Embed Size (px)
Citation preview

Cell Tiss. Res., 153, 497--515 (1974) �9 by Springer-Verlag 1974
Transformations of Hypothalamic Arcuate Neurons
I . Changes A s s o c i a t e d w i th S tages o f t h e E s t r o u s Cycle*
J. C. K i n g and T. H. Wil l iams
Department of Anatomy, College of Medicine, University of Iowa, Iowa City, Iowa, USA
A. A. Geral l
Department of Psychology, Tulane University, New Orleans, La., USA
Received July 15, 1974
Summary. Changes in subcellular structures of arcuate neurons correlated in a consistent way with stages of the estrons cycle of the rat. Associated with the rough endoplasmic reticulum short ribbons of moderately electron dense material appeared at metestrus and circular or elliptical bodies termed "ribbon-rolls" at diestrus and proestrus. Although present in proestrus, the ribbon-rolls were smaller at this stage. In a few neurons in diestrous females and in ovariectomized animals one to seven months before perfusion multiple large ribbon-rolls occupied much of the cytoplasm. Also, frequency of dense granules and lysosomes increased in diestrus. The significance of the ribbon-rolls and changes in other structures during the estrous cycle are discussed.
Key words: Arcuate nucleus - - Hypothalamus - - Sexual cycle - - Ribbon-rolls - - Electron microscopy.
Introduction
Mechanisms for both tonic and phasic release of gonadot rop ins from the p i t u i t a r y appea r to be dependen t upon the in tegr i ty of the basomedia l s t r ip of the h y p o t h a l a m u s (Hals et al., 1962). W h e n this area, which includes the a rcua te nucleus and med ian eminence, is i sola ted surgical ly f rom the r ema inde r of the hypo tha l amus , only tonic release of F S H and L H is re ta ined (Hals and Pupp , 1965). Neurons loca ted more ros t ra l ly appea r to de te rmine phasic secret ion of gonado t rop in by modi fy ing the ac t i v i t y in this basomedia l region (Hals and Gorski, 1967).
I n th is s t u d y we have endeavored to descr ibe u l t r a s t ruc tu r a l changes occurr ing in a rcua te neurons dur ing the estrous cycle. A t t en t i on was focused upon neurons in the a rcua te region because t hey appea r to opera te under gonada l s teroid influence (Lisk and Newlon, 1963 ; Yagi, 1973) in modi fy ing gonado t rop in release. Z a m b r a n o (1969) has shown t h a t the number of g ranu la t ed vesicles in a rcua te neurons peaks dur ing ear ly proes t rus and then subsides to much lower levels
Send o//print requests to: J. King, Dept. of Psychology, Tulane University, New Orleans' La. 70118 (USA). * Acknowledgments. This investigation was supported by: 1) predoctoral fellowship from the National Institute of Mental Health to J.E.K. 2) NIH-(7-ROI-NS-11650-02) to T.H.W. 3) HD 00867-I0 to A.A.G.

498 J .C . King et al.
dur ing estrus . I n e i t he r sex, t h e n u m b e r of g r a n u l a t e d vesicles in these neu rons
increases six m o n t h s a f t e r g o n a d e c t o m y ( Z a m b r a n o a n d de Rober t i s , 1968). I n
c a s t r a t e d m a l e ra ts , B r a w e r (1971) o b s e r v e d " w h o r l s " of e n d o p l a s m i c r e t i c u l u m
(ER) a r r a n g e d concen t r i c a l l y a n d p a r t i a l l y occup ied by e l ec t ron -dense ma te r i a l .
T h e n u m b e r of these g e o m e t r i c a l " c a s t r a t i o n " whor ls in g o n a d e c t o m i z e d ma les
d imin i sh fo l lowing in j ec t ions w i t h androgen . S imi l a r s t r uc tu r e s were e n c o u n t e r e d
v e r y r a re ly in i n t a c t ma le ra ts . Z a m b r a n o a n d de R o b e r t i s (1968) n o t e d t h a t
rough E R c i s te rnae were d i l a t ed one m o n t h a f te r c a s t r a t i o n as c o m p a r e d w i t h
i n t a c t an imals , b u t t h e genera l a r r a n g e m e n t of E R profi les was r e g a r d e d as
s imilar . I n t h e p r e sen t r e sea rch i n f o r m a t i o n was s o u g h t a b o u t changes in t h e f ine
s t r u c t u r e of a r c u a t e neu rons in f ema le ra t s in se lec ted s tages of t h e es t rous cycle, and in o v a r i e c t o m i z e d an imals .
Materials and Methods Subjects were 34 female Sprague-Dawley rats born in the Tulane Laboratory. Two to
three females were housed in cages in a room maintained on a reversed lighting schedule with 12 hrs light and 12 hrs dark. Group I consisted of 27 intact animals sacrificed on different days of the estrous cycle. Vaginal smears taken by daily lavage and stained with Wright's solution were analyzed from the onset of puberty. Proestrus was diagnosed by smears con- taining a predominance of nucleated epithelial cells; diestrus by those with a large percentage of leukocytes; metestrus with cornified cells and leukocytes; and estrus with 90 % or more of the cells in a cornified condition.
Eight animals were perfused on the day of estrus, 5 on the day of metestrus, 7 on diestrus and 7 proestrus. All animals were sacrificed between 100 and 190 days of age. Vaginal, ovarian and uterine tissues were stained with hematoxylin-eosin, and these tissues were evaluated by an observer unfamiliar with the experimental treatments to verify stages of the estrous cycle. The stage of folliculogenesis, recency of ovulation, proliferative stage of the uterus, and condition of the vaginal epithelium were used to assess endocrine condition. Group I I consisted of 7 oil-injected females ovariectomized at 60 days and sacrificed at 100 days of age.
Per/usion technique. Animals were perfused using three aldehyde solutions of increasing concentrations buffered to pH 7.2 with Sorensen's phosphate buffer. The solutions were: 1) a mixture of 1.5% glutaraldehyde and 1% paraformaldehyde; 2) 2.75% glutaraldehyde and 2.5% paraformaldehyde; and 3) 6% glutaraldehyde and 4% paraformaldehyde mixture. In seeking optimal results, variations were introduced in the concentrations of aldehydes and buffer employed. The brain perfusion technique described by Palay et al. (1962) to secure even osmic acid fixation was changed to take advantage of the properties of aldehyde fixatives. To prevent blood clotting, heparin was injected intraperitoneally 15 min prior to the nembutal anaesthetic which was administered by the same route. The lungs were ventilated by endo- tracheal tube up to the moment that perfusion was started. After opening the thorax, the aorta distal to the arch and the vena cava were clamped. Fixative was introduced into the left ventricle by a large short-beveled needle (to minimize back-pressure), or directly into the aorta through a glass cannula introduced through the cut left ventricle. A hydrostatic column of 5ft. was maintained. The right atrium was punctured as soon as it became distended with fluid. The first portion of the perfusate was heated by coiling the tubing in a 37 ~ water bath. The remainder was delivered cold. When a regular flow had been established, the caudal end of the animal was elevated about 4 in. and ice placed over the head and adjacent portion of tubing, and at that t ime the flow rate was also reduced. Animals were perfused for about 30 to 45 min before the head was removed and immersed in the strongest fixative overnight. The brain was removed on the following day.
Sample Preparation /or Electron Microscopy. With the help of a dissecting microscope coronal slices approximately 3/4 mm thick and including portions of the arcuate-median eminence region were removed from the brain with minimum trauma. Three slices corre-

Fine Structure of Arcuate Neurons 499
sponding to anterior, middle and posterior portions of the arcuate nucleus were obtained from each half brain. These blocks of tissue were washed for I/2 hour in a cold phosphate buffered 8% dextrose solution, postfixed in 1.5% osmium tetroxide solution buffered with 0.1 M Sorensen's buffer for 1 hour, washed in a 2.4% sodium chloride solution, dehydrated in graded ethanols, cleared in propylene oxide and embedded in an epon-araldite mixture.
R e s u l t s
Changes in the characteristics of subcellular structures--particularly the membranous ones--were found to correlate in a consistent way with successive stages in the estrous cycle. The pattern of alterations in the key organelles of a representative arcuate neuron at sequential stages of the estrous cycle are dia- grammatically illustrated in Fig. 1.
Estrus. Estrous animals include some sacrificed in early estrus soon after ovulation occurred, and a majority perfused in the afternoon. In the latter animals, the LH surge would have occurred approximately 24 hrs. before sacrifice. In all estrous animals, the rough ER of arcuate neurons is sparse and scattered around the cytoplasm without any orientation in particular. Fig. 2 is a represen- tative micrograph of a neuron at this stage in the cycle. Ribbons and ribbon-rolls are not components of the cytoplasm in early estrus.
In sections through a l l neurons, particles identified as lysosomes are seen. They were counted in electron micrographs that displayed complete sections through perikarya and nuclei, and the average number of lysosomes per neuron section increases from 4.2 per section in early to 5.1 per section in late estrus. The homogenity of the lysosome population in estrous animals is greater at this stage of the cycle than at others. Few of the lysosomes contain dark granules or lucent vacuoles. Smaller dense inclusions of assorted shapes and enclosed in distinct unit membranes could be seen in neuronal cytoplasm of estrous animals. Their mean number was 1.1 per neuron section. Many other small vesicles, both "alveolate" and smooth-walled, located in the vicinity of the Golgi apparatus in estrous animals seldom contained dense inclusions.
Metestrus. When the cytoplasm of neurons is examined in detail, it is found that on the day of metestrus the morphology of arcuate neurons changes in a striking fashion. The most conspicuous feature observed at this phase of the cycle is a new arrangement of the rough endoplasmic reticulum, which forms a widely dispersed network of cisternae as typified in Fig. 3. Associated with the rough ER in many arcuate neurons, are short ribbons of electron-dense material that maintain a width of about 500 /~. As shown in Fig. 4, the accumulated material in short ribbons appears first where the cisternae of two strands of rough ER come together. At this point, the contiguous membranes of the two cisternae lose their ribosomes and diverge to envelop the moderately electro-dense material. With few exceptions, the outer membranes of the rough endoplasmic reticulum (RER) retain their coatings of ribosomes. However, in some cases a second ribbon is formed parallel to the first, with the result that both boundary membranes of a cistern participate in ribbon formation and lose their ribosomes. Complex ribbon formations are rarely seen in metestrus females. One such stack of parallel ribbons is shown in Fig. 5. (All types of ribbons are never completely independent of the RER.)
33 Cell. Tiss. Res., 153

500 J.C. King et al.
~ ~ & ' ~ ~ ~ di~ ~,~:~,d':.:: ~.:~,~;~:~:~::;~: ~ ...... ~ ~ / �9 ..." . . ' . . . ~ ' ~ . : ; - : : . . o i . o ~ , . : ..:, ~ ' . . . . . . . . . . . . . ~ '%.~ t~=:( . : �9 . ~ . . : . ~ . . . . . - ~
...: ~.~ . .~ ~.~I~.?~ . " \:~.../:~-.~.o~ :-~..:i ~://.//.~ '" '~" ' ~ ' '"; " '/~ (:~.?.'2.. /
�9 .. ~ . ~.-...: :...:,..:~ .... ':~\://
:-:i ~ .~< M .+.. ~ . ~ . Proestrus
Fig. 1. Typical appearances of organelles within arcuate neurons of normal female rats in sequential stages of the estrous cycle. For details refer to text. dcv dense-cored vesicles; G Golgi complex; GFB granular filamentous body; L R long "ribbon"; L Y lysosome; M mitochondrion; N nucleolus; N U C nucleus; R E R rough endoplasmic reticulum; R R
"ribbon-roll"; S R short "ribbon"
Another distinctive feature of the cytoplasm in metestrus is a great ly increased population of dense granules associated with the Golgi apparatus, where they partially replace or form an addition to the clear vesicles--coated and un- c o a t e d - f o u n d in the domain of the Golgi appara tus (see Fig. 6).
Dies t rus . The arcuate neuron during diestrus provides an interesting com- parison to the preceding stages. I n particular, the rough E R profiles become most profuse and complicated in form and are often densely packed into coherent Nissl bodies whose parallel arrays of cisternae tend to repeat the contour of the cell membrane (Fig. 7). Here, as in Nissl bodies of m a n y other neurons, the E R membranes were s tudded with ribosomes and many ribosomes occupied the interstices between the cisternal profiles. I n a large sample of arcuate neurons from diestrous animals approximately half surveyed showed this extensive R E R

Fine Structure of Arcuate Neurons 501
Fig. 2. Features of typical arcuate neurons of estrous females. Note stacks of Golgi profiles (G), while the rough EI~ is sparse. There are few granulated vesicles in the vicinity of the Golgi apparatus, bu t other organelles including mitochondria and lysosomes are fairly abundant .
Nuclear invaginations are present in a large percentage of arcuate neurons, x 12800
Fig. 3. Arcuate neurons from a metestrous female. The rough E R has the form of a loose network made up of interconnecting cisternae, x 13600
33*

502 J .C . King et al.
Fig. 4. Arcuate neuron from a metestrous female. Simple ribbon-like structure, composed of moderately electron-dense material located in 400 A gap between the two rough ER profiles. Ribosomes adhere to the outer membranes of the rough ER cisternae but are absent from the segments of membranes (arrows) that are in contact with the ribbon of electron-dense material.
• 44400
Fig. 5. Metestrus. ER profiles are stacked in parallel, and varying amounts of moderately electron-dense material are accumulated between the stacks. Several ribbons in parallel may
be distinguished. • 20000

Fine Structure of Arcuate Neurons 503
Fig. 6. In arcuate neurons of metestrous females there is an increase in the number of dense granules which are found most commonly in close relationship to elements of the Golgi
apparatus (G). x 15200
Fig. 7. Build up of rough ER, particularly near the periphery of the neuron, in diestrous females. X 14100

504 J.C. King et al.
development. The remainder of the perikarya showed no appreciable increase in RER, dense cored vesicles or lysosomes. Since the rough ER pervades most of the peripheral cytoplasm in many neurons, other cytoplasmic organelles including mitochondria, lysosomes and Golgi elements become sequestered close to the nucleus.
Many ribbons composed of moderately electron-dense material are found in the diestrous female. There is no recognizable increase or decrease in density of the membrane bound material. Some of these ribbons, as in metestrus, are more or less straight while others curve to form circular structures two of which are shown in Fig. 8.
In some arcuate neurons ribbons of similar material appeared to aggregate into distinctive circular or elliptical bodies ranging in diameter from 2 to 6 microns. We have coined the term "ribbon-roll" (RR) to describe their geo- metrical forms, examples of which can be seen in Figs. 1 and 9. Many of these ribbon-rolls exhibit a central core of cytoplasm in which few if any inclusions could be identified. The ER membrane adjacent to the central core of cytoplasm is studded with ribosomes as is the outermost membrane of the ribbon-roll. The number of concentric ribbons which makes up the ribbon-rolls varies, but the average is 8 lamellae per roll. I t was the exception rather than the rule to find a single ribbon-roll in the cell. More often they occur in pairs or in groups up to 6 (Fig. 9). Since some ribbon-rolls in a given cell may not have been visualized because they were outside the plane of section, the number observed probably underestimates their actual frequency.
I t should be noted that ribbon-rolls are not distributed randomly among cells of the arcuate nucleus. Cells with this cytoplasmic component are in most cases located close to the median eminence and the infundibular recess. Results of surveys using coronal sections, which are depicted in Fig. 10, show that the cells under discussion are more prevalent where the arcuate nucleus reaches the precinct of the ependyma of the third ventricle and the median eminence.
In diestrous animals, many arcuate neurons exhibit an abundance of lyso- somes, dense cored vesicles and mitochondria. In 17 neurons, selected unsys- tematically without regard to their content of cytoplasmic organelles, the average number of lysosomes in the plane of section was 9.8. This percentage is approxi- mately double the number seen in estrous and metcstrous animals. At this stage of the cycle many of the lysosomes incorporate clear vacuoles and dark spots. The shapes of lysosomes also became more elaborate by the time diestrus arrived so that not only the frequency but also the complexity of lysosomal form is enhanced during diestrus.
The smaller membrane-bound dense inclusions also have a greater frequency in diestrous than in metestrous or estrous animals. These dense inclusions are generally associated with the Golgi region, but some of them arc dispersed among the rough ER cisternac. Examples of these vesicles are shown in Fig. 9.
P r o e s t r u s . The proestrous phase prior to ovulation also had some distinctive features, particularly with respect to appearances of the ribbon-rolls and the images of I~ER cisternae. The profile of the ribbon-roll in proestrus is of reduced complexity as compared with the previous day. Lamellae on average decrease
Fine Structure of Arcuate Neurons 505
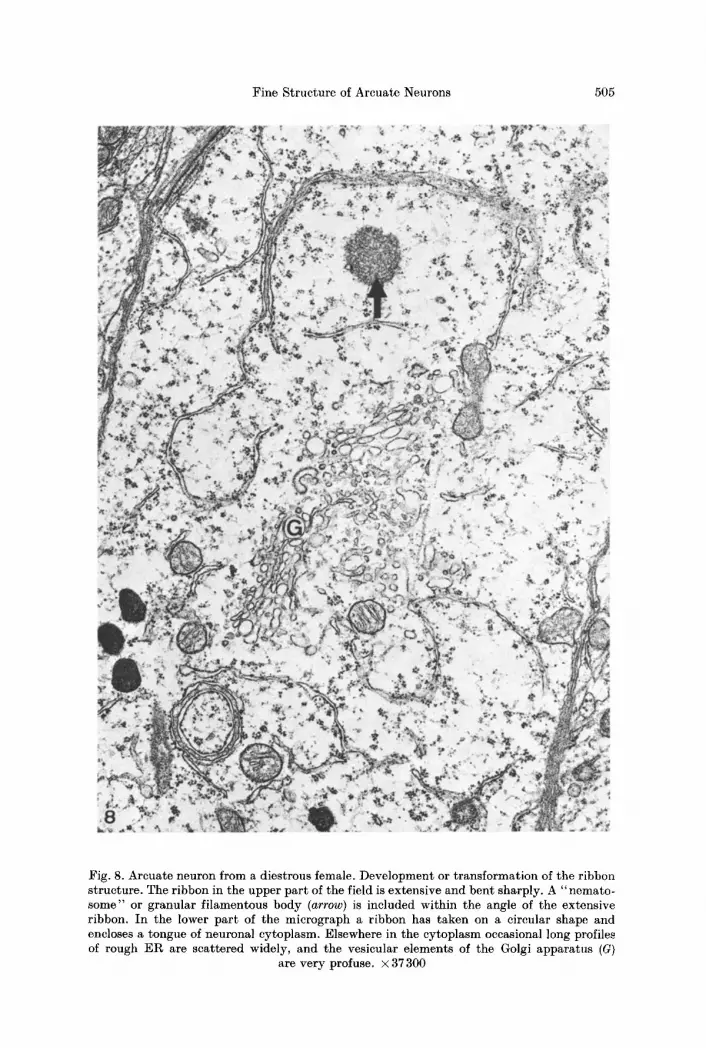
Fig. 8. Arcuate neuron from a diestrous female. Development or transformation of the ribbon structure. The ribbon in the upper part of the field is extensive and bent sharply. A "nemato- some" or granular filamentous body (arrow) is included within the angle of the extensive ribbon. In the lower part of the micrograph a ribbon has taken on a circular shape and encloses a tongue of neuronal cytoplasm. Elsewhere in the cytoplasm occasional long profiles of rough ER are scattered widely, and the vesicular elements of the Golgi apparatus (G)
are very profuse. • 37 300

506 J .C . King et al.
Fig. 9. Portions of three ribbon-rolls (arrows) in an arcuate neuron from a diestrous female. Six rolls were observed in this cell. Some of the membranes are blurred because of the oblique angle of cutting. The largest ribbon-roll is connected to the rough E R complex by several
channels of anastomosing E R cisternae. • 14600
/ ~ r - - ~ . . \ ~ '~ l / / ~ " " \ \
* / I I I J ~ . . . . / C - ~ ~ / I I I I \ ~ . _ . I I \ , / ~ 1 1 / ~ , ' f N
, , I , , i l l . / I / '~ , , - , , / / V I \ \ , . \ I l u I i l~r I \ .,,. \ V I I I I
I / / .:./ \- \ x \ / i / . . ' : ' I ~ : \ - - - - / \ , . - / . . : : : . / \:..... \ \ \ ,
~ -'- ~ . . , r : . : . : ~ : : : . : ! : . , . . . ~ - - - / M E ......
10 Fig. 10. A coronal section through the arcuate-median eminence region shows the distr ibut ion of ribbon-rolls in the arcuate nucleus (arrow). These structures appear only in diestrus and proestrus and are found most frequently close to the median eminence and underlying the ependymal elements of the th i rd ventricle (stippled area). DM dorsomedial nucleus; VM
ventromedial nucleus; ME median eminence

Fine Structure of Arcuate Neurons 507
from about 8 in diestrus to 2 to 4 concentric ribbons in proestrus (Fig. 11). The appearances are compatible with a process of dissassembly of the ribbon-rolls, with membranes less distinct and less orderly.
As in diestrus, there are still large amounts of rough ER in the neuronal cyto- plasm. However, the cisternae tend to be shorter. A prominent feature of the day of proestrus is the unusual number of Golgi profiles set out in curved arrays all around the nucleus and often pervading most of the cytoplasm. Swarms of clear vesicles and many multivesicular bodies are found in close relationship to the Golgi apparatus. However, only a few dense-cored vesicles are present. As in diestrus, lysosomes are abundant during proestrus and some of them contain small lucent vacuoles.
Ovariectomized animals. In adult female rats ovariectomized from 3 weeks to 7 months prior to sacrifice (Figs. 12 and 13), approximately 50% of arcuate neurons contain large amounts of RER, often densely packed into coherent Nissl bodies similar to the kind seen in diestrous females (Fig. 13). Ribbon-rolls and long ribbons are more numerous in the arcuate neurons of ovariectomized than in diestrous females (Fig. 12). The ribbons are either in single or in parallel array and most often are related to ribbon-rolls (Fig. 13). Most ribbon-rolls seen in ovariectomized females are made up of many lamellae packed closely together. However, in some of these structures, the lamellae are interrupted and partially replaced by irregular vacuoles or by rows of vesicles. In addition, some of the membranes bounding the concentric vesicles are sometimes disrupted. One regularly organized ribbon-roll is sometimes found juxtaposed to another that displays evidence of disruption. Although signs suggesting disorganization have been observed in ribbon-rolls from diestrous animals, vacuoles are relatively infrequent and the concentric lamellae are less numerous than in ovariectomized females.
Small dense inclusions associated with the Golgi apparatus are not abundant in arcuate neurons of ovariectomized females. The number per cell approximated tha t seen in estrus. Lysosomes are quite numerous, but contain fewer lucent vacuoles and dark spots than in normal diestrous or proestrous animals.
In intact females throughout the estrous cycle and in ovariectomized females a circular body 0.8 to 2.8 microns in diameter was sometimes observed embedded in the perikaryal cytoplasm. Composed of t iny filaments and granules, the structure lacks a limiting membrane and resembles nematosomes or nucleolus- like bodies (Le Beux, 1971). Small zones of translucency (vacuoles) are present frequently within this structure. Dense bodies adhere to its surface and micro- filaments radiate from its periphery into the "ha lo" or clear area of surrounding cytoplasm. Commonly related organelles include free polysomes and rough ER. Sometimes the granular filamentous bodies are located in neurons containing ribbon-rolls.
From time to t ime a flattening on one side of the granular filamentous body was viewed. A narrow band has sometimes been seen between the flattened edge of a filamentous body and an axosomatic synapse. A similar band parallel to the flattened side of a filamentous body, has been observed in close proximity to a cistern of rough ER. Rarely the circular structure seemed to be wrapped around

508 J .C . King et al.
Fig. 11. Ribbon-roll (RR) from an arcuate neuron of a proestrous female. This ribbon-roll has fewer lamellae than those seen in diestrus. • 25900
Fig. 12. Stacking of simple ribbons in an arcuate neuron from an ovariectomized female. The basic structure of the simple ribbon is repeated twice (arrows). At the end of the assembly,
two cisternae of rough El% (e2 and e3) are joined by a third cistern (el). • 32400

Fine Structure of Arcuate Neurons 509
Fig. 13. Three ribbon-rolls in an arcuate neuron from an ovariectomized female. Many lamellae are arranged concentrically around a central cytoplasmic core. All three roils are interconnected via the cisternae of rough ER (arrows). As compared with arcuate neurons from diestrous females, these ribbon-rolls have more inclusions in the cytoplasmic cores
(arrowheads) and vesiculation is present in the concentric lamellae. • 17 300

510 J.C. King et al.
by an irregular membrane. The morphology of nucleolus-like body does not vary cyclically.
Discussion
Many neuronal organelles varied with the cyclic endocrine condition of the female rat. The cell content of rough endoplasmic reticulum (ER) cisternae progresses from fragmentary to organized assemblies of profiles from estrus to diestrus. Accompanying the increase in the amount of rough ER is a change in its pat tern of distribution in the cytoplasm. The highest degree of organization occurs in diestrus ; on this day the cisternae are oriented in stacks aligned with the contours of the plasma membrane.
At the stage that rough ER had proliferated markedly, some cisternae are seen close together and in parallel, and with ribbons of moderately electron-dense material occupying the ribosome free area between them. Ribosomal coating of the membranes resume at either end of each ribbon. A comparable modification of rough ER has been described as intracytoplasmic confronting or paired cisternae by Kumegawa et al. (1968). We have called them ribbons to focus attention upon the striking accumulation of material between two cisternae. In metestrus, ribbons are rather short and usually composed of two cisternae, whereas in diestrus development of the ribbon-like inclusions had continued so that they become longer and frequently arranged in stacks of 3 to 7 cisternae. The appearance of complex ribbon-rolls during diestrus and smaller ribbon-rolls during proestrus may represent the culmination in the production of ribbon material.
Although the function of the ribbon-roll is enigmatic, it might be useful to tabulate examples of these inclusions drawn from the literature in order to focus attention on the frequency of the phenomenon and its presumed significance. A partial listing of information about circular membranous structures categorized by the presence or absence of ribosomes is shown in Table 1.
Type I membranes have been observed in cells from ovary, testis, arcuate nucleus, ependyma, and pineal gland. This type of configuration contains mem- branes without ribosomes and it appears reasonable to infer tha t development of these structures is correlated with secretory activity of various kinds. For example, they have been reported by Bjersing (1967) in the corpus luteum during times of high progesterone secretion.
Type I I structures have been demonstrated in a variety of organs, including pancreas, adrenals, melanoma, neurons, spermatids, and Rathke 's pouch. Con- centrically arranged membranes contain ribosomes along their entire course, and such elements have sometimes been referred to as " N e b e n k e r n " . In some circumstances, as when adrenocortical cells have been stimulated by ACTH, the rough ER whorls disappear and additional smooth ER appears to be created (Nickerson and Curtis, 1969; Nickerson, 1970), suggesting a possible role for rough ER in the synthesis of smooth ER. There is a deficiency of information about the regulation of these changes within the cell.
Ribbon-rolls have been reported less frequently than the other concentric membranous structures. Brawer (1971) found them in arcuate neurons in castrated but not in intact male rats. He was able to reduce the number of bodies in arcuate neurons in males by the administration of testosterone, thereby providing evidence

Fine Structure of Arcuate Neurons
Table 1
511
Tissue Condition
Type I: Membranous whorls without ribosomes: Parasomes
Carr and Carr (1962)
Christensen (1965)
Blanchette (1966)
Christensen and Fawcett (1966)
Bjersing (1967)
Brawer (1972)
Bucana and Nadakavukaren (1973)
Pevet and Saboureau (1973)
testicular interstitial cells normal in mouse
interstitial cells of guinea pig normal
rabbit lutein cells first stage of pregnancy
mouse testicular interstitial cells normal
corpus luteum of pig
processes of tanycytes
Harderian gland of female hamsters
pineal organs of male hedgehog
early diestrus; time of high progesterone secretion
normal male rat
12 days of life to adulthood
period of quiescence
Type I I : Membranous whorls with ribosomes: "Nebenkern"
Herman and Fitzgerald (1962) regenerating acinar cells of rat pancreas
Nickerson and Curtis (1969) adrenocortical cells in region of fasciculata-reticularis boundary
Nickerson (1970) same as above
Andersen et al. (1971)
Hu (1971)
human foetus cells lining Rathke's pouch
melanoma cells
Castel (1972) tadpole melanotrophs
Cnrgy and Anderson (1972)
Kiski (1972)
Jong-Brink (1973)
Rasmussen (1973)
Drosophila spermatocytes
medulla oblongata of rat, nucleus dorsalis vagi lateralis
albumen gland of Biomphalaria glabrata
spermatid of Drosophila
12 to 15 days after ethionine destruction
normal young male gerbils
normal and after cessa- tion of ACTH treatment
normal
dividing in late phases of mitosis
normal dark adapted and 24 hrs following de- nervation disinhibition
1st stages of spermiogenesis
occasional profiles in normal animal
10 to 36 days following starvation and desiccation
Spermatid at time of elongation
Type I I I : Membranous whorls with ribosomes on innermost and outermost sur/aces with electron dense material between cisternae: Ribbon-Rolls
Brawer (1971) arcuate nucleus, male rats following castration
Dubois and Girod (1971) anterior pituitary, golden during winter hamsters
Ford et al. (1973) arcuate nucleus, adult male rats following chronic morphine treatment

512 J.C. King et al.
for the view of hormonal regulation of " e m p t y i n g " or "fil l ing" of these inclusion bodies. Dubois and Girod (1971) report similar formations in anterior pituitary cells of sexually quiescent hamsters and apparently identical bodies have been located in the arcuate nuclei of male rats treated for several days with increasingly higher doses of morphine (Ford et al., 1973). In the present study, these structures were prevalent during diestrus, following ovariectomy and in much smaller sizes, on the morning of proestrus. In all of these conditions, levels of gonadal hormones are low, suggesting tha t the disappearance of the ribbon-rolls may be correlated with changes in feedback processes. In the case of females, the absence of ribbon- rolls might signal conditions prior to the LH surge. Lampert i and Niewenhuis (personal communication) have also observed these structures in arcuate neurons of female hamsters on Day 3 and 4 of the cycle. Also, formation of ribbon-rolls may be influenced by neural stimulation from rostral areas such as the medial preoptic area as well as hormones, such as LH or other substances.
No knowledge about the content or function of the ribbons and ribbon-rolls in arcuate neurons is available. I t might be hypothesized that the ribbon-rolls are involved in the synthesis of LH-RH. Development of the ribbon-rolls parallels in time the peaks of L H - R H in the median eminence (Chowers and McCann, 1965; Ramirez and Sawyer, 1965). I f not directly involved in the synthesis of L H - R H itself, these structures may represent carrier protein or enzymes associated with L H - R H manufacture or release. Litteria (1973) has recently found tha t H a lysine is incorporated into proteins of medial preoptic, arcuate, periventricular, para- ventricular, and supraoptic nuclei maximally during proestrus and minimally during diestrus.
Another possible function of ribbon-rolls could be synthesis and packing of neurosecretory material in conjunction with the Golgi apparatus. Events related to alterations in rough ER are followed by changes in the Golgi apparatus. In some neurons on the day of diestrus the Golgi apparatus is associated with more vesicles including some very small ones, and contained tubular and cisternal profiles. This proliferation of elements became exceptionally high in proestrus. Occasionally, the Golgi complex is the most prominent and extensive component of the cytoplasm. Multivesicular bodies are often present in their vicinity. Golgi complex and ribbon-like inclusions go through various stages together, suggesting the possibility of a close functional link between them.
A third function of structures could involve detoxification of surplus hormones or their metabolites. Jones and Fawcett (1965) have described hypertrophy of smooth ER in liver cells induced by phenobarbitol. In arcuate neurons, the process of ribbon formation makes its first appearance during metestrus, as indicated by the presence of short ribbons. The transformation of profiles of R E R from short ribbons to longer stack ribbons and finally to ribbon-rolls could be for the purpose of trapping excess substances. However, this hypothesis suffers from the disadvantage tha t the ribbon-rolls are dispersed during proestrus and cleared from the scene by estrus.
Investigators are trying to correlate the sizes of granular vesicles with their presumed contents, and special efforts have been made to categorize nerve profiles in the median eminence, on the basis of mean sizes of granular vesicles (Stoechart et al., 1973). As of now, no reliable ultrastructural criterion for distinguishing

Fine Structure of Arcuate Neurons 513
between vesicles containing neurohormone or neurotransmitter has been identified. The fluctuation in the dense-cored vesicle population observed in the rat arcuate nucleus between proestrus and estrus is of considerable interest. There is overlap between the part of the arcuate nucleus that contains ribbon-rolls, and the region showing dopamine fluorescence (Fuxe and Hokfelt, 1969). Therefore, the fluctua- tion may be related to the storage and release of LH-RH, and/or catecholamine, specifically dopamine.
Oksche et al. (1972) have suggested that there is clustering of aminergic neurons and neurons containing releasing hormones. The fluorescence intensity of dopamine cell bodies in the arcuate nucleus (Fuxe et al., 1967 ; Lichtensteiger, 1969) is correlated with stages of the cycle in the intact female. Cyclic variations are no longer present in the ovariectomized female (Fuxe et al., 1970), nor in constant estrous females (Fuxeeta l . , 1972). However, level of fluorescence intensity can be altered by unilateral stimulation of areuate neurons (Keller and Liehtensteiger, 1971).
The increased numbers of dense-cored vesicles observed in diestrous females may reflect the greater catecholamine or releasing hormone content of areuate neurons at this stage of the cycle. Nonetheless the role of dopamine in the regula- tion of LH-RH is still to be clarified. Increased numbers of dense-cored vesicles were not observed in females ovariectomized one month previously. Zambrano and de Robertis (1968) reported a greater number of granulated vesicles 6 but not one month after ovariectomy.
The increase in the number of variegated inclusions in dense bodies, probably lysosomes, in diestrus may be related to the consumption of over-produced dense- cored vesicles as has been suggested by Zambrano (1969). In proestrus, the population decreased and the inclusions are smaller, clearer, and more uniform.
"Granular filamentous bodies" or "nematosomes" have been described by Le Beux (1971) and reported to be present in areuate neurons in male (Brawer, 1971) and female rats (Adamo, 1972). In the data reported here, granular fila- mentous bodies were present in intact females at all stages of the cycle and in ovariectomized females. Kishi (1972), utilizing enzyme digestion tests, has demonstrated the proteinous character of the nucleolus-like body and suggested that ncmatosomes may be accumulations of protein synthesized by ribosomes.
References
Adamo, N. J. : Ultrastruetural features of the lateral preoptie area, median eminence and areuate nucleus of the rat. Z. Zeilforsch. 127, 483-491 (1972)
Andersen, H., van Bullow, F. A., Mollgard, K.: The early development of the pars distalis of human foetal pituitary gland. Z. Anat. Entwickl.-Gesch. 135, 117-138 (1971)
Bjersing, L.: On the ultrastrueture of granulosa lutein cells in porcine corpus luteum. Z. Zellforsch. 82, 187-211 (1967)
Blanchette, E. J.: Ovarian steroid cells, II. The lutein cell. J. Cell Biol. 81, 517-542 (1966) Brawer, J. R. : The role of the arcuate nucleus in the brain-pituitary-gonad axis. J. comp.
Neurol. 143, 411446 (1971) Brawer, J. R. : The fine structure of the ependymal tanycytes at the level of the arcuate
nucleus. J. comp. Neurol. 145, 2542 (1972) Bucana, C. D., Nadakavukaren, M. J.: Ultrastructural investigation of the postnatal devel-
opment of the hamster Harderian gland. Z. Ze]lforsch. 142, 1-12 (1973)

514 J .C . King et al.
Carr, I., Carr, J . : Membranous whorls in the testicular interstitial cell. Anat. Rec. 144, 143-147 (1962)
Castel, M. : Ultrastructure of the anuran pars intermedia following severance of hypothalamic connection. Z. Zellforsch. 131,545-557 (1972)
Chowers, I., McCann, S. M. : Content of luteinizing hormone-releasing factor and luteinizing hormone during the estrous cycle and after changes in gonadal steroid titers. Endocrino- logy 76, 700-708 (1965)
Christensen, A. K. : The fine structure of testicular interstitial cells in guinea pigs. J. Cell Biol. 26, 911-935 (1965)
Christensen, A. K., Fawcett, D. W. : The fine structure of testicular interstitial cells in mice. Amer. J. Anat. 118, 551-572 (1966)
Curgy, J. J . , Anderson, W. A. : Synth~se d 'ARN dans le chondriome au cours de la spermio- gen~se chez la Drosophile. Z. Zellforsch. 125, 31-44 (1972)
Dubois, P., Girod, C.: Formations lamellaires concentriques dans des cellules ant6hypo- physaires chez le hamster dot6. Z. Zellforsch. 115, 196-211 (1971)
Ford, D. H., Voeller, K., Callegari, B.: Changes in neurons of the median eminence-arcuate region of rats induced by chronic morphine treatment: an electron microscopic study. Anat. Rec. 175,321 (1973)
Fuxe, K., HSkfelt, T.: Catecholamines in the hypothalamus and the pituitary gland. In: Frontiers in neuroendocrinology (Ganong, W. F. and Martini, L., eds.), p. 47-96. New York: Oxford Univ. Press 1969
Fuxe, K., HSkfelt, T., Jonsson, G.: The effect of gonadal steriods on the tubero-infundibular dopamine neurons. Hormonal steroids (Proceedings of the Third International Congress), p. 806-813 (1970)
Fuxe, K., HSkfelt, T., Nilsson, O.: Activity changes in the tuberoinfundibular dopamine neurons of the rat during various stages of the reproductive cycle. Life Sci. 6, 2057-2061 (1967)
Fuxe, K., HSkfelt, T., Nilsson, O. : Effect of constant light and androgen-sterilization on the amine turnover of the tuberoinfundibular dopamine neurons: blockade of cyclic activity and induction of a persistent high dopamine turnover in the median eminence. Acta endocr. (Kbh.) 69, 625-639 (1972)
Halhsz, B., Gorski, R. A.: Gonadotrophic hormone secretion in female rats after partial or total interruption of neural afferents to the media basal hypothalamus. Endocrinology 89, 608-622 (1967)
Halhsz, B., Pupp, L. : Hormone secretion of the anterior pituitary gland after physical inter- ruption of all nervous pathways to the hypophysiotropic area. Endocrinology 77, 553-562 (1965)
Hal~sz, B., Pupp, L., Uhlarik, S. : Hypophysiotrophic area in the hypothalamus. J. Endocr. 25, 147-154 (1962)
Herman, L., Fitzgerald, P. J. : Restitution of pancreatic acinar cells following ethionine. J. Cell Biol. 12, 297-312 (1962)
Hu, F. : Ultrastructnral changes in the cell cycle of cultured melanoma cells. Anat. Rec. 170, 41-55 (1971)
Jones, A. L., Fawcett, D .W. : Hypertrophy of the agranular endoplasmic reticulum in hamster liver induced by phenobarbital (with a review on the functions of this organelle in the liver). J. Histochem. Cytochem. 14, 215-232 (1966)
Jong-Brink, M. : The effects of desiccation and starvation upon the weight, histology, and ultrastructure of the reproductive tract of Biomphalaria glabrata, intermediate host of Schistosoma mansoni. Z. Zellforsch. 186, 229-262 (1973)
Keller, P. J . , Lichtcnsteiger, W.: Stimulation of tubero-infundibular dopamine neurons and gonadotrophin secretion. J. Physiol. (Lond.) 219, 385-401 (1971)
Kishi, K. : Fine structural and cytochemical observations on cytoplasmic neucleolus-like bodies in nerve cells of rat medulla oblongata. Z. Zellforsch. 182, 523-532 (1972)
Kumegawa, M., Cattoni, M., Rose, G. G. : Electron microscopy of oral ceils in vitro II . Sub- surface and intracytoplasmic confronting cisternae in strain KB cells. J. Cell Biol. 86, 443-452 (1968)

Fine Structure of Arcuate Neurons 515
Le Beux, u J. : An ultrastructural study of the neurosecretory cells of the medial vascular prechiasmatic gland, the preoptic recess and the anterior part of the suprachiasmatic area. I. Cytoplasmic inclusions resembling nucleoli. Z. Zellforsch. 114, 404440 (1971)
Lichtensteiger, W. : Cyclic variations of catecholamine content in hypothalamic nerve cells during the estrous cycle of the rat with a concomitant study of the substantia nigra. J. Pharmacol. exp. Ther. 165,204-215 (1969)
Lisk, R. D., Newlon, M. : Estradioh Evidence for its direct effect on hypothalamic neurons. Science 189, 223-224 (1963)
Litteria, M. : In vivo alterations in the incorporation of aH lysine into the medial preoptic nucleus and specific hypothalamic nuclei during the estrous cycle of the rat. Brain Res. 55, 234-237 (1973)
Nickerson, P.A.: Effects of ACTH on membranous whorls in the adrenal gland of the mongolian gerbil. Anat. Rec. I66, 479-490 (1970)
Nickerson, P. A., Curtis, J. C. : Concentric whorls of rough endoplasmic reticulum in adreno- cortical cells of the mongolian gerbil. J. Cell Biol. 40, 859-862 (1969)
Oksche, A., Zimmermann, P., Oehmke, H.-J.: Morphometric studies of tubero-eminential systems controlling reproductive functions. In: Brain-endocrine interaction. Median eminence: Structure and function. Int. Symp. Munich 1971 (K. M. Knigge, D. E. Scott, A. Weindl, eds.), p. 142-153. Basel: Karger 1972
Palay, S. L., McGee-Russell, S. M., Gordon, S., Grillo, M. A.: Fixation of neural tissues for electron microscopy by perfusion with solutions of osmium tetroxide. J. Cell Biol. 12, 385-410 (1962)
Pevet, P., Saboureau, M.: L'@piphyse du Herisson (Erinaceus europaeus L.) male. Z. Zell. forsch. 143, 369-385 (1973)
Ramirez, V.D., Sawyer, C. H.: Fluctuations in LH-RF (Luteinizing Hormone Releasing Factor) during the rat estrous cycle. Endocrinology 76, 282-289 (1965)
Rasmussen, S.W.: Ultrastructural studies of spermatogenesis in Drosophila melanogaster Meigen. Z. Zellforsch. 140, 125-144 (1973)
Stoechart, R., Kreike, A. J., Jansen, H. G.: Sizes of granular vesicles in the rat median eminence, with special reference to the zona granulosa. Z. Zellforsch. 146, 501-515 (1973)
u K. : Changes in firing rates of single preoptic and hypothalamic units following an intravenous administration of estrogen in the castrated female rat. Brain Res. 58, 343-352 (1973)
Zambrano, D.: The arcuate complex of the female rat during the sexual cycle. An electron microscopic study. Z. Zellforsch. 93, 560-570 (1969)
Zambrano, D., de Robertis, D.: The effect of castration upon the ultrastructure of the rat hypothalamus. I1 Areuate nucleus and outer zone of the median eminence. Z. Zellforsch. 87, 409-421 (1968)
34 Cell. Tiss. Res. 153





























