Embed Size (px)
Citation preview

Transendothelial migration enhances integrin-dependenthuman neutrophil chemokinesis
Anjelica L. Gonzalez,*,1 Wafa El-Bjeirami,* Jennifer L. West,† Larry V. McIntire,‡
and C. Wayne Smith**Section of Leukocyte Biology, Department of Pediatrics, Baylor College of Medicine, Houston, Texas, USA;†Department of Bioengineering, Rice University, Houston, Texas, USA; and ‡Coulter Department of BiomedicalEngineering, Georgia Institute of Technology/Emory University, Atlanta, Georgia, USA
Abstract: Transendothelial migration of neutro-phils induces phenotypic changes that influence theinteractions of neutrophils with extravascular tis-sue components. To assess the influence of trans-migration on neutrophil chemokinetic motility, weused polyethylene glycol hydrogels covalentlymodified with specific peptide sequences relevantto extracellular matrix proteins. We evaluatedformyl-Met-Leu-Phe-stimulated human neutrophilmotility on peptides Arg-Gly-Asp-Ser (RGDS) andTMKIIPFNRTLIGG (P2), alone and in combina-tion. RGDS is a bioactive sequence found in anumber of proteins, and P2 is a membrane-acti-vated complex-1 (Mac-1) ligand located in the�-chain of the fibrinogen protein. We evaluated,via video microscopy, cell motility by measuringcell displacement from origin and total accumu-lated distance traveled and then calculated averagevelocity. Results indicate that although adhesionand shape change were supported by hydrogelscontaining RGD alone, motility was not. Mac-1-dependent motility was supported on hydrogelscontaining P2 alone. Motility was enhancedthrough combined presentation of RGD and P2,engaging Mac-1, �V�3, and �1 integrins. Naı̈veneutrophil motility on combined peptide substrateswas dependent on Mac-1, and �4�1 and �6�1 con-tributed to speed and linear movement. Transmi-grated neutrophil motility was dependent on �v�3and �5�1, and �4�1, �6�1, and Mac-1 contributedto speed and linear motion. Together, the datademonstrate that efficient neutrophil migration,dependent on multi-integrin interaction, is en-hanced after transendothelial migration. J. Leu-koc. Biol. 81: 000–000; 2007.
Key Words: transmigration � �1 integrins � inflammation � extra-cellular matrix � motility � �2 integrin
INTRODUCTION
During inflammation, neutrophils interact with proteins of theextracellular matrix (ECM) after migration through vascularendothelium to the site of tissue injury or infection. These
interactions are mediated through integrins, which are het-erodimers of noncovalently associated � and � subunits be-longing to members of the �1, �2, and �3 families [1]. Here, weinvestigate neutrophil motility mediated by specific interac-tions between neutrophil integrins and active domains of ECMproteins and evaluate functional changes of the neutrophil as aresult of transendothelial migration.
Neutrophil integrin affinity and/or avidity may be stimulatedthrough endogenous and bacterial chemokines and chemotac-tic factors, including IL-8, leukotriene B4 (LTB4), formyl-Met-Leu-Phe (fMLF), and anaphylatoxin C5a. In the vasculature,endothelial-derived chemokines (e.g., CXCL8) may stimulatethe neutrophil to arrest and transmigrate, and tissue-derivedchemotactic factors (e.g., bacterial-released fMLF or C5a) in-duce the neutrophil to migrate through the ECM toward thesource of infection. Neutrophil integrins interact with ECMproteins through binding, which not only allows cell adhesionbut also initiates signal transduction cascades that influenceshape change, chemotaxis, cell proliferation, and survival.These phenomena have only recently been linked to specificintegrin-ligand interactions [2–5]. Recently, we investigatedthe ability of ECM protein sequences Arg-Gly-Asp (RGD) andTMKIIPFNRTLIGG (P2) to promote specific, integrin-medi-ated neutrophil adhesion and spreading [6]. That study usedtissue-engineering techniques to evaluate interactions with theECM peptides. Specifically, (poly)ethylene glycol (PEG) diac-rylate derivatives were used to form hydrogels as a nonadhesivebackground for covalent attachment of bioactive moieties.Those experiments demonstrated that neutrophils show littleability to adhere to unmodified PEG hydrogels, but adhesionand spreading are robust on peptide-modified hydrogels. In-corporating RGD or P2, alone or in combination, has enabledrecognition of differential functions of neutrophil integrins onadhesion and spreading. In this case, the PEG hydrogel servesas an inert background [7], rendered bioactive by includingRGD-Ser (RGDS) [8, 9] and P2 peptides [6]. Combined inter-
1 Correspondence: Baylor College of Medicine, Leukocyte Biology, 1100Bates, Suite 6014, Houston, TX 77030-2600, USA. E-mail:[email protected]
Received September 6, 2006; revised October 6, 2006; accepted November8, 2006.
doi: 10.1189/jlb.0906-553
0741-5400/07/0081-0001 © Society for Leukocyte Biology Journal of Leukocyte Biology Volume 81, March 2007 1
Uncorrected Version. Published on December 12, 2006 as DOI:10.1189/jlb.0906553
Copyright 2006 by The Society for Leukocyte Biology.

actions result in adhesion that differs markedly from that seenwith either integrin engaged independently.
A secondary goal of this study is to evaluate phenotypicchanges of the neutrophil as a result of transendothelial mi-gration. Although investigators have observed changes in in-tegrin expression after transendothelial migration [10, 11],little has been done to investigate the functionality of theseintegrins after transendothelial migration. In this study, weprovide evidence that integrin-mediated motility is alteredfollowing neutrophil transendothelial migration.
MATERIALS AND METHODS
Peptides
RGDS peptide was purchased from American Peptide Company Inc. (Sunny-vale, CA, USA). P2 [12] was synthesized using an automated peptide synthe-sizer (Model 431A, PE Applied Biosystems, Foster City, CA, USA) andstandard fmoc chemistry [6].
Development of biomimetic hydrogels
The biomimetic PEG hydrogels were prepared as described previously [6, 13].Briefly, PEG diacrylate was prepared by combining 0.1 mmol/ml dry PEG(10,000 Da, Fluka, Milwaukee, WI, USA), 0.4 mmol/ml acryloyl chloride, and0.2 mmol/ml triethylamine in anhydrous dichloromethane and stored underargon overnight. The resulting PEG diacrylate was then precipitated with ether,filtered, and dried. PEG diacrylate was dialyzed prior to use.
Peptides were conjugated to PEG monoacrylate by reacting the peptide withacryloyl-PEG-N-hydroxysuccinimide (3400 Da, Nektar, Inc., San Carlos, CA,USA) in sodium bicarbonate (pH 8.5) at a 1:1 molar ratio for 2 h. The coupledacryloyl-PEG peptide was lyophilized, and the conjugated PEG peptide wasdialyzed prior to use. All polymers were characterized by proton nuclearmagnetic resonance (Avance 400 Mhz, Bruker, Billerica, MA, USA) and gelpermeation chromatography with an evaporative light-scattering detector usingPEG standards (Polymer Laboratories, Amherst, MA, USA).
Hydrogels were prepared by combining 0.1 g/ml PEG diacrylate and 0.02g/ml (5.2 �mol/ml) acryloyl-PEG-RGDS, 0.02 g/ml (4.0 �mol/ml) acryloyl-PEG-P2, or a combination of 0.01 g/ml acryloyl-PEG-RGDS and 0.01 g/mlacryloyl-PEG-P2 in PBS (pH 7.2) containing 0.1 g/L glucose. The solution wasthen sterilized by filtration (0.2 �m with 0.8 �m prefilter, Pall Corp., AnnArbor, MI, USA). 2,2-Dimethoxy-2-phenylacetophenone (10 �l/ml) in n-vi-nylpyrrolidone (300 mg/ml) was added as the photoinitiator. The resultingsolution was exposed to UV light (365 nm, 10 mW/cm2) for 30 s to convert theliquid polymer solution to a covalently cross-linked hydrogel. The polymerizedgels were then incubated overnight in PBS to allow them to reach theirequilibrium swelling. PEG diacrylate hydrogels without peptides were used ascontrols.
Endotoxin detection
Endotoxin assays were conducted on polymer solution prior to gelation. TheLimulus amoebocyte lysate (LAL) assay was used to determine endotoxinpresence in each component of the PEG hydrogel. Briefly, a 100-�l sample wasplaced in a sterilized glass tube with 100 �l LAL. This solution was thenplaced on a heating block for 1 h. Endotoxin-negative solutions resulted in nogelation, and endotoxin-positive solutions became gelated. Pyrogen-free water(Baxter Healthcare Corp., Deerfield, IL, USA) served as a negative control, andbacterial LPS served as an endotoxin-positive control. All samples of PEGhydrogel reagents were endotoxin-negative.
Neutrophil isolation
Neutrophils were obtained from the whole blood of healthy human volunteers.The method for isolation has been described previously [14]. Briefly, neutro-phils were purified by 6% Dextran sedimentation to remove RBC and byFicoll-Hypaqe gradient centrifugation to remove remaining leukocytes. The
cells were finally suspended in PBS containing 0.1 g/L glucose to be used ata concentration of 1 � 106 cells/ml.
Antibodies and reagents
Neutrophil-stimulating agent fMLF was obtained from Sigma Chemical Co. (St.Louis, MO, USA) and used at 1 � 10�7 M concentration. Mouse antihumanmembrane-activated complex-1 (Mac-1) antibody, 60.1, was received as agenerous gift from Dr. Lora Whitehouse (Repligen, Cambridge, MA, USA) andused at a concentration of 10 �g/ml [14]. Anti-E-selectin mAb CL3 was usedat a concentration of 10 �g/ml, as described previously [15]. Mouse antihumanintegrin �v�3 mAb, LM609, was purchased from Chemicon International(Temecula, CA, USA) and was used at a concentration of 10 �g/ml [16]. Mouseantihuman antibodies against �4 (CD49d), �5 (CD49e), and �6 (CD49f) werepurchased from BD Biosciences PharMingen (San Jose, CA, USA).
HUVEC culture
HUVEC harvest and culture have been described by Burns et al. [17]. Briefly,HUVEC were harvested from human umbilical veins by collagenase perfusion,according to Huang et al. [18] and pooled, plated, and pretreated with 0.2%gelatin (Difco, Detroit, MI, USA). Monolayers are cultured in a 1:1 mixture ofM199 (Gibco BRL, Grand Island, NY, USA) and medium supplemented with10% FBS and 10% bovine calf serum (Hyclone Laboratories, Inc., Logan, UT,USA), 1% penicillin-streptomycin (Gibco BRL), 1% fungizone (Gibco BRL),1% HEPES buffer (Gibco BRL), 1 �g/ml heparin (Sigma Chemical Co.), and50 �g/ml endothelial cell (EC) growth supplement (Collaborative BiomedicalProducts, Bedford, MA, USA).
Neutrophil transendothelial migration
Primary HUVEC were dissociated using trypsin/EDTA, seeded onto gelatin-coated, 3.0 �m pore polycarbonate Transwell� filters (Corning Inc., Corning,NY, USA), which were placed in matching six-well plates and cultured for 4days. HUVEC monolayers were stimulated with IL-1� (10 U/ml) for 4 h at37°C. Following stimulation, the inserts were washed with HBSS, once trans-ferred to six-well plates coated with a thin layer of 1% agarose, whichfacilitated removal of transmigrated neutrophils for further analysis and min-imized additional activation by the plastic wells. Freshly isolated neutrophilsfrom healthy volunteers were placed on top of the insert at a ratio of 4:1(PMN:endothelial) and allowed to transmigrate through the activated HUVECfor 1 h. Nontransmigrated neutrophils were then collected from the top of theinsert (referred to as EC contacted), and transmigrated neutrophils werecollected from the bottom of the insert. Collected cells were counted and usedfor experimentation.
RNA isolation and RT
Total RNA was isolated from transmigrated and nontransmigrated neutrophilsusing Trizol reagent following the manufacturer’s instructions (Life Technolo-gies/Invitrogen, Carlsbad, CA, USA). RT was performed in a 20-�l finalvolume containing 40 U RT AMV, 50 mA260 units primer random p(dN)6, 20U RNase inhibitor, and 1 mM deoxy-unspecified nucleoside 5�-triphosphates(Roche Applied Sciences, Indianapolis, IN, USA). The mixture was subjectedto 25°C for 10 min and 42°C for 60 min and inactivated at 95°C for 5 min. ThecDNA was analyzed immediately or stored at –20°C until use.
TaqMan� assay-based real-time PCR
Primers and probes for human �4, �5, �6, and 18S were acquired from PEApplied Biosystems (Assays-on-Demand). These predesigned and preopti-mized TaqMan gene expression human sequence-based assays are provided ina 20� format and used according to the manufacturer’s instructions. Thepreformulated assay consists of two unlabeled PCR primers (900 nM each finalconcentration) and a dye-labeled TaqMan� minor groove binder probe (250 nMfinal concentration). Real-time TaqMan PCR systems for �4, �5, or �6 weremultiplexed with 18S (internal standard). Each cDNA sample (1 �L) wasanalyzed. All assays were run in triplicates in a 96-well format plate. Real-timefluorescent detection of PCR products was peformed using an ABI 7500 (PEApplied Biosystems) using the following thermocycling conditions: 1 cycle of50°C for 2 min and 95°C for 10 min; 40 cycles of 95°C for 15 s and 60°C for1 min. Data were analyzed by the sequence detection systems (SDS) software
2 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org

(PE Applied Biosystems). Subsequent analysis was performed on the dataoutput from the SDS software using Microsoft Excel. Relative RNA expressionwas determined using the formula Rel Exp � 2–(��Ct), where, for example,��Ct � (Ct �5–CT 18S). The ratio for �5/18S in transmigrated neutrophilsamples was then normalized to the ratio seen in nontransmigrated samplesand expressed as the means SEM.
Flow cytometric analysis
The following antibodies to human antigens were used: CD49d (�4, PE-conjugated, BD Biosciences PharMingen); CD49e (�5, PE-conjugated, BDBiosciences PharMingen); CD49f (�6, PE-conjugated, BD Biosciences Phar-Mingen); CD11b (Mac-1, PE-conjugated, BD Biosciences PharMingen); av (BDTransduction Laboratories, Franklin Lakes, NJ, USA); Alexa Fluor� 546 (goatantimouse IgG, Molecular Probes, Eugene, OR, USA). PE-conjugated, mouseIgG1 was purchased from BD Biosciences PharMingen.
For flow cytometric analysis, neutrophils (1�106 in a volume of 100 �l PBS)were stained in one step with one of the above fluorochrome-labeled antibodies(1 �g). After incubation with the indicated antibodies for 15 min on ice, thecells were washed three times with cold PBS and finally resuspended in 400�l PBS with 1% paraformaldehyde. Data were collected with a FACScan andanalyzed with CellQuest software (BD Biosciences PharMingen).
Cell motility calculations
Cells were determined adherent to specific peptide-containing hydrogels usinga static adhesion assay described previously [6, 14, 19]. Neutrophils, naı̈ve ortransmigrated, stimulated with 10�7 M fMLF, were seeded on hydrogelscontaining various adhesive peptide sequences. Neutrophils were allowed tosettle on the hydrogel and were videotaped for 15 min while viewed under aninverted microscope. Each videotape was analyzed, and frames were digitizedat one frame every 30 s using Image Pro Plus Version 5.1 (Media Cybernetics,Inc., Silver Springs, MD, USA). Neutrophils were selected randomly in variousviewing fields (n�4 fields per sample, approximately three cells per field). Theanalyzed frame dimensions were calibrated using a micrometer. Using theImage Pro Plus Track Object image analysis option with semiautomatedtracking, we tracked the centroid of each cell over the 15-min time period. Cellcentroid x, y coordinates, displacement from origin (DO), total accumulateddistance (DTOT), and average speed of movement (VTOT), were calculated. DO
was used to determine motility. Cells with DO less than 10 �m, the approxi-mate diameter of one cell, were considered immobile. Cells with DO greaterthan 10 �m were considered mobile. VTOT was calculated as DTOT/15 min. Thetrajectory figures presented are representative of typical cell trajectories underspecific cell treatment and substrate conditions.
Statistical analysis
Data were expressed as mean SE for n 30. At least six individuals wereused. Statistical significance was verified using one-way ANOVA calculations
followed by Bonferroni post-test. Pair-wise differences were tested usingStudent’s t-test. Statistical significance is indicated by *, P � 0.05.
RESULTS
Our previous studies demonstrated that hydrogels containing5.2 �mol/ml RGDS or 4.0 �mol/ml P2 peptides providedoptimal neutrophil adhesion after 10�7 M fMLF stimulation[6]. In addition, gels containing combined 2.6 �mol/ml RGDSand 2.0 �mol/ml P2 provided ligand density for optimal ad-hesion. In this study, we used these concentrations to investi-gate the effect of the adhesive peptides on motility.
RGD alone does not support neutrophil motility
To examine whether adhesion to RGDS facilitated motility,stimulated neutrophils on hydrogels containing 2.6 or 5.2�mol/ml RGDS were tracked over a 15-min period after con-tact. Although naive neutrophils (i.e., isolated blood neutro-phils without contact with EC) were adherent and able toundergo shape change on RGD, they did not translocate overthe surface of the hydrogel. Neutrophils, which were immobilewhen seeded on hydrogels, contain 2.6 �mol/ml (data notshown) or 5.2 �mol/ml RGDS (Fig. 1). This failure of loco-motion on RGD-containing hydrogels was also observed withneutrophils that had been exposed for 1 h to monolayers ofactivated EC. Neither the neutrophils collected from the lowercompartment of the chambers (i.e., the transmigrated cells) northose removed from the apical surface (i.e., the nontransmi-grated cells) were migratory (Fig. 1A). Motility in this instancewas determined by displacement (DO) of the neutrophil fromthe point of initial contact. Neutrophils with DO � 10 �m over15 min were considered immobile (Fig. 2A). These observa-tions were consistent among neutrophil samples and treat-ments, and �10% of all neutrophils on RGDS were mobile(Fig. 2B). The consistency of these data indicates that the RGDpeptide alone was not sufficient to support chemokinesis ofnaı̈ve neutrophils or those after contact with activated EC.
Fig. 1. Neutrophil chemokinesis on hydrogels containing RGD, P2, or RGDS and P2 peptide. Neutrophils were collected 1 h after transmigration throughmonolayers of activated HUVEC (IL-1� for 4 h prior to contact of neutrophils). The neutrophils were washed in balanced salt solution and then placed on hydrogelscontaining 5.2 �mol/ml RGDS (A), 4.0 �mol/ml P2 (B), or 2.6 �mol/ml RGDS � 2.0 �mol/ml P2 (C). Chemokinesis induced by addition of fMLF to the mediumwas tracked for 15 min.
Gonzalez et al. Human neutrophil transmigration enhances motility 3
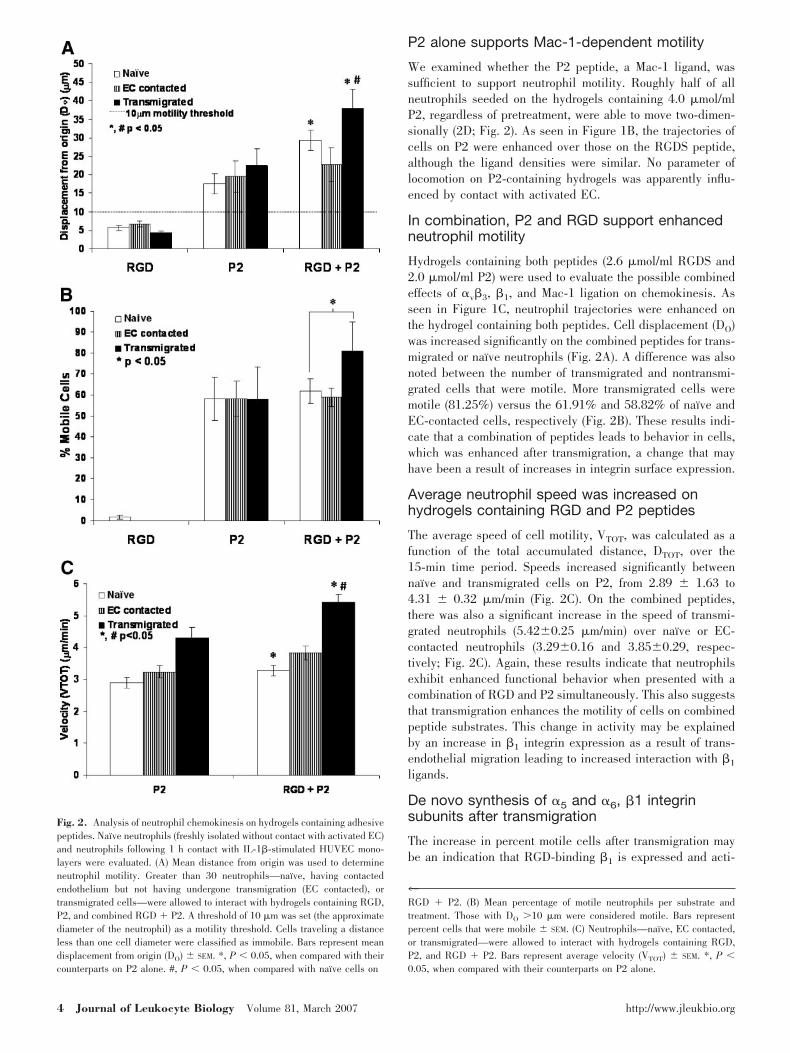
P2 alone supports Mac-1-dependent motility
We examined whether the P2 peptide, a Mac-1 ligand, wassufficient to support neutrophil motility. Roughly half of allneutrophils seeded on the hydrogels containing 4.0 �mol/mlP2, regardless of pretreatment, were able to move two-dimen-sionally (2D; Fig. 2). As seen in Figure 1B, the trajectories ofcells on P2 were enhanced over those on the RGDS peptide,although the ligand densities were similar. No parameter oflocomotion on P2-containing hydrogels was apparently influ-enced by contact with activated EC.
In combination, P2 and RGD support enhancedneutrophil motility
Hydrogels containing both peptides (2.6 �mol/ml RGDS and2.0 �mol/ml P2) were used to evaluate the possible combinedeffects of �v�3, �1, and Mac-1 ligation on chemokinesis. Asseen in Figure 1C, neutrophil trajectories were enhanced onthe hydrogel containing both peptides. Cell displacement (DO)was increased significantly on the combined peptides for trans-migrated or naı̈ve neutrophils (Fig. 2A). A difference was alsonoted between the number of transmigrated and nontransmi-grated cells that were motile. More transmigrated cells weremotile (81.25%) versus the 61.91% and 58.82% of naı̈ve andEC-contacted cells, respectively (Fig. 2B). These results indi-cate that a combination of peptides leads to behavior in cells,which was enhanced after transmigration, a change that mayhave been a result of increases in integrin surface expression.
Average neutrophil speed was increased onhydrogels containing RGD and P2 peptides
The average speed of cell motility, VTOT, was calculated as afunction of the total accumulated distance, DTOT, over the15-min time period. Speeds increased significantly betweennaı̈ve and transmigrated cells on P2, from 2.89 1.63 to4.31 0.32 �m/min (Fig. 2C). On the combined peptides,there was also a significant increase in the speed of transmi-grated neutrophils (5.420.25 �m/min) over naı̈ve or EC-contacted neutrophils (3.290.16 and 3.850.29, respec-tively; Fig. 2C). Again, these results indicate that neutrophilsexhibit enhanced functional behavior when presented with acombination of RGD and P2 simultaneously. This also suggeststhat transmigration enhances the motility of cells on combinedpeptide substrates. This change in activity may be explainedby an increase in �1 integrin expression as a result of trans-endothelial migration leading to increased interaction with �1
ligands.
De novo synthesis of �5 and �6, �1 integrinsubunits after transmigration
The increase in percent motile cells after transmigration maybe an indication that RGD-binding �1 is expressed and acti-
Fig. 2. Analysis of neutrophil chemokinesis on hydrogels containing adhesivepeptides. Naı̈ve neutrophils (freshly isolated without contact with activated EC)and neutrophils following 1 h contact with IL-1�-stimulated HUVEC mono-layers were evaluated. (A) Mean distance from origin was used to determineneutrophil motility. Greater than 30 neutrophils—naı̈ve, having contactedendothelium but not having undergone transmigration (EC contacted), ortransmigrated cells—were allowed to interact with hydrogels containing RGD,P2, and combined RGD � P2. A threshold of 10 �m was set (the approximatediameter of the neutrophil) as a motility threshold. Cells traveling a distanceless than one cell diameter were classified as immobile. Bars represent meandisplacement from origin (DO) SEM. *, P � 0.05, when compared with theircounterparts on P2 alone. #, P � 0.05, when compared with naı̈ve cells on
4RGD � P2. (B) Mean percentage of motile neutrophils per substrate andtreatment. Those with DO 10 �m were considered motile. Bars representpercent cells that were mobile SEM. (C) Neutrophils—naı̈ve, EC contacted,or transmigrated—were allowed to interact with hydrogels containing RGD,P2, and RGD � P2. Bars represent average velocity (VTOT) SEM. *, P �0.05, when compared with their counterparts on P2 alone.
4 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org
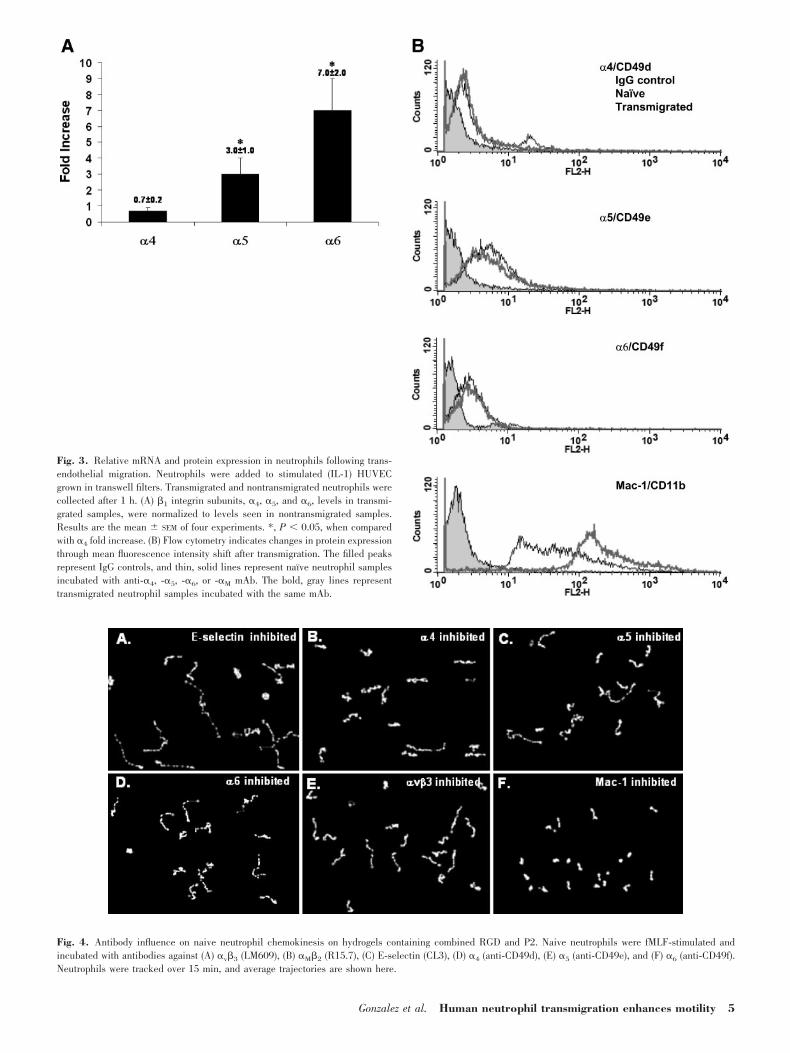
Fig. 3. Relative mRNA and protein expression in neutrophils following trans-endothelial migration. Neutrophils were added to stimulated (IL-1) HUVECgrown in transwell filters. Transmigrated and nontransmigrated neutrophils werecollected after 1 h. (A) �1 integrin subunits, �4, �5, and �6, levels in transmi-grated samples, were normalized to levels seen in nontransmigrated samples.Results are the mean SEM of four experiments. *, P � 0.05, when comparedwith �4 fold increase. (B) Flow cytometry indicates changes in protein expressionthrough mean fluorescence intensity shift after transmigration. The filled peaksrepresent IgG controls, and thin, solid lines represent naı̈ve neutrophil samplesincubated with anti-�4, -�5, -�6, or -�M mAb. The bold, gray lines representtransmigrated neutrophil samples incubated with the same mAb.
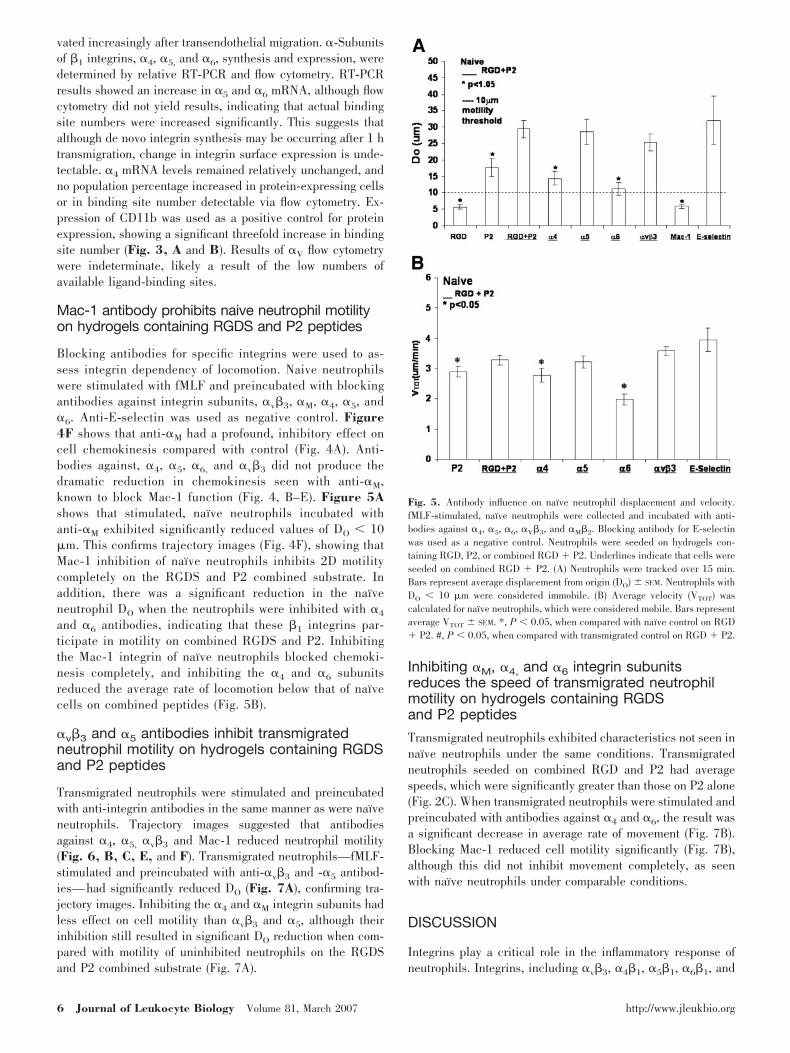
Fig. 4. Antibody influence on naive neutrophil chemokinesis on hydrogels containing combined RGD and P2. Naive neutrophils were fMLF-stimulated andincubated with antibodies against (A) �v�3 (LM609), (B) �M�2 (R15.7), (C) E-selectin (CL3), (D) �4 (anti-CD49d), (E) �5 (anti-CD49e), and (F) �6 (anti-CD49f).Neutrophils were tracked over 15 min, and average trajectories are shown here.
Gonzalez et al. Human neutrophil transmigration enhances motility 5
vated increasingly after transendothelial migration. �-Subunitsof �1 integrins, �4, �5, and �6, synthesis and expression, weredetermined by relative RT-PCR and flow cytometry. RT-PCRresults showed an increase in �5 and �6 mRNA, although flowcytometry did not yield results, indicating that actual bindingsite numbers were increased significantly. This suggests thatalthough de novo integrin synthesis may be occurring after 1 htransmigration, change in integrin surface expression is unde-tectable. �4 mRNA levels remained relatively unchanged, andno population percentage increased in protein-expressing cellsor in binding site number detectable via flow cytometry. Ex-pression of CD11b was used as a positive control for proteinexpression, showing a significant threefold increase in bindingsite number (Fig. 3, A and B). Results of �V flow cytometrywere indeterminate, likely a result of the low numbers ofavailable ligand-binding sites.
Mac-1 antibody prohibits naive neutrophil motilityon hydrogels containing RGDS and P2 peptides
Blocking antibodies for specific integrins were used to as-sess integrin dependency of locomotion. Naive neutrophilswere stimulated with fMLF and preincubated with blockingantibodies against integrin subunits, �v�3, �M, �4, �5, and�6. Anti-E-selectin was used as negative control. Figure4F shows that anti-�M had a profound, inhibitory effect oncell chemokinesis compared with control (Fig. 4A). Anti-bodies against, �4, �5, �6, and �v�3 did not produce thedramatic reduction in chemokinesis seen with anti-�M,known to block Mac-1 function (Fig. 4, B–E). Figure 5Ashows that stimulated, naı̈ve neutrophils incubated withanti-�M exhibited significantly reduced values of DO � 10�m. This confirms trajectory images (Fig. 4F), showing thatMac-1 inhibition of naı̈ve neutrophils inhibits 2D motilitycompletely on the RGDS and P2 combined substrate. Inaddition, there was a significant reduction in the naı̈veneutrophil DO when the neutrophils were inhibited with �4
and �6 antibodies, indicating that these �1 integrins par-ticipate in motility on combined RGDS and P2. Inhibitingthe Mac-1 integrin of naı̈ve neutrophils blocked chemoki-nesis completely, and inhibiting the �4 and �6 subunitsreduced the average rate of locomotion below that of naı̈vecells on combined peptides (Fig. 5B).
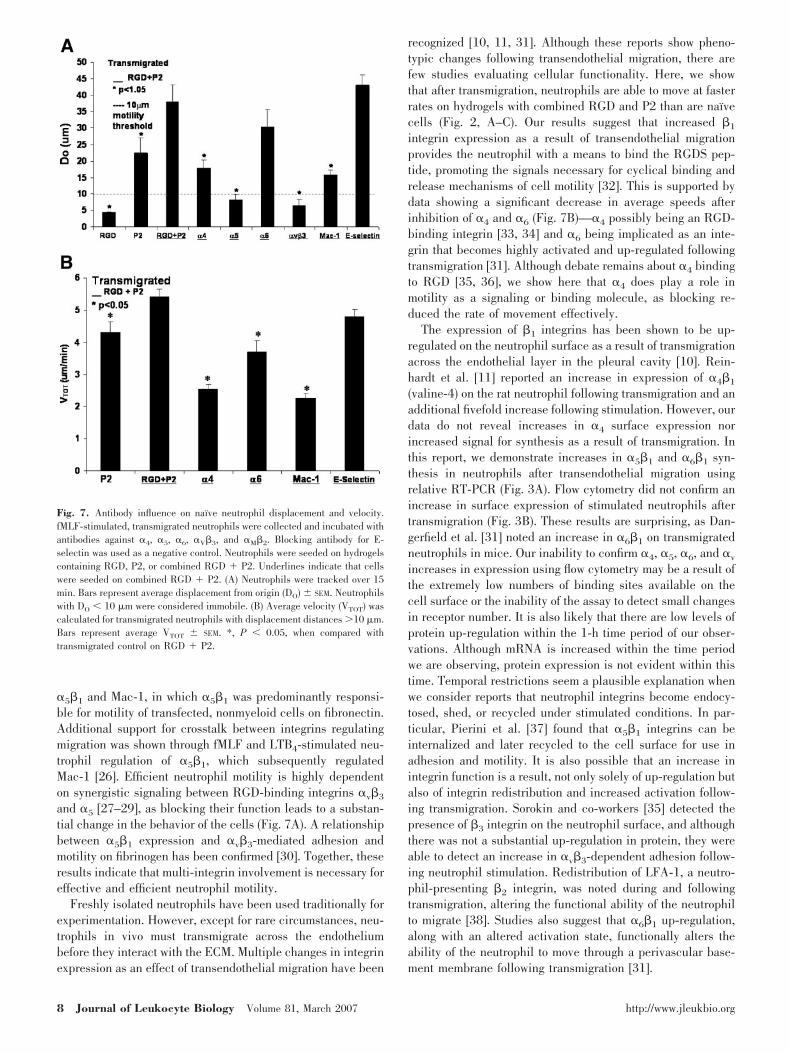
�v�3 and �5 antibodies inhibit transmigratedneutrophil motility on hydrogels containing RGDSand P2 peptides
Transmigrated neutrophils were stimulated and preincubatedwith anti-integrin antibodies in the same manner as were naı̈veneutrophils. Trajectory images suggested that antibodiesagainst �4, �5, �v�3 and Mac-1 reduced neutrophil motility(Fig. 6, B, C, E, and F). Transmigrated neutrophils—fMLF-stimulated and preincubated with anti-�v�3 and -�5 antibod-ies—had significantly reduced DO (Fig. 7A), confirming tra-jectory images. Inhibiting the �4 and �M integrin subunits hadless effect on cell motility than �v�3 and �5, although theirinhibition still resulted in significant DO reduction when com-pared with motility of uninhibited neutrophils on the RGDSand P2 combined substrate (Fig. 7A).
Inhibiting �M, �4, and �6 integrin subunitsreduces the speed of transmigrated neutrophilmotility on hydrogels containing RGDSand P2 peptides
Transmigrated neutrophils exhibited characteristics not seen innaı̈ve neutrophils under the same conditions. Transmigratedneutrophils seeded on combined RGD and P2 had averagespeeds, which were significantly greater than those on P2 alone(Fig. 2C). When transmigrated neutrophils were stimulated andpreincubated with antibodies against �4 and �6, the result wasa significant decrease in average rate of movement (Fig. 7B).Blocking Mac-1 reduced cell motility significantly (Fig. 7B),although this did not inhibit movement completely, as seenwith naı̈ve neutrophils under comparable conditions.
DISCUSSION
Integrins play a critical role in the inflammatory response ofneutrophils. Integrins, including �v�3, �4�1, �5�1, �6�1, and
Fig. 5. Antibody influence on naı̈ve neutrophil displacement and velocity.fMLF-stimulated, naı̈ve neutrophils were collected and incubated with anti-bodies against �4, �5, �6, �V�3, and �M�2. Blocking antibody for E-selectinwas used as a negative control. Neutrophils were seeded on hydrogels con-taining RGD, P2, or combined RGD � P2. Underlines indicate that cells wereseeded on combined RGD � P2. (A) Neutrophils were tracked over 15 min.Bars represent average displacement from origin (DO) SEM. Neutrophils withDO � 10 �m were considered immobile. (B) Average velocity (VTOT) wascalculated for naı̈ve neutrophils, which were considered mobile. Bars representaverage VTOT SEM. *, P � 0.05, when compared with naı̈ve control on RGD� P2. #, P � 0.05, when compared with transmigrated control on RGD � P2.
6 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org

�M�2, are responsible for binding ECM proteins and formingthe adhesive traction forces necessary for migration. Thesemolecules become activated and able to bind their ligands inresponse to conformational changes mediated through chemo-kines and chemotactic factors. fMLF, for example, stimulatesthe neutrophil through p38 MAPK to elicit Mac-1 activationand chemotaxis and �4�1-dependent polarization [20]. UsingfMLF as a stimulant, we were able to evaluate adhesive mech-anisms that support chemokinesis by presenting specific inte-grin ligands. In addition, we were able to determine functionalchanges in neutrophil integrins following transmigration. Thedata presented support the hypothesis that neutrophil chemo-kinesis is mediated by specific interactions between neutrophilintegrins and bioactive domains of ECM proteins. This motilityis enhanced by neutrophil transendothelial migration. Specif-ically, we show that neutrophil 2D chemokinesis is supportedby the Mac-1 ligand, P2, and is enhanced through multi-integrin interactions with RGD and P2 (Figs. 1 and 2). The dataalso demonstrate that phenotypic changes as a result of trans-endothelial migration enhance chemokinesis on a multipep-tide-presenting substrate (Fig. 2, A–C).
In this study we use PEG hydrogel as an inert substrate toeliminate the issues of multiple ligand-binding, nonspecificbinding, and nonspecific activation. We previously publishedthat neutrophil adhesion to PEG was induced through RGDS,supporting adhesion and spreading through �v�3 and �1 inte-grins [6]. In the present study, we find that these firmly adher-ent and widely spread neutrophils are immobile. In this case,�v�3-mediated adhesion and �1-mediated spreading on RGDare not sufficient to promote neutrophil motility (Figs. 1A and2, A and B). Our observations coincide with others who ob-served that the mobility of fMLF-stimulated neutrophils wasinversely related to adhesion and increased spreading on glass,
polyurethane, and octadecyltrichlorosilane [21]. Additionalstudies showed that firm adhesions via �v�3 clusters in the rearof the cell rendered the neutrophil incapable of migration onvitronectin [22]. Clearly, RGD-mediated firm adhesion,whether �V�3- or �1-dependent, is not sufficient to supportmotility.
Unlike RGD, ligation of Mac-1 via P2 is sufficient for 2Dmotility (Figs. 1B and 2, A–C). Others similarly report thatMac-1-dependent motility is possible in the absence of sec-ondary integrin involvement [2, 12, 23, 24]. Such Mac-1-dependent migration has been seen in the lung [2], throughintestinal epithelium [23] and through matrices of fibrinogenand collagen [12, 24]. To the best of our knowledge, this is thefirst instance in which Mac-1-dependent motility has beenshown using a single peptide under isolated conditions, asopposed to whole protein or antibody inhibition of secondaryintegrin ligation. Although Mac-1 integrin-dependent motilityis possible, our results confirm that the addition of RGD tohydrogels containing P2 promotes the involvement of the �v�3
and �1 integrins, prompting neutrophils to move at increasedrates (Figs. 1C and 2, A–C). Integrin-dependent motility is aresult of a balance between cyclical binding and release. Thisbalance allows the cell to form adhesive traction forces to pullit, while releasing from the substrate to allow forward advance-ment. As we see, on combined RGD and P2 hydrogels, thisbalance is preserved, and on RGD alone, it is not, makingmotility impossible. Our results also indicate that engagementof the �1 integrins is not inhibitory, demonstrated by increasesin displacement from the origin on surfaces with RGD and P2together (Figs. 2A and 5A). Again, this suggests that �1 andMac-1 crosstalk provides optimum signaling for efficient mo-tility. Ugarova and co-workers [25] support the idea of integrincrosstalk through observations of functional interplay between
Fig. 6. Antibody influence on transmigrated neutrophil chemokinesis on hydrogels containing combined RGD and P2. Transmigrated neutrophils werefMLF-stimulated and incubated with antibodies against (A) �v�3 (LM609), (B) �M�2 (R15.7), (C) E-selectin (CL3), (D) �4 (anti-CD49d), (E) �5 (anti-CD49e), and(F) �6 (anti-CD49f). Neutrophils were tracked over 15 min, and average trajectories are shown here.
Gonzalez et al. Human neutrophil transmigration enhances motility 7
�5�1 and Mac-1, in which �5�1 was predominantly responsi-ble for motility of transfected, nonmyeloid cells on fibronectin.Additional support for crosstalk between integrins regulatingmigration was shown through fMLF and LTB4-stimulated neu-trophil regulation of �5�1, which subsequently regulatedMac-1 [26]. Efficient neutrophil motility is highly dependenton synergistic signaling between RGD-binding integrins �v�3
and �5 [27–29], as blocking their function leads to a substan-tial change in the behavior of the cells (Fig. 7A). A relationshipbetween �5�1 expression and �v�3-mediated adhesion andmotility on fibrinogen has been confirmed [30]. Together, theseresults indicate that multi-integrin involvement is necessary foreffective and efficient neutrophil motility.
Freshly isolated neutrophils have been used traditionally forexperimentation. However, except for rare circumstances, neu-trophils in vivo must transmigrate across the endotheliumbefore they interact with the ECM. Multiple changes in integrinexpression as an effect of transendothelial migration have been
recognized [10, 11, 31]. Although these reports show pheno-typic changes following transendothelial migration, there arefew studies evaluating cellular functionality. Here, we showthat after transmigration, neutrophils are able to move at fasterrates on hydrogels with combined RGD and P2 than are naı̈vecells (Fig. 2, A–C). Our results suggest that increased �1
integrin expression as a result of transendothelial migrationprovides the neutrophil with a means to bind the RGDS pep-tide, promoting the signals necessary for cyclical binding andrelease mechanisms of cell motility [32]. This is supported bydata showing a significant decrease in average speeds afterinhibition of �4 and �6 (Fig. 7B)—�4 possibly being an RGD-binding integrin [33, 34] and �6 being implicated as an inte-grin that becomes highly activated and up-regulated followingtransmigration [31]. Although debate remains about �4 bindingto RGD [35, 36], we show here that �4 does play a role inmotility as a signaling or binding molecule, as blocking re-duced the rate of movement effectively.
The expression of �1 integrins has been shown to be up-regulated on the neutrophil surface as a result of transmigrationacross the endothelial layer in the pleural cavity [10]. Rein-hardt et al. [11] reported an increase in expression of �4�1
(valine-4) on the rat neutrophil following transmigration and anadditional fivefold increase following stimulation. However, ourdata do not reveal increases in �4 surface expression norincreased signal for synthesis as a result of transmigration. Inthis report, we demonstrate increases in �5�1 and �6�1 syn-thesis in neutrophils after transendothelial migration usingrelative RT-PCR (Fig. 3A). Flow cytometry did not confirm anincrease in surface expression of stimulated neutrophils aftertransmigration (Fig. 3B). These results are surprising, as Dan-gerfield et al. [31] noted an increase in �6�1 on transmigratedneutrophils in mice. Our inability to confirm �4, �5, �6, and �v
increases in expression using flow cytometry may be a result ofthe extremely low numbers of binding sites available on thecell surface or the inability of the assay to detect small changesin receptor number. It is also likely that there are low levels ofprotein up-regulation within the 1-h time period of our obser-vations. Although mRNA is increased within the time periodwe are observing, protein expression is not evident within thistime. Temporal restrictions seem a plausible explanation whenwe consider reports that neutrophil integrins become endocy-tosed, shed, or recycled under stimulated conditions. In par-ticular, Pierini et al. [37] found that �5�1 integrins can beinternalized and later recycled to the cell surface for use inadhesion and motility. It is also possible that an increase inintegrin function is a result, not only solely of up-regulation butalso of integrin redistribution and increased activation follow-ing transmigration. Sorokin and co-workers [35] detected thepresence of �3 integrin on the neutrophil surface, and althoughthere was not a substantial up-regulation in protein, they wereable to detect an increase in �v�3-dependent adhesion follow-ing neutrophil stimulation. Redistribution of LFA-1, a neutro-phil-presenting �2 integrin, was noted during and followingtransmigration, altering the functional ability of the neutrophilto migrate [38]. Studies also suggest that �6�1 up-regulation,along with an altered activation state, functionally alters theability of the neutrophil to move through a perivascular base-ment membrane following transmigration [31].
Fig. 7. Antibody influence on naı̈ve neutrophil displacement and velocity.fMLF-stimulated, transmigrated neutrophils were collected and incubated withantibodies against �4, �5, �6, �V�3, and �M�2. Blocking antibody for E-selectin was used as a negative control. Neutrophils were seeded on hydrogelscontaining RGD, P2, or combined RGD � P2. Underlines indicate that cellswere seeded on combined RGD � P2. (A) Neutrophils were tracked over 15min. Bars represent average displacement from origin (DO) SEM. Neutrophilswith DO � 10 �m were considered immobile. (B) Average velocity (VTOT) wascalculated for transmigrated neutrophils with displacement distances 10 �m.Bars represent average VTOT SEM. *, P � 0.05, when compared withtransmigrated control on RGD � P2.
8 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org

Our studies closely simulate the results of others who haveobserved neutrophils on a HUVEC monolayer moving at ratesof 6 �m/min, although their observation of transmigratedcells below the HUVEC monolayer was at a highly increasedrate of 14 �m/min [39]. The diminished velocities observed inour experiments with transmigrated cells could be a result ofthe texture of the hydrogel surface. The choice of PEG as thepresenting substrate was based on its inert characteristic,resistance to protein adsorption, and relatively easy chemicalmanipulation. As suggested by Tan et al. [40–42], however,the topographical geometry and chemistry of a substrate canhave a dramatic effect on the rate of movement of neutrophils.As our polymer did not vary from study to study, this effectdoes not hinder our ability to observe the differential effects ofmotility as a result of specific integrin and combined integrininteractions.
In summary, the results of this study demonstrate that RGD,as a single substrate for adhesion, is insufficient to supportleukocyte chemokinesis, but the peptide P2 as a single sub-strate for adhesion is sufficient to support Mac-1-dependentchemokinesis. When both peptides are present, neutrophillocomotion is enhanced significantly and dependent on a com-bination of leukocyte integrins. In addition, the results dem-onstrate that neutrophil chemokinetic motility is increasedsignificantly within 1 h of transendothelial migration, and onsubstrates containing RGD and P2 peptides, the increasedlocomotion is dependent on a combination of leukocyte inte-grins, predominately �v�3 and �5�1.
ACKNOWLEDGMENTS
This work was supported by funding from the National Insti-tutes of Health (HL070537, HL007939, HL18672, USDA6250-5100-046). The writers acknowledge the nursing staff atthe Labor and Delivery Units of The Woman’s Hospital ofTexas and the Ben Taub General Hospital for aid in acquiringumbilical cords.
REFERENCES
1. van der Flier, A., Sonnenberg, A. (2001) Function and interactions ofintegrins. Cell Tissue Res. 305, 285–298.
2. Doerschuk, C. M., Winn, R. K., Coxson, H. O., Harlan, J. M. (1990)Cd18-dependent and Cd18-independent mechanisms of neutrophil emi-gration in the pulmonary and systemic microcirculation of rabbits. J. Im-munol. 144, 2327–2333.
3. Luscinskas, F. W., Ding, H., Lichtman, A. H. (1995) P-Selectin andvascular cell-adhesion molecule-1 mediate rolling and arrest, respec-tively, of Cd4(�) T-lymphocytes on tumor-necrosis-factor �-activatedvascular endothelium under flow. J. Exp. Med. 181, 1179–1186.
4. Mileski, W., Harlan, J., Rice, C., Winn, R. (1990) Streptococcus pneu-moniae-stimulated macrophages induce neutrophils to emigrate by aCd18-independent mechanism of adherence. Circ. Shock 31, 259–267.
5. Norman, M. U., Kubes, P. (2005) Therapeutic intervention in inflammatorydiseases: a time and place for anti-adhesion therapy. Microcirculation 12,91–98.
6. Gonzalez, A. L., Gobin, A. S., West, J. L., McIntire, L. V., Smith, C. W.(2004) Integrin interactions with immobilized peptides in polyethyleneglycol diacrylate hydrogels. Tissue Eng. 10, 1775–1786.
7. Gombotz, W. R., Guanghui, W., Horbett, T. A., Hoffman, A. S. (1991)Protein adsorption to poly(ethylene oxide) surfaces. J. Biomed. Mater. Res.25, 1547–1562.
8. Gobin, A. S., West, J. L. (2002) Cell migration through defined, syntheticextracellular matrix analogues. FASEB J. 16, 751–753.
9. Mann, B. K., Gobin, A. S., Tsai, A. T., Schmedlen, R. H., West, J. L.(2001) Smooth muscle cell growth in photopolymerized hydrogels with celladhesive and proteolytically degradable domains: synthetic ECM analogsfor tissue engineering. Biomaterials 22, 3045–3051.
10. Teng, R., Johkura, K., Ogiwara, N., Zhao, X., Cui, L., Iida, I., Okouchi, Y.,Asanuma, K., Sasaki, K. (2003) Morphological analysis of leucocytetransmigration in the pleural cavity. J. Anat. 203, 391–404.
11. Reinhardt, P. H., Ward, C. A., Giles, W. R., Kubes, P. (1997) Emigratedrat neutrophils adhere to cardiac myocytes via �(4) integrin. Circ. Res. 81,196–201.
12. Forsyth, C. B., Solovjov, D. A., Ugarova, T. P., Plow, E. F. (2001) Integrin�(M)�(2)-mediated cell migration to fibrinogen and its recognition pep-tides. J. Exp. Med. 193, 1123–1133.
13. Gobin, A. S., West, J. L. (2003) Effects of epidermal growth factor onfibroblast migration through biomimetic hydrogels. Biotechnol. Prog. 19,1781–1785.
14. Gopalan, P. K., Burns, A. R., Simon, S. I., Sparks, S., McIntire, L. V.,Smith, C. W. (2000) Preferential sites for stationary adhesion of neutro-phils to cytokine-stimulated HUVEC under flow conditions. J. Leukoc.Biol. 68, 47–57.
15. Kishimoto, T. K., Warnock, R. A., Jutila, M. A., Butcher, E. C., Lane, C.,Anderson, D. C., Smith, C. W. (1991) Antibodies against human neutro-phil Lecam-1 (Lam-1/Leu-8/Dreg-56 antigen) and endothelial-cell Elam-1inhibit a common Cd18-independent adhesion pathway in vitro. Blood 78,805–811.
16. Cheresh, D. A., Spiro, R. C. (1987) Biosynthetic and functional propertiesof an Arg-Gly-Asp-directed receptor involved in human melanoma cellattachment to vitronectin, fibrinogen, and von Willebrand factor. J. Biol.Chem. 262, 17703–17711.
17. Burns, A. R., Bowden, R. A., MacDonell, S. D., Walker, D. C., Odebunmi,T. O., Donnachie, E. M., Simon, S. I., Entman, M. L., Smith, C. W. (2000)Analysis of tight junctions during neutrophil transendothelial migration.J. Cell Sci. 113, 45–57.
18. Huang, A. J., Furie, M. B., Nicholson, S. C., Fischbarg, J., Liebovitch,L. S., Silverstein, S. C. (1988) Effects of human neutrophil chemotaxisacross human endothelial cell monolayers on the permeability of thesemonolayers to ions and macromolecules. J. Cell. Physiol. 135, 355–366.
19. Smith, C. W., Hollers, J. C., Patrick, R. A., Hassett, C. (1979) Motility andadhesiveness in human neutrophils—effects of chemotactic factors.J. Clin. Invest. 63, 221–229.
20. Heit, B., Colarusso, P., Kubes, P. (2005) Fundamentally different roles forLFA-1, Mac-1 and �(4)-integrin in neutrophil chemotaxis. J. Cell Sci.118, 5205–5220.
21. Zhou, Y., Doerschuk, C. M., Anderson, J. M., Marchant, R. E. (2004)Biomaterial surface-dependent neutrophil mobility. J. Biomed. Mater. Res.A 69, 611–620.
22. Lawson, M. A., Maxfield, F. R. (1995) Ca2� and calcineurin-dependentrecycling of an integrin to the front of migrating neutrophils. Nature 377,75–79.
23. Blake, K. M., Carrigan, S. O., Issekutz, A. C., Stadnyk, A. W. (2004)Neutrophils migrate across intestinal epithelium using �(2) integrin(CD11b/CD18)-independent mechanisms. Clin. Exp. Immunol. 136,262–268.
24. Saltzman, W. M., Livingston, T. L., Parkhurst, M. R. (1999) Antibodies toCD18 influence neutrophil migration through extracellular matrix. J. Leu-koc. Biol. 65, 356–363.
25. Lishko, V. K., Yakubenko, V. P., Ugarova, T. P. (2003) The interplaybetween integrins �(M)�(2) and �(5)�(1), during cell migration to fi-bronectin. Exp. Cell Res. 283, 116–126.
26. Loike, J. D., Cao, L., Budhu, S., Marcantonio, E. E., El Khoury, J.,Hoffman, S., Yednock, T. A., Silverstein, S. C. (1999) Differential regu-lation of �(1) integrins by chemoattractants regulates neutrophil migrationthrough fibrin. J. Cell Biol. 144, 1047–1056.
27. Ruoslahti, E. (1996) RGD and other recognition sequences for integrins.Annu. Rev. Cell Dev. Biol. 12, 697–715.
28. Sun, Z., Martinez-Lemus, L. A., Trache, A., Trzeciakowski, J. P., Davis,G. E., Pohl, U., Meininger, G. A. (2005) Mechanical properties of theinteraction between fibronectin and �(5)�(1)-integrin on vascular smoothmuscle cells studied using atomic force microscopy. Am. J. Physiol. HeartCirc. Physiol. 289, H2526–H2535.
29. Garcia, A. J., Schwarzbauer, J. E., Boettiger, D. (2002) Distinct activationstates of � 5 � 1 integrin show differential binding to RGD and synergydomains of fibronectin. Biochemistry 41, 9063–9069.
30. Ly, D. P., Zazzali, K. M., Corbett, S. A. (2003) De novo expression of theIntegrin �(5)�(1) regulates �(v)�(3)-mediated adhesion and migration onfibrinogen. J. Biol. Chem. 278, 21878–21885.
Gonzalez et al. Human neutrophil transmigration enhances motility 9

31. Dangerfield, J., Larbi, K. Y., Huang, M. T., Dewar, A., Nourshargh, S.(2002) PECAM-1 (CD31) homophilic interaction up-regulates �(6)�(1) ontransmigrated neutrophils in vivo and plays a functional role in the abilityof �(6) integrins to mediate leukocyte migration through the perivascularbasement membrane. J. Exp. Med. 196, 1201–1211.
32. Vitte, J., Benoliel, A. M., Eymeric, P., Bongrand, P., Pierres, A. (2004)�-1 Integrin-mediated adhesion may be initiated by multiple incompletebonds, thus accounting for the functional importance of receptor cluster-ing. Biophys. J. 86, 4059–4074.
33. Yin, Z., Giacomello, E., Gabriele, E., Zardi, L., Aota, S., Yamada, K. M.,Skerlavaji, B., Doliana, R., Colombatti, A., Perris, R. (1999) Cooperativeactivity of � 4 � 1 and � 4 � 7 integrins in mediating human B-celllymphoma adhesion and chemotaxis on fibronectin through recognition ofmultiple synergizing binding sites within the central cell-binding domain.Blood 93, 1221–1230.
34. Brittain, J. E., Han, J., Ataga, K. I., Orringer, E. P., Parise, L. V. (2004)Mechanism of CD47-induced �(4)�(1) integrin activation and adhesion insickle reticulocytes. J. Biol. Chem. 279, 42393–42402.
35. Sixt, M., Hallmann, R., Wendler, O., Scharffetter-Kochanek, K., Sorokin,L. M. (2001) Cell adhesion and migration properties of �(2)-integrinnegative polymorphonuclear granulocytes on defined extracellular matrixmolecules. Relevance for leukocyte extravasation. J. Biol. Chem. 276,18878–18887.
36. Sechler, J. L., Cumiskey, A. M., Gazzola, D. M., Schwarzbauer, J. E.(2000) A novel RGD-independent fibronectin assembly pathway initiatedby � 4 � 1 integrin binding to the alternatively spliced V region. J. CellSci. 113, 1491–1498.
37. Pierini, L. M., Lawson, M. A., Eddy, R. J., Hendey, B., Maxfield, F. R.(2000) Oriented endocytic recycling of � 5 � l in motile neutrophils. Blood95, 2471–2481.
38. Shaw, S. K., Ma, S., Kim, M. B., Rao, R. M., Hartman, C. U., Froio, R. M.,Yang, L., Jones, T., Liu, Y., Nusrat, A., Parkos, C. A., Luscinskas, F. W.(2004) Coordinated redistribution of leukocyte LFA-1 and endothelial cellICAM-1 accompany neutrophil transmigration. J. Exp. Med. 200, 1571–1580.
39. Luu, N. T., Rainger, G. E., Buckley, C. D., Nash, G. B. (2003) CD31regulates direction and rate of neutrophil migration over and under endo-thelial cells. J. Vasc. Res. 40, 467–479.
40. Tan, J., Saltzman, W. M. (2002) Topographical control of human neutro-phil motility on micropatterned materials with various surface chemistry.Biomaterials 23, 3215–3225.
41. Tan, J., Saltzman, W. M. (1999) Influence of synthetic polymers onneutrophil migration in three-dimensional collagen gels. J. Biomed. Mater.Res. 46, 465–474.
42. Tan, J., Shen, H., Saltzman, W. M. (2001) Micron-scale positioning offeatures influences the rate of polymorphonuclear leukocyte migration.Biophys. J. 81, 2569–2579.
10 Journal of Leukocyte Biology Volume 81, March 2007 http://www.jleukbio.org





























