Embed Size (px)
Citation preview

1997;57:1922-1928. Published online May 1, 1997.Cancer Res Monica Moro, Micaela Pelagi, Giulia Fulci, et al.
αBiotinylated Tumor Necrosis Factor Tumor Cell Targeting with Antibody-Avidin Complexes and
Updated Version http://cancerres.aacrjournals.org/content/57/10/1922
Access the most recent version of this article at:
Citing Articles http://cancerres.aacrjournals.org/content/57/10/1922#related-urls
This article has been cited by 18 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
[email protected] atTo order reprints of this article or to subscribe to the journal, contact the AACR Publications
To request permission to re-use all or part of this article, contact the AACR Publications
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from

CANCER RESEARCH 57, 1922-1928. May 15, 1997]
ABSTRACT
Tumor pretargeting with biotinylated antibodies and avidin, followedby a delayed delivery of radioactive-labeled biotin, is currently used for in
vivodiagnosis and therapy in cancer patients. Herein, we describe the useof a three-step antibody/avidin targeting approach to increase the localconcentration and the persistence of biotinylated human tumor necrosisfactor a (bio-TNF) on a mouse tumor. Mouse RMA lymphoma cells weretransfected with the Thy 1.1 allele (RMA.Thy 1.1) to generate a uniquetumor-associated antigen. In vitro pretargeting of RMA-Thy 1.1 cells with
the biotinylated anti-Thy 1.1 monoclonal antibody 19E12 (blo-19E12) andNeutrAvidin Increased the amount of bio-TNF that bound to the cell(10-20 times in comparison with non-pretargeted cells), as well as its
half.life on the surface (>30 times). Furthermore, cell pretargeting reduced by more than 2 orders of magnitude the LD@,of bio-TNF in acytolytic assay with actinomycin D. Finally, RMA.Thy 1.1 cells, pretreatedin vilro with blo-TNF according to the three-step procedure and injected
into syngeneicC57/BL6mice, were less tumorigenic than controls.These results indicate that the three-step targeting approach markedly
increases the amount and the persistence of bio-TNF on the cell surfaceand that cell-bound blo.TNF can trigger cytolytic effects in vitro andantitumor effects in vivo.Tumor pretargeting with biotinylated antibodiesand avidin could be a novel strategy for increasing the therapeutic indexof TNF.
INTRODUCTION
TNF3 is a macrophage-denved cytokine originally identified for itsability to induce hemorragic necrosis of transplanted solid tumors in
mice and for its cytotoxic activity against some tumor cell lines invitro (1, 2). In addition, TNF exerts a wide variety of other biologicaleffects on normal cells and is a primary mediator in many inflamma
tory, immunological, and pathophysiological reactions (3, 4). In spiteof its efficient antitumor activity observed in murine models, itstherapeutic application in humans is hampered by severe systemictoxicity and harmful side effects (5).
Bioactive TNF is a Mr 50,000 homotrimeric protein that slowly
dissociates into inactive subunits at picomolar levels (6—9). Most
biological effects exerted by trimeric TNF are mediated by the clustering of two membrane receptors (TNF-R55 and TNF-R75) of Mr55,000—60,000and 75,000—80,000,respectively(10). TNF-R75 isthought to play a crucial role in mediating the systemic toxicity ofTNF, because human TNF, which binds only the mouse TNF-R55, isconsiderably less toxic than mouse TNF in mice ( 11). On the otherhand, human TNF can exert antitumor effects in mice, suggesting thatTNF-R55 can provide sufficient signals for mediating antitumoreffects (12).
Received I 1/7/96; accepted 3/24/97.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
I This work was supported by Associazione Italiana Ricerca sul Cancro (AIRC). M. M.
was recipient of a fellowship from AIRC.2 To whom requests for reprints should be addressed.
3 The abbreviations used are: TNF, tumor necrosis factor; TNF-R55, TNF receptor
type I (p55); TNF-R75, TNF receptor type 2 (p75); bio-TNF, biotinylated human TNF;bio-19E12. biotinylated l9E12; NAy, NeutrAvidin; mAb. monoclonal antibody; FACS,fluorescence-activated cell sorter.
These notions led to the development of new approaches to overcome the problem of systemic toxicity in cancer therapy, based on theadministration of TNF mutants that have largely lost their ability tobind human TNF-R75 (13). Other strategies aimed at increasing thetherapeutic potential of TNF have been attempted [e.g., strategiesbased on tumor targeting with antibody-TNF conjugates (14) or withTNF gene-modified tumor-infiltrating lymphocytes (15) or based onlocoregional treatments (16, 17)]. Although the success of the TNFtargeting approaches still remains to be proved, the remarkable resultsobtained with locoregional treatments strengthen the hypothesis that alocalization of TNF action could increase its therapeutic index incancer therapy.
Tumor pretargeting with biotinylated antibodies and avidin orstreptavidin, followed by a delayed delivery of radioactivity-labeledbiotin, is currently used for in vivo cancer diagnosis in patients (18)and for therapy (19). The good localization of the radiolabeled biotinobserved with carcinoembryonic antigen-positive tumors, cerebralgliomas, ocular melanoma, and neuroendocrine tumors pretargetedwith various mAbs (18, 20—22)implies that avidin can efficientlylocalize at tumor sites in patients. Therefore, we hypothesize thattumor avidinization strategies could be exploited to deliver to thetumor site also low doses (nontoxic) of biotinylated compounds, suchas bio-TNF or other biotinylated cytokines, taking advantage of thehigh affinity of the biotin-avidin interaction.
In this work, we try to address this hypothesis, and we show that byantibody/avidin pretargeting it is possible to increase the binding andthe time of persistence of TNF on tumor cells. Moreover, we showthat cell-bound bio-TNF is still able to induce cytolytic effects invitro, as well as to decrease the tumorigenicity of a mouse lymphomain vivo.
MATERIALS AND METHODS
Generation of RMA.Thy 1.1 Cells. RMA lymphoma cells carrying theThy 1.2 allelewere culturedin RPMI 1640,5% FCS, 2 mMglutamine,100IU/ml penicillin, 100 @.tg/mlstreptomycin, and 50 nM 2-mercaptoethanol at37°Cin 5% CO2. 95% air.
The cDNA encoding for the Thy 1.1 allele was cloned by reverse transcription-PCR from BWa@ thymoma cells using the following oligonucleotides:
5'-AAGTCCGAATfCTfGGCACCATG-3' (5' primer) and 5'-GCTCCT
G1T1'AAGCTT000CC-3' (3' primer). The PCR product was cloned into themammalian expression vector pRSI-neo. For transfection, 15 @gof DNA wereelectroporated into l0@ RMA lymphoma cells using a Bio-Rad Gene Pulserapparatus(250 V. 960 mF). TransfectedRMAcells that survivedselectionwith 1.8 mg/mi geneticin and that expressed Thy 1.1 were selected by cellsorting and subcloned. The RMA-Thy 1.1 clone 2 was selected for further
study. This clone was found to stably express Thy 1.1 antigen as observed byFACS analysis with the anti-Thy 1.1 mAb l9El2 of cells recovered from anestablished mouse tumor after 15 days after inoculation. The Thy 1. 1 antigen
density on the transfected RMA cells was similar to that of the Thy I .2 antigenexpressed by nontransfected cells. The mAb l9El2IFhy I.1 complex does notinternalize even after prolonged incubation at 37°C.
Preparation of bio-TNF and mAb bio-19E12. Sixty @lof a 0.5 mg/mlhuman TNF (l0@ units/mg; Polymed Medical Centre, GmbH, Glattburg, Germany) in double-distilled water were mixed with 6 pJ of a I M sodium
1922
Tumor Cell Targeting with Antibody-Avidin Complexes and Biotinylated TumorNecrosis Factor a@
Monica Moro, Micaela Pelagi, Giulia Fulci, Giovanni Paganelli, Paolo Dellabona, Giulia Casorati,Antonio G. Siccardi, and Angelo Corti2
Dipartimento di Ricerca Biologica e Tecnologica, San Raffaele Hospital Scientific institute, via Olgettina 58, 20132 Milan (M. M., M. P., G. F., P. D., G. C., A. C.]; Europeaninstitute of Oncology, 20141 Milan 1G. P.]: and Department ofBiology and Geneticsfor Medical Sciences, University ofMilan, 20132 Milan (A. G. S.], Italy
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from

Table IComparison of various TNF biotinylationprotocolsCytolytic
activity―BindingBiotin/TNFto
avidin(molar-NAV+NAV―agarosecCoderatio)pH(units/mg)(units/mg)(%)TNFA01.0
X [email protected] X10'0bio-TNFB666.87.0
X [email protected] [email protected]@ND―NDbio-TNFD668.83.1X [email protected] [email protected] 106<1.0 X10'>99bio-TNFF2758.83.0X l0@ND>99
ThREE-STEP TUMOR TARGETING WITH BIO-TNF
carbonate buffer, pH 6.8, and 6 @lof D-biotinyl-6-aminocapmic acid N-
hydroxysuccinimide ester (SocietáProdotti Antibiotici S.p.A, Milan, Italy) atvarious concentrations in DMSO. The mixtures were incubated for 3 h at roomtemperature and mixed with 7.5 @lof 1 MTris-HCI, pH 7.5. After incubationfor 1 h at room temperature, 240 @dof RPMI containing 10% FCS, 2 nmiglutamine, 100 lU/mI penicillin, 100 @g/mlstreptomycin, and 250 ng/mlamphotericin B were added to the mixture. The product was dialyzed overnightagainst 2 liters of 9 g/liter sodium chloride at 4°Cand kept at —20°C.Theproportion of biotinylated TNF over the total number of TNF molecules wasestimated by affinity chromatography on avidin (monomeric)-agarose gel(Sigma Chemical Co., St. Louis, MO). One ,.@gof bio-TNF in 100 @lof PBS
containing 5 gfliter BSA was loaded onto the column (1 ml of resin), washedwith PBS, and eluted with 0.1 Mglycine-HC1,pH 2.0. The TNF antigen wasdetected in the bound and unbound fractions by TNF ELISA.
The anti-Thy 1.1mAb l9El2 (kindly provided by Dr. A. Livingstone, BaselInstitute for Immunology, Basel, Switzerland) was biotinylated as follows: Iml of a mAb 19EI2 solution, I mg/mi in sodium bicarbonate, pH 8.5, wasmixed with 34 @dof a sulfo-NHS-LC-biotin (Pierce Chemical Co., Rockford,IL) solution, 1 mg/mI in water, incubated for 30 mm at 24°C,and dialyzedovernight against 2 liters of PBS at 4°C.The product (mAb bio-l9El2) waskept at —20°C.
TNF ELISA. TNF ELISA was carried out by sandwich assay with ananti-TNF mAb (mAb78) and a rabbit polyclonal antiserum as describedpreviously (9).
TNF Cytolytic Assays. Bioactive TNF was measured by a cytolytic assayusing mouse WEHI 164clone 13(23) or RMA-Thy 1.1cells. WHEI cells werecultured in RPMI 1640 containing 10% FCS, 2 mM glutamine, 100 lU/mIpenicillin, 100 @g/mlstreptomycin, and 0.25 @.tg/mlamphotericin B (RPMIFCS) at 37°Cin a humidifiedatmospherewith 5% CO2.Fifty-pi aliquotsofTNF standard solutions or samples, diluted with RPMI-FCS, were added induplicate to the wells of 96-well flat-bottomed polystyrene cell culture plates,followed by 50-pi aliquots of 1.2 X 106 cell suspension in RPMI-FCScontaining 2 pg/mi actinomycin D and 80 m@ilithium chloride. After 24 hincubation at 37°Cin 5% C02, live cells were stained by adding 20 @.dof a 2.5mg/mI 3(4,5-dimethylthiazolyl-2-yl)2,5-diphenyltetrazoliumbromide (Calbiochem, San Diego, CA) solution and incubating for 4 h at 37°C.Then, a l00-pialiquot of a 33% N,N-dimethylformamide, 200 g/liter SDS solution in water,brought to pH 4.7 with acetic acid, was added to each well and furtherincubated for 24 h to dissolve the dye. The absorbance of each well was thenread by a microplate reader at 570 nm, using a 650 nm filter as reference.Bioactive TNF in the samples was calculated by interpolating the absorbancevalues on a calibration curve. The cytolytic activity of TNF on RMA-Thy 1.1
cells was carried out essentially as described above except that cells werecultivated in RPMI, 5% FCS, 2 m'vi glutamine, 100 IU/ml penicillin, [email protected]/mlstreptomycin, 250 ng/ml amphotericin B, 50 flM2-mercaptoethanol, and500 @g/mlG4l8 and that 30 ng/ml actinomycin D were added to the mediumduring assay incubation and lithium chloride was not.
Three-Step Targeting of RMA Cells with bio-TNF and in Vitro Cyto.
toxicity Assay. Fifty-pA aliquots of a RMA-Thy 1.1 cell suspension concentration (I X l06/ml) in PBS containing 2% FCS (PBS/FCS) were seeded in
round-bottomed microtiter plate wells, mixed with 1 @lof a 0.5 mg/mlbio-l9El2 solution in PBS/FCS, and incubated for 10 mm on ice. The cells
were washed twice by adding 200 s.d/wellof PBS/FCS and centrifuged for 2mmat300Xg at4°C.Thecellsineachwellwereresuspendedbygentlevortexing and subjected to a series of incubation and washings steps, as above,
with 10 i.tg/mi NAV (a deglycosylated avidin with neutral isoelectric point;Pierce) in PBSIFCS for 10 mm, followed by TNF or bio-TNF at variousconcentrations for 1 h. After the final wash, the targeted cells were resus
pendedin RPMI,5% FCS, 2 nmiglutamine,100 IU/ml penicillin, 100 @.tg/mlstreptomycin, 250 ng/ml amphotericin B, 50 flM 2-mercaptoethanol, 500 @tg/ml
G4l8, and 30 ng/ml actinomycin D and incubated for 24 h at 37°Cin ahumidified atmosphere with 5% CO2. Live cells were stained with 3(4,5-dimethylthiazolyl-2-yl)2,5-diphenyltetrazolium bromide as described above.
Cytofluorometric Analysis. The TNF antigen present on the surface ofRMA-Thy 1.1 after three-step targeting with bio-TNF or TNF was measured
by cytofluorometric analysis. Cells were pretreated with bio-l9El2, NAy, andbio-TNF or TNF as described above. After the third step, cells were washedwith PBS-FCS, incubated on ice with rabbit anti-TNF antiserum (1:1000, 10mm), washed again with PBS/FCS, and further incubated with goat antirabbit
a As measured by cytolytic assay using RMA-Thy 1 .1 cells (see “Materials and
Methods―).Bioactive TNF in the samples was calculated by interpolating the absorbancevalues on a calibration curve set up using TNF (l0' units/mg) standard solutions atvarious concentrations.
bThecytolytic activityof 10ng/mlbio-TNFwasmeasuredin thepresenceof 10 @g/m1NAy.
C Measured as described in “Material and Methods.―
d ND, not determined.
FITC (1:1000, 10 mm). After the final washing, the cells were analyzed usinga FACScan (Becton Dickinson, Oxnard, CA).
In Vivo Studies. RMA-Thy 1.1 cells were pretreated in vitro with biol9El2 (10 @.tg/mlfor 10 mm), NAV (10 @g/mlfor 10 mm), and bio-TNF orTNF (l0@units/ml for 1 h) as described above, washed twice with 9 g/litersodium chloride, and resuspended in the same solution. All steps were carried
out on ice, although preliminary experiments showed that the antigen does notinternalize. The cells were then injected s.c. in the left side of C571BL6 mice(Charles River, Milan, Italy; 5 X 10―cells per mouse, 5 mice per group). Cell
viability was >95% as checked by FACS analysis using propidium iodide
staining. The tumor growth was monitored twice weekly by measuring the
longitudinal and lateral diameters using calipers. Control groups includedanimals that were treated with cells in which the bio-19E12 or NAV or bothwere omitted. The same experiment was carried out with nu/nu mice (CharlesRiver).
RESULTS
Recombinant Human TNF Can Be Efficiently Biotinylatedwithout Loss of Biological Activity. Human TNF contains six lysineresidues and one a-amino group (24) that could react with D-biotinyl
6-aminocaproic acid N-hydroxysuccinimide ester. Because biotinylation of some of these residues could be critical for biological activityof TNF, preliminary experiments were carried out to optimize thebiotinylation reaction and preserve the biological activity. As shownin Table I, TNF markedly loses activity when biotinylated at pH 8.8with high biotin!FNF molar ratios. The loss of activity was evengreater when measured in the presence of an excess of NAy. Incontrast, a good recovery of both activity and biotinylation wasobtained at pH 6.8. Electrospray mass spectrometry analysis showedthat this product consists of a mixture of molecules with 0, 1, and 2biotins per subunit.4 The product obtained at pH 6.8 was used for
subsequent studies.Three-Step Targeting of Tumor Cells Increases both the
Amount and the Persistence of Cell Surface-bound Bio-TNF. Tostudy the targeting of bio-TNF onto tumor cells using the avidinpretargeting approach, a mouse model for in vitro and in vivo studies
was developed. RMA-lymphoma cells carrying the Thy 1.2 allelewere transfected with the Thy 1.1 allele (RMA-Thy I .1) to generate aunique tumor-associated antigen. Reverse transcription-PCR analysisshowed that both TNF-R55 and TNF-R75 genes are transcribed inthese cells (data not shown). The TNF targeting onto cell membraneswas carried out by a three-step incubation procedure with the anti-Thy1.1 mAb l9El2-biotin conjugate (bio-l9El2; first step), NAV (sec
4 F. Magni, personal communication.
1923
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from

1-STEP 2-STEP 3-STEP
+mAb bio-19E12 +NAV +bio-TNF
!L cola.
ond step) and bio-TNF (third step), schematically represented in Fig. a significant cell death, whereas no or little effect was observed when1. The effect of cell pretargeting on the binding of bio-TNF was NAV was omitted or when TNF was used in the place of bio-TNF.quantified by cytofluorometric analysis using rabbit anti-TNF poly- This effect was neutralized by an anti-TNF mAb, mAb78 (Fig. 5,clonal antibodies. As shown in Fig. 2, whereas RMA-Thy 1.1 cells inset). Interestingly, although bio-TNF induced cytolysis of pretarbind TNF or bio-TNF to the same extent in the absence of bio-l9El2 geted cells even alter a very short exposure (15 mm), no effectsand in the absence of NAV (Fig. 2, B—Dand F—H,respectively), cells occurred with TNF in the same conditions (Fig. 6). These results,pretargeted with bio-l9El2 and NAV bind 10—20-foldmore bio-TNF(Fig. 2E) than TNF (Fig. 2A). These results indicate that NAV bound
to the cell surface through bio-19E12 acts specifically and efficiently A + bio-19E12 261 E + bio-19E12as an additional receptor for bio-TNF. + NAV + NAV
. . + TNF + bio-TNF
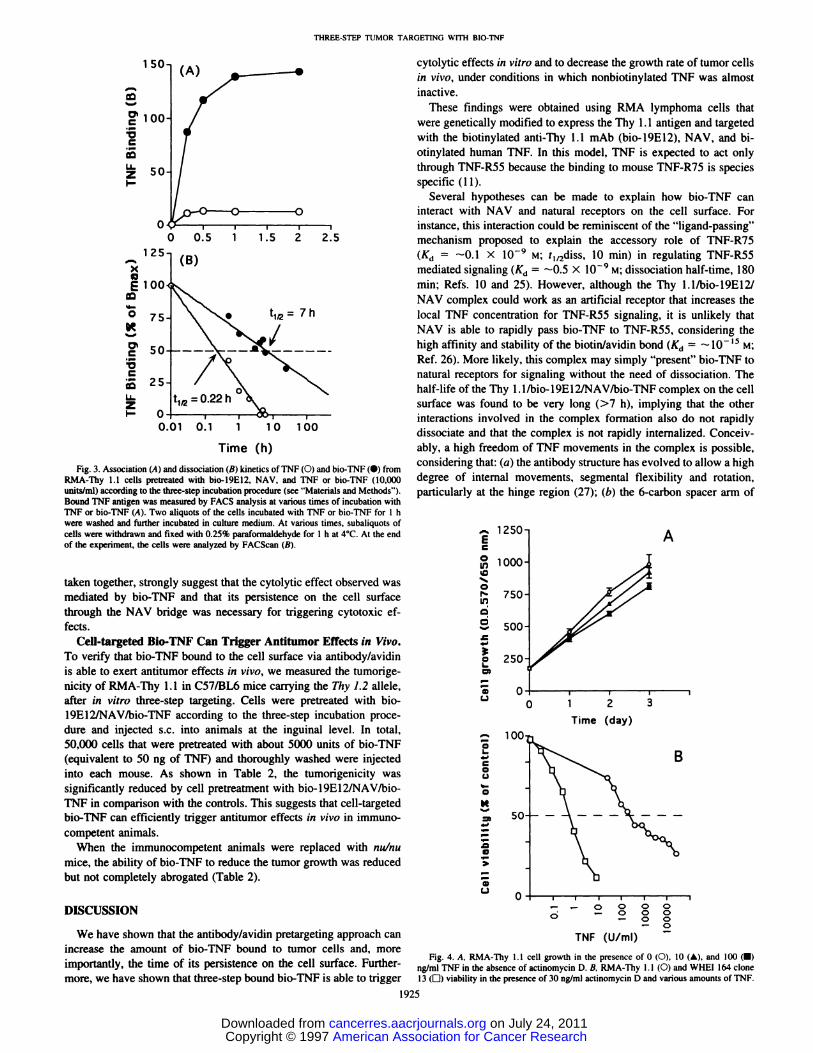
The associatlon and dissoclatlon kinetics of TNF and blo-TNF frompretargeted cells were also evaluated by cytofluorometric analysis.Although both TNF and bio-TNF can bind in less than 1 h (Fig. 3A),their half-lives on the cell surface are different: 0.22 and 7 h, respeclively (Fig. 3B). Thus, whereas TNF bound to natural receptors _________________rapidly disappears, presumably by dissociation and/or internalization, @oou@i jo2 103@bio-TNF bound via the artificial antibody/avidin bridge persists for a + bio.1 9E12@ F + bio-19E12considerably longer time (>30-fold increase). + TNF + bio-TNF
In conclusion, immunofluorescence studies in the RMA-Thy 1.1model indicate that the antibody/avidin pretargeting approach increases the amount of bio-TNF bound to the cell surface, as well as its
persistence.
Cell-targeted Bio-TNF Is Bioactive in a Cytolytic Assay. The ______________effects of soluble TNF on RMA-Thy 1.1 cell were studied first. In -@ @o @o' io@ io3 104the absence of protein synthesis inhibitors, cell growth was onlymoderately inhibited even after incubation for several days at 2relatively high TNF concentrations (Fig. 4A), indicating that RMAThy 1.1 cells are resistant to the cytolytic effect of TNF. Incontrast, in the presence of 30 ng/ml of actinomycin D, a 50%decrease of cell viability was observed after 24 h incubation with
3—5ng/ml of TNF (500 units/ml; Fig. 4B), indicating that RMA
Thy 1.1 cells become sensitive to the cytolytic effects of TNF inthe presence of a transcription inhibitor. This is in keeping with thenotion that TNF rapidly induces protective factors in many celllines and that induction requires protein synthesis (12). In the
presence of actinomycin D, however, RMA-Thy 1.1 cells are stillrelatively resistant to TNF compared with WHEI cells (the reference for TNF cytolytic assays), which are 1000-fold more sensitive(Fig. 4B).
To verify that three-step bound bio-TNF can interact with the “j@@@@@@@@@ i0@natural receptors and trigger cytolytic effects, we measured the cellviability of RMA-Thy 1.1 cells in the presence of actinomycin D after Fluoresce nce Intensity
targeting with bio-TNF. Fig.2.BindingofTNF(A—D)orbio-TNF(E—F)toRMA-Thy1.I cellspretreatedwithAs shown in Fig. 5, the combination of bio-TNF and NAV caused or withoutbio-l9El2 andNAV,as measuredby FACSanalysis.
1924
+ NAV
+TNF+ NAV
+ bio-TNF
+ TNF
THREE-STEP TUMOR TARGETING WITH BIO-TNF
.4. bio-TNF
TNF-Rs
Thy 1.1
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from
0.01 0.1 1 10 100
THREE-STEP TUMOR TARGETING WITH BIO-TNF
cytolytic effects in vitro and to decrease the growth rate of tumor cellsin vivo, under conditions in which nonbiotinylated TNF was almostinactive.
These findings were obtained using RMA lymphoma cells thatwere genetically modified to express the Thy 1.1 antigen and targeted
with the biotinylated anti-Thy 1.1 mAb (bio-l9El2), NAV, and biotinylated human TNF. In this model, TNF is expected to act onlythrough TNF-R55 because the binding to mouse TNF-R75 is speciesspecific (11).
Several hypotheses can be made to explain how bio-TNF caninteract with NAV and natural receptors on the cell surface. Forinstance, this interaction could be reminiscent of the “ligand-passing―mechanism proposed to explain the accessory role of TNF-R75(Kd _ —0.1X i0@ M; t1,@diss,10 mm) in regulating TNF-R55mediated signaling (Kd = —0.5X l0@ M;dissociation half-time, 180mm; Refs. 10 and 25). However, although the Thy 1.l/bio-l9El2/NAV complex could work as an artificial receptor that increases thelocal TNF concentration for TNF-R55 signaling, it is unlikely thatNAV is able to rapidly pass bio-TNF to TNF-R55, considering thehigh affinity and stability of the biotin/avidin bond (Kd = —l0 ‘@Ref. 26). More likely, this complex may simply “present―bio-TNF tonatural receptors for signaling without the need of dissociation. Thehalf-life of the Thy 1.l/bio-l9El2/NAV/bio-TNF complex on the cellsurface was found to be very long (>7 h), implying that the otherinteractions involved in the complex formation also do not rapidlydissociate and that the complex is not rapidly internalized. Conceivably, a high freedom of TNF movements in the complex is possible,considering that: (a) the antibody structure has evolved to allow a highdegree of internal movements, segmental flexibility and rotation,particularly at the hinge region (27); (b) the 6-carbon spacer arm of
1 50
100
50
1@
@iiiii:LI.zI-
xEl
0
C
VC
U-zI-
0 0.5 1 1.5 2 2.5
Time (h)
Fig. 3. Association (A)and dissociation (B) kinetics ofTNF(O) and bio-TNF(•) fromRMA-Thy 1.1 cells pretreated with bio-19E12, NAV, and TNF or bio-TNF (10,000units/mi) according to the three-step incubation procedure (see “Materialsand Methods―).Bound TNF antigen was measured by FACS analysis at various times of incubation withTNF or bio-TNF (A). Two aliquots of the cells incubated with TNF or bio-TNF for 1 hwere washed and further incubated in culture medium. At various times, subaliquots ofcells were withdrawn and fixed with 0.25% paraformaldehyde for 1 h at 4°C.At the endof the experiment, the cells were analyzed by FACScan (B).
taken together, strongly suggest that the cytolytic effect observed wasmediated by bio-TNF and that its persistence on the cell surfacethrough the NAV bridge was necessary for triggering cytotoxic effects.
Cell-targeted Bio-TNF Can Trigger Antitumor Effects in Vivo.To verify that bio-TNF bound to the cell surface via antibody/avidinis able to exert antitumor effects in vivo, we measured the tumorigenicity of RMA-Thy 1.1 in C571BL6 mice carrying the Thy 1.2 allele,after in vitro three-step targeting. Cells were pretreated with bio19E12/NAV/bio-TNF according to the three-step incubation procedure and injected s.c. into animals at the inguinal level. In total,50,000 cells that were pretreated with about 5000 units of bio-TNF(equivalent to 50 ng of TNF) and thoroughly washed were injected
into each mouse. As shown in Table 2, the tumorigemcity wassignificantly reduced by cell pretreatment with bio-19E12/NAV/bioTNF in comparison with the controls. This suggests that cell-targetedbio-TNF can efficiently trigger antitumor effects in vivo in immunocompetent animals.
When the immunocompetent animals were replaced with nu/numice, the ability of bio-TNF to reduce the tumor growth was reducedbut not completely abrogated (Table 2).
DISCUSSION
We have shown that the antibody/avidin pretargeting approach canincrease the amount of bio-TNF bound to tumor cells and, moreimportantly, the time of its persistence on the cell surface. Furthermore, we have shown that three-step bound bio-TNF is able to trigger
A
3Time (day)
,- 0 0 00. _ 0 0 0
0 00
TNF (U/mi)
E
0U,‘C
0NLI,
.@
0I-a,
aC-)
aI.
aaC.)
asQ
a
aC-)
Fig. 4. A, RMA-Thy 1.1 cell growth in the presence of 0 (0), 10 (A), and 100 ()ng/ml TNF in the absence of actinomycin D. B. RMA-Thy 1.1 (0) and WHEI 164 clone13 (0) viability in the presence of 30 ng/ml actinomycin D and various amounts of TNF.
1925
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from

50
THREE-STEP TUMOR TARGETING WITH BIO-TNF
the biotin labels of antibody and TNF molecules could contribute toconfer further points of flexibility and rotation to the complex; (c)TNF biotinylation at pH 6.8 may have favored the reaction with thea-amino group of the NH2-terminal region, known to be located in aflexible segment (7) and not necessary for receptor binding (28); and(d)theThy 1.1antigenis likely to moveandrotatein themembrane.It is thus possible that the high freedom of relative movements andangle formation of the complex allows cell-bound TNF to interactwith membrane receptors without the need to dissociate from it. Athird hypothesis could also be made that takes into account that atleast part of the TNF subunits can slowly dissociate during the 24 hincubation in the cytolytic assay (9, 29, 30) and reassociate intosoluble trimers that, in turn, may freely interact with the receptors. Itis also possible that all of these mechanisms contribute, to a differentextent, to the overall effects observed.
The in vitro cytolysis, observed in the presence of actinomycin D,and the reduced tumorigenicity in mice of RMA-Thy 1.1 cell pretreated with bio-19E12INAV/bio-TNF not only document the functional activity of targeted bio-TNF in interacting with natural receptors but also suggest that local stimulation of the mouse TNF-R55 byTNF is sufficient for triggering antitumorigenic effects in vivo because TNF-R75 is species specific (1 1).
The reduced tumorigenicity of RMA-Thy 1.1 cells could be due toboth direct cytotoxicity and indirect antitumor effects triggered bybio-TNF. Because human TNF is noncytolytic and only moderatelyable to inhibit the growth of RMA cells when tested in vitro in theabsence of actinomycin D, we favor the hypothesis that three-steptargeted bio-TNF acts in vivo through indirect mechanisms. Moreover, the fact that the tumor growth rate was still reduced after 20—30
aI-
aaC.)
aII
a
aC..)
125
75
25100 150
1000
Time (mm)
Fig. 6. RMA-Thy 1.1 cell viability after the three-step targeting with TNF (0) orbio-1'NF (•).Three-step incubations were carried out as described in the legend of Fig.5 except that 10,000 units/mi TNF or bio-TNF were incubated for various times priorcytolytic assay.
days [i.e., when TNF is likely to be absent from most (if not all) tumor
cells] further supports the hypothesis of an indirect effect of bio-TNF.For instance, indirect mechanisms could rely on: (a) TNF-inducedsecretion of cytokine by the tumor cells and/or peritumoral tissue; (b)induction of inflammation and of a nonadaptive immune response,
mainly mediated by phagocytes of myeloid lineage; or (c) formationof a capsule that may reduce tumor growth (31—34).Furthermore, inview of the known effects of TNF on tumor vascular endothelial cells(35, 36), inhibition by TNF of neovascularization of blood vessels ofthe tumor might also contribute to the reduced oncogenicity of RMAcells. Interestingly, using the nude mice model, we found that theability of cell bound bio-TNF to delay tumor growth was reduced in
comparison with the CS7IBL6 mice model but not completely abrogated, supporting the concept that effector cells other than T cellsmight play a role in the observed antitumor effects.
Previous reports showed that some mouse tumors can be cured withTNF only several days after inoculation because TNF exerts antitumoreffects mainly by affecting vascular endothelial cells (12). Althoughour in vivo RMA model was useful to show the potential of targeting
TNF to tumors, further work aimed at developing appropriate modelsbased on established, highly vascularized tumors and at optimizing thetime of injection and the doses of each reagent, alone or combinationwith other cytokines or drugs (e.g., IFN--y or NF-kB inhibitors), isnecessary to further explore the therapeutic potential of this approach.
It has been reported that cross-linking of Thy 1 with a mAb caninduce primary T cells, as well as T cell lymphomas, to secrete
interleukin 2. We think that it is very unlikely that the antitumoractivity observed in our experimental system was due to Thy 1.1
cross-linking by mAb bio-l9E12, for the following reasons: first,control experiments showed that treatments with bio-l9E12 alone orin combination with NAV was not sufficient to delay the in vivo
growth of RMA cells (Table 2); second, in vitro cross-linking withplastic-bound 19E12 does not activate RMA-Thy 1.1—2 to secrete
measurable amounts of interleukin 2.@The finding that a local TNF action can reduce the RMA-Thy 1.1
cell growth in vivo is in agreement with previous findings that expression of the TNF gene in tumor cells correlates with reducedtumorigenicity and invasiveness (37) and strengthens the concept thatstrategies aimed at localizing TNF action might be of therapeuticvalue. The approach described here may have some remarkable advantages over targeting with conventional antibody-cytokine conjugates prepared by chemical synthesis or genetic engineering technol
A) TNF
10000 100000
TNF (U/mi)
800@
700
600
500@
400
300
@uu.
100
, I I. B) bio-TNF
700•
600'
500
400@
300
200.
Fig. 5. RMA-Thy 1.1 cell viability after the three-step targeting with TNF (A) andbio-TNF (B) and incubation for 24 h in the presence of 30 ng/ml actinomycin D.Three-step incubations were carried out on ice with 10 @g/mlbio-l9E12 (10 mm) or with0 (0) or 10 @sg/ml(@) NAV (10 mm), TNF(A) or bio-TNF(B) at various doses (1 h). Cellviability was measured as described in “Materialsand Methods.―inset. cell viability afterthree-step targeting with 30,000 units/mI bio-TNF (+ NAV) in the absence (D) andpresence (@) of 2 p@g/mlof the neutralizing anti-TNF mAb78. 5 M. Moro, unpublished results.
1926
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from
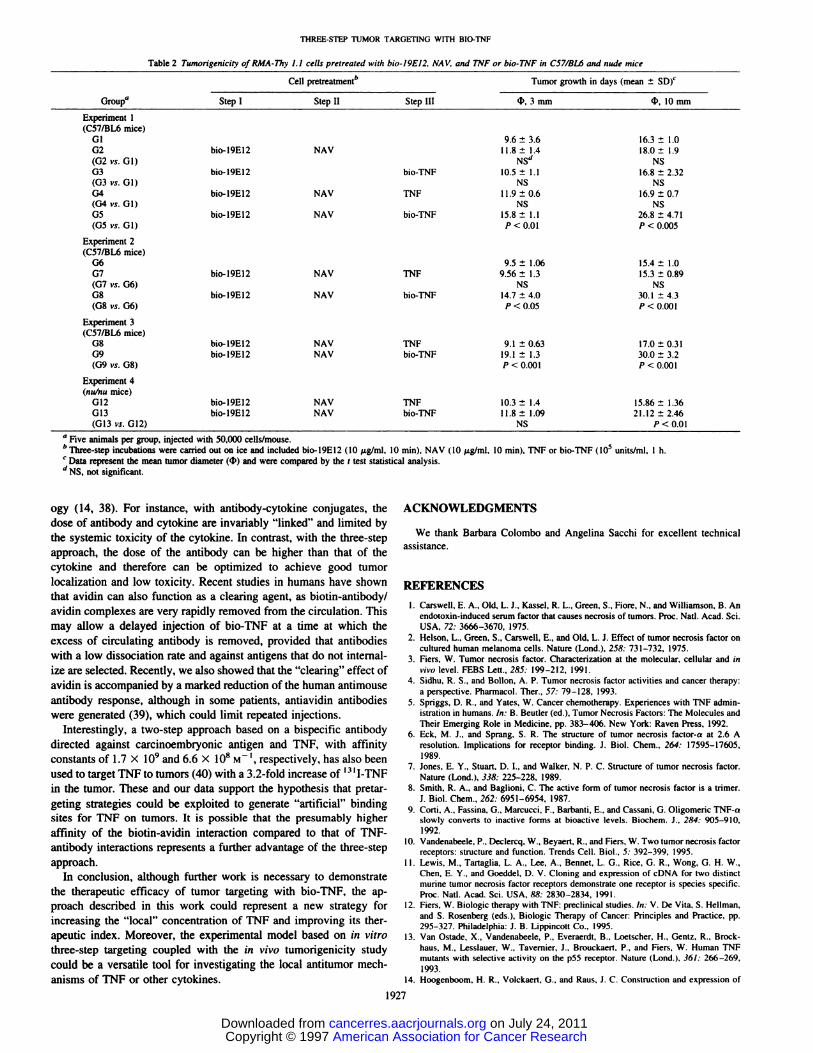
Table 2 Tumorigenicity oIRMA-Thy 1.1 cells pretreated with bio-19E12, NAy, andTNF or bio-TNF in C57/BL6 and nudemiceGroup―Cell
pretreatment―Tumor growth in days (mean ±SD)cStep
IStep IIStep III4'. 3 mm 4, 10mmExperiment
1(C57/BL6mice)019.6
±3.6 16.3 ±1.002bio-19E12NAV11.8±1.4 18.0±1.9(G2vs.Gl)NSd
NS03bio-19E12bio-TNF10.5± 1.1 16.8 ±2.32(03
vs.01)NSNSG4bio-19El2NAVTNF1
1.9 ±0.6 16.9 ±0.7(G4vs.Gl)NSNS05bio-19E12NAVbio-TNF15.8
±1.1 26.8 ±4.71(05vs. 01)P < 0.01 P <0.005Experiment
2(C571BL6mice)069.5±1.0615.4±1.0G7bio-l9E12NAV1'NF9.56
±1.3 15.3 ±0.89(G7vs. 06)NSNS08bio-l9El2NAVbio-TNF14.7
±4.0 30.1 ±4.3(08vs. 06)P < 0.05 P <0.001Experiment
3(C57IBL6mice)08bio-19El2NAVTNF9.1
±0.63 17.0 ±0.31G9bio-l9El2NAVbio-TNF19.1
±1.3 30.0 ±3.2(G9vs. 08)P < 0.001 P <0.001Experiment
4(nu/numice)012bio-l9E12NAVTNF10.3
±1.4 15.86 ±1.36013bio-19El2NAVbio-TNFI1.8 ±1.09 21. 12 ±2.46(013
vs. 012)NS P < 0.01
ThREE-sTEP TUMOR TARGETING WITH BlO-TNF
a Five animals per group, injected with 50,000 cells/mouse.
b@fl..@_@@stepincubations were carried out on ice and included bio-l9El2 (10 gig/mi, 10 mm), NAV (10 @sg/mi, 10 miii), TNF or bio-TNF (l0@ units/mI, I h.
C Data represent the mean tumor diameter (c1@) and were compared by the t test statistical analysis.
d NS, not significant.
ogy (14, 38). For instance, with antibody-cytokine conjugates, thedose of antibody and cytokine are invariably “linked―and limited bythe systemic toxicity of the cytokine. In contrast, with the three-stepapproach, the dose of the antibody can be higher than that of thecytokine and therefore can be optimized to achieve good tumorlocalization and low toxicity. Recent studies in humans have shownthat avidin can also function as a clearing agent, as biotin-antibody/avidin complexes are very rapidly removed from the circulation. Thismay allow a delayed injection of bio-TNF at a time at which theexcess of circulating antibody is removed, provided that antibodieswith a low dissociation rate and against antigens that do not internalize are selected. Recently, we also showed that the “clearing―effect ofavidin is accompanied by a marked reduction of the human antimouseantibody response, although in some patients, antiavidin antibodieswere generated (39), which could limit repeated injections.
Interestingly, a two-step approach based on a bispecific antibodydirected against carcinoembryonic antigen and TNF, with affinityconstants of 1.7 x 10@and 6.6 X 108 M @,respectively, has also beenused to target TNF to tumors (40) with a 3.2-fold increase of ‘31I-TNFin the tumor. These and our data support the hypothesis that pretar
geting strategies could be exploited to generate “artificial―bindingsites for TNF on tumors. It is possible that the presumably higheraffinity of the biotin-avidin interaction compared to that of TNF
antibody interactions represents a further advantage of the three-stepapproach.
In conclusion, although further work is necessary to demonstratethe therapeutic efficacy of tumor targeting with bio-TNF, the approach described in this work could represent a new strategy forincreasing the “local―concentration of TNF and improving its therapeutic index. Moreover, the experimental model based on in vitrothree-step targeting coupled with the in vivo tumorigenicity studycould be a versatile tool for investigating the local antitumor mechanisms of TNF or other cytokines.
ACKNOWLEDGMENTS
We thank Barbara Colombo and Angelina Sacchi for excellent technicalassistance.
REFERENCES
1. Carswell, E. A., Old, L. J., Kassel, R. L., Green, S., Fiore, N., and willi@son. B. Anendotoxin-induced serum factor that causes necrosis of tumors. Proc. NatI. Acad. Sci.USA, 72: 3666—3670, 1975.
2. Helson, L., Green, S., Carswell, E., and Old, L. J. Effect of tumor necrosis factor oncultured human melanoma cells. Nature (Land.), 258: 731—732,1975.
3. Fiers, W. Tumor necrosis factor. Characterization at the molecular, cellular and invivo level. FEBS Left., 285: 199—212,1991.
4. Sidhu, R. S., and Bollon, A. P. Tumor necrosis factor activities and cancer therapy:a perspective. Pharmacol. Ther., 57: 79—128, 1993.
5. Spriggs, D. R., and Yates, W. Cancer chemotherapy. Experiences with TNF administration in humans. in: B. Beutler (ed), Tumor Necrosis Factors: The Molecules and
Their Emerging Role in Medicine, pp. 383—406.New York: Raven Press, 1992.6. Eck, M. J., and Sprang, S. R. The structure of tumor necrosis factor-a at 2.6 A
resolution. Implications for receptor binding. J. Biol. Chem., 264: 17595—17605,1989.
7. Jones, E. Y., Stuart, D. I., and Walker, N. P. C. Structure of tumor necrosis factor.Nature (Land.). 338: 225—228,1989.
8. Smith, R. A., and Baglioni, C. The active form of tumor necrosis factor is a trimer.J.Biol.Chem.,262:6951—6954,1987.
9. Corti, A., Fassina. G., Marcucci, F., Barbanti, E., and Cassani, G. Oligomeric TNF-aslowly converts to inactive forms at bioactive levels. Biochem. J., 284: 905—910,1992.
10. Vandenabeele, P., Declercq, W., Beyaert, R., and Fiers, W. Two tumor necrosis factorreceptors: structure and function. Trends Cell. Biol., 5: 392—399,1995.
11. Lewis, M., Tartaglia, L. A., Lee, A., Bennet, L. G., Rice, G. R., Wong, G. H. W.,Chen, E. Y., and Goeddel, D. V. Cloning and expression of cDNA for two distinctmurine tumor necrosis factor receptors demonstrate one receptor is species specific.Proc. NatI. Aced. Sd. USA, 88: 2830—2834, 1991.
12. Fiers, W. Biologic therapy with TNF: preclinical studies. in: V. De Vita, S. Hellman,and S. Rosenberg (eds.), Biologic Therapy of Cancer: Principles and Practice, pp.295—327.Philadelphia: J. B. Lippincott Co., 1995.
13. Van Ostade, X., Vandenabeele, P., Everaerdt, B., Loetscher, H., Gentz, R., Brockhaus, M., Lesslauer, W., Tavernier, J.. Brouckaert, P., and Fiers, W. Human TNFmutants with selective activity on the p55 receptor. Nature (Lond.), 36/: 266—269,1993.
14. Hoogenboom. H. R., Volckaert, G., and Raus, J. C. Construction and expression of
1927
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from

THREE.STEP TUMOR TARGETING @1THBIO-TNF
antibody-tumor necrosis factor fusion proteins. Mol. Immunol., 28: 1027—1037,1991.15. Hwu, P., and Rosenberg, S. A. The genetic modification of T cells for cancer therapy:
an overview of laboratory and clinical trials. Cancer Detect. Prey., 18: 43—50,1994.16. Lejeune, F., Lienard, D., Eggermont, A., Schraffordt Koops, H., Kroon, B., Gerain,
J., Rosenkaimer, F., and Schmitz, P. Clinical experience with high-dose tumor
necrosis factor a in regional therapy of advanced melanoma. Circ. Shock, 43:191—197,1994.
17. Lienard, D., Eggermont, A. M., Schraffordt Koops, H., Kroon, B. B., Rosenkaimer,F., Autier, P., and Lejeune, F. J. Isolated perfusion of the limb with high-dose tumournecrosis factor-a (TNF-a), interferon-gamma (IFN-gamma) and melphalan for melanoma stage III. Results of a multi-centre pilot study. Melanoma Res., 4: 21—26,1994.
18. Paganelli, G., Magnani, P., Siccardi, A., and Fazio, F. Clinical application of theavidin-biotin system for tumor targeting. in: D. Goldenberg (ed), Cancer Therapywith Radiolabeled Antibodies, pp. 239—253.Boca Raton, FL: CRC Press, 1995.
19. Paganelli, G., Grana, C., Chinol, M., De Cicco, C., Cremonesi, M., Tarditi, L.,Franceschini, R., Zoboli, S., Dc Braud, F., and Siccardi, A. 0. Antibody guided
radiotherapy in malignant glioma patients with Y-90-biotin using a 3-step pretargeting approach. Advances in the application of monoclonal antibodies in clinicaloncology. The Thirteenth International Hammersmith Conference, 27th-30th May1996, p. 36. Chania, Greece, 1996.
20. Paganelli, G., Magnani, P., Zito, F., Villa, E., Sudati, F., Lopalco, L., Rossetti, C.,Malcovati, M., Chiolerio, F., Seccamani, E., Siccardi, A. G., and Fazio, F. Three-stepmonoclonal antibody tumor targeting in carcinoembryonic antigen-positive patients.Cancer Res., 51: 5960—5966, 1991.
21. Modorati, 0., Brancato, R., Paganelli, 0., Magnani, P., Pavoni, R., and Fazio, F.Immunoscintigraphy with three step monoclonal pretargeting technique in diagnosisof uveal melanoma: preliminary results. Br. J. Ophthalmol., 78: 19—23,1994.
22. Colombo, P., Paganelli, 0., Magnani, P., Songini, C., Fazio, F., and Faglia, G.lmmunoscintigraphy with anti-chromogranin A antibodies in patients with endocrine/neuroendocrine tumors. J. Endocr. Invest., 16: 841—843, 1993.
23. Espevik, T., and Nissen-Meyer, J. A highly sensitive cell line, WEHI 164 clone 13,for measuring cytotoxic factor/tumor necrosis factor from human monocytes. J. Immunol. Methods, 95: 99—105,1986.
24. Pennica, D., Nedwin, G. E., Hayflick, J. S., Seeburg, P. H., Derynck, R., Palladino,M. A., Kohr, W. J., Aggarwal, B. B., and Goeddel, D. V. Human tumor necrosisfactor: precursor, structure, expression and homology to lymphotoxin. Nature(Lond.), 321: 724—729,1984.
25. Tartaglia, L. A., Pennica, D., and Goeddel, D. V. Ligand passing: the 75 kDa tumornecrosis factor (TNF) receptor recruits TNF for signaling by the 55 kDa TNFreceptor. J. Biol. Chem., 268: 18542—18548, 1993.
26. Green, N. Avidin and streptavidin. Methods Enzymol., 184: 51—67,1990.27. Nezlin, R. Intemal movements in immunoglobulin molecules. Adv. Immunol., 48:
1—39,1990.28. Lin, L. S. The design and activities of TNF muteins. in: B. Beutler (ed), Tumor
Necrosis Factors: The Molecules and Their Emerging Role in Medicine, pp. 33—48.New York: Raven Press, 1992.
29. Poiesi, C., Albertini, A., Ghielmi, S., Cassani, G., and Corti, A. Kinetic analysis ofTNF-a oligomer-monomer transition by surface plasmon resonance and immunochemical methods. Cytokine, 5: 539—545, 1993.
30. Alzani, R., Cozzi, E., Corti, A., Temponi, M., Trizio, D., Gigli, M., and Rizzo, V.Mechanism of suramin-induced deoligomerization of tumor necrosis factor a. Biochemistry, 34: 6344—6350, 1995.
31. Talmadge, J. E., Tribble, H. R., Penmngton, R. W., Phillips, H., and Wiltrout, R. H.Immunomodulatory and immunotherapeutic properties of recombinant ‘y-interferonand recombinant tumor necrosis factor in mice. Cancer Res., 47: 2563—2570,1987.
32. Beyaert, R., De Potter, C., Vanhaesebroeck, B., Van Roy, F., and Fiers, W. Inductionof inflammatory cell infiltration and necrosis in normal mouse skin by the combinedtreatment of tumor necrosis factor and lithium chloride. Am. J. Pathol., 138: 727—739,1991.
33. Palladino, M. A., Jr., Shalaby, M. R., Kramer, S. M., Ferraiolo, B. L., Baughman,R. A., Delco, A. B., Crase, D., Marafino, B., Aggarwal, B. B., Figari, I. S., Liggitt,D., and Patton, J. S. Characterization of the antitumor activities of human tumornecrosis factor-a and the comparison with other cytokines: induction of tumorspecific immunity. J. Immunol., 138: 4023—4032,1987.
34. Havell, E. A., Fiers, W., and North, R. J. The antitumor function of tumor necrosisfactor (TNF). I. Therapeutic action of TNF against an established murine sarcoma isindirect, immunologically dependent, and limited by severe toxicity. J. Exp. Med.,167: 1067—1085,1988.
35. Sato, N., Goto, T., Haranaka, K., Satomi, N., Nasiuchi, H., Mano-Hirano, Y., andSawasaki, Y. Actions of tumor necrosis factor on cultured vascular endothelial cells:morphologic modulation, growth inhibition, and cytotoxicity. J. NatI. Cancer Inst.,76:1113—1121,1986.
36. Watanabe, N., Niitsu, Y., Umeno, H., Kuriyama, H., Neda, H., Yamauchi, N., Maeda,M., and Urushizaki, I. Toxic effect of tumor necrosis factor on tumor vasculature inmice. Cancer Res., 48: 2179—2183, 1988.
37. Vanhaesebroeck, B., Mareel, M., Van Roy, F., Grooten, J., and Fiers, W. Expressionof the tumor necrosis factor gene in tumor cells correlates with reduced tumorigenicity and reduced invasiveness in vivo. Cancer Res., 5!: 2229—2238,1991.
38. Yang, J., Raju, R., Sharma, R., and Xiang, J. A genetically engineered fusion proteinM4ITNF with increased bifunctional activity refolded in the presence of proteindisulfide isomerase. Hum. Antib. Hybrid., 6: 129—136,1995.
39. Paganelli, G., Sidoli, A., Chinol, M., Corti, A., Maggiolo, M., Baroni, S., andSiccardi, A. G. Three-step pretargeting approach reduces the human anti-mouseantibody response in patients submitted to radioimmunoscintigraphy and radioimmunotherapy. Eur. J. Nucl. Med., in press, 1997.
40. Robert, B., Mach, J. P., Mani, J. C., Ychou, M., Folli, S., Artus, J. C., and Pelegrin,A. Cytokine targeting in tumors using a bispecific antibody directed against carci
noembryonic antigen and tumor necrosis factor a. Cancer Res., 56: 4758—4765,1996.
1928
American Association for Cancer Research Copyright © 1997 on July 24, 2011cancerres.aacrjournals.orgDownloaded from





























