Embed Size (px)
Citation preview

Tilapia (Oreochromis niloticus) Vitellogenins:Development of Homologous and HeterologousELISAs and Analysis of Vitellogenin PathwayThrough the Ovarian Follicle
PAP NDIAYE1, JEAN FORGUE2, VALERIE LAMOTHE3, CHANTAL CAUTY4,PHILIPPE TACON4, PIERRETTE LAFON5, BLANDINE DAVAIL2,ALEXIS FOSTIER4, FRANC-OISE LE MENN2, AND JESUS NUNEZ6�1IFAN, Universite Cheikh Anta Diop, Dakar, Senegal2Laboratoire de Genomique et de Physiologie des Poissons, Universite deBordeaux I, 33405 Talence, France3Unite Micronutriments Reproduction Sante, ENITAB, 1 cours du general deGaulle, 33175 Gradignan cedex, France4INRA SCRIBE, campus de Beaulieu, 35042 Rennes cedex, France5Laboratoire des Regulations Neuroendocriniennes, EA 2972, Universite deBordeaux I, 33405 Talence cedex, France6IRD, Groupe Aquaculture Continentale Mediterraneenne et Tropicale (GAMET),UR 175 CAVIAR, Av. Agropolis, 34033 Montpellier cedex, France
ABSTRACT Vitellogenin (VTG) of Oreochromis niloticus was again purified, due to theconflicting results found in the literature. Three purification processes have been used:electrophoresis and electro-elution, double chromatography (gel filtration and ion-exchangechromatography) and single ion-exchange chromatography. Using SDS-PAGE we confirmed in allcases the presence of two polypeptidic forms of plasma VTG of 130 kDa (VTG1) and 170 kDa (VTG2).We raised polyclonal antibodies against each VTG form and we demonstrated the complete cross-reactivity of each antibody with both forms of VTG by Enzyme Immuno-Assay (EIA) and Westernblots. The homologous ELISAs developed exhibited a detection limit of 6 ng.ml�1, equivalent to60 ng.ml�1 of plasma VTG and allowed us to quantify the total plasma VTG of O. niloticus with highspecificity and sensitivity. Using photonic and electron immunomicroscopy, we followed the pathwayof VTG into the ovarian follicle (OF) demonstrating that VTG enters the oocyte at stage 3 of OFdevelopment, at the same time as cortical alveoli and lipid globules appear. Heterologous ELISAsperformed on other cichlid species allowed us to quantify plasma VTG in Oreochromis aureus andSarotherodon melanotheron and to detect it in Hemichromis fasciatus, Hemichromis bimaculatusand Tilapia zillii, constituting a reliable tool for monitoring the presence of xeno-estrogens in theenvironment of these fish species. J. Exp. Zool. 305A:576–593, 2006. r 2006 Wiley-Liss, Inc.
The tilapia Oreochromis niloticus belongs to thePerciform order and Cichlidae family. This specieswas originally restricted to the Nile Basin and thenorth tropical areas of Africa, from Senegal to thecentral Great Lakes (Philipart and Ruwet, ’82).This species, well adapted to wide variations ofecological factors, has been able to colonizeextremely diverse biotopes (Pullin and Lowe-McConnell, ’82). At present, O. niloticus is widelyspread throughout Africa, South, Central andNorth America as well as Asia, owing to itsaquaculture. Even more, the species is now reared
in temperate countries using power plants hotwater effluents. Its world production was aroundone million tonnes in 2003 (FAO statistics) and iswidely tipped to become an even bigger player inthe international aquaculture area (Young andMuir, 2002).
Published online 13 April 2006 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jez.a.290.
Received 15 August 2005; Accepted 24 January 2006
�Correspondence to: J. Nunez, IRD, Groupe Aquaculture Con-tinentale Mediterraneenne et Tropicale (GAMET), UR 175, CAVIAR,Av. Agropolis, 34033 Montpellier cedex, France. E-mail: [email protected]
r 2006 WILEY-LISS, INC.
JOURNAL OF EXPERIMENTAL ZOOLOGY 305A:576–593 (2006)

Despite numerous publications concerning thereproductive cycle, endocrinology of reproduction,fecundity and sexual behavior or strategy ofreproduction of this species (Babiker and Ibrahim,’79; Fishelson and Yaron, ’83; Baroiller andJalabert, ’89; Kestemont et al., ’89; Tacon et al.,’95, ’96, 2000; Baroiller et al., ’97; Hines et al., ’99;Duponchelle et al., ’99, 2000), few data areavailable about gametogenesis (Dadzie, ’74; Alveset al., ’83; Nakamura et al., ’93; Francolini et al.,2003) and particularly vitellogenesis. In fish, as inother oviparous vertebrates, embryonic and earlylarval development relies on egg yolk nutrients.The main yolk precursor, synthesized by the liverunder estrogenic induction and transported in theplasma, is vitellogenin (VTG), a large phospholi-poglycoprotein, which is incorporated specificallyinto oocytes by receptor-mediated endocytosis(Stifani et al., ’90; Chan et al., ’91; NunezRodriguez et al., ’96; Davail et al., ’98; Hiramatsuet al., 2004).
Fish VTG has been quantified for the first timeby ELISA in Solea vulgaris (Nunez Rodriguezet al., ’89) and since then for various other fishfamilies. Published studies on O. niloticus VTGstructure are conflicting. In previous work (Chanet al., ’91; Lee et al., ’92; Buerano et al., ’95), it hasbeen reported that the two VTGs present in thisspecies were immunochemically different. Thisencouraged us to start the purification of O.niloticus VTGs to be able to detect or to quantifyall forms of VTGs present over again.
The aim of this study was to obtain efficientimmunological tools in order (i) to investigate thedegree of homology between the two forms of VTG(VTG1 and VTG2) possibly present in this species,(ii) to follow the process of VTG internalizationinto the oocyte by immunocytochemistry, (iii) toquantify VTG during a sexual cycle using a specifichomologous ELISA and (iv) to develop hetero-logous ELISAs for other related Cichlid species.
MATERIAL AND METHODS
Fish, hormonal treatment, blood samplingand yolk extraction
Females and males of O. niloticus, weighingfrom about 30 g for immature juveniles to about120 g for the breeders, were obtained from eitherthe Nianga fish farm (Dagana, Senegal), Institutdes Savanes, (Bouake, Ivory Coast) or GAMETexperimental facility (Montpellier, France) andtransported to laboratories under oxygen aeration.
They were reared in 200-l aquaria provided withaerated dechlorinated tap water in natural orexperimental conditions of temperature (241C)and photoperiod, and fed once daily. After accli-matization for 2 weeks, 12 juvenile males wereinjected intra-peritoneally every 2 days during20 days with 4.5mg.g�1 of body weight of17b-estradiol (Sigma) using a stock solution ofestradiol in absolute ethanol (30 mg ml�1) diluted10-fold in cotton oil. Fish blood was collected bycaudal puncture in heparinized plastic centrifugetubes just before the first estrogen injection (t0),every 2 days before each estrogen injection and2 days after the last injection (t14). The blood wascentrifuged immediately at 10,000 g for 5 min, at41C. Plasma samples were pooled, aliquoted andstored at �201C. Vitellogenic follicles were col-lected on vitellogenic females, opened understereo-microscope and yolk samples diluted inNaCl. 12% were centrifuged for 5 min, at 2,000g.The intermediate translucent phase was used asyolk extract. Wild mature males were caught inthe Senegal river, their plasma immediatelyprepared as described above and stored at�201C. Ten females weighing between 50 and150 g were sampled at 3-days postspawning inter-vals, between days 1 and 27 of the normalreproductive cycle. The fish were caught, anesthe-tized, blood-sampled for VTG ELISA and weighed.Following decapitation, the ovaries were removedand weighed for calculation of the gonadosomaticindex (GSI) (ovarian weight�100/body weight).
Other Cichlidae species—Oreochromis aureus,Sarotherodon melanotheron, Tilapia dageti, Tila-pia guineensis, Tilapia zillii, Hemichromis bima-culatus and Hemichromis fasciatus—were caughtin the wild in Senegal and plasma of vitellogenicfemales was sampled.
Microscopy
As soon as the female was caught and anesthe-tized with MS222, the bulb was sectioned, and theheart still beating, the abdominal cavity wasopened for fixation of ovaries, one for photonicmicroscopy, the other for electron microscopy.
For photonic microscopy, one ovary was sec-tioned and immersed in Bouin Holland’s fluid.Ovarian lamellae were isolated under a stereo-microscope. After 72 hr fixation and then dehydra-tion in graded ethanol, pieces were embedded incytoparaffin using butanol as vehicle. For vitello-genic ovarian follicles (OFs), blocks were treatedduring 48 hr with ethanol-60%/glycerine (9:1) in
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 577
J. Exp. Zool. DOI 10.1002/jez.a

order to facilitate the slicing of yolk (Baker, ’45).Serial sections of 7 mm thickness were stained withGroat eosin–hematoxylin, Masson trichrome orHeidenhain azan.
For electron microscopy, the second ovary wastreated at the same time by fixative solution(glutaraldehyde 3%, para-formaldehyde 2%, CaCl28 mM, sodium cacodylate 0.1 M, pH 7.3) either byimmersion (juvenile) or perfusion of the ovarianartery for 30 min (vitellogenic female) and quicklydissected under a stereo-microscope. A comple-mentary fixation of small pieces of ovarianlamellae of about 1 mm3 was performed overnightin fresh fixative at room temperature underdegassed atmosphere. Osmolarity of the fixativefluid was 320 mOsm, when O. niloticus plasma was300 mOsm. Except for immunocytochemistry, apostfixation was performed for 1 hr in 1% OsO4 incacodylate buffer containing 1.5% potassium fer-ricyanide. The fixed samples were then dehy-drated through graded ethanol and propyleneoxide, and embedded in Epon 812 resin.
The semi-thin sections were stained with 0.1%toluidine blue. Ultra-thin sections were contrastedby uranyl acetate and lead citrate.
Electrophoresis
SDS-PAGE (SDS 0.1% polyacrylamide gel 7.5%)was performed using Miniprotean II device(Biorad) with SDS-Tris-glycine buffer, for 1 hrat 120 V for two gels, according to Laemmli (’70).b-mercapto-ethanol was used as reducing agent.Relative molecular weights of separated proteinswere determined by comparing their mobility withstandard proteins using an electrophoresis cali-bration kit (Biorad HMW). Gels were stained withCoomassie Brilliant blue G 250.
VTG purification
All procedures were performed at 41C. In someexperiments, aprotinin and leupeptin or PMSF(1 mM) were used as anti-protease agents.
Electro-elution procedure
Plasma from an end-course estrogenized male(t14) was run on SDS-PAGE. After Coomassie bluestaining, each of the two bands correspondingto the putative VTGs (VTG2 and VTG1) was cutout, minced in 0.5 ml electro-elution buffer((NH4)HCO3 0.4 M, SDS 2%, pH 8.2) and imme-diately put in an electro-elution chamber. Proteinswere electro-eluted through a 10,000 Da cutoffmembrane by a 50 V, 10 mA current. At the end of
the electro-elution, protein was concentrated upto 500 mg.ml�1 on Amicon cell (10,000 Da cutoffmembrane), aliquoted and stored at �301C.
Double chromatography procedure
Gel filtration. Three milliliters of VTG-inducedplasma (t14) were mixed with 2 ml of chromato-graphy buffer (Tris 100 mM, CaCl2 1 mM, pH 7.8)and centrifuged during 5 min at 10,000g. Thesupernatant was deposited onto a 100� 2.5 cm gelfiltration equilibrated column of Sepharose6B (Pharmacia LKB). A 60 ml.h�1 buffer flowrate was applied and fractions of 10 ml werecollected. Fractions containing VTG were identi-fied by SDS-PAGE, and were pooled and con-centrated to 5 ml on the Amicon cell (10,000 Dacutoff membrane).
Ion-exchange chromatography. Previously sepa-rated proteins were applied to a 50� 1.8 cm2
column of DEAE-trysacryl M (IBF) equilibratedwith the previous buffer (flow rate: 36 ml.h�1).After 2 hr of washing, bound plasma proteins wereeluted by 200 ml of a linear gradient of Tris buffer(Tris 100–300 mM, CaCl2 1 mM, pH 7.8). Fractionscontaining VTG were selected by SDS-PAGE,pooled and concentrated using the Amicon cell(10,000 Da cutoff membrane) up to a final concen-tration of 2 mg.ml�1, aliquoted and stored at�301C.
Single chromatography procedure
Single DEAE chromatography. One milliliter ofinduced plasma (t14) was applied on a columncontaining 1.6 ml of ion-exchange resin DEAEFractogel (Merck) equilibrated with Tris buffer(Tris 50 mM, CaCl2 2 H2O 2 mM, pH 8.2) at a flowrate of 36 ml.h�1. After washout of unboundmolecules, the bound proteins were eluted with160 ml of linear gradient of NaCl (NaCl 0–500 mM,Tris 50 mM, CaCl2 2 mM, pH 8.2) at the same flowrate. Eluted fractions (3 ml) containing VTG wereselected by SDS-PAGE, pooled, aliquoted andstored at �301C.
For each procedure, absorbance at 280 nm ofevery eluted fraction was measured.
Immunization and anti-serum harvesting
Twelve-week-old rabbits received injections ofemulsified VTG solution (50% VTG at 200mg.ml�1
in 9% NaCl, and 50% Sigma Freund’s completeadjuvant). Each 1 ml subcutaneous injection,performed in 8–10 points on the back, was givenevery week during 4 weeks and twice during thenext month. Blood was sampled before immuniza-
P. NDIAYE ET AL.578
J. Exp. Zool. DOI 10.1002/jez.a

tion protocol and 1 week after each injection, andserum was obtained after 6 hr of blood clottingat 201C.
Immunizations were performed using VTGpurified by double chromatography (putativeVTG1) and by electro-elution (putative VTG2).
Before use, anti-sera were harvested with VTG-free lyophilized liver and plasma obtained fromwild males. One milliliter of anti-serum wasincubated with 10 mg of lyophilized liver and10 mg of lyophilized plasma for 3 hr at 201Cfollowed by 15 min centrifugation at 2,000g. Thesupernatant was incubated again overnight at 41Cwith the same amount of lyophilized liver andplasma and centrifuged for 15 min at 2,000g and41C. The final supernatant was recovered as anti-serum and stored at �201C.
Immunochemical techniques
Antigen–antibody dilution test
The binding kinetics and the specific activity ofantibodies were performed using Enzyme Immu-no-Assay (EIA). A range of diluted antigen coatedonto microtitration plates (Nunc maxisorp) wascrossed with a range of diluted anti-sera. Theimmunocomplexes were incubated with peroxi-dase anti-peroxidase (PAP) complexes. The perox-idase activity was revealed in the dark by adding200ml per well of orthophenylene-diamine solution(0.05%) in citric acid buffer (0.1 M, pH 5) in thepresence of 0.05% H2O2, giving an orange color.The reaction was stopped after 20 min by adding50ml per well of H2SO4, 4 M.
Immunocytochemistry
Photonic immunocytochemistry
Dewaxed paraffin slices were treated succes-sively with lithium carbonate (1%), hydrogenperoxide (30%), distilled water and phosphatebuffer (PB) (NaHPO4 0.1 M, pH 7.4). Saturationof non-specific binding sites was achieved byincubation in blocking solution (1% normal pigserum in PB) during 20 min. The slides were thenincubated for 1 hr with the primary antibody(anti-VTG1 1:1,000 in blocking solution), washedin PB and incubated for 1 hr in horse–radishperoxidase (HRP) conjugated with swine anti-rabbit IgG (1:250 in blocking solution). Antigen–antibody complexes were revealed as above.
Electron immunocytochemistry
Ultra-thin sections were deposited on gold gridsand incubated for 15 min in PB. Saturation of non-
specific binding sites was performed during 20 minin PB containing 0.3% of normal goat serum and0.1% of ovalbumin. After washing six times withPB, grids were incubated for 1 hr in PB containinganti-VTG1 (1:1,000). Then, after washing threetimes, the grids were incubated for 40 min in PBcontaining (1:100) goldlabelled goat anti-rabbitIgG (Amersham). Finally, after washing threetimes, grids were dried and observed under anelectron microscope (Phillips 200).
Western blot
After electrophoresis, the proteins were trans-ferred onto PVDF (polyvinyldifluoride) membranes(Amersham). Before use, PVDF membranes wereimmersed quickly in methanol and distilled water,and protein transfer was performed using Biorad’sprocedure for 1 hr at 100 V.
Western blot was processed under shackingin TBS-T (Tris 20 mM, NaCl 137 mM, pH 7.6,Tween-20 0.3%) at room temperature, and eachwashing step was performed four times in TBST-Tfor 10 min. PVDF blots were blocked overnight at41C in 5% defatted milk in TBS-T. Selectedantibody was then added in the blocking solution(1:40,000 or 1:60,000) for 2 hr. After washing,HRP-conjugated goat anti-rabbit IgG (Dako) wasadded at 1:10,000 in blocking solution for 1 hr.Western blots were revealed after washing withchemioluminescence reagents (Chemiglow, TMWEST).
ELISA procedure
The assay was performed in 96-well NuncMaxisorp microtitration plates as follows usingchromatography-purified putative VTG1 or electo-eluted purified putative VTG2.
1. Antigen coating: Purified VTG or lyophilizedVTG-free male plasma (100–400 ng.ml�1 insodium carbonate buffer, 0.05 M, pH 9.6) wascoated overnight at 41C.
2. Pre-incubation: In parallel, samples and serialdilutions of the reference purified VTG (from1000 to 7.81 ng) diluted in PBST-NPS (0.01 MPB, 0.15 M NaCl, 0.05% Tween 20, pH 7.4,containing 4% of normal pig serum) were pre-incubated in separate tubes overnight at 371Cwith specific antibody diluted (1:40,000 to1:160,000) in the same buffer.
3. Saturation: After removing the coating solu-tion, non-specific binding was lowered byadding PBST-NPS for 30 min at 371C.
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 579
J. Exp. Zool. DOI 10.1002/jez.a

4. Washing: After every step of the assay, thewells were extensively washed three timeswith PBS-T.
5. Incubation: Each pre-incubation solution wasdispensed in duplicate into the microplatewells, and PBST-NPS added into the fournon-specific binding wells. After incubationfor 2 hr at 371C, the plates were washed.
6. Second antibody incubation: Each well receivedswine anti-rabbit immunoglobulins (Dako) di-luted in PBST-NPS (1:2,500) during 45 min at371C, then washed.
7. PAP incubation: Each well received PAP com-plexes (Dako) diluted in PBST-NPS (1:5,000)for 45 min at 371C, then washed.
8. Revelation: The peroxidase activity wasrevealed as described for antigen–antibodydilution test.
9. Absorbance measurement: The absorbance ofeach well was measured after 10 min at 492 nmusing a Titertech multiskan plate reader.
The results of absorbance measurements wereexpressed using two models. In the non-linearmodel, results were expressed as Bi/B0� 100 as afunction of log (dose), where Bi represented thebinding of each standard or sample point and B0
the maximum binding obtained when no free VTGwas present in the well. The linear model wasperformed by logit–log transformation (Rodbardand Lewald, ’70) expressed as logit (Bi/B0) orlog (B0�Bi/Bi�N) as a function of the log(dose),where N was the non-specific binding. Theparallelism between regression curves was testedby covariance analysis (Snedecor and Cochran,’89; Sokal and Rohlf, ’96) with the Statgraphics 4.0Software package.
RESULTS
Kinetics of new proteins appearance
Total plasma proteins concentration exhibited ahuge increase, from around 45 mg.ml�1 inthe control plasma (t0) to around 130 mg.ml�1 atthe end of hormonal treatment (t14). SDS-PAGE(7.5% gel) of treated fish plasma (Fig. 1) showedthat the dramatic increase of total proteinswas mainly linked to the emergence of two newbands, of 130 and 170 kDa. They appeared fromday 2 of treatment and became particularlyintense from day 6, corresponding to the third17b-estradiol injection, to the end of the treat-ment (day 14), corresponding to the seventh17b-estradiol injection.
Purification of Nile tilapia VTG
Purification by electro-elution
The two major bands induced by estrogenictreatment were assumed to represent two VTGforms. The electro-eluted fractions submitted toSDS-PAGE (Fig. 2) showed that each purifiedprotein appeared as a single band located atthe same position as in the electrophoregramof 17b-estradiol-treated fish plasma. Both VTG
Fig. 1. SDS-PAGE follow-up of 17b-estradiol treatment.tn: Plasma sample control with n corresponding to thenumber of days of treatment (0–14).
Fig. 2. Electro-eluted VTGs SDS-PAGE. kDa: molecularmass markers; contr; Pl: free-VTG male plasma; Pl1E2:plasma from 17b-estradiol-treated fish, diluted (1:100); VTG1,VTG2: electro-eluted VTGs.
P. NDIAYE ET AL.580
J. Exp. Zool. DOI 10.1002/jez.a

fractions exhibited almost no plasma proteincontamination.
Purification by double chromatography
The elution profile of the gel filtration on 6BSepharose (Fig. 3a) of 17b-estradiol-treated juve-nile male plasma showed three successive peaks,peak 1 corresponding to the void volume. Electro-phoretic control of a selection of eluted fractions(Fig. 3b) indicated that the rising part of peak 2contained a major band of 130 kDa as well as othercontaminating plasma proteins.
Fractions containing the highest amounts ofVTG (27–34) were pooled, concentrated on theAmicon cell (membrane cutoff 50,000 Da) anddeposited on top of a DEAE-M trisacryl ion-exchange column. The elution profile is shown in
Figure 3c. The first peak corresponded to theunbound material eluted in the void volume. Thefollowing peaks (2, 3 and 4) corresponded tothe peaks eluted by application of the linear Trisbuffer gradient.
A selection of eluted fractions were subjected toSDS-PAGE (Fig. 3d). The 130 kDa containingfractions were located in the decreasing partof peak 3, with a maximum intensity observedin fraction 52. Eluted fractions from 50 to 56considered as VTG1 were pooled, concentrated onthe Amicon cell and frozen at �201C.
Purification by single Fractogelion-exchange chromatography
The elution buffer volume recommended by themanufacturer was increased by 100 times in
Fig. 3. (a) Sepharose 6B gel filtration chromatogram. Solid black points correspond to eluted fractions tested on SDS-PAGE.(b): SDS-PAGE of Sepharose 6B gel filtration fractions. Numbers indicate the eluted fractions from Sepharose 6B gel filtrationchromatography. MW: molecular mass markers (kDa); PL1E2: plasma from E2-treated fish. (c): Ion-exchange chromatogram.Solid black points (13, 37, 50, 52, 64) correspond to eluted fractions tested on SDS-PAGE. Gray points (51, 53, 54, 55, 56) andgray point 52 (with between brackets scripts) correspond to vitellogenic eluted fractions pooled to obtain VTG1. (d): SDS-PAGEof DEAE Tris-acryl fractions. Numbers indicate eluted fractions from DEAE Tris-acryl chromatography. MW: molecular massmarkers (kDa); Pl1E2: plasma from 17b-estradiol-treated fish; pool chr.1: pool of gel filtration vitellogenic fractions applied onthe ion-exchange column.
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 581
J. Exp. Zool. DOI 10.1002/jez.a

proportion of the large amount of VTGs induced inestradiol-treated fish. The elution profile (Fig. 4a)revealed three different peaks. The first onecorresponded to the unbound material eluted inthe void volume. The two other well-separatedpeaks were eluted following the application of alinear gradient (0–1 M) NaCl.
The electrophoretic control is shown in Figure 4b.A 130 kDa band was eluted with peak 2 in frac-tion 35 as putative VTG1. It appeared wellisolated with few plasma proteins contamination.A 170 kDa band was eluted in peak 3 and appearedstrongly contaminated. We selected fraction 62 asputative VTG2.
Charaterization of the VTGs
Electrophoretic characterization
Samples of the different VTG preparations wererun simultaneously in SDS-PAGE. Electrophore-tic properties of the two 130 and 170 kDa proteins
remained identical in the presence (lanes b, d, fand h) or in the absence (lanes a, c, e and g) ofb-mercapto-ethanol as reducing agent (Fig. 5).This demonstrated that the heavier 170 kDa bandwas not a dimer of the lighter 130 kDa one as itseemed the case for Salvelinus alpestris VTG(lanes g and h) tested in parallel.
Immunochemical characterization
Two antibodies have been developed in rab-bits, against the 170 kDa (putative VTG2)electro-eluted protein and against 130 kDa (puta-tive VTG1) protein purified by double chromato-graphy (Sepharose 6B and DEAE-Trisacryl).
Characterization by Western blotting. Both anti-bodies have been tested on Western blots. Bothantibodies recognized yolk proteins contained inO. niloticus yolk extract that definitively provestheir yolk origin. They also recognized the twoestradiol-induced new bands exhibited by treatedmales or vitellogenic females (Fig. 6).
Fig. 4. (a): Fractogel ion-exchange chromatography. Solidblack points correspond to eluted fractions tested on SDS-PAGE. Gray points with between brackets numbers corre-spond to VTG1 (fraction 35) and to VTG2 (fraction 62). (b):SDS-PAGE of Fractogel chromatography fractions. kDa:Molecular mass marquers; Pl C: VTG-free male controlplasma; Pl1E2: plasma from 17b-estradiol-treated fish. Num-bers indicate the fraction number of eluted fractions fromFractogel chromatography.
Fig. 5. SDS-PAGE of Oreochromis niloticus (a–f) andSalvelinus alpestris (g, h) electro-eluted VTGs. (a, c, e, g).Without b-mercapto-ethanol; (b, d, f, h) with b-mercapto-ethanol.
Fig. 6. Blots (a) and corresponding Western blots (b)revealed with (A) anti-VTG1 chr. antibody and (B) anti-VTG2 el. antibody. (1) Oreochromis niloticus vitellus extract;(2) plasma from vitellogenic female; (3) plasma from 17b-estradiol-treated fish; (4) VTG-free male control plasma.
P. NDIAYE ET AL.582
J. Exp. Zool. DOI 10.1002/jez.a

Each of two antibodies recognized both forms ofVTG (Fig. 7), VTG1 purified by double chromato-graphy (a) and VTG2 purified by electro-elution(b). Furthermore, the lack of any signal in wildmale plasma, used as negative control, showedthat antibody-harvesting procedures eliminatedthe rabbit gamma globulins directed towardscontaminating plasma proteins. We demonstratedthat each antibody strongly recognized its specificVTG, and the other form.
Characterization by EIA. The good parallelismof logit–log transformed binding curves (Fig. 8)between serial dilutions of O. niloticus VTG1 and
VTG2 purified by DEAE Fractogel, and VTG1
purified by double chromatography, using electro-eluted VTG2 for coating and anti-VTG2 electro-eluted as antibody, demonstrated the cross-reac-tivity between all kinds of VTGs and anti-VTGs.
Same kind of results were obtained with serialdilutions of electro-eluted VTG2, using VTG1
purified by double chromatography for coatingand anti-VTG1 purified by double chromatographyas antibody (results not shown), indicating thatsimilar cross-reactivity existed between the twoVTGs and anti-VTG1.
Homologous ELISA of O. niloticus
ELISA was established using one or the otherantibody obtained, each of them exhibiting a goodrecognition of the two VTGs as well as the vitellusextract components. The specificity was attestedby the parallel curves obtained using anti-VTG1
(Fig. 9) between O. niloticus VTG standard, serialvitellogenic female plasma dilutions and the lackof displacement with VTG-free wild male plasma.
The detection limit was 6 ng.ml�1 after over-night coating with 400 ng.ml�1 induced maleplasma, specific antibody diluted 1:40,000 andincubation time of 2 hr at 371C. This was equiva-lent to 60 ng.ml�1 in plasma, since it has to bediluted 1:10 to avoid any non-specific competition(results not shown). Inter- and intra-assay varia-tions were similar around 12% in 50% of binding.
This ELISA allowed us to measure plasma VTGduring a sexual cycle of wild females duringgonadal development between two successivespawnings (Fig. 10). VTG plasma profile and GSI
Fig. 8. Logit–log tranformation of binding curves usinganti-VTG2 antibody of three different VTG preparations:VTG Chr (VTG1 prepared by DEAE chromatography seeFig. 3c and d), VTG2 FG and VTG1 FG (VTG prepared byDEAE Fractogel chromatography (see Fig. 4a and b).
Fig. 7. Western blots with (a) anti-VTG1 chr. antibody;(b) anti-VTG2 el. antibody. C pl: VTG-free male controlplasma; pl1E2: plasma from 17b-estradiol-treated fish; chr.VTG1: VTG1 obtained by double chromatography; el. VTG2:VTG2 obtained by electro-elution.
Fig. 9. Homologous ELISA for Oreochromis niloticus.Binding curves of VTG standard (VTG1 FG), mature femaleand VTG-free male serial plasma dilutions using anti-VTG1
antibody.
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 583
J. Exp. Zool. DOI 10.1002/jez.a

exhibit similar profiles: they appeared to decreaseslightly during the first 6 days of the mothermouthbrooding (MG), they levelled out from day12 to day 21 during the mother guardian phase (G)and then continuously increased until day 27,at the spawning ending the cycle.
Heterologous ELISAs of various Cichlidae
In order to assess VTG homology betweendifferent tilapia species, serial dilutions of femaleplasma from seven different species within thefour closerelated genera of the Cichlidae Family(Sarotherodon, Oreochromis, Tilapia and Hemi-chromis) have been tested in the O. niloticus VTGELISA system (Fig. 11). Slopes of the serialdilution curves and statistical significance arepresented in Table 1.
Parallelism of regression lines indicated similarinteraction between heterologous VTG andO. niloticus VTG antibody and the different VTGsof the species tested. Only two species (O. aureusand S. melanotheron) gave parallel curves toO. niloticus VTG. From the other species tested,three gave parallel slopes between them(H. fasciatus, H. bimaculatus and Tilapia zilii)but different from O. niloticus VTG. Finally,T. dagetti had a completely unparallel slope withall other species tested. As a negative control,plasma from a wild male of S. melanotheron gaveno displacement (slope 5�0.001).
Histology, electron microscopy andimmunocytochemistry
The data presented here deal with the descrip-tion in O. niloticus of the two Stages 3 and 4, of OFdevelopment reflecting the accumulation of yolk inthe primary oocyte.
The diameters of Stage 3 ranged from 200mm(early) to 600mm (late). At the photonic level(Fig. 12a), the nucleus exhibited numerous nu-cleoli illustrating the amplification of the nuclearorganizer genes, which started during the pre-vious previtellogenic stages. The ooplasm wastypically organized in three concentric rings. Theouter ring was constituted by cortical alveolilocated at the vicinity of numerous Golgi
Fig. 11. Logit–log transformed competition curves ob-tained with serial dilutions of plasma of different vitellogenicfemale tilapia species. O.n.: Oreochromis niloticus; O.a.:Oreochromis aureus; S.m. F: Sarotherodon melanotheronfemale; H.b.: Hemichromis bimaculatus; H.f.: Hemichromisfasciatus; S.m. M: Sarotherodon melanotheron male; T.d.:Tilapia dageti; T.z.: Tilapia zillii.
TABLE 1. Slope comparison of binding curves obtainedwith the serial plasma dilutions of tilapia species
after logit–log transformation
O. n. O. a. S. m. H. b. H. f. T. z. T. d.
O. n. 0.324 0.120 0.000 0.002 0.001 0.000O. a. NS 0.279 0.000 0.000 0.002 0.000S. m. NS NS 0.000 0.000 0.006 0.000H. b. ��� ��� ��� 0.722 0.762 0.000H. f. ��� ��� ��� NS 0.556 0.000T. z. ��� ��� �� NS NS 0.000O. n. ��� ��� ��� ��� ��� ���
P values of paired comparisons (upper triangle), and significance levels(lower triangle). See Fig. 11 for other details.
Fig. 10. Changes in GSI and plasma VTG concentrationduring an inter-spawning interval in wild females of Oreo-chromis niloticus. S: spawning, MB: mouthbrooding period,fr: fry release, G: guarding period. Bars indicate standarddeviation, n 5 10 for all samples.
P. NDIAYE ET AL.584
J. Exp. Zool. DOI 10.1002/jez.a
complexes. The median ring enclosed numerousgranules. The inner ring was constituted by thefirst appearance of lipid globules. At the outersurface of the oocyte, a thin dark layer appearedlocated under the granulosa epithelium (Fig. 12b),due to the onset of deposition of the zonaradiata externa.
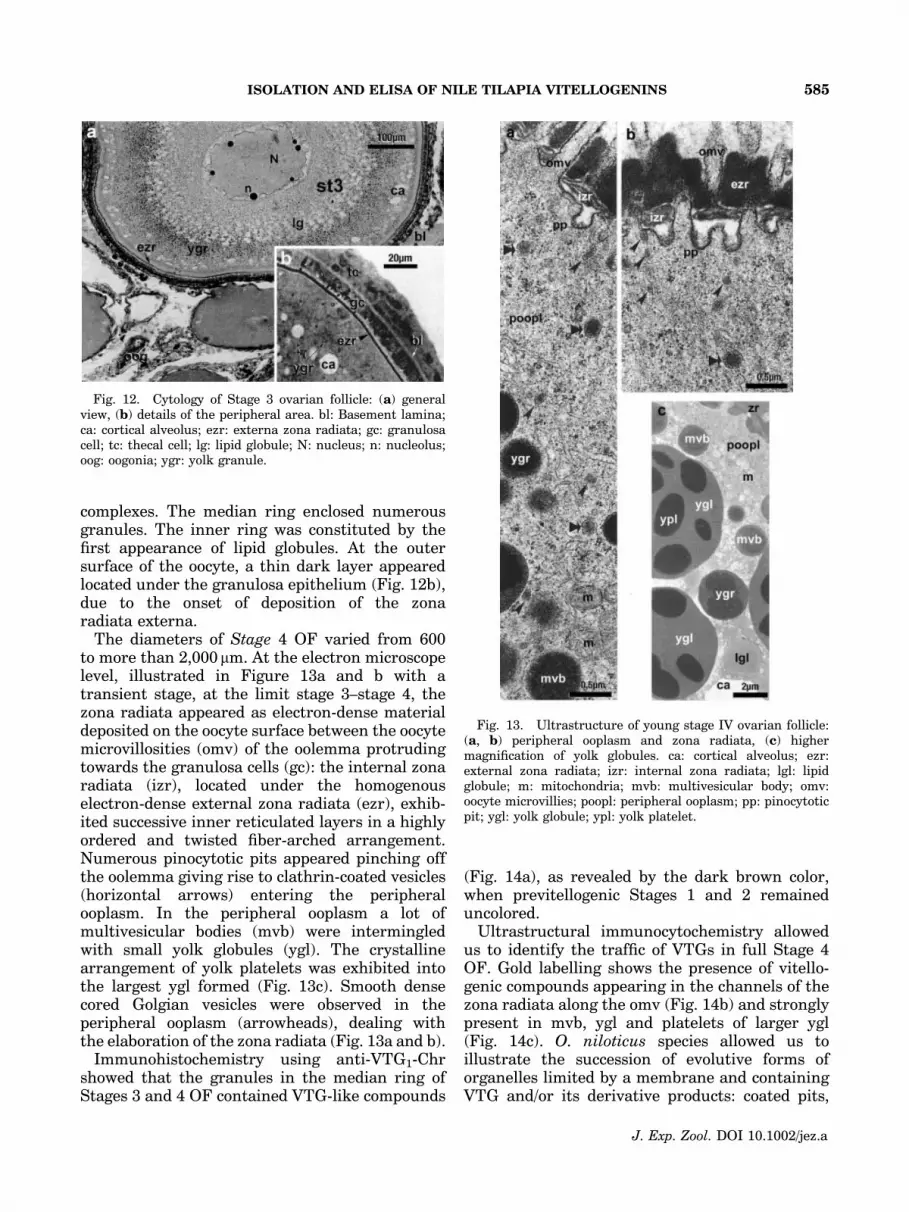
The diameters of Stage 4 OF varied from 600to more than 2,000 mm. At the electron microscopelevel, illustrated in Figure 13a and b with atransient stage, at the limit stage 3–stage 4, thezona radiata appeared as electron-dense materialdeposited on the oocyte surface between the oocytemicrovillosities (omv) of the oolemma protrudingtowards the granulosa cells (gc): the internal zonaradiata (izr), located under the homogenouselectron-dense external zona radiata (ezr), exhib-ited successive inner reticulated layers in a highlyordered and twisted fiber-arched arrangement.Numerous pinocytotic pits appeared pinching offthe oolemma giving rise to clathrin-coated vesicles(horizontal arrows) entering the peripheralooplasm. In the peripheral ooplasm a lot ofmultivesicular bodies (mvb) were intermingledwith small yolk globules (ygl). The crystallinearrangement of yolk platelets was exhibited intothe largest ygl formed (Fig. 13c). Smooth densecored Golgian vesicles were observed in theperipheral ooplasm (arrowheads), dealing withthe elaboration of the zona radiata (Fig. 13a and b).
Immunohistochemistry using anti-VTG1-Chrshowed that the granules in the median ring ofStages 3 and 4 OF contained VTG-like compounds
(Fig. 14a), as revealed by the dark brown color,when previtellogenic Stages 1 and 2 remaineduncolored.
Ultrastructural immunocytochemistry allowedus to identify the traffic of VTGs in full Stage 4OF. Gold labelling shows the presence of vitello-genic compounds appearing in the channels of thezona radiata along the omv (Fig. 14b) and stronglypresent in mvb, ygl and platelets of larger ygl(Fig. 14c). O. niloticus species allowed us toillustrate the succession of evolutive forms oforganelles limited by a membrane and containingVTG and/or its derivative products: coated pits,
Fig. 13. Ultrastructure of young stage IV ovarian follicle:(a, b) peripheral ooplasm and zona radiata, (c) highermagnification of yolk globules. ca: cortical alveolus; ezr:external zona radiata; izr: internal zona radiata; lgl: lipidglobule; m: mitochondria; mvb: multivesicular body; omv:oocyte microvillies; poopl: peripheral ooplasm; pp: pinocytoticpit; ygl: yolk globule; ypl: yolk platelet.
Fig. 12. Cytology of Stage 3 ovarian follicle: (a) generalview, (b) details of the peripheral area. bl: Basement lamina;ca: cortical alveolus; ezr: externa zona radiata; gc: granulosacell; tc: thecal cell; lg: lipid globule; N: nucleus; n: nucleolus;oog: oogonia; ygr: yolk granule.
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 585
J. Exp. Zool. DOI 10.1002/jez.a

then in coated vesicles, mvb, yolk granules, ygland finally large ygl with platelets.
DISCUSSION
VTG purification
In the past two decades, purification of VTGfrom estradiol-induced fish plasma using variouschromatographic techniques was published in alarge number (Mananos et al., ’94b; Bon et al., ’97;Korsgaard and Pedersen, ’98; Mosconi et al., ’98;Parks et al., ’99; Brion et al., 2000; Holbech et al.,2001; Takemura and Kim, 2001; Fukada et al.,
2003; Hennies et al., 2003; Malgalhaes et al.,2004). Analysis of this literature showed thatcombination of chromatographic techniques leadsto good results and encouraged us to use thisapproach.
We chose to undertake the purification of VTGfrom plasma without a preliminary concentrationstep. Indeed, previous reports concerningO. niloticus VTG purification mentioned the pre-cipitation of VTG by EDTA prior to purification byanion-exchange chromatography on DEAE Sepha-dex (Chan et al., ’91) or electro-elution from PAGE(Buerano et al., ’95). This procedure, used toconcentrate the VTG before its purification, was
Fig. 14. Immunocytochemistry: (a) photonic immunocytochemistry of VTG labelling in ovary; (b, c) electron immunocy-tochemistry of VTG labelling in OF. ca: cortical alveolus; ezr: external zona radiata; izr: internal zona radiata; mvb:multivesicular body; omv: oocyte microvillies; poopl: peripheral ooplasm; st: stage; yst: young stage; sloped arrow: pinocytotic pit.
P. NDIAYE ET AL.586
J. Exp. Zool. DOI 10.1002/jez.a

largely used for various teleosts (Norberg andHaux, ’85; Copeland and Thomas, ’88; Ding et al.,’89; Silversand et al., ’93). It is now welldocumented that any change in the plasma, suchas addition of distilled water or EDTA-Mg21,modifies the interactions between VTG andCa21, and disrupts the quaternary structure ofthe VTG, which is then generally precipitated withaggregation of its subunits (Ding et al., ’89). Thefollowing process of redissolving the VTG does notlead, in all the species studied, to the initial nativeprotein tri-dimensional structure (Ansari et al.,’71; De Vlaming et al., ’80; Kanungo et al., ’90;Tyler and Sumpter, ’90).
Gel filtration was performed as the first of a two-steps procedure, taking advantage of the largedifference in electrophoretic mobility between thetwo new estrogen-induced bands as seen on SDS-PAGE. Surprisingly, as shown in Figure 3b, theband of higher molecular mass corresponding toVTG2 did not appear during the elution and wasundetectable on PAGE. This phenomenon has notyet been described in the literature. The hypoth-esis which could be advanced might be that VTG2
remained adsorbed on the Sepharose gel, due to itshydrophobic properties. Nevertheless, VTG1, wellpurified after DEAE chromatography, was used toraise the anti-VTG1 antibody.
In other respects, by operating the two otherpurification techniques—electro-elution and sin-gle DEAE chromatography—we isolated twoforms of VTGs, VTG1 and VTG2, exhibiting thesame electrophoretic migration patterns in SDS-PAGE as in the induced plasma after estradioltreatment.
VTG characterization
Electrophoretic identification
In the recent literature, the existence of eitherone or two forms of VTG in O. niloticus has beendiscussed several times.
In a previous work (Chan et al., ’91), thepresence in female plasma of two native and verydistinct protein bands, Vg and H, was shown, theformer only being identified as VTG. Indeed, theauthors claimed that Vg was the only estrogen-inducible phosphoprotein in O. niloticus, theprotein H being unlabelled when E2-induced fishwere injected with 32P-orthophosphate. Theirapparent molecular weights in native conditionswere respectively around 300 and 400 kDa. Sincein native PAGE the electrophoretic mobility is notlinearly related to the molecular mass (Brion
et al., 2000), these two proteins could be equiva-lent to VTG1 and VTG2 identified in our study as130 and 180 kDa proteins.
Lee et al. (’92), studying the degree of homologybetween VTG from various Perciformes claimedthat O. niloticus VTG appeared in a singlemolecular form under native PAGE conditions,but exhibited two polypeptide forms of 220 and130 kDa under denaturing-reducing conditions.This was in part questioned by Buerano et al.(’95) who have identified in the same species twoforms of VTG both in native and in SDS-PAGEconditions. The two prominent polypeptides,EIpp2 (120 kDa) and EIpp1 (185 kDa), detectedby these authors in denaturing conditions couldcorrespond, respectively, to our VTG1 and VTG2.
The presence of two VTGs was demonstrated inclosely related species. Two VTGs of 130 and180 kDa were shown on SDS-PAGE by Ding et al.(’89) in estrogen-treated O. aureus and confirmedby Lim et al. (’91). Other works (Kishida andSpecker, ’93, ’94) identified by SDS-PAGE, inOreochromis mossambicus, two plasmatic vitello-genic components designated as tVTG-130 andtVTG-200 according to their apparent molecularmasses. In O. mossambicus as well, Takemuraand Kim (2001) identified two VTGs with appar-ent molecular masses of 140 and 210 kDa bySDS-PAGE.
In summary, we conclude that in O. niloticus,as well as in closely related species, two formsof vitellogenic polypeptides exist in maturefemales and in estrogen-treated males: VTG1
and VTG2.
Immunological characterizationby Western blot
We raised two antibodies in rabbits, one againstthe putative VTG1 purified by a two-step chroma-tography and the other against putative VTG2
purified by electro-elution. We harvested the anti-VTG1 antibody with VTG-free plasma from wildmale caught in the Senegal river. Results showedthat the harvesting procedure removed the rabbitgamma globulins directed towards the few plasmaproteins contaminating the purified VTG1.
The two antibodies recognized O. niloticusvitellus extract, which means that 130 and180 kDa proteins are plasma precursors of yolk.Each antibody recognized the two VTGs whateverthe purification protocol used.
Western blot demonstrated that the storage ofthe purified VTGs at �801C and a fortiori at
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 587
J. Exp. Zool. DOI 10.1002/jez.a

�201C lead to a rapid degradation, numerousbands of lower molecular mass being labelled byanti-VTGs on Western blots. The quicker theVTGs are used after their purification, the lessthey are broken down. If experimentations wereperformed within around 24 hr following VTGsisolation, results would be the same with orwithout the use of anti-proteolytic agents duringthe VTGs purification. Nevertheless, we haveobserved that VTGs purified by electro-elutionappeared less degradable than VTGs purified bychromatography. This observation was cleared uptaking into account the presence of a trypsin-likeprotease in the plasma of O. niloticus (Inaba et al.,’97) whose activity is strongly inhibited byleupeptin and aprotinin: during isolation of pro-teins by electrophoresis which is quicker than bychromatography, the time of contact of VTGs withprotease is shorter than during isolation bychromatography, and the action of the trypsin-like protease is lower.
Immunological characterization by EIA
The cross-reactivity previously demonstratedbetween the two antibodies and their correspond-ing antigens was largely confirmed by crossed EIAs.
In contrast with previous data, our results showthat both antibodies cross-react with each VTGform. It has been shown (Chan et al., ’91) that anantibody directed against the purified Vg (corre-sponding to VTG1) did not recognize the heavierband H (corresponding to VTG2) induced at thesame time by estrogen treatment of males.Similarly, Buerano et al. (’95) found that the twovitellogenic polypeptides (EIpp1 and EIpp2) wereimmunochemically different. We may concludefrom those diverging results that each of ourpolyclonal antibodies recognize a larger diversityof epitopes, including some shared by both formsof VTG.
Use of homologousand heterologous ELISAs
One or the other of the two specific antibodiesrecognizing the two forms of VTG present in thisspecies can be used to quantify by ELISA the totalplasma VTG of O. niloticus. The O. niloticus VTGELISA exhibited high specificity and sensitivityand was comparable to other ELISAs developedfor other fresh water or marine fish species, Soleavulgaris (Nunez Rodriguez et al., ’89), Acipenserbaeri (Cuisset et al., ’89), Dicentrarchus labrax(Mananos et al., ’94a), Onchorhynchus mykiss
(Mourot and Le Bail, ’95; Bon et al., ’97), Zoarcesviviparous (Korsgaard and Pedersen, ’98), Sparusaurata (Mosconi et al., ’98), Pimephales promelas(Parks et al., ’99), Salmo trutta (Sherry et al., ’99),Danio rerio (Holbech et al., 2001), Cyprinus carpio(Hennies et al., 2003), Acanthogobius flavimanus(Ohkubo et al., 2003), Lepomis macrochirus(Cheek et al., 2004), C. carpio, D. rerio and Oryziaslatipes (Nilsen et al., 2004; Nozaka et al., 2004).
We have used this ELISA to quantify plasmaVTG during wild female sexual cycle, i.e., duringthe inter-spawning period. We have shown thatthe plasma VTG profile was in correlation with theGSI, reflecting itself the development of thevitellogenic oocytes batch spawned at the end ofthe cycle (Tacon et al., ’96). All the data obtainedunderlined that the MG and guard of the fry bythe female play an inhibitory role upon ovariandevelopment (Tacon et al., 2000).
This assay might be used to quantify VTG in twoother species (S. melanotheron and O.aureus),since plasma dilution curves of heterologousELISAs were parallel to O. niloticus VTG stan-dard curve, indicating that the VTG structure inthese three species was similar. Nevertheless, thelack of parallelism between O. niloticus and theother species tested is not a damning result, andthe presence of VTG in males or fry can bequalitatively tested in order to denounce thepresence of xeno-estrogens in the environment.
Similar heterologous immunoreactions havebeen shown in other fish families. A carp anti-VTG recognized VTG of common carp (C. carpio),roach (Rutilis rutilis) and bream (Abramis brama)using RIA (Tyler and Sumpter, ’90). Anothercommon carp anti-VTG used in ELISA recognizedVTG of fathead minnow (P. promelas) (Mylchreestet al., 2003). More recently, both anti-rainbowtrout (O. mykiss) VTG and anti-masu salmon(O. masu) VTG also recognized cutthroat trout(O. clarki), white-spotted char (Salvelinus leuco-maensis) and Sakalin taimen (Hucho perryi) VTGs(Fukada et al., 2001).
Endocytosis of VTG into the OF
We have traced the uptake of VTG by the OFduring vitellogenesis at the photonic and electro-nic microscope level using VTG immunodetectionwith anti-VTG1 antibody. An ultrastructuralapproach confirms in this species that Stage 3 ischaracterized by the onset of oocyte vitellogenesis,where VTG starts to enter the OF at the sametime when lipid globules and cortical alveoli
P. NDIAYE ET AL.588
J. Exp. Zool. DOI 10.1002/jez.a

appear (Le Menn et al., 1999). Once again, weobserved in this fish that initiation of yolkaccumulation into the oocyte was concomitantwith deposition of the zona radiata externa. Atthis stage, yolk is present in the oocyte, but lipidaccumulation is largely predominant over VTGendocytosis: type I vitellogenesis. In the followingStage 4, the yolk components are the same as inStage 3, but the proportion of lipid globules andygl is inverted and characterizes type II vitello-genesis. It corresponds to the rapid growth phaseof the OF development, where endocytosis of VTGis considerable and during which the zona radiatainterna is deposited.
These observations concerning O. niloticus arein agreement with those previously described inother teleostean or chondrichtyan species: Fundu-lus heteroclitus (Wallace and Selman, ’81; Selmanand Wallace, ’83, ’86), Salmo trutta (Breton et al.,’83), Solea vulgaris (Nunez Rodriguez, ’85), D.labrax (Zanuy et al., ’89), Acipenser baeri (Cuissetet al., ’89) and Gobius niger (Le Menn et al., ’99).
Our ultrastructural approach coupled with im-munocytochemistry detection of the VTG allowsus to claim that in all species studied the yolkentering the oocyte occurs earlier than photonichistology leads to believe.
What gene(s) code(s) for VTGsin O. niloticus?
In animals, VTGs are members of the large lipidtransfer protein superfamily, which could arisefrom an ancestral gene ensuring a pivotal eventin the intra-cellular and extra-cellular transfer oflipid and liposoluble substances (Babin et al., ’99).Recent results have demonstrated the presence ofmultiple VTG genes (vtg) in some teleost fishspecies (Buisine et al., 2002; Wang et al., 2005).Comparative genomic analyses suggest two wholegenome duplication events occurring one beforeand one after the divergence of ray-finned andlobe-finned fishes, prior to the teleost radiation(Jaillon et al., 2004; Van de Peer, 2004; Dehal andBoore, 2005; Sawaguchi et al., 2005; Woods et al.,2005). In teleost fish, the current model for vtgamplification has been demonstrated in Salmoni-dae, where ancestral vtg duplications gave rise totwo paralogous gene clusters, VtgA and VtgB(Buisine et al., 2002). Salmonids harboring a lowvtg copy number reflect an ancestral state,whereas salmonids harboring a high vtg copynumber reflect the latest state of differentiation.The ultimate state of differentiation appears in all
Oncorhynchus species which have lost, in thecourse of evolution, VtgB-related genes, andexhibit a unique VtgA locus containing a set of20 highly similar genes tandemly arranged (Tri-chet et al., 2000). The zebrafish (D. rerio) genomecontains at least seven vtg encoding heterologousVTGs with three distinct types mapped onto twodifferent chromosomes. Northern blot resultsshow that all seven vtg were expressed predomi-nantly in the liver and adipose tissue of female fishor E2-induced males (Wang et al., 2005). In thisspecies, lipovitellin I, phosvitin and lipovitellin II,parts of the yolk derivative proteins (Wahli, ’88),are encoded by vtg1 (vg1) type, which is mosthighly expressed.Two vtg transcripts have beencloned in F. heteroclitus (LaFleur et al., ’95;GenBank acession number UO7055) in Oryziaslatipes (Tong et al., 2004; GenBank accessionnumbers: AB064320 and AB074891) and inAcanthogobius flavimanus (Ohkubo et al., 2003).Multiple vtg transcripts have been demonstratedin Gambusia affinis (Sawaguchi et al., 2005). A vtgtranscript was characterized from the tilapia,O. aureus (Lim et al., 2001), and the structureof the corresponding gene determined (Teo et al.,’98). However, in a previous study (Lee et al., ’94),the authors found the presence of twomajor groups of OaVg cDNA exhibiting a 7%intra-specific divergence in the vtg family.Takenaltogether, these results support the presence ofat least two vtg-related genes in the genome ofOreochromis genus, in accordance with the char-acterization of two forms of plasma VTG in O.aureus (Ding et al., ’89), in O. mossambicus(Kishida and Specker, ’93; Takemura and Kim,2001) and in O. niloticus (this study).
CONCLUSION
In this paper we have clearly identified twoforms of VTG in O. niloticus by SDS-PAGE. Thesetwo proteins have apparent molecular masses of130 kDa (VTG1) and 170 kDa (VTG2). They arepresent in mature female or in estrogen-treatedfish plasma.
We have raised two antibodies against each formof VTG. Using EIA and Western blotting, wehave demonstrated the complete cross-reactivityof these antibodies with both forms of VTG. Eachof them is able to recognize each of the twoplasma VTGs.
With anti-VTG1, using photonic and electronimmunocytochemistry, we have confirmed on thisspecies that oocyte yolk accumulation is initiated
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 589
J. Exp. Zool. DOI 10.1002/jez.a

at OF Stage 3, as soon as the oocyte synthesis ofcortical alveoli occurs and lipid globules appear inthe ooplasm. On this species, as for numerousprevious ones, we have seen that initiation ofoocyte vitellogenesis is concomitant with firstdeposition of the zona radiata.
Using one or the other antibody, we havedeveloped specific ELISAs with high sensitivityallowing in the plasma the detection of the VTGsas a whole. We have used the ELISA to follow thevariation of plasma VTG during wild femalesexual cycle and demonstrated that VTG profilewas perfectly correlated with the evolution ofthe GSI, with a pause during the maternal MGand guard.
We demonstrated that using O. niloticus VTG ascoating and O. niloticus anti-VTG as antibody,this ELISA might be used for several other relatedspecies to quantify (O. aureus and S. melanother-on) or detect (H. fasciatus, H. bimaculatus andT. zillii) plasma VTG. These heterologous ELISAsconstitute a reliable tool to monitor the presenceof xeno-estrogens in the environment of thesefish species. Indeed, recent studies show thatO. niloticus (Leanos-Castaneda et al., 2002; Par-ente et al., 2004) as well as other Cichlidae(Okoumassoun et al., 2002) may be submitted toxenobiotic aggressions in their natural habitat asknown for many other fish species (Sumpter, ’98;Kime et al., ’99; Hutchinson and Pickford, 2002).
ACKNOWLEDGMENTS
We wish to thank Pr. Patrick Babin (Labor-atoire de Genomique et Physiologie des Poissons,Universite de Bordeaux I) for critical readingof the manuscript and for granting access to hislab, Christophe Cochet from the IRD-GAMET forhelp with VTG induction and Pr. Patrick Cottin(ISTAB, Universite de Bordeaux I) for helpfuladvices on chromatrography.
LITERATURE CITED
Alves MM, Dos Santos HS, Lopes RA, Petnusci SO, HaiyashiC. 1983. Rhythm of development in the oocyte of the tilapiaOreochromis niloticus L. (Pisces: Cichlidae); a morpho-metric and histochemical study. Gegenbaurs Morph Jahrb129:575–592.
Ansari AQ, Dolphin PJ, Lazier CB, Munday KA, Akhtar M.1971. Chemical composition of an œstrogen-induced cal-cium-binding glycolipophosphoprotein in Xenopus laevis.Biochem J 122:107–113.
Babiker MM, Ibrahim H. 1979. Studies on the biology ofreproduction in the Cichlid Tilapia nilotica L.: gonadalmaturation and fecundity. J Fish Biol 14:437–448.
Babin PJ, Bogerd J, Kooiman FP, Van der Horst DJ. 1999.Apollipophorin II/I, apolipoprotein B, vitellogenin, andmicrosomal triglyceride transfer protein genes are derivedfrom a common acestor. J Mol Evol 49:150–160.
Baker JR. 1945. Cytological technique: the principles andpractice of methods used to determine the structure of themetozoan cell. London: Methuen & Co. 211p.
Baroiller JF, Jalabert B. 1989. Contribution of research inreproductive physiology to the culture of tilapias. AquatLiving Resour 2:105–116.
Baroiller JF, Desprez D, Carteret Y, Tacon P, Borel F,Hoareau MC, Melard C, Jalabert B. 1997. In: FitzsimmonsK, editor. Influence of environmental and social factors onthe reproductive efficiency in three tilapia species, Oreo-chromis niloticus, O. aureus, and the red tilapia (red Floridastrain). Orlando, FL. p 238–252.
Bon E, Barbe U, Nunez Rodriguez J, Cuisset B, Pelissero C,Sumpter J, Le Menn F. 1997. Serum vitellogenin levelsduring the annual reproductive cycle of the female rainbowtrout (Onchorhynchus mykiss): establishment and valida-tion of an ELISA. Comp Biochem Physiol A 117:75–84.
Breton B, Fostier A, Zohar Y, Le Bail PY, Billard R. 1983.Gonadotropine glycoproteique maturante et œstradiol-17beta pendant le cycle reproducteur chez la truite Fario(Salmo trutta) femelle. Gen Comp Endocrinol 49:220–231.
Brion F, Rogerieux F, Noury P, Migeon B, Flammarion P,Thybaud E, Porcher J-M. 2000. Two-step purificationmethod of vitellogenin from three teleost fish species:rainbow trout (Oncorhynchus mykiss), gudgeon (Gobiogobio) and chub (Leuciscus cephalus). J Chrom B 737:3–12.
Buerano CC, Inaba K, Natividad FF, Morisawa M. 1995.Vitellogenins of Oreochromis niloticus: identification, isola-tion, and biochemical and immunochemical characteriza-tion. J Exp Zool 273:59–69.
Buisine N, Trichet V, Wolff J. 2002. Complex evolution ofvitellogenin genes in salmonid fishes. Mol Genet Genomics268:535–542.
Chan SL, Tan CH, Pang MK, Lam TJ. 1991. Vitellogeninpurification and development of assay for vitellogeninreceptor in oocyte membranes of the tilapia (Oreochromisniloticus, Linnaeus 1766). J Exp Zool 257:96–109.
Cheek AO, King WV, Burse JR, Borton DL, Sullivan CV. 2004.Bluegill (Lepomis macrochirus) vitellogenin: purificationand enzyme-linked immunosorbent assay for detection ofendocrine disruption by papermill effluent. Comp BiochemPhysiol C 137:249–260.
Copeland PA, Thomas P. 1988. The measurement of plasmavitellogenin levels in a marine teleost, the spotted seatrout(Cynoscion nebulosus) by homologous radioimmunoassay.Comp Biochem Physiol B 91:17–23.
Cuisset B, Pelissero C, Le Menn F, Nunez Rodriguez J. 1989.In: Williot P, editor. ELISA for Siberian sturgeon (Acipenserbaeri B.) vitellogenin. Bordeaux, France: CEMAGREF.p 107–111.
Dadzie S. 1974. Stades de developement de Tilapia mossam-bica. Ghana J Sci 14:23–31.
Davail B, Pakdel F, Bujo H, Perazzolo LM, Waclawek M,Schneider W, Le Menn F. 1998. Evolution of oogenesis: thereceptor for vitellogenin from the rainbow trout. J Lipid Res39:1929–1937.
De Vlaming VL, Wiley HS, Delahunty G, Wallace RA. 1980.Goldfish (Carassius auratus) vitellogenin: induction, isola-tion, properties and relationship to yolk proteins. CompBiochem Physiol B 67:613–623.
P. NDIAYE ET AL.590
J. Exp. Zool. DOI 10.1002/jez.a

Dehal P, Boore JL. 2005. Two rounds of whole genomeduplication in the ancestral vertebrate. PLoS Biol 3:e314.
Ding JL, Hee PL, Lam TJ. 1989. Two forms of vitellogenin inthe plasma and gonads of male Oreochromis aureus. CompBiochem Physiol 93:363–370.
Duponchelle F, Cecchi P, Corbin D, Nunez J, Legendre M.1999. Spawning season variations of female Nile Tilapia,Oreochromis niloticus, from man-made lakes of Coted’Ivoire. Environ Biol Fishes 56:375–387.
Duponchelle F, Cecchi P, Corbin D, Nunez J, Legendre M.2000. Variations in fecundity and egg size of female Niletilapia, Oreochromis niloticus, from man-made lakes of Coted’Ivoire. Envirom Biol Fishes 57:155–170.
Fishelson L, Yaron Z. 1983. In: Fishelson L, Yaron Z, editors.Steroid hormones in two Tilapia species: Oreochromisaureus and Oreochromis niloticus. Nazareth, Israel: TelAviv University. p 67–80.
Francolini M, Lora Lamia C, Bonsignorio D, Cotelli F. 2003.Oocyte development and egg envelope formation in Oreo-chromis niloticus, a mouth-brooding cichlid fish. J Sub-microsc Cytol Pathol 35:49–60.
Fukada H, Haga A, Fujita T, Hiramatsu N, Sullivan CV, HaraA. 2001. Development and validation of chemiluminescentimmunoasay for vitellogenin in five salmonid species. CompBiochem Physiol A 130:163–170.
Fukada H, Fujiwara Y, Takahashi T, Hiramatsu N, SullivanCV, Hara A. 2003. Carp (Cyprinus carpio) vitellogenin:Purification and development of a simultaneous chemilu-minescent immunoassay. Comp Biochem Physiol A 134:615–623.
Hennies M, Wiesmann M, Allner B, Sauerwein H. 2003.Vitellogenin in carp (Cyprinus carpio) and perch (Percafluviatilis): purification, characterization and developmentof an ELISA for the detection of estrogenic effects. Sci TotEnviron 309:93–103.
Hines GA, Boots LR, Wibbels T, Watts A. 1999. Steroid levelsand steroid metabolism in relation to early gonadal devel-opment in the tilapia Oreochromis niloticus (Teleostei:Cyprinoidei). Gen Comp Endocrinol 114:235–248.
Hiramatsu N, Chapman RW, Lindzey JK, Haynes MR,Sullivan CV. 2004. Molecular characterization and expres-sion of vitellogenin receptor from white perch (Moroneamericana). Biol Reprod 70:1720–1730.
Holbech H, Andersen L, Petersen GI, Korsgaard B, PedersenKL, Bjerregaard P. 2001. Development of an ELISA forvitellogenin in whole body homogenate of zebrafish (Daniorerio). Comp Biochem Physiol C 130:119–131.
Hutchinson TH, Pickford DB. 2002. Ecological risk assess-ment and testing for endocrine disruption in the aquaticenvironment. Toxicology 181–182:383–387.
Inaba K, Buerano CC, Natividad FF, Morisawa M. 1997.Degradation of vitellognins by 170 kDa Trypsin-likeprotease in the plasma of the tilapia, Oreochromis niloticus.Comp Biochem Physiol B 118:85–90.
Jaillon O, Aury JM, Brunet F, Petit JL, Stange-Thomann N,Mauceli E, Bouneau L, Fischer C, Ozouf-Costaz C, Bernot A,Nicaud S, Jaffe D, Fisher S, Lutfalla G, Dossat C, SegurensB, Dasilva C, Salanoubat M, Levy M, Boudet N, CastellanoS, Anthouard V, Jubin C, Castelli V, Katinka M, Vacherie B,Biemont C, Skalli Z, Cattolico L, Poulain J, De Berardinis V,Cruaud C, Duprat S, Brottie P, Coutanceau JP, Gouzy J,Parra G, Lardier G, Chapple C, McKernan KJ, McEwan P,Bosak S, Kellis M, Volff JN, Guigo R, Zody MC, Mesirov J,Lindblad-Toh K, Birren B, Nusbaum C, Kahn D, Robinson-
Rechavi M, Laudet V, Schachter V, Quetier F, Saurin W,Scarpelli C, Wincker P, Lander ES, Weissenbach J, RoestCrollius H. 2004. Genome duplication in the teleost fishTetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 431:946–957.
Kanungo J, Petrino TR, Wallace RA. 1990. Oogenesis inFundulus heteroclitus. VI. Establishment and verification ofconditions for vitellogenin incorporation by oocytes in vitro.J Exp Zool 254:313–321.
Kestemont P, Petrino TR, Falter U. 1989. Les methodes deproduction d’alevins de Tilapia nilotica. FAO. Report no.ADCP/REP/89/46.
Kime DE, Nash JP, Scott AP. 1999. Vitellogenesis as abiomarker of reproductive disruption by xenobiotics. Aqua-culture 177:345–352.
Kishida M, Specker JL. 1993. Vitellogenin in tilapia (Oreo-chromis mossambicus): induction of two forms by estradiol,quantification in plasma and characterization in oocyteextract. Fish Physiol Biochem 12:171–182.
Kishida M, Specker JL. 1994. Vitellogenin in the surfacemucus of tilapia (Oreochromis niloticus): possibility foruptake by the free-swimming embryos. J Exp Zool 268:259–268.
Korsgaard B, Pedersen KL. 1998. Vitellogenin in Zoarcesviviparus: purification, quantification by ELISA and induc-tion by estradiol-17b and 4-nonylphenol. Comp BiochemPhysiol C 120:159–166.
Laemmli UK. 1970. Cleavage of structural proteins duringassembly of the head of bacteriophage T4. Nature 227:680–685.
LaFleur JGJ, Byrne BM, Kanungo J, Nelson LD, GreenbergRM, Wallace RA. 1995. Fundulus heteroclitus vitellogenin:the deduced primary structure of a piscine precursor tononcrystalline, liquid-phase yolk protein. J Mol Evol 41:505–521.
Le Menn F, Davail B, Pelissero C, Ndiaye P, Bon E, PerazzoloL, Nunez Rodriguez J. 1999. In: Norberg B, Kjesbu O,Taranger G, Andersson E, Stefansson S, editors. A newapproach to fish oocyte vitellogenesis, July 4–7, Bergen,Norway. p 281–284.
Leanos-Castaneda O, Van der Kraak G, Lister A, Sima-Alvarez R, Gold-Bouchot G. 2002. o,p0-DDT induction ofvitellogenesis and its inhibition by tamoxifen in Nile tilapia(Oreochromis niloticus). Marine Environ Res 54:703–707.
Lee BH, Lim EH, Lam TJ, Ding JL. 1994. Two major groups ofvitellogenin cDNA clones from Oreochromis aureus (Stein-dachner). Biochem Mol Biol Int 34:75–83.
Lee KBH, Lim EH, Lam TJ, Ding JL. 1992. Vitellogenindiversity in the Perciformes. J Exp Zool 264:100–106.
Lim EH, Ding JL, Lam TJ. 1991. Estradiol-induced vitello-genin gene expression in a teleost fish, Oreochromis aureus.Gen Comp Endocrinol 82:206–214.
Lim EH, Teo BY, Lam TJ, Ding JL. 2001. Sequencee analysisof a fish vitellogenin cDNA with a large phosvitin domain.Gene 277:175–186.
Malgalhaes I, Ledrich M-L, Pihan J-C, Falla J. 2004. One-step,non-denaturing purification method of carp (Cyprinuscarpio) vitellogenin. J Chrom B 799:87–93.
Mananos E, Nunez J, Zanuy S, Carrillo M, Le Menn F. 1994a.Sea bass (Dicentrarchus labrax L.) vitellogenin. II—Valida-tion of an enzyme-linked immunosorbent assay (ELISA).Comp Biochem Physiol B 107:217–223.
Mananos E, Zanuy S, Le Menn F, Carrillo M, Nunez J. 1994b.Sea bass (Dicentarchus labrax L.) vitellogenin. I—Induction,
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 591
J. Exp. Zool. DOI 10.1002/jez.a

purification and partial characterization. Comp BiochemPhysiol B 107:205–216.
Mosconi G, Carnevali O, Carletta R, Nabissi M, Polzonetti-Magni AM. 1998. Gilthead seabream (Sparus aurata)vitellogenin: purification, partial characterization, andvalidation of an enzyme-linked immunosorbent assay(ELISA). Gen Comp Endocrinol 110:252–261.
Mourot B, Le Bail PY. 1995. Enzyme-linked immunosorbentassay (ELISA) for rainbow trout (Oncorhynchus mykiss)vitellogenin. J Immunoassay 16:365–377.
Mylchreest E, Snajdr S, Korte JJ, Ankley GT. 2003.Comparison of ELISAs for detecting vitellogenin in thefathead minnow (Pimephales promelas). Comp BiochemPhysiol C 134:251–257.
Nakamura M, Specker JL, Nagahama Y. 1993. Ultrastructuralanalysis of the developing follicle during early vitellogenesisin tilapia, Oreochromis niloticus, with special reference tothe steroid-producing cells. Cell Tissue Res 272:33–39.
Nilsen B, Berg K, Eidem J, Kristiansen S, Brion F, Porcher J,Goksøyr A. 2004. Development of quantitative vitellogenin-ELISAs for fish test species used in endocrine disruptorscreening. Anal Bioanal Chem 378:621–633.
Norberg B, Haux C. 1985. Induction, isolation and character-ization of the lipid content of plasma vitellogenin from twoSalmo species: Rainbow trout (Salmo gairdneri) and seatout (Salmo trutta). Comp Biochem Physiol B 81:869–876.
Nozaka T, Abe T, Matsuura T, Sakamoto T, Nakano N, MaedaM, Kobayashi K. 2004. Development of vitellogenin assayfor endocrine disrupters using medaka (Oryzias latipes).Environ Sci 11:99–121.
Nunez Rodriguez J. 1985. Hepatic synthesis and plasma levelsof vitellogenin during the sexual cycle of the sole, Soleavulgaris, Barcelona, Spain. 23p.
Nunez Rodriguez J, Kah O, Geffard M, Le Menn F. 1989.Enzyme-linked immunosorbent assay (ELISA) for solevitellogenin. Comp Biochem Physiol B 92:741–746.
Nunez Rodriguez J, Bon E, Le Menn F. 1996. Vitellogeninreceptors during vitellogenesis in rainbow trout, Oncor-hynchus mykiss. J Exp Zool 274:163–170.
Ohkubo N, Mochida K, Adachi S, Hara A, Hotta K, NakamuraY, Matsubara T. 2003. Development of enzyme-linkedimmunosorbent assays (ELISAs) for two forms of vitello-genin in Japanese common goby (Acanthogobius flavima-nus). Gen Comp Endocrinol 131:353–364.
Okoumassoun L-E, Brochu C, Deblois C, Akponan S, MarionM, Averill-Bates D, Denizeau F. 2002. Vitellogenin in tilapiamale fishes exposed to organochlorine pesticides in OuemeRiver in republic of Benin. Sci Tot Environ 299:163–172.
Parente TEM, De-Oliveira ACAX, Silva IB, Araujo FG,Paumgartten FJR. 2004. Induced alkoxyresorufin-O-deal-kylases in tilapias (Oreochromis niloticus) from Guanduriver, Rio de Janeiro, Brazil. Chemosphere 54:1613–1618.
Parks LG, Cheek AO, Denslow ND, Heppell SA, McLachlan JA,LeBlanc GA, Sullivan CV. 1999. Fathead minnow (Pime-phales promelas) vitellogenin: Purification, characterizationand quantitative immunoassay for the detection of estro-genic compounds. Comp Biochem Physiol C 123:113–125.
Philipart JC, Ruwet JC. 1982. In: Pullin RSV, Lowe-McConnell RH, editors. Ecology and distribution of Tilapias.Manila, Philippines: ICLARM. p 15–59.
Pullin RSV, Lowe-Mc Connell RH. 1982. In: Pullin RSV,Lowe-Mc Connell RH, editors. General discussion on thebiology and culture of tilapia. Manila, Philippines: ICLARM.p 3–13.
Rodbard D, Lewald JE. 1970. Computer analysis of radioli-gand assay and radioimmunoassay data. Acta EndocrinolSuppl 147:79–103.
Sawaguchi S, Koya Y, Yoshizaki N, Ohkubo N, Andoh T,Hiramatsu N, Sullivan CV, Hara A, Matsubara T. 2005.Multiple vitellogenins (Vgs) in mosquitofish (Gambusiaaffinis): identification and characterization of three func-tional Vg genes and their circulating and yolk proteinnproducts. Biol Reprod 72:1045–1060.
Selman K, Wallace RA. 1983. Oogenesis in Fundulus hetero-clitus. III. Vitellogenesis. J Exp Zool 226:441–457.
Selman K, Wallace RA. 1986. Gametogenesis in Fundulusheteroclitus. Am Zool 26:173–192.
Sherry J, Gamble A, Fielden M, Hodson P, Burnison B,Solomon K. 1999. An ELISA for brown trout (Salmo trutta)vitellogenin and its use in bioassays for environmentalestrogens. Sci Tot Environ 225:13–31.
Silversand C, Hyllner SJ, Haux C. 1993. Isolation, immuno-chemical detection, and observations of the instability ofvitellogenin from four teleosts. J Exp Zool 267:587–597.
Snedecor GW, Cochran WG. 1989. Statistical methods. Ames:Iowa State University Press. 491p.
Sokal R, Rohlf F. 1996. Biometry. New York: W. H. Freemanand Co. 887p.
Stifani S, Le Menn F, Nunez Rodriguez J, Schneider WJ. 1990.Regulation of oogenesis: the piscine receptor for vitellogen-in. Biochem Biophys Acta 1045:271–279.
Sumpter JP. 1998. Xenoendocrine disrupters - environmentalimpacts. Toxicol Lett 102–103:337–342.
Tacon P, Fostier A, Le Bail PY, Prunet P, Jalabert B. 1995.Relations between maternal behaviour, ovarian develop-ment, and endocrine status, in the mouthbrooding female ofOreochromis niloticus. In: Goetz FWTP, editor. Austin, TX:University of Texas, Austin. 276p.
Tacon P, Ndiaye P, Cauty C, Le Menn F, Jalabert B. 1996.Relationships between the expression of maternal behaviourand ovarian development in the mouthbrooding cichlid fish(Oreochromis niloticus). Aquaculture 146:261–275.
Tacon P, Baroiller J-F, Le Bail PY, Prunet P, Jalabert B. 2000.Effect of egg deprivation on sex steroids, gonadotropin,prolactin, and growth hormone profiles during the repro-ductive cycle of the mouthbrooding cichlid fish (Oreochromisniloticus). Gen Comp Endocrinol 117:54–65.
Takemura A, Kim BH. 2001. Effects of estradiol-17(beta)treatment on in vitro and in vivo synthesis of two distinctvitellogenins in tilapia. Comp Biochem Physiol A 129:641–651.
Teo BY, Tan NS, Lim EH, Lam TJ, Ding JL. 1998. A novelpiscine vitellogenin gene: structural and functional analysesof estrogen-inducible promoter. Mol Cell Endocrinol 146:103–120.
Tong Y, Shan T, Poh YK, Yan T, Wang H, Lam SH, Gong Z.2004. Molecular cloning of zebrafish and medaka vitello-genin genes and comparision of their expression in responseto 17b-estradiol. Gene 328:25–36.
Trichet V, Buisine N, Mouchel N, Moran P, Pendas AM, LePennec JP, Wolff J. 2000. Genomic analysis of thevitellogenin locus in rainbow trout (Oncorhynchusmykiss) reveals a complex history of gene amplifica-tion and retroposon activity. Mol Gen Genomics 263:828–837.
Tyler CR, Sumpter JP. 1990. The development of a radio-immunoassay for carp, Cyprinus carpio, vitellogenin. FishPhysiol Biochem 8:129–140.
P. NDIAYE ET AL.592
J. Exp. Zool. DOI 10.1002/jez.a

Van de Peer Y. 2004. Tetraodon genome confirms Takifugufindings: most fish are ancient polyploids. Genome Biol5:250.
Wahli W. 1988. Evolution and expression of vitellogenin genes.Trends Genet 4:227–232.
Wallace RA, Selman K. 1981. Cellular and dynamic aspects ofoocyte growth in teleosts. Am Zool 21:325–343.
Wang H, Tan JTT, Emelyanov A, Korzh V, Gong Z.2005. Hepatic and extrahepatic expression of vitello-genin genes in the zebrafish, Danio rerio. Gene 356:91–100.
Woods IG, Wilson C, Friedlander B, Chang P, Reyes DK, NixR, Kelly PD, Chu F, Postlethwait JH, Talbot WS. 2005. Thezebrafish gene map defines ancestral vertebrate chromo-somes. Genome Res 15:1307–1314.
Young JA, Muir JF. 2002. Tilapia: Both fish and fowl? MarResour Econ 17:163–173.
Zanuy S, Le Menn F, Carrillo M, Nunez Rodriguez J. 1989. In:Carrillo M, Zanuy S, Planas J, editors. Ultrastructuralaspect of sea-bass (Dicentrarchus labrax L.) follicles duringvitellogenesis. Almunecar, Granada, Spain: Universitat deBarcelona. 77p.
ISOLATION AND ELISA OF NILE TILAPIA VITELLOGENINS 593
J. Exp. Zool. DOI 10.1002/jez.a





























