Embed Size (px)
Citation preview

750 Research Paper
The regulation of Cdc20 proteolysis reveals a role for the APCcomponents Cdc23 and Cdc27 during S phase and early mitosisSusanne Prinz, Ellen S. Hwang, Rosella Visintin and Angelika Amon
Background: In eukaryotic cells, a specialized proteolysis machinery thattargets proteins containing destruction-box sequences for degradation and thatuses a ubiquitin ligase known as the anaphase-promoting complex/cyclosome(APC) plays a key role in the regulation of mitosis. APC-dependent proteolysistriggers the separation of sister chromatids at the metaphase–anaphasetransition and the destruction of mitotic cyclins at the end of mitosis. Recently,two highly conserved WD40-repeat proteins, Cdc20 and Cdh1/Hct1, havebeen identified as substrate-specific regulators of APC-dependent proteolysisin the budding yeast Saccharomyces cerevisiae. Here, we have investigated thecell cycle regulation of Cdc20 and Cdh1/Hct1.
Results: Whereas the levels of CDH1/HCT1 RNA and Cdh1/Hct1 protein areconstant throughout the cell cycle, CDC20 RNA and Cdc20 protein arepresent only during late S phase and mitosis and Cdc20 protein is unstablethroughout the entire cell cycle. The instability of Cdc20 depends on CDC23and CDC27, which encode components of the APC. During the G1 phase, adestruction box within Cdc20 mediates its instability, but during S phase andmitosis, although Cdc20 destruction is still dependent on CDC23 and CDC27,it does not depend on the Cdc20 destruction box.
Conclusions: There are remarkable differences in the regulation of Cdc20 andCdh1/Hct1. Furthermore, the APC activator Cdc20 is itself a substrate of theAPC-dependent proteolysis machinery, and the APC subunits Cdc23 andCdc27 have a role in the degradation of Cdc20 during S phase and earlymitosis that is not mediated by its destruction box.
BackgroundProgression through mitosis requires the precisely timedubiquitin-dependent degradation of specific substrates.Degradation of an as yet unidentified protein in eggextracts from Xenopus [1], of Pds1 in the budding yeastSaccharomyces cerevisiae [2] and Cut2 in Schizosaccharo-myces pombe [3] is required for the metaphase–anaphasetransition. Proteolysis of mitotic cyclins (cyclins A and B inhigher eukaryotes, Cdc13 in S. pombe, and Clb cyclins inS. cerevisiae; reviewed in [4]) is important for the inactiva-tion of mitotic kinase during exit from mitosis. Destruc-tion of Ase1 — a protein associated with the mitoticspindle — promotes disassembly of the mitotic spindleduring exit from mitosis [5]. The efficient degradation ofthese proteins requires that they have a specific aminoacid sequence termed the destruction box [6], which isusually located near the amino terminus of a protein.Deletion of this 9 amino acid motif from substrates pre-vents them from becoming multi-ubiquitinated and thusrapidly degraded (reviewed in [4,7–9]).
The machinery responsible for ubiquitinating mitoticcyclins and other proteins has been identified. It is
comprised of the ubiquitin-conjugating enzymes Ubc4and UbcX [10–13] and a ubiquitin ligase known as theanaphase promoting complex/cyclosome (APC) [11,14].The APC is composed of multiple subunits that includethe four previously identified proteins Cdc16, Cdc23,Cdc27 and Apc1/BimE [11,15–19] and the cullin homologApc2 [20,21]. Following ubiquitination, substrates aredegraded by the 26S proteasome (reviewed in [22]).
The stability of substrates of the APC-dependent proteol-ysis machinery varies greatly during the cell cycle. Sub-strates are stable during S phase, G2 and early mitosis(during which, for example, Clb2 has a half-life of morethan 2 hours), but are unstable during exit from mitosisand G1 (Clb2 has a half-life of less than 1 minute duringG1) [2,5,23–25]. Work using extracts from Xenopus andclam has shown that APC activity is a target for cell cycleregulation [11,14].
Regulation of the APC-dependent proteolysis machineryis complex. In S. cerevisiae, inactivation of APC-dependentproteolysis at the G1–S phase transition requires kinasesactivated by the G1 phase Cln cyclins [24]. Repression
Address: Whitehead Institute for BiomedicalResearch, 9 Cambridge Center, Cambridge,Massachusetts 02142, USA.
Correspondence: Angelika AmonE-mail: [email protected]
Received: 16 February 1998Revised: 20 April 1998Accepted: 7 May 1998
Published: 4 June 1998
Current Biology 1998, 8:750–760http://biomednet.com/elecref/0960982200800750
© Current Biology Ltd ISSN 0960-9822

during S phase and early mitosis requires the continuousactivity of Cln-associated and Clb-associated kinases [26].Activation of APC-dependent proteolysis during mitosisalso depends on cyclin-dependent kinase activity. Recon-stitution of cyclin B proteolysis in vitro using partially puri-fied components suggests that cyclin B–Cdc2 kinaseactivates APC-dependent proteolysis [14,27–29].Recently, several regulators of the APC-dependent prote-olysis machinery have been identified. Protein kinase Ahas been shown to inhibit APC activity [17,30] whereasthe protein kinase Cdc5/Polo/Polo-like kinase activatesthe APC [30–32]. Two highly conserved WD40-repeatproteins, Cdc20 and Cdh1/Hct1, have also been identifiedas activators of APC-dependent proteolysis [31,33,34]. Incdc20-1 mutants, degradation of Pds1 is defective but Clb2and Ase1 are degraded properly. Deletion of CDH1/HCT1results in stabilization of Clb2 and Ase1 but not of Pds1.Overexpression of CDC20 or CDH1/HCT1 is sufficient toinduce the APC-dependent proteolysis of the appropriatetarget in stages of the cell cycle when substrates are nor-mally stable. These findings define Cdc20 and Cdh1/Hct1as substrate-specific rate limiting activators of APC-dependent proteolysis.
As Cdc20 and Cdh1/Hct1 are critical regulators of APC-dependent proteolysis, it is important to understand howthese proteins are themselves regulated. Here, we reportthat the levels of Cdh1/Hct1 RNA and protein do not fluc-tuate during the cell cycle. In contrast, the levels of Cdc20RNA and protein are cell cycle regulated. CDC20 RNA isdetectable only during pheromone arrest, late S phase andmitosis. The transcriptional controls operative on CDC20might be similar to those operative on the gene encodingthe mitotic cyclin Clb2, as transcription of both CDC20 andCLB2 requires active mitotic kinase. Furthermore, Cdc20protein is unstable throughout the cell cycle. This instabil-ity is in part dependent on two APC subunits, Cdc23 andCdc27, suggesting that APC-dependent proteolysis ispartly responsible for the instability of Cdc20. Unlike otherAPC substrates, proteolysis of Cdc20 depends on CDC23and CDC27 throughout the cell cycle. During G1, the insta-bility of Cdc20 is mediated by a destruction box located inthe amino-terminal part of the protein. In contrast, theinstability of Cdc20 during S phase and mitosis, thoughdependent on CDC23 and CDC27, does not depend on thedestruction box within Cdc20. Our results suggest a role forthe APC in protein degradation during S phase and earlymitosis. Remarkably, for the degradation of Cdc20, thisactivity does not depend on the destruction box in Cdc20necessary for its degradation during G1.
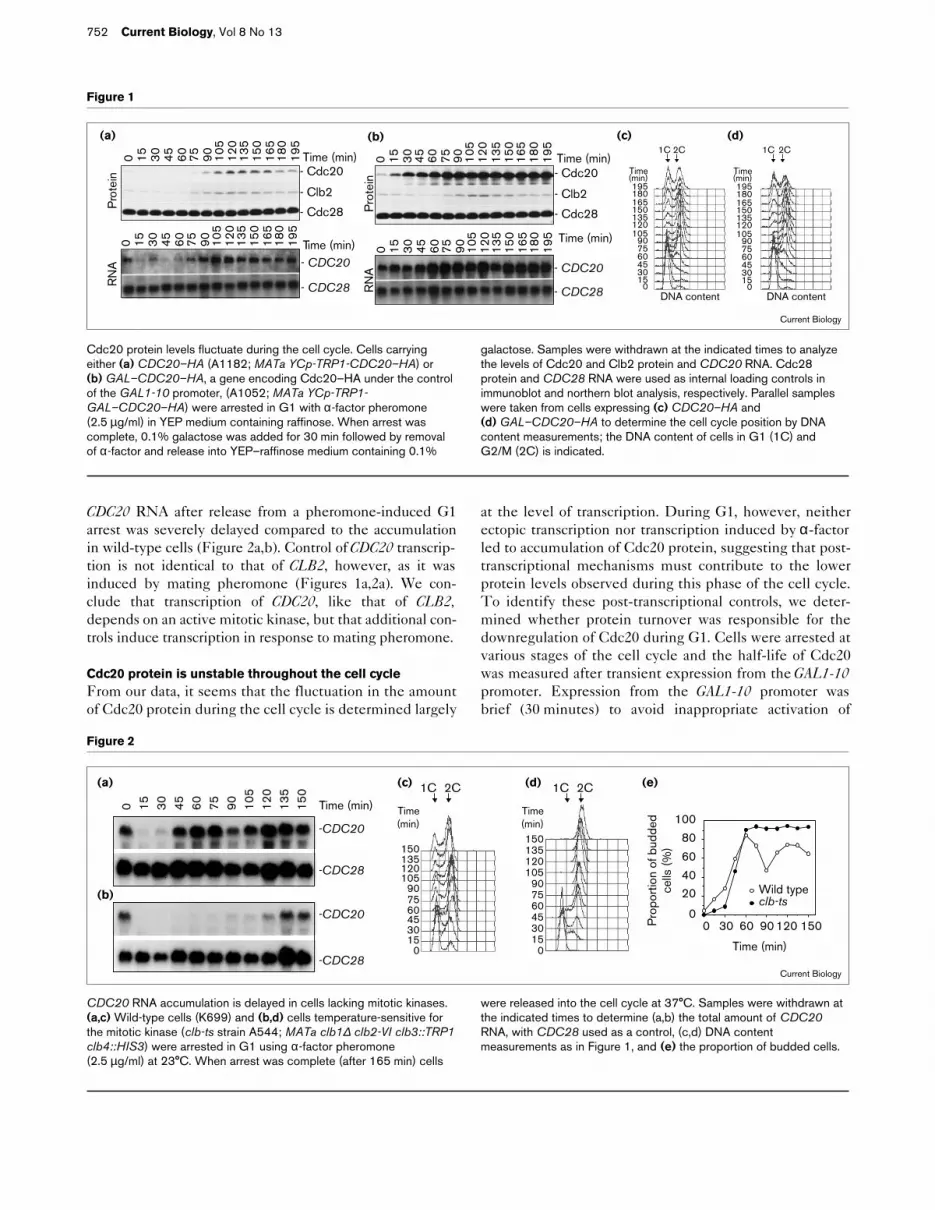
ResultsCdc20 protein levels are cell cycle regulatedAs a first step towards understanding how Cdc20 activityis regulated, we analyzed the levels of Cdc20 RNA andprotein during the cell cycle. To this end, we engineered a
gene (CDC20–HA) encoding Cdc20 tagged with a triplehemagglutinin epitope and under the control of the wild-type CDC20 promoter — a gene which complements thegrowth defect of a temperature sensitive cdc20-1 mutant.Cells carrying CDC20–HA were synchronized in G1 byaddition of the mating pheromone α-factor. Cells werethen released from the block and Cdc20 RNA and proteinlevels were analyzed. CDC20 RNA levels were high duringthe α-factor-induced G1 arrest, declined during S phaseand accumulated again during G2 and mitosis(Figures 1a,2a). The increased RNA levels in thepheromone-induced G1 arrest were not observed as cellsentered the subsequent G1 phase (Figure 1a,2a), suggest-ing that they were specific to the pheromone-induced G1arrest. Despite high levels of CDC20 RNA in pheromone-arrested cells, Cdc20 protein was absent during G1(Figure 1a) suggesting that post-transcriptional mecha-nisms downregulate Cdc20 protein during G1. Through-out the rest of the cell cycle, Cdc20 protein levels had acell cycle profile similar to that of CDC20 RNA and that ofthe mitotic cyclin protein Clb2 (Figure 1a). Cdc20 proteinwas absent during S phase, peaked when cells were inmitosis and declined as cells entered G1 (compareFigure 1a and 1c). We conclude that Cdc20 RNA andprotein levels fluctuate during the cell cycle, beingmaximal during mitosis.
To determine the degree to which transcriptional controlwas responsible for the fluctuation in Cdc20 protein levelsduring the cell cycle, we expressed CDC20 from the galac-tose-inducible GAL1-10 promoter. To allow moderate ratesof transcription, low levels of galactose (0.1%) were added.Expression of CDC20 from the GAL1-10 promoter afterrelease from a pheromone-induced G1 arrest gave levels ofCDC20 RNA that were mostly constant throughout the cellcycle (Figure 1b). Cdc20 protein levels were mostly con-stant throughout the cell cycle under these conditions,except during G1, when Cdc20 protein levels were signifi-cantly lower than in other stages of the cell cycle(Figure 1b, time point 0). Our results suggest that transcrip-tional regulation of CDC20 plays an important role inrestricting Cdc20 protein to late S phase and mitosis.During G1, additional post-transcriptional mechanisms helpdownregulate the Cdc20 protein, because neither ectopictranscription from the GAL1-10 promoter nor α-factor-induced transcription led to accumulation of Cdc20 protein.
Similar transcriptional controls may be operative onCDC20 and CLB2The similar cell cycle oscillation of CDC20 and CLB2 RNAprompted us to investigate whether CDC20 and CLB2 tran-scription are under similar controls. Transcription of CLB2depends on an active mitotic kinase; when grown at therestrictive temperature, strains temperature-sensitive for themitotic kinase (clb-ts strains) fail to accumulate CLB2 tran-scripts [35]. Similarly, in the clb-ts strain, accumulation of
Research Paper The regulation of Cdc20 proteolysis Prinz et al. 751
CDC20 RNA after release from a pheromone-induced G1arrest was severely delayed compared to the accumulationin wild-type cells (Figure 2a,b). Control of CDC20 transcrip-tion is not identical to that of CLB2, however, as it wasinduced by mating pheromone (Figures 1a,2a). We con-clude that transcription of CDC20, like that of CLB2,depends on an active mitotic kinase, but that additional con-trols induce transcription in response to mating pheromone.
Cdc20 protein is unstable throughout the cell cycleFrom our data, it seems that the fluctuation in the amountof Cdc20 protein during the cell cycle is determined largely
at the level of transcription. During G1, however, neitherectopic transcription nor transcription induced by α-factorled to accumulation of Cdc20 protein, suggesting that post-transcriptional mechanisms must contribute to the lowerprotein levels observed during this phase of the cell cycle.To identify these post-transcriptional controls, we deter-mined whether protein turnover was responsible for thedownregulation of Cdc20 during G1. Cells were arrested atvarious stages of the cell cycle and the half-life of Cdc20was measured after transient expression from the GAL1-10promoter. Expression from the GAL1-10 promoter wasbrief (30 minutes) to avoid inappropriate activation of
752 Current Biology, Vol 8 No 13
Figure 1
0 15 30 45 60 75 90 105
120
135
150
165
180
195
Time (min)- Cdc20
- Clb2
- Cdc28
(a)0 15 30 45 60 75 90 10
512
013
515
016
518
019
5
Time (min)- CDC20
- CDC28
Pro
tein
RN
A
0 15 30 45 60 75 90 105
120
135
150
165
180
195
Time (min)- Cdc20
- Clb2
- Cdc28
(b)
Pro
tein
RN
A0 15 30 45 60 75 90 10
512
013
515
016
518
019
5 Time (min)
- CDC20
- CDC28
1C 2C
90
Time(min)
015
456075
105
135150
180
30
120
165
195
DNA contentDNA content
1C 2C
Time(min)
015
45607590
105
135150
180
30
120
165
195
(c) (d)
Current Biology
Cdc20 protein levels fluctuate during the cell cycle. Cells carryingeither (a) CDC20–HA (A1182; MATa YCp-TRP1-CDC20–HA) or(b) GAL–CDC20–HA, a gene encoding Cdc20–HA under the controlof the GAL1-10 promoter, (A1052; MATa YCp-TRP1-GAL–CDC20–HA) were arrested in G1 with α-factor pheromone(2.5 µg/ml) in YEP medium containing raffinose. When arrest wascomplete, 0.1% galactose was added for 30 min followed by removalof α-factor and release into YEP–raffinose medium containing 0.1%
galactose. Samples were withdrawn at the indicated times to analyzethe levels of Cdc20 and Clb2 protein and CDC20 RNA. Cdc28protein and CDC28 RNA were used as internal loading controls inimmunoblot and northern blot analysis, respectively. Parallel sampleswere taken from cells expressing (c) CDC20–HA and(d) GAL–CDC20–HA to determine the cell cycle position by DNAcontent measurements; the DNA content of cells in G1 (1C) andG2/M (2C) is indicated.
Figure 2
1C 2C
Time (min)
150
90
015
456075
105
135
30
120
Time (min)150
90
015
456075
105
135
30
120
1C 2C
1501209060300
Time (min)
Pro
port
ion
of b
udde
d ce
lls (%
)
Wild typeclb-ts
0
2040
60
80
100
(a) (c)
(b)
(d) (e)
Time (min)
-CDC20
0 15 30 45 60 75 90 105
120
135
150
-CDC20
-CDC28
-CDC28
Current Biology
CDC20 RNA accumulation is delayed in cells lacking mitotic kinases.(a,c) Wild-type cells (K699) and (b,d) cells temperature-sensitive forthe mitotic kinase (clb-ts strain A544; MATa clb1∆ clb2-VI clb3::TRP1clb4::HIS3) were arrested in G1 using α-factor pheromone(2.5 µg/ml) at 23°C. When arrest was complete (after 165 min) cells
were released into the cell cycle at 37°C. Samples were withdrawn atthe indicated times to determine (a,b) the total amount of CDC20RNA, with CDC28 used as a control, (c,d) DNA contentmeasurements as in Figure 1, and (e) the proportion of budded cells.

Research Paper The regulation of Cdc20 proteolysis Prinz et al. 753
APC-dependent proteolysis. Half-life measurementsrevealed that Cdc20 was unstable throughout the cellcycle. The protein had a half-life of less than 3 minutes inexponentially growing cells (Figure 3a), during S phase(Figure 3d), during G2 and early mitosis (Figure 3e,f), andin cells arrested in anaphase (Figure 3h,i). During early G1,
Cdc20 seemed to be even less stable (Figure 3b). Theincreased instability of Cdc20 during G1 might contributeto the low levels of protein observed even when CDC20 isexpressed from the GAL1-10 promoter. Interestingly, incells arrested due to inactivation of the APC (Figure 3g),the half-life of Cdc20 was prolonged.
The endogenous Cdc20 protein was as unstable as Cdc20transiently expressed from the GAL1-10 promoter.Endogenous Cdc20 had a half-life of less than 5 minutesin cells arrested using pheromone and those arrested inS phase and early mitosis (Figure 4). Our results show thatCdc20 is a short lived protein throughout the cell cycle butthat it is partially stabilized in cells with a defective APC.
The instability of Cdc20 throughout the cell cycle dependson an active APCThe finding that Cdc20 was partially stabilized in an arrestdue to inactivation of the APC raised the possibility thatCdc20 instability was at least partly mediated by the APC.Cdc20 might be degraded by the APC only during exitfrom mitosis and during G1 — as are all other known APCsubstrates. The finding, however, that Cdc20, unlikeother APC substrates, was unstable throughout the cellcycle raised the possibility that Cdc20 was degraded bythe APC throughout the cell cycle. To distinguishbetween these possibilities, we analyzed the conse-quences of inactivating APC on the turnover of Cdc20 incells arrested at various stages of the cell cycle. Wearrested wild-type cells and cells defective for the APC(cells carrying a temperature sensitive cdc23-1 mutation)using either α-factor pheromone (Figure 5b), hydroxyurea(Figure 5c), or nocodazole (Figure 5d) at the permissivetemperature (23°C). When arrest was complete, cells wereshifted to 36°C to inactivate the cdc23-1 gene product and
Figure 3
Cdc20 is unstable throughout the cell cycle. Wild-type cells carryingGAL–CDC20–HA were either (a) left to cycle, or arrested at 36°C inYEP–raffinose medium at the indicated cell cycle stages using either(b) 5 µg/ml α-factor, (d) 10 mg/ml hydroxyurea or (f) 15 µg/mlnocodazole. (c,e,g–i) Various temperature-sensitive cell division cycle(cdc) mutants carrying GAL–CDC20–HA were arrested at 36°C atthe indicated cell cycle stages. After 3 h, galactose was added for30 min to induce the production of Cdc20. Then glucose (2%) andcycloheximide (1 mg/ml) were added to repress transcription andtranslation, respectively. Samples were taken at the indicated times todetermine the amount of Cdc20 protein and to determine cell cycleposition by DNA content measurements.
Cycling
α-factor(Early G1 phase)
cdc4-1(Late G phase)
Hydroxyurea(S phase)
cdc13-1(G2/M phase)
Nocodazole(G2/M phase)
cdc23-1(G2/M phase)
cdc5-1(Anaphase)
cdc15-2
(a)
(b)
(c)
(d)
(e)
(f)
(g)
(h)
(i)
(Anaphase)
- Cdc28
- Cdc20
50 10 30 Time (min)
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
Time(min)
1C 2C
Current Biology
Figure 4
The endogenous Cdc20 protein is unstable during G1, S phase andearly mitosis. Wild-type cells carrying CDC20–HA (A1182) werearrested using 15 µg/ml nocodazole, 10 mg/ml hydroxyurea or 5 µg/mlα-factor. The half-life of Cdc20–HA was measured as described inMaterials and methods, taking samples to determine (a) the level ofCdc20–HA protein at the indicated times after a pulse-chase with[35S]methionine and (b) the DNA content of arrested cells.
(b)
- Cdc20
50 15 4050 15 40α-factor(a)
Time (min)50 15 40Hydroxyurea Nocodazole
α-factor Hydroxyurea Nocodazole
1C 2C1C 2C1C 2CCurrent Biology

the half-life of Cdc20 was measured after transient expres-sion from the GAL1-10 promoter (see Materials andmethods). The cdc23-1 mutation caused a significantincrease in the half-life of Cdc20 in all the cell cycle stagesanalyzed: in exponentially growing cells (Figure 5a), andcells arrested in G1 (Figure 5b), S phase (Figure 5c), andmitosis (Figure 5d). Defects in the APC subunit Cdc27[11,19] also caused stabilization of Cdc20. In temperature-sensitive cdc27-A mutants, Cdc20 was stabilized in all thecell cycle stages analyzed (Figure 5a–d). The finding thatthroughout the cell cycle proteolysis of Cdc20 dependedon the APC subunits Cdc23 and Cdc27 indicates thatAPC-mediated proteolysis is responsible for the instabilityof Cdc20 throughout the cell cycle.
We have previously shown that prolonged overexpressionof CDC20 induces ectopic activation of APC-dependent
proteolysis [33]. Thus, expression of CDC20 from theGAL1-10 promoter, though brief, might inappropriatelyactivate APC-dependent proteolysis in stages of the cellcycle when it is normally inactive. This would result inCdc20 appearing to be unstable throughout the cell cycle.To test this possibility, we constructed a strain thatexpressed the genes for both the APC substrates Pds1 andCdc20 under the control of the GAL1-10 promoter. Afterbrief transient expression from the GAL1-10 promoter, wemeasured the half-life of Pds1 and Cdc20. Whereas Cdc20was unstable during G1, S phase and mitosis, Pds1 stabil-ity was characteristic of that of other APC substrates — itwas unstable during G1, but stable during S phase andearly mitosis (Figure 6). This result shows that the insta-bility of Cdc20 throughout the cell cycle that is depen-dent on CDC23 and CDC27 was not due to inappropriateactivation of APC-dependent proteolysis brought about
754 Current Biology, Vol 8 No 13
Figure 5
- Cdc28
- Cdc20
50 10 3050 10 30
cdc23-1
0
30
Time(min)
1C 2C
0
30
Time(min)
1C 2C
cdc23-1
(d)
- Cdc28
- Cdc20
Time (min)50 10 3050 10 30cdc23-1
0
30
Time(min)
1C 2C
0
30
Time(min)
1C 2C
(c)50 10 30
cdc27-A
cdc27-A
50 10 30
0
30
Time(min)
1C 2C
0
30
Time(min)
1C 2C
cdc27-A0
30
Time(min)
1C 2C
0
30
Time(min)
1C 2C
cdc23-1
(b)
- Cdc28
- Cdc20
50 10 3050 10 30Wild type
Wild type Wild type
cdc23-150 10 30
cdc27-A
- Cdc28
- Cdc20
50 10 3050 10 30Wild type
Wild type
Current Biology
Wild typecdc23-1(a)
Time (min)
Time (min)Time (min)
50 10 30cdc27-A
1C 2C
0
30
Time(min)
0
30
Time(min)
1C 2C
0
30
Time(min)
1C 2C
0
30
Time(min)
1C 2C
cdc27-A
cdc23-1 cdc27-Acdc23-1Wild type Wild typecdc27-A
The instability of Cdc20 during the cell cycle depends on CDC23 andCDC27. Wild-type cells (A1052), cdc23-1 mutants (A1178) andcdc27-A mutants (A1391) carrying GAL–CDC20–HA were either(a) grown to exponential phase, or arrested with either (b) 5 µg/mlα-factor, (c) 10 mg/ml hydroxyurea, or (d) 15 µg/ml nocodazole inYEP–raffinose medium at 23°C. After 3 h, galactose was added for
30 min followed by a shift to 36°C for another 30 min. Transcriptionand translation were repressed by the addition of glucose (2%) andcycloheximide (1 mg/ml), respectively (at time zero). Samples weretaken at the indicated times to determine the amount of Cdc20 proteinand the cell cycle position by DNA content measurements.

Research Paper The regulation of Cdc20 proteolysis Prinz et al. 755
by transient ectopic expression of CDC20 from theGAL1-10 promoter. Our results suggest that, in contrast toother APC substrates, APC-dependent proteolysis ofCdc20 is active throughout the cell cycle.
A Cdc20 destruction box contributes to its instabilityduring G1 but not during other stages of the cell cycleCdc20 contains two destruction-box-like motifs in itsamino terminus (Box1 and Box2; Figure 7a). To deter-mine whether the instability of Cdc20 that is dependenton CDC23 and CDC27 is mediated by these destructionboxes, we constructed Cdc20 proteins lacking either oneor both of the boxes. Deletion of Box2 did not cause achange in the half-life of Cdc20 in cells arrested in G1, Sphase or mitosis (Figure 7b–e). In contrast, deletion ofBox1 stabilized Cdc20 but, surprisingly, not throughoutthe cell cycle. Deletion of Box1 caused stabilization inG1 (Figure 7c) to the same extent as did inactivation of
the APC (Figure 5b). In contrast, deletion of Box1 hadno stabilizing effects during S phase or mitosis(Figure 7d,e). Deleting both Box1 and Box2 did not sta-bilize Cdc20 any more than did deleting Box1 alone,confirming that Box2 did not contribute to the instabilityof Cdc20 (Figure 7b–e).
Our data suggest that the APC subunits Cdc23 and Cdc27are required for degradation of Cdc20 throughout the cellcycle. APC-dependent proteolysis acts on Cdc20 in twoways, however. APC-dependent proteolysis that is medi-ated by the Cdc20 destruction box is required for degrad-ing Cdc20 during G1, whereas a previously unidentifiedfunction of the APC subunits Cdc23 and Cdc27, which isnot mediated by the destruction box of Cdc20, is neces-sary for degrading Cdc20 during S phase and early mitosis.
Cdh1/Hct1 RNA and protein levels are constant throughoutthe cell cycleThe role of the Cdc20 homolog Cdh1/Hct1 in the degra-dation of the mitotic cyclin Clb2 and the mitotic-spindle-associated protein Ase1 is thought to be analogous to that
Figure 6
Cdc20, but not Pds1, is unstable throughout the cell cycle. Various cdcmutants and wild-type cells carrying both GAL–CDC20–HA and aGAL–PDS1–HA fusion were arrested as described in the legend toFigure 3. The half-lives of Cdc20 and Pds1 were measured aftertransient expression from the GAL1-10 promoter. Samples were taken atthe indicated times to determine the amount of Cdc20 and Pds1 proteinand to determine cell cycle position by DNA content measurements.
Cycling
α-factor(Early G1phase)
Hydroxyurea(S phase)
cdc13-1(G2/M phase)
Nocodazole(G2/M phase)
cdc23-1(G2/M phase)
- Cdc28
- Cdc20100 30 60 Time (min)
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
Pds1
Pds1
Pds1
Pds1
- Cdc28
- Cdc20 Pds1
Pds1
0
30
0
30
0
30
0
30
0
30
0
30
Time(min)
1C 2C
Current Biology
Figure 7
The destruction box of Cdc20 contributes to the instability of Cdc20during G1. (a) Alignment of the destruction boxes of Clb2, Pds1 andCut2 with two putative destruction boxes of Cdc20, Box1 and Box2;the residue number of the first amino acid in each sequence isindicated and identical residues are shown in bold. (b–e) Cells(A1242) carrying GAL–CDC20–HA or the same construct lackingeither the region encoding Box1 (∆Box1), the region encoding Box2(∆Box2), or both these regions (∆Box1+2) were either (b) grown toexponential phase, or arrested using either (c) α-factor,(d) hydroxyurea, or (e) nocodazole. The half-life of Cdc20 and Cdc20lacking one or both destruction boxes was determined as described inthe legend to Figure 3.
Wild type ∆ Box1Time (min)0 5 10300 5 10300 5 10300 5 1030- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
- Cdc20
- Cdc28
∆ Box1+2∆ Box2
Cycling
α-factor
Hydroxyurea
Nocodazole
(b)
(c)
(d)
(e)
Clb2 25 RLALNNVTNPds1 85 RLPLAAKDNCut2 33 RAPLGSTKQ
52 RTVLGGKSTCdc20 Box1 17 RSVLSIASP
Box2 60 RPSLQASAN
(a)
Current Biology

of Cdc20 in the degradation of Pds1 [33,34]. To determinewhether the cell cycle control of Cdh1/Hct1 and Cdc20are similar, we analyzed the levels of Cdh1/Hct1 RNA andprotein during the cell cycle. Cells carrying a functionalMYC-tagged CDH1/HCT1 gene (CDH1–MYC; see Materi-als and methods) were synchronized in G1 by addition ofthe mating pheromone α-factor. After release from theblock, Cdh1/Hct1 RNA and protein levels were analyzed.As shown in Figure 8, the levels of both Cdh1/Hct1 RNAand protein were constant throughout the cell cycle. It isworth noting that the migration of Cdh1/Hct1 protein inpolyacrylamide gels containing sodium dodecyl sulfate(SDS) varies during the cell cycle. Cdh1/Hct1 proteinextracted from G1-phase cells migrates faster thanCdh1/Hct1 obtained from mitotic cells (compare the 0 and75 minute time points in Figure 8b). We do not know thereason for this differential migration, but phosphorylationhas been shown to alter migration of proteins inSDS–polyacrylamide gels. Our result suggests thatalthough Cdc20 might perform a similar role in the degra-dation of Pds1 to that of Cdh1/Hct1 in the degradation ofClb2 and Ase1, there are remarkable differences in theirregulation during the cell cycle.
DiscussionTranscriptional controls help to restrict Cdc20 activity tolate stages of the cell cycleWe have characterized the multiple controls operative onCdc20 that restrict its activity to late stages of the cellcycle. In contrast to the levels of Cdh1/Hct1, which areconstant throughout the cell cycle, Cdc20 levels fluctuateduring the cell cycle. The amount of Cdc20 in a cell isregulated at the level of transcription, which appears to beresponsible mainly for restricting Cdc20 protein to latestages of the cell cycle. When CDC20 is expressed at lowlevels from the GAL1-10 promoter, Cdc20 protein ispresent at constant levels throughout the cell cycle assoon as cells enter S phase. The transcriptional controlsoperative on the CDC20 promoter might be similar tothose operative on the CLB2 and SWI5 promoters [35].
Transcription of CDC20, like that of CLB2 and SWI5,depends on an active mitotic kinase. Thus, the mitotickinases activate APC-dependent proteolysis by inducingtranscription of the critical activator CDC20.
Transcription of CDC20 is also distinct from that of CLB2and SWI5. CDC20 transcription is induced by matingpheromone which may be important to ensure continuousAPC-dependent proteolysis activity during pheromonearrest [36]. We have previously shown that CDC20 isrequired for Pds1 degradation during a pheromone-induced G1 arrest [33]. During G1, however, Cdc20 ishighly unstable and does not accumulate to levels that aredetectable by immunoblot analysis (Figure 1) [31]. Thefinding that CDC20 transcription is highly induced duringthe pheromone arrest might help to explain this apparentdiscrepancy. Although Cdc20 is highly unstable duringG1, the protein is nevertheless synthesized at high levelsduring the arrest (Figure 4). Thus, Cdc20 might be able toaffect APC-dependent protein degradation. Before beingdegraded by the APC, Cdc20 might modify the APC.Alternatively, Cdc20 might target substrates such as Pds1for degradation and by doing so becomes degraded itselfby the APC.
Cdc20 is degraded by a pathway dependent on CDC23 andCDC27 and by a pathway independent of CDC23 and CDC27 The half-life of Cdc20 is less than 3 minutes in exponen-tially growing cells. In cdc23-1 or cdc27-A mutants, the half-life of Cdc20, although prolonged compared to wild-typecells, is still short. One possible explanation for thisfinding is that the defective APC components expressedby cdc23-1 and cdc27-A mutants are not completely inac-tive. We do not think that this is likely as other APC sub-strates are dramatically stabilized in cdc23-1 and cdc27-Amutants under similar experimental conditions (S.P. andA.A., unpublished observations) [15]. We thereforebelieve that APC-dependent proteolysis is not the onlymechanism responsible for degrading Cdc20: there mustalso be a mechanism that is independent of CDC23 and
756 Current Biology, Vol 8 No 13
Figure 8
Cdh1/Hct1 RNA and protein levels areconstant during the cell cycle. (a) Extractsfrom wild-type cells (no tag, K699) and cellscarrying CDH1–MYC (CDH1–MYC, A1342)were probed with an anti-Myc antibody tovisualize Cdh1–Myc. (b–d) Cells carryingCDH1–MYC (A1342) were arrested in G1using α-factor pheromone (2.5 µg/ml). After2 h, cells were released from the block andsamples were withdrawn at the indicatedtimes to analyze the amount of (b) Cdh1/Hct1protein and (c) CDH1/HCT1 RNA. (d) Inparallel, samples were taken to determine cellcycle position by DNA content measurements.
(a)
- Cdh1
- No
tag
- Cdh
1–M
yc
66-97-kDa
Time (min)
(c)R
NA
0 15 30 45 60 75 90 105
120
135
150
165
- Cdh1/Hct1
- Cdc28
(b)
Pro
tein
(d) 1C 2C
90
Time(min)
0
456075
105
135150
180
30
120
165
0 15 30 45 60 75 90 105
120
135
150
165
Time (min)
- CDH1/HCT1
- CDC28
Current Biology

CDC27, about which we know next to nothing. It isunclear whether it is ubiquitin dependent and which partof Cdc20 mediates this mode of proteolysis, but it appearsto be active throughout the cell cycle. The half-life ofCdc20 in a cdc23-1 or cdc27-A mutant is similar whethercells are arrested in S phase, mitosis or G1.
Two modes of protein degradation dependent on CDC23and CDC27We have shown that the instability of Cdc20 during G1 isdependent on the APC subunits Cdc23 and Cdc27. Thisfinding, together with the finding that during G1 theinstability of Cdc20 is dependent on its destruction box,strongly suggests that APC-dependent proteolysis isresponsible for the instability of Cdc20 during G1. Similarfindings were recently reported by Shirayama et al. [31].Here, we have also shown that the instability of Cdc20during S phase and early mitosis is independent of thisdestruction box, although the APC components Cdc23and Cdc27 are required for the degradation of Cdc20during these cell cycle stages. It is possible that Cdc23 andCdc27 are not only APC components but that they are alsocomponents of an as yet unidentified complex that medi-ates protein degradation during S phase and early mitosis.Given that the instability of Cdc20 during G1 shows allthe characteristics of APC-dependent proteolysis and thatdegradation during S phase and early mitosis requires theAPC subunits Cdc23 and Cdc27, however, we favor theidea that the requirement of CDC23 and CDC27 for Cdc20degradation during S phase and early mitosis reflects arequirement for APC-dependent proteolysis during thesecell cycle stages. We do not know yet whether Cdc23 andCdc27 directly ubiquitinate Cdc20 during S phase andearly mitosis. Irrespective of whether the effects of inacti-vating CDC23 or CDC27 on Cdc20 stability are direct orindirect, however, our data demonstrate a previouslyunknown function for Cdc23 and Cdc27, and possibly theAPC, during S phase and early mitosis.
We propose that APC has at least two functions. One ismediated by destruction boxes in its substrates, the otherone is not. The proteolysis that is dependent on destruc-tion boxes, as previously reported for numerous APC sub-strates, is cell cycle regulated, being active during G1 butinactive during S phase and early mitosis. For Cdc20,another proteolytic activity is mediated by Cdc23 andCdc27, but this is not dependent on either of Cdc20’sdestruction boxes. Throughout the remainder of this dis-cussion, we will term this activity the ‘S/early M,CDC23/CDC27-dependent proteolysis machinery’. Weknow little about the S/early M, CDC23/CDC27-depen-dent proteolysis machinery. The part of Cdc20 that medi-ates this mode of protein degradation has to be identified.Cryptic motifs similar to destruction boxes or new motifscould mediate this activity. Furthermore, the degradationmachinery itself needs to be characterized. Whether other
proteins are degraded by the S/early M CDC23/CDC27-dependent proteolysis machinery also remains to be deter-mined. It is tempting to speculate that proteins importantfor restricting DNA replication to once per cell cycle[37,38] are degraded by this machinery.
Our data indicate that the S/early M, CDC23/CDC27-dependent proteolysis machinery is active during S phaseand early mitosis but might not be very active during G1.Deletion of Box1 of Cdc20 prolongs the half-life of Cdc20during G1 almost to the extent seen in cdc23-1 mutants,suggesting that proteolysis mediated by this destructionbox is the main mechanism responsible for the instabilityof Cdc20 during the G1 phase. Our data also suggest thatthe S/early M, CDC23/CDC27-dependent proteolysismachinery might not be as active as the APC-dependentprotein degradation machinery that requires destructionboxes. Cdc20 appears less stable in G1-arrested cells thanin cells arrested using hydroxyurea or nocodazole(Figures 3,5,6).
Is there a role for the APC during S phase and early mitosis?We found that Cdc20 is unstable throughout the cell cycleand that this instability depends on two APC components,Cdc23 and Cdc27. During different stages of the cellcycle, however, Cdc23 and Cdc27 function with differentmodes. Because of the complex pattern of protein degra-dation responsible for keeping Cdc20 unstable throughoutthe cell cycle and because the part of Cdc20 mediating theS/early M, CDC23/CDC27-dependent proteolysis has notbeen identified, we have not been able to directly analyzethe consequences of expressing Cdc20 that is stablethroughout the cell cycle on cell cycle progression. Dele-tion of the destruction box stabilizes Cdc20 only duringG1 which has no consequences on cell cycle progressionwhen it is expressed under the control of the CDC20 pro-moter (data not shown). Regulating the amount of Cdc20protein might, however, be critical for cell cycle progres-sion and cell viability. We have previously shown thatectopic expression of CDC20 from the GAL1-10 promoteris lethal [33]. Cells cease to divide, but do not arrest at aspecific stage of the cell cycle. Overexpression of Cdc20 isalso sufficient to cause a bypass of the DNA damage andmitotic spindle assembly checkpoint arrest [39]. Thus,keeping Cdc20 protein levels under tight control is notonly critical for normal cell cycle progression but mightalso be important during a cell cycle arrest induced byDNA damage or by mitotic spindle damage.
Cdc20 versus Cdh1/Hct1 regulationAlthough Cdc20 and Cdh1/Hct1 are highly related proteinsand probably perform similar roles in the degradation ofPds1, and Clb2 and Ase1, respectively, their roles and reg-ulation during the cell cycle differ remarkably. Cdc20 playsa critical role in the activation of Pds1 degradation [31,33].CDC20 is also required to initiate Clb degradation: cdc20-1
Research Paper The regulation of Cdc20 proteolysis Prinz et al. 757

758 Current Biology, Vol 8 No 13
mutants arrest in metaphase with high levels of stable Clb2[31] (S.P. and A.A., unpublished observations) whichreflects the requirement of Pds1 destruction for the initia-tion of the proteolytic program responsible for degradingClb cyclins. Cdc20 also performs an essential functionduring exit from mitosis. Deletion of PDS1 in a cdc20-1mutant causes cdc20-1 cells to escape the metaphase arrest,but cells go on to arrest in anaphase instead [31,40]. CDC20may also play a role in mitotic spindle dynamics. At therestrictive temperature, cdc20-1 mutants form abnormalmitotic spindles containing excessive numbers of micro-tubules [41,42]. Thus, Cdc20 plays a central role in the ini-tiation of the APC-dependent proteolytic program thattriggers the metaphase–anaphase transition and exit frommitosis. In contrast, Cdh1/Hct1 is dispensable for themetaphase–anaphase transition but plays an important,though not essential, role during exit from mitosis and G1.Cdh1/Hct1 triggers degradation of mitotic cyclins and Ase1during these stages of the cell cycle [33,34].
The different roles of Cdc20 and Cdh1/Hct1 in the regula-tion of APC-mediated proteolysis and the differences intheir regulation might allow us to explain the substrate-specific differences in APC-mediated protein degradation.It could explain why APC-mediated proteolysis, althoughactivated at the metaphase–anaphase transition, com-pletely degrades Pds1 at this transition, but does not com-plete degradation of mitotic cyclins until exit from mitosis(at the telophase–G1 transition). The Cln-dependent andClb-dependent kinases are continuously required toprevent activation of APC-dependent proteolysis that ismediated by a destruction box during S phase and earlymitosis [24,26]. We speculate that Cdh1/Hct1, whoseprotein levels are constant throughout the cell cycle, isinhibited by Cln-dependent and Clb-dependent kinases.In contrast, Cdc20, which is not present during S phaseand early mitosis might be less susceptible to theinhibitory activity of kinases dependent on Cln and Clb.Therefore, when synthesized during mitosis, Cdc20 initi-ates the proteolytic program responsible for progressionthrough mitosis even in the presence of high levels of Cln-dependent and Clb-dependent kinase activity. In contrast,Cdh1/Hct1 cannot become active until mitotic kinases areat least partially inactivated. But how is mitotic kinaseinactivation initiated? Cdc20 might initiate inactivation byone or more of several mechanisms, including promotingaccumulation of the inhibitor of cyclin-dependent kinasesSic1 or inducing degradation of a fraction of mitoticcyclins, which would then allow activation of Cdh1/Hct1and accumulation of Sic1.
ConclusionsOur results suggest that RNA and protein levels of theAPC activator Cdh1/Hct1 are constant throughout thecell cycle. In contrast, the APC activator Cdc20 is regu-lated at multiple levels. CDC20 RNA levels are cell cycle
regulated, being maximal during mitosis. The transcrip-tional controls operative on CDC20 might be similar tothose regulating CLB2, as transcription of both CLB2 andCDC20 depends on an active mitotic kinase. Further-more, Cdc20 is unstable throughout the cell cycle andthis instability is partly dependent on two APC subunits,Cdc23 and Cdc27, suggesting that it depends on theAPC-dependent proteolysis machinery. Surprisingly, twotypes of proteolytic mechanisms that are dependent onCDC23 and CDC27 act on Cdc20. One mechanism, whichis dependent on the destruction box of Cdc20, is activeduring exit from mitosis and G1, whereas the secondmode is responsible for the instability of Cdc20 duringS phase and early mitosis and does not require the Cdc20destruction box. Our results suggest that there is a previ-ously unidentified activity for the APC-dependent prote-olysis machinery that is independent of destruction boxesand that acts during S phase and early mitosis.
Materials and methodsPlasmid and strainsAll strains were derivatives of strain W303 (K699; MATa ade2-1leu2-3 ura3 trp1-1 his3-11,15 can1-100 GAL psi+). DNA manipula-tions were performed according to published methods [43]. To createCDC20–HA, DNA encoding a triple HA tag was inserted in frame atthe BstEII site 40 bp downstream of the initiation codon in the CDC20gene or at the MluI site 510 bp downstream of it. Functionality of thefusions was determined by their ability to complement the growthdefect of a temperature-sensitive cdc20-1 mutant strain. The fusionswere then cloned such that they were under the control of theGAL1-10 promoter as described [33] and placed into plasmidYCplac22 [44]. The destruction boxes within Cdc20 (Box1, aminoacids 17 to 25; Box2, amino acids 60 to 68) were deleted using theChameleon double-stranded, site-directed mutagenesis kit (Strata-gene) according to the manufacturers’ instructions. To createCDH1–MYC, a triple MYC tag was integrated at the initiation codon ofthe chromosomal CDH1/HCT1 locus as described [45].
Growth conditionsConditions for growth and release of synchronous cultures from arrestby α-factor (Figures 1,2,8) were as described [46], except cells werereleased into YEP (yeast extract, peptone) medium containing raffinoseand 0.1% galactose in the experiment described in Figure 1. To deter-mine the half-life of Cdc20 and versions of Cdc20 lacking either one orboth destruction boxes in cells arrested at various stages of the cellcycle (Figures 3,6,7), cells were grown in YEP–raffinose medium at23°C. Cells were then shifted to 37°C for 3 h and 5 µg/ml α-factor,10 mg/ml hydroxyurea or 15 µg/ml nocodazole was added as appropri-ate. Thereafter, galactose was added for 30 min. Then 2% glucose and1 mg/ml cycloheximide were added to repress transcription and trans-lation, respectively. The amount of Cdc20 was then analyzed byimmunoblot analysis.
To determine the half-life of the endogenous Cdc20 protein(Figure 4), cells were arrested with α-factor, hydroxyurea or nocoda-zole in medium containing 0.1 mM methionine. Cells from a 20 mlculture were then harvested by filtration, washed with two volumes ofmedium lacking methionine and containing glucose (–metD medium)and resuspended in 2 ml –metD medium. After a 4 min incubation,cells were labeled with 0.5 mCi 35S-Translabel (ICN; 85%[35S]methionine, 15% [35S]cysteine) for 7 min. Cells were then cen-trifuged and resuspended at time zero in YEPD (yeast extract,polypeptone and dextrose) medium containing 2 mM methionine,2 mM cysteine and 1 mg/ml cycloheximide.

Research Paper The regulation of Cdc20 proteolysis Prinz et al. 759
To determine whether the half-life of Cdc20 was dependent on a func-tional APC throughout the cell cycle (Figure 5), cells were arrested at23°C (the permissive temperature for the cdc23-1 and the cdc27-Amutations) using α-factor (5 µg/ml), hydroxyurea (10 mg/ml), or noco-dazole (15 µg/ml). After 3 h, galactose was added for 30 min, fol-lowed by a shift to 36°C for another 30 min. Then 2% glucose and1 mg/ml cycloheximide were added to repress transcription and trans-lation, respectively. The amount of Cdc20 was then analyzed byimmunoblot analysis.
TechniquesImmunoblot analysis of the total amount of Clb2, Cdc28, Cdc20–HAand Cdh1–Myc was determined as described [47]. The dilutions usedwere 1:2000 for anti-Clb2, anti-HA and anti-Cdc28 antibodies and1:500 for anti-Myc antibody. Equal loading of gel lanes was shown byprobing blots with an anti-Cdc28 antibody. All other techniques wereperformed as described ([35] and references therein). Immunoprecipi-tation of 35S-labeled Cdc20 was performed as described [48] using anHA-affinity matrix distributed by BABCO.
AcknowledgementsWe thank Steve Bell, Terry Orr-Weaver, Brian Cali and Jacqueline Lopez forhelpful comments and discussions during the course of this project and fortheir critical reading of the manuscript. This research was supported by anErwin Schroedinger Fellowship to S.P. and a National Institutes of Healthgrant GM 56800-01 to A.A.
References1. Holloway SL, Glotzer M, King RW, Murray AW: Anaphase is initiated
by proteolysis rather than by the inactivation of maturation-promoting factor. Cell 1993, 73:1393-1402.
2. Cohen-Fix O, Peters J-M, Kirschner MW, Koshland D: Anaphaseinitiation in Saccharomyces cerevisiae is controlled by the APC-dependent degradation of the anaphase inhibitor Pds1p. GenesDev 1996, 10:3081-3093.
3. Funabiki H, Yamano H, Kumada K, Nagao K, Hunt T, Yanagida M:Cut2 proteolysis required for sister-chromatid separation infission yeast. Nature 1996, 381:438-441.
4. King RW, Deshaies RJ, Peters J-M, Kirschner MW: How proteolysisdrives the cell cycle. Science 1996, 274:1652-1659.
5. Juang Y-L, Huang J, Peters J-M, McLaughlin ME, Tai C-Y, Pellman D:APC-mediated proteolysis of Ase1 and the regulation of themitotic spindle. Science 1997, 275:1311-1314.
6. Glotzer M, Murray AW, Kirschner MW: Cyclin is degraded by theubiquitin pathway. Nature 1991, 349:132-138.
7. Murray AW: Cyclin ubiquitination: the destructive end of mitosis.Cell 1995, 81:149-152.
8. Glotzer M: The only way out of mitosis. Curr Biol 1995, 5:970-972.9. Deshaies RJ: The self-destructive personality of a cell cycle
transition. Curr Opin Cell Biol 1995, 7:781-789. 10. Hershko A, Ganoth D, Sudakin V, Dahan A, Cohen LH, Luca FC, et
al.: Components of a system that ligates cyclin to ubiquitin andtheir regulation by the protein kinase Cdc2. J Biol Chem 1994,269:4940-4946.
11. King RW, Peters J-M, Tugendreich S, Rolfe M, Hieter P, KirschnerMW: A 20S complex containing CDC27 and CDC16 catalyzes themitosis-specific conjugation of ubiquitin to cyclin B. Cell 1995,81:279-288.
12. Aristarkhov A, Eytan E, Moghe A, Admon A, Hershko A, Ruderman JV:E2-C, a cyclin-selective ubiquitin carrier protein required for thedestruction of mitotic cyclins. Proc Natl Acad Sci USA 1996,93:4294-4299.
13. Yu H, King RW, Peters J-M, Kirschner MW: Identification of a novelubiquitin-conjugating enzyme involved in the mitotic cyclindegradation. Curr Biol 1996, 6:455-466.
14. Sudakin V, Ganoth D, Dahan A, Heller H, Hershko J, Luca FC, et al.:The cyclosome, a large complex containing cyclin selectiveubiquitin ligase activity targets cyclins for destruction at the endof mitosis. Mol Biol Cell 1995, 6:185-198.
15. Irniger S, Piatti S, Michaelis C, Nasmyth K: Genes involved in sisterchromatid separation are needed for B-type cyclin proteolysis.Cell 1995, 87:269-277.
16. Tugendreich S, Tomkiel J, Earnshaw W, Hieter P: CDC27Hscolocalizes with CDC16Hs to the centrosome and mitotic spindleand is essential for the metaphase to anaphase transition. Cell1995, 81:261-268.
17. Yamashita YM, Nakaseko Y, Samejima I, Kumada K, Yamada H,Michaelson D, Yanagida M: 20S cyclosome complex formation andproteolytic activity inhibited by the cAMP/PKA pathway. Nature1996, 384:276-279.
18. Peters J-M, King RW, Hoog C, Kirschner MW: Identification of BIMEas a subunit of the anaphase promoting complex. Science 1996,274:1199-1201.
19. Zachariae W, Shin TH, Galova G, Obermaier B, Nasmyth K:Identification of subunits of the anaphase-promoting complex ofSaccharomyces cerevisiae. Science 1996, 274:1201-1204.
20. Yu H, Peters J-M, King RW, Page AM, Hieter P, Kirschner MW:Identification of a cullin homolog region in a subunit of theanaphase-promoting complex. Science 1998, 279:1219-1222.
21. Zachariae W, Shevchenko A, Andrews PD, Galova M, Stark MJR,Mann M, Nasmyth K: Mass spectometric analysis of the anaphasepromoting complex from budding yeast: identification of asubunit related to cullins. Science 1998, 279:1216-1219.
22. Hochstrasser M: Ubiquitin, proteasomes, and the regulation ofintracellular protein degradation. Curr Biol 1995, 7:215-223.
23. Evans T, Rosenthal ET, Youngblom J, Distel D, Hunt T: Cyclin: aprotein specified by maternal mRNA in sea urchin eggs that isdestroyed at each cleavage division. Cell 1983, 33:389-396.
24. Amon A, Irniger S, Nasmyth K: Closing the cell cycle circle in yeast:G2 cyclin proteolysis initiated at mitosis persists until the activationof G1 cyclins in the next cell cycle. Cell 1994, 77:1037-1050.
25. Brandeis M, Hunt T: The proteolysis of mitotic cyclins inmammalian cells persists from the end of mitosis until the onsetof S phase. EMBO J 1996, 15:5280-5289.
26. Amon A: Regulation of B-type cyclin proteolysis by Cdc28-associated kinases in budding yeast. EMBO J 1997, 16:2693-2702.
27. Felix MA, Labbe J C, Doree M, Hunt T, Karsenti E: Triggering ofcyclin degradation in interphase extracts of amphibian eggs bycdc2 kinase. Nature 1990, 346:379-382.
28. Luca FC, Shibuya EK, Dohrmann CD, Ruderman JV: Both cyclinAD60 and BD97 are stable and arrest cells in M-phase, but only
cyclin BD97 turns on cyclin destruction. EMBO J 1991, 10:4311-4320.
29. Lahav-Baratz S, Sudakin V, Ruderman JV, Hershko A: Reversiblephosphorylation controls the activity of cyclosome associatedcyclin-ubiquitin ligase. Proc Natl Acad Sci USA 1995, 92:9303-9307.
30. Kotani S, Tugendreich S, Fujii M, Jorgensen P-M, Watanabe N, HoogC, et al.: PKA and MPF-activated Polo-like kinase regulateanaphase-promoting complex activity and mitotic progression.Mol Cell 1998, 1:371-380.
31. Shirayama M, Zachariae W, Ciosk R, Nasmyth K: The Polo-likekinase Cdc5p and the WD-repeat protein Cdc20/fizzy areregulators and substrates of the anaphase promoting complex inSaccharomyces cerevisiae. EMBO J 1998, 17:1336-1349.
32. Descombes P, Nigg EA: The polo-like kinase Plx1 is required for Mphase exit and destruction of mitotic regulators in Xenopus eggextracts. EMBO J 1998, 17:1328-1335.
33. Visintin R, Prinz S, Amon A: CDC20 and CDH1: a family ofsubstrate-specific activators of APC-dependent proteolysis.Science 1997, 278:460-463.
34. Schwab M, Lutum AS, Seufert W: Yeast Hct1 is a regulator of Clb2cyclin proteolysis. Cell 1997, 90:683-693.
35. Amon A, Tyers M, Futcher B, Nasmyth K: Mechanisms that help theyeast cell cycle clock tick: G2 cyclins transcriptionally activate G2cyclins and repress G1 cyclins. Cell 1993, 74:993-1007.
36. Irniger S, Nasmyth K: The anaphase-promoting complex isrequired in G1-arrested yeast cells to inhibit B-type cyclinaccumulation and to prevent uncontrolled entry into S phase. JCell Sci 1997, 110:1523-1531.
37. Heichman KA, Roberts JM: The yeast CDC16 and CDC27 genesrestrict DNA replication to once per cell cycle. Cell 1996, 85:39-48.
38. Heichman KA, Roberts JM: CDC16 controls initiation atchromosome replication origins. Mol Cell 1998, 1:457-463.
39. Hwang LH, Lau LF, Smith DL, Mistrot CA, Hardwick KG, Hwang ES,et al.: Budding yeast Cdc20: a target of the spindle checkpoint.Science 1998, 279:1041-1044.

40. Yamamoto A, Guacci V, Koshland D: Pds1, an inhibitor of anaphasein budding yeast, plays a critical role in the APC and checkpointpathway(s). J Cell Biol 1996, 133:99-110.
41. Sethi N, Monteagudo MC, Koshland D, Hogan E, Burke DJ: TheCDC20 gene product of Saccharomyces cerevisiae, a betatransducin homologue, is required for a subset of microtubule-dependent cellular processes. Mol Cell Biol 1991, 11:5592-5602.
42. O′Toole ET, Mastronarde DN, Giddins TH, Winey M, Burke DJ,McIntosh JR: Three-dimensional analysis and ultrastructuraldesign of mitotic spindles from the cdc20 mutant ofSaccharomyces cerevisiae. Mol Biol Cell 1997, 8:1-11.
43. Maniatis T, Fritsch EF, Sambrook J: Molecular Cloning: A LaboratoryManual. New York: Cold Spring Harbor Laboratory Press; 1982.
44. Gietz RD, Sugino A: New yeast–E. coli shuttle vectors constructedwith in vitro mutagenised yeast genes lacking six-base pairrestriction sites. Genes 1988, 74:527-535.
45. Schneider BL, Seufert W, Steiner B, Yang QH, Futcher AB: Use ofpolymerase chain reaction epitope tagging for protein tagging inSaccharomyces cerevisiae. Yeast 1995, 11:1265-1274.
46. Surana U, Amon A, Dowzer C, Mcgrew J, Byers B, Nasmyth K:Destruction of the CDC28/CLB kinase is not required formetaphase/anaphase transition in yeast. EMBO J 1993, 12:1969-1978.
47. Schwob E, Böhm T, Mendenhall MD, Nasmyth K: The B-type cyclinkinase inhibitor p40SIC1 controls the G1 to S phase transition inS. cerevisiae. Cell 1994, 79:233-244.
48. Moll T, Tebb G, Surana U, Robitsch H, Nasmyth K: The role ofphosphorylation and the Cdc28 protein kinase in cell cycle-regulated nuclear important of the S. cerevisiae transcriptionfactor SWI5. Cell 1991, 66:743-758.
760 Current Biology, Vol 8 No 13
Because Current Biology operates a ‘Continuous PublicationSystem’ for Research Papers, this paper has been publishedon the internet before being printed. The paper can beaccessed from http://biomednet.com/cbiology/cub — forfurther information, see the explanation on the contents page.





























