Embed Size (px)
Citation preview
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/7199644
Thetickprotectiveantigen,4D8,isaconservedproteininvolvedinmodulationoftickbloodingestionandreproduction
ARTICLEinVACCINE·JUNE2006
ImpactFactor:3.62·DOI:10.1016/j.vaccine.2006.02.046·Source:PubMed
CITATIONS
97
READS
30
8AUTHORS,INCLUDING:
JosedelaFuente
UniversityofCastilla-LaMancha
499PUBLICATIONS11,860CITATIONS
SEEPROFILE
ConsueloAlmazán
AutonomousUniversityofTamaulipas
61PUBLICATIONS2,369CITATIONS
SEEPROFILE
AtilioJoséMangold
InstitutoNacionaldeTecnologíaAgropecu…
189PUBLICATIONS2,885CITATIONS
SEEPROFILE
ChristianGortázar
UniversityofCastilla-LaMancha
352PUBLICATIONS7,312CITATIONS
SEEPROFILE
Allin-textreferencesunderlinedinbluearelinkedtopublicationsonResearchGate,
lettingyouaccessandreadthemimmediately.
Availablefrom:AtilioJoséMangold
Retrievedon:03February2016

Vaccine 24 (2006) 4082–4095
The tick protective antigen, 4D8, is a conserved protein involvedin modulation of tick blood ingestion and reproduction�
Jose de la Fuente a,b,∗, Consuelo Almazan a, Uriel Blas-Machado c, Victoria Naranjo b,Atilio J. Mangold d, Edmour F. Blouin a, Christian Gortazar b, Katherine M. Kocan a
a Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, United Statesb Instituto de Investigacion en Recursos Cinegeticos IREC (CSIC-UCLM-JCCM), Ronda de Toledo s/n, 13071 Ciudad Real, Spainc Diagnostic Laboratory, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, United States
d Instituto Nacional de Tecnologıa Agropecuaria, Estacion Experimental Agropecuaria Rafaela, CC 22, CP2300 Rafaela, Santa Fe, Argentina
Received 24 December 2005; received in revised form 15 February 2006; accepted 21 February 2006Available online 9 March 2006
Abstract
s6wwp(©
K
1
amt
ccag
a
HO
d
0d
The gene that encodes the tick protective antigen, 4D8, was cloned from 10 species belonging to 6 genera, and the nucleotide and amino acidequences were analyzed. 4D8 nucleotide and protein sequences were conserved among these tick species with identity/similarity between5–98 and 60–98%, respectively. The function of 4D8 was characterized by RNA interference (RNAi) in five tick species. After the ticksere allowed to feed, degeneration of gut, salivary glands and reproductive tissues was observed, and tick survival, weight and ovipositionere significantly reduced. 4D8 RNAi effected >90% reduction in oviposition in all tick species tested. Because of the critical role that 4D8lays during tick feeding and oviposition, which ultimately results in the reduction of tick progeny, we proposed the generic name “subolesin”Latin, suboles: offspring, progeny) for tick 4D8 proteins and subA for the subolesin-encoding gene.
2006 Elsevier Ltd. All rights reserved.
eywords: Tick vaccine; Evolution; RNA interference; Reproduction; Subolesin
. Introduction
Ticks are ectoparasites of wild and domestic animalsnd humans, and are considered to be second worldwide toosquitoes as vectors of human diseases and the most impor-
ant vector of pathogens in North America [1].Control of tick infestations has been difficult, and suc-
essful tick control has been limited [2]. Recent efforts toontrol tick infestations have focused on integrated pest man-gement in which different control methods are adapted in aeographic area against one tick species with consideration
� The GenBank accession numbers for sequences of tick 4D8 cDNA clonesre AY652654, AY652657, DQ159961–DQ159972.∗ Corresponding author at: Veterinary Pathobiology, Center for Veterinaryealth Sciences, 250 McElroy Hall, Oklahoma State University, Stillwater,K 74078, United States. Tel.: +1 405 744 0372; fax: +1 405 744 5275.
E-mail addresses: jose [email protected],[email protected] (J. de la Fuente).
to their environmental effects (reviewed by [3]). The recentdevelopment of vaccines against Boophilus spp. has providednew possibilities for identification of protective antigens foruse in vaccines for control of tick infestations (reviewed by[4,5]). Control of ticks by vaccination would avoid environ-mental contamination and the selection of drug resistant ticksthat result from repeated acaricide applications. Tick vaccinescould also be designed to include multiple tick and pathogenantigens that may target a broad range of both tick speciesand tick-borne pathogens (reviewed by [4]). Tick vaccinesdecrease tick populations and could affect the transmissionof tick-borne pathogens by decreasing the vector capacityof ticks and interfering with the development of pathogens(reviewed by [4]).
The discovery of tick protective antigens has been thelimiting step in the development of tick vaccines. Althoughseveral tick proteins have been proposed as putative protec-tive antigens, few have been tested in vaccination experi-
264-410X/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2006.02.046

J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095 4083
ments as recombinant proteins (reviewed by [4,5]). Amongthem, Boophilus microplus Bm86 and Bm95 gut proteins ofunknown function [6–8], Bm91 peptidase [9], and BmT1trypsin inhibitor [10]; Rhipicephalus appendiculatus 64Pcement protein [11] and immunoglobulin-binding proteins[12]; Haemaphysalis longicornis p29 salivary gland puta-tive extracellular matrix protein [13], HL34 of unknownfunction [14], serine protease inhibitor serpin-2 [15] andP27/30 troponin I-like protein [16]; Ixodes scapularis 4D8and 4E6 of unknown function and 4F8 nucleotidase [17–20]and Amblyomma hebraeum AhEF engorgement factor [21]have affected tick infestations. The results of these studieshave demonstrated the feasibility of controlling tick infesta-tions by use of multiple gene products that target various tickphysiological processes.
The tick protective antigen, 4D8, was discovered bycDNA expression library immunization (ELI) and analysisof expressed sequenced tags (EST) in a mouse model of tickinfestations for identification of cDNAs protective againstI. scapularis [17,18]. The gene and protein sequences werefound to be conserved in invertebrates and vertebrates and thegene was expressed in all tick developmental stages and inadult tissues of several tick species, suggesting a conservedfunction for 4D8 [19]. Immunization with recombinant I.scapularis 4D8 reduced larval, nymphal and adult tick infes-tations, supporting the use of 4D8 for development of tickv
toip
fbmm
2
2
gSnstraMeTar
ratory (OSU) or at the “Finca Galiana” (University of Castilla,La Mancha, Spain) with the approval and supervision of Insti-tutional Animal Care and Use Committees. Off-host tickswere maintained in a 12 h light:12 h dark photoperiod at22–25 ◦C and 95% relative humidity.
2.2. Cloning and sequence analysis of 4D8 cDNAs anddeduced proteins
Total RNA was extracted from homogenized tick samplesusing TRI Reagent (Sigma, St. Louis, MO, USA), except forI. ricinus and B. microplus RNA which were extracted usingthe RNA Instapur kit (Eurogentec, Seraing, Belgium) and theRNeasy mini kit (Qiagen, Valencia, CA, USA), respectively,according to the manufacturer’s instructions. The final RNApellet was resuspended in 50–100 �l diethyl pyrocarbonate-treated distilled deionized sterile water. RT-PCR reactionswere performed using the Access RT-PCR system (Promega,Madison, WI, USA). One microliter RNA was reverse tran-scribed in a 50 �l reaction mixture (1.5 mM MgSO4, 1×avian myeloblastosis virus (AMV) RT/Thermus flavus (Tfl)reaction buffer, 0.2 mM each deoxynucleoside triphosphate(dNTP), 5 U AMV RT, 5 u Tfl DNA polymerase (Promega),10 pmol of each primer) at 48 ◦C for 45 min. After 2 min incu-bation at 94 ◦C, PCR was performed in the same reactionmixture for 35 cycles with specific primers and amplificationctosoed4RsrwptobDfi(dDRi
pBWdGn
accines [19,20].The analysis of gene function in ticks has been limited until
he recent application of RNA interference (RNAi) technol-gy through double-stranded (ds)RNA-mediated gene silenc-ng [22–32]. Nevertheless, the function of some of the tickrotective antigens discovered remains unknown.
In the study reported herein, 4D8 sequences were clonedrom different tick genera and its function was characterizedy RNAi in several tick species. The results of these experi-ents suggest that 4D8 is a conserved protein involved in theodulation of tick blood ingestion and reproduction.
. Materials and methods
.1. Ticks
I. scapularis, A. americanum, D. variabilis and R. san-uineus adult ticks were obtained from the Oklahomatate University (OSU) Tick Rearing Facility. Larvae andymphs were fed on rabbits and adult ticks were fed onheep. D. marginatus, H. punctata and Hy. m. margina-um ticks were field collected in south-central Spain. I.icinus and B. microplus strains Santa Luisa and Munoznd Cepich and Susceptible were kindly provided by Drs.ilan Labuda (Institute of Zoology, Slovak Academy of Sci-
nces, Bratislava, Slovakia), Robert J. Miller (Cattle Feverick Research Laboratory, USDA, Edinburgh, TX, USA)nd Rodrigo Rosario (INIFAP, Juitepec, Morelos, Mexico),espectively. Animals were housed at the Tick Rearing Labo-
onditions (Table 1). Oligonucleotide primers for amplifica-ion and cloning of tick 4D8 cDNAs were designed basedn the sequence of the I. scapularis 4D8 (GenBank acces-ion no. AY652654; [19]) and on the sequences that werebtained from other tick species during the course of thexperiments. RA4D85 and RA4D83 oligonucleotides wereesigned based on the sequence of the R. appendiculatusD8 (RaGI TC107) after BLAST [33] search of the TIGR. appendiculatus Gene Index (http://www.tigr.org/tigr-cripts/tgi/T index.cgi?species=r appendiculatus). Controleactions were performed using the same procedures butithout RT to control for DNA contamination in the RNAreparations and without RNA added to control contamina-ion of the PCR reaction. PCR products were electrophoresedn 1% agarose gels to check the size of amplified fragmentsy comparison to a DNA molecular weight marker (1 kbNA ladder, Promega). Amplified fragments were resin puri-ed (Wizard, Promega) and cloned into the pGEM-T vectorPromega) for sequencing both strands by double-strandedye-termination cycle sequencing (Core Sequencing Facility,epartment of Biochemistry and Molecular Biology, Nobleesearch Center, Oklahoma State University). At least two
ndependent clones were sequenced for each PCR.Multiple sequence alignment was performed using the
rogram AlignX (Vector NTI Suite V 5.5, InforMax, Northethesda, MD, USA) with an engine based on the Clustal
algorithm [34]. Nucleotides were coded as unordered,iscrete characters with five possible character states: A, C,, T, or N and gaps were coded as missing data. Phyloge-etic analyses were implemented with 4D8 DNA and protein

4084 J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095
Table 1Primer sets and RT-PCR conditions used for this study
Tick species Upstream/downstream primer sequences (5′–3′) PCR conditionsa
I. scapularis 4D8R5: GCTTGCGCAACATTAAAGCGAAC 62 ◦C, 30 sI. ricinus 4D8-R: TGCTTGTTTGCAGATGCCCATCA 68 ◦C, 1 minA. americanum 4D8R5: GCTTGCGCAACATTAAAGCGAAC 58 ◦C, 30 s
4D833: TTTGGTCGTACGTAAACTTGACAAATGTG 68 ◦C, 1 min
B. microplus, Santa Luisa 4D8R5: GCTTGCGCAACATTAAAGCGAAC 52 ◦C, 30 s4D833: TTTGGTCGTACGTAAACTTGACAAATGTG 68 ◦C, 1 min
Cepich, Munoz and susceptible BM4D85: ACATGACTGGGACCCCTTGCAC 55 ◦C, 30 sBM4D83: CTGTTCTGCGAGTTTGGTAGATAG 68 ◦C, 1 min
R. sanguineus, D. variabilis, D. marginatus,Hy. m. marginatum, H. punctata
RA4D85: ATGGCTTGTGCGACATTAAAGCGGAC 59 ◦C, 30 sRA4D83: TTACGACAAATAGCTGGGCGTAGC 68 ◦C, 1 min
a PCR conditions are shown for annealing and extension in RT-PCR.
sequences using MEGA Version 3.0 [35]. D. melanogaster(GenBank accession number AAF50569) and A. gambiae(EAA04195) sequences were used as outgroups. Maxi-mum parsimony (MP) analyses were conducted with equalweights for all characters and substitutions, heuristic searcheswith 10 random additions of input taxa and tree bisection-reconnection (TBR) branch-swapping. Minimum evolution(ME) and neighbor Joining (NJ) trees were constructed basedon p-distances and pairwise deletion of gaps. Stability oraccuracy of inferred topology(ies) were assessed via boot-strap analysis of 1000 replications.
Conserved domains in proteins were analyzed by search-ing CDD, a curated Entrez database of conserved domainalignments [36] at NCBI web site. Analysis of proteinstructure and function was done by the PredictProteinserver (http://www.predictprotein.org) [37] through Loc3d(http://cubic.bioc.columbia.edu/db/LOC3d/).
2.3. Generation of dsRNA
Oligonucleotide primers containing T7 promotersequences for in vitro transcription and synthesis of dsRNAwere synthesized to amplify DNA encoding tick 4D8 (450–555 bp) and the I. scapularis 4A8 (584 bp; GenBank acces-sion no. CD052546) sequences. The primer sequences wereas follows: I. scapularis 4D8, D8T75: 5′-TAATACGACTCA-CTATAGGGTACTATGGCTTGCGCAACATTAAAG-3′and D8T73: 5′-TAATACGACTCACTATAGGGTACTTTAT-GACAAATAGCTTGGAG-3′; A. americanum 4D8, D8AA-T75: 5′-TAATACGACTCACTATAGGGTACTGACTGGG-ATCCCCTGCACAGT-3′ and D8AAT73: 5′-TAATACGAC-TCACTATAGGGTACTCAAGCTTGGTGGAGAGCACG-3′; R. sanguineus 4D8, D8RST75: 5′-TAATACGACTCACT-ATAGGGTACTGACTGGGACCCGCTGCACAGT-3′ andD8RST73: 5′-TAATACGACTCACTATAGGGTACTCAA-
F hown ina
ig. 1. Alignment of tick 4D8 protein sequences. Protein sequences are ssterisks. Numbers correspond to the sequence of I. scapularis 4D8.
the single letter amino acid code. Identical amino acids are shown with

J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095 4085
GCTTGGTGGAAAGCACG-3′; D. variabilis 4D8, D8AA-T75 and D8DVT73: 5′-TAATACGACTCACTATAGGGTA-CTCGAGCTTGGTGGAAAGGACG-3′; D. marginatus4D8, D8AAT75 and D8DMT73: 5′-TAATACGACTCACTA-TAGGGTACTCGAGCTTGGTGGAAAGCACG-3′. RT-PCR and dsRNA synthesis reactions were performed aspreviously described [30], using the Access RT-PCR system(Promega) and the Megascript RNAi kit (Ambion, Austin,TX, USA). The dsRNA was purified and quantified byspectrometry.
2.4. Injection of ticks with dsRNA
I. scapularis, A. americanum, R. sanguineus, D. variabilisand D. marginatus ticks were injected with approximately0.2–0.5 �l of dsRNA (6 × 1010 to 2 × 1011 molecules per�l) in the lower right quadrant of the ventral surface of theexoskeleton of ticks [30]. The injections were done with aHamilton syringe with a 1 in., 33 gauge needle. Control tickswere injected with I. scapularis 4A8 dsRNA, injection buffer(10 mM Tris–HCl, pH 7, 1 mM EDTA) or were left uninjected[30]. Fifty ticks were used in each group, except with D.marginatus for which 30 ticks were used per group. The tickswere held in a humidity chamber for 1 day after which theywere allowed to feed on a sheep with male ticks. Female ticksthat fed to repletion or those that were removed from the sheepaaa
2i
gaia
described above using oligonucleotide primers for cloningof 4D8 or designed for the preparation of dsRNA [30]. Tick�-actin was amplified as a control gene unrelated to thesequences targeted by RNAi [30]. Control reactions wereperformed using the same procedures but without RT to testfor DNA contamination in the RNA preparations and withoutRNA added to test for contaminations of the PCR reaction.PCR products were electrophoresed on 1% agarose gels tocheck the size of amplified fragments by comparison to aDNA molecular weight marker (1 kb DNA ladder, Promega).
Immunohistochemistry. I. scapularis female guts weredissected and fixed in formaldehyde and embedded in paraf-fin for immunohistochemical analysis of 4D8 expression[19]. Sections (4 �m) were prepared and mounted on micro-scope slides that were stored at 4 ◦C. Tissue sections weredeparaffinized and dehydrated twice for 5 min in xylene,100% ethanol, 95% ethanol, followed by a 5 min wash in80% ethanol. For antigen retrieval, slides were incubatedwith 0.05% pronase (DakoCytomation, Glostrup, Denmark)diluted in TBS, pH 7.2 during 15 min. The slides wereincubated for 1 h with rabbit 4D8 antiserum [19] diluted1:400 in PBS, pH 7.2. A preimmune rabbit serum and amonospecific rabbit serum prepared with total tick IDE8proteins were used as negative and positive controls, respec-tively [19]. The slides were blocked in PBS/0.5% Tween20, pH 7.2 (PBST) with 10% goat serum and 5% skimmlUnH1tisiFt
TN larity fo
S ng the pU ithin th
fter 10 days of feeding were collected, weighed and evalu-ted for oviposition by weighing the egg mass oviposited byll ticks in the group.
.5. Analysis to confirm gene silencing by RT-PCR andmmunohistochemistry of tick tissue sections
RT-PCR. Salivary glands and guts were dissected fromroups of five I. scapularis, A. americanum, R. sanguineusnd D. variabilis ticks from mock-injected and dsRNA-njected groups after feeding. Total RNA was isolated andnalyzed for transcription of target genes by RT-PCR as
able 2ucleotide (above diagonal) and amino acid (below diagonal) identity/simi
equences were aligned and percent identity/similarity was determined usiSA). Values for B. microplus strain sequence comparisons are contained w
ilk for 1 h and then incubated for 1 h with peroxidase-abeled goat anti-rabbit IgG (KPL, Inc., Gaithersburg, MD,SA) diluted 1:3000 in PBST. To inactivate the endoge-ous peroxidase activity, slides were incubated with 3%2O2 in PBS, pH 7 and 10% ethanol for 1 h prior to amin incubation with the substrate 3′,3′-diaminobenzidine
etrahydrochloride Fast DAB set (Sigma) followed by stain-ng with hematoxylin for 2 min. After each treatment, thelides were rinsed twice for 5 min in PBST, unless otherwisendicated. All incubations were done at room temperature.or microscopic examination, the slides were rinsed with dis-
illed water and dehydrated two times for 2 min each in 95%
r all pairwise comparisons of tick 4D8 sequences
rogram AlignX (Vector NTI Suite V 5.5, InforMax, North Bethesda, MD,e squared area.

4086 J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095
Fig. 2. Phylogenetic and evolutionary relationship of 4D8 sequences. (A) A strict consensus of nine maximum parsimony trees from the analysis of 4D8 DNAsequences. Numbers above branches indicate percent support for 1000 bootstrap replicates with 10 random additions of input taxa. (B) Condensed tree of thephylogenetic relationships of 4D8 DNA sequences using the minimum-evolution criterion. The length of the 4D8 amino acid (aa) sequences were mapped ontothe tree.
Fig. 3. Silencing of 4D8 expression at mRNA level. The expression of 4D8 was analyzed by RT-PCR in female ticks injected with species-specific 4D8 dsRNA(+) or with injection buffer alone (−) and compared with the expression of tick �-actin (ACT).

J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095 4087
Table 3Effect on tick feeding, oviposition and mortality of targeting 4D8 expression by RNAi
Tick species Tick weight (mg)a Eggs/tick (mg)b Tick mortality (%)c
4D8 dsRNA Control 4D8 dsRNA Control 4D8 dsRNA Control
I. scapularis 95 ± 48* 149 ± 48* 0 34 62** 46**
A. americanum 224 ± 189* 629 ± 103* 0 27 81** 22**
R. sanguineus 53 ± 58* 199 ± 32* 7 85 62** 10**
D. variabilis 141 ± 152* 573 ± 103* 3 243 40** 0**
D. marginatus 0.04 ± 0.03* 38 ± 20* 0 100 100** 80**
a Ticks that completed feeding and those removed after 10 days of feeding were weighed individually and average ± S.D. calculated and compared between4D8 dsRNA- and mock-injected control ticks by Student’s t-test (*P < 0.01).
b Egg masses oviposited by all ticks on each group were weighed and divided by the total number of ticks recovered after feeding.c Tick mortality was evaluated as the ratio of dead female ticks after 10 days of feeding to the total number of female ticks and was compared between 4D8
dsRNA- and mock-injected ticks by χ2–test (**α < 0.01).
ethanol, 100% ethanol, and finally xylene, and mounted inpermount.
2.6. Light microscopy studies of tick gut, salivary glandand reproductive tissue development
Female and male D. variabilis were used to evaluate theeffect of 4D8 RNAi on the development of guts, salivaryglands and reproductive tissues. Ticks were injected with4D8 dsRNA or injection buffer alone as described above anddivided into groups of 50 ticks each. Twenty four hours afterinjection, ticks were used to infest sheep. Unattached tickswere removed 2 days after infestation and ticks were allowed
to feed for eight additional days and then removed from thehost. Ten ticks were cut in half and fixed for microscopystudies. Ticks halves were fixed in 2% glutaraldehyde in0.1 M sodium cacodylate buffer (pH 7.2). Ticks were thenpost-fixed in 2% cacodylate-buffered osmium tetroxide (pH7.2), dehydrated in a graded series of ethanol and embed-ded in epoxy resin after Kocan et al. [38]. Thick sections(1.0 �m) were cut and stained with Mallory’s stain [39]for light microscopic observation. Sections were observedand photographed using a light microscope and a 3-chipdigital camera. Tissue development was arbitrarily ratedand compared between 4D8 dsRNA-injected and controlticks.
FArP
ig. 4. Silencing of 4D8 expression at the protein level. The expression of 4D8 wasmonospecific rabbit serum prepared with total IDE8 cells tick proteins (A) and a
espectively. Ticks were injected with injection buffer alone (C) or a species-specificositive staining of 4D8 was visualized using a peroxidase-labeled goat anti-mouse
analyzed by immunohistochemistry of gut sections of I. scapualris females.preimmune rabbit serum (B) were used as positive and negative controls,
4D8 dsRNA (D) and analyzed using a rabbit anti-4D8 polyclonal antiserum.IgG (arrows).

4088 J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095
Fig. 5. Role of 4D8 in tick growth and development. (A) Example of the extreme differences observed in 4D8 dsRNA- and mock-injected ticks after feeding.(B) Distribution of female tick weight after feeding. Maximum, minimum and average tick weight is shown for each group. Abbreviations: Is, I. scapularis;Aa, A. americanum; Dv, D. variabilis; Rs, R. sanguineus.
2.7. Statistical analysis
The weight of female ticks after feeding was comparedby Student’s t-test between 4D8 dsRNA- and mock-injectedticks. Tick mortality was evaluated as the ratio of dead femaleticks after 10 days of feeding to the total number of femaleticks and was compared between 4D8 dsRNA and mock-injected ticks by χ2-test as implemented in Mstat 4.01.
3. Results
3.1. The sequence of 4D8 is conserved among tickspecies
The sequence of 4D8 was cloned and analyzed from 10tick species belonging to 6 different genera (Fig. 1). Sequence
identity/similarity varied among tick species between 65–98and 60–98% at the nucleotide and protein levels, respec-tively (Table 2), with regions of extensive similarity betweenamino acids 1–32, 39–83 and 112–184 of the I. scapularis4D8 sequence (Fig. 1). The 4D8 sequence was also highlyconserved among strains of B. microplus (99–100% identity;Table 2). Preliminary analysis of protein structure suggestedthat 4D8 is an intracellular protein but did not result in con-clusive predictions of the putative function of 4D8.
Phylogenetic analysis of 4D8 tick sequences was con-ducted using 4D8 sequences derived from Drosophilamelanogaster and Anopheles gambiae as outgroups.The results of maximum parsimony (MP), minimumevolution (ME) and neighbor joining (NJ) analysisof DNA and protein sequences gave similar results(Fig. 2 and data not shown). 4D8 DNA sequences pro-
F tissue di bars) and . SalivaO f maturn 1 beingo s: ND,i
ig. 6. Light microscopy studies of tick gut, salivary gland and reproductivenjected with 4D8 dsRNA (black bars) or with injection buffer alone (whiteevelopment. Males were also examined for the presence of mature spermvaries were rated 1–5, 1 being undeveloped and 5 indicating the presence oecrotic and 5 being normal with no necrosis. Mature sperm was rated 1–5,f ticks analyzed reflect absence of tissue in the cross section. Abbreviationn replete female ticks because they would have degenerated by this time.
evelopment. Average (±S.D.) rating for 5–10 female and male D. variabilisd examined microscopically for gut, salivary gland and reproductive tissuery glands were rated 1–5, 1 being the most shrunken and 5 being normal.e eggs. Guts and male reproductive tissues were rated 1–5, 1 being the mostabsent and 2–5 indicating the relative quantity. Differences in the number
not determined because salivary glands were not observed which is normal
J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095 4089
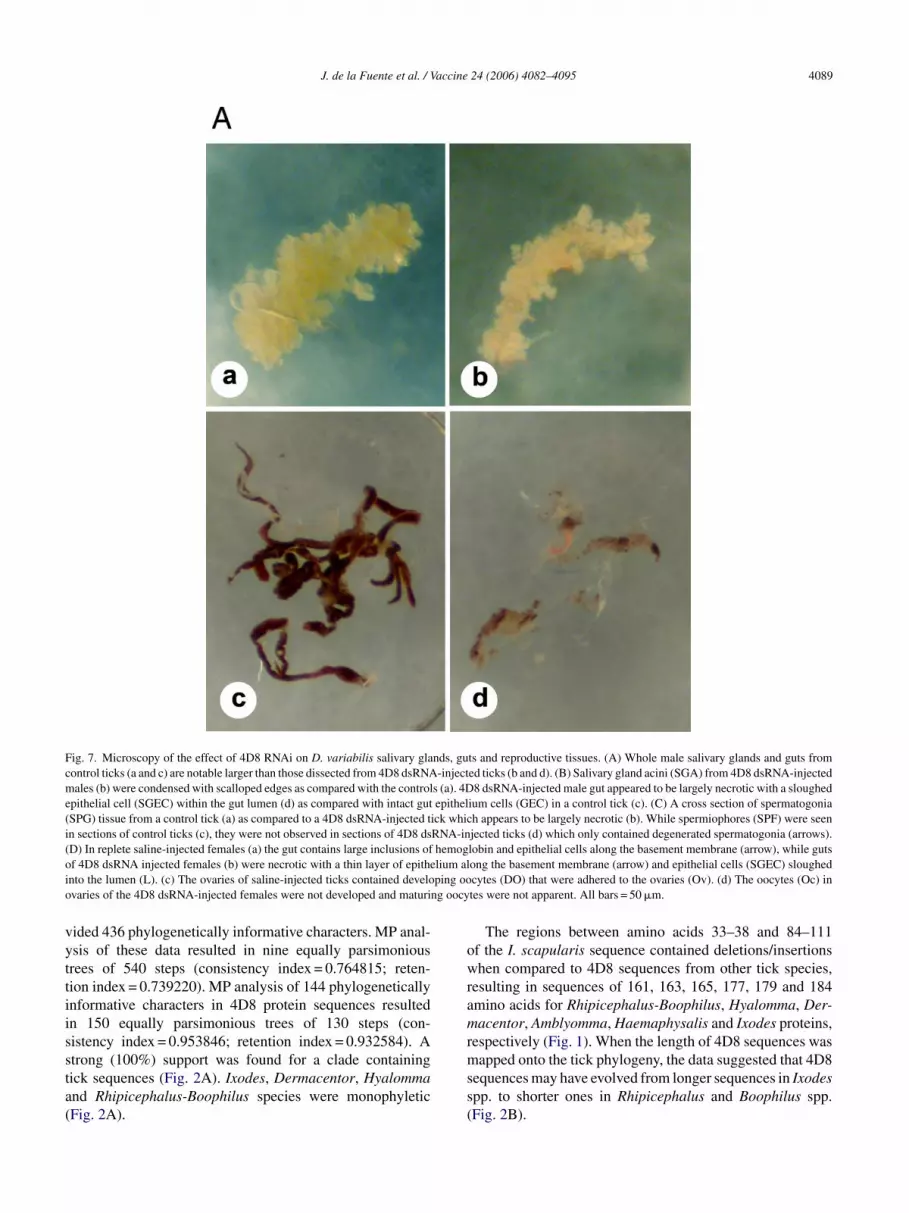
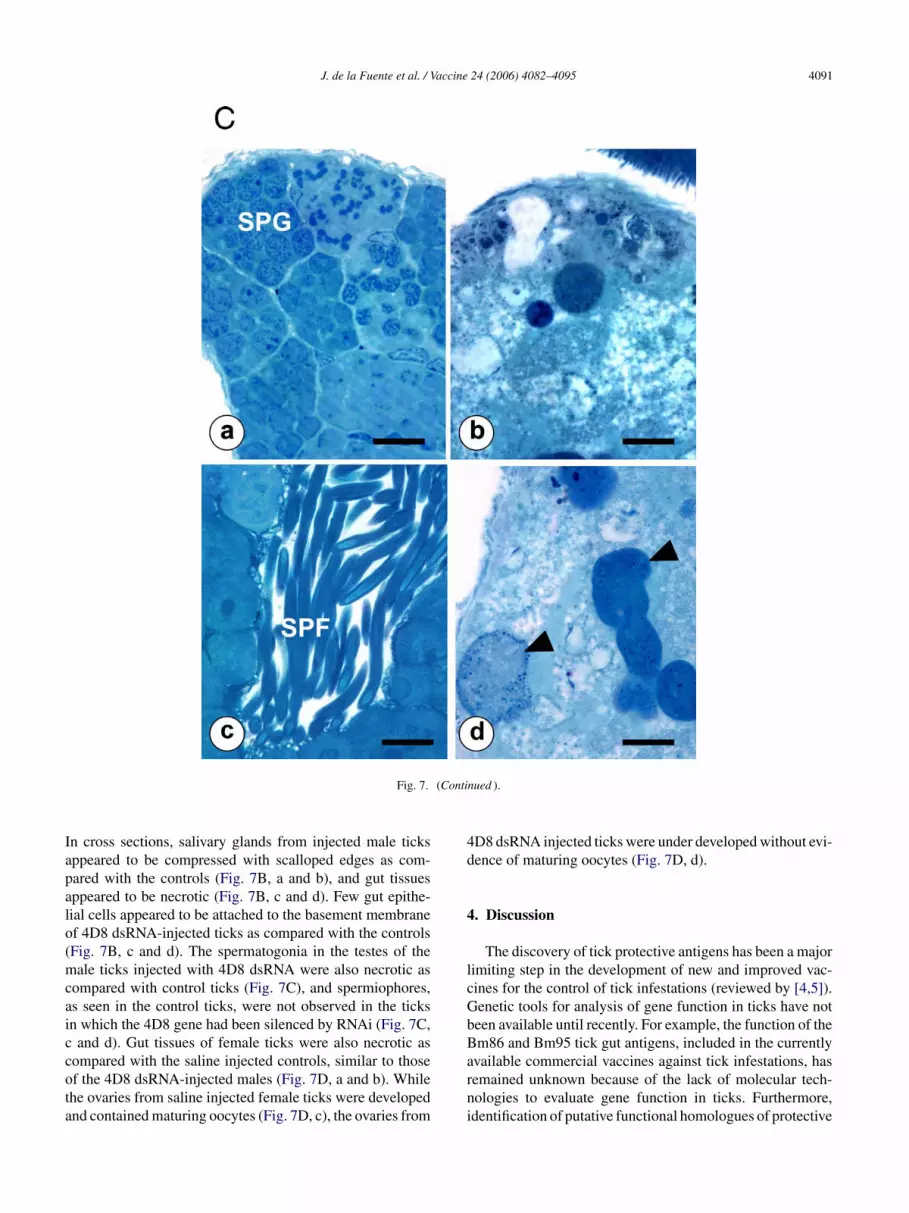
Fig. 7. Microscopy of the effect of 4D8 RNAi on D. variabilis salivary glands, guts and reproductive tissues. (A) Whole male salivary glands and guts fromcontrol ticks (a and c) are notable larger than those dissected from 4D8 dsRNA-injected ticks (b and d). (B) Salivary gland acini (SGA) from 4D8 dsRNA-injectedmales (b) were condensed with scalloped edges as compared with the controls (a). 4D8 dsRNA-injected male gut appeared to be largely necrotic with a sloughedepithelial cell (SGEC) within the gut lumen (d) as compared with intact gut epithelium cells (GEC) in a control tick (c). (C) A cross section of spermatogonia(SPG) tissue from a control tick (a) as compared to a 4D8 dsRNA-injected tick which appears to be largely necrotic (b). While spermiophores (SPF) were seenin sections of control ticks (c), they were not observed in sections of 4D8 dsRNA-injected ticks (d) which only contained degenerated spermatogonia (arrows).(D) In replete saline-injected females (a) the gut contains large inclusions of hemoglobin and epithelial cells along the basement membrane (arrow), while gutsof 4D8 dsRNA injected females (b) were necrotic with a thin layer of epithelium along the basement membrane (arrow) and epithelial cells (SGEC) sloughedinto the lumen (L). (c) The ovaries of saline-injected ticks contained developing oocytes (DO) that were adhered to the ovaries (Ov). (d) The oocytes (Oc) inovaries of the 4D8 dsRNA-injected females were not developed and maturing oocytes were not apparent. All bars = 50 �m.
vided 436 phylogenetically informative characters. MP anal-ysis of these data resulted in nine equally parsimonioustrees of 540 steps (consistency index = 0.764815; reten-tion index = 0.739220). MP analysis of 144 phylogeneticallyinformative characters in 4D8 protein sequences resultedin 150 equally parsimonious trees of 130 steps (con-sistency index = 0.953846; retention index = 0.932584). Astrong (100%) support was found for a clade containingtick sequences (Fig. 2A). Ixodes, Dermacentor, Hyalommaand Rhipicephalus-Boophilus species were monophyletic(Fig. 2A).
The regions between amino acids 33–38 and 84–111of the I. scapularis sequence contained deletions/insertionswhen compared to 4D8 sequences from other tick species,resulting in sequences of 161, 163, 165, 177, 179 and 184amino acids for Rhipicephalus-Boophilus, Hyalomma, Der-macentor, Amblyomma, Haemaphysalis and Ixodes proteins,respectively (Fig. 1). When the length of 4D8 sequences wasmapped onto the tick phylogeny, the data suggested that 4D8sequences may have evolved from longer sequences in Ixodesspp. to shorter ones in Rhipicephalus and Boophilus spp.(Fig. 2B).

4090 J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095
Fig. 7. (Continued ).
3.2. The silencing of 4D8 expression affects female ticksurvival, engorgement and oviposition
RNA interference was used to study the function of 4D8in five species of ticks (Table 3). Female ticks were injectedwith species-specific 4D8 dsRNA and compared with neg-ative control ticks that were either injected with control4A8 dsRNA, mock injected with injection buffer alone ornot injected. In the initial series of experiments, differenceswere not observed between 4A8 dsRNA injected or untreatedticks and those injected with injection buffer alone ([30] anddata not shown). Therefore, in subsequent experiments ticksinjected with an equal volume of injection buffer alone servedas negative controls.
The silencing of 4D8 expression after RNAi was demon-strated at the mRNA and protein levels by RT-PCR andimmunohistochemistry, respectively (Figs. 3 and 4). Theinjection of species-specific 4D8 dsRNA resulted in thereduction of tick weight, oviposition and survival after feed-ing (Table 3 and Fig. 5A and B). These results were con-
sistent in five tick species belonging to four different genera(Table 3).
The most pronounced effect of 4D8 RNAi was on ovipo-sition, which was reduced in over 90% in all tick speciesevaluated (Table 3). The effect on oviposition was not a directconsequence of the reduction in tick weight because someticks that did not lay eggs in the 4D8 dsRNA-injected groupshad a weight similar to control ticks (Fig. 5B).
3.3. The silencing of 4D8 expression affects gut, salivarygland and reproductive tissues in D. variabilis ticks
RNAi was done in female and male D. variabilis to evalu-ate the effect of 4D8 gene silencing on gut, salivary gland andreproductive tissues. The development of gut, salivary glandand reproductive tissues was affected in female and male ticksinjected with 4D8 dsRNA but not in control ticks injectedwith saline (Fig. 6). Whole guts and salivary glands dissectedfrom the 4D8 dsRNA-injected ticks after they had fed werenotably smaller as compared with the controls (Fig. 7A).
J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095 4091
Fig. 7. (Continued ).
In cross sections, salivary glands from injected male ticksappeared to be compressed with scalloped edges as com-pared with the controls (Fig. 7B, a and b), and gut tissuesappeared to be necrotic (Fig. 7B, c and d). Few gut epithe-lial cells appeared to be attached to the basement membraneof 4D8 dsRNA-injected ticks as compared with the controls(Fig. 7B, c and d). The spermatogonia in the testes of themale ticks injected with 4D8 dsRNA were also necrotic ascompared with control ticks (Fig. 7C), and spermiophores,as seen in the control ticks, were not observed in the ticksin which the 4D8 gene had been silenced by RNAi (Fig. 7C,c and d). Gut tissues of female ticks were also necrotic ascompared with the saline injected controls, similar to thoseof the 4D8 dsRNA-injected males (Fig. 7D, a and b). Whilethe ovaries from saline injected female ticks were developedand contained maturing oocytes (Fig. 7D, c), the ovaries from
4D8 dsRNA injected ticks were under developed without evi-dence of maturing oocytes (Fig. 7D, d).
4. Discussion
The discovery of tick protective antigens has been a majorlimiting step in the development of new and improved vac-cines for the control of tick infestations (reviewed by [4,5]).Genetic tools for analysis of gene function in ticks have notbeen available until recently. For example, the function of theBm86 and Bm95 tick gut antigens, included in the currentlyavailable commercial vaccines against tick infestations, hasremained unknown because of the lack of molecular tech-nologies to evaluate gene function in ticks. Furthermore,identification of putative functional homologues of protective

4092 J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095
Fig. 7. (Continued ).
genes in different tick species is difficult when the proteinfunction is unknown. RNAi provides a way to character-ize tick protective antigens such as 4D8, which do not havesequence similarity to previously recognized protein domainsof known biological function [17–19].
The cloning and sequence analysis of 4D8 from 10 tickspecies belonging to six different genera demonstrated that4D8 gene and protein sequences are highly conserved amongIxodidae. Furthermore, a high degree of sequence conserva-tion was also found among geographic strains of B. microplus,in which sequence divergence was found in other genes suchas the tick protective antigen Bm86 [5,8,40,41]. Furthermore,the sequence of I. scapularis 4D8 was similar to proteins ofunknown function identified in other invertebrate and verte-brate organisms, suggesting that the protein is highly con-served throughout evolution [19]. The conservation of 4D8sequences among invertebrate and vertebrate organisms mayraise the question of safety when using 4D8 for immunizationwith the potential of inducing autoimmune responses dam-aging to the host. However, immunization with intracellularproteins has been proposed for other organisms, which maysuggest a low risk to induce autoimmune responses againstintracellular proteins in vertebrate hosts [42–44].
The phylogeny of ticks obtained with 4D8 sequences wassimilar to the phylogenies obtained with other mitochondrialand nuclear genes (reviewed by [45]). This gene may, there-
fore, be useful for studying the phylogeny and evolution ofticks. Our results agreed with recent hypotheses on the evolu-tion of ticks (reviewed by [45,46]) and suggest that 4D8 haveevolved to shorter sequences in ticks and other organisms(Fig. 8).
Fig. 8. Scheme of the evolutionary model of 4D8 sequences from a commonancestor to shorter sequences in ticks and other organisms. 4D8 amino acid(aa) sequences were derived from Almazan et al. [19] and from this study.

J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095 4093
Vaccination experiments with 4D8 cDNA and recombi-nant protein resulted in the control of larval, nymphal andadult tick infestations through induction of tick mortality andreductions in tick weight, molting and oviposition [17–20].These experiments suggested that 4D8 is involved in impor-tant biological pathways related to tick feeding and repro-duction.
To further investigate the function of 4D8, we conducteda series of RNAi experiments in five tick species from fourdifferent genera. The RNAi experiments reported herein,extended to R. sanguineus, D. variabilis and D. margina-tus the previous application of this technology to other tickspecies [22–32]. Injection of 4D8 dsRNA into ticks resultedin the silencing of 4D8 expression at the mRNA and proteinlevels. Based on these studies, 4D8 appears to be involved inthe modulation of tick blood ingestion and reproduction. Themost pronounced effect of 4D8 RNAi was on tick ovipositionwhich was consistent among all tick species studied.
Female and male D. variabilis were used to analyze theeffect of silencing the 4D8 gene expression on tick guts, sali-vary glands and reproductive tissues. The silencing of 4D8expression after RNAi affected gut, salivary gland and repro-ductive organ development. Tick guts and salivary glands arethe major site of pathogen infection, development and trans-mission [47–49]. The effect of 4D8 silencing on reproductivetissues in both female and male ticks further reinforces therc
Cimtpapi
io(s
eiaoosifcwdnm
These results point to the potential use of subolesin in devel-opment of new and novel tick vaccines that may both con-trol tick infestations and affect transmission of tick-bornepathogens.
Acknowledgments
This research was supported by the Oklahoma Agri-cultural Experiment Station (project 1669), the SitlingtonEndowed Chair for Food Animal Research (K.M. Kocan,Oklahoma State University) and the Instituto de Cien-cias de la Salud, Spain (ICS-JCCM) (project 03052-00).Consuelo Almazan was funded by Pfizer Animal Health,Kalamazoo, MI, USA and a grant-in-aid from the CONA-CYT and Promep (University of Tamaulipas), Mexico. V.Naranjo was founded by Consejerıa de Educacion, JCCM,Spain. Drs. Milan Labuda (Institute of Zoology, SlovakAcademy of Sciences, Bratislava, Slovakia), Robert J. Miller(Cattle Fever Tick Research Laboratory, USDA, Edinburg,TX, USA), Rodrigo Rosario (INIFAP, Juitepec, Morelos,Mexico), Francisco Ruiz-Fons (IREC, Ciudad Real, Spain)and Jerry Bowman (Tick Rearing Facility, Department ofEntomology and Plant Pathology, Oklahoma State Uni-versity, OK, USA) are acknowledged for providing ticksand tick material. The staff of the Fundacion General andFar
R
ole for this protein in the control of tick developmental pro-esses and reproduction.
Expression of I. scapularis, human, D. melanogaster andaenorhabditis elegans 4D8 homologues has been detected
n a variety of adult and immature tissues [19,50,51]. Further-ore, studies in D. melanogaster and C. elegans have shown
hat 4D8 may be involved in the control of developmentalrocesses in these organisms [50,51]. These results are ingreement with the findings reported herein in ticks and sup-ort the role of 4D8 in the development of tissues involvedn tick blood ingestion and utilization and reproduction.
Because of the critical role that 4D8 plays during tick feed-ng and oviposition, which ultimately results in the reductionf tick progeny, we proposed the generic name “subolesin”Latin, suboles: offspring, progeny) for tick 4D8 proteins andubA for the subolesin-encoding gene.
In summary, we have provided evidence through RNAixperiments in several tick species that subolesin is involvedn the modulation of tick blood ingestion and utilizationnd reproduction. The pleiotropic effects on tick organsf subolesin silencing, together with the high conservationf subolesin nucleotide and protein sequences among tickpecies, as well as other invertebrate and vertebrate organ-sms suggest a highly specialized and conserved biologicalunction for 4D8, which affects different physiological pro-esses in the tick. Vaccination with recombinant subolesinould likely impact both the feeding capacity and repro-uctive ability of ticks, thereby impacting the immediateumber of ticks feeding and the amount of blood that theyay intake as well as the overall tick numbers over time.
inca Galiana, University of Castilla—La Mancha, Spainre acknowledged for support and technical assistance,espectively.
eferences
[1] Parola P, Raoult D. Tick-borne bacterial diseases emerging in Europe.Clin Microbiol Infect 2001;7:80–3.
[2] Graf J-F, Gogolewski R, Leach-bing N, Sabatini GA, Molento MB,Bordin EL, et al. Tick control: an industry point of view. Parasitology2004;129:S427–42.
[3] Ginsberg HS. Integrated pest management and allocation of controlefforts for vector-borne diseases. J Vector Ecol 2001;26:32–8.
[4] de la Fuente J, Kocan KM. Advances in the identification and char-acterization of protective antigens for development of recombinantvaccines against tick infestations. Expert Rev Vaccines 2003;2:583–93.
[5] Willadsen P. Anti-tick vaccines. Parasitology 2004;129:S1–S21.[6] Willadsen P, Riding GA, McKenna RV, Kemp DH, Tellam RL,
Nielsen JN, et al. Cloning and expression of a protective antigenfrom the cattle tick Boophilus microplus. Proc Natl Acad Sci USA1989;86:9657–61.
[7] Rodrıguez M, Rubiera R, Penichet M, Montesino R, Cremata J,Falcon V, et al. High level expression of the Boophilus microplusBm86 antigen in the yeast Pichia pastoris forming highly immuno-genic particles for cattle. J Biotechnol 1994;33:135–46.
[8] Garcıa-Garcıa JC, Montero C, Redondo M, Vargas M, Canales M,Boue O, et al. Control of ticks resistant to immunization with Bm86in cattle vaccinated with the recombinant antigen Bm95 isolated fromthe cattle tick, Boophilus microplus. Vaccine 2000;18:2275–87.
[9] Willadsen P, Smith D, Cobon G, McKenna RV. Comparative vaccina-tion of cattle against Boophilus microplus with recombinant antigenBm86 alone or in combination with recombinant Bm91. ParasiteImmunol 1996;18:241–6.

4094 J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095
[10] Andreotti R, Gomes A, Malavazi-Piza KC, Sasaki SD, SampaioCA, Tanaka AS. BmTI antigens induce a bovine protective immuneresponse against Boophilus microplus tick. Int Immunopharmacol2002;2:557–63.
[11] Trimnell AR, Hails RS, Nuttall PA. Dual action ectoparasitevaccine targeting “exposed” and “concealed” antigens. Vaccine2002;20:3560–8.
[12] Wang H, Nuttall PA. Immunoglobulin-binding proteins in ticks: newtarget for vaccine development against a blood-feeding parasite. CellMol Life Sci 1999;56:286–95.
[13] Mulenga A, Sugimoto C, Sako Y, Ohashi K, Musoke A, ShubashM, et al. Molecular characterization of a Haemaphysalis longi-cornis tick salivary gland-associated 29-kilodalton protein and itseffect as a vaccine against tick infestation in rabbits. Infect Immun1999;67:1652–8.
[14] Tsuda A, Mulenga A, Sugimoto C, Nakajima M, Ohashi K,Onuma M. cDNA cloning, characterization and vaccine effect anal-ysis of Haemaphysalis longicornis tick saliva proteins. Vaccine2001;19:4287–96.
[15] Imamura S, da Silva Vaz Junior I, Sugino M, Ohashi K, Onuma M.A serine protease inhibitor (serpin) from Haemaphysalis longicornisas an anti-tick vaccine. Vaccine 2005;23:1301–11.
[16] You MJ. Immunization of mice with recombinant P27/30 pro-tein confers protection against hard tick Haemaphysalis longicornis(Acari: Ixodidae) infestation. J Vet Sci 2005;6:47–51.
[17] Almazan C, Kocan KM, Bergman DK, Garcia-Garcia JC, BlouinEF, de la Fuente J. Identification of protective antigens for the con-trol of Ixodes scapularis infestations using cDNA expression libraryimmunization. Vaccine 2003;21:1492–501.
[18] Almazan C, Kocan KM, Bergman DK, Garcia-Garcia JC, Blouin EF,de la Fuente J. Characterization of genes transcribed in an Ixodes
[
[
[
[
[
[
[
[
[
[28] Pal U, Li X, Wang T, Montgomery RR, Ramamoorthi N, DesilvaAM, et al. TROSPA, an Ixodes scapularis receptor for Borreliaburgdorferi. Cell 2004;119:457–68.
[29] Miyoshi T, Tsuji N, Islam MK, Kamio T, Fujisaki K. Genesilencing of a cubilin-related serine proteinase from the hard tickHaemaphysalis longicornis by RNA interference. J Vet Med Sci2004;66:1471–3.
[30] de la Fuente J, Almazan C, Blouin EF, Naranjo V, Kocan KM.RNA interference screening in ticks for identification of protectiveantigens. Parasitol Res 2005;96:137–41.
[31] Soares CA, Lima CM, Dolan MC, Piesman J, Beard CB, Zeid-ner NS. Capillary feeding of specific dsRNA induces silencing ofthe isac gene in nymphal Ixodes scapularis ticks. Insect Mol Biol2005;14:443–52.
[32] Ramamoorthi N, Narasimhan S, Pal U, Bao F, Yang XF, Fish D,et al. The Lyme disease agent exploits a tick protein to infect themammalian host. Nature 2005;436:573–7.
[33] Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic localalignment search tool. J Mol Biol 1990;215:403–10.
[34] Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improvingthe sensitivity of progressive multiple sequence alignment throughsequence weighing, positions-specific gap penalties and weightmatrix choice. Nucl Acid Res 1994;22:4673–80.
[35] Kumar S, Tamura K, Nei M. MEGA3: integrated software for molec-ular evolutionary genetics analysis and sequence alignment. BriefingsBioinform 2004;5:150–63.
[36] Marchler-Bauer A, Anderson JB, DeWeese-Scott C, Fedorova ND,Geer LY, He S, et al. CDD: a curated Entrez database of conserveddomain alignments. Nucl Acids Res 2003;31:383–7.
[37] Rost B, Yachdav G, Liu J. The PredictProtein server. Nucl AcidsRes 2004;32(Web Server issue):W321–6.
[
[
[
[
[
[
[
[
[
[
[
scapularis cell line that were identified by expression library immu-nization and analysis of expressed sequence tags. Gene Ther MolBiol 2003;7:43–59.
19] Almazan C, Blas-Machado U, Kocan KM, Yoshioka JH, BlouinEF, Mangold AJ, et al. Characterization of three Ixodesscapularis cDNAs protective against tick infestations. Vaccine2005;23:4403–16.
20] Almazan C, Kocan KM, Blouin EF, de la Fuente J. Vaccination withrecombinant tick antigens for the control of Ixodes scapularis adultinfestations. Vaccine 2005;23:5294–8.
21] Weiss BL, Kaufman WR. Two feeding-induced proteins from themale gonad trigger engorgement of the female tick Amblyommahebraeum. Proc Natl Acad Sci USA 2004;101:5874–9.
22] Aljamali MN, Sauer JR, Essenberg RC. RNA interference: applica-bility in tick research. Exp Appl Acarol 2002;28:89–96.
23] Aljamali MN, Bior AD, Sauer JR, Essenberg RC. RNA interfer-ence in ticks: a study using histamine binding protein dsRNAin the female tick Amblyomma americanum. Insect Mol Biol2003;12:299–305.
24] Karim S, Ramakrishnan VG, Tucker JS, Essenberg RC, Sauer JR.Amblyomma americanum salivary glands: double-stranded RNA-mediated gene silencing of synaptobrevin homologue and inhibitionof PGE2 stimulated protein secretion. Insect Biochem Mol Biol2004;34:407–13.
25] Karim S, Ramakrishnan VG, Tucker JS, Essenberg RC, Sauer JR.Amblyomma americanum salivary gland homolog of nSec1 is essen-tial for saliva protein secretion. Biochem Biophys Res Commun2004;324:1256–63.
26] Karim S, Miller NJ, Valenzuela J, Sauer JR, Mather TN. RNAi-mediated gene silencing to assess the role of synaptobrevin andcystatin in tick blood feeding. Biochem Biophys Res Commun2005;334:1336–42.
27] Narasimhan S, Montgomery RR, DePonte K, Tschudi C, Marcan-tonio N, Anderson JF, et al. Disruption of Ixodes scapularis anti-coagulation by using RNA interference. Proc Natl Acad Sci USA2004;101:1141–6.
38] Kocan KM, Hair JA, Ewing SA. Ultrastructure of Anaplasmamarginale Theiler in Dermacentor andersoni Stiles and Dermacentorvariabilis (Say). Am J Vet Res 1980;41:1966–76.
39] Richardson KC, Jarret L, Finke FH. Embedding in epoxy resinsfor ultrathin sectioning in electron microscopy. Stain Technol1960;35:313–23.
40] Garcıa-Garcıa JC, Gonzalez IL, Gonzalez DM, Valdes M, MendezL, Lamberti J, et al. Sequence variations in the Boophilusmicroplus Bm86 locus and implications for immunoprotection incattle vaccinated with this antigen. Exp Appl Acarol 1999;23:883–95.
41] de la Fuente J, Garcıa-Garcıa JC, Gonzalez DM, Izquierdo G, Ocha-gavia ME. Molecular analysis of Boophilus spp. (Acari: Ixodidae)tick strains. Vet Parasitol 2000;92:209–22.
42] Elad D, Segal E. Immunogenicity in calves of a crude ribosomalfraction of Trichophyton verrucosum: a field trial. Vaccine 1995;13:83–7.
43] Silva CL. The potential use of heat-shock proteins to vacci-nate against mycobacterial infections. Microbes Infect 1999;1:429–35.
44] Cassataro J, Velikovsky CA, Giambartolomei GH, Estein S, BrunoL, Cloeckaert A, et al. Immunogenicity of the Brucella melitensisrecombinant ribosome recycling factor-homologous protein and itscDNA. Vaccine 2002;20:1660–9.
45] Barker SC, Murrell A. Systematics and evolution of ticks with alist of valid genus and species names. Parasitology 2004;129:S15–36.
46] de la Fuente J. The fossil record and the origin of ticks (Acari:Parasitiformes: Ixodida). Exp Appl Acarol 2003;29:331–44.
47] Kocan KM, Stiller D, Goff WL, Claypool PL, Edwards W, EwingSA, et al. Development of Anaplasma marginale in male Derma-centor andersoni transferred from parasitemic to susceptible cattle.Am J Vet Res 1992;53:499–507.
48] Kocan KM, Goff WL, Stiller D, Claypool PL, Edwards W, Ewing S,et al. Persistence of Anaplasma marginale (Rickettsiales: Anaplas-mataceae) in male Dermacentor andersoni (Acari: Ixodidae) trans-

J. de la Fuente et al. / Vaccine 24 (2006) 4082–4095 4095
ferred successively from infected to susceptible calves. J Med Ento-mol 1992;29:657–68.
[49] Kocan K, de la Fuente J, Blouin EF, Garcia-Garcia JC. Anaplasmamarginale (Rickettsiales: Anaplasmataceae): recent advances indefining host–pathogen adaptations of a tick-borne rickettsia. Par-asitology 2004;129:S285–300.
[50] Maeda I, Kohara Y, Yamamoto M, Sugimoto A. Large-scale analy-sis of gene function in Caenorhabditis elegans by high-throughputRNAi. Curr Biol 2001;11:171–6.
[51] Pena-Rangel MT, Rodriguez I, Riesgo-Escovar JR. A misex-pression study examining dorsal thorax formation in Drosophilamelanogaster. Genetics 2002;160:1035–50.





























