Embed Size (px)
Citation preview

Epilepsy & Behavior xxx (2010) xxx–xxx
ARTICLE IN PRESS
Contents lists available at ScienceDirect
Epilepsy & Behavior
journal homepage: www.elsevier .com/locate /yebeh
The sleep–wake cycle in adult rats following pilocarpine-induced temporallobe epilepsy
Gabriela Matos a, Rodrigo Tsai b, Marcus Vinícius Baldo c, Isac de Castro d, Koichi Sameshima e,Angela Cristina Valle a,*
a School of Medicine, Department of Pathology, University of São Paulo, São Paulo, Brazilb Institute of Statistics, University of Campinas, Campinas, Brazilc Institute of Biomedical Sciences, Department of Physiology and Biophysics, University of São Paulo, São Paulo, Brazild Division of Neurology, Federal University of São Paulo, São Paulo, Brazile School of Medicine, Department of Radiology, University of São Paulo, São Paulo, Brazil
a r t i c l e i n f o a b s t r a c t
Article history:Received 23 September 2009Revised 12 November 2009Accepted 18 November 2009Available online xxxx
Keywords:Sleep–wake cycleEpilepsyPilocarpineStatus epilepticusSeizures
1525-5050/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.yebeh.2009.11.015
* Corresponding author. Address: Department ofPaulo, Av. Dr. Arnaldo, 455, Room 4301, CerqueiraBrazil. Fax: +55 11 30617040.
E-mail address: [email protected] (A.C. Valle).
Please cite this article in press as: Matos G et al.(2010), doi:10.1016/j.yebeh.2009.11.015
The relationship between sleep and epilepsy is both complex and clinically significant. Temporal lobe epi-lepsy (TLE) influences sleep architecture, while sleep plays an important role in facilitating and/or inhib-iting possible epileptic seizures. The pilocarpine experimental model reproduces several features ofhuman temporal lobe epilepsy and is one of the most widely used models in basic research. The aimof the present study was to characterize, behaviorally and electrophysiologically, the phases of sleep–wake cycles (SWC) in male rats with pilocarpine-induced epilepsy. Epileptic rats presented spikes inall phases of the SWC as well as atypical cortical synchronization during attentive wakefulness and par-adoxical sleep. The architecture of the sleep–wake phases was altered in epileptic rats, as was the integ-rity of the SWC. Because our findings reproduce many relevant features observed in patients withepilepsy, this model is suitable to study sleep dysfunction in epilepsy.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
The relationship between sleep–wake states and epilepsy hasbeen known since ancient times. In recent decades, an astonishingnumber of publications have illustrated the complexity of theinteractions between clinical seizures and sleep [1–3]. It has beenwell established that frequent sleep cycle disruption results inworsening of seizures, and further impairment of sleep can beresponsible for intractability of epilepsy [4,5]. Furthermore, evi-dence has suggested that the presence of epilepsy itself can disruptsleep, even when the seizures occur only during wakefulness [6].
Many researchers have attempted to achieve a better under-standing of how seizures are triggered by sleep disruptions andalso how they can compromise sleep architecture. According toprevious reports, some epileptic events are activated by sleep, par-ticularly non-rapid eye movement (NREM) sleep [7,8]. There is rec-iprocity in the system in which epilepsy alters the organization ofthe sleep–wake cycle and sleep architecture both acutely throughthe disruption of continuity and chronically by impairing the orga-
ll rights reserved.
Pathology, University of SãoCésar 01246903, São Paulo,
The sleep–wake cycle in adult
nization and microstructure of sleep [5,6,9,10]. Likewise, sleepdeprivation leads to sensitization to seizures [11–14].
The sleep–epilepsy relationship differs, however, in accordancewith the kind of epilepsy or epileptic syndrome. Even though themechanisms are well established for some forms of idiopathic epi-lepsies such as absence epilepsy, which usually occur during thesleep state, the mechanisms for symptomatic partial epilepsy arestill unclear [8,13,15,16].
Experimental models have been widely established as essentialtools to provide the basis for understanding epilepsy [17–19]. Oneof the most valuable experimental models is the induction of tem-poral lobe epilepsy with pilocarpine [20,21]. In rats, this drug in-duces limbic seizures, which become secondarily generalized,leading to status epilepticus [20]. Nevertheless, the occurrence ofsustained seizure activity is accompanied by widespread damagein the forebrain of both rats and mice after systemic injections ofhigh doses of cholinergic agonists [20,22,23]. Consequently, theanimals become devoid of important function controls, includingthose related to biological rhythms.
Although this model has been widely used to understand epi-leptic mechanisms, several behavioral aspects related to thesleep–wake cycle are not completely known. Likewise, the preciseinteraction that leads the phases of the sleep–wake cycle to triggerseizures has not been elucidated using this model. Very few studieshave been designed to clarify some of these aspects [21,24–26]. In
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav

2 G. Matos et al. / Epilepsy & Behavior xxx (2010) xxx–xxx
ARTICLE IN PRESS
an elaborating study, Ferreira et al. [24] investigated the epilepticdisturbances along the sleep–wake phases in rats submitted to sta-tus epilepticus (SE) in early life. Although abnormal potentialswere present in every phase of the sleep–wake cycle, regardlessof coincidence with motor manifestations, no significant altera-tions were found in the architecture of cycles of these animals[24]. Thus, this could be attributed to a high level of neuroplasticityduring brain development that prevents major physiological defi-cits in the adult phase. However, there are no available data con-cerning the effects of pilocarpine-induced epilepsy on sleep–wake phases in adult epileptic rats. This lack of information callsfor further study into the main features of the sleep–wake cyclearchitecture in pilocarpine-induced epilepsy, which could contrib-ute to a better understanding of many other variables such as sei-zure frequency or even therapeutic approaches, as these factors areprobably dependent on the sleep–wake phase.
Thus far, no systematic studies have been performed to evaluatethe recovery and precise reorganization of the sleep–wake archi-tecture after the establishment of epilepsy following pilocarpine-induced SE in adult rats. Therefore, to gain a better understandingof the interactions between circadian rhythms and epilepsy, theaim of this study was to present a comprehensive behavioral andelectrophysiological characterization of the states and phases ofthe sleep–wake cycle in chronic epileptic rats with pilocarpine-in-duced temporal lobe epilepsy (TLE).
2. Methods
Twelve male Wistar rats (200–250 g), six nonepileptic (NON-EPI) and six epileptic (EPI), were selected for the present study.The animals were singly housed in individual plastic cages in atemperature (22 ± 2 �C)- and light/dark cycle (12 hours light onat 7 AM)-controlled environment with free access to food andwater. This study was approved by the local Animal Care EthicalCommittee (CAPPESQ-HC/USP No. 665/05).
2.1. Pilocarpine-induced status epilepticus
To develop the pilocarpine model of epilepsy, SE was inducedwith a single dose (360 mg/kg, IP) of pilocarpine 30 minutes aftersubcutaneous injection of methylscopolamine (0.1 mg/kg, IP)[20,21,27]. A nonepileptic group was injected with saline instead(0.9% NaCl solution). To obtain the pattern of occurrence for epilep-tic seizures in terms of frequency in light/dark periods, the animalswere continuously monitored with a digital video camera systemfor 10 days after the 40th day following SE. The seizures recordedwere similar to Racine’s kindled stages IV (bilateral forelimb clonusand rearing) and V (stage IV with additional loss of postural tone).
2.2. Electrode implantation
Fifty days after SE was induced by systemic administration ofpilocarpine, both epileptic and nonepileptic rats were subjectedto stereotaxic surgical procedures for electrode implantation. Un-der ketamine anesthesia, the animals were bilaterally implantedwith 150-lm-diameter nichrome bipolar electrodes over neocorti-cal area 3 (AP: �1.5 mm, ML: ±3.0 mm) according to skull refer-ences [28,29] that make it possible to implant the electrodesover specific cytoarchitectonic and functional areas through 1-mm holes in the bone [30,31]. Similar but straight electrodes par-allel and isolated except at the end cut cross section were stereo-taxically implanted into the CA1 hippocampal field (AP:�3.6 mm, ML: ±1.2 mm, DV: �3.0 mm) according to the Paxinosand Watson atlas [32] (Fig. 1). In addition to the electrodes for elec-tro-oscillographic recordings, we implanted electrodes in each
Please cite this article in press as: Matos G et al. The sleep–wake cycle in adult(2010), doi:10.1016/j.yebeh.2009.11.015
external canthus of both eyes, in the trapezius muscles, and inthe muscle pad that moves the snout (rostrum) and vibrissae to re-cord movements of the eye, head, and rostrum + vibrissae, respec-tively. Electrodes were implanted while taking extreme care toreduce edema and other brain injury that could interfere in theelectro-oscillographic recordings.
2.3. Electro-oscillographic recordings
Ten days after the surgical procedure, the rats were placed in aFaraday cage and connected to the input panel of a 21-channel Ni-hon–Kohden electroencephalograph (Neurofax EEG 4400) for 24hours of continuous recording. Animals were habituated to thecables and to the recording environment for 3 days before theirparticipation in experiments. Electro-oscillographic recordingswere monitored during the light and dark cycles, each lasting 12hours (food and water ad libitum). The behavior of the rats wascontinuously monitored during recordings. The recordings weresubmitted to visual analysis for identification of each of thesleep–wake phases [28,33–35] by a researcher who was blind tothe experiment. The electro-oscillographic recordings were per-formed in a seizure-free condition.
2.4. Histology
On completion of the experiments, each animal was sacrificedwith an overdose of the anesthetic. Brains were then removed forNissl staining to confirm correct electrode positioning in thehippocampus.
2.5. Statistical analysis
To analyze the architecture of sleep–wake phases throughoutthe 24-hour recording, a MANOVA with a Tukey post hoc testwas used. To explore the organization of sleep–wake cycles andtheir pattern of occurrence along the light–dark cycle, a two-wayANOVA was performed with a post hoc Tukey test. For all teststhe significance level was set at 5%.
3. Results
This study represents both ethological and detailed quantitativeanalyses of the sleep–wake cycles of epileptic and nonepilepticrats.
3.1. Visual analysis
The 24-hour continuous video camera monitoring alloweddelineation of a seizure distribution pattern along light–dark peri-ods. The majority of seizures occurred during the light period(58%), primarily in the afternoon (72.4%) in chronic epileptic rats,whereas seizures during the night (42%) occurred randomly(Fig. 2).
The recordings were subjected to visual analysis to identifyeach sleep–wake phase according to previously described specifi-cations [28,33,34].
Nonepileptic animals manifested all the typical electrophysio-logical features of the sleep–wake phases (Figs. 3a, c, e, g, i),whereas epileptic rats showed some important differences with re-spect to their occurrence. The attentive wakefulness (AW) of theseanimals was characterized by high electromyographic activity inthe trapezium muscles, in addition to increased rostrum and eyemovements during exploratory behavior (Fig. 3b). The corticaldesynchronization was often disrupted by unexpected isolated orclustered low-frequency waves with high voltage, while the hippo-
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav
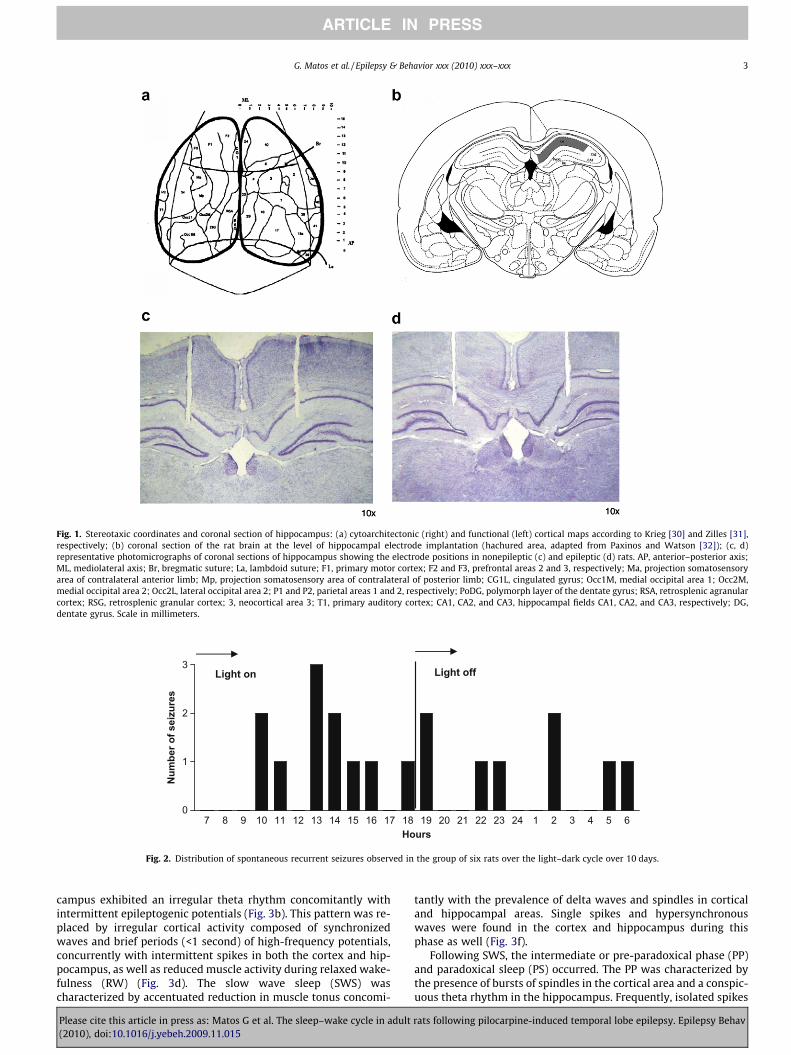
Fig. 1. Stereotaxic coordinates and coronal section of hippocampus: (a) cytoarchitectonic (right) and functional (left) cortical maps according to Krieg [30] and Zilles [31],respectively; (b) coronal section of the rat brain at the level of hippocampal electrode implantation (hachured area, adapted from Paxinos and Watson [32]); (c, d)representative photomicrographs of coronal sections of hippocampus showing the electrode positions in nonepileptic (c) and epileptic (d) rats. AP, anterior–posterior axis;ML, mediolateral axis; Br, bregmatic suture; La, lambdoid suture; F1, primary motor cortex; F2 and F3, prefrontal areas 2 and 3, respectively; Ma, projection somatosensoryarea of contralateral anterior limb; Mp, projection somatosensory area of contralateral of posterior limb; CG1L, cingulated gyrus; Occ1M, medial occipital area 1; Occ2M,medial occipital area 2; Occ2L, lateral occipital area 2; P1 and P2, parietal areas 1 and 2, respectively; PoDG, polymorph layer of the dentate gyrus; RSA, retrosplenic agranularcortex; RSG, retrosplenic granular cortex; 3, neocortical area 3; T1, primary auditory cortex; CA1, CA2, and CA3, hippocampal fields CA1, CA2, and CA3, respectively; DG,dentate gyrus. Scale in millimeters.
Light off
7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 1 2 3 4 5 60
1
2
3
Hours
Num
ber o
f sei
zure
s
Light on
Fig. 2. Distribution of spontaneous recurrent seizures observed in the group of six rats over the light–dark cycle over 10 days.
G. Matos et al. / Epilepsy & Behavior xxx (2010) xxx–xxx 3
ARTICLE IN PRESS
campus exhibited an irregular theta rhythm concomitantly withintermittent epileptogenic potentials (Fig. 3b). This pattern was re-placed by irregular cortical activity composed of synchronizedwaves and brief periods (<1 second) of high-frequency potentials,concurrently with intermittent spikes in both the cortex and hip-pocampus, as well as reduced muscle activity during relaxed wake-fulness (RW) (Fig. 3d). The slow wave sleep (SWS) wascharacterized by accentuated reduction in muscle tonus concomi-
Please cite this article in press as: Matos G et al. The sleep–wake cycle in adult(2010), doi:10.1016/j.yebeh.2009.11.015
tantly with the prevalence of delta waves and spindles in corticaland hippocampal areas. Single spikes and hypersynchronouswaves were found in the cortex and hippocampus during thisphase as well (Fig. 3f).
Following SWS, the intermediate or pre-paradoxical phase (PP)and paradoxical sleep (PS) occurred. The PP was characterized bythe presence of bursts of spindles in the cortical area and a conspic-uous theta rhythm in the hippocampus. Frequently, isolated spikes
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav
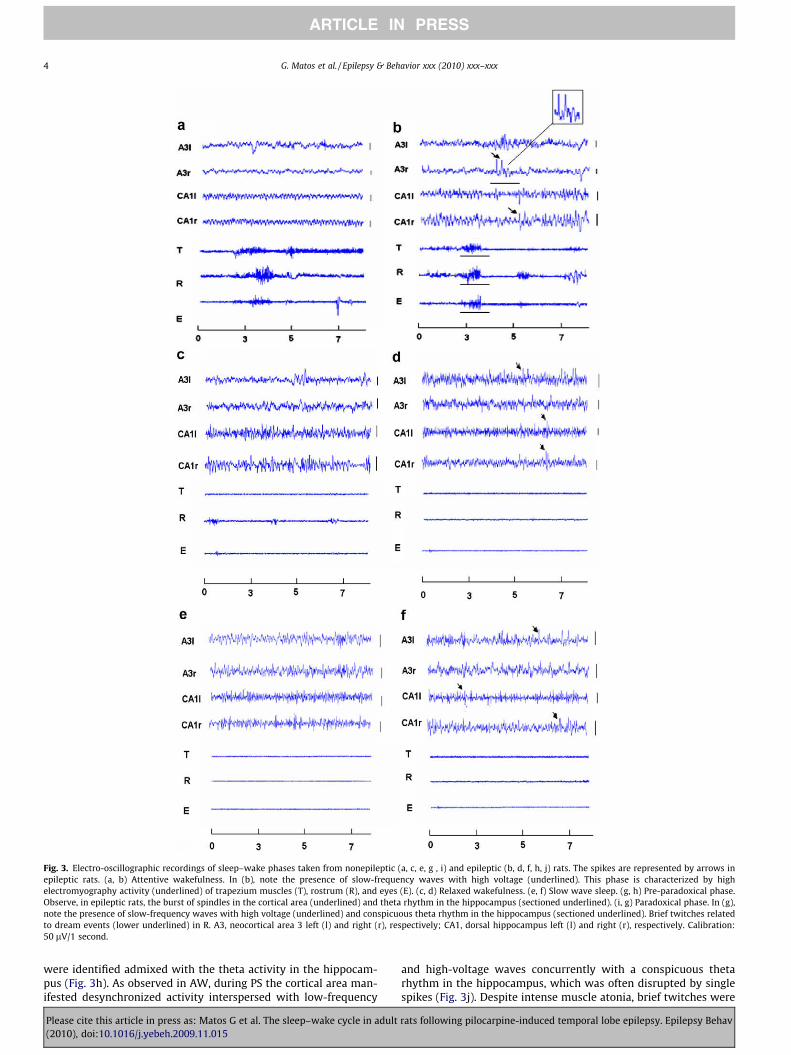
Fig. 3. Electro-oscillographic recordings of sleep–wake phases taken from nonepileptic (a, c, e, g , i) and epileptic (b, d, f, h, j) rats. The spikes are represented by arrows inepileptic rats. (a, b) Attentive wakefulness. In (b), note the presence of slow-frequency waves with high voltage (underlined). This phase is characterized by highelectromyography activity (underlined) of trapezium muscles (T), rostrum (R), and eyes (E). (c, d) Relaxed wakefulness. (e, f) Slow wave sleep. (g, h) Pre-paradoxical phase.Observe, in epileptic rats, the burst of spindles in the cortical area (underlined) and theta rhythm in the hippocampus (sectioned underlined). (i, g) Paradoxical phase. In (g),note the presence of slow-frequency waves with high voltage (underlined) and conspicuous theta rhythm in the hippocampus (sectioned underlined). Brief twitches relatedto dream events (lower underlined) in R. A3, neocortical area 3 left (l) and right (r), respectively; CA1, dorsal hippocampus left (l) and right (r), respectively. Calibration:50 lV/1 second.
4 G. Matos et al. / Epilepsy & Behavior xxx (2010) xxx–xxx
ARTICLE IN PRESS
were identified admixed with the theta activity in the hippocam-pus (Fig. 3h). As observed in AW, during PS the cortical area man-ifested desynchronized activity interspersed with low-frequency
Please cite this article in press as: Matos G et al. The sleep–wake cycle in adult(2010), doi:10.1016/j.yebeh.2009.11.015
and high-voltage waves concurrently with a conspicuous thetarhythm in the hippocampus, which was often disrupted by singlespikes (Fig. 3j). Despite intense muscle atonia, brief twitches were
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav
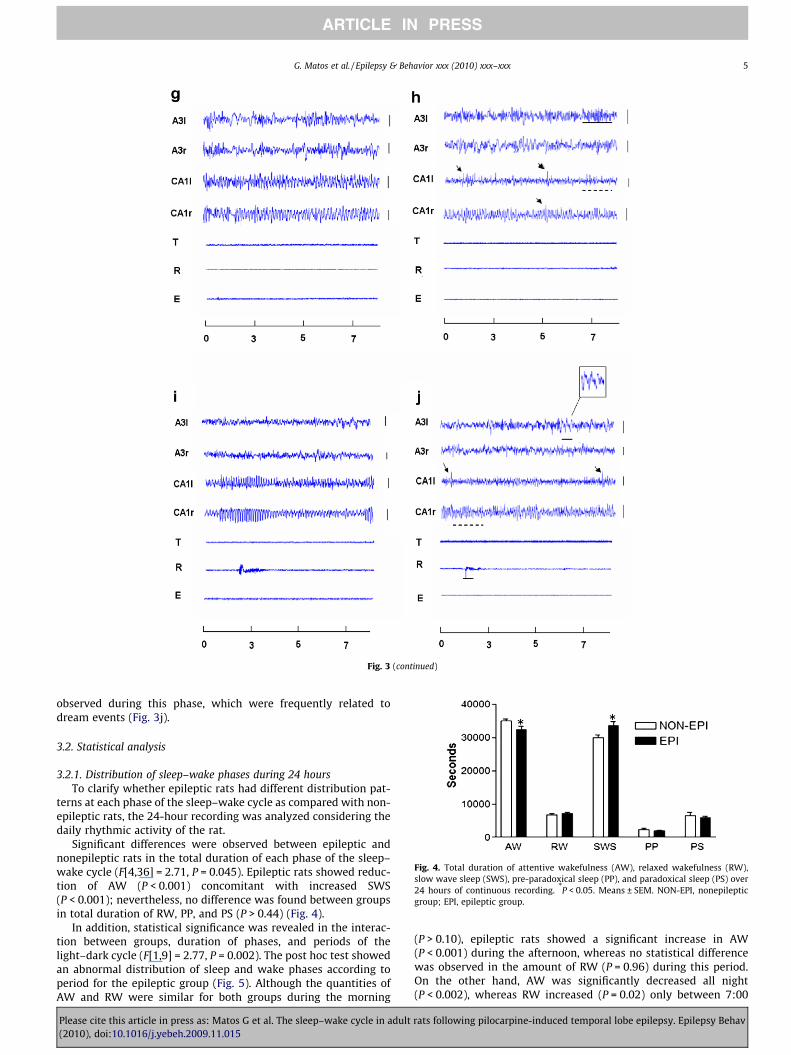
Fig. 3 (continued)
Fig. 4. Total duration of attentive wakefulness (AW), relaxed wakefulness (RW),slow wave sleep (SWS), pre-paradoxical sleep (PP), and paradoxical sleep (PS) over24 hours of continuous recording. *P < 0.05. Means ± SEM. NON-EPI, nonepilepticgroup; EPI, epileptic group.
G. Matos et al. / Epilepsy & Behavior xxx (2010) xxx–xxx 5
ARTICLE IN PRESS
observed during this phase, which were frequently related todream events (Fig. 3j).
3.2. Statistical analysis
3.2.1. Distribution of sleep–wake phases during 24 hoursTo clarify whether epileptic rats had different distribution pat-
terns at each phase of the sleep–wake cycle as compared with non-epileptic rats, the 24-hour recording was analyzed considering thedaily rhythmic activity of the rat.
Significant differences were observed between epileptic andnonepileptic rats in the total duration of each phase of the sleep–wake cycle (F[4,36] = 2.71, P = 0.045). Epileptic rats showed reduc-tion of AW (P < 0.001) concomitant with increased SWS(P < 0.001); nevertheless, no difference was found between groupsin total duration of RW, PP, and PS (P > 0.44) (Fig. 4).
In addition, statistical significance was revealed in the interac-tion between groups, duration of phases, and periods of thelight–dark cycle (F[1,9] = 2.77, P = 0.002). The post hoc test showedan abnormal distribution of sleep and wake phases according toperiod for the epileptic group (Fig. 5). Although the quantities ofAW and RW were similar for both groups during the morning
Please cite this article in press as: Matos G et al. The sleep–wake cycle in adult(2010), doi:10.1016/j.yebeh.2009.11.015
(P > 0.10), epileptic rats showed a significant increase in AW(P < 0.001) during the afternoon, whereas no statistical differencewas observed in the amount of RW (P = 0.96) during this period.On the other hand, AW was significantly decreased all night(P < 0.002), whereas RW increased (P = 0.02) only between 7:00
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav
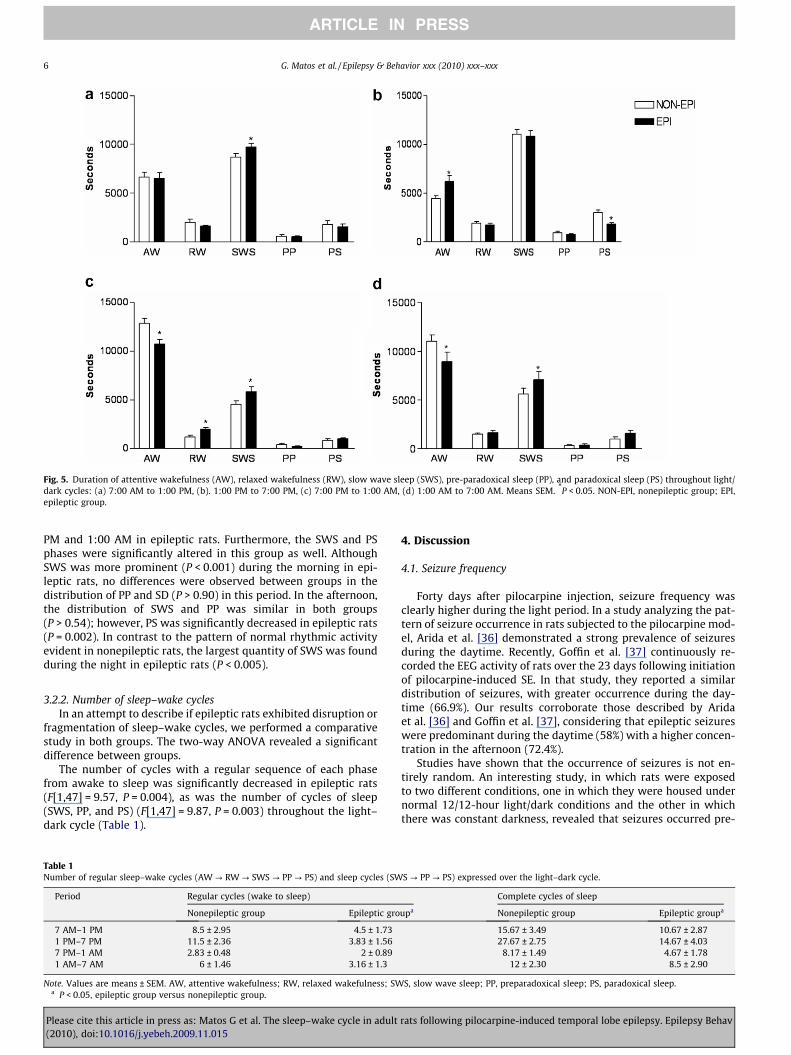
Fig. 5. Duration of attentive wakefulness (AW), relaxed wakefulness (RW), slow wave sleep (SWS), pre-paradoxical sleep (PP), and paradoxical sleep (PS) throughout light/dark cycles: (a) 7:00 AM to 1:00 PM, (b). 1:00 PM to 7:00 PM, (c) 7:00 PM to 1:00 AM, (d) 1:00 AM to 7:00 AM. Means SEM. *P < 0.05. NON-EPI, nonepileptic group; EPI,epileptic group.
6 G. Matos et al. / Epilepsy & Behavior xxx (2010) xxx–xxx
ARTICLE IN PRESS
PM and 1:00 AM in epileptic rats. Furthermore, the SWS and PSphases were significantly altered in this group as well. AlthoughSWS was more prominent (P < 0.001) during the morning in epi-leptic rats, no differences were observed between groups in thedistribution of PP and SD (P > 0.90) in this period. In the afternoon,the distribution of SWS and PP was similar in both groups(P > 0.54); however, PS was significantly decreased in epileptic rats(P = 0.002). In contrast to the pattern of normal rhythmic activityevident in nonepileptic rats, the largest quantity of SWS was foundduring the night in epileptic rats (P < 0.005).
3.2.2. Number of sleep–wake cyclesIn an attempt to describe if epileptic rats exhibited disruption or
fragmentation of sleep–wake cycles, we performed a comparativestudy in both groups. The two-way ANOVA revealed a significantdifference between groups.
The number of cycles with a regular sequence of each phasefrom awake to sleep was significantly decreased in epileptic rats(F[1,47] = 9.57, P = 0.004), as was the number of cycles of sleep(SWS, PP, and PS) (F[1,47] = 9.87, P = 0.003) throughout the light–dark cycle (Table 1).
Table 1Number of regular sleep–wake cycles (AW ? RW ? SWS ? PP ? PS) and sleep cycles (SW
Period Regular cycles (wake to sleep)
Nonepileptic group Epileptic gro
7 AM–1 PM 8.5 ± 2.95 4.5 ± 1.731 PM–7 PM 11.5 ± 2.36 3.83 ± 1.567 PM–1 AM 2.83 ± 0.48 2 ± 0.891 AM–7 AM 6 ± 1.46 3.16 ± 1.3
Note. Values are means ± SEM. AW, attentive wakefulness; RW, relaxed wakefulness; SWa P < 0.05, epileptic group versus nonepileptic group.
Please cite this article in press as: Matos G et al. The sleep–wake cycle in adult(2010), doi:10.1016/j.yebeh.2009.11.015
4. Discussion
4.1. Seizure frequency
Forty days after pilocarpine injection, seizure frequency wasclearly higher during the light period. In a study analyzing the pat-tern of seizure occurrence in rats subjected to the pilocarpine mod-el, Arida et al. [36] demonstrated a strong prevalence of seizuresduring the daytime. Recently, Goffin et al. [37] continuously re-corded the EEG activity of rats over the 23 days following initiationof pilocarpine-induced SE. In that study, they reported a similardistribution of seizures, with greater occurrence during the day-time (66.9%). Our results corroborate those described by Aridaet al. [36] and Goffin et al. [37], considering that epileptic seizureswere predominant during the daytime (58%) with a higher concen-tration in the afternoon (72.4%).
Studies have shown that the occurrence of seizures is not en-tirely random. An interesting study, in which rats were exposedto two different conditions, one in which they were housed undernormal 12/12-hour light/dark conditions and the other in whichthere was constant darkness, revealed that seizures occurred pre-
S ? PP ? PS) expressed over the light–dark cycle.
Complete cycles of sleep
upa Nonepileptic group Epileptic groupa
15.67 ± 3.49 10.67 ± 2.8727.67 ± 2.75 14.67 ± 4.03
8.17 ± 1.49 4.67 ± 1.7812 ± 2.30 8.5 ± 2.90
S, slow wave sleep; PP, preparadoxical sleep; PS, paradoxical sleep.
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav

G. Matos et al. / Epilepsy & Behavior xxx (2010) xxx–xxx 7
ARTICLE IN PRESS
dominantly during the daytime in both conditions regardless of alack of circadian rhythm [38]. The authors suggested that seizuresare not only influenced by external stimuli and that there may be adirect influence of an endogenous circadian stimulus.
In fact, there are many conflicting data concerning the role ofthe biological clock in the occurrence of seizures. Very few studieshave sought to elucidate the influence of suprachiasmatic nuclei onlimbic seizures. Sanabria et al. [39] demonstrated abnormalexpression of c-fos in the suprachiasmatic nuclei of animals sub-jected to 30 minutes of a single photic stimulus during the earlysubjective night after 7 days of total darkness, showing a clearfunctional change in response to an external stimulus. On the otherhand, a study that examined the effects of temporal lobe seizureson the daily rhythm of locomotor activity in rats showed that, de-spite the changes in spontaneous locomotor activity rhythmparameters, histological analysis indicated no significant decreasein neuronal density in suprachiasmatic nuclei [26].
The interaction between epilepsy and circadian rhythms is verycomplex and cannot be attributed to a single factor or region of thecentral nervous system. Therefore, further studies are required toreveal the mechanisms underlying the relationship between occur-rence of seizures and circadian rhythms.
4.2. Visual analysis of electro-oscillographic recordings
Although all behavioral and electrophysiological features ofsleep–wake phases were present in both epileptic and nonepilepticrats, our results disclosed important details about pilocarpine-in-duced SE that warrant close attention.
In the epileptic group, the spikes detected in both cortical andhippocampal areas during the sleep–wake cycle could be attrib-uted to a dysfunction of compromised structures following pilocar-pine-induced SE. In fact, some studies have shown interictal spikesin acute and chronic epileptic rats following SE in the pilocarpinemodel [21,24,25,37].
Another important electro-oscillographic finding in epilepticanimals was the intrusion of isolated or clustered low-frequencywaves with high voltage in the PS and AW phases. These atypicalwaves could be explained by lesions in some important centersresponsible for generating and sustaining desynchronization,which are characteristic of AW and PS. It is well known that pilo-carpine-induced SE provokes extensive damage in the central ner-vous system during the acute [40] and chronic [20,21,41] periods.Nevertheless, there is little evidence indicating that a specific orlocalized lesion could be responsible for these atypical waves. San-abria et al. [42] identified important alterations in neocortical areasof epileptic animals following pilocarpine-induced SE. Theseauthors demonstrated conspicuous alterations in superficial layers(II–IV), with relative enlargement of layers V and VI compared withthe nonepileptic group. At this point, we emphasize that the corti-cal area studied in the present work was equivalent to that ana-lyzed by Sanabria et al. Therefore, it is plausible to suggest thatthe electro-oscillographic abnormalities found in our recordingscould be, at least in part, due to cortical lesions. However, only acomprehensive study including both cortical and subcortical areascould clarify the mechanisms underlying this phenomenon.
4.3. Distribution of sleep–wake phases along the light–darkphotoperiod
Considering the rat daily rhythmic activity, the data in thisstudy from comparison of epileptic and nonepileptic groups re-vealed important characteristics concerning the distribution ofsleep–wake phases along the light–dark photoperiod. During the24-hour seizure-free recordings, epileptic animals manifestedabnormal distribution of AW and SWS.
Please cite this article in press as: Matos G et al. The sleep–wake cycle in adult(2010), doi:10.1016/j.yebeh.2009.11.015
Although it is generally accepted that rodents usually present thehighest level of activity during the dark cycle, epileptic rats showedthe opposite biological rhythm. These animals presented an in-creased quantity of SWS and RW, with a concomitant decrease inAW during the night. It is interesting to note that in epileptic rats,the highest level of RW was observed between 7 PM and 12 AM, aperiod in which rats are expected to exhibit more exploratory behav-ior. This inversion of the rest–activity cycle in epileptic rats is diffi-cult to explain at this moment inasmuch as our study was notfocused on alterations in circadian structures resulting from SE fol-lowing pilocarpine injection. Additionally, only a few studies haveattempted to explain this interaction in the pilocarpine model, andthus data supporting our findings are still scarce [24].
Furthermore, most of the methodologies adopted to classify thesleep–wake cycle in experimental models have been based only onthree phases: wakefulness, slow wave sleep, and paradoxical sleep.In general, RW is ignored during staging as one of the phases of thesleep–wake cycle [10,43]. In fact, the relaxed awake state is animportant phase that reflects the transition from alertness to sleep,in which alertness mechanisms decrease, whereas those related tosynchronization are activated [28,33,44]. One of the most importantcharacteristics related to RW is the high susceptibility to the occur-rence of specific kinds of seizures, which are strongly recognized asbeing dependent on synchronizing structures for their generation[45,46].
Moreover, epileptic rats unexpectedly showed increased SWSduring the night; they also displayed an enhanced total duration ofthis phase during the morning when compared with nonepilepticrats. One possible cause could be an impaired capability to shiftsleep–wake phases according to the alternating light–dark cycles.The epileptic rats were indifferent to most of the exogenous clues.In fact, studies have shown that epilepsy can influence the organiza-tion of biological rhythms, changing their response to external stim-uli [26,38,39,47,48]. The increased SWS observed in both night andmorning periods seemed to be balanced by the strong increase inAW during the afternoon. In addition, epileptic rats showed a signif-icant reduction of PS in the same period, which is also in agreementwith the hypothesis of possible dysfunction in the regulation of thecyclicity of the sleep–wake cycle. In fact, in an experiment with thekindling model in rats, Bastlund et al. [10] showed a disturbance insleep architecture accompanied by neuronal loss in the dorsomedialhypothalamus, a region important for normal sleep regulation.
Notwithstanding all the important alterations described above,a discernible fragmentation of the sleep–wake cycle was evident inepileptic rats. The number of cycles that exhibited a regular se-quence of each phase from awake to sleep was significantly de-creased in these animals. In fact, it is well established thattemporal lobe epilepsy affects the distribution of the standardsleep parameters (total sleep time, time in bed, sleep efficiency,sleep latency, wake after sleep onset, amounts of REM and NREMsleep stages, and REM latency) and the phasic events (e.g., K-com-plexes, delta bursts, intermittent alpha rhythm, vertex sharp tran-sients, arousal), respectively [5,6,9,49–51] designated asmacrostructure [52] and microstructure of sleep [52,53].
Because our data reproduce many relevant features observedclinically in patients with epilepsy, we suggest our work as a suit-able model to study the role of sleep dysfunction in epilepsy. Thiswork underscores the importance of considering all aspects relatedto alterations in the sleep–wake cycle for studies that use pilocar-pine as a model for temporal lobe epilepsy.
Acknowledgments
The authors express their cordial thanks to Eloisa Vicente andAdilson da Silva Alves for their excellent assistance during theproject.
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav

8 G. Matos et al. / Epilepsy & Behavior xxx (2010) xxx–xxx
ARTICLE IN PRESS
References
[1] Méndez M, Radtke RA. Interactions between sleep and epilepsy. J ClinNeurophysiol 2001;18:106–27.
[2] Malow BA. The interaction between sleep and epilepsy. Epilepsia2007;48(Suppl. 9):6–8.
[3] Kotagal P, Yardi N. The relationship between sleep and epilepsy. Semin PediatrNeurol 2008;15:42–9.
[4] Baldy-Moulinier M. Inter-relationships between sleep and epilepsy. In: PedleyTA, Meldrum BS, editors. Recent advances in epilepsy, vol.3. Edinburgh: Churchill Livingstone; 1986. p. 37–55.
[5] Crespel A, Coubes P, Baldy-Moulinier M. Sleep influence on seizures andepilepsy effects on sleep in partial frontal and temporal lobe epilepsies. ClinNeurophysiol 2000;111(Suppl. 2):S54–9.
[6] Bazil CW, Castro LH, Walczak TS. Reduction of rapid eye movement sleep bydiurnal and nocturnal seizures in temporal lobe epilepsy. Arch Neurol2000;57:363–8.
[7] Bazil CW, Walczak TS. Effects of sleep and sleep stage on epileptic andnonepileptic seizures. Epilepsia 1997;38:56–62.
[8] Herman ST, Walczak TS, Bazil CW. Distribution of partial seizures during thesleep wake cycle: differences by seizure onset site. Neurology2001;56:1453–9.
[9] Touchon J, Baldy–Moulinier M, Billiard M, Besset A, Cadilhac J. Sleeporganization and epilepsy. In: Degen R, Rodin EA, editors. Epilepsy, sleep andsleep deprivation. Amsterdam: Elsevier; 1991. p. 73–81.
[10] Bastlund JF, Jennum P, Mohapel P, Penschuck S, Watson WP. Spontaneousepileptic rats show changes in sleep architecture and hypothalamic pathology.Epilepsia 2005;46:934–8.
[11] Degen R. A study of the diagnostic value of waking and sleep EEGs after sleepdeprivation in epileptic patients on anticonvulsive therapy.Electroencephalogr Clin Neurophysiol 1980;49:577–84.
[12] Shouse MN. Sleep deprivation increases susceptibility to kindled and penicillinseizure events during all waking and sleep states in cats. Sleep 1988;11:162–71.
[13] Broughton R. Sleep and sleep deprivation studies in epilepsy, EEG handbook.In: Wada JA, Ellingsen RJ, editors. Clinical neurophysiology ofepilepsy. Amsterdam: Elsevier; 1990. p. 89–119.
[14] Badawy RA, Curatolo JM, Newton M, Berkovic SF, Macdonell RA. Sleepdeprivation increases cortical excitability in epilepsy: syndrome-specificeffects. Neurology 2006;67:1018–22.
[15] Sato S, Dreifuss F, Penry JK. The effect of sleep on spike–wave discharges inabsence seizures. Neurology 1973;2:1335–45.
[16] Hofstra WA, Grootemarsink BE, Dieker R, van der Palen J, de Weerd AW.Temporal distribution of clinical seizures over the 24-h day: a retrospectiveobservational study in a tertiary epilepsy clinic. Epilepsia 2009;50:2019–26.
[17] Dichter MA, Ayala GF. Cellular mechanisms of epilepsy: a status report.Science 1987;237:157–64.
[18] Sharma AK, Reams RY, Jordan WH, Miller MA, Thacker HL, Snyder PW. Mesialtemporal lobe epilepsy: pathogenesis, induced rodent models and lesions.Toxicol Pathol 2007;35:984–99.
[19] Curia G, Longo D, Biagini G, Jones RS, Avoli M. The pilocarpine model oftemporal lobe epilepsy. J Neurosci Methods 2008;172:143–57.
[20] Turski WA, Cavalheiro EA, Schwarz M, Czuczwar S, Kleinrok Z, Turski L. Limbicseizures produced by pilocarpine in rats: behavioral, electroencephalographicand neuropathological study. Behav Brain Res 1983;9:315–35.
[21] Cavalheiro EA, Leite JP, Bortolotto ZA, Turski WA, Ikonomidou C, Turski L. Long-term effects of pilocarpine in rats: structural damage of the brain triggerskindling and spontaneous recurrent seizures. Epilepsia 1991;32:778–82.
[22] Olney JW, de Gubareff T, Sloviter RS. ‘‘Epileptic” brain damage in rats inducedby sustained electrical stimulation of the perforant path. II. Ultrastructuralanalysis of acute hippocampal pathology. Brain Res Bull 1983;10:699–712.
[23] Turski WA, Cavalheiro EA, Bortolotto ZA, Mello LM, Schwarz M, Turski L. Seizuresproduced by pilocarpine in mice: a behavioral, electroencephalographic andmorphological analysis. Brain Res 1984;321:237–53.
[24] Ferreira BL, Valle AC, Cavalheiro EA, Timo-Iaria C. Prevalence of epilepticseizures during the wakefulness–sleep cycle in rats. Dev Neurosci1999;21:339–404.
[25] Ferreira BL, Valle AC, Cavalheiro EA, Timo-Iaria C. Absence-like seizures inadult rats following pilocarpine-induced status epilepticus early in life. Braz JMed Biol Res 2003;36:1685–94.
[26] Stewart LS, Leung LS. Temporal lobe seizures alter the amplitude and timing ofrat behavioral rhythms. Epilepsy Behav 2003;4:153–60.
Please cite this article in press as: Matos G et al. The sleep–wake cycle in adult(2010), doi:10.1016/j.yebeh.2009.11.015
[27] Cavalheiro EA. The pilocarpine model of epilepsy. Ital J Neurol Sci1995;16:33–7.
[28] Timo-Iaria C, Negrão N, Schmidek WR, Hoshino K, Lobato de Menezes CE, Lemeda Rocha T. Phases and states of sleep in the rat. Physiol Behav1970;5:1057–62.
[29] Timo-Iaria C, Negrão N, Pereira WC. Spindling during natural and barbituratesleep, in the isolated cerebrum and in recruiting in the rat brain. In: Kao FF,Koizumi K, Vassale M, editors. Research in physiology. Bologna: A. Gaggi;1971. p. 567–80.
[30] Krieg WJS. Connections of the cerebral cortex. I. The albino rat: a topography ofthe cortical areas. J Comp Neurol 1946;84:221–75.
[31] Zilles K. Anatomy of the neocortex, cytoarchitecture and myeloarchitecture.In: Kolb B, Tess RC, editors. The cerebral cortex of the rat. Cambridge: MITPress; 1991.
[32] Paxinos G, Watson C. The rat brain in stereotaxiccoordinates. Sydney: Academic Press; 1997.
[33] Valle AC, Timo-Iaria C, Fraga JL, Sameshima K, Yamashita R. Theta waves andbehavioral manifestations of alertness and dreaming activity in the rat. Braz JMed Biol Res 1992;25:745–50.
[34] Valle AC. Análise eletrofisiológica do hipocampo durante o sonodessincronizado [thesis]. Federal University of São Paulo; 1995.
[35] Andersen ML, Valle AC, Timo-Iaria C, Tufik S. Implantação de elétrodos para oestudo eletrofisiológico do ciclo vigília-sono no rato. São Paulo: CLR BalieiroEditores Ltda; 2001. p. 43–59.
[36] Arida RM, Scorza FA, Peres CA, Cavalheiro EA. The course of untreated seizuresin the pilocarpine model of epilepsy. Epilepsy Res 1999;34:99–107.
[37] Goffin K, Nissinen J, Van Laere K, Pitkänen A. Cyclicity of spontaneousrecurrent seizures in pilocarpine model of temporal lobe epilepsy in rat. ExpNeurol 2007;205:501–5.
[38] Quigg M, Clayburn H, Straume M, Menaker M, Bertram 3rd EH. Effects ofcircadian regulation and rest–activity state on spontaneous seizures in a ratmodel of limbic epilepsy. Epilepsia 2000;41:502–29.
[39] Sanabria ERG, Scorza AS, Bortolotto ZA, Calderazzo-Filho LS, Cavalheiro EA.Disruption of light-induced c-Fos immunoreactivity in the suprachiasmaticnuclei of chronic epileptic rats. Neurosci Lett 1996;216:105–8.
[40] Covolan L, Mello LE. Temporal profile of neuronal injury following pilocarpineor kainic acid-induced status epilepticus. Epilepsy Res 2000;39:133–52.
[41] Mello LE, Covolan L. Spontaneous seizures preferentially injure interneurons inthe pilocarpine model of chronic spontaneous seizures. Epilepsy Res1996;26:123–9.
[42] Sanabria ER, Silva AV, Spreafico R, Cavalheiro EA. Damage, reorganization, andabnormal neocortical hyperexcitability in the pilocarpine model of temporallobe epilepsy. Epilepsia 2002;43(Suppl. 5):96–106.
[43] Vataev SI. The effects of repeated audiogenic convulsions on the organizationof sleep in rats. Neurosci Behav Physiol 2006;36:729–35.
[44] De Gennaro L, Ferrara M, Bertini M. The boundary between wakefulness andsleep: quantitative electroencephalographic changes during the sleep onsetperiod. Neuroscience 2001;107:1–11.
[45] Janz D. The grand mal epilepsies and the sleeping–waking cycle. Epilepsia1962;3:69–109.
[46] Shouse MN, Farber PR, Staba RJ. Physiological basis: how NREM sleepcomponents can promote and REM sleep components can suppress seizuredischarge propagation. Clin Neurophysiol 2000;111(Suppl. 2):9–18.
[47] Persinger MA, Koren SA. Persistent elevation of nocturnal activity in rodentsfollowing apparent recovery from lithium/pilocarpine-induced limbicseizures. Percept Mot Skills 1998;86:1243–8.
[48] Quigg M, Clayburn H, Straume M, Menaker M, Bertram 3rd EH. Hypothalamicneuronal loss and altered circadian rhythm of temperature in a rat model ofmesial temporal lobe epilepsy. Epilepsia 1999;40:1688–96.
[49] Baldy-Moulinier M. Temporal lobe epilepsy and sleep organization. In:Sterman MB, Passouant P, editors. Sleep and epilepsy. New York: AcademicPress; 1982. p. 347–59.
[50] Crespel A, Baldy-Moulinier M, Coubes P. The relationship between sleep andepilepsy in frontal and temporal lobe epilepsies: practical andphysiopathologic considerations. Epilepsia 1998;39:150–7.
[51] De Almeida CA, Lins OG, Lins SG, Laurentino S, Valença MM. Sleep disorders intemporal lobe epilepsy. Arq Neuropsiquiatr 2003;61:979–87.
[52] Terzano MG, Parrino L. Origin and significance of the cyclic alternating pattern(CAP). Sleep Med Rev 2000;4:101–23.
[53] Trentin MM, Pinho AP, da Costa JC. Epilepsias e Parassônias. J Epilepsy ClinNeurophysiol 2004;10(Suppl. 2):7–19.
rats following pilocarpine-induced temporal lobe epilepsy. Epilepsy Behav





























