Embed Size (px)
Citation preview

NeuroToxicology 33 (2012) 416–423
The similar neurotoxic effects of nanoparticulate and ionic silverin vivo and in vitro
Niels Hadrup a,*, Katrin Loeschner b, Alicja Mortensen a, Anoop K. Sharma a,Klaus Qvortrup c, Erik H. Larsen b, Henrik R. Lam a,d
a Division of Toxicology and Risk Assessment, National Food Institute, Technical University of Denmark, Mørkhøj Bygade 19, DK-2860 Søborg, Denmarkb Division of Food Chemistry, National Food Institute, Technical University of Denmark, Mørkhøj Bygade 19, DK-2860 Søborg, Denmarkc Department of Biomedical Sciences, University of Copenhagen, Blegdamsvej 3, DK-2200 Copenhagen N, Denmarkd Environment and Toxicology, DHI, Agern Alle 5, DK-2970 Hørsholm, Denmark
A R T I C L E I N F O
Article history:
Received 6 December 2011
Accepted 10 April 2012
Available online 15 April 2012
Keywords:
Neurotoxicology
Nanoparticle
Silver
Dopamine
Noradrenaline
5-Hydroxytryptamine
A B S T R A C T
We compared the neurotoxic effects of 14 nm silver nanoparticles (AgNPs) and ionic silver, in the form of
silver acetate (AgAc), in vivo and in vitro. In female rats, we found that AgNPs (4.5 and 9 mg AgNP/kg bw/
day) and ionic silver (9 mg Ag/kg bw/day) increased the dopamine concentration in the brain following
28 days of oral administration. The concentration of 5-hydroxytryptamine (5-HT) in the brain was
increased only by AgNP at a dose of 9 mg Ag/kg bw/day. Only AgAc (9 mg Ag/kg bw/day) was found to
increase noradrenaline concentration in the brain. In contrast to the results obtained from a 28-day
exposure, the dopamine concentration in the brain was decreased by AgNPs (2.25 and 4.5 mg/kg bw/day)
following a 14-day exposure. These data suggest that there are differential effects of silver on dopamine
depending on the length of exposure. In vitro, AgNPs, AgAc and a 12 kDa filtered sub-nano AgNP fraction
were used to investigate cell death mechanisms in neuronal-like PC12 cells. AgNPs and the 12 kDa
filtered fraction decreased cell viability to a similar extent, whereas AgAc was relatively more potent.
AgNPs did not induce necrosis. However, apoptosis was found to be equally increased in cells exposed to
AgNPs and the 12 kDa filtered fraction, with AgAc showing a greater potency. Both the mitochondrial and
the death receptor pathways were found to be involved in AgNP- and AgAc-induced apoptosis. In
conclusion, 14 nm AgNPs and AgAc affected brain neurotransmitter concentrations. AgNP affected 5-HT,
AgAc affected noradrenaline, whereas both silver formulations affected dopamine. Furthermore,
apoptosis was observed in neuronal-like cells exposed to AgNPs, a 12 kDa filtered fraction of AgNP, and
AgAc. These findings suggest that ionic silver and a 14 nm AgNP preparation have similar neurotoxic
effects; a possible explanation for this could be the release and action of ionic silver from the surface of
AgNPs.
� 2012 Elsevier Inc. All rights reserved.
Contents lists available at SciVerse ScienceDirect
NeuroToxicology
1. Introduction
Antibacterial silver nanoparticles (AgNPs) have been developedfor multiple applications, including the coating of refrigerators andbaby bottles (Woodrow Wilson International Center for Scholars,2011). Several findings suggest there are potential neurotoxico-logical effects of AgNPs. First, the central nervous system (CNS) ishighly sensitive to metals such as manganese or mercury(Carpenter, 2001; Clarkson, 1987). Second, silver in its ionic formhas been shown to have effects on the nervous system. Decreasedactivity in mice and concomitant granular silver deposits,especially in brain areas involved in motor control, were foundin mice orally administered AgNO3 for four months (Rungby and
* Corresponding author. Tel.: +45 35887595.
E-mail address: [email protected] (N. Hadrup).
0161-813X/$ – see front matter � 2012 Elsevier Inc. All rights reserved.
http://dx.doi.org/10.1016/j.neuro.2012.04.008
Danscher, 1984). Third, AgNP itself can impact the CNS. AgNPs (50–100 nm) have been reported to disrupt and cross the rat bloodbrain barrier (Tang et al., 2008; Sharma et al., 2010). In vivo effectsof AgNPs include edema (Sharma et al., 2010), and changes in thebrain mRNA levels of genes linked to increased oxidative stress(Rahman et al., 2009). Disruption and inflammation also have beendemonstrated in in vitro blood–brain barrier models followingincubation with AgNP (Tang et al., 2010; Trickler et al., 2010).
The toxic effects of nanoparticles have been attributed to ahighly reactive environment on the particle surface and aconcomitant high surface to mass ratio (Oberdorster et al.,2005). In support of this, differences in toxicity have beendemonstrated for AgNPs and ionic silver in some settings. Inmouse spermatogonia germline stem cells, AgNPs but not Ag2CO3
inhibited mitochondrial metabolic activity (Braydich-Stolle et al.,2005). In mouse fibroblast cells, AgNP, but not silver ions, inducedapoptosis (Hsin et al., 2008). In addition, AgNPs were more potent

Fig. 1. A transmission electron micrograph (TEM) of the stock suspension of AgNPs.
The AgNPs used in the experiments were not aggregated.
N. Hadrup et al. / NeuroToxicology 33 (2012) 416–423 417
than silver ions in inducing the production of reactive oxygenspecies in a human T-cell line (Eom and Choi, 2010). However, incontrast to specific particle surface effects, an alternative/supplementary mechanism of AgNP toxicity could be the releaseof reactive free ions from the surface. In support of this, datasuggest that AgNP preparations exert effects similar to those ofionic silver. The reduction of mitochondrial reductase activity wasfound to be similar in a rat macrophage cell line exposed to AgNO3
and AgNPs (Carlson et al., 2008). Additionally, a similar genome-wide gene expression profile was found in an in vitro humanintestinal epithelium Caco-2 cell line following incubation withAgNO3 or AgNPs (Bouwmeester et al., 2011).
To expand our knowledge of the neurotoxic mechanisms ofsilver and to investigate differences between nanoparticulate andionic silver formulations, we employed in vivo and in vitro
techniques. Neurotoxicity can be manifestations of changedneurotransmitter metabolism, specifically, manganese, mercuryand lead have been demonstrated to influence neurotransmitterdynamics in the brain (Barbeau, 1984; Bondy et al., 1979;Donaldson, 1984). In the present investigation we tested whetherthe concentrations of multiple brain neurotransmitters werealtered following the oral administration of 14 nm AgNPs or ionicsilver to rats. Cell death is linked to disturbances in neurotrans-mitter brain concentrations in such conditions as Parkinson’sdisease and Alzheimer’s disease (Mattson et al., 1999; Ehringer andHornykiewicz, 2012). We therefore utilized neuronal-like PC12cells to test whether there was a difference in cell deathmechanisms following exposure to 14 nm AgNPs, ionic silver ora 12 kDa filtered sub-nano fraction of the AgNP preparation.
2. Materials and methods
2.1. Silver suspensions
The AgNPs were kindly produced by Dr. Xueyun Gao from theInstitute of High Energy Physics, at the Chinese Academy ofScience in Beijing. The AgNPs were produced and characterizedas previously described (Zhang et al., 1996; Loeschner et al.,2011). Briefly, 14 nm AgNPs were stabilized with 11.5 mg/mLpolyvinylpyrrolidone (PVP). The size distribution of the AgNPswas determined by dynamic light scattering. The volumedistribution of the size had two peaks. The first peak(representing approximately 90% of the particle volume) hadits maximum at 14 � 2 nm with a peak width of 4 � 1 nm. Thesecond peak had its maximum at 50 � 9 nm with a peak width of42 � 3 nm. The second peak represented 11% of the total volume ofnanoparticles, but less than 0.1% of the total particle number.Through inductively coupled plasma mass spectrometry (ICP-MS)analysis, the AgNP concentration was determined to be 450 mg Ag/mL. Transmission electron microscopy (TEM) was employed toensure that no aggregation or agglomeration had occurred (Fig. 1).During the in vivo and in vitro experimental periods, dynamic lightscattering was utilized to further ensure the size distribution of theAgNPs in suspension (specifically this was done seven times duringa 150 day period. Data not shown). Specifically for the in vitro
experiments, a 12 kDa filtered fraction was prepared by centrifu-gation of the AgNP suspension through a cellulose acetate filter(VectaSpin Micro Centrifuge Filter, Whatman, England) at 8117 � g
for 30 min in a micro-centrifuge (Eppendorf MiniSpin, EppendorfAG, Hamburg, Germany). The free Ag ion concentration in thefiltrate constituted 11% of the total silver in the AgNP preparation.Ionic silver was used in the form of silver acetate (AgAc; Sigma, St.Louis, Prod. no. 204374). To obtain a vehicle similar to the AgNPsuspension, AgAc was dissolved in 11.5 mg/mL PVP. The AgAcconcentration in the solution was 708 mg/mL.
2.2. Neurotransmitter analysis
The total brain concentrations of dopamine, noradrenaline and5-hydroxytryptamine (5-HT) were measured in homogenatesobtained from a previous animal investigation (Hadrup et al.,2011). Administered orally by gavage (gavage volume: 10 mL/kgbw), female Wistar rats (n = 6/group) were given a vehicle control(PVP, 11.5 mg/mL), 2.25 mg Ag/kg bw/day, 4.5 mg Ag/kg bw/day,9 mg Ag/kg bw/day or 14 mg AgAc/kg bw/day (equal to 9 mg Ag/kgbw/day) for 28 days. Additional groups, used to determine anoptimal dose, included administration of the vehicle control, 2.25or 4.5 mg AgNP/kg bw/day for only 14 days. Animals were notaffected in general appearance or behavior. Terminal body weightwas not significantly different although body weight gain wasslightly decreased in AgAc administered rats (Hadrup et al., 2011).The rat studies were conducted under conditions approved by theDanish Animal Experiments Inspectorate and the in-house AnimalWelfare Committee. After anaesthethesia in CO2/O2 and euthana-sia by decapitation, brains were quickly transferred to ice-cold0.32 M sucrose (9 mL/whole brain) and homogenized with anUltraTurrax T25 homogenizer (IKA, Staufen, Germany) at24,000 rpm in an ice bath. Next, 1000 mL of homogenate wasadded to 1000 mL of 0.2 M perchloric acid. The perchloric acid-deproteinized samples were then centrifuged (20 min, 0–4 8C,10,000 � g). For the noradrenaline and dopamine measurements,3,4-dihydroxybenzylamine (Sigma, D 7012) was added to thesamples as an internal standard, then the samples were purified onaluminum oxide (Anton and Sayre 1962) and eluted by addition of0.2 M HClO4. For the analysis of 5-HT, N-v-methyl-5-HT (Sigma, M1514) was added to the samples as an internal standard.Noradrenaline, dopamine and 5-HT concentrations were analyzedusing high performance liquid chromatography (HPLC) withelectrochemical detection (Lam et al., 1992). The instrumentationconsisted of a Hewlett-Packard Ti-SERIES 1100 liquid chromato-graph equipped with an ESA MD-150 analytical column (3 mmID � 15 cm, C-18, 5 mm). Separation was achieved at ambienttemperature with a 5% (v/v) acetonitrile modified 75 mM sodiumphosphate buffer (1.7 octenylsulfonic acid, 100 mL triethylamine/Land 25 mM EDTA; pH 3) and a flow rate of 0.8 ml/min. Signals weredetected using a dual potentiostat electrochemical detector (ESAModel 5200A Coulochem II and ESA Model 5011 high sensitivity

N. Hadrup et al. / NeuroToxicology 33 (2012) 416–423418
analytical cell with an E1 of �150 mV and an E2 of +220 mV; guardcell potential: +350 mV).
2.3. Cell culture
PC12 cells were obtained from American Type Culture Center(Prod no. CRL 1721, lot no. 58078699, ATCC, Manassas, VA). Thecells were grown in Dulbecco’s modified Eagle’s medium/Ham’s F-12 (1:1) solution, containing 15 mM 4-(2-hydroxyethyl)-1-piper-azineethanesulfonic acid and L-glutamine (Lonza, Walkersville,MD), 15% horse serum (Lonza, Walkersville, MD), 2.5% fetal bovineserum (Sigma, St. Louis, MO), 1% (25 U/mL) penicillin/streptomy-cin, an additional 1% (2 mM) glutamine (Gibco, Carlsbad, CA) and0.075% (w/v) sodium-bicarbonate (Sigma, St. Louis, MO), at 37 8C ina humidified atmosphere with 5% CO2. Polystyrene culturesurfaces (Sarstedt, Newton, NC, USA) were coated with 1% collagen(Prod. no. 5409, Inamed Biomaterials, Freemont, CA, USA) tofacilitate cell adhesion. Cells were subcultured by scraping and twopassages through a 27 G 3/4 syringe before re-seeding at3 � 104 cells/cm2. Experiments were done on cells that hadundergone fewer than six passages.
2.4. In vitro evaluation of cell viability and death mechanisms
For viability measurements, PC12 cells were seeded at3 � 104 cells/cm2 in 12-well plates (cell medium volume: 1 mL/well) and cultured for 24 h. Cells were then incubated for 4–48 hwith 0.5, 5 and 10 mg/mL unfiltered AgNP (NP in figures), 0.06, 0.6and 1.2 mg Ag/mL (concentration levels are after filtration andcorresponds to unfiltered AgNP groups described above) of the12 kDa filtered fraction (F (NP 0.5), F (NP 5) and F (NP 10) in figures)or 0.05, 0.5, 5 or 10 mg Ag/mL of AgAc. Viability was evaluated bycounting adhered and floating cells in the presence of Trypan Blue(Sigma, St. Louis) at a 1:1 concentration by counting with a Burker-Turk counting chamber using an Olympus CK2 inverted micro-scope (Olympus, Essex, UK). Blue cells appeared dead andtransparent cells were considered to be alive. The samples werenot blinded during this procedure. The experiment was conductedindependently twice with an n = 3 wells for each condition.
For the necrosis detection, PC12 cells were cultured at3 � 104 cells/cm2 in 12-well polystyrene plates (cell mediumvolume: 1 mL/well) for 24 h and then incubated with nanoparticlesat 5 or 10 mg/mL for 24 h (2 repetitions, n = 1 in each). The positivecontrol for necrosis detection was induced by three cycles offreezing and thawing. The culture medium was removed andcentrifuged 5 min at 1300 � g to remove floating cells. An equalvolume of 2� Laemmli buffer was added to 500 mL supernatantand samples were incubated for 30 min at 37 8C. Western blottingwas completed as previously described (Hadrup et al., 2007). Ananti-High-mobility group protein B1 (HMGB1) antibody was usedat a 3000-fold dilution (Sigma, St. Louis).
Apoptosis was determined using the terminal deoxynucleotidyltransferase dUTP nick end labeling (TUNEL) assay. For this assay,cells were seeded at 6 � 104 cells/cm2 on collagen coated Lab-TekII-CC2 eight-well glass slides (Prod. no. 154941, Nunc, Roskilde,Denmark) (cell medium volume: 0.5 mL/well). Cells were allowedto attach and grow for 24 h, then treated with Ag with or without20 mM of a caspase 8 inhibitor (cat. no. 218840, Merck, Darmstadt,Germany), or 20 mM of a caspase 9 inhibitor (cat. no. 218841,Merck, Darmstadt, Germany). Next, cells were cultured for 24 h,washed with PBS (pH 7.4), and then fixed in 4% formaldehyde inPBS (pH 7.4) for 15 min at room temperature. Following twowashes in PBS, the cells were post-fixed in a pre-cooledethanol:acetic acid (2:1) solution for 5 min at �20 8C, washedanother two times in PBS, and then subjected to TUNEL with anApoptag peroxidase in situ apoptosis detection kit (Prod. no. S7100,
Millipore, Billerica, MA, USA), according to the manufacturer’sinstructions. The results of the assay were evaluated using lightmicroscopy (Leica DMR Microscope, Leica Microsystems, Wetzlar,Germany). Twelve areas per culture well were selected in astandardized fashion by moving the specimen in fixed steps (1 mmleft and 1 mm toward the front using the scale controls of themicroscope). The total number of healthy and apoptotic cells ofeach area were counted manually in a 10 cm � 10 cm frame on thecomputer screen in images recorded with a Leica DFC 290 digitalcamera (Leica Microsystems, Wetzlar, Germany). The data for thetwelve areas were summed, and the percentage of apoptotic cellswas calculated. The data for individual slides were obtained onindividual days (n = 3–12, as noted in the figure legends) and thenplotted using GraphPad Prism (GraphPad Software, La Jolla, CA).
2.5. Determination of intracellular silver
For the determination of silver uptake, the PC12 cells wereseeded at 3 � 104 cells/cm2, grown for 24 h and then incubatedwith 5 mg/mL AgNP for 24 h (n = 3 wells) or for 30 min on ice to testfor unspecific binding (n = 2). The PC12 cells incubated with thevehicle for 24 h at 37 8C served as a control (n = 1). Cells were thenwashed twice in PBS, released by scraping, and collected bycentrifugation for 5 min at 1300 � g. Cell pellets were digested in1 mL of concentrated nitric acid (PlasmaPure, SCP Science, Quebec,Canada) in a microwave oven (Multiwave, Anton Paar, Graz,Austria) at elevated temperature and pressure (�250 8C, �70 bar).The silver concentration in samples diluted with deionized waterwas determined using a quadrupole-based ICP-MS (7500ce,Agilent Technologies, Japan). The concentration was quantifiedagainst an external calibration curve (1000 mg/mL Ag stocksolution; SCP Science, Quebec, Canada) with internal standardiza-tion (1000 mg/mL rhodium (Rh) stock solution; SCP Science,Quebec, Canada). The silver isotope 107Ag was chosen for the ICP-MS measurements because of its low detection limit.
2.6. The detection of intracellular AgNP using transmission electron
microscopy (TEM)
PC12 cells were exposed to 10 mg/mL AgNP for 24 h (n = 1),washed twice with PBS, fixed in a 2% (v/v) glutaraldehyde in 0.05 Msodium phosphate buffer (pH 7.2) solution for 1.5 h, scraped, andcentrifuged. The sample was then rinsed in 0.15 M sodiumcacodylate buffer (pH 7.2) and subsequently postfixed in a 1%(w/v) OsO4 in 0.12 M sodium cacodylate buffer (pH 7.2) solutionfor 2 h. Next, the specimen was dehydrated in a gradated series ofethanol solutions, transferred to propylene oxide and embedded inEpon according to standard procedures. Two consecutive sections,approximately 80 nm thick, were cut with a Reichert-Jung UltracutE microtome and collected on copper grids with Formvarsupporting membranes. One section was stained with uranylacetate and lead citrate and one section was left unstained. Bothsections contained cross sections of approximately twenty cellsand were randomly scanned at the appropriate magnification forthe presence of AgNP. This was completed with a Philips CM 100TEM, operated at an accelerating voltage of 80 kV, and an OSISVeleta side-mounted digital slow scan 2k � 2k CCD camera system.Digital images were recorded with the analySIS software package(SoftImaging Systems, Germany), and one hundred positions wereanalyzed for the presence of AgNP.
2.7. Statistical analyses
The data are presented as the mean � standard error of the mean(SEM). For the neurotransmitter measurements significant differ-ences were determined using an ANOVA with a Bonferroni’s multiple

Ta
ble
1B
rain
ne
uro
tra
nsm
itte
rco
nce
ntr
ati
on
s.
14
da
yin
ve
stig
ati
on
28
da
yin
ve
stig
ati
on
Tre
atm
en
tV
eh
icle
Ag
NP
Ag
NP
Ve
hic
leA
gN
PA
gN
PA
gN
PA
gA
c
Do
se(m
gA
g/k
gb
w/d
ay
)0
2.2
54
.50
2.2
54
.59
.09
.0
Bra
ind
op
am
ine
2.6
8�
0.0
72
.20�
0.0
9**
*2
.32�
0.0
2**
3.9
6�
0.2
04
.24�
0.2
85
.32�
0.1
0**
*4
.74�
0.1
9*
5.2
6�
0.1
9**
(nm
ol/
gb
rain
tiss
ue)
Bra
inn
ora
dre
na
lin
e1
.78�
0.1
12
.17�
0.1
62
.18�
0.0
81
.70�
0.0
81
.73�
0.0
61
.98�
0.1
11
.69�
0.0
42
.02�
0.0
5*,§
(nm
ol/
gb
rain
tiss
ue)
Bra
in5
-HT
1.5
3�
0.2
51
.62�
0.1
11
.68�
0.0
90
.18�
0.0
20
.19�
0.0
20
.22�
0.0
40
.26�
0.0
2**
0.2
3�
0.0
2(n
mo
l/g
bra
inti
ssu
e)
N. Hadrup et al. / NeuroToxicology 33 (2012) 416–423 419
comparisons test (control vs. AgNP groups, control vs. AgAc, and AgNP9 mg/kg bw/day vs. AgAc). For the viability data (control vs. AgNPgroups, control vs. the 12 kDa filtered fraction and control vs. AgAc),significant differences were determined using an ANOVA with aDunnet’s post-test. For measurements of apoptosis (control vs. AgNP,the 12 kDa filtered fraction or AgAc groups, and AgNP or AgAc vs.
caspase inhibitor 8 and 9 groups), a t-test without Bonferronicorrection was utilized. Statistics were completed using GraphPadPrism 5.0 (GraphPad Software, La Jolla, CA).
3. Results
3.1. The effect of AgNPs and AgAc on brain neurotransmitter
concentrations
Rats treated with AgNPs or AgAc for 28 days show an increaseddopamine concentration in the brain when compared with thecontrols (mean � SEM; Control: 4.0 � 0.2 nmol/g brain tissue vs.
AgNP 4.5 mg/kg bw/day: 5.3 � 0.1 nmol/g brain tissue, p < 0.001;Control vs. AgNP 9.0 mg/kg bw/day: 4.8 � 0.2 nmol/g brain tissue,p < 0.05; Control vs. AgAc 9 mg Ag/kg bw/day: 5.3 � 0.2 nmol/g braintissue, p < 0.01; Table 1). In a 14-day treatment for dose determina-tion, results show that AgNPs decreased the dopamine concentrationin the brain (mean � SEM; Control: 2.7 � 0.1 nmol/g brain tissue vs.
AgNP 2.25 mg/kg bw/day: 2.2 � 0.1 nmol/g brain tissue, p < 0.001;Control vs. AgNP 4.5 mg/kg bw/day: 2.3 � 0.2 nmol/g brain tissue,p < 0.01; Table 1). We note that the 14 and 28 day investigations wereconducted as two independent experiments. Thus, taking differentanimal batches into account, the difference in dopamine levels incontrol animals between the two experiments cannot be directlyattributed to differences in treatment period length. Following 28days of silver administration, concentration of 5-HT and noradrena-line in the brain were both altered. The noradrenaline brainconcentration was increased following AgAc administration, butnot following AgNP administration (Control: 1.7 � 0.08 nmol/g braintissue vs. AgAc 9 mg Ag/kg bw/day: 2.0 � 0.05 nmol/g brain tissue,p < 0.05, AgAc 9 mg Ag/kg bw/day: 2.0 � 0.05 nmol/g brain tissue vs.
AgNP 9.0 mg/kg bw/day: 1.7 � 0.05 nmol/g brain tissue, p < 0.05;Table 1). The 5-HT concentration was increased following AgNP butnot AgAc administration (Control: 0.18 � 0.02 nmol/g brain tissue vs.
AgNP 9.0 mg/kg bw/day: 0.26 � 0.01 nmol/g brain tissue, p < 0.01;Table 1). The brain weight was not affected by AgNP or AgActreatment (data not shown).
3.2. The levels of apoptosis and necrosis in PC12 cells following
exposure to AgNPs, the 12 kDa filtered fraction or AgAc
Cell death was evaluated by Trypan Blue staining of cellsexposed to AgNP, the 12 kDa filtered fraction and AgAc (Fig. 2). Theresults are similar between cells exposed to AgNP and the 12 kDafiltered fraction. After 24 h of exposure to 10 mg/mL AgNPs or acorresponding concentration of the 12 kDa filtered fraction, thenumber of dead cells increased (10 and 7 fold, respectively). After48 h of exposure to 5 and 10 mg/mL AgNPs or a correspondingconcentration of the 12 kDa filtered fraction, the number of deadcells was found to be increased (5 mg/mL: 8 and 3.5 fold,respectively; 10 mg/mL: 16 and 8 fold, respectively). In addition,the number of living cells decreased after exposure to 10 mg/mLAgNP (10 fold) or a corresponding concentration of the 12 kDafiltered fraction (5 fold). With regard to AgAc exposure, after 4 h ofincubation the number of dead cells was increased (5 fold) at aconcentration of 5 mg Ag/mL. After 24 h of incubation with 5 mgAg/mL, the number of dead cells was increased (12 fold) and thenumber of living cells was decreased (3.5 fold). After 48 h ofincubation, the number of dead cells exposed to 5 mg Ag/mL wasincreased (23 fold) and the number of living cells was decreased
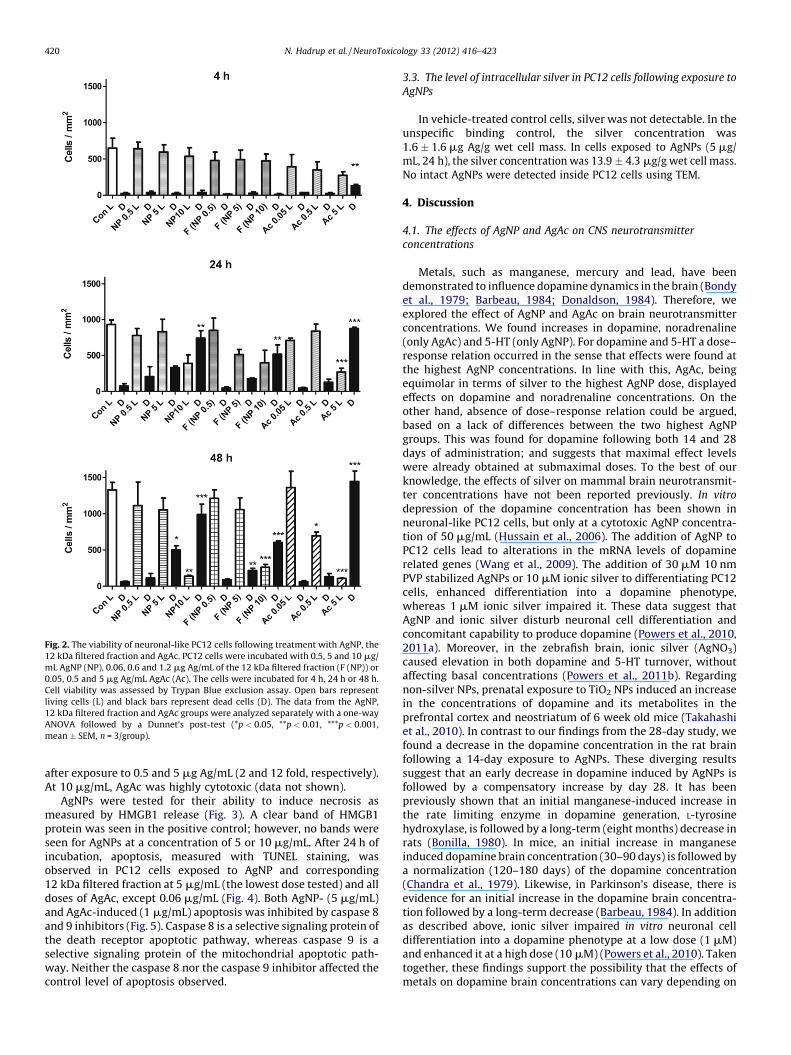
Fig. 2. The viability of neuronal-like PC12 cells following treatment with AgNP, the
12 kDa filtered fraction and AgAc. PC12 cells were incubated with 0.5, 5 and 10 mg/
mL AgNP (NP), 0.06, 0.6 and 1.2 mg Ag/mL of the 12 kDa filtered fraction (F (NP)) or
0.05, 0.5 and 5 mg Ag/mL AgAc (Ac). The cells were incubated for 4 h, 24 h or 48 h.
Cell viability was assessed by Trypan Blue exclusion assay. Open bars represent
living cells (L) and black bars represent dead cells (D). The data from the AgNP,
12 kDa filtered fraction and AgAc groups were analyzed separately with a one-way
ANOVA followed by a Dunnet’s post-test (*p < 0.05, **p < 0.01, ***p < 0.001,
mean � SEM, n = 3/group).
N. Hadrup et al. / NeuroToxicology 33 (2012) 416–423420
after exposure to 0.5 and 5 mg Ag/mL (2 and 12 fold, respectively).At 10 mg/mL, AgAc was highly cytotoxic (data not shown).
AgNPs were tested for their ability to induce necrosis asmeasured by HMGB1 release (Fig. 3). A clear band of HMGB1protein was seen in the positive control; however, no bands wereseen for AgNPs at a concentration of 5 or 10 mg/mL. After 24 h ofincubation, apoptosis, measured with TUNEL staining, wasobserved in PC12 cells exposed to AgNP and corresponding12 kDa filtered fraction at 5 mg/mL (the lowest dose tested) and alldoses of AgAc, except 0.06 mg/mL (Fig. 4). Both AgNP- (5 mg/mL)and AgAc-induced (1 mg/mL) apoptosis was inhibited by caspase 8and 9 inhibitors (Fig. 5). Caspase 8 is a selective signaling protein ofthe death receptor apoptotic pathway, whereas caspase 9 is aselective signaling protein of the mitochondrial apoptotic path-way. Neither the caspase 8 nor the caspase 9 inhibitor affected thecontrol level of apoptosis observed.
3.3. The level of intracellular silver in PC12 cells following exposure to
AgNPs
In vehicle-treated control cells, silver was not detectable. In theunspecific binding control, the silver concentration was1.6 � 1.6 mg Ag/g wet cell mass. In cells exposed to AgNPs (5 mg/mL, 24 h), the silver concentration was 13.9 � 4.3 mg/g wet cell mass.No intact AgNPs were detected inside PC12 cells using TEM.
4. Discussion
4.1. The effects of AgNP and AgAc on CNS neurotransmitter
concentrations
Metals, such as manganese, mercury and lead, have beendemonstrated to influence dopamine dynamics in the brain (Bondyet al., 1979; Barbeau, 1984; Donaldson, 1984). Therefore, weexplored the effect of AgNP and AgAc on brain neurotransmitterconcentrations. We found increases in dopamine, noradrenaline(only AgAc) and 5-HT (only AgNP). For dopamine and 5-HT a dose–response relation occurred in the sense that effects were found atthe highest AgNP concentrations. In line with this, AgAc, beingequimolar in terms of silver to the highest AgNP dose, displayedeffects on dopamine and noradrenaline concentrations. On theother hand, absence of dose–response relation could be argued,based on a lack of differences between the two highest AgNPgroups. This was found for dopamine following both 14 and 28days of administration; and suggests that maximal effect levelswere already obtained at submaximal doses. To the best of ourknowledge, the effects of silver on mammal brain neurotransmit-ter concentrations have not been reported previously. In vitro
depression of the dopamine concentration has been shown inneuronal-like PC12 cells, but only at a cytotoxic AgNP concentra-tion of 50 mg/mL (Hussain et al., 2006). The addition of AgNP toPC12 cells lead to alterations in the mRNA levels of dopaminerelated genes (Wang et al., 2009). The addition of 30 mM 10 nmPVP stabilized AgNPs or 10 mM ionic silver to differentiating PC12cells, enhanced differentiation into a dopamine phenotype,whereas 1 mM ionic silver impaired it. These data suggest thatAgNP and ionic silver disturb neuronal cell differentiation andconcomitant capability to produce dopamine (Powers et al., 2010,2011a). Moreover, in the zebrafish brain, ionic silver (AgNO3)caused elevation in both dopamine and 5-HT turnover, withoutaffecting basal concentrations (Powers et al., 2011b). Regardingnon-silver NPs, prenatal exposure to TiO2 NPs induced an increasein the concentrations of dopamine and its metabolites in theprefrontal cortex and neostriatum of 6 week old mice (Takahashiet al., 2010). In contrast to our findings from the 28-day study, wefound a decrease in the dopamine concentration in the rat brainfollowing a 14-day exposure to AgNPs. These diverging resultssuggest that an early decrease in dopamine induced by AgNPs isfollowed by a compensatory increase by day 28. It has beenpreviously shown that an initial manganese-induced increase inthe rate limiting enzyme in dopamine generation, L-tyrosinehydroxylase, is followed by a long-term (eight months) decrease inrats (Bonilla, 1980). In mice, an initial increase in manganeseinduced dopamine brain concentration (30–90 days) is followed bya normalization (120–180 days) of the dopamine concentration(Chandra et al., 1979). Likewise, in Parkinson’s disease, there isevidence for an initial increase in the dopamine brain concentra-tion followed by a long-term decrease (Barbeau, 1984). In additionas described above, ionic silver impaired in vitro neuronal celldifferentiation into a dopamine phenotype at a low dose (1 mM)and enhanced it at a high dose (10 mM) (Powers et al., 2010). Takentogether, these findings support the possibility that the effects ofmetals on dopamine brain concentrations can vary depending on
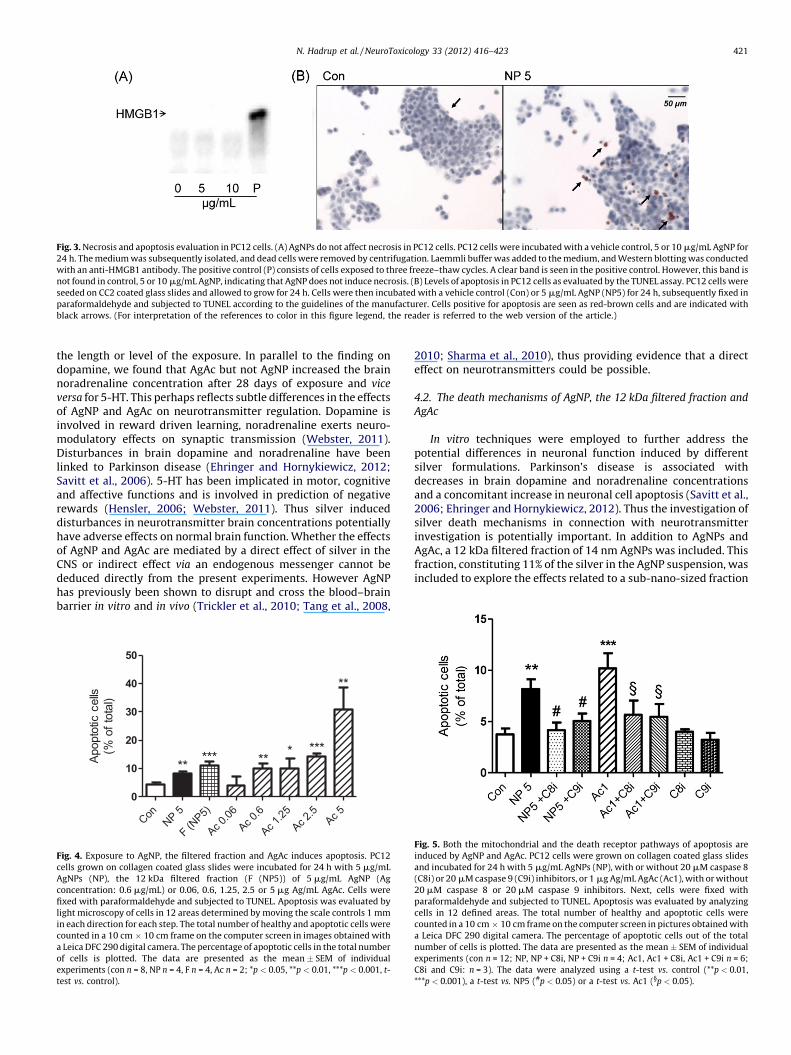
Fig. 3. Necrosis and apoptosis evaluation in PC12 cells. (A) AgNPs do not affect necrosis in PC12 cells. PC12 cells were incubated with a vehicle control, 5 or 10 mg/mL AgNP for
24 h. The medium was subsequently isolated, and dead cells were removed by centrifugation. Laemmli buffer was added to the medium, and Western blotting was conducted
with an anti-HMGB1 antibody. The positive control (P) consists of cells exposed to three freeze–thaw cycles. A clear band is seen in the positive control. However, this band is
not found in control, 5 or 10 mg/mL AgNP, indicating that AgNP does not induce necrosis. (B) Levels of apoptosis in PC12 cells as evaluated by the TUNEL assay. PC12 cells were
seeded on CC2 coated glass slides and allowed to grow for 24 h. Cells were then incubated with a vehicle control (Con) or 5 mg/mL AgNP (NP5) for 24 h, subsequently fixed in
paraformaldehyde and subjected to TUNEL according to the guidelines of the manufacturer. Cells positive for apoptosis are seen as red-brown cells and are indicated with
black arrows. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
N. Hadrup et al. / NeuroToxicology 33 (2012) 416–423 421
the length or level of the exposure. In parallel to the finding ondopamine, we found that AgAc but not AgNP increased the brainnoradrenaline concentration after 28 days of exposure and vice
versa for 5-HT. This perhaps reflects subtle differences in the effectsof AgNP and AgAc on neurotransmitter regulation. Dopamine isinvolved in reward driven learning, noradrenaline exerts neuro-modulatory effects on synaptic transmission (Webster, 2011).Disturbances in brain dopamine and noradrenaline have beenlinked to Parkinson disease (Ehringer and Hornykiewicz, 2012;Savitt et al., 2006). 5-HT has been implicated in motor, cognitiveand affective functions and is involved in prediction of negativerewards (Hensler, 2006; Webster, 2011). Thus silver induceddisturbances in neurotransmitter brain concentrations potentiallyhave adverse effects on normal brain function. Whether the effectsof AgNP and AgAc are mediated by a direct effect of silver in theCNS or indirect effect via an endogenous messenger cannot bededuced directly from the present experiments. However AgNPhas previously been shown to disrupt and cross the blood–brainbarrier in vitro and in vivo (Trickler et al., 2010; Tang et al., 2008,
Con
NP 5
F (NP5)
Ac 0.
06
Ac 0.
6
Ac 1.
25
Ac 2.
5Ac 5
0
10
20
30
40
50
**
******
*****A
popto
tic c
ells
(% o
f to
tal)
Fig. 4. Exposure to AgNP, the filtered fraction and AgAc induces apoptosis. PC12
cells grown on collagen coated glass slides were incubated for 24 h with 5 mg/mL
AgNPs (NP), the 12 kDa filtered fraction (F (NP5)) of 5 mg/mL AgNP (Ag
concentration: 0.6 mg/mL) or 0.06, 0.6, 1.25, 2.5 or 5 mg Ag/mL AgAc. Cells were
fixed with paraformaldehyde and subjected to TUNEL. Apoptosis was evaluated by
light microscopy of cells in 12 areas determined by moving the scale controls 1 mm
in each direction for each step. The total number of healthy and apoptotic cells were
counted in a 10 cm � 10 cm frame on the computer screen in images obtained with
a Leica DFC 290 digital camera. The percentage of apoptotic cells in the total number
of cells is plotted. The data are presented as the mean � SEM of individual
experiments (con n = 8, NP n = 4, F n = 4, Ac n = 2; *p < 0.05, **p < 0.01, ***p < 0.001, t-
test vs. control).
2010; Sharma et al., 2010), thus providing evidence that a directeffect on neurotransmitters could be possible.
4.2. The death mechanisms of AgNP, the 12 kDa filtered fraction and
AgAc
In vitro techniques were employed to further address thepotential differences in neuronal function induced by differentsilver formulations. Parkinson’s disease is associated withdecreases in brain dopamine and noradrenaline concentrationsand a concomitant increase in neuronal cell apoptosis (Savitt et al.,2006; Ehringer and Hornykiewicz, 2012). Thus the investigation ofsilver death mechanisms in connection with neurotransmitterinvestigation is potentially important. In addition to AgNPs andAgAc, a 12 kDa filtered fraction of 14 nm AgNPs was included. Thisfraction, constituting 11% of the silver in the AgNP suspension, wasincluded to explore the effects related to a sub-nano-sized fraction
Fig. 5. Both the mitochondrial and the death receptor pathways of apoptosis are
induced by AgNP and AgAc. PC12 cells were grown on collagen coated glass slides
and incubated for 24 h with 5 mg/mL AgNPs (NP), with or without 20 mM caspase 8
(C8i) or 20 mM caspase 9 (C9i) inhibitors, or 1 mg Ag/mL AgAc (Ac1), with or without
20 mM caspase 8 or 20 mM caspase 9 inhibitors. Next, cells were fixed with
paraformaldehyde and subjected to TUNEL. Apoptosis was evaluated by analyzing
cells in 12 defined areas. The total number of healthy and apoptotic cells were
counted in a 10 cm � 10 cm frame on the computer screen in pictures obtained with
a Leica DFC 290 digital camera. The percentage of apoptotic cells out of the total
number of cells is plotted. The data are presented as the mean � SEM of individual
experiments (con n = 12; NP, NP + C8i, NP + C9i n = 4; Ac1, Ac1 + C8i, Ac1 + C9i n = 6;
C8i and C9i: n = 3). The data were analyzed using a t-test vs. control (**p < 0.01,
***p < 0.001), a t-test vs. NP5 (#p < 0.05) or a t-test vs. Ac1 (§p < 0.05).

N. Hadrup et al. / NeuroToxicology 33 (2012) 416–423422
of our AgNP preparation. The overall viability and apoptosis levelsof PC12 cells showed that AgNP and the 12 kDa filtered fraction hadequal toxicity. This suggests that the sub-nano fraction of the AgNPpreparation is responsible for the decreased viability and increasedapoptosis in the neuronal-like cells. In addition, the overallviability data and the specific apoptosis data on cells treated withAgAc suggest that ionic silver is toxic at doses two- to tenfold lowerthan AgNP. This provides further evidence that the toxic effect ofAgNP is mediated by the sub-nano/ionic fraction. The effect ofAgNP on necrosis markers, to our knowledge, has not beenpreviously investigated in neuronal-like cells. However, in mono-cytes, necrosis was induced by 60 nm AgNPs (Foldbjerg et al.,2009). Thus, the lack of necrosis induction by 14 nm AgNPs in thepresent investigation, suggests a cell type-specific or AgNP size-specific effect. The involvement of the death receptor ormitochondrial pathways was investigated by co-incubation ofAgNPs or AgAc and inhibitors of caspase 8 and 9. Both inhibitorsabrogated AgNP- or AgAc-induced apoptosis, suggesting that boththe death receptor and mitochondrial pathways play a role in thismechanism of cell death. Apoptosis induced by AgNP and ionicsilver via the mitochondrial pathway has been demonstrated inseveral non-neural cell types (Wei et al., 2010; Lee et al., 2011; Piaoet al., 2011; Foldbjerg et al., 2009; Miura and Shinohara, 2009) andin human U251 glioblastoma cells (AshaRani et al., 2009).However, in contrast to our findings, none of these reports showthe additional involvement of the death receptor pathway.According to findings in Parkinson’s disease (Savitt et al., 2006;Ehringer and Hornykiewicz, 2012), the decrease in brain dopamineconcentrations observed following 14 days of AgNP administrationcould be explained by an increase in apoptosis of dopaminergicneurons. In contrast, it is less obvious how increased apopotosis isconnected with increases in rat brain neurotransmitter concen-trations, as observed following 28 days of AgNP or AgAcadministration. In vitro data have indicated that ionic silverimpaired neuronal cell differentiation into a dopamine phenotypeat a low dose (1 mM) and enhanced it at a high dose (10 mM)(Powers et al., 2010). These data, together with our data showingdose–response related increases in apoptosis, suggest that certainconcentrations of silver cause concomitant increases in dopamineand apoptosis. An alternative explanation is that apoptosis in sometypes of neurons cause other types of neurons to increase theproduction of neurotransmitters in the mammalian brain.
4.3. The detection of silver inside PC12 cells
Following a 24 h incubation of PC12 cells with 5 mg/mL of AgNP,we found silver in the PC12 cells. Data from ice-incubated cellsshow that cell surface binding could be excluded, suggesting thatsilver is internalized following exposure to AgNPs. TEM was thenapplied to determine whether intact AgNPs were internalized.Internalization has been demonstrated previously in non-neuronalcells exposed to 30–50 nm AgNPs (Carlson et al., 2008; Wei et al.,2010; Hackenberg et al., 2011). In our study, no AgNPs could bedetected within the PC12 cells, indicating that the employed AgNPsare not internalized into neuronal-like cells. Instead, the AgNPsmay dissolve rapidly upon internalization, or it may be that freesilver ions act on the cells.
4.4. The evidence for and against a specific nano-formulation effect of
silver
Several lines of data suggest dissimilar effects of AgNPs andionic silver. AgNPs but not Ag ions affected mitochondrialmetabolism in mammalian germline stem cells (Braydich-Stolleet al., 2005). With regard to apoptosis, AgNP had an opposite effectfrom that of non-nanosilver in a mouse fibroblast cell line (Hsin
et al., 2008). AgNPs induced more reactive oxygen species whencompared with Ag ions in a human T-cell line (Eom and Choi,2010). Together, these findings suggest that a specific nano-effectcan occur in some instances, but such conclusions cannot be drawnfrom our data. Our findings point to similar neurotoxic effects ofboth nano-formulated and ionic silver (and the filtered subnanofraction) and that these effects are mediated by the free silver ionsin the suspension. In line with these results, data on otherbiological aspects support similar mechanisms for AgNPs and ionicsilver. Gene expression was similar in an in vitro model for humanintestinal epithelium following incubation with AgNO3 and AgNPsthat consisted of 6–17% free ions (Bouwmeester et al., 2011).Several apoptotic induction studies point to similar effects ofAgNPs and Ag ions but with ions being more potent. This suggeststhat it is the free ions in the AgNP preparations that exert theeffects seen in AgNP-treated cells (Miura and Shinohara, 2009;Foldbjerg et al., 2009; Li et al., 2010). Thus at present there are datato support specific effects of nano-formulation, and other datapointing toward effects mediated by released ions, suggesting thatboth mechanisms can occur together.
5. Conclusion
In conclusion, 14 nm AgNPs and AgAc affected brain neuro-transmitter concentrations. AgNP affected 5-HT, AgAc affectednoradrenaline, whereas both silver formulations affected dopa-mine. In vitro apoptotic death mechanisms were similar for AgNPs,the 12 kDa filtered fraction, and AgAc. These findings suggest thationic silver and a 14 nm AgNP preparation have similar neurotoxiceffects; A possible explanation for this could be the release andaction of ionic silver form the surface of AgNPs.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Acknowledgements
The authors wish to thank the Danish Food Industry Agency forfunding the project. Annette Landin, Karen Roswall and Mary-AnnGleie are thanked for their excellent technical assistance. The CoreFacility for Integrated Microscopy, Faculty of Health SciencesUniversity of Copenhagen is acknowledged for allowing access tothe TEM.
References
AshaRani PV, Low Kah MG, Hande MP, Valiyaveettil S. Cytotoxicity and genotoxicity ofsilver nanoparticles in human cells. ACS Nano 2009;3:279–90.
Barbeau A. Manganese and extrapyramidal disorders (a critical review and tribute toDr. George C. Cotzias). Neurotoxicology 1984;5:13–35.
Bondy SC, Anderson CL, Harrington ME, Prasad KN. The effects of organic and inorganiclead and mercury on neurotransmitter high-affinity transport and release mecha-nisms. Environ Res 1979;19:102–11.
Bonilla E. L-Tyrosine hydroxylase activity in the rat brain after chronic oral adminis-tration of manganese chloride. Neurobehav Toxicol 1980;2:37–41.
Bouwmeester H, Poortman J, Peters RJ, Wijma E, Kramer E, Makama S, et al. Charac-terization of translocation of silver nanoparticles and effects on whole-genomegene expression using an in vitro intestinal epithelium coculture model. ACS Nano2011;5:4091–103.
Braydich-Stolle L, Hussain S, Schlager JJ, Hofmann MC. In vitro cytotoxicity of nano-particles in mammalian germline stem cells. Toxicol Sci 2005;88:412–9.
Carlson C, Hussain SM, Schrand AM, Braydich-Stolle LK, Hess KL, Jones RL, et al. Uniquecellular interaction of silver nanoparticles: size-dependent generation of reactiveoxygen species. J Phys Chem B 2008;112:13608–19.
Carpenter DO. Effects of metals on the nervous system of humans and animals. Int JOccup Med Environ Health 2001;14:209–18.
Chandra SV, Shukla GS, Saxena DK. Manganese-induced behavioral dysfunction and itsneurochemical mechanism in growing mice. J Neurochem 1979;33:1217–21.
Clarkson TW. Metal toxicity in the central nervous system. Environ Health Perspect1987;75:59–64.

N. Hadrup et al. / NeuroToxicology 33 (2012) 416–423 423
Donaldson J. Involvement of manganese in physiological and biochemical processes:an overview. Neurotoxicology 1984;5:1–3.
Ehringer H, Hornykiewicz O. Distribution of noradrenaline and dopamine (3-hydro-xytyramine) in the human brain and their behavior in diseases of the extrapyra-midal system. Klin Wochenschr 2012;38:1236–9.
Eom HJ, Choi J. p38 MAPK activation. DNA damage, cell cycle arrest and apoptosis asmechanisms of toxicity of silver nanoparticles in Jurkat T cells. Environ Sci Technol2010;44:8337–42.
Foldbjerg R, Olesen P, Hougaard M, Dang DA, Hoffmann HJ, Autrup H. PVP-coated silvernanoparticles and silver ions induce reactive oxygen species, apoptosis andnecrosis in THP-1 monocytes. Toxicol Lett 2009;190:156–62.
Hackenberg S, Scherzed A, Kessler M, Hummel S, Technau A, Froelich K, et al. Silvernanoparticles: evaluation of DNA damage, toxicity and functional impairment inhuman mesenchymal stem cells. Toxicol Lett 2011;201:27–33.
Hadrup N, Loeschner K, Bergstrom A, Wilcks A, Gao X, Vogel U, et al. Subacute oraltoxicity investigation of nanoparticulate and ionic silver in rats. Arch Toxicol 2011.
Hadrup N, Petersen JS, Windfeld S, Risom L, Andersen CB, Nielsen S, et al. Differentialdown-regulation of aquaporin-2 in rat kidney zones by peripheral nociceptin/orphanin FQ receptor agonism and vasopressin type-2 receptor antagonism. JPharmacol Exp Ther 2007;323:516–24.
Hensler JG. Serotonergic modulation of the limbic system. Neurosci Biobehav Rev2006;30:203–14.
Hsin YH, Chen CF, Huang S, Shih TS, Lai PS, Chueh PJ. The apoptotic effect of nanosilveris mediated by a ROS- and JNK-dependent mechanism involving the mitochondrialpathway in NIH3T3 cells. Toxicol Lett 2008;179:130–9.
Hussain SM, Javorina AK, Schrand AM, Duhart HM, Ali SF, Schlager JJ. The interaction ofmanganese nanoparticles with PC-12 cells induces dopamine depletion. Toxicol Sci2006;92:456–63.
Lam HR, Lof A, Ladefoged O. Brain concentrations of white spirit components andneurotransmitters following a three week inhalation exposure of rats. PharmacolToxicol 1992;70:394–6.
Lee YS, Kim DW, Lee YH, Oh JH, Yoon S, Choi MS, et al. Silver nanoparticles induceapoptosis and G2/M arrest via PKCzeta-dependent signaling in A549 lung cells.Arch Toxicol 2011.
Li PW, Kuo TH, Chang JH, Yeh JM, Chan WH. Induction of cytotoxicity and apoptosis inmouse blastocysts by silver nanoparticles. Toxicol Lett 2010;197:82–7.
Loeschner K, Hadrup N, Qvortrup K, Larsen A, Gao X, Vogel U, et al. Distribution of silverin rats following 28 days of repeated oral exposure to silver nanoparticles or silveracetate. Part Fibre Toxicol 2011;8:18.
Mattson MP, Pedersen WA, Duan W, Culmsee C, Camandola S. Cellular and molecularmechanisms underlying perturbed energy metabolism and neuronal degenerationin Alzheimer’s and Parkinson’s diseases. Ann N Y Acad Sci 1999;893:154–75.
Miura N, Shinohara Y. Cytotoxic effect and apoptosis induction by silver nanoparticlesin HeLa cells. Biochem Biophys Res Commun 2009;390:733–7.
Oberdorster G, Oberdorster E, Oberdorster J. Nanotoxicology: an emerging disciplineevolving from studies of ultrafine particles. Environ Health Perspect 2005;113:823–39.
Piao MJ, Kang KA, Lee IK, Kim HS, Kim S, Choi JY, et al. Silver nanoparticles induceoxidative cell damage in human liver cells through inhibition of reduced glutathi-one and induction of mitochondria-involved apoptosis. Toxicol Lett 2011;201:92–100.
Powers CM, Badireddy AR, Ryde IT, Seidler FJ, Slotkin TA. Silver nanoparticlescompromise neurodevelopment in PC12 cells: critical contributions of silverion, particle size, coating, and composition. Environ Health Perspect2011a;119:37–44.
Powers CM, Levin ED, Seidler FJ, Slotkin TA. Silver exposure in developing zebrafishproduces persistent synaptic and behavioral changes. Neurotoxicol Teratol2011b;33:329–32.
Powers CM, Wrench N, Ryde IT, Smith AM, Seidler FJ, Slotkin TA. Silver impairsneurodevelopment: studies in PC12 cells. Environ Health Perspect2010;118:73–9.
Rahman MF, Wang J, Patterson TA, Saini UT, Robinson BL, Newport GD, et al. Expressionof genes related to oxidative stress in the mouse brain after exposure to silver-25nanoparticles. Toxicol Lett 2009;187:15–21.
Rungby J, Danscher G. Hypoactivity in silver exposed mice. Acta Pharmacol Toxicol(Copenh) 1984;55:398–401.
Savitt JM, Dawson VL, Dawson TM. Diagnosis and treatment of Parkinson disease:molecules to medicine. J Clin Invest 2006;116:1744–54.
Sharma HS, Hussain S, Schlager J, Ali SF, Sharma A. Influence of nanoparticles on blood-brain barrier permeability and brain edema formation in rats. Acta Neurochir Suppl2010;106:359–64.
Takahashi Y, Mizuo K, Shinkai Y, Oshio S, Takeda K. Prenatal exposure to titaniumdioxide nanoparticles increases dopamine levels in the prefrontal cortex andneostriatum of mice. J Toxicol Sci 2010;35:749–56.
Tang J, Xiong L, Wang S, Wang J, Liu L, Li J, et al. Influence of silver nanoparticles onneurons and blood–brain barrier via subcutaneous injection in rats. Appl Surf Sci2008;255:502–4.
Tang J, Xiong L, Zhou G, Wang S, Wang J, Liu L, et al. Silver nanoparticles crossingthrough and distribution in the blood–brain barrier in vitro. J Nanosci Nanotechnol2010;10:6313–7.
Trickler WJ, Lantz SM, Murdock RC, Schrand AM, Robinson BL, Newport GD, et al. Silvernanoparticle induced blood–brain barrier inflammation and increased permeabil-ity in primary rat brain microvessel endothelial cells. Toxicol Sci 2010;118:160–70.
Wang J, Rahman MF, Duhart HM, Newport GD, Patterson TA, Murdock RC, et al.Expression changes of dopaminergic system-related genes in PC12 cells inducedby manganese, silver, or copper nanoparticles. Neurotoxicology 2009;30:926–33.
Webster RA. Neurotransmitters, drugs, and brain function. John Wiley & Sons, Ltd;2011.
Wei L, Tang J, Zhang Z, Chen Y, Zhou G, Xi T. Investigation of the cytotoxicity mechanismof silver nanoparticles in vitro. Biomed Mater 2010;5:1–6.
Woodrow Wilson International Center for Scholars. Project on emerging nanotechnol-ogies., 2011 www.nanotechprojects.org/inventories/consumer.
Zhang Z, Zhao B, Hu L. PVP Protective Mechanism of Ultrafine Silver Powder Synthe-sized by Chemical Reduction Processes. J Solid State Chem 1996;121:105–10.





























