Embed Size (px)
Citation preview

THE SPINY RAT PROECHIMYS GUYANNENSIS AS MODEL OFRESISTANCE TO EPILEPSY: CHEMICAL CHARACTERIZATION OF
HIPPOCAMPAL CELL POPULATIONS AND PILOCARPINE-INDUCEDCHANGES
P. F. FABENE,a L. CORREIA,a;b R. A. CARVALHO,b E. A. CAVALHEIROb andM. BENTIVOGLIOa*
aDepartment of Morphological and Biomedical Sciences, Faculty of Medicine, University of Verona, Strada Le Grazie 8,37134 Verona, Italy
bLaboratory of Experimental Neurology, Universidade Federal de Sa¬o Paulo/Escola Paulista de Medicina, Sa¬o Paulo, Brazil
AbstractöAt variance with pilocarpine-induced epilepsy in the laboratory rat, pilocarpine administration to the tropicalrodent Proechimys guyannensis (casiragua) elicited an acute seizure that did not develop in long-lasting status epilepticusand was not followed by spontaneous seizures up to 30 days, when the hippocampus was investigated in treated andcontrol animals. Nissl staining revealed in Proechimys a highly developed hippocampus, with thick hippocampal com-missures and continuity of the rostral dentate gyri at the midline. Immunohistochemistry was used to study calbindin,parvalbumin, calretinin, GABA, glutamic acid decarboxylase, and nitric oxide synthase expression. The latter was alsoinvestigated with NADPH-diaphorase histochemistry. Cell counts and densitometric evaluation with image analysis wereperformed. Di¡erences, such as low calbindin immunoreactivity con¢ned to some pyramidal cells, were found in thenormal Proechimys hippocampus compared to the laboratory rat. In pilocarpine-treated casiraguas, stereological cellcounts in Nissl-stained sections did not reveal signi¢cant neuronal loss in hippocampal sub¢elds, where the examinedmarkers exhibited instead striking changes. Calbindin was induced in pyramidal and granule cells and interneuronsubsets. The number of parvalbumin- or nitric oxide synthase-containing interneurons and their staining intensity weresigni¢cantly increased. Glutamic acid decarboxylase67-immunoreactive interneurons increased markedly in the hilus anddecreased in the CA1 pyramidal layer. The number and staining intensity of calretinin-immunoreactive pyramidal cellsand interneurons were signi¢cantly reduced.
These ¢ndings provide the ¢rst description of the Proechimys hippocampus and reveal marked long-term variations inprotein expression after an epileptic insult, which could re£ect adaptive changes in functional hippocampal circuitsimplicated in resistance to limbic epilepsy. ß 2001 IBRO. Published by Elsevier Science Ltd. All rights reserved.
Key words: epilepsy, calcium-binding protein, nitric oxide, GABA hippocampal interneuron, principal neuron.
Spiny rats of the genus Proechimys (family Echimydae,suborder Hystricomorpha) are mammals inhabiting theground of moist neotropical forests in Central andSouth America. These nocturnal terrestrial rodentshave a high population density in their habitat; theydo not dig burrows and move around rather sluggishlywithin a small home range (Emmons, 1982). Proechimysguyannensis, the common Brazilian name of which iscasiragua, received some attention as natural host ofinfectious agents (e.g. Everard and Tikasingh, 1973). Inaddition, this animal raised interest for its remarkable
genome size, which is the highest recorded so far inmammals and harbors several repetitive DNA families(Garagna et al., 1997). To our knowledge, however, theorganization of the casiragua brain has been hithertoneglected.
In the framework of comparative studies on animalmodels of experimental epilepsy, the susceptibility ofcasiragua to epileptogenic treatments was tested, andthe ¢ndings pointed out a striking resistance of theseanimals to several di¡erent paradigms of experimentalepilepsy (Carvalho, 1999).
The potential interest of Proechimys as animal modelof anti-convulsant mechanisms prompted the presentinvestigation of the hippocampus under normal condi-tions and after treatment with pilocarpine. This choliner-gic muscarinic agent was used in our investigation sincein the laboratory rat systemic injections of pilocarpineresult acutely in status epilepticus, which lasts up to24 h and is followed, after a seizure-free period ofapproximately 15 days, by a chronic phase characterizedby recurrent seizures with no spontaneous remission(Turski et al., 1983; Cavalheiro et al., 1991).
979
*Corresponding author. Tel. : +39-045-8027158; fax: +39-045-8027163.E-mail address: [email protected] (M. Bentivoglio).Abbreviations: CB, calbindin; CR, calretinin; DG, dentate gyrus;
EEG, electroencephalography; GAD, glutamic acid decarboxy-lase; ir, immunoreactive; NADPH-d, nicotinamide adenine dinu-cleotide phosphate-diaphorase; NGS, normal goat serum; NHS,normal horse serum; nNOS, neuronal nitric oxide synthase;NOS, nitric oxide synthase; OD, optical density; PB, phosphatebu¡er; PBS, phosphate-bu¡ered saline; PV, parvalbumin.
NSC 4983 5-7-01
www.elsevier.com/locate/neuroscience
Neuroscience Vol. 104, No. 4, pp. 979^1002, 2001ß 2001 IBRO. Published by Elsevier Science Ltd
Printed in Great Britain. All rights reservedPII: S 0 3 0 6 - 4 5 2 2 ( 0 1 ) 0 0 1 3 8 - 5 0306-4522 / 01 $20.00+0.00

For the investigation of pilocarpine-induced changes,we analyzed the electrographic and behavioral responseof casiraguas during the acute post-treatment phase, andthe behavior of these animals was then monitored for1 month, to ascertain whether they developed chronicseizures or were resistant to pilocarpine-induced epilepto-genesis. We then examined the hippocampus and pur-sued a chemical characterization of neurons in di¡erenthippocampal sub¢elds of the animals treated with pilo-carpine and control cases.
For the identi¢cation of cell subsets, we exploited theimmunoreactivity to parvalbumin (PV), calbindin D28k(CB) and calretinin (CR). These calcium-binding pro-teins represent e¡ective and sensitive markers of hip-pocampal cells (Celio, 1990) and, in particular, ofpopulations of inhibitory interneurons, extensively inves-tigated in the laboratory rat (see Freund and Buzsaki,1996 for review). Direct visualization of GABAergic ele-ments was based on antibodies that recognize the GABAsynthetic enzyme, glutamic acid decarboxylase (GAD),or GABA itself. We also examined the elements contain-ing nitric oxide synthase (NOS), the enzyme that synthe-sizes the gaseous free radical nitric oxide (NO), which isexpressed by GABAergic interneurons of the laboratoryrat hippocampus (Valtschano¡ et al., 1993). This part ofthe study was conducted with NADPH-diaphorase(NADPH-d) histochemistry, which reveals NOS activityin aldehyde-¢xed nervous tissue (Matsumoto et al.,1993), and with antibodies that recognize the neuronalNOS isoform (nNOS or NOS-I).
EXPERIMENTAL PROCEDURES
Animals
The study was based on adult male P. guyannensis (3^4months old; body weight: 210^240 g): four naive animals,eight animals treated with systemic administration of pilocar-pine, and ¢ve vehicle-injected control animals. Three pilocar-pine-treated animals were used for electroencephalography(EEG), and the study of the hippocampus was based on theother 14 animals (Table 1). The experiments were conductedfollowing the principles of the NIH Guide for the Use andCare of Laboratory Animals, and the Brazilian and EuropeanCommunities Council (86/609/EEC) directives. All e¡orts weremade to minimize the number of animals used and avoid anysu¡ering or stress.
The casiraguas were bred in a colony established at the Uni-versidade Federal de Sa¬o Paulo/Escola Paulista de Medicinafrom animals originally deriving from the Amazon basin.They were kept under controlled environmental parameters
and veterinarian control, and were habituated to the experi-menters for at least 2 weeks prior to all the procedures employedin the present study.
EEG recording, treatment and histology
Under pentobarbital anesthesia (50 mg/kg i.p.), bipolartwisted wire electrodes were implanted stereotaxically in thedorsal hippocampus of the animals destined for EEG recording,and ¢xed to the skull with dental acrylic cement. Surface recordswere led from jeweler screws positioned bilaterally over the sen-sorimotor cortex. The animals were allowed to recover fromsurgery for 1 week prior to recording and treatment.
Pilocarpine or vehicle injections were performed followingprotocols similar to those described in previous studies (Turskiet al., 1983, 1984; Cavalheiro et al., 1991; Obenaus et al., 1993).To minimize the peripheral e¡ects of pilocarpine, the cholinergicantagonist methyl-scopolamine (1 mg/kg, s.c. ; Sigma, St. Louis,MO, USA) was administered to all casiraguas. After 30 min theanimals were randomly divided in two groups: eight cases(including the three animals in which electrodes had beenimplanted) were injected i.p. with pilocarpine (300 mg/kg;Sigma) diluted in 0.1 M phosphate-bu¡ered saline (PBS),pH 7.4; the other ¢ve animals were injected i.p. with PBS ascontrol.
After EEG recording, the animals were again deeply anesthe-tized (Tionembutal, 50 mg/kg, i.p.), and perfused transcardiallywith PBS followed by 4% paraformaldehyde. The brains werecryoprotected and cut on a freezing microtome; sections werecollected and stained with Cresyl Violet to verify the electrodeplacement. These cases were not destined for further anatomicalinvestigation to avoid confounding factors represented by thedamage caused by the electrodes themselves. The EEG datawere compared with archival material of recordings obtainedfrom Wistar rats treated with the same paradigm.
Following a post-injection survival of 30 days, the ¢vevehicle-treated and ¢ve pilocarpine-treated animals, as well asfour untreated cases (Table 1), were anesthetized as indicatedabove and perfused through the heart with 50^100 ml of PBSfollowed by 250^300 ml of ¢xative solution: 4% paraformalde-hyde in 0.1 M phosphate bu¡er (PB), pH 7.4, was used in 13animals, whereas one of the naive animals was perfused with asolution of mixed aldehydes (2.5% glutaraldehyde and 0.5%paraformaldehyde in PB). The brains were dissected out andsoaked for cryoprotection in 30% sucrose in PBS until theysank. Sections were then cut with a freezing microtome at a40-Wm thickness throughout the rostrocaudal extent of the hip-pocampus. The sections were stored individually, and serieswere sampled collecting one every sixth section in sequentialorder starting at random from one of the ¢rst six sections. Inthis way each series of sections was collected allowing the di¡er-ent portions of the hippocampus to be sampled with the sameprobability (Coggeshall and Lekan, 1996). Sections derived frompilocarpine-treated animals were always processed for immuno-histochemistry or histochemistry together with those of matchedcontrol cases and in the same solutions. The sections were thenmounted on gelatinized slides, dehydrated, cleared in xylene andcoverslipped with Entellan (Merck, Darmstadt, Germany). In allcases, one series of sections was stained with Cresyl Violet.
Table 1. Immunohistochemical procedures and NADPH-d histochemistry utilized in PBS-treated control animals and pilocarpine-treatedanimals
No. cases Treatment (i.p.) PV CR CB NADPH-d NOS GAD67 GABA
4 ^ � � � �2 PBS � � � �1 PBS � � � � �1 PBS � �1 PBS � �5 pilocarpine � � � � �
NSC 4983 5-7-01
P. F. Fabene et al.980

Immunohistochemistry and NADPH-d histochemistry
Serial sections were processed free-£oating for immunohisto-chemistry or histochemistry (Table 1). Sections from the animalsperfused with 4% paraformaldehyde were processed for PV(n = 12), CR (n = 12), CB (n = 11), and GAD (n = 8) immunohis-tochemistry, or NADPH-d histochemistry (n = 13); in one casesome sections were processed for NOS immunohistochemistry.GABA immunohistochemistry was performed in the animal per-fused with mixed aldehydes, processing the adjacent sectionswith anti-GAD antibodies.
The following primary antibodies were used: monoclonalanti-PV antibodies (Sigma) diluted 1:3000, monoclonal anti-CB antibodies (Swant, Bellinzona, Switzerland) diluted 1:2000,polyclonal anti-CR antibodies raised in rabbit (Swant), diluted1:2500, anti-nNOS rabbit polyclonal antibodies (BoehringerMannheim, Indianapolis, IN, USA), diluted 1:1000; polyclonalrabbit anti-GABA antibodies (Chemicon International, Teme-cula, CA, USA) diluted 1:10 000; anti-GAD67 rabbit polyclonalantibodies (Chemicon), diluted 1:2000. The latter antibodieswere reported to immunolabel mainly neuronal perikarya inthe laboratory rat brain, at variance with anti-GAD65=67 anti-bodies which label both cell bodies and neuropil (Esclapez et al.,1993). However, in preliminary experiments, antibodies againstthe GAD67 isoform were found to be more e¡ective for bothperikarya and neuropil than anti-GAD65=67 antibodies in theProechimys brain (Fabene and Bentivoglio, unpublished obser-vations), and were therefore used for the purpose of the presentanalysis. The antibodies were diluted in 1% normal horse serum(NHS; for monoclonal), or 1% normal goat serum (NGS; forpolyclonal), 0.3% Triton X-100 and 0.1% NaN3 in PBS; anti-GABA and anti-GAD67 antibodies were diluted in 1% NGS inPBS omitting Triton X-100.
For immunohistochemical processing, the sections were pre-incubated in 5% NHS (for monoclonal antibodies), or 5% NGS(for polyclonal antibodies) in 0.3% Triton X-100 and 0.1%NaN3 in PBS for 1 h and then incubated in the primary anti-bodies for 24^48 h (monoclonal) or 48^72 h (polyclonal). Thesections were then incubated in biotinylated horse anti-mouse(for monoclonal) or goat anti-rabbit (for polyclonal) IgGs,diluted 1:200 in 1% NHS or NGS, 0.3% Triton X-100 and0.1% NaN3 in PBS. Finally, the sections were reacted in theavidin^biotin^peroxidase solution (ABC kit, Vectastain, VectorLabs, Burlingame, CA, USA), using 3,3P-diaminobenzidine(Sigma) as chromogen in the last step of the procedure. Controlsections were processed as above omitting each of the primaryantibodies, and no immunostaining was observed in this mate-rial.
The sections destined to NADPH-d histochemistry wererepeatedly washed with PB and then preincubated in 0.2% Tri-ton X-100 in PB. After 30 min they were incubated inL-NADPH (Sigma; 1 mg/ml), nitroblue tetrazolium (Sigma;0.5 mg/ml) and 0.2% Triton X-100 in PB at 37³C for 1^2 h.
Data analysis
After the systemic injections, the behavior of the animals wasobserved by two independent investigators over a period of 10 h.During the following month, the animals were monitored with avideo system (Arida et al., 1999).
The histological material was studied under the microscopewith bright-¢eld or dark-¢eld illumination. For quantitativeanalysis, the data derived from the pilocarpine-treated animalswere compared to those obtained from the matched PBS-treatedcases. Cell counts and densitometric evaluation of the labelingintensity were performed blindly as to the experimental groupassignment.
In order to assess the e¡ect of pilocarpine treatment on neuro-nal numbers in di¡erent sub¢elds of the hippocampus, stereo-logical cell counts were performed in four control and fourpilocarpine-treated casiraguas. Cells were counted in Cresyl Vio-let-stained sections, at intervals corresponding to every 18thsection through the entire extent of the dorsal hippocampus.The counts were made using a rectangular frame of 150U100Wm and a 40U objective, within two frames per hippocampalarea (a total of 20 frames per section), placed randomly in thestrata oriens, pyramidale and radiatum of CA1, CA2 and CA3,respectively, as well as in the polymorphic zone of the dentategyrus (DG), here referred to as hilus. Cells within each framewere counted using the `optical dissector' method (Coggeshalland Lekan, 1996), excluding those within the uppermost focalplane. Only neurons with evident nucleoli were included in thecounts, discarding glial cells identi¢ed as smaller cells withoutevident nucleoli and with intensely stained nuclei. The data werestatistically analyzed using one way analysis of variance, fol-lowed by the post-hoc Bonferroni test. Di¡erences were consid-ered statistically signi¢cant at P6 0.05.
The distribution of cell bodies positive to each marker wascharted from the three cases per group which provided the bestmaterial in terms of staining, in four sections per case regularlyspaced through the hippocampus and at corresponding antero-posterior levels, using a digital camera and Neurolucida soft-ware (Microbright¢eld Inc., Colchester, VT, USA).
The number of PV-, CR-, GAD-immunopositive or NADPH-d-stained cell bodies, as well as the perikaryal areas of some ofthe immunopositive neuronal populations, were evaluated inthree sections per animal at corresponding anteroposterior lev-els, and in four animals per group, using Neurolucida softwarefor image analysis. In addition, the number of GABA- or GAD-immunostained neurons was evaluated in three pairs of adjacentsections, regularly spaced through the dorsal hippocampus ofthe naive animal perfused with mixed aldehydes.
Densitometric analysis of the intensity of PV-, CR-immuno-stained, and NADPH-d-positive hippocampal cells was per-formed using for each marker three regularly spaced sectionsderived from the three control cases and three pilocarpine-treated ones used for the charting of labeled cells. In this part
Abbreviations used in the ¢gures
alv alveusAmy amygdaloid complexCA1, CA2, CA3 hippocampal ¢elds CA1, CA2, CA3cc corpus callosumccs splenium of the corpus callosumdf dorsal fornixDG dentate gyrusDGcr cresta of the DGDGlb, DG lateral bladeDGmb, DG medial bladeDGpo polymorphic layer of the DGdhc dorsal hippocampal commissureec external capsuleFC fasciola cinerea¢ ¢mbriahf hippocampal ¢ssure
mo stratum molecularerf rhinal ¢ssureSF septo¢mbrial nucleussg stratum granulareslm stratum lacunosum-moleculareslu stratum lucidumso stratum orienssp stratum pyramidalesps super¢cial pyramidal layersr stratum radiatumsub subiculumSUBd dorsal subiculumV3 third ventriclevhc ventral hippocampal commissureVL lateral ventricle
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 981

of the study, a total of 100 PV-immunoreactive (ir) neurons and100 NADPH-d-stained neurons were randomly sampled in eachgroup. Densitometric analysis of CR immunostaining was basedon a total of 400 cells (including non-pyramidal neurons
sampled randomly from the stratum lacunosum-moleculare ofCA1 and pyramidal cells sampled randomly in CA3). Densitom-etry was performed with a JVC CCD KY-F58 digital cameraand the image analysis software Image Pro Plus0 4.0 for Win-
Fig. 1. Electroencephalographic recordings from the hippocampus (HPC) and cerebral cortex (CX) illustrating the sequenceof alterations observed after the administration of pilocarpine (PILO) to Proechimys (right) compared with the ¢ndingsobserved under the same paradigm in the laboratory rat (left). The Wistar rat data, which derive from the laboratory archivalmaterial of the Department of Experimental Neurology of the UNIFESP-EPM (Sa¬o Paulo, Brazil) and illustrate ¢ndingsrepeatedly reported in the literature (see text for details), are shown here to clarify the di¡erences with the Amazon spiny ratin the acute response to pilocarpine administration. Note the occurrence of spikes in the hippocampal recordings 10 min afterpilocarpine injection, and the electrographic seizure at 25 min. Electrographic status epilepticus is observed at 1 h. Statusepilepticus persists 2 h later in the Wistar rat, whereas a progressive normalization of the EEG can be observed in Proe-chimys. At 6 and 24 h after pilocarpine injection, note the presence of spiking activity in the recordings from the Wistar rat,
whereas those obtained from Proechimys can be considered similar to the pre-treatment pattern.
NSC 4983 5-7-01
P. F. Fabene et al.982

dows (Media Cybernetics, Silver Spring, MD, USA), using forall measurements constant light illumination and a 25U objec-tive. In each section the zero value of optical density (OD) wasassigned to the background tissue. The mean OD value wasobtained in each group from the raw data of all measurements.
The statistical signi¢cance of the di¡erence between vehicle-
and pilocarpine-treated rats in the number of each population ofstained cells (mean total number of labeled neurons, or meannumber of neurons labeled in each hippocampal region), as wellas in the staining intensity (mean OD value) of each marker wasevaluated using the two-tailed Student's t-test, with signi¢canceat P6 0.05.
Fig. 2. Low power photomicrographs of Nissl-stained sections through the brain of P. guyannensis at the level of the dorsalhippocampus and diencephalon (A^C), and photomicrographs illustrating the features observed in the hippocampus at themidline (D^I): note the contiguity of the ventral laminae of the DG and the continuity of the subiculum. Scale bars = 0.2 mm
(A; also applies to B and C); 0.5 mm (D; also applies to E^I).
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 983

Fig. 3. Low-power views of the dorsal hippocampus of P. guyannensis, illustrating the pattern of immunohistochemical label-ing of CB (A, B), PV (C, D), CR (E, F) and of NADPH-d positivity (G, H) 1 month after treatment: a control animalinjected with PBS is shown in the left column, and one pilocarpine (Pilo)-treated case is shown in the right column. Theimages have been obtained with the technique of `histography' as used by Celio (1990), projecting the section on the slidesdirectly on photographic paper which is subsequently developed and ¢xed. The images are, therefore, a negative of the origi-
nal and the immunostained structures (brown in the original) appear white. Scale bar = 0.5 mm, H also applies to A^G.
NSC 4983 5-7-01
P. F. Fabene et al.984

Fig. 4 (Caption overleaf).
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 985

RESULTS
Behavioral and EEG observations after treatment
The EEG ¢ndings we observed in casiraguas are com-pared in Fig. 1 to those typically obtained in Wistar ratsand corresponding to those reported in previous studies(Turski et al., 1983, 1984; Cavalheiro et al., 1991; ). Inthe laboratory rat pilocarpine-induced acute seizuresare characterized by a burst of paroxysmal spiking activ-ity in the hippocampus, that spreads to cortical record-ings, and cortical paroxysmal activity is ¢rst observed30 min after the injection (Fig. 2). In P. guyannensisspreading to the cortex was instead observed almostimmediately, 10 min after pilocarpine injection (Fig. 1).During status epilepticus, high voltage spikes are thenobserved for several hours in Wistar rats, while theseelectrographic changes were documented in casiraguasonly in the ¢rst 3 h after pilocarpine administration(Fig. 1).
For the study of the behavioral response to pilocar-pine, the seizure severity was scored using the stages ofRacine's scale (Racine, 1972). As repeatedly reported inprevious studies (Turski et al., 1983, 1984; Cavalheiroet al., 1991), after pilocarpine injection Wistar ratsshow continuous mastication, salivation and clonicmovements of the vibrissae and limbs of increasingseverity. Wistar rats then rapidly reach stage 5, withtonic^clonic seizures and loss of balance, evolving intypical limbic status epilepticus that lasts several hours.From 15 to 30 min after pilocarpine injections, the casir-aguas exhibited behavioral abnormalities typical of stage2, with mastication and head nodding, that did notevolve in more severe seizures. The animals remainedinstead for 1^2 h with the four limbs on the £oor,extended ¢ngers and tonic extension of the tail, withoutclonic movements of head or limbs, and then recoveredto a normal behavior. None of these cases exhibitedbehavioral changes during the following month, untilperfusion. The PBS-treated casiraguas of the controlgroup did not show behavioral abnormalities after theinjections.
We also wish to mention that one additional casiraguainvestigated in the same session rapidly reached stage 5seizure and died a few minutes after the onset of tonic^clonic seizures; this animal was not included in thepresent series because it was not perfused. Another pilo-carpine-injected casiragua did not display any behavioralabnormality and was discarded.
General features of the hippocampal formation andanalysis of Cresyl Violet-stained sections
Under macroscopic inspection, the Proechimys brainexhibited a highly developed temporal region and adeep rhinal ¢ssure, which was very obvious also undermicroscopic examination (Fig. 2A^C).
In Nissl-stained sections, a large hippocampal forma-tion was observed in the Proechimys brain, together withrelatively large temporal lobe structures, such as theamygdaloid complex (Fig. 2A^C). Peculiar featureswere observed in the hippocampus at the midline,where both the dorsal and ventral hippocampal commis-sures were represented by thick bundles (Fig. 2D^I). Inparticular, the ventral hippocampal commissure paral-leled in size the corpus callosum (see, for example,Fig. 2D). At variance with the ¢ndings observed in thelaboratory rat (Paxinos and Watson, 1986; Swanson,1992), at rostral levels, the ventral granular layers ofthe DG of the right and left sides were continuous(Fig. 2E, F); close contiguity of the subicular region oftwo sides was observed proceeding posteriorly (Fig. 2F,G).
All the classical cytoarchitectonic subdivisions of thehippocampus were clearly identi¢ed in the Proechimys,and the laboratory rat atlas (Swanson, 1992) was used asreference. We include under the term hippocampus theCA1, CA2 and CA3 sub¢elds of the Ammon's horn andthe DG.
No obvious cytoarchitectural abnormalities were seenin the Nissl-stained sections of the hippocampus of pilo-carpine-treated casiraguas. In these cases, some pyknoticpyramidal cells were detected in the CA2 and CA3 sub-¢elds, but cell loss was not evident in any of the hippo-campal regions. In support of these observations, thestereological analysis did not reveal at the post-hoc com-parison any statistically signi¢cant di¡erence in the num-ber of neurons of the analyzed hippocampal layers andsub¢elds of the pilocarpine-treated animals versus thevehicle-treated ones.
In the sections processed for immunohistochemistry orhistochemistry, consistent ¢ndings were observed in thenaive and PBS-treated animals, that will be, therefore, allde¢ned as normal cases.
Calcium-binding proteins
Similarities, as well as major di¡erences with the pat-terns of immunopositivity to calcium-binding proteinsdescribed in the hippocampus of the laboratory rat
6Fig. 4. Photomicrographs showing the distribution of CB-immunopositive cells in the dorsal hippocampus of control (PBS)and pilocarpine (Pilo)-treated P. guyannensis. A and B illustrate the hippocampal midline and C represents at higher magni¢-cation the cluster of neurons indicated by the arrow in B. D and E illustrate the CA2 hippocampal sub¢eld and F illustratesat higher power the clusters of cells indicated by the arrow in E. G and H illustrate CB immunoreactivity in the DG and Jillustrates at higher magni¢cation part of the granular layer (the same point is marked by the stars in H and J as reference).Note at all these sites the marked induction of CB immunoreactivity after pilocarpine treatment. I illustrates at low powerCB-ir neurons in the entorhinal cortex in the normal animal: note the high number of intensely ir neurons, at variance withthe paucity of immunostained cell bodies detected in the hippocampus of the same animal (A, D, G). Scale bars = 250 Wm
(A; also applies to B, D, E, G, H, I); 75 Wm (C; also applies to F, J).
NSC 4983 5-7-01
P. F. Fabene et al.986

Fig. 5. Schematic representation of the distribution and density of CB-ir neurons in the Proechimys in four representativelevels of the hippocampus in rostrocaudal (A^D) order. In this ¢gure and in Figs. 7, 10 and 13 the section outlines are
derived from the atlas of the laboratory rat hippocampus (Swanson, 1992).
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 987

(Celio, 1990; Freund and Buzsaki, 1996), were observedin the normal Proechimys hippocampus. In addition,marked di¡erences were detected after pilocarpinetreatment. The features of immunolabeling were con-sistent in each group of normal or pilocarpine-treatedcasiraguas.
CB immunoreactivity
Normal animals. CB immunohistochemistry resultedin a very low signal in the normal Proechimys hippo-campus (Figs. 3A and 4A, D, G). Only a few immuno-positive pyramidal neurons, exhibiting a light staining,were detected. These CB-positive cells were sparsely dis-tributed in the pyramidal cell layer of CA1, CA2 andCA3 throughout the anteroposterior extent of theAmmon's horn (Fig. 5A^D).
The low CB immunoreactivity in the hippocampusunder basal conditions was not due to technical failuresince marked CB immunopositivity was observed inother brain regions. For example, intense CB immuno-staining was evident in non-pyramidal neurons scatteredthroughout the cerebral cortex (Fig. 4I) and CB-immuno-positive cells were very abundant in the thalamus.
Pilocarpine-treated animals. A striking increase of CBimmunoreactivity in both cell bodies and neuropil wasdetected after pilocarpine treatment in distinct regions ofthe hippocampus. CB induction in the hippocampal for-mation, in which CB immunoreactivity ¢lled the granulecell layer and the molecular layer of the entire DG, waswell evident even at low power observation (Fig. 3B).
Intensely CB-positive principal cells and interneuronswere observed in the sections derived from the pilocar-pine-treated animals. In particular, dark immunopositiv-ity was detected in non-pyramidal neurons scattered inthe strata oriens and pyramidale of CA1, and clusteredclose to the midline (Fig. 4B, C). CB-positive pyramidalneurons were also sparsely distributed in the CA3 pyra-midal layer (Fig. 5C). In addition, intensely CB-immu-nostained cells were seen in CA2, and were representedby pyramidal neurons clustered in the pyramidal layer
and by non-pyramidal cells of the stratum oriens, inwhich the perikaryal staining extended also to proximaldendrites (Figs. 4E, F and 5B^D). Marked CB immuno-positivity was seen in granule cells that ¢lled the entiregranular layer of the DG (Figs. 4H and 5B^D), andsome of these neurons were very darkly stained (Fig. 4L).
Due to the low CB immunoreactivity under basal con-ditions, the induction of this protein after pilocarpineadministration was very obvious. Therefore, the di¡er-ences of CB immunostaining in the control versus pilo-carpine-treated group did not require quantitativeanalysis.
PV immunoreactivity
Normal animals. At variance with CB, marked PVimmunopositivity was seen in the hippocampus of con-trol animals even at low power (Fig. 3C). Both neuronalperikarya and the neuropil were PV-immunolabeled(Fig. 6A, B, E). The neuropil immunostaining includedpunctate PV-ir elements, which were di¡usely distributedthroughout the hippocampus, and exhibited a distinctperisomatic distribution in the DG granular layer(Fig. 6B), and in the pyramidal layer of CA3 (Fig. 6E)and CA1 (Fig. 6G).
All the PV-ir cell bodies exhibited the features of non-pyramidal neurons (Fig. 6B, E, G), and in some of thesecells the immunopositivity extended in the proximal por-tions of the dendritic arbor. The PV-immunostainedperikarya were especially numerous in the alveusthroughout the extent of the hippocampus (Figs. 6Aand 7; Table 2). A relatively high density of PV-ir cellbodies was also observed in the strata oriens and pyra-midale of CA1 and CA3 of the dorsal hippocampus, aswell as in all sub¢elds of the ventral hippocampus,whereas PV-stained perikarya were rarely found in thestratum lacunosum-moleculare (Figs. 6A and 7; Table 2).PV-ir cells were also sparsely distributed in the lateraland medial blades of the DG and in the hilus (Fig. 7C).
The PV-ir neurons displayed a great variety of shapesand sizes: large and round immunostained perikarya(mean area: 321.47 Wm2) were detected in the stratum
Table 2. Mean number þ S.D. of PV- and CR-ir neurons
Hippocampal sub¢elds P. guyannensis : control (PBS) versus pilocarpine-treated (pilo)
PV (PBS) PV (pilo) CR (PBS) CR (pilo)
Dorsal hippocampus CA1 alv 62.6 þ 6.4 128.4 þ 44.5so 25.6 þ 6.1 81.7 þ 40.8sp 47.3 þ 11.9 65.7 þ 15.8sr 13.6 þ 5.7 22.7 þ 7.6slm 2 þ 1.7 1.6 þ 1.5 297.3 þ 68.13 168.3 þ 39.5
CA2 sp 10 þ 4.5 13.7 þ 5.5CA3 so 24.3 þ 3.2 52.3 þ 16
sp 15.3 þ 3.1*** 30 þ 2.6 73.6 þ 4.04*** 11.6 þ 3.5sr 5.7 þ 2.5 19 þ 9.1
dentate gyrus DGmo 1.7 þ 1.5* 7.3 þ 2.5DGsg 2.3 þ 0.5 4 þ 3.6DGpo 9.3 þ 0.6* 67.3 þ 18.8 121 þ 40.5* 16 þ 2
*P6 0.05, **P6 0.01, ***P6 0.001 (see text for individual P values). alv, alveus; so, stratum oriens; sp, stratum pyramidale ; sr, stratumradiatum; slm, stratum lacunosum-moleculare; DGmo, stratum moleculare of the DG; DGsg, stratum granulare of the DG; DGpo, poly-morphic layer of the DG.
NSC 4983 5-7-01
P. F. Fabene et al.988

Fig. 6. Photomicrographs illustrating PV immunoreactivity in control (PBS) and pilocarpine (PILO)-treated casiraguas in thehippocampus and DG at low power (A, C), in CA3 (E, F), and CA1 (G, H). B and D represent at higher magni¢cationparts of the ¢elds shown in A and C, respectively (the arrows point to the same cells as reference). Note at all these sites theincrease in the density of PV-ir neurons and immunostaining intensity after pilocarpine treatment (C, D, F, H) in respect to
controls (A, B, E, G). Scale bars = 250 Wm (A, C); 50 Wm (B, D); 200 Wm (E^H).
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 989

Fig. 7. Schematic representation of the distribution and density of PV-ir neurons in the four selected representative sectionsof the hippocampus.
NSC 4983 5-7-01
P. F. Fabene et al.990
radiatum of CA3, while the alveus contained smallerPV-ir cells (mean area: 285.49 Wm2).
Pilocarpine-treated animals. PV immunoreactivitywas intense also in the pilocarpine-treated animals(Fig. 3D), and an increase in the density of stained neu-rons compared to controls was evident (Fig. 6C, D, F,H). In addition, PV immunostaining of hippocampalneuronal cell bodies and neuropil appeared more intensethan in normal cases (Fig. 6D, F).
The study of the distribution of PV-ir neurons con-¢rmed that immunostained cells had increased after pilo-carpine treatment. The immunostained cells were verynumerous in the alveus (Figs. 6C, H and 7), in the DG
granular layer and in the hilus (Figs. 6C, D and 7B^D).In addition, clusters of PV-ir neurons were found alongthe midline (Figs. 6C and 7B, C), and the density of theimmunopositive cells as well as neuropil immunostainingwere high in the strata oriens and pyramidale of CA3and CA1 (Figs. 6F, H and 7).
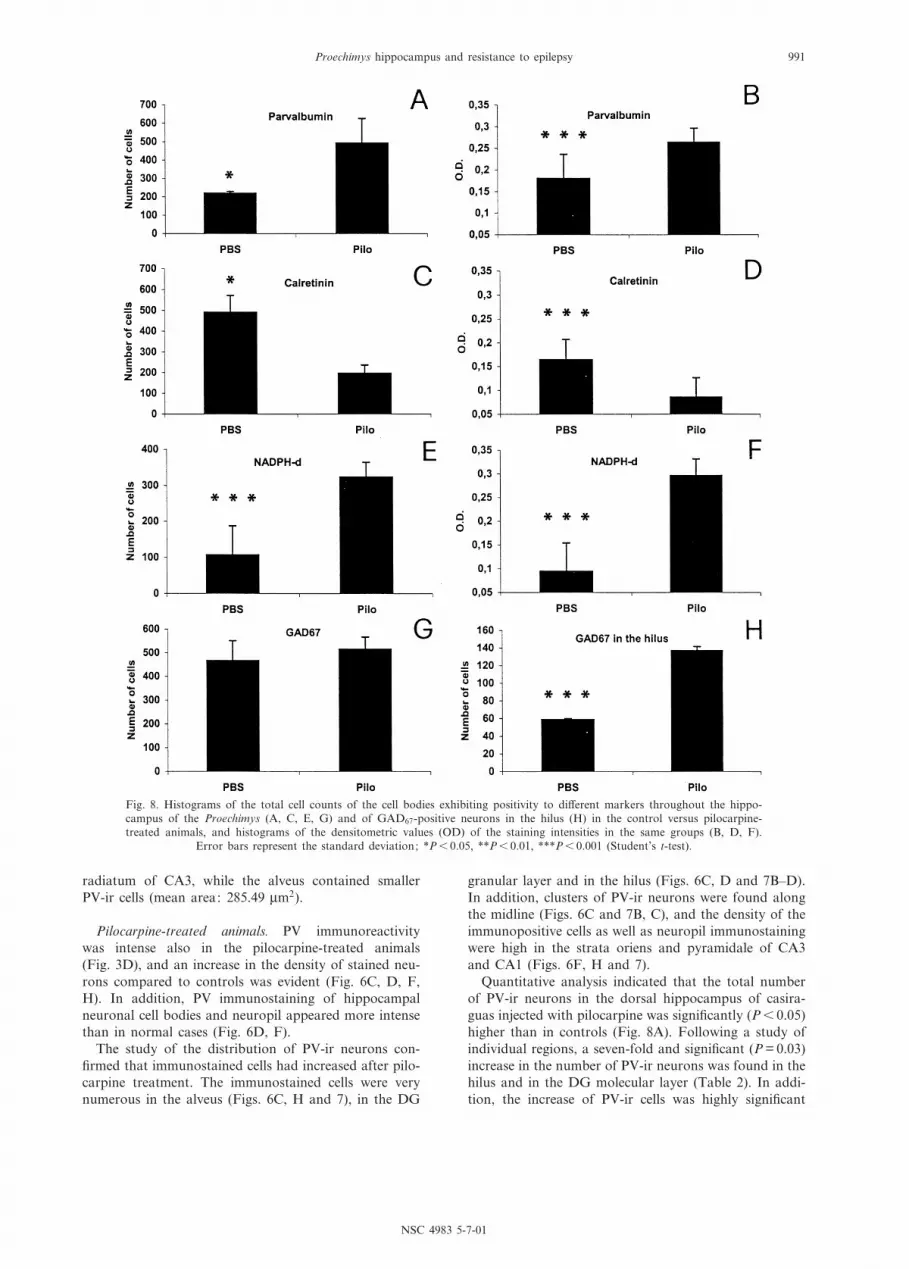
Quantitative analysis indicated that the total numberof PV-ir neurons in the dorsal hippocampus of casira-guas injected with pilocarpine was signi¢cantly (P6 0.05)higher than in controls (Fig. 8A). Following a study ofindividual regions, a seven-fold and signi¢cant (P = 0.03)increase in the number of PV-ir neurons was found in thehilus and in the DG molecular layer (Table 2). In addi-tion, the increase of PV-ir cells was highly signi¢cant
Fig. 8. Histograms of the total cell counts of the cell bodies exhibiting positivity to di¡erent markers throughout the hippo-campus of the Proechimys (A, C, E, G) and of GAD67-positive neurons in the hilus (H) in the control versus pilocarpine-treated animals, and histograms of the densitometric values (OD) of the staining intensities in the same groups (B, D, F).
Error bars represent the standard deviation; *P6 0.05, **P6 0.01, ***P6 0.001 (Student's t-test).
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 991
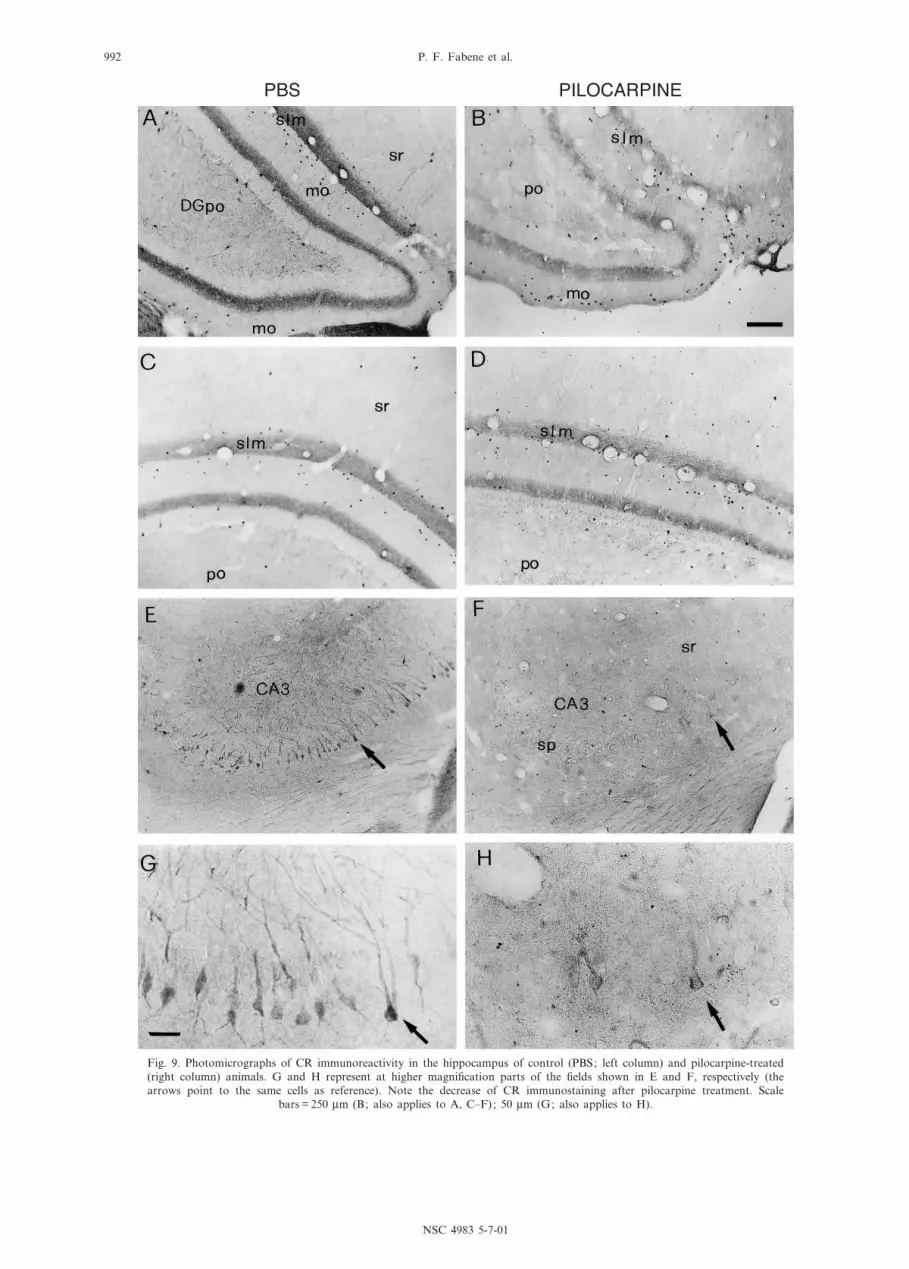
Fig. 9. Photomicrographs of CR immunoreactivity in the hippocampus of control (PBS; left column) and pilocarpine-treated(right column) animals. G and H represent at higher magni¢cation parts of the ¢elds shown in E and F, respectively (thearrows point to the same cells as reference). Note the decrease of CR immunostaining after pilocarpine treatment. Scale
bars = 250 Wm (B; also applies to A, C^F); 50 Wm (G; also applies to H).
NSC 4983 5-7-01
P. F. Fabene et al.992

Fig. 10. Schematic representation of the distribution and density of CR-ir neurons in the four selected representative sectionsof the hippocampus.
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 993
(P = 0.0005) in the CA3 pyramidal layer (Table 2). Thenumber of PV-labeled neurons was at least twice thatseen in controls also in the alveus, as well as in the strataoriens and radiatum of CA1 and CA3 but, probably dueto the high standard deviation, this di¡erence failed toreach a statistical signi¢cance (Table 2).
The densitometric evaluation showed that the intensityof PV immunostaining in neurons was highly signi¢-cantly increased (P6 0.000003) in the pilocarpine-treatedcases in respect to the control ones (Fig. 8B).
CR immunoreactivity
Normal animals. CR immunohistochemistry resultedin intense staining in the normal Proechimys hippocam-
pus, with a distinct regional prevalence. In particular,neuropil immunopositivity clearly delineated the DGand the stratum lacunosum-moleculare also at lowpower observation (Figs. 3E and 9A). Both principalneurons and interneurons exhibited CR immunostaining.(Fig. 9A, C, E). Numerous CR-ir pyramidal cells (meanarea: 310.35 Wm2) were seen in CA3 (Figs. 9E and 10B,C) and the immunopositivity extended along the den-dritic arborizations of these neurons (Fig. 9G). A fewpyramidal cells were CR-immunolabeled also in CA2(Fig. 10B, C). CR-ir non-pyramidal cells were seen inthe stratum radiatum of CA1, and were very numerousin the stratum lacunosum-moleculare (Fig. 10). Theseneurons were small-sized (mean area: 45.79 Wm2) withintensely immunostained perikarya but a limited labeling
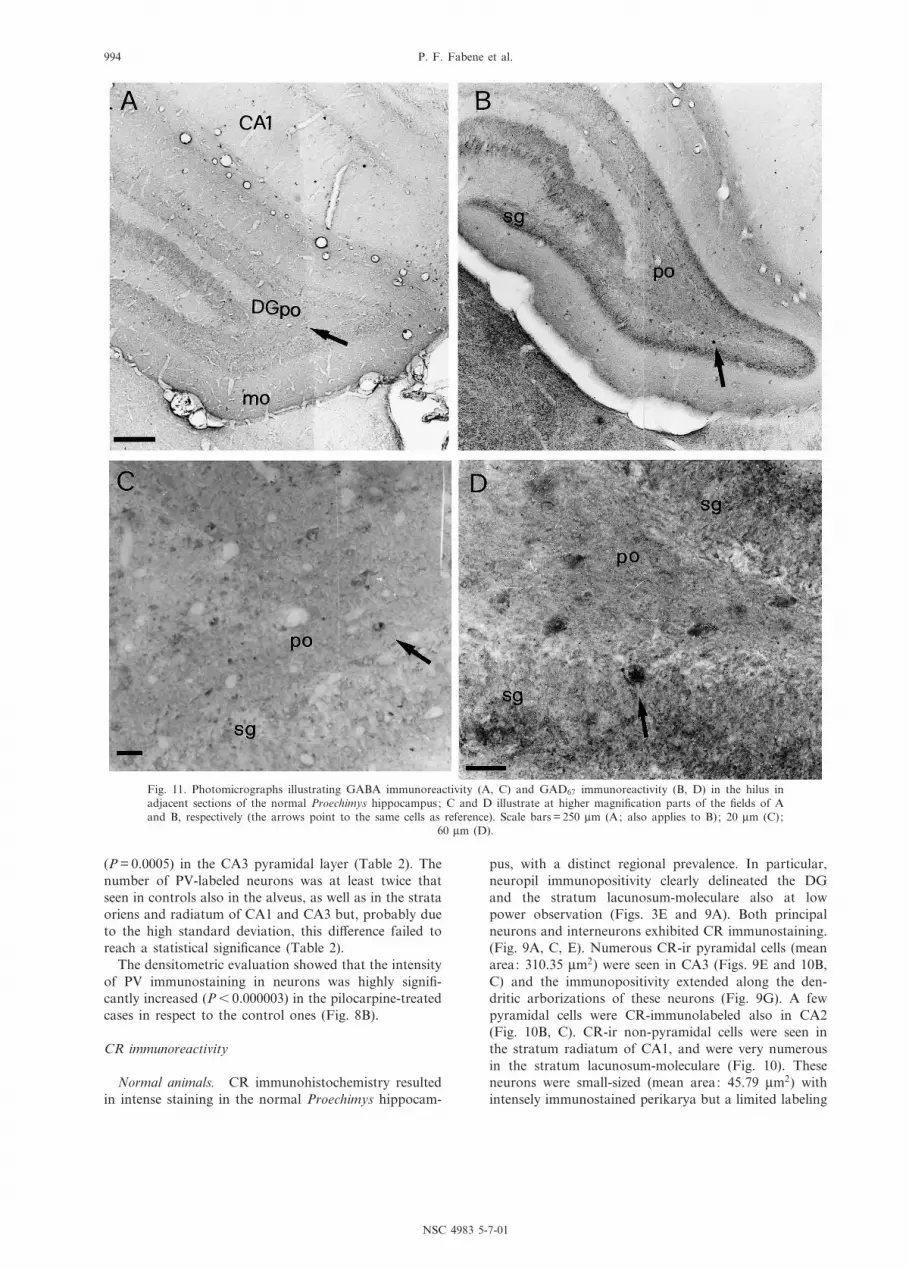
Fig. 11. Photomicrographs illustrating GABA immunoreactivity (A, C) and GAD67 immunoreactivity (B, D) in the hilus inadjacent sections of the normal Proechimys hippocampus; C and D illustrate at higher magni¢cation parts of the ¢elds of Aand B, respectively (the arrows point to the same cells as reference). Scale bars = 250 Wm (A; also applies to B); 20 Wm (C);
60 Wm (D).
NSC 4983 5-7-01
P. F. Fabene et al.994

of proximal dendrites. CR-positive interneurons werealso found in the polymorphic and molecular layers ofthe DG medial blade and in the molecular layer of theDG lateral blade (Fig. 10C, D). In addition, CR-positivecells were sparsely distributed in the stratum oriens of allhippocampal sub¢elds, and in the stratum radiatum ofCA1 and CA3 (Fig. 10). Besides the stratum lacunosum-moleculare, the neuropil immunopositivity outlined aband in the inner molecular layer (Fig. 9A).
Pilocarpine-treated animals. CR protein expression inthe hippocampus exhibited high sensitivity to pilocarpinetreatment that resulted in an overall decrease of CRimmunostaining (Figs. 3F and 9B). In particular, CRimmunopositivity was preserved only in a few pyramidalcells of CA3 (Figs. 9F, H and 10).
The charting of the distribution of labeled cells showeda marked decrease of CR-immunopositive interneuronsin the stratum lacunosum-moleculare, especially in theCA1 and CA3 sub¢elds of the dorsal hippocampus,and in the DG (Fig. 10).
The loss of CR immunoreactivity, both in terms ofnumber of positive cells and staining intensity, was con-¢rmed by the quantitative analysis. The total number ofCR-ir cells in the hippocampus was more than two-foldlower in the pilocarpine-treated cases (196 þ 40.7) than inthe control ones (492 þ 80.7), and such di¡erence wassigni¢cant (P6 0.02) (Fig. 8C). When individual regionswere considered, the decrease of CR-ir neurons wasfound to be signi¢cant (P6 0.05) in the hilus, and highlysigni¢cant (P = 0.00008) in the CA3 pyramidal layer; inboth these regions the number of CR-immunostainedcells was approximately seven times lower in the pilocar-pine-treated animals than in controls (Table 2).
The densitometric analysis showed that the intensity ofCR immunoreactivity in neurons was highly signi¢cantlydecreased (P6 0.0001) in the pilocarpine-treated animalsin respect to the control ones (Fig. 8B).
GABA and GAD immunoreactivities
Normal animals. GABA-ir cell bodies were abundant
throughout the Proechimys hippocampus. The immuno-reactivity was mostly con¢ned to cell bodies (Fig. 11A,B), and extended in some neurons along the proximaldendritic stem. GABA-ir perikarya exhibited consistentlya non-pyramidal shape and a great variety of sizes. Inparticular, numerous GABA-ir cells were found in thealveus, and in the strata lacunosum-moleculare and radi-atum of CA1 and CA3. The GABA-ir cells in CA3 wereamong the largest of those stained in the hippocampus.In addition, GABA-ir neurons were distributed through-out the DG.
GABA-immunostained puncta were very abundant,and displayed a perisomatic distribution in the pyramidallayer of CA1, CA2 and CA3 and in the DG granularlayer. A GABAergic network of preterminal and termi-nal elements was also observed around the hippocampalcommissures and in the subicular area.
As mentioned above, in our material GAD67 immuno-reactivity was found to stain both cell bodies and ¢bers,and neuropil labeling was especially evident in the DG,where the granular layer exhibited intense immunoposi-tivity (Fig. 11B, D). In general, due to the marked neuro-pil staining the signal-to-noise ratio of perikaryallabeling was lower than in the GABA-immunoreactedmaterial. However, the quantitative comparison betweenGABA- and GAD-immunostained neurons revealed sim-ilar numbers of immunopositive neurons in hippocampalregions, and the statistical evaluation did not show sig-ni¢cant di¡erences between GABA-ir and GAD-ir cellsin normal animals (Table 3).
Pilocarpine-treated animals. GABA immunohisto-chemistry was not performed in tissue obtained fromanimals injected with pilocarpine. In the GAD67-immuno-processed sections, no major qualitative di¡erences wereseen in the features of staining of cell bodies and neuro-pil after pilocarpine treatment in respect to the controlcases. It should be mentioned, however, that the qualityof the immunohistochemical preparations was not opti-mal, and quantitative analysis of the immunostainingintensity was, therefore, not performed.
Cell counts were instead pursued. No signi¢cant di¡er-
Table 3. Mean number þ S.D. of GAD-ir and NADPH-d-stained neurons
Hippocampal sub¢elds P. guyannensis : control (PBS) versus pilocarpine-treated (pilo)
GABA(untreated)
GAD67
(PBS)GAD67
(pilo)NADPH-d(PBS)
NADPH-d(pilo)
Dorsal hippocampus CA1 alv 30.7 þ 11.7 31.3 þ 12.7 54.7 þ 9.1 18 þ 4.4 21 þ 1.7so 78.3 þ 10.9 94.7 þ 15 95.3 þ 23.1 12 þ 3 17.7 þ 3.2sp 37.3 þ 5.9 32.3 þ 4.9** 9.7 þ 5.9 9 þ 1** 27.3 þ 3.2sr 58.7 þ 6.5 45.7 þ 13.1 32 þ 4 12 þ 2* 23.3 þ 3.8slm 46.3 þ 9.4 47.7 þ 10.7 55.3 þ 2.1 11.3 þ 4.9* 60 þ 10.5
CA2 sp 5.7 þ 1.5 10 þ 4 12 þ 1 0 þ 0* 8.7 þ 2.3CA3 so 58 þ 16.8 62 þ 11.5 53.3 þ 13.1 8 þ 3.5 12.7 þ 6.8
sp 19.7 þ 2.5 17.3 þ 2.1 22.3 þ 5.5 10.7 þ 0.6* 24 þ 1sr 56 þ 27.9 51.3 þ 26 39.3 þ 2.5 11 þ 4.4 16.3 þ 6.1
dentate gyrus DGmo 5.3 þ 7.6 7.3 þ 4.1 1.3 þ 1.1 0 þ 0** 19.3 þ 3.5DGsg 6 þ 3.6 8 þ 4.4 3.7 þ 1.1 1.7 þ 1.5* 18.3 þ 7.5DGpo 57 þ 4.4 59 þ 1*** 137 þ 4.6 13 þ 5.6* 74.7 þ 13.8
*P6 0.05, **P6 0.01, ***P6 0.001 (see text for individual P values).
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 995

ence (P = 0.42) was found in the total number of GAD67-ir cells in the hippocampus of casiraguas injected withpilocarpine (516 þ 51.2) versus that found in controls(466.7 þ 85.2) (Fig. 8G), but intergroup regional di¡er-ences were observed (Table 3). In the hilus, the numberof GAD67-ir neurons increased dramatically and signi¢-cantly (P6 0.001) after pilocarpine injection in respect tocontrols (Fig. 8H, Table 3). In the CA1 pyramidal layer,the number of GAD67-ir cells was instead about threetimes lower after pilocarpine injection (Table 3), andthis di¡erence was signi¢cant (P6 0.01). In the alveus,the number of GAD67-ir neurons was increased in thepilocarpine-treated cases in respect to controls but, prob-ably due to the high standard deviation, this di¡erencedid not reach a statistical signi¢cance.
NADPH-d positivity and NOS immunoreactivity
Normal animals. In the casiragua hippocampusNADPH-d staining was seen in both cell bodies andneuropil (Figs. 3G and 12A), but was lighter than inother brain areas, the description of which is, however,beyond the scope of the present study. NADPH-d-stained cells were sparsely distributed throughout thehippocampal regions (Figs. 12A and 13). The histochem-ical positivity ¢lled non-pyramidal perikarya and exhib-
ited a distinct perisomatic distribution in the neuropilof the granular layer, outlining an intensely stainedband also in the inner molecular layer of the DG(Fig. 12A, C).
In the study of sections processed for nNOS immuno-histochemistry, a general correspondence was foundin the distribution of NADPH-d staining and NOSimmunopositivity. Quantitative analysis of the twomarkers was not performed since the staining protocolshad not been applied to a series of adjacent sections.
Pilocarpine-treated animals. In the observation of theoverall pattern of NADPH-d staining, the main di¡er-ence between the control and pilocarpine-treated caseswas represented by enhancement of the histochemicalpositivity in the latter animals, especially in CA3(Fig. 3H). The charting of the distribution of stainedcells pointed out a marked increase of these neuronsthroughout the hippocampal formation, and especiallyin the strata oriens, pyramidale and lacunosum-molecu-lare of CA1 (Fig. 13B, C). The increase was also evidentin the DG and marked in the hilus (Fig. 13, Table 3),where intensely stained cells were very numerous(Fig. 12B, D).
Quantitative evaluation indicated that the total num-ber of NADPH-d-stained neurons in the hippocampus
Fig. 12. The photomicrographs illustrate NADPH-d histochemical positivity in the hippocampus of control (left) and pilocar-pine-treated (right) animals. C and D represent at higher magni¢cation parts of the ¢elds shown in A and B, respectively (thearrows point to the same cells as reference). Note the quantitative increase of positive neurons and staining intensity in thehilus after pilocarpine treatment and the marked perisomatic distribution of stained puncta in the stratum granulare. Scale
bars = 250 Wm (A; also applies to B); 85 Wm (C; also applies to D).
NSC 4983 5-7-01
P. F. Fabene et al.996

Fig. 13. Schematic representation of the distribution and density of NADPH-d-positive neurons in the four selected represen-tative sections of the hippocampus.
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 997

(Fig. 8E) was signi¢cantly (P6 0.001) increased(323.3 þ 9.1 in the pilocarpine-treated animals versus106.7 þ 10.3 in controls). Regional di¡erences were alsofound (Table 3). In particular, in the CA1 pyramidallayer the number of positive neurons was about threetimes higher after pilocarpine injection than in controls,and the di¡erence was highly signi¢cant (P6 0.005). Atwo-fold and highly signi¢cant (P = 0.004) increase ofstained neurons was found in the stratum radiatum ofCA1 after pilocarpine injection. The increase of positivecells after pilocarpine treatment was also signi¢cant inthe stratum lacunosum-moleculare (P6 0.01) and in thepyramidal layer (P = 0.002) of CA3. Marked di¡erenceswere detected in the DG. Thus, stained neurons were notevident in the DG molecular layer of control cases,whereas they were consistently observed in the pilocar-pine-treated animals. In the DG granular layer, stainedcells were very few in control cases but signi¢cantly(P = 0.04) more numerous after pilocarpine injection; asigni¢cant (P = 0.02) increase of about six times wasfound in the hilus of the pilocarpine-treated cases.
DISCUSSION
The hippocampus of a tropical rodent, the spiny ratP. guyannensis, was here investigated for the ¢rst time,focusing on the chemical features of neuronal popula-tions in the normal adult brain and 1 month after aseizure episode provoked by pilocarpine administration.Such response exhibited marked di¡erences in respect tothe model of temporal lobe epilepsy typically induced bypilocarpine in the laboratory rat (Cavalheiro, 1994). Inparticular, in the animals we investigated the acute seiz-ure elicited by pilocarpine did not evolve in long-lastingstatus epilepticus and was not followed by the onset ofspontaneous recurrent seizures. These ¢ndings are inagreement with those obtained in a large series of experi-ments in Proechimys that included a follow-up of2 months after pilocarpine injections (Carvalho, 1999).In this previous investigation casiraguas were found tobe resistant also to kindling and to seizures induced bythe systemic administration of kainic acid (Carvalho,1999). These ¢ndings argue against the possibility thatthe resistance to pilocarpine-induced status epilepticuscould be due to a low sensitivity of Proechimys to thepharmacological action of this cholinergic agent.
Our survey of hippocampal cell subsets exploited thechemical characterization of neurons as a tool to revealthe occurrence of long-term changes elicited by the epi-leptic insult. Although this approach does not fullyunravel the organization of the casiragua hippocampus,several interesting cytological aspects were observed. It isalso worth noting that the hippocampal commissuralsystem, which exhibits species di¡erences in mammals(Van Groen and Wyss, 1988), was found to be highlydeveloped in Proechimys. Thus, the relative sizes of thehippocampal commissures and corpus callosum suggestthat a crosstalk between limbic structures could play acrucial role in the interhemispheric communication of theProechimys brain. In this respect, it is interesting to recall
that hippocampal commissural neurons have been impli-cated in direct and feed-forward inhibitory mechanismsand in the termination of epileptic afterdischarge (Braginet al., 1997).
Chemically characterized cell populations in theProechimys hippocampus
Features conserved in di¡erent mammalian species,such as PV expression in interneurons, and variationacross species of CB, PV or CR expression in neuronalsubsets, were documented in the hippocampus byprevious studies (see, for example, Seress et al., 1991;Murakawa and Kosaka, 1999). On line with these inves-tigations, the present study pointed out similarities anddistinctive features of hippocampal cell populations ofthe spiny rat Proechimys.
In particular, in the laboratory rat CB is expressed byneurons in all layers and sub¢elds of the hippocampusand DG (Celio, 1990; Toth and Freund, 1992), repre-sented by granule cells, interneurons of CA1 and CA3,and a subset of pyramidal neurons located in the innerhalf of CA1 and CA2. In the normal Proechimys hippo-campus, CB immunoreactivity was instead strikinglylow, and was only detected in pyramidal neuronssparsely distributed in the Ammon's horn.
In contrast, in the laboratory rat PV is expressed ininterneurons comprising basket cells and axo-axoniccells, distributed through the strata oriens and radiatumof various hippocampal regions, as well as in the gran-ular and molecular layers of the DG and in the dentatehilus (Kosaka et al., 1987; Celio, 1990). PV was found tobe expressed in interneurons also in the Proechimys hip-pocampus, and these cells were widely distributed andprevailed in the strata oriens and pyramidale. At var-iance with the laboratory rat, in Proechimys numerousPV-ir neurons were also seen in the alveus.
In Wistar rats, small-sized CR-positive cells, mostlylocated between the pyramidal layer and the stratumradiatum, were described in CA1, CA2 and CA3; it isnotable that in all mammalian species examined so far,CR was reported to be expressed in non-principal cells ofthe hippocampus, and in a subset of mossy hilar cellsand granule cells of some species, whereas CR-positivehippocampal pyramidal cells were not described in pre-vious studies (Gulyas et al., 1992; Miettinen et al., 1992;Resibois and Roger, 1992; Seress et al., 1993; Nitsch andOhm, 1995; Murakawa and Kosaka, 1999). In Pro-echimys, the CR-immunopositive cells included insteadinterneurons located in the stratum lacunosum-molecu-lare of CA1 and in the dentate hilus, as well as pyramidalcells of the CA3 sub¢eld. It has been reported that CR-containing excitatory a¡erents deriving from supramam-millary neurons regulate the activity of granule cells(Magloczki et al., 1994; Kiss et al., 2000) and, on thebasis of the present observations, this input could alsoplay a role in the Proechimys hippocampus.
In the laboratory rat, NADPH-d-positive cells are rep-resented by GABAergic interneurons sparsely distributedin several ¢elds of the Ammon's horn and in the DG(Valtschano¡ et al., 1993), and concentrated in the den-
NSC 4983 5-7-01
P. F. Fabene et al.998

tate hilus, as well as in the stratum radiatum of CA3(Miettinen et al., 1995; Kotti et al., 1997). Similar fea-tures of distribution were seen in the Proechimys hippo-campus under basal conditions, but in this speciesNADPH-d-stained cells populated also the alveus.
The understanding of the signi¢cance of such neuro-chemical features would require a detailed identi¢cationof the di¡erent hippocampal cell types, and of the inter-cellular synaptic relationships at the electron microscopiclevel. However, several lines of evidence have indicatedthat most, if not all, hippocampal interneurons areGABAergic despite their heterogeneity (Freund andBuzsaki, 1996). Studies on the co-localization of theexamined markers with GAD were not performed inour investigation since, in preliminary experimentsbased on GAD67 and PV immuno£uorescence, theGAD signal in the Proechimys hippocampal cells wasfound to be too low to detect unequivocally double-immunolabeled neurons (unpublished observations).However, the occurrence and distribution of GABAergicinterneurons directly visualized by GAD67 and GABAindicated that GABAergic inhibition is a basic principleof organization also in the hippocampus of this species.
Changes in hippocampal cell populations of theProechimys after pilocarpine-induced acute seizures
Cell loss in the hippocampus has been repeatedlyreported in the laboratory rat after pilocarpine-inducedstatus epilepticus. These degenerative changes a¡ect espe-cially CA3 and CA1, and profoundly involve the hilus,as demonstrated by quantitative stereological analyses,that showed marked neuronal loss in these regions1 month after pilocarpine-induced status epilepticus(Mello et al., 1993; Liu et al., 1994). In the presentstudy, cell counts did not show in casiragua a signi¢cantloss of neurons in hippocampal sub¢elds and hilus1 month after the acute seizure elicited by pilocarpine.Pyknotic neurons, indicative of neurodegenerativeevents, were observed in the pyramidal layer of CA2and CA3, and paralleled a decrease of CR immunoreac-tivity at the same locations. Altogether the data suggest aselective vulnerability of pyramidal cell subsets of thecasiragua hippocampus to the seizure episode, but indi-cate that the neuropathological sequelae of the acutepilocarpine-induced seizures did not result in signi¢cantloss of hippocampal cells. However, the response of theanimals to pilocarpine did not evolve in limbic statusepilepticus, which represents a major pathogenetic factorof neuron damage in the hippocampus.
The most striking cytological consequences of theacute seizure were represented in Proechimys by markedvariations in the phenotype of hippocampal neurons.These changes involved induction and/or up-regulationof CB and PV, alterations in the number of GAD-pos-itive interneurons, and down-regulation of CR in bothinterneurons and principal cells. Such long-term conse-quences of epileptic injury were especially marked inCA1, CA3 and in the hilus, i.e. the regions which, asmentioned above, are most susceptible to pilocarpine-induced seizure damage, and are also a¡ected in
human cases of temporal lobe epilepsy (Meldrum,1993; Proper et al., 2000).
The present data deserve special interest in view ofthe debated issue of the susceptibility to epilepsy ofhippocampal cells, concerning in particular the expres-sion of proteins that can bu¡er intracellular calcium. Apositive correlation was found between the neuronal CBor PV content and the relative resistance of hippocampalneuronal subsets to seizure activity (Sloviter, 1989;Sloviter et al., 1991). The signi¢cance of this ¢ndingwas recently questioned, since targeted deletion of cal-cium-binding proteins in genetically manipulated micewas not found to play a role in neurodegenerative pro-cesses triggered by kainate-induced seizures (Bouilleret etal., 2000).
The present observation that calcium-binding proteinexpression undergoes changes after exposure to a con-vulsant challenge does not necessarily implicate a criticalrole of calcium bu¡ering in these events. Our data showinstead that PV, CB and CR can reveal changes in thecell protein content in response to stimuli that alter theneuron excitability. Therefore, increased or decreasedprotein immunolabeling could re£ect variations in func-tional networks, potentially counteracting perturbationsin calcium homeostasis and/or other factors of cellimbalance.
In particular, in the casiraguas injected with pilocar-pine the number of PV-ir interneurons increased in thealveus, in the dentate hilus and throughout the Ammon'shorn. In a previous study, profound loss of PV interneur-ons was found in kainate-treated rats that exhibitedmarkedly hyperexcitable ¢eld-potential response in theDG to perforant path stimulation, indicating that theseinterneurons could play a major role in the inhibition ofgranule cell activity (Buckmaster and Dudek, 1997). PV-containing interneurons were found instead to be selec-tively vulnerable in a di¡erent model of experimentalseizures (Suzukawa et al., 1999). The present data sup-port a role of interneurons that express PV, as well as ofPV induction in interneurons, in counteracting epilepto-genic events.
We also demonstrated that CB expression was dramat-ically induced in the casiragua hippocampus 1 monthafter pilocarpine administration, when intense CB immu-nostaining was detected in interneurons, CA2 pyramidalcells and granule cells of the entire DG. This pattern ofdistribution is similar to that observed in the laboratoryrat under basal conditions (Celio, 1990; Toth andFreund, 1992), indicating that CB gene expression ispart of the repertoire of hippocampal neuronal subsets,in which CB protein content can be regulated by func-tional demands.
Experiments comparing the damage observed in CA1neurons and granule cells in CB-knockout mice andwild-type littermates after kainate-induced seizures haverecently involved CB in the vulnerability of CA1 cells toseizure-induced injury but not in the resistance of dentategranule cells (Gary et al., 2000). It has been reported thatseveral weeks after kainate-induced seizures CB immuno-reactivity was decreased in the CA1/CA2 pyramidal cellsand lost in granule cells (Yang et al., 1997). In the
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 999

human epileptic DG, CB-containing interneurons werefound to be preserved, and exhibited marked hypertro-phy and dendritic changes, whereas dentate granule cellslost their CB content (Magloczki et al., 2000). Infusionof recombinant CB in tissue derived from human epilep-tic hippocampus indicated that CB can regulate Ca2�
entry in neurons by controlling the Ca2�-dependent inac-tivation of voltage-gated Ca2� channels (Na«gerl et al.,2000).
It is also interesting to note that in our study CB andPV induction in hippocampal neuronal subsets was par-alleled by a down-regulation of CR immunoreactivity ininterneurons, in CA3 pyramidal cells, as well as in thehilus. In the hippocampus of the laboratory rat CR isexpressed in interneurons specialized in the control ofother interneurons (Gulyas et al., 1996). Such synapticrelationships remain to be veri¢ed in Proechimys, but thepresent data suggest that a decreased inhibition of CR-containing interneurons could ultimately result inincreased activity of dentate granule cells. In contrast,the present ¢nding of a selective decrease of CR-ir neu-rons and CR protein expression in the hippocampus ofepilepsy-resistant animals, and especially in the hilus,indicates that CR-containing hippocampal cell subsetsare involved in the rearrangements that defend thebrain against a convulsant challenge.
Contrasting reports have appeared on the selectivedamage or resistance of CR-positive cells in the epileptichuman hippocampus. Thus, in the hippocampus of tem-poral lobe epilepsy patients with Ammon's horn sclero-sis, sparing of CR neurons (Blu«mcke et al., 1996), orsigni¢cant loss of these cells (Magloczki et al., 2000)were described. CR-containing excitatory a¡erents togranule cells deriving from supramammillary neurons(Magloczki et al., 1994; Kiss et al., 2000) were alsofound to be involved in this rearrangement and woulddeserve further investigation in the Proechimys hippo-campus.
Striking regional variations were observed in Pro-echimys in the distribution of GAD67-ir hippocampalneurons, which increased markedly in the hilus anddecreased in the pyramidal layer of CA1 after pilocar-pine injections. In the laboratory rat, selective loss ofhilar neurons expressing GAD mRNA was reported upto 2 months of pilocarpine-induced seizures (Obenaus etal., 1993). However, GAD65 and GAD67 isoforms werefound to be up-regulated at the gene and protein levels inthe remaining GABAergic neurons of the hippocampalformation, pointing out a considerable plasticity of thiscell subset (Esclapez and Houser, 1999). In the laborato-ry rat, adaptive changes of the GABAergic system to anepileptic insult also involve acutely granule cells, inwhich GAD67 mRNA induction (Schwarzer and Sperk,1995) and transient GABA immunoreactivity (Makiuraet al., 1999) were reported after limbic seizures provokedby kainic acid. However, acute changes were not exam-ined in Proechimys in the present investigation, and stud-ies based on in situ hybridization could provide furtherinformation on variations induced by epileptic injury in
GAD-containing hippocampal cells of this animal spe-cies.
Marked induction of NADPH-d positivity wasdetected in the casiragua hippocampal neurons, espe-cially those located in the alveus and dentate hilus,after exposure to the epileptic insult. In Wistar rats,NADPH-d-positive cells, especially those located in thehilus and CA1, exhibit vulnerability to status epilepticusinduced by the systemic administration of kainic acid(Miettinen et al., 1995; Kotti et al., 1997). A signi¢cantloss of NADPH-d-stained hippocampal neurons, and inparticular in the hilus, was also reported in pilocarpine-induced chronic epilepsy in Wistar rats (Hamani et al.,1999). The present data favor a potential role of NOS-containing hippocampal neurons in anti-epileptic rear-rangements, as supported by ¢ndings indicating thatthe free radical NO is implicated in anti-convulsantmechanisms (Przegalinski et al., 1994; Jayakumar etal., 1999). It should also be considered that NOSinduction in neurons has been repeatedly reported afterdi¡erent kinds of injuries, and the neurotoxic or neuro-protective signi¢cance of this phenomenon is stilldebated (Wu, 2000). The present investigation suggestsnot only that NOS was induced in hippocampal cell sub-sets in response to a functional challenge, but also thatthese NOS-containing neurons could be involved in pro-tective plasticity phenomena.
Finally, it is worth emphasizing that NOS-positivecells increased in the hilus of pilocarpine-treated casesin parallel with an increase of neurons expressing GADand PV, suggesting that they could represent, at least inpart, the same cell populations. The changes in the hilusare of special interest in view of the crucial role playedby hilar cells in the modulation of the activity of dentategranule cells.
Concluding remarks
Our study showed that in a natural animal model ofresistance to epilepsy both inhibitory and excitatory hip-pocampal cell subsets undergo marked plastic changesafter an epileptic insult. In particular, pilocarpine admin-istration elicited in casiragua ictal activity followed bylong-term variations of protein expression in subsets ofinterneurons and principal cells. Thus, the present ¢nd-ings indicate that a complex rearrangement in both theinhibitory regulation and excitatory activity of the Pro-echimys hippocampus occurs in response to a convulsantagent. These variations could be implicated in mecha-nisms of resistance to epilepsy and protection of the hip-pocampus against an epileptogenic injury.
AcknowledgementsöThis work was supported in part by grantsof the CNR, MURST and CNPq. The authors are very gratefulto Dr. T. Freund and Dr. Zs. Magloczky for their critical read-ing of the manuscript. The authors also wish to thank Dr. A.Vercelli for making image analysis facilities available in the ini-tial steps of this study and Dr. E. Tongiorgi for his help in thestereological analysis.
NSC 4983 5-7-01
P. F. Fabene et al.1000

REFERENCES
Arida, R.M., Scorza, F.A., de Araujo Peres, C., Cavalheiro, E.A., 1999. The course of untreated seizures in the pilocarpine model of epilepsy.Epilepsy Res. 34, 99^107.
Blu«mcke, I., Beck, H., Nitsch, R., Eickho¡, C., Che¥er, B., Celio, M.R., Schramm, J., Elger, C.E., Wolf, H.K., Wiestler, O.D., 1996. Preservationof calretinin-immunoreactive neurons in the hippocampus of epilepsy patients with Ammon's horn sclerosis. J. Neuropathol. Exp. Neurol. 55,329^341.
Bouilleret, V., Schwaller, B., Schurmans, S., Celio, M.R., Fritschy, J.M., 2000. Neurodegenerative and morphogenic changes in a mouse model oftemporal lobe epilepsy do not depend on the expression of the calcium-binding proteins parvalbumin, calbindin, or calretinin. Neuroscience 97,47^58.
Bragin, A., Penttonen, M., Buzsaki, G., 1997. Termination of epileptic afterdischarge in the hippocampus. J. Neurosci. 17, 2567^2579.Buckmaster, P.S., Dudek, F.E., 1997. Neuron loss, granule cell axon reorganization, and functional changes in the dentate gyrus of epileptic
kainate-treated rats. J. Comp. Neurol. 385, 385^404.Carvalho, R.A., 1999. Caracter|©zac°a¬o de modelos de epilepsia do lobo temporal em Proechimys guyannensis. Ph.D. Thesis, Universidade Federal
de Sa¬o Paulo, Brazil.Cavalheiro, E.A., 1994. Status epilepticus and secondary epileptogenesis in the pilocarpine model of epilepsy. In: Wolf, P. (Ed.), Epileptic Seizures
and Syndromes. Libbey and Co., London, UK, pp. 523^532.Cavalheiro, E.A., Leite, J.P., Bortolotto, Z.A., Turski, W.A., Ikonomidou, C., Turski, L., 1991. Long-term e¡ects of pilocarpine in rats: structural
damage of the brain triggers kindling and spontaneous recurrent seizures. Epilepsia 32, 778^782.Celio, M.R., 1990. Calbindin D28k and parvalbumin in the rat nervous system. Neuroscience 35, 375^475.Coggeshall, R.E., Lekan, H.A., 1996. Methods for determining numbers of cells and synapses: a case for more uniform standards of review.
J. Comp. Neurol. 364, 6^15.Emmons, L.H., 1982. Ecology of Proechimys (Rodentia, Echimyidae) in South-Eastern Peru. Trop. Ecol. 23, 280^290.Esclapez, M., Houser, C., 1999. Up-regulation of GAD65 and GAD67 in remaining hippocampal GABA neurons in a model of temporal lobe
epilepsy. J. Comp. Neurol. 412, 488^505.Esclapez, M., Tillarakatne, N.J.K., Tobin, A.J., Houser, C.R., 1993. Comparative localization of mRNAs encoding two forms of glutamic acid
decarboxylase with nonradioactive in situ hybridization methods. J. Comp. Neurol. 331, 339^362.Everard, C.O.R., Tikasingh, E.S., 1973. Ecology of the rodents, Proechimys guyannensis trinitatis and Oryzomys capito velutinus. Trinidad J.
Mammal. 54, 875^886.Freund, T.F., Buzsaki, G., 1996. Interneurons of the hippocampus. Hippocampus 6, 345^470.Garagna, S., Perez-Zapata, A., Zuccotti, M., Mascheretti, S., Marziliano, N., Redi, C.A., Aguilera, M., Capanna, E., 1997. Genome composition
in Venezuelan spiny-rats of the genus Proechimys (Rodentia, Echimyidae). I. Genome size, C-heterochromatin and repetitive DNAs in situhybridization patterns. Cytogenet. Cell Genet. 78, 36^43.
Gary, D.S., Sooy, K., Chan, S.L., Christakos, S., Mattson, M.P., 2000. Concentration- and cell type-speci¢c e¡ects of calbindin D28k onvulnerability of hippocampal neurons to seizure-induced injury. Mol. Brain Res. 75, 89^95.
Gulyas, A.I., Miettinen, R., Jacobowitz, D.M., Freund, T.F., 1992. Calretinin is present in non-pyramidal cells of the rat hippocampus. I. A newtype of neuron speci¢cally associated with the mossy ¢ber system. Neuroscience 48, 1^27.
Gulyas, A.I., Hajos, N., Freund, T.F., 1996. Interneurons containing calretinin are specialized to control other interneurons in the rat hippo-campus. J. Neurosci. 16, 3397^3411.
Hamani, C., Tenorio, F., Mendez-Otero, R., Mello, L.E.A.M., 1999. Loss of NADPH-diaphorase-positive neurons in the hippocampal formationof chronic pilocarpine-epileptic rats. Hippocampus 9, 303^313.
Jayakumar, A.R., Sujatha, R., Paul, V., Puviarasan, K., Jayakumar, R., 1999. Involvement of nitric oxide and nitric oxide synthase activity inanticonvulsive action. Brain Res. Bull. 48, 387^394.
Kiss, J., Csaki, A., Bokor, H., Shanabrough, M., Leranth, C., 2000. The supramammillo-hippocampal and supramammillo-septal glutamatergic/aspartatergic projections in the rat: a combined [3H]D-aspartate autoradiograhic and immunohistochemical study. Neuroscience 97, 657^669.
Kosaka, Y., Katsumaru, H., Hama, K., Wu, J.-Y., Heizmann, C.W., 1987. GABAergic neurons containing the Ca2�-binding protein parvalbuminin the rat hippocampus and dentate gyrus. Brain Res. 419, 119^130.
Kotti, T., Halonen, T., Sirvio« , J., Riekkinen, P., Miettinen, R., 1997. Comparison of NADPH diaphorase histochemistry, somatostatin immu-nohistochemistry, and silver impregnation in detecting structural and functional impairment in experimental status epilepticus. Neuroscience80, 105^117.
Liu, Z., Nagao, T., Desjardins, G.C., Gloor, P., Avoli, M., 1994. Quantitative evaluation of neuronal loss in the dorsal hippocampus in rats withlong-term pilocarpine seizures. Epilepsy Res. 17, 237^247.
Magloczki, Zs., Acsadi, L., Freund, T.F., 1994. Principal cells are the postsynaptic targets of supramammillary a¡erents in the hippocampus of therat. Hippocampus 4, 322^334.
Magloczki, Zs., Wittner, L., Borhegyi, Zs., Halasz, P., Vajda, J., Czirjak, S., Freund, T.F., 2000. Changes in the distribution and connectivity ofinterneurons in the epileptic human dentate gyrus. Neuroscience 96, 7^25.
Makiura, Y., Suzuki, F., Chevalier, E., Onteniente, B., 1999. Excitatory granule cells of the dentate gyrus exhibit a double inhibitory neuro-chemical content after intrahippocampal administration of kainate in adult mice. Exp. Neurol. 159, 73^83.
Matsumoto, T., Nakane, M., Pollock, J.S., Kuk, J.E., Forstermann, U., 1993. A correlation between soluble brain nitric oxide synthase andNADPH-diaphorase activity is only seen after exposure of the tissue to ¢xative. Neurosci. Lett. 155, 61^64.
Meldrum, B.S., 1993. Excitotoxicity and selective neuronal loss in epilepsy. Brain Pathol. 3, 405^412.Mello, L.E.A.M., Cavalheiro, E.A., Tan, A.M., Kupfer, W.R., Pretorius, J.K., Babb, T.L., Finch, D.M., 1993. Circuit mechanisms of seizures in
the pilocarpine model of chronic epilepsy: cell loss and mossy ¢ber sprouting. Epilepsia 34, 985^995.Miettinen, R., Gulyas, A.I., Baimbridge, K.G., Jacobowitz, D.M., Freund, T.F., 1992. Calretinin is present in non-pyramidal cells of the rat
hippocampus. II. Co-existence with other calcium binding proteins and GABA. Neuroscience 48, 29^43.Miettinen, R., Kotti, T., Halonen, T., Riekkinen, P.Sr., 1995. NADPH diaphorase-containing nonpyramidal cells in the rat hippocampus exhibit
di¡erential sensitivity to kainic acid. Eur. J. Neurosci. 7, 1822^1825.Murakawa, R., Kosaka, T., 1999. Diversity of calretinin immunoreactivity in the dentate gyrus of gerbils, hamsters, guinea pigs, and laboratory
shrews. J. Comp. Neurol. 411, 413^430.Na«gerl, U.V., Mody, I., Jeub, M., Lie, A.A., Elger, C.E., Beck, H., 2000. Surviving granule cells of the sclerotic human hippocampus have reduced
Ca2� in£ux because of a loss of calbindin-D28k in temporal lobe epilepsy. J. Neurosci. 20, 1831^1836.Nitsch, R., Ohm, T.G., 1995. Calretinin immunoreactive structures in the human hippocampal formation. J. Comp. Neurol. 360, 475^487.Obenaus, A., Esclapez, M., Houser, C.R., 1993. Loss of glutamate decarboxylase mRNA-containing neurons in the rat dentate gyrus following
pilocarpine-induced seizures. J. Neurosci. 13, 4470^4485.
NSC 4983 5-7-01
Proechimys hippocampus and resistance to epilepsy 1001

Paxinos, G., Watson, C., 1986. The Rat Brain in Stereotaxic Coordinates, 2nd edn. Academic Press, Sydney.Proper, E.A., Oestreicher, A.B., Jansen, G.H., Veelen, C.W.M., van Rijen, P.C., Gispen, W.H., de Graan, P.N.E., 2000. Immunohistochemical
characterization of mossy ¢ber sprouting in the hippocampus of patients with pharmaco-resistant temporal lobe epilepsy. Brain 123, 19^30.Przegalinski, E., Baran, L., Siwanowicz, J., 1994. The role of nitric oxide in the kainate-induced seizures in mice. Neurosci. Lett. 170, 74^76.Racine, R.J., 1972. Modi¢cation of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 32, 281^
284.Resibois, A., Roger, J.H., 1992. Calretinin in rat brain: an immunohistochemical study. Neuroscience 46, 101^134.Schwarzer, C., Sperk, G., 1995. Hippocampal granule cells express glutamic acid decarboxylase-67 after limbic seizures in the rat. Neuroscience 69,
705^709.Seress, L., Gulyas, A.I., Freund, T.F., 1991. Parvalbumin- and calbindin D28k-immunoreactive neurons in the hippocampal formation of the
macaque monkey. J. Comp. Neurol. 313, 162^177.Seress, L., Nitsch, R., Leranth, C., 1993. Calretinin immunoreactivity in the monkey hippocampal formation. I. Light and electron microscopic
characteristics and co-localization with other calcium-binding proteins. Neuroscience 55, 775^796.Sloviter, S.S., 1989. Calcium-binding protein (calbindin-D28k) and parvalbumin immunocytochemistry : localization in the rat hippocampus with
speci¢c reference to the selective vulnerability of hippocampal neurons to seizure activity. J. Comp. Neurol. 280, 183^196.Sloviter, S.S., Sollas, A.L., Barbaro, N.M., Laxer, K.D., 1991. Calcium-binding protein (calbindin-D28K) and parvalbumin immunocytochemistry
in the normal and epileptic human hippocampus. J. Comp. Neurol. 308, 381^396.Suzukawa, J., Omori, K., Okugawa, G., Fujiseki, Y., Heizmann, C.W., Inagaki, C., 1999. Long-lasting c-fos and NGF mRNA expressions and
loss of perikaryal parvalbumin immunoreactivity in the development of epileptogenesis after ethacrynic acid-induced seizure. Brain Res. 834,89^102.
Swanson, L.W., 1992. Brain Maps: Structure of the Rat Brain. Elsevier, Amsterdam.Toth, K., Freund, T.F., 1992. Calbindin D28K-containing nonpyramidal cells in the rat hippocampus: their immunoreactivity for GABA and
projection to the medial septum. Neuroscience 49, 793^805.Turski, W.A., Cavalheiro, E.A., Schwartz, M., Czuczvar, S.J., Kleinrok, Z., Turski, L., 1983. Limbic seizures produced by pilocarpine in rats:
behavioural, electroencephalographic and neuropathological study. Behav. Brain Res. 9, 315^335.Turski, W.A., Cavalheiro, E.A., Bortolotto, Z.R., Mello, L.M., Schwartz, M., Turski, L., 1984. Seizures produced by pilocarpine in mice:
a behavioral, electroencephalographic and morphological analysis. Brain Res. 478, 385^390.Valtschano¡, J.G., Weinberg, R.J., Kharazia, V.N., Nakane, M., Schmidt, H.H.H.W., 1993. Neurons in rat hippocampus that synthesize nitric
oxide. J. Comp. Neurol. 331, 111^121.Van Groen, T., Wyss, J.M., 1988. Species di¡erences in hippocampal commissural connections: studies in rat, guinea pig, rabbit, and cat. J. Comp.
Neurol. 267, 322^334.Wu, W., 2000. Response of nitric oxide synthase to neuronal injury. In: Steinbusch, H.W.M., De Vente, J., Vincent, S.R. (Eds.), Functional
Neuroanatomy of the Nitric Oxide System Handbook of Chemical Neuroanatomy, vol. 17. Elsevier, Amsterdam, pp. 315^353.Yang, Q., Wang, S., Hamberger, A., Celio, M.R., Haglid, K.G., 1997. Delayed decrease of calbindin immunoreactivity in the granule cell-mossy
¢bers after kainic acid-induced seizures. Brain Res. Bull. 43, 551^559.
(Accepted 21 March 2001)
NSC 4983 5-7-01
P. F. Fabene et al.1002


























![Intermolecular interactions between cucurbit[7]uril and pilocarpine](https://img.pdfslide.net/doc/110x75/63595009ca12adf96502f793/intermolecular-interactions-between-cucurbit7uril-and-pilocarpine.jpg)


