Embed Size (px)
Citation preview

Trophic interactions between introduced laketrout (Salvelinus namaycush) and native Arcticcharr (S. alpinus) in a large Fennoscandiansubarctic lakeAntti P. Eloranta1, Petri Nieminen1, Kimmo K. Kahilainen2,31Department of Biological and Environmental Sciences, University of Jyv€askyl€a, P.O. Box 35, FIN-40014, Jyv€askyl€a, Finland2Kilpisj€arvi Biological Station, University of Helsinki, K€asivarrentie 14622, FIN-99490, Kilpisj€arvi, Finland3Department of Environmental Sciences, University of Helsinki, P.O. Box 65, FIN-00014, Helsinki, Finland
Accepted for publication January 31, 2014
Abstract – Introduced fishes may have major impacts on community structure and ecosystem function due tocompetitive and predatory interactions with native species. For example, introduced lake trout (Salvelinusnamaycush) has been shown to replace native salmonids and induce major trophic cascades in some NorthAmerican lakes, but few studies have investigated trophic interactions between lake trout and closely related nativeArctic charr (S. alpinus) outside the natural distribution of the former species. We used stomach content and stableisotope analyses to investigate trophic interactions between introduced lake trout and native Arctic charr in largesubarctic Lake Inarij€arvi in northern Finland. Both salmonids had predominantly piscivorous diets at >280 mm totallength and were mainly caught from the deep profundal zone. However, lake trout had a more generalist diet andshowed higher reliance on littoral prey fish than Arctic charr, whose diet consisted mainly of pelagic planktivorouscoregonids. According to length at age and condition data, lake trout showed slightly faster growth but lowercondition than Arctic charr. The results indicate that introduced lake trout may to some extent compete with andprey upon native Arctic charr, but currently have only a minor if any impact on native fishes and food webstructure in Inarij€arvi. Future monitoring is essential to observe potential changes in trophic interactions betweenlake trout and Arctic charr in Inarij€arvi, as well as in other European lakes where the two salmonids currentlycoexist.
Key words: Invasive species; niche segregation; piscivory; predation; resource competition; stable isotope analysis
Introduction
Invasive species have become a major threat to manynative species and even whole ecosystems worldwide(Mack et al. 2000; Sakai et al. 2001). In fresh waters,invasive fishes can have deleterious effects on nativefish fauna through resource competition, predation,parasitism and hybridisation (Gozlan et al. 2010;Cucherousset & Olden 2011). Stocking of non-nativepredatory fishes can have multidimensional impactson lake ecosystem function such as increasing top-down effects, altering nutrient cycling and decreasingor increasing links between aquatic and terrestrial
ecosystems (Eby et al. 2006). Thus, evaluating thepotential impacts of introduced fishes on native faunais an important aspect of conservation and manage-ment of lakes and their fish stocks, particularly inoligotrophic high-latitude lakes, which are consideredespecially sensitive to environmental changes (Shin-dler & Smol 2006; Jeppesen et al. 2010).Salmonids have been widely introduced to many
lakes and rivers due to their high recreational andcommercial value (Rahel 2002; Crawford & Muir2008). However, introduced salmonids may pose amajor threat to native fishes and cause trophic cas-cades with potential impacts at the whole community
Correspondence: Antti P. Eloranta; Norwegian Institute for Nature Research (NINA), P.O. Box 5685 Sluppen, NO-7485 Trondheim, Norway. E-mail: [email protected]
doi: 10.1111/eff.12132 181
Ecology of Freshwater Fish 2015: 24: 181–192 � 2014 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd
ECOLOGY OFFRESHWATER FISH

and ecosystem levels (Simon & Townsend 2003; Ebyet al. 2006). For example, many native brown trout(Salmo trutta) populations in northern Fennoscandianstreams are currently threatened due to resource com-petition with introduced brook trout (Salvelinus fonti-nalis) (Korsu et al. 2010), whereas in western NorthAmerican lakes, predation by introduced lake trout(Salvelinus namaycush) has been shown to limit thedistribution of bull trout (Salvelinus confluentus)(Donald & Alger 1993) and cause significant popula-tion declines of cutthroat trout (Onchorhynchusclarki) (Ruzycki et al. 2003). In the latter case, preda-tion by lake trout on planktivorous cutthroat trout hasinduced a four-level trophic cascade in Yellowstonelake (Tronstad et al. 2010) and also influenced preda-tor–prey interactions in surrounding terrestrial eco-systems by increasing predation by grizzly bears(Ursus arctos) on elk (Cervus elaphus) calves due tothe loss of native cutthroat trout in small tributaries(Middleton et al. 2013). In addition to ecologicalimpacts, introduced salmonids can also have evolu-tionary impacts as several species can hybridise andintrogress with native species, as has been observedbetween European whitefish (Coregonus lavaretus)and vendace (Coregonus albula) (Kahilainen et al.2011a), between Atlantic salmon (Salmo salar) andbrown trout (Salmo trutta) (H�orreo et al. 2011), andbetween Arctic charr (Salvelinus alpinus) and laketrout (Wilson & Hebert 1993; Wilson & Bernatchez1998).Lake trout and Arctic charr are generalist salmo-
nids that have partly overlapping natural distributionin northernmost North America (Johnson 1980;Martin & Olver 1980). There, lake trout seems tohave displaced Arctic charr from several lakes, exceptfrom the coldest and most oligotrophic lakes in theHigh Arctic (Hershey et al. 1999, 2006). The two sal-monids may coexist in large, deep lakes, but competefor food and habitat resources due to their rather sim-ilar (mainly piscivorous) feeding behaviour (Johnson1980; Martin & Olver 1980). Lake trout is an effi-cient piscivore because it can attain a large body size(>1 m total length) and may therefore prey heavilyupon sympatric salmonids (Ruzycki et al. 2003),including Arctic charr (Swanson et al. 2010). Hence,introduced lake trout are predicted to have competi-tive and predatory impacts on native Arctic charr inlakes where the species do not naturally coexist. Suchnegative impacts on northern Arctic charr populationsmay become more common, particularly if climatechange expands the spatial extent and duration ofpreferred thermal habitat for lake trout (cf. Clineet al. 2013).Lake trout has been introduced to several lakes in
Europe (Martin & Olver 1980; Crossman 1995),including large subarctic Lake Inarij€arvi (hereafter
Inarij€arvi) in northern Finland, where stocking hasbeen conducted since 1972 (Salonen & Mutenia2007). Introduced lake trout have been reported toreproduce in some alpine and Fennoscandian lakes(Langeland 1994; Crossman 1995; Delacoste et al.1997), with potential negative effects on native sal-monids (cf. Eby et al. 2006). Although some studies(e.g. Fraser & Power 1989; Tronstad et al. 2010)have been performed in natural distribution areas,there has been a lack of detailed studies on the ecol-ogy and impacts of introduced lake trout in Euro-pean lakes. Potential influences of lake troutintroductions on native fish fauna and whole lakeecosystem function are difficult to assess and predictas in most cases, data are not available for thereceiving lake prior to introductions. However, stableisotope analysis (SIA) may provide a powerfulmethod to study the prevailing niche segregationbetween introduced and native fishes (Cucheroussetet al. 2012).We used stomach contents and stable isotope
analyses, together with trophically transmitted para-sites (i.e. Diphyllobothrium sp. tapeworms), to inves-tigate the trophic niche of introduced lake trout andnative Arctic charr in Inarij€arvi prior to the end oflake trout stockings in 2009–2010. Our study focusedon large (total length >280 mm) lake trout and Arcticcharr because all lake trout stocked into Inarij€arvihave been of age 2–3 years and total length 200–350 mm (E. Salonen, Finnish Game and FisheriesResearch Institute, personal communication). Thetwo species show generally similar feeding behaviour(Martin & Olver 1980; Fraser & Power 1989),although lake trout are assumed to show a somewhatmore generalist foraging behaviour, consuming agreater diversity and size of prey than Arctic charrand also preying upon native Arctic charr (Johnson1980; Martin & Olver 1980). We hypothesised thatlake trout and Arctic charr in Inarij€arvi should showsignificant dietary overlap and thus compete for simi-lar food resources. If temporally stable, this hypothes-ised niche overlap should be evident from all thedietary metrics used (stomach contents, stable iso-topes and trophically transmitted parasites), reflectingthe trophic niche in different temporal resolutions,from a few hours or days (stomach contents) up toseveral years (parasites). Besides their trophic interac-tions, we also compared condition and growth of laketrout and Arctic charr to assess the performance ofthese species in Inarij€arvi. We expected that the spe-cies with a higher condition and growth rate wouldbe competitively superior to the other. Our study pro-vides detailed information about the current fish com-munity and food web structure in Inarij€arvi, therebyserving as a baseline for potential future studies afterthe end of the lake trout stocking era.
182
Eloranta et al.

Material and methods
Study lake
Inarij€arvi is a large (surface area = 1043 km2), deep(maximum depth = 92 m) and oligotrophic (TN =180 lg�l�1; TP = 9 lg�l�1) lake in northern Finland(N 68°580; E 27°400; Fig. 1). The lake drains to theArctic Ocean via the Paatsjoki River system, and thewater level is regulated (with maximum amplitude of2.4 m) by hydroelectric power plants. The littoralzone covers approximately 60% of the total surfacearea (delimited from bathymetric maps along thedetermined 8-m light-compensation depth contour[LI-1400 light meter, Li-Cor Inc., Lincoln, NE, USA;wavelength 400–700 nm]). During summer stratifica-tion, the epilimnion usually extends down to around15 m depth (see Fig. S1).The fish community in Inarij€arvi consists of 10
native fishes including whitefish, Arctic charr, browntrout, grayling (Thymallus thymallus), burbot (Lotalota), pike (Esox lucius), perch (Perca fluviatilis),nine-spined stickleback (Pungitius pungitius), three-spined stickleback (Gasterosteus aculeatus) andminnow (Phoxinus phoxinus). Whitefish is highlypolymorphic in this region, and sympatric littoral(large sparsely rakered, LSR), pelagic (denselyrakered, DR) and profundal (small sparsely rakered,SSR) morphs occur in Inarij€arvi (Harrod et al. 2010;Kahilainen et al. 2011b). Introduced fishes includevendace, land-locked salmon (Salmo salar m. sebago)and lake trout. Vendace was introduced to this regionin the 1950s, and its population in Inarij€arvi boomed
in the 1980s, with detrimental effects on the nativepelagic whitefish morph (Amundsen et al. 1999;Salonen et al. 2007). Vendace is known to reproduceand hybridise with native whitefish in the region(Kahilainen et al. 2011a). Land-locked salmon mayreproduce in the River Ivalojoki which feeds intoInarij€arvi, but hybridisation with native brown trouthas not been observed (Salonen & Mutenia 2007).Lake trout have been stocked annually to Inarij€arviduring 1972–2012, but have not been observed toreproduce naturally or to hybridise with Arctic charr(Salonen & Mutenia 2007). Annual stockings of allthree introduced fishes have now ceased and thuslake trout should die out unless they are able toreproduce in Inarij€arvi.
Sampling
Lake trout and Arctic charr were mainly caught using5- to 6-m-high and 60-m-long gill nets of knot-to-knot mesh sizes 40–60 mm from southern and centralparts of Inarij€arvi in September 2009 and 2010(Fig. 1). Potential prey fishes and some additionalArctic charr were caught using three sets of 1.8-m-high and 30-m-long gill nets of mesh sizes 12, 15,20, 25, 30, 35, 45 and 60 mm. Gill nets were setovernight in pelagic (0–5 m below the surface), lit-toral (0–5 m depth) and profundal (>15 m depth)habitats. Some prey fishes were sampled using asmall pelagic trawl (height 4 m, width 8 m, cod-endmesh size 3 mm) towed with two 20- to 40-hp out-board motorboats (for details, see Malinen et al.2014). All lake trout and all but three >280 mm
Fig. 1. The location of Lake Inarij€arvi in northern Finland. The exact sampling area is marked with an ellipse in the right-hand panel.
183
Trophic interactions between lake trout and Arctic charr

Arctic charr were caught with the 5- to 6-m-high gillnets from the profundal zone (15–25 m depth).Each fish was transported on ice to a field labora-
tory, where they were identified, measured (totallength, �1.0 mm) and weighed (wet mass, �0.1 g).Whitefish morphs and vendace were identified bytheir distinct head and gill raker morphologies(Kahilainen & Østbye 2006; Kahilainen et al. 2011b).Sagittal otoliths were removed from each lake troutand Arctic charr. Stomachs were removed and imme-diately frozen at �20°C for later dietary analyses. Inaddition, the number of Diphyllobothrium sp. tape-worm cysts on the stomach wall of each lake troutand Arctic charr was counted. Diphyllobothrium sp.parasites are transmitted to piscivorous fish via pela-gic intermediate hosts (copepods and planktivorousprey fishes); thus their abundance in fish tissues canbe used as a proxy for long-term use of pelagic prey(Knudsen et al. 2008, 2014). From each lake troutand Arctic charr, as well as from random subsamples(n = 30–50) of potential prey fishes, a small block ofdorsal muscle was dissected and stored at �20°Cprior to SIA.Qualitative samples of pelagic zooplankton and lit-
toral and profundal benthic macroinvertebrate preywere collected for SIA in September 2009. Zooplank-ton were sampled with replicate (n = 5–8) semi-hori-zontal hauls from 15 m depth up to the surface usinga 50-lm mesh plankton net. Zooplankton SIA sam-ples were subsequently sieved through 50-, 100- and200-lm meshes to separate large cladocerans and co-pepods from detritus and other unwanted material.Benthic macroinvertebrates were collected for SIAalong a transect by taking three replicate sampleswith an Ekman grab (area 272 cm2) from depths of1, 2, 3, 5, 10, 15, 20, 30 and 40 m. Additional quali-tative samples were taken from the shallow littoralzone (0–1 m depth) using a 500-lm mesh kick net.All macroinvertebrate samples were sieved through a500-lm mesh prior to sorting and identification inthe laboratory. All zooplankton and benthic macroin-vertebrates were stored at �20°C for later SIA prepa-ration. The mean � SD d13C and d15N values forlittoral and profundal benthic macroinvertebrateswere calculated from samples collected from depthsof 0–3 and 15–40 m, respectively.
Stomach content analyses
Stomach content analyses (SCA) were conducted toinvestigate taxonomic composition and relativeimportance of recently ingested prey items and subse-quently evaluate interspecific differences in diets oflake trout and Arctic charr. Stomach fullness of eachlake trout and Arctic charr was visually estimatedwith a scale from 0 (empty) to 10 (fully extended),
and the relative contribution of each prey taxon tototal stomach fullness was evaluated according toHynes (1950). From each stomach, total lengths(accuracy 1.0 mm) of undigested prey fishes weremeasured using a ruler. Degree of prey selectivity bylake trout and Arctic charr was estimated usingIvlev’s (1961) electivity index, which compares therelative proportions of prey fish species i in the pred-ator stomach contents and in the environment. Here,availability of each prey fish species was based onnumerical catches from 1.8-m-high gill net sets andfrom pelagic trawl. Index values range between �1and +1, with values under �0.5 indicating avoidanceof the prey item, values �0.49 to 0.49 indicatingrandom selection and values over 0.5 indicatingactive selection.Dietary overlap between lake trout and Arctic charr
was tested with a nested analysis of similarities(ANOSIM) using the PAST 2.12 program (Hammeret al. 2001). Levin’s index (1968) was calculated toestimate dietary niche width, whereas a proportionalsimilarity (PSi) index (Bolnick et al. 2002) was cal-culated to evaluate individual dietary specialisationwithin populations of lake trout and Arctic charr. ThePSi index approaches 1 when the individual has asimilar diet to that of the whole population.
Stable isotope analyses
Stable isotope analyses (SIA) were used to investi-gate the general structure of the Inarij€arvi food weband particularly to estimate overlap in long-term tro-phic niches of lake trout and Arctic charr. All frozenSIA samples were freeze-dried (Alpha 1-4 LD Plus,Martin Christ Gefriertrocknungsanlagen GmbH, Oste-rode, Germany) for 48 h, homogenised and weighed(0.500–0.600 mg) into tin cups prior to analysis witha FlashEA 1112 elemental analyser coupled to aThermo Finnigan DELTAplus Advantage mass spec-trometer at the University of Jyv€askyl€a. In each run,the standard deviation of an internal working stan-dard (pike white muscle tissue) was <0.24& for d13Cand 0.20 & for d15N. Prior to subsequent analyses,raw d13C values from fish muscle tissue were cor-rected for lipids according to Kiljunen et al. (2006).The Stable Isotope Analysis in R (SIAR, v. 4.1.3;
Parnell et al. 2010) package was used in R (v. 2.14.2;R Development Core Team 2012) to estimate relativecontributions of different prey fishes to the long-termdiets of lake trout and Arctic charr. SIAR uses aBayesian approach to consider uncertainties withrespect to variation in trophic fractionation and in iso-topic compositions of consumers and food sources(Parnell et al. 2010). Here, pelagic coregonids (i.e.vendace and DR whitefish), LSR whitefish, nine-spined stickleback and burbot were chosen as puta-
184
Eloranta et al.

tive food sources due to predominant piscivorousdiets of lake trout and Arctic charr observed here bySCA as well as in a nearby lake inhabited by Arcticcharr (Kahilainen & Lehtonen 2003). The commonlyused trophic fractionation factors of 0.4 � 1.3& ford13C and 3.4 � 1.0& for d15N were used (Post2002). These estimates correspond well to theobserved fractionation of d13C and d15N betweeninvertebrates and whitefish morphs sampled from thisregion (Harrod et al. 2010).The Stable Isotope Bayesian Ellipses in R (SIBER;
Jackson et al. 2011) method was used in SIAR to esti-mate isotopic niche widths of lake trout and Arcticcharr. Bayesian inference is used by SIAR to calculatestandard ellipse areas (SEA) in d13C–d15N space. Highoverlap in SEA ellipses was interpreted as strongresource competition, whereas lack of overlap or awide ellipse area (i.e. isotopic niche) was interpretedas reduced resource competition between the two sal-monids. Compared with traditional metrics (e.g. totalconvex hull area; Layman et al. 2012), the estimatedisotopic niche areas are less sensitive to differences insamples sizes and estimates the core niche of thepopulation or species regardless of possible outliers.
Condition and growth
The condition of lake trout and Arctic charr was esti-mated using the Fulton (1904) condition factor. Theage of lake trout and Arctic charr was determinedunder a stereomicroscope by submerging clear
otoliths in clean water to clarify annual rings. Oto-liths that were difficult to interpret were burned andcracked according to Christensen (1964). Growth ofthe two species was subsequently examined by calcu-lating and testing the mean � SD total length in eachyear class. Growth rate was also determined usingthe von Bertalanffy (1938) nonlinear growth model:
LT ¼ L1j1� e�kðt�t0Þj
where LT is the expected total length at time t, L∞ isthe asymptotic total length (i.e. theoretical totallength to which a fish would grow if time permitted),k is the rate at which the growth curve approachesthe asymptote and t0 is the hypothetical time at whichLT was zero (i.e. hatching time). Theoretical annualgrowth rate (x) of lake trout and Arctic charr wasfinally estimated as L∞ * k (mm�year�1; Gallucci &Quinn 1979).
Statistical analyses
Depending on normality and homoscedasticity of thedata, either t-test or Mann–Whitney U-test was usedto compare total length and mass, condition factor,mean total length at age, stomach fullness, mean totallength of prey fish, PSi indices, numbers of parasitecysts in stomach wall and muscle d13C and d15N val-ues between lake trout and Arctic charr. These statis-tical analyses were performed using PASW Statistics20 for Windows (SPSS Inc., Chicago, IL, USA).
Table 1. Size, condition, parasite load, stomach content data (SCA) and stable isotope data (SIA) measured from >280 mm lake trout and Arctic charr caughtfrom Lake Inarij€arvi, northern Finland, in September 2009–2010
Lake trout Arctic charr
StatisticsMean � SD Range Mean � SD Range
Size and parasitesn 79 53Total length (mm) 441 � 78 281 to 590 458 � 91 281 to 703 t = 1.15, df = 130, P = 0.254Total mass (g) 788 � 407 155 to 1634 1173 � 851 199 to 4540 U = 1483, n = 132, P = 0.005Condition factor K 0.83 � 0.10 0.46 to 1.03 1.04 � 0.12 0.75 to 1.31 U = 312, n = 132, P < 0.001Diphyllobothrium sp. (n) 2.5 � 4.1 0 to 24 14.2 � 19.2 0 to 97 U = 885, n = 132, P < 0.001
SCAn 79 (26) 44 (21)Stomach fullness (0–10) 2.2 � 2.3 0 to 10 1.4 � 2.5 0 to 9 U = 1606, n = 132, P = 0.018Levins B 5.1 1.3PSi (0–1) 0.31 � 0.12 0.04 to 0.53 0.46 � 0.21 0.05 to 0.61 U = 293, n = 73, P = 0.002Prey fish length (mm) 116 � 48 24 to 195 124 � 43 41 to 172 U = 278, n = 55, P = 0.412
SIAn 79 53d13C (&) �24.9 � 2.6 �28.9 to �20.8 �28.2 � 1.5 �31.5 to �23.6 U = 579, n = 132, P < 0.001d15N (&) 11.3 � 0.5 10.5 to 12.6 11.6 � 0.8 8.1 to 12.6 U = 1249, n = 132, P < 0.001SEAB 2.9 2.3 to 3.6 2.7 2.0 to 3.5
Sample sizes (n, number of empty stomachs in SCA in parentheses) and details of statistical tests (significant P-values are marked in bold) are also shown.Abbreviations: SEAB = Bayesian standard ellipse area (mean and 95% Bayesian credibility intervals); PSi = proportional similarity index (approaches 1 whenindividual has similar diet to that of the population).
185
Trophic interactions between lake trout and Arctic charr
Results
A total of 79 lake trout and 53 Arctic charr of totallength >280 mm were caught from Inarij€arvi in2009–2010. The species did not significantly differ in
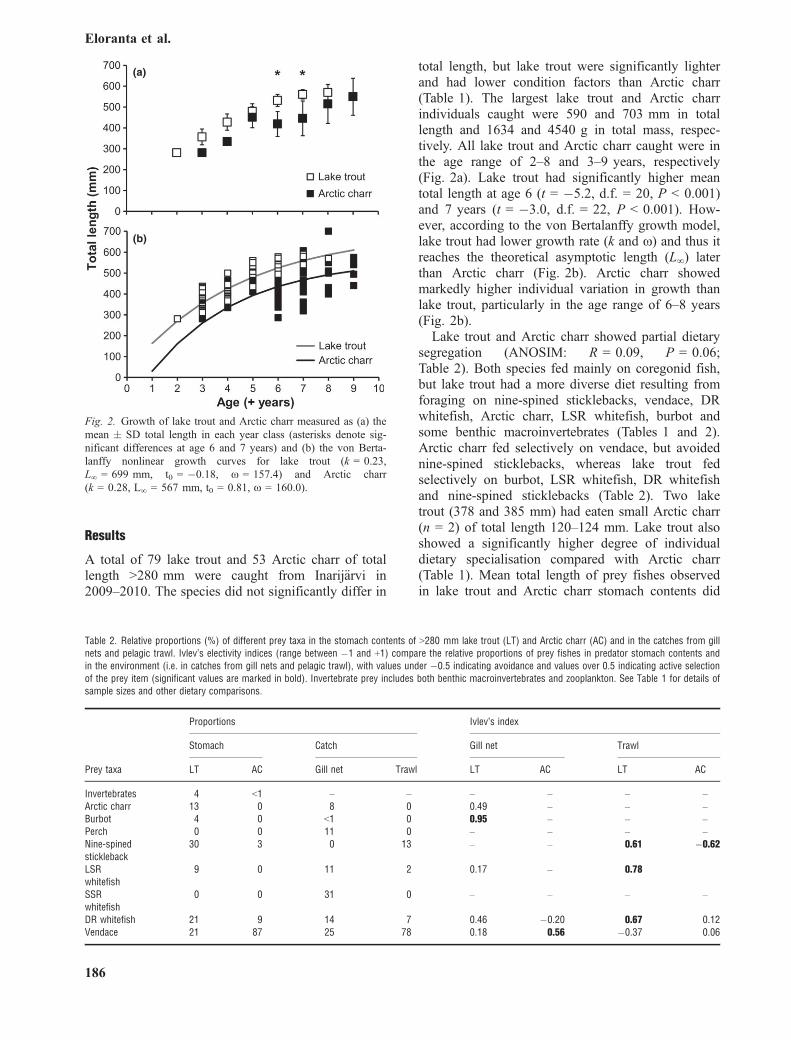
total length, but lake trout were significantly lighterand had lower condition factors than Arctic charr(Table 1). The largest lake trout and Arctic charrindividuals caught were 590 and 703 mm in totallength and 1634 and 4540 g in total mass, respec-tively. All lake trout and Arctic charr caught were inthe age range of 2–8 and 3–9 years, respectively(Fig. 2a). Lake trout had significantly higher meantotal length at age 6 (t = �5.2, d.f. = 20, P < 0.001)and 7 years (t = �3.0, d.f. = 22, P < 0.001). How-ever, according to the von Bertalanffy growth model,lake trout had lower growth rate (k and x) and thus itreaches the theoretical asymptotic length (L∞) laterthan Arctic charr (Fig. 2b). Arctic charr showedmarkedly higher individual variation in growth thanlake trout, particularly in the age range of 6–8 years(Fig. 2b).Lake trout and Arctic charr showed partial dietary
segregation (ANOSIM: R = 0.09, P = 0.06;Table 2). Both species fed mainly on coregonid fish,but lake trout had a more diverse diet resulting fromforaging on nine-spined sticklebacks, vendace, DRwhitefish, Arctic charr, LSR whitefish, burbot andsome benthic macroinvertebrates (Tables 1 and 2).Arctic charr fed selectively on vendace, but avoidednine-spined sticklebacks, whereas lake trout fedselectively on burbot, LSR whitefish, DR whitefishand nine-spined sticklebacks (Table 2). Two laketrout (378 and 385 mm) had eaten small Arctic charr(n = 2) of total length 120–124 mm. Lake trout alsoshowed a significantly higher degree of individualdietary specialisation compared with Arctic charr(Table 1). Mean total length of prey fishes observedin lake trout and Arctic charr stomach contents did
(a)
(b)
Fig. 2. Growth of lake trout and Arctic charr measured as (a) themean � SD total length in each year class (asterisks denote sig-nificant differences at age 6 and 7 years) and (b) the von Berta-lanffy nonlinear growth curves for lake trout (k = 0.23,L∞ = 699 mm, t0 = �0.18, x = 157.4) and Arctic charr(k = 0.28, L∞ = 567 mm, t0 = 0.81, x = 160.0).
Table 2. Relative proportions (%) of different prey taxa in the stomach contents of >280 mm lake trout (LT) and Arctic charr (AC) and in the catches from gillnets and pelagic trawl. Ivlev’s electivity indices (range between �1 and +1) compare the relative proportions of prey fishes in predator stomach contents andin the environment (i.e. in catches from gill nets and pelagic trawl), with values under �0.5 indicating avoidance and values over 0.5 indicating active selectionof the prey item (significant values are marked in bold). Invertebrate prey includes both benthic macroinvertebrates and zooplankton. See Table 1 for details ofsample sizes and other dietary comparisons.
Prey taxa
Proportions Ivlev’s index
Stomach Catch Gill net Trawl
LT AC Gill net Trawl LT AC LT AC
Invertebrates 4 <1 – – – – – –Arctic charr 13 0 8 0 0.49 – – –Burbot 4 0 <1 0 0.95 – – –Perch 0 0 11 0 – – – –Nine-spinedstickleback
30 3 0 13 – – 0.61 �0.62
LSRwhitefish
9 0 11 2 0.17 – 0.78
SSRwhitefish
0 0 31 0 – – – –
DR whitefish 21 9 14 7 0.46 �0.20 0.67 0.12Vendace 21 87 25 78 0.18 0.56 �0.37 0.06
186
Eloranta et al.

not significantly differ, but lake trout showed a largerrange in size of prey fishes (Table 1). Despite lowercondition, fewer lake trout stomachs were empty andfewer Diphyllobothrium sp. parasite cysts wereobserved in the stomach wall of lake trout than ofArctic charr (Table 1).Lake trout had a more littoral trophic niche com-
pared with Arctic charr as indicated by the significantlyhigher muscle d13C values (Table 1, Fig. 3a). Laketrout showed slightly lower d15N values compared withArctic charr (Table 1, Fig. 3a). Two individual Arcticcharr had exceptionally high d13C and low d15N val-ues, probably due to predominant foraging on littoralbenthic macroinvertebrates (Fig. 3b). Like SCA, theresult from the SIAR isotopic mixing model suggestedhigh consumption of nine-spined sticklebacks and LSRwhitefish by lake trout and pelagic coregonids (vend-ace and DR whitefish) by Arctic charr (Fig. 3c). Isoto-pic niches of lake trout and Arctic charr showed nosignificant difference in size, but overlapped onlyslightly because lake trout had a generally more littoralniche than Arctic charr (Table 1, Fig. 3b).
Discussion
Our study demonstrates that introduced lake trout canhave a largely piscivorous diet and prey to someextent upon small Arctic charr in lakes situated out-side the natural distribution area where the speciesco-occur (cf. Martin & Olver 1980; Fraser & Power1989). Although both lake trout and Arctic charr weremainly piscivores and caught from the deep profundalzone, the SCA and SIA data indicate that lake troutwere more generalist with respect to composition andsize of prey fishes and also relied more heavily onlittoral prey resources than the more pelagic Arcticcharr. The more specialised foraging on planktivo-rous coregonids probably explains the higher abun-dance of trophically transmitted Diphyllobothrium sp.parasites in Arctic charr. The initially higher growthrate but subsequently lower condition of lake troutmay be due to their hatchery-reared origin. Futuremonitoring is required for evaluation of potentialchanges in trophic interactions between lake trout andArctic charr in Inarij€arvi.Contrary to our main hypothesis, the SCA, SIA
and parasite data give congruent evidence for partialniche segregation between introduced lake trout andnative Arctic charr in large Inarij€arvi. Lake trout evi-dently used more littoral and diverse food resourcesthan Arctic charr. When dominating in the fish com-munity, Arctic charr have been observed to have awide trophic niche but generally prefer littoral foodand habitat resources (e.g. Karlsson & Bystr€om 2005;Eloranta et al. 2010). However, competitive andpredatory interactions with sympatric salmonids such
(a)
(c)
(b)
Fig. 3. Stable isotope data showing (a) the mean � SD d13Cand d15N values of lake trout and Arctic charr muscle tissueand of the putative invertebrate and fish prey, (b) the isotopicniches (standard ellipse areas) of lake trout (grey ellipse) andArctic charr (black ellipse) and (c) dietary proportions based onthe SIAR mixing model (mean � 95% Bayesian credibilityintervals). See Table 1 for further details of sample sizes and sta-tistics.
187
Trophic interactions between lake trout and Arctic charr

as brown trout may force Arctic charr to use a morepelagic or profundal trophic niche (Kahilainen &Lehtonen 2003; Eloranta et al. 2013 and referencestherein). Hence, the observed pelagic niche use byArctic charr may partly result from increased compe-tition for littoral resources following the introductionsof lake trout. Unlike in some North American lakeswhere piscivorous lake trout has been demonstratedto force Arctic charr to feed on invertebrate prey(Fraser & Power 1989), these two salmonids wereboth piscivorous top predators in the Inarij€arvi foodweb. However, lake trout preyed upon different spe-cies and sizes of prey fishes, including nine-spinedstickleback, whitefish, vendace, burbot and smallArctic charr, whereas Arctic charr had a less diversediet consisting mainly of small pelagic coregonids,particularly vendace. Moreover, lake trout evidentlyconsumed larger prey fishes than Arctic charr (John-son 1980; Martin & Olver 1980). Inclusion of morebenthic-associated fishes, such as burbot, in the dietof lake trout may also indicate differences in foragingtactics between the two salmonids. However, theobserved selective predation by lake trout on burbotmay partly result from low catchability of burbotwhen using gill nets and a pelagic trawl, meaningthey appeared selected because they were sampledfrom lake trout stomachs, but their estimated densi-ties in the lake could have been biased low.Our results from SCA and SIA suggest that the
wide dietary niche of lake trout is largely due tomarked individual differences in niche use (cf.Bolnick et al. 2002; Morbey et al. 2006). Some indi-vidual lake trout had exceptionally high or low mus-cle d13C values indicating predominantly littoral orpelagic diets, respectively, and also high individualvariation in feeding habitats (Morbey et al. 2006).The high d13C values are probably due to specialisedforaging on nine-spined sticklebacks, whereas thelow d13C values of most Arctic charr and also somelake trout probably indicate a long-term diet domi-nated by pelagic coregonids such as vendace and DRwhitefish. The selection of these prey fishes is likelyassociated with their high abundance in the pelagicareas in Inarij€arvi, as reported for nearby lakes (Jen-sen et al. 2008).Lake trout may benefit from a generalist and lit-
toral trophic niche as it may be adaptive for potentialannual fluctuations in density of pelagic prey fishes,particularly that of vendace (Salonen et al. 2007;Bøhn et al. 2008). Correspondingly, lake trout hasbeen shown to shift to a more littoral diet (consistingmainly of round goby, Neogobius melanostomus) dueto reduced pelagic prey fish (rainbow smelt, Osmerusmordax) abundances in Lake Ontario, North America(Rush et al. 2012). On the other hand, in Inarij€arvi,invasive vendace may have provided Arctic charr
with a profitable, alternative prey after the boom ofthis coregonid species in late 1980s (Salonen et al.2007). The littoral food and habitat resources in Inar-ij€arvi may also have been reduced due to water levelregulation for hydropower, which may have increasedthe relative contribution of pelagic production andsubsequently induced native top predators such asArctic charr to shift to a more pelagic niche (cf. Lueket al. 2013). The littoral trophic niche of some indi-vidual lake trout may also result from a delayed orlimited ability to leave the shallow littoral habitatwhich probably resembles the hatchery conditionsmore than the open-water pelagic and profundal habi-tats. Nevertheless, our study gives further support forthe commonly observed niche plasticity of introducedspecies (Sakai et al. 2001) and interactive niche seg-regation between coexisting salmonids (e.g. Nilsson1965; Jonsson et al. 2008).Lake trout may have replaced Arctic charr from
many high-latitude lakes in North America, poten-tially due to strong competitive and/or predatoryinteractions (Johnson 1976; Hershey et al. 1999).Although there is no sound evidence for majorimpacts on Inarij€arvi Arctic charr and the lake foodweb, our results indicate that introduced lake troutcan prey upon juvenile Arctic charr and thus poten-tially affect their recruitment and niche use (cf.Damsg�ard & Ugedal 1997; Keyse et al. 2007).Increased predation risk and resource competition fol-lowing lake trout introductions may both explain theearly shift of Inarij€arvi Arctic charr to a piscivorousniche (already at 130 mm total length). However,predatory and competitive impacts are difficult toevaluate due to lack of data on Arctic charr abun-dance and trophic ecology prior to lake trout intro-duction. Hence, future research is essential forevaluating potential changes in Arctic charr recruit-ment and population size as well as in the wholeInarij€arvi food web following the putative extinctionof lake trout.The congruent results from SCA and SIA demon-
strate temporally stable piscivorous niches for intro-duced lake trout and native Arctic charr. Lake troutalso showed generally higher individual variation inniche use and stronger reliance on pelagic foodresources than Arctic charr. The raw d13C and d15Nvalues were effectively transformed into more easilyinterpreted dietary estimates by the SIAR mixingmodel, indicating substantial foraging on nine-spinedsticklebacks by lake trout and on pelagic coregonids(vendace and DR whitefish) by Arctic charr, as wasalso suggested by the SCA data. Although our studylacks data on potential seasonal fluctuations in taxo-nomic and isotopic compositions of pelagic and ben-thic invertebrates (cf. Syv€aranta et al. 2006), theresults from SIAR are still reliable due to the piscivo-
188
Eloranta et al.

rous diets of lake trout and Arctic charr almostentirely lacking invertebrate prey, and also due to theobserved seasonal constancy of Arctic charr musclestable isotope values (Eloranta et al. 2010). Theresults from SCA and SIA were further supported bythe parasite data that indicated a generally higherabundance and prevalence of Diphyllobothrium sp.tapeworms in Arctic charr than in lake trout that usedless pelagic prey. The observed differences in abun-dance and prevalence of Diphyllobothrium sp. furthersupport the observed individual differences in nicheuse within the Arctic charr and lake trout populations(Bolnick et al. 2002).The length at age of lake trout was slightly higher
than that of Arctic charr, at least partly reflecting thehatchery-reared origin of the former species. The vonBertalanffy growth model indicated slower growth (kand x) for lake trout. The generally poorer conditionof lake trout may indicate inferior ability to forage inthe natural environment, although stomach fullnessestimates suggest more active foraging for lake troutthan Arctic charr. The hatchery-reared origin of laketrout may also explain the observed low individualvariation in growth. Despite the relatively slowgrowth and low condition, successful reproductionand/or introgression of lake trout could pose a seriousthreat to native Arctic charr and food web structurein Inarij€arvi. However, we observed no maturefemales and only one mature male lake trout of totallength 405 mm. Despite this lack of evidence fromInarij€arvi (Salonen & Mutenia 2007), introduced laketrout has been reported to reproduce naturally in sev-eral European lakes (Langeland 1994; Crossman1995; Delacoste et al. 1997). Hence, future monitor-ing of population size and age structure of Inarij€arvilake trout is essential for observing potential naturalreproduction and hybridisation as well as whole eco-system level impacts.Like many other fish stocking programmes (Gozlan
et al. 2010), the initial justification for lake troutintroductions in Inarij€arvi was to compensate lossesin salmonid stocks resulting from hydropower devel-opment and to diversify a relatively species-poor eco-system (Mutenia & Salonen 1994; Salonen &Mutenia 2007). Since its introduction in 1972, laketrout has become an economically important speciesfor commercial and recreational fishing (Salonen &Mutenia 2007). Lake trout has made a significantaddition to fish catches particularly in winter whenother salmonid species reportedly swim less actively(local fisherman T. Aarnipuro, personal communica-tion). The annual stockings of non-native and nativesalmonids may become problematic because they canmaintain higher predator populations than would nat-urally occur, especially in oligotrophic, unproductivelakes (Johnson & Martinez 2000). The high stocking
rate of piscivorous salmonids may also have strongtop-down effects on pelagic prey fish populations(mainly vendace and DR whitefish). Besides potentialeconomic impacts, ecological consequences of laketrout stockings in Inarij€arvi may show a lag phase sothat potential natural reproduction of the species andany impacts on the whole ecosystem may only appeardecades or even centuries after stocking has ended(Gozlan et al. 2010). For example, this may happen inthe future due to climate change, if the failure of laketrout reproduction is related to cold-water tempera-tures. This scenario may be supported by observationsfrom the natural distribution areas, where lake troutcommonly inhabit more southern and warmer lakesthan Arctic charr (Hershey et al. 2006). For example,in Lake Superior, the number of days with availablepreferred thermal habitat (10 � 2°C) for (lean) laketrout has been observed to increase at a mean rate ofsix days per decade between 1979 and 2006 (Clineet al. 2013). Similar shifts in thermal niche availabilityfor lake trout may also occur in European lakes due topotential climate change.To conclude, the present coexistence of lake trout
and Arctic charr in Inarij€arvi may be supported by theobserved partial dietary segregation along the littoral–pelagic resource axis, as well as by the evidentlyunsuccessful natural reproduction of lake trout limit-ing population growth. Further research is needed toevaluate potential future changes in the food webstructures and ecosystem function following the intro-duction of lake trout as well as some other potentialinvasive fishes in Inarij€arvi and other European high-latitude lakes (cf. Jeppesen et al. 2010).
Acknowledgements
We are grateful to Jouni Kervinen, Pekka Antti-Poika, OlliSaari and Viljami Kinnunen for help during the fieldwork andTapio Aarnipuro for providing additional fish samples. Wealso thank Roger I. Jones for checking the grammar and tworeferees for their constructive comments on the manuscript.Muddusj€arvi Research Station and the Municipality of Inarikindly provided fish processing facilities and equipment dur-ing the fieldwork. The study was supported by project grantsfrom the Ministry of Agriculture and Forestry and Academyof Finland (140903), and by personal grants to APE from theJenny and Antti Wihuri Foundation, the Ellen and ArtturiNyyss€onen Foundation and the Ella and Georg EhrnroothFoundation, to PN from the Vanamo Biological Society ofFinland and from the Olvi Foundation and to KKK from theEuropean Regional Development Fund (A30205).
References
Amundsen, P.-A., Staldvik, F.J., Reshetnikov, Y.S., Kashulin,N., Lukin, A., Bøhn, T., Sandlund, O.T. & Popova, O.A.
189
Trophic interactions between lake trout and Arctic charr

1999. Invasion of vendace Coregonus albula in a subarcticwatercourse. Biological Conservation 88: 405–413.
von Bertalanffy, L. 1938. A quantitative theory of organicgrowth (inquiries on growth laws II). Human Biology 10:181–213.
Bolnick, D.I., Yang, L.H., Fordyce, J.A., Davis, J.M. &Svanb€ack, R. 2002. Measuring individual level resource spe-cialization. Ecology 83: 2936–2941.
Christensen, J.M. 1964. Burning of otoliths, a technique forage determination of soles and other fish. Journal du ConseilInternational pour l’Exploration de la Mer 29: 73–81.
Cline, T.J., Bennington, V. & Kitchell, J.F. 2013. Climatechange expands the spatial extent and duration of preferredthermal habitat for Lake Superior fishes. PLoS One 8:e62279.
Crawford, S.S. & Muir, A.M. 2008. Global introductions ofsalmon and trout in the genus Oncorhynchus: 1870–2007.Reviews in Fish Biology and Fisheries 18: 313–344.
Crossman, E.J. 1995. Introduction of the lake trout (Salvelinusnamaycush) in areas outside its native distribution: a review.Journal of Great Lakes Research 21: 17–29.
Cucherousset, J. & Olden, J.D. 2011. Ecological impacts ofnon-native freshwater fishes. Fisheries 36: 215–230.
Cucherousset, J., Bouletreau, S., Martino, A., Roussel, J.-M.& Santoul, F. 2012. Using stable isotope analyses to deter-mine the ecological effects of non-native fishes. FisheriesManagement and Ecology 19: 111–119.
Damsg�ard, B. & Ugedal, O. 1997. The influence of predationrisk on habitat selection and food intake by Arctic charr,Salvelinus alpinus (L.). Ecology of Freshwater Fish 6:95–101.
Delacoste, M., Baran, P., Lascaux, J.M., Abad, N. & Besson,J.P. 1997. Bilan des introductions de salmonid�es dans les lacset ruisseaux d’altitude des Hautes-Pyr�en�ees. Bulletin FrancaisDe La Peche Et De La Pisciculture 344/354: 205–219.
Donald, D.B. & Alger, D.J. 1993. Geographic distribution,species displacement, and niche overlap for lake trout andbull trout in mountain lakes. Canadian Journal of Zoology71: 238–247.
Eby, L.A., Roach, W.J., Crowder, L.B. & Stanford, J.A. 2006.Effects of stocking-up freshwater food webs. Trends in Ecol-ogy and Evolution 21: 576–584.
Eloranta, A.P., Kahilainen, K.K. & Jones, R.I. 2010. Sea-sonal and ontogenetic shifts in diet of Arctic charr Salveli-nus alpinus in a subarctic lake. Journal of Fish Biology77: 80–97.
Eloranta, A.P., Knudsen, R. & Amundsen, P.-A. 2013. Nichesegregation of coexisting Arctic charr (Salvelinus alpinus)and brown trout (Salmo trutta) constrains food web couplingin subarctic lakes. Freshwater Biology 58: 207–221.
Fraser, N.C. & Power, G. 1989. Influences of lake trout onlake-resident Arctic char in Northern Quebec, Canada.Transactions of the American Fisheries Society 118: 36–45.
Fulton, T.W. 1904. The rate of growth of fishes. AnnualReport of the Fishery Board of Scotland 22: 141–241.
Gallucci, V.F. & Quinn, T.J. 1979. Reparameterizing, fitting,and testing a simple growth model. Transactions of theAmerican Fisheries Society 108: 14–25.
Gozlan, R.E., Britton, J.R., Cowx, I. & Copp, G.H. 2010. Cur-rent knowledge on non-native freshwater fish introductions.Journal of Fish Biology 76: 751–786.
Hammer, Ø., Harper, D.A.T. & Ryan, P.D. 2001. PAST: pale-ontological statistics software package for education anddata analysis. Palaentologia Electronica 4: 1–9.
Harrod, C., Mallela, J. & Kahilainen, K.K. 2010. Phenotype-environment correlations in a putative whitefish adaptiveradiation. Journal of Animal Ecology 79: 1057–1068.
Hershey, A.E., Gettel, G.M., McDonald, M.E., Miller, M.C.,Mooers, H., O’Brien, W.J., Pastor, J., Richards, C. & Schu-ldt, J.A. 1999. A geomorphic–trophic model for landscapecontrol of Arctic lake food webs. BioScience 49: 887–897.
Hershey, A.E., Beaty, S., Fortino, K., Keyse, M., Mou, P.P.,O’Brien, W.J., Ulseth, A.J., Gettel, G.A., Lienesch, P.W.,Luecke, C., McDonald, M.E., Mayer, C.H., Miller, M.C.,Richards, C., Schuldt, J.A. & Whalen, S.C. 2006. Effect oflandscape factors on fish distribution in arctic Alaskan lakes.Freshwater Biology 51: 39–55.
H�orreo, J.L., Ayll�on, J., Perez, J., Beall, E. & Garcia-Vazquez,E. 2011. Interspecific hybridization, a matter of pioneering?Insights from Atlantic salmon and brown trout. Journal ofHeredity 102: 237–242.
Hynes, H.B.N. 1950. The food of freshwater sticklebacks(Gasterosteus aculeatus and Pygosteus pungitius), with areview of methods used in studies of the food of fishes.Journal of Animal Ecology 19: 36–58.
Ivlev, V.S. 1961. Experimental ecology of the feeding offishes. New Haven: Yale University Press, viii + 302 pp.
Jackson, A.L., Inger, R., Parnell, A.C. & Bearhop, S. 2011.Comparing isotopic niche widths among and within commu-nities: SIBER – Stable Isotope Bayesian Ellipses in R. Jour-nal of Animal Ecology 80: 595–602.
Jensen, H., Kahilainen, K.K., Amundsen, P.-A., Gjelland,K.Ø., Tuomaala, A., Malinen, T. & Bøhn, T. 2008. Preda-tion by brown trout (Salmo trutta) along a diversifying preycommunity gradient. Canadian Journal of Fisheries andAquatic Sciences 65: 1831–1841.
Jeppesen, E., Meerhoff, M., Holmgren, K., Gonz�ales-Bergonz-oni, I., Teixeira-de Mello, F., Declerck, S.A.J., De Meester,L., Søndergaard, M., Lauridsen, T.L., Bjerring, R., Conde-Porcuna, J.M., Mazzeo, N., Iglesias, C., Reizenstein, M.,Malmquist, H.J., Liu, Z., Balayla, D. & Lazzaro, X. 2010.Impacts of climate warming on lake fish community struc-ture and potential effects on ecosystem function. Hydrobio-logia 646: 73–90.
Johnson, L. 1976. Ecology of arctic populations of lake trout,Salvelinus namaycush, lake whitefish, Coregonus clupeafor-mis, Arctic char, S. alpinus, and associated species in unex-ploited lake of the Canadian Northwest Territories. Journalof the Fisheries Research Board of Canada 33: 2459–2488.
Johnson, L. 1980. The Arctic charr, Salvelinus alpinus.In: Balon, E.K., ed. Charrs: salmonid fishes of the genusSalvelinus. The Hague: Dr. W. Junk Publishers, pp. 15–98.
Johnson, B.M. & Martinez, P.J. 2000. Trophic economics oflake trout management in reservoirs of differing productiv-ity. North American Journal of Fisheries Management 20:127–143.
Jonsson, B., Jonsson, N., Hindar, K., Northcote, T.G. &Engen, S. 2008. Asymmetric competition drives lake use ofcoexisting salmonids. Oecologia 157: 553–560.
Kahilainen, K. & Lehtonen, H. 2003. Piscivory and preyselection of four predator species in a whitefish dominatedsubarctic lake. Journal of Fish Biology 63: 659–672.
190
Eloranta et al.

Kahilainen, K. & Østbye, K. 2006. Morphological differentia-tion and resource polymorphism in three sympatric whitefishCoregonus lavaretus (L.) forms in a subarctic lake. Journalof Fish Biology 68: 63–79.
Kahilainen, K.K., Østbye, K., Harrod, C., Shikano, T., Mali-nen, T. & Meril€a, J. 2011a. Species introduction promoteshybridization and introgression in Coregonus: is there signof selection against hybrids? Molecular Ecology 20: 3838–3855.
Kahilainen, K.K., Siwertsson, A., Gjelland, K.Ø., Knudsen,R., Bøhn, T. & Amundsen, P.-A. 2011b. The role of gillraker number variability in adaptive radiation of coregonidfish. Evolutionary Ecology 25: 573–588.
Karlsson, J. & Bystr€om, P. 2005. Littoral energy mobilizationdominates energy supply for top consumers in subarcticlakes. Limnology and Oceanography 50: 538–543.
Keyse, M.D., Fortino, K., Hershey, A.E., O’Brien, W.J., Lie-nesch, P.W., Luecke, C. & McDonald, M.E. 2007. Effects oflarge lake trout (Salvelinus namaycush) on the dietary habitsof small lake trout: a comparison of stable isotopes (d15Nand d13C) and stomach content analyses. Hydrobiologia579: 175–185.
Kiljunen, M., Grey, J., Sinisalo, T., Harrod, C., Immonen, H.& Jones, R.I. 2006. A revised model for lipid-normalizingd13C values from aquatic organisms, with implications forisotope mixing models. Journal of Applied Ecology 43:1213–1222.
Knudsen, R., Amundsen, P.-A., Nilsen, R., Kristoffersen, R. &Klemetsen, A. 2008. Food borne parasites as indicators oftrophic segregation between Arctic charr and brown trout.Environmental Biology of Fishes 83: 107–116.
Knudsen, R., Siwertsson, A., Adams, C.E., Newton, J. &Amundsen, P.-A. 2014. Similar patterns of individual nicheuse are revealed by different time-integrated trophic tracers(stable isotopes and parasites). Ecology of Freshwater Fish23: 259–268.
Korsu, K., Huusko, A. & Muotka, T. 2010. Invasion of northEuropean streams by brook trout: hostile takeover or pre-adapted habitat niche segregation? Biological Invasions 12:1363–1375.
Langeland, A. 1994. Successful introduction of lake trout(Salvelinus namaycush) in three Norwegian lakes. In: Cowx,I.G., ed. Rehabilitation of Freshwater Fisheries. Oxford:Fishing News Books, pp. 289–292.
Layman, C.A., Ara�ujo, M.S., Boucek, R., Hammerschlag-Peyer,C.M., Harrison, E., Jud, Z.R., Matich, P., Rosenblatt, A.E.,Vaudo, J.J., Yeager, L.A., Post, D.M. & Bearhop, S. 2012.Applying stable isotope to examine food-web structure: anoverview of analytical tools. Biological Reviews 87: 545–562.
Levins, R. 1968. Evolution in changing environments: sometheoretical explorations. New Jersey: Princeton UniversityPress, 132 pp.
Luek, A., Morgan, G.E., Wissel, B., Gunn, J.M. & Ramcha-ran, C.W. 2013. Impaired littoral energy pathways cause ashift to pelagic resources by fish in recovering lake foodwebs. Ecology of Freshwater Fish 22: 348–360.
Mack, R.N., Simberloff, D., Lonsdale, W.M., Evans, H.,Clout, M. & Bazzaz, F.A. 2000. Biotic invasions: causes,epidemiology, global consequences, and control. EcologicalApplications 10: 689–710.
Malinen, T., Tuomaala, A., Lehtonen, H. & Kahilainen, K.K.2014. Hydroacoustic assessment of mono- and polymorphicCoregonus density and biomass in subarctic lakes. Ecologyof Freshwater Fish 23: 424–437.
Martin, N.V. & Olver, C.H. 1980. The lake charr, Salvelinusnamaycush. In: Balon, E.K., ed. Charrs: salmonid fishes ofthe genus Salvelinus. The Hague: Dr. W. Junk Publishers,pp. 205–277.
Middleton, A.D., Morridon, T.A., Fortin, J.K., Robbins, C.T.,Proffitt, K.M., White, P.J., McWhirter, D.E., Koel, T.M.,Brimeyer, D.G., Fairbanks, W.S. & Kauffman, M.J. 2013.Grizzly bear predation links the loss of native trout to thedemography of migratory elk in Yellowstone. Proceedingsof the Royal Society of London. Series B. Biological Sci-ences 280: 20130870.
Morbey, Y.E., Addison, P., Shuter, B.J. & Vascotto, K. 2006.Within-population heterogeneity of habitat use by lake troutSalvelinus namaycush. Journal of Fish Biology 69: 1675–1696.
Mutenia, A. & Salonen, E. 1994. Rehabilitation of the fisher-ies of Lake Inari, northern Finland. In: Cowx, I.G., ed.Rehabilitation of Freshwater Fisheries. Oxford: FishingNews Books, pp. 280–289.
Nilsson, N.-A. 1965. Food segregation between salmonid spe-cies in North Sweden. Report of the Institute for FreshwaterResearch, Drottningholm 2: 58–78.
Parnell, A.C., Inger, R., Bearhop, S. & Jackson, A.L. 2010.Source partitioning using stable isotopes: coping with toomuch variation. PLoS One 5: e9672.
Post, D.M. 2002. Using stable isotopes to estimate trophicposition: models, methods, and assumptions. Ecology 83:703–718.
R Development Core Team. 2012. R: a language and environ-ment for statistical computing. Vienna, Austria: R Founda-tion for Statistical Computing.
Rahel, F.J. 2002. Homogenization of freshwater faunas.Annual Review of Ecology and Systematics 33: 291–315.
Rush, S.A., Paterson, G., Johnson, T.B., Drouillard, K.G.,Haffner, G.D., Hebert, C.E., Arts, M.T., McGoldrick, D.J.,Backus, S.M., Lantry, B.F., Lantry, J.R., Schaner, T. & Fisk,A.T. 2012. Long-term impacts of invasive species on anative top predator in a large lake system. Freshwater Biol-ogy 57: 2342–2355.
Ruzycki, J.R., Beauchamp, D.A. & Yule, D.L. 2003. Effectsof introduced lake trout on native cutthroat trout in Yellow-stone lake. Ecological Applications 13: 23–37.
Sakai, A.K., Allendorf, F.W., Holt, J.S., Lodge, D.M., Molof-sky, J., With, K.A., Baughman, S., Cabin, R.J., Cohen, J.E.,Ellstrand, N.C., McCauley, D.E., O’Neil, P., Parker, I.M.,Thompson, J.N. & Weller, S.G. 2001. The population biol-ogy of invasive species. Annual Review of Ecology andSystematics 32: 305–332.
Salonen, E. & Mutenia, A. 2007. Alien fish species in northern-most Finland. Riista- ja kalatalous – Tutkimuksia 2: 1–16(Report of the Finnish Game and Fisheries Research Institute).
Salonen, E., Amundsen, P.-A. & Bøhn, T. 2007. Boom andbust development of invading vendace Coregonus albula inthe subarctic Inari-Pasvik watershed (Finland, Norway andRussia). Advances in Limnology 60: 331–342.
Shindler, D.W. & Smol, J.P. 2006. Cumulative effects of cli-mate warming and other human activities on freshwaters
191
Trophic interactions between lake trout and Arctic charr

of Arctic and subarctic North America. Ambio 35: 160–168.
Simon, K.S. & Townsend, C.R. 2003. Impacts of freshwaterinvaders at different levels of ecological organisation, withemphasis on salmonids and ecosystem consequences. Fresh-water Biology 48: 982–994.
Swanson, H.K., Kidd, K.A. & Reist, J.D. 2010. Effects ofpartially anadromous Arctic charr (Salvelinus alpinus)populations on ecology of coastal Arctic lakes. Ecosystems11: 261–274.
Syv€aranta, J., H€am€al€ainen, H. & Jones, R.I. 2006. Within-lakevariability in carbon and nitrogen stable isotope signatures.Freshwater Biology 51: 1090–1102.
Tronstad, L.M., Hall, R.O. Jr, Koel, T.M. & Gerow, K.G.2010. Introduced lake trout produced four-level trophic cas-cade in Yellowstone lake. Transaction of the American Fish-eries Society 139: 1536–1550.
Wilson, C.C. & Bernatchez, L. 1998. The ghost of hybridspast: fixation of arctic charr (Salvelinus alpinus) mitochon-drial DNA in an introgressed population of lake trout (S. na-maycush). Molecular Ecology 7: 127–132.
Wilson, C.C. & Hebert, P.D.N. 1993. Natural hybridizationbetween Arctic char (Salvelinus alpinus) and lake trout (S.namaycush) in the Canadian Arctic. Canadian Journal ofFisheries and Aquatic Sciences 50: 2652–2685.
Supporting Information
Additional Supporting Information may be found inthe online version of this article:Fig. S1. Water temperature profile measured from
Lake Inarij€arvi, northern Finland, during summerstratification on 8 September 2009.
192
Eloranta et al.





























