransferaseactivity. Molecular Pharmacology 11 (3), 241–255, 1975.
Peschke E, Wolgast S, Bazwinsky I, Klaus Ponicke K, Muhlbauer E. Increased melatoninsynthesis in pineal glands of rats in streptozotocin induced type 1 diabetes. Journalof Pineal Research 45 (4), 439–448, 2008.
Picinato MC, Haber EP, Carpinelli AR, Cipolla-Neto J. Daily rhythm of glucose-inducedinsulin secretion by isolated islets from intact and pinealectomized rat. Journal ofPineal Research 33 (3), 172–177, 2002a.
Picinato MC, Haber EP, Cipolla-Neto J, Curi R, de Oliveira Carvalho CR, Carpinelli AR.Melatonin inhibits insulin secretion and decreases PKA levels without interferingwith glucose metabolism in rat pancreatic islets. Journal of Pineal Research 33 (3),156–160, 2002b.
Picinato MC, Hirata AE, Cipolla-Neto J, Curi R, Carvalho CR, Anhê GF, Carpinelli AR.Activation of insulin and IGF-1 signaling pathways by melatonin through MT1receptor in isolated rat pancreatic islets. Journal of Pineal Research 44 (1), 88–94,2008.
Price DM, Wloka MT, Chik CL, Ho AK. Mitogen-activated protein kinase phosphatase-1(MKP-1) preferentially dephosphorylates p42/44MAPK but not p38MAPK in ratpinealocytes. Journal of Neurochemistry 101 (6), 1685–1693, 2007.
Ramm G, Larance M, Guilhaus M, James DE. A role for 14-3-3 in insulin-stimulatedGLUT4 translocation through its interaction with the RabGAP AS160. The Journal ofBiological Chemistry 281 (39), 29174–29180, 2006.
Saltiel AR, Kahn CR. Insulin signaling and the regulation of glucose and lipidmetabolism. Nature 414 (6865), 799–806, 2001.
Simonneaux V, Ribelayga C. Generation of the melatonin endocrine message inmammals: a review of the complex regulation of melatonin synthesis bynorepinephrine, peptides and other pineal transmitters. Pharmacological Reviews55 (2), 325–395, 2003.
Sugden D, Vanecek J, Klein DC, Thomas TP, AndersonWB. Activation of protein kinase Cpotentiates isoprenaline-induced cyclic AMP accumulation in rat pinealocyte.Nature 314 (6009), 359–361, 1985.
Towbin H, Staehlin J, Gordon J. Electrophoretic transfer of proteins from polyacrylamidegels to nitrocellulose sheets: procedure and some applications. Proceedings of theNational Academy of Sciences of the United States of America 76 (9), 4350–4354,1979.
Wu ZL, O'Kane TM, Connors TJ, Marino MJ, Schaffhauser H. The phosphatidylinositol 3-kinase inhibitor LY 294002 inhibits GlyT1-mediated glycine uptake. Brain Research1227, 42–51, 2008.





























