Embed Size (px)
Citation preview

Title: Measuring developmental noise in bilateral traits.
Authors and Institutions:
Darryl J. Holman1, Laura L. Newell-Morris1, Carrie M. Kuehn, Christina M. Giovas1,
Robert E. Jones3, Ann Streissguth2
1 Department of Anthropology
2 Department of Psychiatry and Behavioral Science
University of Washington
Seattle, WA 98195 USA
3 Center for Demography and Ecology
University of Wisconsin-Madison
Madison, WI 53706 USA
Total Number of Pages: 33 Number of Figures: ? Number of Tables: ? Number of Footnotes: ? Number of References: ? Abbreviated title: Measuring developmental noise
Send Proof to: Darryl Holman Department of Anthropology University of Washington Box 353100 Seattle, WA 98195 USA Phone: (206) 543-7586 Email: [email protected]
Keywords: Fluctuating asymmetry, Directional asymmetry, Antisymmetry, measurement error,
developmental homeostasis.

2
Introduction
The body plan of most animals includes a major longitudinal axis of symmetry. Most structures
develop along this axis as nearly identical antimers, that is, they are expressed as mirror images of one
another. The genome of the developing embryo is believed to contribute equally to both sides of these
bilateral structures, so that there is an expectation that antimers will be symmetrical. Usually the
structures do not achieve perfect symmetry; instead, phenotype is expressed with small deviations from
perfect symmetry, that represent a departure from an ideal developmental plan encoded by genes (Van
Valen 1962).
For a half-century biologists have used departures from perfect symmetry as a means of
exploring the nature of development, developmental stability, and the effects of endogenous and
exogenous stresses on development. Many exogenous stressors are predicted to elevate asymmetry in
individuals, especially in areas or structures where developmental homeostasis is low and easily
disrupted (Livshits and Kobyliansky 1991). Endogenous disturbances like chromosomal abnormalities,
Mendelian-transmitted disease, and mulitfactorial hereditary defects have been shown to increase the
asymmetry of traits such as dermatoglyphic ridge counts (Kobylianski et al. 2000).
The tool most commonly used in investigations of asymmetry has been the distribution of left-
minus-right (LMR) values of a character. This distribution has been used to quantify two types of
bilateral asymmetry: directional asymmetry (DA) and fluctuating asymmetry (FA). Directional
asymmetry is the condition in which the trait on one side has a larger measure, on average, than its
counterpart. Directional asymmetry is usually quantified as the mean of the LMR distribution. A mean
of zero implies no DA and values that differ significantly from zero are evidence for DA.
Fluctuating asymmetry quantifies random deviations from perfect symmetry that occur when
antimeres are not perfect mirror images of each other. A number of indices have been developed for
quantifying fluctuating asymmetry. Nearly all of them are based on the variance (or standard deviation)
of the distribution of LMR differences (Palmer and Strobeck 1986). A variance near zero in this
distribution reflects perfect symmetry.

3
A third type of asymmetry, called antisymmetry (AS) arises when there is consistent DA within
individuals, but that the direction of the asymmetry is distributed randomly among individuals. In other
words, one of the antimeres tends to be consistently larger for each individual, but among individuals
there is no apparent bias for one side or the other. A well-known example of AS in nature is the
asymmetric claws on the male fiddler crab. Subtle examples of AS are not as widely recognized,
perhaps because the tools to distinguish AS from other types of asymmetry have not been available.
In this paper, we develop a new approach for investigating asymmetries. Rather that using the
LMR distribution as the basis for quantifying asymmetry, our approach is to estimate a distribution of
developmental noise along with the distribution of the trait itself. We base this approach, to the extent
possible, on the processes believed to bring about the expression of bilateral traits. As a result, the
parameters estimated for particular traits should provide insight into developmental processes and
disruptions by which asymmetries arise.
We begin by discussing some of problems and limitations inherent in the traditional methods for
studying asymmetries and developmental noise. Next, we describe the new method for directly
investigating developmental stability that is applicable to a wide variety traits, and solves many of the
problems encountered by the traditional analytic methods. Finally, we provide a number of examples of
the new method.
Standard Approaches to Measuring Asymmetry
Current approaches to the study developmental noise depend almost entirely on estimating
indices of FA and DA. A plethora of indices have been proposed, and nearly all are based on the
distribution of LMR differences between bilateral traits. Detailed descriptions of different methods, and
comparisons among them can be found elsewhere (Palmer and Stroebeck, 1986; Palmer, 1994; Møller
and Swaddle, 1997). For our purposes it is sufficient to point out that the most basic forms for
measures of DA and FA are Mean(L – R) and Var(L – R), respectively. Additionally, for a trait with
no DA, Mean(| L – R |) is another way to characterize the dispersion of the LMR distribution, and can
form the basis of a measure of FA.

4
A fundamental problem with current methods for assessing FA is that they are not based on a
specific etiologic model of developmental noise and measurement error. It is the non-etiologic nature of
the FA indices that leads, in part, to analytical difficulties and hinders interpretation. Some of the
problems of using and interpreting FA (and to a lesser extent, DA) are as follows:
Different index, different result: Different FA indices can produce different results on the
same data, making interpretation unclear. No single index is considered clearly superior under all
circumstances. Palmer and Stroebeck (1986), after examining many different indices, and after
analyzing different simulated data sets, found no universally superior index. They recommend computing
and reporting several indices. Clearly, a single index that provides an unambiguous result is preferred to
multiple, and possibly conflicting, indices.
Non-normal traits and discrete traits: A universal approach has not been developed for
working with traits that are not normally distributed. As discussed later in this paper we should exploit
the very reasonable assumption that developmental noise is normally distributed. Yet, the LMR
distribution is not, in general, normally distributed. For example, if the bilateral trait is distributed as a
negative exponential, then the resulting distribution of LMR differences is a Laplace (or double
exponential) distribution (Johnson et al. 1995). Ad hoc versions of FA indices have arisen in response.
A related problem arises in working with meristic (discrete) traits that have low trait counts.
Indices of FA are based on continuous traits, so that ad hoc adjustments are typically used with meristic
traits (Palmer and Stroebeck 1986; M∅ller and Swaddle 1997).
Incompletely observed traits: A difficulty arises when the bilateral trait under investigation is
incompletely observed. For example, traits that are measured as the timing to some bilateral event
(suture closure, tooth emergence, etc.) can entail right-censoring of one or both of the measurement
pairs. Additionally, these traits are usually measured over an interval, rather than as an exact time to the
event. The interval encloses the unobserved time. This type of observation referred to as interval-
censored in the statistical literature. Methods to assess asymmetry in interval-censored traits have never
been developed. For this reason some type of bilateral traits—particularly those that are timings to
some developmental stage—have rarely been explored.
Exploring asymmetry. The traditional methods are not conducive to exploring the causes and
nature of asymmetries. For example, there is no reason to believe that FA cannot be measured as a

5
signal on top of AS. Likewise, directional effects can be considered a continuous spectrum that begins
with DA (where a size bias arises 100% to one side) to AS (where the size bias occurs 50% to one
side). Finally, an understanding of how FA, DA, and AS arise and are affected by endogenous and
exogenous stresses requires that we simultaneously model all three asymmetries. Furthermore, we
should be able to incorporate the effects of covariates on the properties of asymmetries. Current
methods lack this degree of flexibility.
Measurement Error: It was recognized early on that measurement error can severely affect
estimates of FA (REF). This is because the variance in the LMR distribution is the sum of the variance
arising out of true differences and the variance resulting from errors of measurement. Some methods
have been developed that disentangle FA from measurement error given repeated measures of each
trait. Palmer and Stroebeck (1986) introduced a two-way mixed analysis of variance (ANOVA)
framework (see also, Møller and Swaddle 1997; Boklage 1992). The ANOVA framework provides
for systematic tests of hypothesis about fluctuating and directional asymmetry, and can be used to
examine effects of other covariates (e.g. sex, treatment). The technique yields a single error-adjusted
variance computed using repeated measurements (Palmer 1994). More recently, Van Dongen and
colleagues (1999) have proposed a mixed regression model that yields the same results as the
ANOVA, but provides a more direct interpretation of estimated parameters, allows for more flexibility
for hypothesis testing, and can be used to estimate individual error-corrected estimates of FA.
Methods
Measuring developmental noise
We have developed a new approach to quantifying developmental noise in bilateral traits. The
approach is motivated by simple biological principles, and we believe it can be broadly applied to the
study of asymmetries. Briefly, the difference between our approach and previous approaches is that our

6
method simultaneously estimates two parametric distributions from pairs of bilateral traits: (1) a
distribution of developmental noise and (2) a single distribution for the underlying trait. The first
distribution, developmental noise, is expected to be normally distributed (although even this assumption
is easy to relax). In the absence of directional asymmetry, the distribution of developmental noise will
have a mean of zero and a variance that characterizes the developmental noise. The second distribution
represents the distribution of the underlying phenotype, unmodified by developmental noise. This
distribution need not be (and arguably should rarely be) a normal distribution. Some of the difficulty
encountered using the traditional approaches is that distribution of the trait and the distribution of
developmental noise are confounded.
We are able to estimate both distributions using maximum likelihood to estimate parameters.
The method should have broad applicability for a wide variety of traits and missing data situations.
Simple extensions of the method allow for quantification of the effects of covariates. The covariates can
be modeled as affecting the bilateral trait in several ways. They can affect the variance in developmental
noise, akin to increase or decrease in fluctuating asymmetry. Covariates effects can also be modeled as
affecting the mean of the underlying trait. For example, disturbances might retard the rate of
development, resulting in longer mean times to developmental events. Third, the covariates might affect
the variance of the underlying trait. For example, the expression of a trait in individuals exposed to an
environmental perturbation might be more variable, but with the same mean as individuals not exposed.
Finally, covariates can affect directional asymmetry in a number of ways.
In the following section we provide details of the new approach, beginning with the simplest
case of a continuous trait. We then generalize the basic model to incorporate discrete traits, directional
asymmetry, time-varying covariates, measurement error, and incomplete observations. The result is a
flexible approach that overcomes many of the disadvantages of the standard methods used to quantify
FA.
We begin with a single bilaterally symmetric trait, denoted by random variable T. Trait T is
distributed according to probability density function (PDF) fT(t; µT, σT), where µT and σT are the
location and scale parameters respectively. For now, we need not specify any particular distribution of
the trait T, although a normal or a lognormal distribution tends to be used for many continuous traits
(Wright 1968). For all traits, it is difficult or impossible to measure specific values of ti (the value of t

7
for the i-th individual) because of the presence an unknown amount of developmental noise and
measurement error superimposed on ti. Hence, T is a latent random variable because it cannot be
measured directly.
Instead of measuring ti for individual i, we can measure li and ri, the left and right measurements
of the bilateral trait. Aside from a common but unmeasureable value of ti, measurements li and ri also
include developmental noise and possibly measurement error. Call A a random variable that describes
all sources of departures from the true values of T. In particular, A includes developmental noise (which
will be represented by random variable D) and measurement error (which will be represented by
random variable E).
Random variables for the left and right measurements (L and R respectively) share a common
value T and the bilateral measurements provide information on two independent realizations of A.
(1) l l l
r r r
L T A T D ER T A T D E
= + = + += + = + +
.
Note that we do not subscript T for each side. This results from the assumption that the trait is a single
phenotype that is expressed identically on both sides. When A is subdivided into its two components, E
can frequently be assumed normally distributed with a mean of zero. The variance of A is σA2 = σD
2 +
σE2, where σD
2 is the variance attributable to developmental noise, and σE2 is the variance in
measurement error.
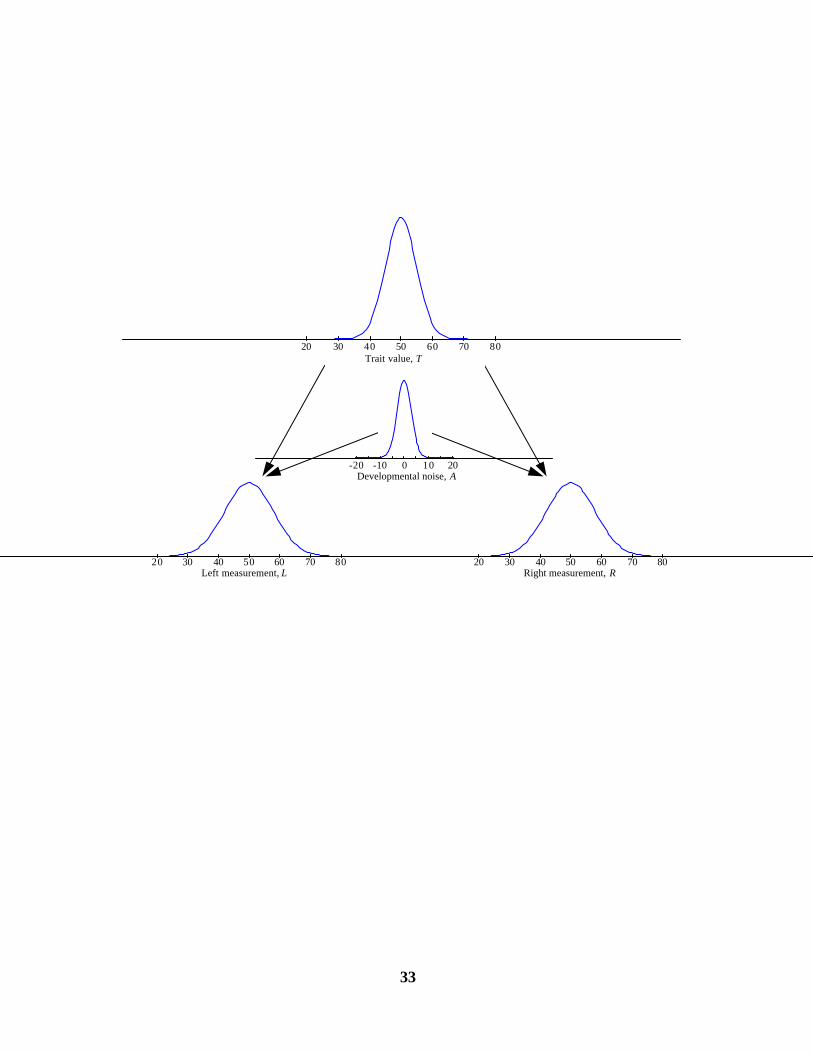
A graphical representation is given in Figure 1. The top of the figure shows the distribution of
the trait, T. Added to T are two values drawn from the distribution of developmental noise (and
perhaps measurement error) shown as the distributions in the center. The result are the two distributions
of measured values (L and R) shown on the bottom.
We begin with the simplest model in which E is minimal, which corresponds to the case where
traits can be measured in a way that ensures a very small measurement error. This assumption is used
when repeated measures are not available for each side to separate A into its two components. The
PDF of A is denoted g(a|0, σA); usually we can assume A is normally distributed with a mean of zero
and a variance of σA2.

8
Suppose for a moment that ti, ali, and ari could be measured directly in a sample of N
individuals. For each individual, we would have a single realization from T, and two independent
realizations from A, one left and right, with a mean of zero and a common variance. It would the be
simple to estimate parameters for the three distributions fT(t|µT, σT), g(aL|0, σA), and g(aR|0, σA). If the
distributions were Normal, estimates of µT, σT could be obtained as a mean and standard deviation
from the array of N measurements t = (t1, t2, ... tN). Likewise, σa could be found as the standard
deviation among N measurements aL = (aL1, aL2, ... aLN) and aR= (aR1, aR2, ... aRN). An alternative
method of finding µT, σT and σA is maximum likelihood. The likelihood for the i-th individual would be
constructed as the product of values of the three distributions as
(2) ( , , | , , ) ( | , ) ( |0, ) ( |0, )i T T A i Li Ri T i T T A Li A A Ri AL t a a f t g a g aµ σ σ = µ σ σ σ .
The likelihood over all N individuals would be the product of the individual likelihoods, or
(3) 1
( , , | , , ) ( | , ) ( |0, ) ( |0, )N
T T A T i T T A Li A A Ri Ai
L f t g a g a=
µ σ σ = µ σ σ σ∏L Rt a a .
The values of µT, σT and σA that maximize L for a given set of observations are the maximum likelihood
estimates for these parameters.
One advantage of using the likelihood over the least-squares estimation is that any parametric
distribution can be specified for fT( ). For that matter, any parametric distribution can be used for gA( ),
as well. Additional advantages come from being able to build complex models to incorporate, for
example, measurement error, directional asymmetry, and the effects of covariates on the distributions, as
done in later sections.
So far, we have made the assumption that ti, aLi and aRi could be measured directly. In
practice we only have measurements li and ri from which to make inferences about parameters of fT( )
and gA( ). To construct a likelihood that only makes use of li and ri, suppose we could measure ti, li
and ri, but not aLi, and aRi. Then, given that li = ti + aLi and ri = ti + aRi, likelihood (3) would be
changed to

9
(4) 1
( , , | , , ) ( | , ) ( |0, ) ( |0, )N
T T A T i T T A i i A A i i Ai
L f t g l t g r t=
µ σ σ = µ σ − σ − σ∏t l r .
In fact, even ti cannot be measured directly. Instead T can be treated as a latent variable. A likelihood
can be taken as the expectation over all possible values of T as
(5) 1 0
( , , | , ) ( | , ) ( |0, ) ( |0, )N
T T A T T T A i A A i Ai
L f t g l t g r t dt∞
=
µ σ σ = µ σ − σ − σ∏∫l r .
The left-hand side of (5) is maximized with respect to µT, σT and σA, to find maximum likelihood
estimates for these parameters.
Likelihood (5) provides the means for estimating the distribution of the latent trait T through
parameters µT and σT. At the same time, the parameter σA is found that reflects the sum of the
developmental noise and the error of measuring li and ri.
Meristic Traits: Meristic traits are measured as a discrete integral number of repeated
structures. Some meristic traits used in the study of developmental stability are ridge counts in finger
and palm prints (Livshits and Kobyliansky 1987; Livshits and Smouse 1993; Fraser 1994), bristle
counts on fruit flies and vein counts in insect wings (M∅ller and Swaddle 1997). Typically, meristic
traits with high counts are analyzed as if continuous. Meristic traits can be modeled as having a discrete
distribution for fT( ). Likelihood (5) can be modified for discrete distributions, but still with a continuous
distribution of developmental noise, as
(6)
[ ][ ]
1 1
01
01
( , , | , ) ( | , ) ( |0, ) ( |0, )
( | , ) ( |0, ) ( 1 |0, ) ( |0, ) ( 1 |0, )
i i
i i
l rN
T T A T T T A A A Ati l r
N
T T T A i A A i A A i A A i Ati
L f t g l t dl g r t dr
f t G l t G l t G r t G r t
+ +∞
==
∞
==
µ σ σ = µ σ − σ − σ
= µ σ − σ − + − σ − σ − + − σ
∑∏ ∫ ∫
∑∏
l r,
where GA( ) is the cumulative density function (CDF) that corresponds to gA( ). For example, fT( )
might be well described by a binomial or a poisson distribution.
An alternative analysis for meristic traits is to assume that T is, in fact, a continuous underlying
trait, but one that can only be measured down to an integral number of structures (M∅ller and Swaddle

10
1997). Hence, the true value of ti is an unknown value in the interval [ti, ti+1). Under this assumption,
the methods for analyzing interval censored observations, discussed next, may be directly applied.
Extensions for Chronometric and Censored Traits: Bilateral morphological traits are
usually taken as a pair of measures, li and ri. Some traits, particularly those associated with timing to
some morphological transition (chronometric traits), may be right-censored or observed over intervals
of time. These traits typically involve timing of the initial appearance or the completion of a structure.
Examples are epiphysial closure, suture closure, and the emergence of teeth (which is used as an
example later).
Two common characteristics of such chronometric traits are 1) they are usually observed over
intervals, so that exact measures are not known, 2) bilateral measures for some individuals are
incomplete (right-censored) because one or both of the events fails to occur before the study is
complete, the subject withdraws from the study, the subject dies, etc. All of these types of observations
fall under the rubric of interval censoring. Such traits come as two measures defining the smallest
observed interval over which the event occurred. For a pair of bilateral chronometric traits measured
for the ith individual, we obtain [loi, lci) and [roi, rci), where the subscript o denotes the opening time and
the subscript c denotes the close of the half open interval.
A likelihood is constructed by integrating over all possible values of the trait:
(7)
[ ][ ]
1 0
1 0
( , , | , , , ) ( | , ) ( |0 , ) ( |0, )
( | , ) ( |0, ) ( |0 , ) ( |0, ) ( |0, )
ci ci
oi o i
l rN
T T a T T T A a A ai l r
N
T T T A ci a A oi a A ci a A oi ai
L f t g l t dl g r t drdt
f t G l t G l t G r t G r t dt
∞
=
∞
=
µ σ σ = µ σ − σ − σ
= µ σ − σ − − σ − σ − − σ
∏∫ ∫ ∫
∏∫
o c o cl l r r
Where GA( ) is the cumulative density function (CDF) that corresponds to gA( ). Likelihood (7)
provides enormous flexibility for different study designs and missing data situations. A right-censored
observation for one or both sides is defined as an observation for which a minimum possible time is
known, but the maximum time (c) is not observed. This type of observation is handled by simply setting
the value of the maximum time (c) to infinity. Another situation occurs when the event has occurred on
one or both sides prior to the first observation time, as might occur in a cross-sectional or a mixed
longitudinal-cross-sectional study. In that situation the minimum time for that observation (o) is set to

11
zero, and the maximum time (c) is set to the time of the first observation (Wood et al. 1992; Holman
and Jones 1998).
Paleontological samples present some analytical difficulties because individual specimens may
have fragmented or broken measures for one or both sides, or may simply be missing one of the two
measurements. Depending on the process by which missing observations arise, excluding incomplete
specimens can result in biased parameter estimates. Under some mild statistical assumptions,1 we can
use the same technique as we did for interval censored observations. A missing side is handled as
having a minimum length of zero and a maximum length of infinity. The missing side adds no information
to the distribution gA( ) on the missing side, but the side that is not absent contributes to a better
determination of fT( ) and gA( ). When one or both sides of a structure are incompletely preserved (i.e.
a portion has broken off and has been lost), the missing measurement may be treated as a right-
censored observation, taking the minimum length as the measured size and the maximum length as
infinity (or the maximum length possible in a confined grave).
Measurement Error: An important issue in the recent FA literature is the development of
methods to disentangle measurement error from developmental noise, as the measurement error for
some traits may swamp the developmental noise (Greene 1984; Palmer and Strobeck 1986; 1997;
Fields et al. 1995). The effects of measurement error and developmental instability can be disentangled
when repeated measures are available for each trait. With repeated measures we can refine the
methods so that the distribution gA( ) is now separated into a distribution of measurement error, gE( ),
and the distribution of developmental noise, gD( ).
Given a sample of N individuals, where the i-th individual has mi repeated observations, an
expectation is computed over the distribution of developmental noise and the distribution of
measurement error. Likelihood (5) modified to incorporate repeated measures yields,
1 The condition is that the probability of a measurement being missing should be independent of the parameters being estimated. This condition might be violated, for example, if the most asymmetric specimens have the highest probability of missing a measurement. See Rubin and Little (19??) and Tanner (1996) for a more technical discussion of this issue.

12
(8)1 10
( , , , | , ) ( | , ) ( |0, ) ( |0 , ) ( |0, )imN
T T D E T T T E E D ij D D ij Di jx
L f t g e g l t e g r t e dedt∞ ∞
= =
µ σ σ σ = µ σ σ − − σ − − σ∏ ∏∫ ∫l r .
The other likelihoods are similarly modified to incorporate measurement error.
Directional Asymmetry: The biological basis of DA is not clearly understood. In some cases
it appears to be an adaptive trait; other instances of DA seem to arise from developmental stress
(Møller and Swaddle 1997). Analytical methods that quantify asymmetry should incorporate some
method of quantifying DA in order to avoid serious biases resulting from violating assumptions about the
underlying distribution. Also, many traits display DA, so we should explicitly model it. We model DA
as a shift by parameter d in the trait to the left and minus d in the trait to the right. That is, the mean of
the gA( ) distribution has a parameter d on the left and -d on the right in place of 0. Likelihood (5) is
modified to
(9) 1 0
( , , | , ) ( | , ) ( | , ) ( | , )N
T T A T T T A i A A i Ai
L f t g l t d g r t d dt∞
=
µ σ σ = µ σ − σ − − σ∏∫l r .
A second facet to DA is its effect on the variance of developmental noise on each side. For
example, in cases of extreme DA, the larger side may exhibit a larger variance in developmental noise
instead of, or in addition to, a shift in the location. One simple way of modeling this is to include a
separate scale parameter (σA) for the left and the right side. The likelihood modified for both types of
DA is
(10) 1 0
( , , , | , ) ( | , ) ( | , ) ( | , )l r l r
N
T T A A T T T A i A A i Ai
L f t g l t d g r t d dt∞
=
µ σ σ σ = µ σ − σ − − σ∏∫l r .
A number of hypotheses about directional affects can be tested using this likelihood. To do so, one or
more parameters are constrained so that: (1) there is no mean directional effect (d = 0, σAl = σAr); (2)
there is no DA, but there is an asymmetrical dispersion of developmental noise (d = 0, σAl ≠ σAr); (3)
there is DA, but no asymmetry in dispersion of developmental noise (d ≠ 0, σAl = σAr); (4) there is both
directional asymmetry and asymmetrical dispersion of developmental noise (d ≠ 0, σAl ≠ σAr).
Antisymmetry. It is desirable to be able to distinguish AS from directional asymmetry and
developmental noise. The presence of AS does not rule out measuring useful information on

13
developmental noise. Additionally, questions like the effect of stressors on antisymmetric traits might be
asked. Currently, these question are almost entirely unexplored. It is reasonable to suppose that
developmental noise will leave a signature superimposed on AS. Until recently, there have been few
methods for the analysis of traits exhibiting AS. Van Dongen, Lens and Molenberghs (1999)
introduced a method using finite mixture models to estimate FA, DA, and AS. We adopt a similar
mixture model approach to quantify AS, but within the framework of estimating developmental noise.
Under this approach, AS is assumed to represent a form of DA that is distributed between both sides.
For an individual observation, the largest side will usually, but not necessarily always, reflect the
direction in which development is biased. (It is conceivable that developmental noise added to the
smaller side or removed from the larger side will result in a side reversal of the side with the largest trait.)
Incorporating AS into the estimation framework requires a single new parameter, p, that
quantifies the probability that the directional bias in size (d) and the directional bias in dispersion of
developmental noise (σA1) will occur on the left side. Likelihood (10) is modified to
(11)1 2
1 2 2 11 0
( , , , , | , )
( | , ) ( | , ) ( | , ) (1 ) ( | , ) ( | , )
T T A A
N
T T T A i A A i A A i A A i Ai
L p
f t pg l t d g r t d p g l t d g r t d dt∞
=
µ σ σ σ =
µ σ − σ − − σ + − − − σ − σ ∏∫
l r.
"Perfect" antisymmetry results when p = 0.5. In this case, the directional bias is divided uniformly
between the left and the right side. When p = 0, there is no antisymmetry and the likelihood reduces to
(9), the case of directional asymmetry. Values of p between 0 and 0.5 yield a hybrid antisymmetry-
directional asymmetry. Thus, antisymmetry is one endpoint of a continuous spectrum that begins with
directional asymmetry. Now we can explicitly test the hypotheses that a trait exhibits antisymmetry (p =
0.5). When p is constrained to 0, the directional asymmetry hypotheses can then be examined.
Quantifying Effects of Covariates: Suppose we have individual-level covariates such as
gender or whether or not individuals are exposed to a particular stress. Call xi = (x i1, x i2, . . ., x in) an
array of n such variables (or covariates) for the i-th individual. The xi covariates can be modeled as
affecting several different components of the model. For example, the might affect the variance in
developmental noise by substituting σAi=σAexp(xi'β A), where β A = (β1A, β2A, . . ., βnA) is an array of
beta coefficients that quantify the effects of covariates on the distribution of developmental noise. The
parameter σAi is used instead of σA in the likelihood.

14
The xi covariates might also affect other aspects of growth. For example, the size of the trait
itself might be affected some particular stress. This can be examined by replacing µT for an individual
measure µTi = µTexp(xi'β T), where β T = (β1T, β2T, . . ., βnT) is an array of beta coefficients that
quantify the effects of covariates on the mean of trait T. (Alternatively, the linear parameterization µTi =
µT + xi'β T can be used). Another possible effect of covariates might be to change the magnitude of
directional asymmetry, which is accomplished by substituting di = dexp(xi'β d) in place of d, where β d
= {β1d, β2a, . . ., βnd} is an array of beta coefficients.
Subjects
Asymmetry and developmental noise are estimated for several types of traits in two different
samples. The first is a sample of Javanese children for whom we examine asymmetry in developmental
noise in the time to emergence of deciduous teeth. The second is a sample of individuals with known
prenatal exposure to alcohol, as well as a group of unexposed controls. We examine the effect of
alcohol exposure on two dermatoglyphic traits: a-b ridge counts, and atd angle.
Deciduous tooth emergence in Javanese children. Data on bilateral emergence of
deciduous teeth come from the Ngaglik project, a three-year prospective investigation of maternal and
child health and nutrition, breastfeeding, and birth spacing dynamics carried out in the late 1970’s in
Central Java, Indonesia (Ngaglik Study Team 1978; Hull 1983). A description and previous analyses
of the dental data can be found elsewhere (Holman and Jones 1991, 1998; Hull 1983). Briefly, 468
children in two rural villages were examined for clinical emergence of deciduous teeth prospectively over
the two and one-half year course of the study. Children recruited into the study were from zero to six
months of age, but none had emerged a tooth at the first observation. Exact dates of birth and dates of
dental examinations are known. Interviews were scheduled every 35 days (one Javanese month);
however, about 5% were of emergence events were observed over a larger interval because one or
more visits were missed. Hence, times to emergence are always observed as the minimum time (to) and
the maximum possible time (tc) of emergence. Together, [to, tc) defined an interval within which
emergence occurs; for right censored observations, tc is set to infinity.

15
The timing of tooth emergence is modeled as being lognormally distributed beginning at one
month in utero. The use of a lognormal distribution for tooth emergence
Emergence age is shifted by a constant of eight months, back to one month after conception,
corresponding to the time when the dental lamina are initially formed (Ten Cate, 1998). Using
conception, or shortly thereafter, as the starting point for tooth emergence was suggested by Kihlberg
and Koski (1954) and Hayes and Mantel (1958), and has been used or discussed in a number of
previous studies (Holman and Jones, 1998; Magnússon, 1982; Smith et al., 1994). The distribution of
developmental noise is modeled as a normal distribution. A single trait, emergence of the lower central
deciduous incisor, is analyzed here.
a-b ridge count in children exposed to prenatal alcohol. Right and left hand prints were
obtained from individuals affected with fetal alcohol syndrome (FAS) or fetal alcohol effects (FAE) and
a control sample of individuals who reported that their mothers do not drink alcohol. The affected
sample was 193 subjects diagnosed with fetal alcohol syndrome (FAS) (29F, 39M) or fetal alcohol
effects (FAE) (52F, 73M). All affected individuals had been referred to the Fetal Alcohol Syndrome
Diagnostic Clinic at the University of Washington for diagnostic evaluation during 1994-95 for further
follow-up. The patients were typically referred for cognitive and behavioral problems, and had a
confirmed or suspected history of prenatal alcohol exposure. (Striessguth et al. 1996). At the time of
recruitment, ages of subjects ranged from 6-51 years of age, 74% were 6-11 years, 62%, 12-20 years
and 36%, 21-51 years.
The control sample of 190 subjects (80F, 110M) was recruited from students (19-35 years
old) at the University of Washington during the periods 1995-96 and 2001). All were screened for
eligibility by interview. Participants had no prior diagnosis as FAS/E, had no familial history of heritable
diseases or congenital defects, and reported that their mother did not consume alcohol.
Handprints were taken using the standard carbon-paper-tape technique (Reed and Meier
1990). The prints were scanned into digital images at 400 dots per inch resolution. Prints were
assesed for left and right a-b ridge count. Intraobserver error in ridge count error was XXX ridges and
for ATD angle was XXX degrees. After the prints were read the case number was used to identify the
individual characteristics like sex and diagnosis (FAS/E or Control).

16
Numerical methods and model selection
Maximum likelihood estimates of parameters were found by numerically evaluating likelihoods
using the mle programming language (Holman 2000). Estimates of the standard errors for all
parameters are found by the method of Nelson (1982), which involves inverting a numerical
approximation of Fisher's information matrix. Akaike Information Criterion (AIC) is used to select the
between competing forms of each models (Akaike 1973, 1992; Burnham and Anderson 1998). The
model that minimizes AIC is taken to be the most parsimonious model. (Alternatively, likelihood ratio
tests can be used, and we provide loglikelihoods for that purpose.)
Results
Developmental noise in tooth emergence. Parameter estimates for these interval- and
right-censored observations were found as those that maximize likelihood (7), modified to include the
directional asymmetry parameter (d) as in likelihood (9).
Parameter estimates for the simplest model that includes two parameters of the underlying trait
(µT, σT) and developmental noise (σA) are given in Table 1. Developmental noise was clearly identified
along with the underlying trait of tooth emergence time.
The bottom panel is an analysis that includes DA. Significant DA was found to the left. Clearly,
some of the variance in developmental noise found in the first analysis was an artifact of DA. Even after
when DA is estimated, developmental noise is still detectable. The difference between the two analyses
in Akaike Information Criteria indicate that the second analysis provides a better fit to the observations
even with one extra parameter.
a-b ridge count. Alcohol is a known teratogen and its effects on morphology and especially,
neurobehavior are well documented (Streissguth et al. 1994). Alcohol presents a potentially robust
stressor to the developing embryo/fetus; as a result, developmental instability is predicted to be
increased in the phenotype of an individual exposed prenatally. Kieser (1992) documented greater

17
odontometric asymmetry compared to controls in 112 non-FAS children of alcoholic mothers.
Significantly higher FA was found for the intercore ridge count in macaque monkeys experimentally
exposed to alcohol in utero (Newell-Morris, Mauer and Clarren 1997), the a-b ridge count in patients
diagnosed with FAS (Wilber, Newell-Morris and Streissguth 1993), and females diagnosed with
FAS/E (Kuehn et al. 2001).
For this analysis we treat the a-b ridge count as a continuous, normally distributed trait, and the
distribution of developmental noise as normally distributed. Parameters were estimated using likelihood
(5), modified for some analyses to include the directional asymmetry parameter (d) and for some the
effect of alcohol exposure on the distribution of developmental noise (βa). The results of a series of
analyses are given in Table 3.
The first analysis estimates a mean ridge count of 43.0 and a standard deviation of 4.29. These
figures are close to the means and standard deviations estimated for the right (µ: 42.0, σ: 5.29) and left
ridge counts (µ: 44.1, σ: 5.17) separately. The standard deviation in developmental noise for the
combined sample (affected and controls) was 3.13. In the second analysis, the effect of alcohol
exposure on the distribution of developmental noise (βa) is estimated. The value of 0.53 (SE 0.13) is
significantly different from zero, and indicates an increase in the standard deviation of the distribution of
developmental noise of about 70 percent. (A likelihood ratio test indicates that this second model fits
significantly better). For the third analysis, we add a parameter for directional asymmetry (d), which is
highly significant, and the AIC difference or likelihood ratio test indicates that this third model fits the
data much better. The directional asymmetry is slightly less than one excess ridge to the left. In this
third analysis, the βa parameter is still significant at p < 0.05, and indicates an increase in the standard
deviation of the distribution of developmental noise of about 62 percent after controlling for DA.
This analysis shows that modeling of DA is important when measuring the effects of stressors on
developmental noise. Although the biological mechanisms for subtle directional asymmetry (as exhibited
in the a-b ridge count trait) is poorly understood, there is evidence that stressors may act to amplify the
degree of DA (Graham, Freeman and Emlen 1994). A natural extension to the current analysis would
be to explicitly model an effect of prenatal exposure on the DA parameter (as outlined in the Statistical
Methods Section).

18
References cited
Akaike H (1973, 1992) Information theory and an extension of the maximum likelihood principle. In
Petrov BN and Csaki F (eds.), Second International Symposium on Information Theory.
pp. 268-81. Budapest: Hungarian Academy of Sciences. Reprinted in Kotz S and Johnson N
(eds.). Breakthroughs in Statistics. New York:Springer Verlag, pp. 610-24.
Arrieta MI, Criado B, Martinez B, Lobato MN, Gil A and Lostao CM. Fluctuating dermatoglyphic
asymmetry: genetic and prenatal influences. Ann Hum Biol 20: 557-63.
Babler WJ (1978) Prenatal selection and dermatoglyphic pattern. Am J Phys Anthropol 48: 21-8.
Babler WJ (1979) Quantitative differences in morphogenesis of human epidermal ridges. In: Wertelecki
W, Plato CC and Paul NW (eds). Dermatoglyphics-FiftyYears Later. New York, Wiley-
Liss, pp.199-208.
Babler WJ (1991) Embryonic development of epidermal ridges and their configurations. In: Plato CC,
Garruto RM and Schaumann BA (eds.). Dermatoglyphics: Science in Transition. New
York, Wiley-Liss, pp. 95-112.
Boklage CE (1992) Method and meaning in the analysis of developmental asymmetries. J Hum Ecol
Special Issue 2: 147-56.

19
Bookstein FL, Sampson PD, Streissguth AP and Connor PD (2001) Geometric morphometrics of
corpus callosum and subcortical structures in the fetal-alcohol-affected brain. Teratology 64:4-
32.
Bratley P, Fox BL and Schrage LE (1983) A Guide to Simulation New York: Springer-Verlag.
Brown NA, McCarthy A, Wolpert L (1991) Development of handed body asymmetry in mammals. In.
Bock GR and Marsh J (eds.) Biological Asymmetry and Handedness. Chichester:Wiley. pp.
198-201. (Documents the sensitive period of development and handedness).
Burnham KP and Anderson DR (1998) Model Selection and Inference: A Practical Information-
Theoretic Approach. New York: Springer Verlag.
Connor PD, Sampson PD, Bookstein FL, Barr HM and Streissguth AP (2000) Direct and indirect
effects of prenatal alcohol damage on executive function. Develop Neuropsychol 18: 331-54.
Cummins H. (1936) Dermatoglyphic stigmata in Mongolian idiocy. Anat Rec 64 (Suppl. 3):11.
Cummins H and Midlo C (1943) Finger Prints, Palms and Soles. South Berlin, Mass: Research
Publishing Co.
Dell DA and Munger BL (1986) The early embyogenesis of papillary (sweat duct) ridges in primate
glabrous skin: the dermatoglyphic map of cutaneous mechanoreceptors and dermatoglyphics. J
Comp Neurol 244: 511-32.
Fields SJ, Spiers M, Hershkovitz I, Livshits G (1995) Reliability of reliability coefficients in the
estimation of asymmetry Am J Phys Anthropol 96: 83-7.
Fraser FC (1994) Developmental instability and fluctuating asymmetry in man. In Markow TA (ed.)
Developmental Instability: Its Origins and Evolutionary Implications. Netherlands: Kulwer
Academic Publishers. pp. 319-34.

20
Geoffe WL, Ferrier GD and Rogers J (1994) Global optimization of statistical functions with simulated
annealing. J Econ 60: 65-99.
Greene (1984) Fluctuating dental asymmetry and measurement error. Am J Phys Anthropol 65: 283-
9.
Halgrímsson B(1993) Fluctuating asymmetry in Macaca fascicularis: A study of the etiology of
developmental noise. Int J Primatol 14:421-43.
Hayes RL, Mantel N. 1958. Procedures for computing the mean age of eruption of human teeth. J Dent
Res 37:938-947.
Holman DJ and Jones RE (1991) Longitudinal analysis of deciduous tooth emergence in Indonesian
children: I. Life table methodology. Am J Hum Biol 3: 389-403.
Holman DJ and Jones RE (1998) Longitudinal analysis of deciduous tooth emergence II: Parametric
survival analysis in Bangladeshi, Guatemalan, Japanese and Javanese children. Am J Phys
Anthropol 105: 209-30.
Holman DJ (2000) mle: A Programming Language for Building Likelihood Models. Version 2
(Software and manual). http://faculty.washington.edu/~djholman/mle/
Holt SB (1968) The Genetics of Dermal Ridges. Springfield, Illinois: Charles C. Thomas.
Hull VJ (1983) The Ngaglik study: An inquiry into birth interval dynamics and maternal and child health
in rural Java. World Health Stat. Q. 36: 100-118.
Johnson NL, Kotz S, Balakrishnan N (1995) Continuous Univariate Distributions, Volume 2. New
York: John Wiley and Sons.
Kahn HS, Ravindranath R, Valdez R and Venkat Narayan KM (2001) Am J Epidemol 153:338-44.

21
Kieser JA (1992) Fluctuating odontometric asymmetry and maternal alcohol consumption. Ann Hum
Biol 19: 513-20.
Kihlberg J, Koski KK. 1954. On the properties of the tooth eruption curve. Suom Hammaslaeaek
Toim 50(Suppl 2):6-9.
Kobyliansky E, Bejerano M, Yakovenko K and Katznelson B-M (1999) Relationship between genetic
anomalies of different levels and deviations in dermatoglyphic traits. Part V. Dermatoglyphic
peculiarities of males and females with Cystic Fibrosis. Family study. Zeitschrift fur Morph u
Anthropol 82: 249-302.
Kobyliansky E, Bejerano M, Yakovenko K and Katznelson B-M (2000) Relationship between genetic
anomalies of different levels and deviations in dermatoglyphic traits. Newsl Am Dermatoglyph
Assoc 21: 3-60.
Kuehn C, Newell-Morris L, Reason L and Streissguth A (2001) Fluctuating dermatoglyphic asymmetry
in individuals exposed to alcohol prenatally. Am J Hum Biol 13: 127-8 (Abstract).
Lacroix B, Wolff-Quenot M-J and Haffen K (1984) Early hand morphology: an estimation of fetal age.
Early Hum Develop 9: 127-36.
Livshits G and Kobyliansky E (1987) Dermatoglyphic traits as possible markers of developmental
processes in humans. Am J Med Genet 26: 111-22.
Livshits G and Kobyliansky E (1991) Fluctuating asymmetry as a possible measure of developmental
homeostasis in humans: A review. Hum Biol 63: 441-66.
Livshits G and Smouse PE (1993) Multivariate fluctuating asymmetry in Israeli adults. Hum Biol 65:
547- 78.

22
Magnússon TE. 1982. Emergence of primary teeth and onset of dental stages in Icelandic children.
Community Dent Oral Epidemiol 10:91-97.
Malhotra KC, Majumdar L, Reddy BM (1991) Regional variation in ridge count asymmetry on hand.
In Reddy BM, Roy SB, Sarkar BN (eds.) Dermatoglyphics Today. Calcutta:Anthropological
Survey of India and Indian Statistical Institute. pp. 153-66.
Malina RM and Buschang PH (1984) Anthropomorphic asymmetry in normal and mentally retarded
males. Ann Hum Biol 11: 515-31.
Markow TA (1992) Genetics and developmental stability: an integrative conjecture on aetiology and
neurobiology of schizophrenia. Editorial. Psychol Medicine 22: 295-305.
Markow TA and Wandler K (1986) Fluctuating dermatoglyphic asymmetry and the genetics of liability
to schizophrenia. Psychiatry Research 19: 323-8.
Mellor CS (1992) Dermatoglyphic evidence of fluctuating asymmetry in schizophrenia. Brit J Psychiat
160:467-72.
M∅ller and Swaddle (1997) Asymmetry, Developmental Stability and Evolution. Oxford: Oxford
University Press.
Ngaglik Study Team (1978) Birth interval dynamics in village Java: The methodology of the Ngaglik
study. Methodology Series No. 4. Population Studies Center, Gadjah Mada University,
Yogyakarta, Indonesia.
Nelson W (1982) Applied Life Data Analysis. New York: John Wiley and Sons.
Newell-Morris L, Fahrenbruch C and Sackett GP (1989) Prenatal psychosocial stress, dermatoglyphic
asymmetry and pregnancy outcome in die pig-tailed macaque (Macaca nemestrina). Biol
Neonate 56: 61-75.

23
Newell-Morris L, Fahrenbruch C and Yost C (1982) Nonhuman primate dermatoglyphics: implications
for human biomedical research. In: Bartsocas C. (ed.), Progress in Dermatoglyphic
Research, NY: Alan R. Liss, pp. 189-202.
Newell-Morris L and Weinker TF (1991) The dermatoglyphics of nonhuman primates: A neglected
resource. In: Plato CC, Garuto RM and Schumann B (eds.), Dermatoglyphics: Science in
Transition (Birth Defects Original Article Series 27), pp. 267-90.
Newell-Morris LL, Mauer T and Clarren SK (1997) Maternal consumption of alcohol and association
with dermatoglyphic asymmetry in a monkey model. Am J Phys Anthropol 24 (Suppl):177
(abstract).
Noreen EW (1989) Computer Intensive Methods for Testing Hypotheses. New York: John Wiley
and Sons.
Okajima M and Newell-Morris L (1988) Development of dermal ridges in the volar skin of fetal
pigtailed macaques (Macaca nemestrina) Am J Anat 183: 323-37.
Palmer AR and Strobeck C (1986) Fluctuating asymmetry: Measurement, analysis, pattern. Ann Rev
Ecol System 17: 391-421.
Parsons PA (1990) Fluctuating asymmetry: an epigenetic measure of stress. Biol Rev 65: 131-45.
Reed T and Meier RJ (1990) Taking Dermatoglyphic Prints: A Self-Instruction Manual. Suppl
Newsletter, American Dermatoglyphics Association.
Rose RJ, Reed T and Bogle A (1987) Asymmetry of a-b ridge count and behavioral discordance of
monozygotic twins. Behav Genet 17: 125-40.
Rubin and Little ()

24
Sampson PD, Sreissguth AP, Bookstein FL, Little RE, Clarren SK, Dehaene P, Hanson JW and
Graham JM Jr. (1997) Incidence of fetal alcohol syndrome and prevalence of alcohol-related
neurodevelopmental disorder. Teratology 56: 317-26.
Schaumann B and Alter M (1976) Dermatoglyphics in Medical Disorders. New York: Springer
Verlag.
Smith AFM and Roberts GO (1993) Bayesian comuptation via the Gibbs sampler and related Markov
chain Monte Carlo methods. J Royal Stat Soc B 55: 3-23.
Smith BH, Garn SM and Cole PM (1982) Problems of sampling and inference in the study of
fluctuating asymmetry. Amer J Phys Anthropol 58: 281-9.
Smith BH, Crummett TL, Brandt KL. 1994. Ages of eruption of primate teeth: A compendium for aging
individuals and comparing life histories. Yrbk Phys Anthropol 37:177-231.
Steele J and Mays S (1995) Handedness and directional asymmetry in the long bones of the human
upper limb. Interntl J Osteoarch 5: 39-49.
Streissguth AP, Sampson PD, Carmichael Olson H, Bookstein FL, Barr HM, Scott M and Feldman J
(1994) Maternal drinking during pregnancy and attention/ memory perfomance in 14-year-old
children: A longitudinal prospective study. Alcohol: Clin and Exper Res 18: 202-18.
Ten Cate AR. 1998. Oral Histology: Development, Structure and Function. Fifth edition. St. Louis:
Mosby.
Tanner MA (1996) Tools for Statistical Inference: Methods for the Exploration of Posterior
Distributions and Likelihood Functions, 3rd edition. New York: Springer-Verlag.

25
Van Dongen S, Lens L and Molenberghs G (1999) Mixture analysis of asymmetry: Modelling
directional asymmetry, antisymmetry and heterogeneity in fluctuating asymmetry. Ecology
Letters 2:387-96.
Van Valen L (1962) A study of fluctuating asymmetry. Evolution 16: 125-42.
Weiss KM (1990) Duplication with variation: Metameric logic in evoluation from genes to morphology.
Yrbk Phys Anthropol 33: 1-23.
Wilber E, Newell-Morris L and Streissguth A (1993) Dermatoglyphic asymmetry in fetal alcohol
syndrome. Biol Neonate 64: 1-6.
Wood JW, Holman DJ, Weiss KM, Buchanan AV and LeFor B (1992) Hazards models for human
population biology. Yrbk Phys Anthropol 35: 37-63.
Wright S (1968) Evolution and the Genetics of Populations. Vol. 1. Chicago: University of Chicago
Press.

26
Acknowledgments
We are grateful to V. Hull for facilitating work in Indonesia, Matthew Johnson and Letitia
Reason for assisting with handprints. This research was supported by grants from NSF
(BNS8115586) (REJ), the Alcohol and Drug Abuse Institute, University of Washington (LNM), ...
27
Tables
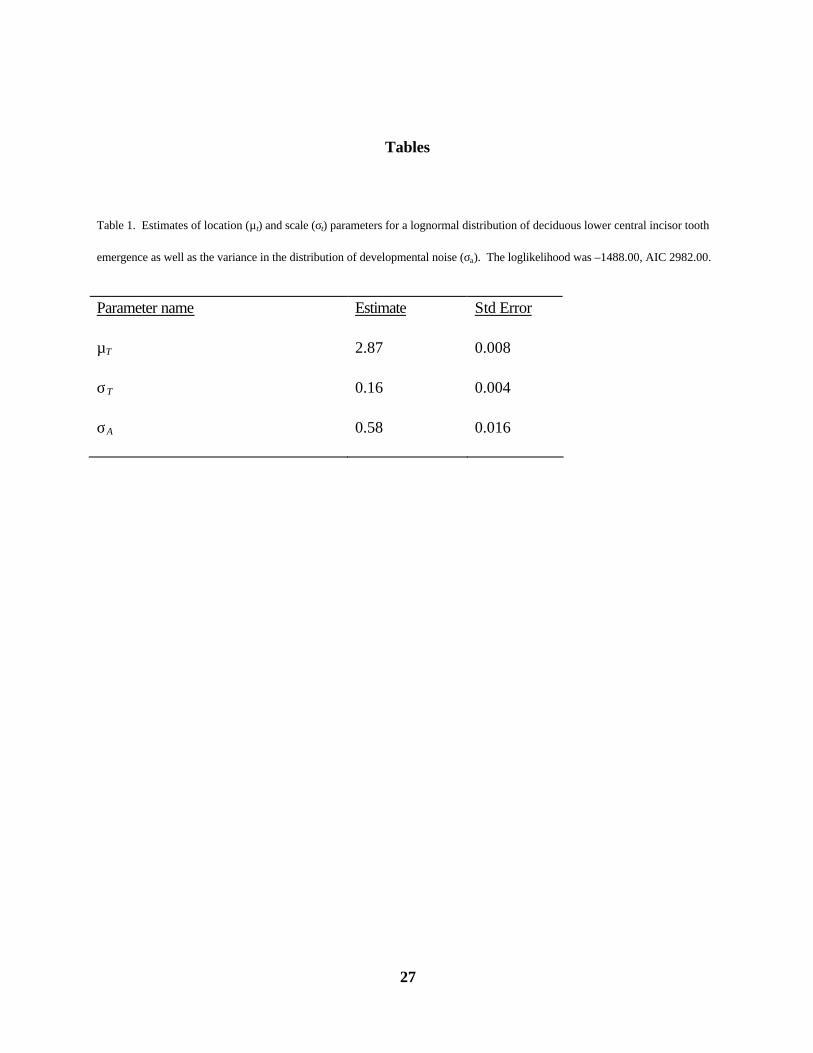
Table 1. Estimates of location (µt) and scale (σt) parameters for a lognormal distribution of deciduous lower central incisor tooth
emergence as well as the variance in the distribution of developmental noise (σa). The loglikelihood was –1488.00, AIC 2982.00.
Parameter name Estimate Std Error
µT 2.87 0.008
σT 0.16 0.004
σA 0.58 0.016

28
Table 2. Estimates of location (µt) and scale (σt) parameters for a lognormal distribution of deciduous lower central incisor tooth
emergence as well as the variance in the distribution of developmental noise (σa) and directional asymmetry (d); loglikelihood is –
1484.58, AIC is 2977.15.
Parameter Estimate Std Error
µT 2.87 0.008
σT 0.17 0.004
σA 0.16 0.04?
d 0.056 0.02?

29
Table 3. Estimates of location (µt) and scale (σt) parameters for a normal distribution of a-b ridge count as well as the variance in
the distribution of developmental noise (σa); loglikelihood is 578.0, AIC is 1162.0.
Parameter Estimate Std Error
µT 43.03 0.49
σT 4.29 0.39
σA 3.13 0.13

30
Table 4. Estimates of location (µt) and scale (σt) parameters for a normal distribution of a-b ridge count as well as the variance in
the distribution of developmental noise (σa), effect of prenatal alcohol exposure on the variance in developmental noise (βa);
loglikelihood is 571.7, AIC is 1151.5.
Parameter Estimate Std Error
µT 43.02 0.50
σT 4.31 0.40
σA 2.15 0.24
βa 0.53 0.13

31
Table 5. Estimates of location (µt) and scale (σt) parameters for a normal distribution of a-b ridge count as well as the variance in
the distribution of developmental noise (σa), effect of prenatal alcohol exposure on the variance in developmental noise (βa), and
directional asymmetry (d); loglikelihood is 561.0, AIC is 1132.2.
Parameter Estimate Std Error
µT 43.02 0.49
σT 4.41 0.39
σA 1.99 0.20
βa 0.48 0.12
d 0.87 0.20

32
Figure Captions
Figure 1. The logic of a distribution of developmental noise imposed on a single underlying trait. The distribution of left and right
measurements (L and R, bottom) are constructed from the value of the latent trait (T, top) and the distribution of developmental
noise (A, middle).
33
20 30 40 50 60 70 80Trait value, T
20 30 40 50 60 70 80Left measurement, L
20 30 40 50 60 70 80Right measurement, R
-20 -10 0 10 20Developmental noise, A





























