Embed Size (px)
Citation preview

Cell Tissue Res (1983) 228:345-355 Cell and Tissue Research �9 Springer-Verlag 1983
The rate-limiting step in hydrosmotic response of frog urinary bladder
A freeze-fracture study at different temperatures and medium pH
J. Chevalier*, M. Parisi, and J. Bourguet with the technical assistance of R. Gobin D6partement de Biologie, Centre d'Etudes Nud~aires de Saclay, France
Summary. The ADH-induced water fluxes and the associated appearance of intramembranous particle aggregates in the luminal membrane of frog urinary bladders have been correlated in a time course study. Plots of the onset and reversal of the oxytocin-induced hydrosmotic response were sigmoidal in shape, symmetrical and slowed by low temperature to the same degree. Parallel freeze- fracture studies showed that the mean size distribution of the aggregates was constant at different temperatures and at different times during hormonal stimulation and washout. No qualitatively different picture of aggregate formation was detected at low temperature: this suggests that the insertion and removal of individual aggregates into or from the apical plasma membrane is a rather rapid process, both at 20 and at 6.5~ As in the case of water permeability, both aggregate appearance and disappearance were similarly slowed by lowering the temperature.
A similar time-course study of the inhibition of the hydrosmotic response by acidification of the medium was also made. In this case, lowering the incubation temperature induced a clear dissociation between net water flow and the surface area occupied by the aggregates. For the first time, a low water permeability was found associated with a high aggregate surface area in the apical membrane, indicating that cellular acidification induces an impairment of aggregate function rather than a reduction of surface area.
Key words" Urinary bladder (frog) - In t ramembranous particle aggregate turnover - Antidiuretic hormone - Water permeability
Antidiuretic hormone (ADH) increases water permeability in amphibian urinary bladder. This action is not instantaneous. After a short latent period, water permeability reaches its maximum level in 10-15min at room temperature (Bourguet 1978). The time-course is remarkably constant and follows a sigmoi'dal curve with a half-time of about 6-7 min (Bourguet 1968). The comparison of time
Send offprint requests to: J. Chevalier DB-LPPC, C.E.N. Saclay, F 91191 Gif-sur-Yvette Cedex, France
* J.C. is a career investigator at the Institut National de la Sant~ et de la Recherche Medicale, INSERM V. 48
0302-766X/83/0228/0345/$02.20

346 J. Chevalier et al.
courses and reaction half times to cyclic AMP, the intracellular messenger of ADH (Orloffand Handler 1967), as well as to other agonists (Bentley 1964; Ripoche et al. 1973), indicates that the apparent rate-limiting step in t h e A D H action should occur after the increase in concentration of cyclic AMP (Ripoche et al. 1973). The nature and the role of this limiting step remain unknown. However, it is temperature- dependent (Bourguet 1966) and a high value was observed for the temperature coefficient (Qlo) calculated from the half time data (Bourguet 1966). Various agonists induce variations in water permeability that show a similar temperature- dependent time course. They are thus likely to interfere with events taking place before the presumed limiting step. Other agonists appear to interfere with events occurring after the rate-limiting step. For example, recent experiments performed at different temperatures, indicate that cellular acidification reversibly inhibits the increase in water permeability induced by different agents by interacting with an apparent temperature independent, post-cyclic AMP step (Parisi et al. 1981). This step must occur after the rate-limiting step.
Freeze-fracture studies have demonstrated the appearance of aggregates of intramembranous particles (most probably proteins) in the apical border of epithelial cells challenged with ADH, cyclic AMP or other agonists (Bourguet et al. 1976; Brown et al. 1980; Chevalier et al. 1974, 1979b; Harmanci et al. 1978; Kachadorian et al. 1975; Wade et al. 1977). Strong experimental evidence indicates that these aggregates are closely related to the ADH-induced water pathway. They appear to be inserted into the apical membrane from preformed cytoplasmic vesicles in response to the hormonal stimulus (Muller et al. 1980), but it is not known if their insertion represents the rate limiting step of the reaction and if aggregates appear progressively in proportion to the increase in net water flux.
We therefore initiated a morpho-functional time-course study of the hy- drosmotic response of the frog urinary bladder at different temperatures and compared the results obtained with those observed following acidification of the incubation medium. We observed that particle aggregate structure was quali- tatively similar under all experimental conditions. The insertion of particles aggregates into the apical membrane and their removal was, however, markedly slower at low temperature. Their number and surface area, however, always remained proportional to the net water flow, suggesting that once the aggregates had been inserted in the apical membrane, there was no longer any significant time delay in the increase in net water flow. Inhibition of the net water flow by medium acidification on the other hand, remained quite rapid at low temperature and was not accompanied by disappearance of the apical particle aggregates. Thus, under these conditions, we have made the first observation of a high aggregate density associated with a low water permeability.
Materials and methods
Frogs (Rana esculenta) originating from Central Europe were kept at 20 ~ C in running tap water for at least 5 days before sacrifice. The bladders were mounted between two lucite chambers, the serosal face of the tissue being bathed with a buffer solution containing Na+= ll4.5mEq/l; K + =5mEq/l; Ca ++ = lmEq/1; C1- = ll9mEq/l; HCO3 = 2.5mEq/l; pH = 8.1 when bubbled with air. The mucosal bath was made hypotonic by reducing the NaC1 concentration to 5.6 raM. In some experiments, the pH of the medium was changed and stabilized at a new value by adding the necessary amount of HC1

The rate limiting step in hydrosmotic action 347
or NaOH with a microsyringe device, as described elsewhere (Parisi et al. 1981). Experiments were performed either at room temperature (20+ 1 ~ C) or at 6.5~ in a thermostatically controlled bath.
Urinary bladders were stimulated by a maximal concentration (2 �9 10- s M) of oxytocin, a synthetic structural analog of ADH (Syntocinon, Sandoz Pharmaceuticals, Basel, Switzerland). The minute by minute net water flow was recorded as already described (Bourguet and Jard 1964). Samples for freeze fracture were obtained by fixing the bladder, immediately after net flow measurement as soon as the appropriate permeability level had been reached. Due to the marked slowing down of some responses at low temperature, incubation times were thus usually longer in the last condition. Preparations were fixed for 15min with 2 ~ glutaraldehyde in 0.1 M cacodylate buffer (the pH was adjusted to that of the incubation buffer used and the solutions were at the experimental temperature), cryoprotected in glycerol Ringer solution 30 % v/v for 40 min and subsequently frozen in Freon 22 chilled by liquid nitrogen. Samples were then processed as usual for freeze-fracture (Chevalier et al. 1974) in a Balzers 301 cryopump freeze etch unit (Balzers, Lichtenstein) at a 4 �9 10-8 Torr high vacuum. The total cell surface area occupied by the aggregates and the size distribution of the aggregates were calculated using a Kontron MOP-AM 03 image analysing computer (Reichert-Jung, Paris) on pictures at a final magnification of 45 500 x. At least 4 different experiments were performed in each condition. On the average, 11 different cells, and an apical surface area of 490+ 40 Ixm 2, taken at random, were analysed in each case. In this work the term "aggregate" refers to units composed of intramembranous particles condensed in closely spaced parallel linear arrays. Two groups of particle arrays, at angles to one another, were thus counted as two aggregates. Aggregate mean size was obtained by dividing the total surface area of the aggregates by their number. These particle condensations are very often associated into "islets" forming long ribbons (see Fig. 2) or stellar arrangements. A group of joining aggregates was considered as an islet when its total surface area was greater than 2.4. 10 -21xm 2 (50mm 2 on micrographs).
Results
Figure 1 illustrates the time-course of the hydrosmotic response of frog urinary bladder to oxytocin at 20 and at 6.5 ~ C. It also shows the effects of medium acidification when applied at both temperatures at the peak of the reaction. Freeze fracture studies were performed at different times after both the addition and the washout of the hormone and after acidification of the incubating medium.
t-.-oxY - o - pH 6 ~
~, oI'AjI~ 11~,: r ~, I I
0 2 0
I.-.- O X Y �9 - o - p_H.~66
p,
- I - - - I I I I i ,, 0 2 0 4 0 6 0 8 0 1 0 0
T i m e ( m i n )
Fig. 1. Changes in net water flow after oxytocin (2.2 - 10- 8 M) addition and washout (filled circles), and during pH inhibition of oxytocin response (open circles). At least 6 experiments were performed under each experimental condition. Incubation temperature was either 20~ (left curves) or ~ 7 ~ (right curves). Arrowheads indicate the conditions that were selected for freeze-fracture studies

348 J. Chevalier et al.
Fig. 2a, b. Protoplasmic and external fracture faces of granular cells fixed at 6.5 ~ C. Typical particle aggregates and the corresponding groove networks are observed, similar to those observed at 20 ~ C. As shown in these pictures, particularly large areas of contiguous particle aggregates are observed in this condition. They form either long ribbons - as here or stellar arrangements (not shown)
I. Morphological aspects oJ preparations at low temperature
In general , no differences were observed in the apical and baso la te ra l m e m b r a n e s t ructure or i n t r a m e m b r a n o u s par t ic le shape and d is t r ibu t ion at r oom and low temperatures . Aggrega tes o f i n t r a m e m b r a n o u s part icles, which are observed at the apical bo rde r o f oxytoc in t rea ted cells, exhibi ted the same appea rance at 6.5 ~ C as at 20~ (Fig. 2). A t these two exper imenta l temperatures , g lu tara ldehyde-f ixed aggregates appea red on the P face whereas the complemen ta ry groove ne tworks were visible on the E face. In some cases, a well defined o r thogona l pa t te rn was clearly visible inside these groove ne tworks (Fig. 2b). A t the peak o f the hydrosmot i c response to oxytoc in the d is t r ibut ions o f aggregate size at the two exper imenta l t empera tu res did not differ s tat is t ical ly (Fig. 3), the mean aggregate size being close to 4 �9 l0 -3 ~tm 2.
When we s tar ted this work at low tempera tures , we hoped to slow down events sufficiently to be able to observe t ransient steps which might explain the mechanisms o f aggregate fo rma t ion and d isappearance , but in fact we never observed any special phenomena which could be related to these mechanisms. The size d is t r ibu t ion o f aggregates remained cons tan t dur ing the t ime-course o f the hydrosmot i c response o f the b ladder to oxytoc in (Fig. 3). The islets o f aggregates, however, were 1.5 t imes more numerous dur ing the onset than ei ther at the peak of the response or dur ing the decrease o f water permeabi l i ty (Table 1).

The rate limiting step in hydrosmotic action 349
o30[ o 2o
E Z
1(:
G 0 - 2 2 - 4 4-6 6 - 8 8-10 10-12 12-14 14-16 16-18
Aggregate size category (,10 -3}
>18
Fig. 3. Aggregate size distribution, as the percentage of the total area occupied by aggregates, under different conditions. For each aggregate size category, bars represent (from left to right) experiments performed under the following conditions: a) At the peak of the response to oxytocin (2.2 �9 10- s M), at 20-21 ~ C; b) at the peak of the response to oxytocin at 6-7 ~ C, c) at 50 ~o of the maximal flux during the increase of the response to oxytocin at 6-7 ~ C and d) at 50 ~ of the maximal flux during the decrease of the response after oxytocin washout at 6-7 ~ C. No significant differences were observed
Table 1. Fraction of the apical surface area occupied by aggregates and islets at different temperatures and at different times after oxytocin (2.2 x 10 - s M) stimulation. Correlation with the corresponding water fluxes
Condition ~ C Net water Aggregate Aggregate Number o f islets Number o f islets f l u x surface surface
x 100 • 100 (~tl �9 m i n - t Total ~ Total surface Aggregate surface �9 cm- 2) surface • ~tl surface • p.l ( p m 2) ( i . tm 2)
At rest 6-7 (n = 3)
50 ~ of maximal 6-7 response (onset) (n = 5)
Maximal flux 6-7 (n = 5)
50 ~ of maximal 6-7 response (washout) (n = 8)
Maximal flux 20 (n = 8)
- 0.01 _+ 0.05 -
0.85_+0.12 0 .72+0.24 0.85_+0.27 0.090+0.022 0.132+0.012(1) a
1.39+ 0.29 0.98_+ 0.14 0.68_+ 0.07 0.077-+ 0.012 0.092-+ 0.011 (2) a
0.83_+ 0.12 0.48-+ 0.17 0.58+ 0.25 0.059_+ 0.031 0.098+ 0.022
3.19-+ 0.40 0.99-+ 0.21 0.32_+ 0.07 0.085_+ 0.021 0.096-+ 0.09
a Mean difference (1-2) = 0.040_+ 0.010; P < 0 . 0 5

350 J. Chevalier et al.
II. Correlation between the observed net f lux of water and intramembranous particle aggregates under dif[erent conditions of temperatures and pH in the incubation medium
Two fragments of the same bladder were stimulated by oxytocin at 6-7 ~ C and the net water flux recorded. Glutaraldehyde fixation was performed under two of the following conditions in each pair of experiments: a) when the response had reached 50 ~0 of the maximum, b) at the peak of the hormonal action and c) when, after oxytocin washout, the flux had again fallen to 50 ~o of the maximal value (see Fig. 1). Bladders were also fixed at rest at 6 7 ~ C and, in one control series at room temperature, either at rest or at the peak of the oxytocin action.
The fraction of the cell surface area occupied by the aggregates and the corresponding water permeabilities at 6.5 ~ C (given in Table 1), represent the mean values of 5 different samples in each case (except in the resting condition where n = 3). Considerable variation in net water flux and cell surface area occupied by aggregates can be observed for individual bladders as indicated by the high values of the standard errors of means. This scatter was however considerably reduced by expressing the aggregate surface per unit of net water flow. A comparable value was then obtained in all cases either for the onset, for the peak of the response or for the washout (Table 1). The distribution of the aggregate size remained constant either at the maximum of the response, at 50 ~o of the maximum during the onset or at 50 % of the maximum during the wash-out, and comparable to that recorded at room temperature (Fig. 3).
We studied the effect of acidification on water fluxes and aggregates at low temperatures in the following way: two fragments of the same bladder were stimulated with oxytocin (2 �9 10- 8 M, a maximal concentration) at 6.5 ~ C. At the peak of the hormonal action, oxytocin was washed out of one fragment while the serosal pH was reduced to 6 in the other (Fig. 1). Four minutes later, both preparations were fixed and processed for freeze-fracture studies. At that moment, the control channel (4rain, oxytocin-washout) did not show any appreciable reduction in flux while in the experimental fragment (pH 6), the water flux was strongly inhibited (Table 2, Fig. 1). In contrast to what occurs at 20 ~ C, when pH inhibition was performed at 6-7 ~ C a clear dissociation between water fluxes and the number of surface of aggregates was observed: while the water permeability was strongly reduced, the number of aggregates remained almost unchanged (Table 2). The structure of these aggregates was, at the resolution level of the freeze-fracture technique, indistinguishable from that observed at 20~ (Bourguet et al. 1981).
III. EJ]ect of temperature oJ' the medium on the relationship between net water f lux and aggregates
Previous experimental evidence supports the idea that, at room temperature, the amount of aggregates is proportional to the observed increase in water permeability (Kachadorian et al. 1978). However, in our experimental conditions, the relationship between the two phenomena did not appear to be the same at 20 ~ C as at 6 7 ~ C (Table 1). At the peak of the response to oxytocin, the net water movement was higher at 20 ~ C than at 6.5 ~ C and differed by a factor of 2.3 in this experimental series. Despite these different intensities in hydrosmotic permeability, aggregate
The rate limiting step in hydrosmotic action 351
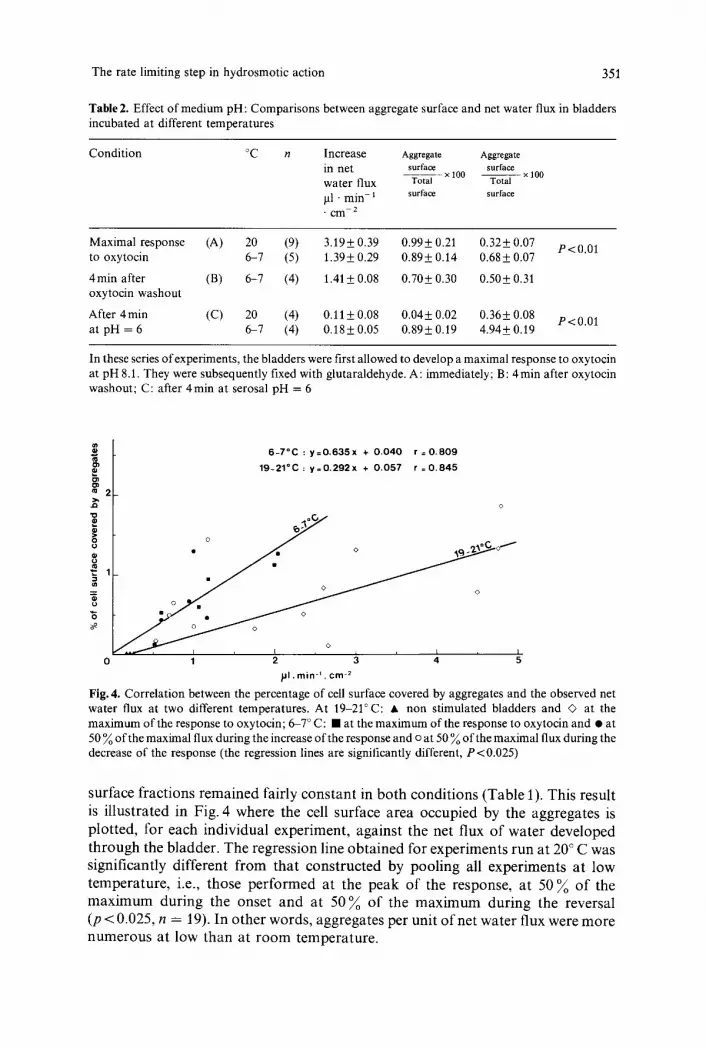
Table 2. Effect of medium pH: Comparisons between aggregate surface and net water flux in bladders incubated at different temperatures
Condition ~ n Increase A g g r e g a t e A g g r e g a t e
in net surface surface x 100 x 100
water flux Total Total pl �9 rain- 1 surface surface " c m 2
Maximal response (A) 20 (9) 3.19+ 0.39 0.99+ 0.21 0.32+ 0.07 P<0.01 to oxytocin 6-7 (5) 1.39+ 0.29 0.89+ 0.14 0.68_+ 0.07
4rain after (B) 6-7 (4) 1.41 + 0.08 0.70+ 0.30 0.50+ 0.31 oxytocin washout
After 4 min (C) 20 (4) 0.11 + 0.08 0.04+ 0.02 0.36_+ 0.08 P<0.01 at pH = 6 6-7 (4) 0.18_+ 0.05 0.89+ 0.19 4.94+ 0.19
In these series of experiments, the bladders were first allowed to develop a maximal response to oxytocin at pH 8.1. They were subsequently fixed with glutaraldehyde. A: immediately; B: 4 min after oxytocin washout; C: after 4min at serosal pH = 6
g
c0 2
>~ o
"C 1
m
6 - 7 ~ : y = O . 6 3 5 x + 0 . 0 4 0 r = 0 . 8 0 9
1 9 - 2 1 ~ y ~ O , 2 9 2 x + 0 . 0 5 7 r = 0 . 8 4 5
�9
�9 o � 9 1 4 9 o
1 2 3 4 5
~.11. r a i n -~ . c m -2
Fig. 4. Correlation between the percentage of cell surface covered by aggregates and the observed net water flux at two different temperatures. At 19-21 ~ �9 non stimulated bladders and ~ at the maximum of the response to oxytocin; 6-7 ~ C: �9 at the maximum of the response to oxytocin and �9 at 50 ~o of the maximal flux during the increase of the response and o at 50 ~o of the maximal flux during the decrease of the response (the regression lines are significantly different, P<0.025)
s u r f a c e f r a c t i o n s r e m a i n e d fa i r ly c o n s t a n t in b o t h c o n d i t i o n s ( T a b l e 1). T h i s r e s u l t
is i l l u s t r a t e d in Fig . 4 w h e r e the cell s u r f a c e a r e a o c c u p i e d by t he a g g r e g a t e s is
p l o t t e d , fo r e a c h i n d i v i d u a l e x p e r i m e n t , a g a i n s t t h e ne t f lux o f w a t e r d e v e l o p e d
t h r o u g h the b l a d d e r . T h e r e g r e s s i o n l ine o b t a i n e d fo r e x p e r i m e n t s r u n a t 20 ~ C was
s i g n i f i c a n t l y d i f f e r e n t f r o m t h a t c o n s t r u c t e d b y p o o l i n g all e x p e r i m e n t s a t l ow
t e m p e r a t u r e , i.e., t h o s e p e r f o r m e d a t t he p e a k o f t he r e s p o n s e , a t 50 ~o o f the
m a x i m u m d u r i n g t he o n s e t a n d a t 5 0 ~ o f t h e m a x i m u m d u r i n g t he r e v e r s a l
(p < 0 . 0 2 5 , n = 19). I n o t h e r w o r d s , a g g r e g a t e s pe r u n i t o f n e t w a t e r f lux we re m o r e n u m e r o u s a t low t h a n a t r o o m t e m p e r a t u r e .

352 J. Chevalier et al.
Discussion
As previously stated, ADH action is not instantaneous and from the time-course of the responses to ADH and cyclic-AMP the existence of a temperature dependent limiting step, that would be located after the accumulation of cyclic AMP, has been proposed (Bourguet 1966, 1968; Ripoche 1967). The reversal of hormonal action is also affected by medium temperature and similar results have been observed in the case of cyclic AMP (Ripoche 1967). Consequently, the onset and reversal of water permeability variations remain rather symmetrical, suggesting that a similar temperature dependent factor affects both processes.
Medium temperature and intramembranous particle aggregates
At the peak of the hydrosmotic response, the aggregate size distribution did not differ in cells fixed at 20 ~ or at 6.5 ~ C. This result confirms previous observations by Kachadorian et al. (1981) in toad urinary bladder. Furthermore, we have now observed that the aggregate size distribution is not different at other periods of hormonal stimulation and washout. The aggregate mean size was, at both 20 ~ C and 6.5~ close to 4-10-3~1T12, a value smaller than that reported for toad bladder (Kachadorian et al. 1981). Nevertheless, this figure may be influenced by subjective factors when deciding whether two adjacent aggregates should be counted as one or two. Because of this problem we consider that the fractional area occupied by aggregates, is the best measure of the aggregation phenomenon since it is not affected by subjective considerations. The fact that the aggregate surface per unit of water flux was similar during the onset of the hormonal action, at the peak of the response and during the washout (column 3, Table 1) suggests that aggregation appearance and disappearance and water permeability are both and similarly affected by low temperature.
Possible mechanisms Jor the insertion and removal oJ aggregates
Although there is some evidence that aggregates are inserted in the apical membrane from preexisting cytoplasmic structures (Muller et al. 1980), much less information exists on the mechanism of their removal. One possible mechanism could be the dispersion of the aggregated particles as it has been observed during the inhibition of net water flow by some non ionic detergents (Chevalier et al. 1979a; Parisi et al. 1979a, b). Even at a low temperature, however, under which condition aggregate involution is markedly slowed down, we never observed evidence for such a mechanism.
A significant increase in the number of islets of aggregates during the onset of the hormonal action was, however, observed. This increase probably results from the fact that the long tubular structures of aggregated particles that can be seen in the cytoplasmic carrier membranes (Humbert et al. 1977; Wade 1980) can fractionate to variable extents during transfer to their final location in the apical membrane. Some step of this process appears to be altered at low temperature.

The rate limiting step in hydrosmotic action 353
The effect of medium pH
Medium acidification, in the presence of a permeant buffer such as bicarbonate, strongly inhibits the response to ADH and cyclic AMP (Gulyassy and Edelman 1965, 1967). At 20~ this inhibition is paralleled by the disappearance of the intramembranous particle aggregates (Parisi et al. 1979a, b). pH inhibition of net water flow was also observed at low temperatures and, unlike hormonal washout, was not slowed down by the cold: the observed inhibition was as rapid as that recorded at 20 ~ C (Parisi et al. 1981). Our observations show that this inhibition is not correlated with aggregate disappearance.
There is strong experimental evidence that the aggregates are closely related to the increase in water permeability. Nevertheless, the present results show that medium acidification at low temperature can dissociate high water permeability from the presence of aggregates. We describe in further detail the action of medium acidification in a separate paper (Parisi et al. 1981); there is evidence that hydrogen ion concentration affects one of the very last steps in the chain of events started by the hormone-receptor interaction. This step must be later than the temperature- dependent rate-limiting reaction described here (Parisi et al. 1981). If we continue to accept that the aggregates represent the ADH-induced water channels, the present results can be explained if cellular acidification modifies the fine structure of the aggregates and reduces their permeability. Time course data strongly suggest such a mechanism (Parisi et al. 1981).
Medium temperature and the magnitude and time course of the hydrosmotic response
The magnitude of the hydrosmotic response to oxytocin is dependent on medium temperature (Bourguet 1966; Hays and Leaf 1962) and different attempts have been made to assess the apparent energy of activation for the ADH-stimulated water flux (Hays and Leaf 1962; Eggena 1972; Hays et al. 1971). Kachadorian et al. (1981) reported that the intramembranous particle aggregates and hormonal stimulated water flow in toad urinary bladder are simultaneously reduced with a reduction in temperature. The present results, in frog urinary bladder, however, show that at the peak of the response of the bladder to oxytocin the amplitudes of the net water movements at 20 ~ or at 5-7 ~ C differed by a factor of 2.3 while the aggregate surface fractions remained by similar (Table 2). The discrepancy between our results and those of Kachadorian et al. (1981), if not due to species differences, could arise from the different experimental protocols. In our conditions the net water flux was recorded every minute and bladders fixed only when the water movement equilibrated at the peak of the response. In the case of experiments at a low temperature this occurred about 50 rain after oxytocin stimulation, while at 20 ~ C, on the other hand, 13 to 16rain were sufficient. If water fluxes are measured in sac preparations for only two ten minute periods after oxytocin action and the preparations then fixed, at a low temperature, the response will be far from the maximum. This is evident if we look at Fig. 1 and consider the net flux evolution between 10 and 20rain after oxytocin at 20~ and 6.5~ Another point to consider, at least in our experiments, is the great scatter of individual values. This is

354 J. Chevalier et al.
spec ia l ly c l ea r f r o m Fig. 4. F u r t h e r e x p e r i m e n t s in w h i c h the w a t e r f lux
m e a s u r e m e n t c o n d i t i o n s a re ca r e fu l l y c o n t r o l l e d in p a i r e d e x p e r i m e n t s a re
p r o b a b l y n e c e s s a r y to c la r i fy th i s p o i n t . N e v e r t h e s s , f r o m o u r r e su l t s a n d a c c e p t i n g
t h a t t he s u r f a c e o f t he a g g r e g a t e s r e p r e s e n t s a g o o d e s t i m a t i o n o f t he a v a i l a b l e
su r f ace for w a t e r p e r m e a t i o n , i t m a y b e c o n c l u d e d t h a t t he ef fec t o f t e m p e r a t u r e o n
t h e w a t e r f low t h r o u g h e a c h p e r m e a t i o n u n i t is o f i m p o r t a n c e .
References
Bentley PJ (1964) Physiological properties of the isolated frog bladder in hyperosmotic solutions. Comp Biochem Physiol 12:233-239
Bourguet J (1966) Influence de la temperature sur la cinetique de l'augmentation de la perm6abilit~ zi l'eau de la vessie de grenouille sous l'action de l'ocytocine. J Physiol (Paris) 58:476
Bourguet J (1968) Cin6tique de la perm6abilisation de la vessie de grenouille par l'ocytocine. R61e du Y,5'-ad6nosine monophosphate cyclique. Biochim Biophys Acta 150:104-112
Bourguet J, Jard S (1964) Un dispositif automatique de mesure et d'enregistrement du flux net d'eau A travers la peau e t la vessie des amphibiens. Biochim Biophys Acta 88:442-444
Bourguet J, Chevalier J, Hugon JS (1976) Alterations in membrane associated particle distribution during antidiuretic challenge in frog urinary bladder epithelium. Biophysical J 16:627-639
Bourguet J, Chevalier J, Parisi M (198l) On the role of intramembranous particle aggregates in the hydrosmotic action ofantidiuretic hormone. In: Ussing HH et al. (ed) Alfred Benzon Symposium 15: "Water transport across epithelia". Munksgaard, Copenhagen 1981, pp 404-421
Brown D, Grosso A, De Sousa RC (1980) lsoproterenol-induced intramembrane particle aggregation and water flux in toad epidermis. Biochim Biophys Acta 596:158 164
Chevalier J, Bourguet J, Hugon JS (1974) Membrane associated particles. Distribution in frog urinary bladder epithelium at rest and after oxytocin treatment. Cell Tissue Res 152:129 140
Chevalier J, Bourguet J, Parisi M (1979a) New evidence on the role of intramembranous particle aggregates as the ADH induced water pathways: the effect of a low HLB surfactant, cemulsol NP- EO6 in: "Hormonal control of epithelial transport". INSERM Symposium Series 85:147 158
Chevalier J, Parisi M, Bourguet J (1979 b) Particle aggregates during antidiuretic action: some comments on their formation. Biol Cellulaire 35:207 210
Eggena P (1972) Temperature dependence of vasopressin action in the toad bladder. J Gen Physiol 59:519 533
Gulyassy PF, Edelman IS (1965) Hydrogen-ion dependence of antidiuretic action of vasopressin, oxytocin and deaminooxytocin. Biochim Biophys Acta 102:185-197
Gulyassy PF, Edelman IS (1967) Effect of pH and theophylline on uptake, elution and antidiuretic action of cyclic AMP. Am J Physiol 212:740-745
Harmanci MC, Kachadorian WA, Valtin H, Di Scala VA (1978) Antidiuretic hormone-induced intramembranous alterations in mammalian collecting ducts. Am J Physiol 235:F440-F443
Hays RM, LeafA (1962) The state of water in the isolated toad bladder in the presence and absence of vasopressin. J Gen Physiol 45:933-948
Hays RM, Franki N, Soberman R (1971)Activation energy for water diffusion across the toad bladder: evidence against the pore enlargement hypothesis. J Clin Invest 50:1016-1018
Humbert F, Montesano R, Grosso A, De Sousa RC, Orci L (1977) Particle aggregates in plasma and intracellular membranes of toad bladder granular cells. Experientia 33:1364-1367
Kachadorian WA, Wade JB, Discala VA (1975) Vasopressin-induced structural change in toad bladder luminal membrane. Science 190:67 69
Kachadorian WA, Casey C, Discala VA (1978) Time course ofADH-induced intramembranous particle aggregation in toad urinary bladder. Am J Physiol 234:F461-F465
Kachadorian WA, Muller J, Rudich SW, Discala VA (1981) Relation of ADH effects to altered membrane fluidity in toad urinary bladder. Am J Physiol 240:F63 F69
Muller J, Kachadorian WA, Discala VA (1980) Evidence that ADH-stimulated intramembranous particle aggregates are transferred from cytoplasmic to luminal membranes in the toad bladder epithelial cells. J Cell Biol 85:83 95

The rate limiting step in hydrosmotic action 355
Orloff J, Handler JS (1967) The role of adenosine 3'-5'-phosphate in the action of antidiuretic hormone. Am J Med 42:757-768
Parisi M, Chevalier J, Bourguet J (1979a) Influence of mucosal and serosal pH on antidiuretic action of frog urinary bladder. Am J Physiol 237:F483-F489
Parisi M, Ripoche P, Chevalier J, Bourguet J (1979b) A low HLB surfactant (NP-EO6) differently modifies water, sodium, urea and nicotinamide permeation in frog urinary bladder. In: "Hormonal control of epithelial transport". INSERM Symposium Series 85:289-300
Parisi M, Montereano R, Chevalier J, Bourguet J (1981) Cellular pH and water permeability control in frog urinary bladder: a possible action on the water pathway. Biochim Biophys Acta 648:267-274
Ripoche P (1967) Cin6tique fi basse temperature de la perm+abilisation ~i l'eau de la vessie de grenouille par l'ocytocine, le 3',5'-ad6nosine monophosphate et la th/~ophylline. M6moire pouv le Dipl6me d'Etudes Sup6rieures de Sciences Naturelles. Facult6 des Sciences. Universit6 de Paris
Ripoche P, Bourguet J, Parisi M (1973) The effect of hypertonic media on water permeability of frog urinary bladder. Inhibition by catecholamines and prostaglandin E 1. J Gen Physiol 51:1113~124
Wade JB (1980) Hormonal modulation of epithelial structure. Current topics in membrane and transport 13:123-147
Wade JB, Kachadorian WA, Discala VA (1977) Freeze-fracture electron microscopy: relationship of membrane structural features to transport physiology. Am J Physiol 232:F77-F83
Accepted September 21, 1982





























