Embed Size (px)
Citation preview

Current Cancer Therapy Reviews, 2006, 2, 73-79 73
1573-3947/06 $50.00+.00 © 2006 Bentham Science Publishers Ltd.
Tumor Control by Manipulation of the Human Anti-Apoptotic SurvivinGene
Zakir Khan, Pratiksha Bhadouria, Radha Gupta and Prakash S. Bisen*,†
Department of Biotechnology, J.C. Bose Institute of Life Sciences, Bundelkhand University, Jhansi, U.P. India
Abstract: Survivin is a relatively unique member of the inhibitor of apoptosis protein (IAP) family. It contains a single
baculovirus IAP repeat (BIR) domain. It is involved in the control of cell cycle and inhibition of apoptosis. Survivin is of
interest because it is specifically up-regulated in cancer cells and completely down-regulated and undetectable in normal
adult tissues. Thus, survivin has proved to be a promising therapeutic target for normal anti-cancer therapy. Survivin
protects the fast dividing tumor cells against default apoptosis to facilitate aberrant mitosis. Down-regulation of survivin
with multiple approaches, suppress tumor progression and induce apoptosis on its own or in combination with
chemotherapy and radiotherapy.
Keywords: Survivin, IAPs, Apoptosis, Caspases, Ribozyme, p53.
INTRODUCTION
Survivin, a member of inhibitor of apoptosis protein(IAP) family, is characterized by a unique structure thatdiscriminates it from other members of the IAP family [1,2].It contains only a single BIR repeat and lacks a carboxyterminal Ring finger domain [3-5]. Survivin is a criticalregulator of multiple processes, including proliferation andapoptosis and its expression appears to be a consistentfeature of hyperproliferative lesions contributing to theformation of hyperplasia [6,7]. It is expressed in G2/M phaseof cell cycle to support rapidly dividing cell machinery [1,3]and helps in proper segregation of chromosomes during celldivision [6,7]. Survivin is abundantly expressed inembryonic tissues and in a wide range of cancer tissues butundetectable in normal differentiated tissues [3].Overexpression of survivin in cancer may overcome cellcycle checkpoints to facilitate aberrant progression oftransformed cells through mitosis [1,2]. To make an idealdrug target, an apoptosis regulator should be preferentiallyexpressed in tumor cells but not in normal tissues, andinterference with its expression/ function should be sufficientto facilitate cell death, either alone, or in combination withchemotherapeutic drugs or ultraviolet/ -irradiation. It hasbeen observed that the human survivin gene may fulfill bothof these prerequisites [8].
Several chemotherapeutic agents kill tumor cells throughapoptosis [8]. It has been found that survivin expression ishigh when tumor cells are treated with anticancer agents[8,9] by counteracting the effect of chemotherapeutic agentsand provides resistance to apoptosis [8,9]. Down regulationof survivin sensitize tumor cells to apoptosis induced bychemotherapy and radiotherapy in many types of cancer[10]. Exploitation of survivin signaling pathway may offernew therapeutic alternatives for cancer treatment.
*Address correspondence to this author at the J.C. Bose Institute of Life
Sciences, Bundelkhand University, Jhansi-284128, U.P., India; E-mail:
†Present Address: Jaipur National University, Jaipur, India
Differential expression of survivin in cancer, comparedto most normal tissues, makes survivin a candidate for amolecular marker of cancer [1,3]. There is good evidencethat survivin may provide a quick prognostic indicator foridentifying patients at risk of recurrent disease [11]. Survivinand/or its auto-antibody present in biological fluids of cancerpatients could provide them a potential diagnostic tool [12].
STRUCTURE AND DISTRIBUTION OF SURVIVIN
Survivin is a 16.3 kD protein consisting of 142 aminoacids. The gene encoding survivin is of 14.5 kb, which islocated at the telomeric region of the chromosome 17 [11]. Ithas four exons and three introns (Fig. 1). Survivin is thesmallest member of the IAP family having two defineddomains including N-terminal Zn
+2 –binding domain linked
to 65 Ao amphipathic C-terminal alpha-helix [12,13]. It is
homodimeric, arranged through hydrophobic surface of theBIR domain of each survivin monomer [4,5]. Several splicevariants of survivin have been identified, such as survivin-delta Ex3 lacking exon 3 and survivin-2B retaining a part ofintron 2 as a cryptic exon (Fig. 1). A complex regulatorybalance exists amongst the isoforms of survivin that maydetermine the response to pro-apoptotic stimuli in tissues andseveral types of cancer [14]. Survivin-deltaEx3 has a uniquecarboxyl terminus sequence containing nuclear localizationsignal, which is found exclusively in the survivin-deltaEx3for an implicated sub-cellular targeting includingmitochondria and nucleus. Both survivin and survivin-deltaEx3 carry anti-apoptotic properties, whereas, survivin-2B is deficient in anti-apoptotic properties which may enablesurvivin-2B as a naturally occurring antagonist of anti-apoptotic survivin variants [15,16].
In mitosis, survivin has been shown to localize to variouscomponents of the mitotic apparatus, such as centrosomesand possibly microtubules. The location of survivin isabnormal in tumor cells as it is present throughout thecytoplasm [13,16].
FUNCTIONS OF SURVIVIN
There are two major types of pathway, extrinsic andintrinsic, that cause programmed cell death. An extrinsic
Not For Distribtion

74 Current Cancer Therapy Reviews, 2006, Vol. 2, No. 1 Bisen et al.
pathway is initiated by ligation of death receptors on the cellsurface (CD95/ Fas receptor, TNF-alpha) and acts throughthe activation of initiator caspase 8. An intrinsic pathway istriggered by multiple death signal either intracellular (un-repaired DNA damage) or environmental culminating toderegulation of the mitochondrial function. As a result, thepermeability of the outer mitochondrial membrane isincreased leading to release of proteins, cyt-c and Smac/DIABLO [2,15,16]. These proteins activate initiator caspase-9 that mediates apoptosome formation [2].
Molecular mechanism(s) of survivin action are not fullyelucidated and are at least in some aspects, controversial.Nevertheless, it is well accepted that survivin is an inhibitorof apoptosis and interferes with cell-cycle progression andmicrotubule stability. In general, mammalian IAPs, blockapoptosis by direct or indirect inhibition of initiator caspase-9 or terminal effector caspases 3 and 7 (Figs. 1 and 2).Studies suggested that survivin inhibits the intrinsic pathwayof apoptosis by interacting with post-mitochondrial events[17-19]. Several hypotheses exist to explain the mechanismsof anti-apoptotic activity of survivin. They might directlybind and inhibit caspases like any another IAPs, such as,XIAP [17]. Smac/DIABLO may act as a pro-apoptoticprotein through its participation in the activation of caspase-9 (Apoptosome formation). It may inhibit apoptosis throughantagonizing the pro-apoptotic ability due to its affinity withSmac/DIABLO [19] (Figs. 2 and 3).
In addition to anti-apoptotic function, survivin alsoregulates cell division because of its presence on the mitoticmachinery of dividing cells. Targeted survivin has resultedin aberrant mitotic progression, leading to failed cytokinesisand multinucleation [6,18,20-22]. Survivin is indispensableduring embryonic development. The homozygous deletion inmice leads to inevitable lethality at day 4-5 due to defects inmitotic spindles formation [13]. The apparent requirement ofsurvivin in normal cell division suggests that overexpressionof survivin in tumors could perturb normal cell cycle control.
FACTORS INDUCING SURVIVIN EXPRESSION
The expression of survivin is undetectable in normaldifferentiated cells but slowly expressed in fast dividing
normal cells, such as, CD34+ bone marrow derived stemcells, basal epithelial cells thymocytes and basal epithelialcells of normal uterine cervix [18,23,24]. Survivinexpression is very high in most cancers, particularly colon,lung, breast, brain and melanoma [12]. The molecularmechanisms of survivin overexpression in cancer seem to becomplex and are only partially understood. It is very likelythat multiple pathways are involved in the reactivation of thesurvivin gene.
Several molecular mechanisms in survivinoverexpression in tumor cells have been elucidated. In thenormal ovaries, survivin exon 1 is silenced by methylationbut it becomes demethylated and transcriptionally active inovarian cancer [12]. In neuroblastoma, amplification of17q25, comprising survivin locus has been reported [23,24].The transcriptional factor such as, p53 has been reported toregulate survivin-expression in various human cancer celllines [25,26]. In many cancers such as, gastric, pancreatic,prostate, lung, and epidermoid carcinomas, a correlationbetween p53 accumulation and survivin expression has beendemonstrated [27,28].
DOWN REGULATION OF SURVIVIN
Anti-Sense Technology
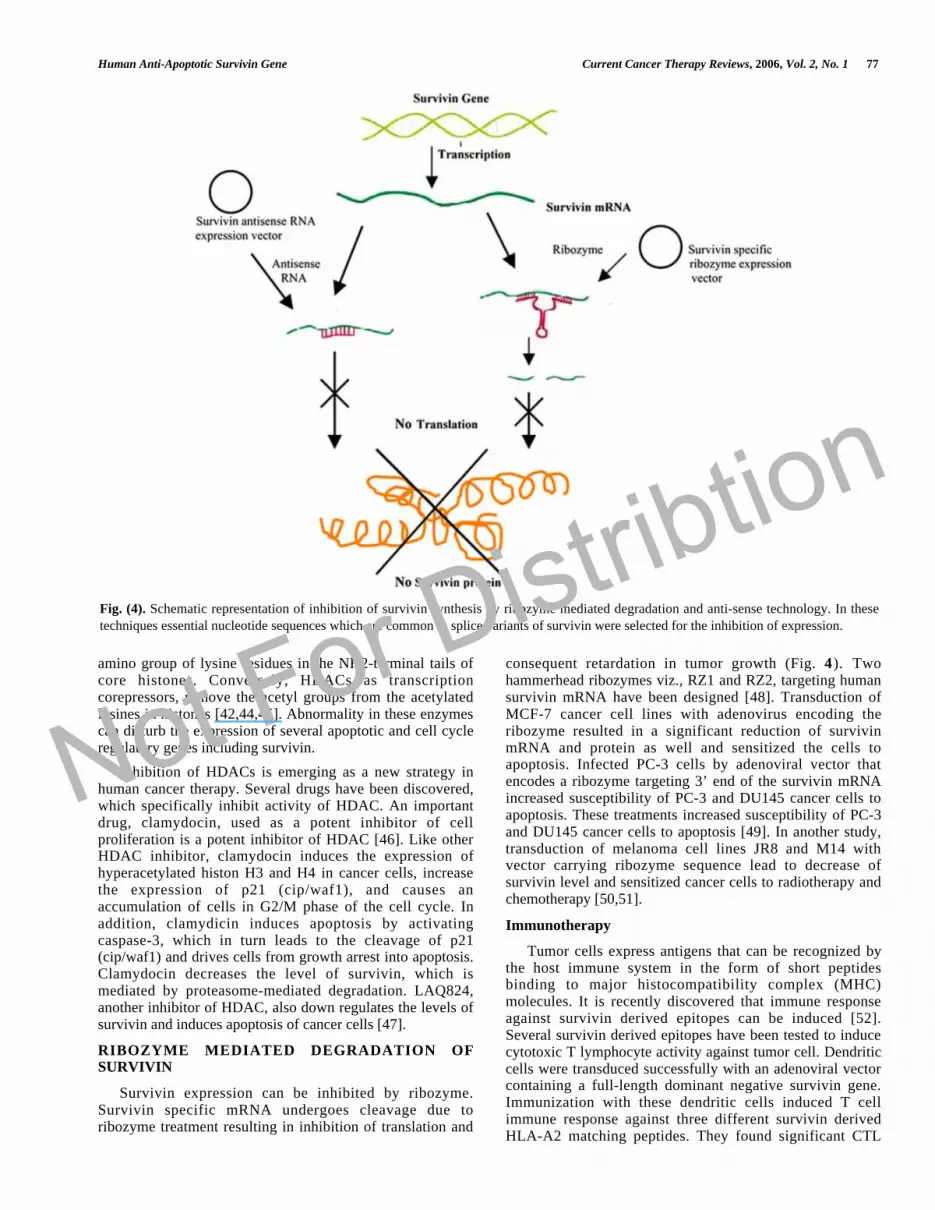
Anti-sense technology, with its potential to selectivelycontrol gene expression and cellular phenotype, is provinguseful as a therapeutic application. Several types of anti-sense approaches (viz. antisense oligonucleotides, antisenseRNA, and small interfering RNA) can be used to inhibitexpression of a target genes (Fig. 4).
Down regulation of survivin by a targeted anti-senseoligonucleotide appears to be an effective gene therapy tocombat several cancers (Fig. 4). Transfection of cancer cellwith antisense of survivin enhances sensitivity of tumor cellsto chemotherapy and radiotherapy. Several anti-survivinoligonucleotides have been tested for their ability to blocksurvivin expression in tumor cell lines. Anti senseoligonucleotide 4003 was found to be the most effective ingrowth inhibition and apoptosis in lung carcinoma cell lines[29].
Fig. (1). Survivin and its splice variants, Survivin–2B, Survivin EX 3. Pre-mRNA of survivin has four exons and an extra exon 2B (A).
Survivin-2B m-RNA has four exons and an extra exon-2B (B). Survivin mRNA has all four exons, but not exon-2B (C). Survivin EX 3
mRNA has three exons (exon 1, 2, 4) but not exon 3 and extra exon-2B (D).
Not For Distribtion

Human Anti-Apoptotic Survivin Gene Current Cancer Therapy Reviews, 2006, Vol. 2, No. 1 75
Most of the mesothelioma cancer cells underwentapoptosis and were more sensitive to chemotherapy andradiotherapy when treated with 20-mer phosphorothioateanti-sense oligonucleotide targeting nucleotides 232-251 ofsurvivin mRNA [30]. Growth of lymphoma cell lines wassignificantly inhibited by using anti-sense oligonucleotide(ASO) [31]. Treatment of colon cancer by adenoviral anti-sense vectors (pAd-CMV-SAS), resulted in an increase ofthe Go/G1 phase population in the cell cycle and increasedtheir sensitivity to chemotherapeutic drugs in vitro, [32]. In
PC-3 prostate cancer cell treatment with anti-sense survivincDNA caused nuclear fragmentation, hypodiploidy, cleavageof a 32- kDa proform caspase-3 to active caspase-3 andproteolysis of the caspase substrate poly (ADP) ribosepolymerase [33]. In another study, similar observation hasbeen seen with human neuroblastoma cell line SK-N-MC[34].
Expression of survivin gene can be checked byinterference RNA including both interfering RNA (siRNA)and short hairpin RNA (shRNA). These interfering RNA can
Fig. (2). Apoptotic Pathways and the site of surviving antiapoptotic action.
Intrinsic and extrinsic apoptotic pathways are shown that coverage into a common downstream pathway of effecter caspase activation. In
intrinsic pathway Cyt-c released from mitochondria in response to several stresses. Cyt-c interacts with Apaf-1 and procaspase-9 in presence
of dATP to form apoptosome complex that leads to activation of procaspase-9. Active caspase-9 activates effector caspases-3, 7, and 6,
which in turn induce apoptosis. Survivin most probably blocks, directly and/or indirectly, caspase 9 activation. It may directly inhibit initiator
caspase-9 and effector caspases-3, 7. Smac/DIABLO is a proapoptotic protein that inhibits activity of IAPs (XIAP). Survivin antagonize the
activity of Smac/DIABLO and may helps in the action of another IAPs (XIAP).
Not For Distribtion

76 Current Cancer Therapy Reviews, 2006, Vol. 2, No. 1 Bisen et al.
be chemically synthesized and transfected into cells ordirectly expressed intracellularly from a plasmid DNA orfrom an adenoviral vector [35]. Transfection of cancer linesby adenoviral vector harboring a tandem-type Si-RNAexpression unit targeting survivin resulted in geneknockdown and induced apoptosis [36,37]. These cancercells, once infected with Adv-siSurv, displayed remarkablyattenuated growth potential, both in vitro and in vivo.Moreover, intratumoral injection of Adv-siSurv significantlysuppressed tumor growth in a genograft model using U251glioma cells. A short hairpin RNA (sh- RNA), containingtwo 20 to 21 bp reverse repeat motifs of survivin targetsequence with 4 to 8 bp spacer, effectively down regulateexpression survivin in liver cancer cell lines Hep G2andSMMC-7721, after transfection [38]. These findings suggestthat targeting of survivin using antisense technology mayhave a potential role in the selective therapy of cancer.
Dominant Negative Construct
In this technology an essential amino acid of the proteinis replaced by another amino acid, which leads to the loss offunction. But this nonfunctional protein has same target asnormal protein. Dominant negative mutant compete withnormal protein for their target and suppress the function ofnormal protein. Several dominant negative constructs havebeen discovered for survivin, from which T34A mutant ofsurvivin is best known [10]. Transduction of breast, cervical,prostate, lung and colorectal cancer cell lines with pAd-T34A (replication deficient adenovirus vector encoding a
nonphosphorylable The34Ala mutant survivin) increasedcyt-c release from mitochondria, processing of caspase-3 tothe active subunits of approximately 17 and 19 kDa,increased caspase-3 catalytic activity, and facilitated tumorcell apoptosis induced by taxol and adriamycin anticancerdrugs [39]. In malignant HeLa cells, trasfection with survivinmutant (survivin-N and survivin T34A) could partiallyreverse the malignancy of HeLa cells [40].
When dominant negative mutant of survivin, replacingthe cysteine residue at amino acid 84 with alanine(Cys84Ala) was introduced into BCG-823 and MKN-45gastric cell lines, the trasfectants exhibited abnormalmorphology, with decreased cell growth and increased rateof spontaneous apoptosis [41]. In PC-3 prostate cancer cellline, transfection with C84A mutant was sufficient tovisualize all biochemical hallmarks of apoptosis includinghypoploid DNA content, caspase 3 activities, and cleavageof caspase substrates [33].
Acetylation and Deacetylation Pathway
In eukaryotic cells, histone acetylation/deacetylation isimportant in transcriptional regulation [42]. Histone acetyltransferases (HATs) are recruited by transcription factors
and
are associated with activation of transcription, whereashistone deacetylases (HDACs) are involved in transcriptionalsilencing. The histone
acetylation is tightly controlled by the
dynamic equilibrium between competing HATs and HDACs
[42,43]. Histone acetyltransferases, as transcriptioncoactivators, catalyze the addition of acetyl groups on the e-
Fig. (3). Models of Survivin in the inhibition of apoptosis. Survivin can directly interact with caspases and inhibit them to suppress
apoptosis (A,B). XIAP is a strong inhibitor of apoptosis, which interacts directly with caspases and inhibits them. Smac/DIABLO is the
negative regulator of XIAP. Survivin may interact with Smac/DIABLO that ultimately leads to inhibition of apoptosis (C).
Not For Distribtion
Human Anti-Apoptotic Survivin Gene Current Cancer Therapy Reviews, 2006, Vol. 2, No. 1 77
amino group of lysine residues in the NH2-terminal tails ofcore histones. Conversely, HDACs as transcriptioncorepressors, remove the acetyl groups from the acetylatedlysines in histones [42,44,45]. Abnormality in these enzymescan disturb the expression of several apoptotic and cell cycleregulatory genes including survivin.
Inhibition of HDACs is emerging as a new strategy inhuman cancer therapy. Several drugs have been discovered,which specifically inhibit activity of HDAC. An importantdrug, clamydocin, used as a potent inhibitor of cellproliferation is a potent inhibitor of HDAC [46]. Like otherHDAC inhibitor, clamydocin induces the expression ofhyperacetylated histon H3 and H4 in cancer cells, increasethe expression of p21 (cip/waf1), and causes anaccumulation of cells in G2/M phase of the cell cycle. Inaddition, clamydicin induces apoptosis by activatingcaspase-3, which in turn leads to the cleavage of p21(cip/waf1) and drives cells from growth arrest into apoptosis.Clamydocin decreases the level of survivin, which ismediated by proteasome-mediated degradation. LAQ824,another inhibitor of HDAC, also down regulates the levels ofsurvivin and induces apoptosis of cancer cells [47].
RIBOZYME MEDIATED DEGRADATION OFSURVIVIN
Survivin expression can be inhibited by ribozyme.Survivin specific mRNA undergoes cleavage due toribozyme treatment resulting in inhibition of translation and
consequent retardation in tumor growth (Fig. 4). Twohammerhead ribozymes viz., RZ1 and RZ2, targeting humansurvivin mRNA have been designed [48]. Transduction ofMCF-7 cancer cell lines with adenovirus encoding theribozyme resulted in a significant reduction of survivinmRNA and protein as well and sensitized the cells toapoptosis. Infected PC-3 cells by adenoviral vector thatencodes a ribozyme targeting 3’ end of the survivin mRNAincreased susceptibility of PC-3 and DU145 cancer cells toapoptosis. These treatments increased susceptibility of PC-3and DU145 cancer cells to apoptosis [49]. In another study,transduction of melanoma cell lines JR8 and M14 withvector carrying ribozyme sequence lead to decrease ofsurvivin level and sensitized cancer cells to radiotherapy andchemotherapy [50,51].
Immunotherapy
Tumor cells express antigens that can be recognized bythe host immune system in the form of short peptidesbinding to major histocompatibility complex (MHC)molecules. It is recently discovered that immune responseagainst survivin derived epitopes can be induced [52].Several survivin derived epitopes have been tested to inducecytotoxic T lymphocyte activity against tumor cell. Dendriticcells were transduced successfully with an adenoviral vectorcontaining a full-length dominant negative survivin gene.Immunization with these dendritic cells induced T cellimmune response against three different survivin derivedHLA-A2 matching peptides. They found significant CTL
Fig. (4). Schematic representation of inhibition of survivin synthesis by ribozyme mediated degradation and anti-sense technology. In these
techniques essential nucleotide sequences which are common in splice variants of survivin were selected for the inhibition of expression.
Not For Distribtion

78 Current Cancer Therapy Reviews, 2006, Vol. 2, No. 1 Bisen et al.
activity against HLA-A2-positive MCF-7 tumor cells thatexpress survivin [53].
Recently, an HLA-24-restricted immunogenic peptide-based vaccination (survivin-2B 80-88, AYACNTSTL) toneutralize survivin-2B splice variant, which is abnormallyexpressed in various types of tumor tissues and tumor celllines has been subjected to phase I trial on patients withadvanced or recurrent colorectal cancer [54]. Immunotherapyof cancer based on survivin may give promising hope.
CONCLUSION
Survivin belongs to an IAP family of proteins that playsan important role in cell cycle progression and cancer cellviability. It is one of the proteins that is specially expressedin most cancer cells. It may, therefore, be a good marker forcancer detection and may serve as an attractive therapeutictarget. Survivin is expressed in the majority of variouscancers studied. Various strategies to target survivin incancer cells are currently under investigation with promisingresults in vivo and in vitro models both. Moreover, somecurrently explored anti-cancer agents might mediate theiranti-cancer effects by inhibiting the survivin pathway.
REFERENCES
[1] Li FZ, Ambrosin G, Chu EY, et al. Control of apoptosis and
mitotic spindle checkpoint by survivin. Nature 1998; 396: 580-3.[2] Deveraux QL, Reed JC. IAP family proteins-suppressor of
apoptosis. Genes Dev 1999; 13: 1253-62.[3] Ambrosini G, Adida C, Altieri DC. A noble anti-apoptotic gene,
survivin, is expressed in cancer and lymphoma. Nature Med 1997;3: 917-21.
[4] Chantalat L, Scoufias DA, Kleman JP, Jung B, Dideberg O,Margolis RL. Crystal structure of survivin reveals a bow tie-shaped
dimmer with two unusual alpha helical extentions. Mol Cell 2000;6: 183-9.
[5] Verdicia MA, Huang H, Dutil E, Kaiser DA, Hunter T, Noel JP.Structure of human anti-apoptotic protein survivin reveals a
dimeric arrangement. Nat Struct Biol 2000; 7: 602-8.[6] Kaitan S, Mendosa M, Jantch-Plunger V, Glotar M. INCENP and
Aurora like kinase form a complex essential for chromosomsegregation and efficient completion of cytokinesis. Curr Biol
2000; 10: 1072-81.[7] Susanne MA Lens, Wolthuis RMS, Klompmaker R, et al. Survivin
is required for a sustained spindle checkpoint arrestin response tolack of tension. EMBO 2003; 22: 2934-2947.
[8] Zaffaroni N, Pennati M, Collela G, et al. Expression of anti-apoptotic gene survivin correlates with taxol resistance in human
ovarian cancer. Cell Mol Life Sci 2002; 59: 1406-12.[9] Taran J, Master Z, Yu JI, Rak J, Dumont DJ, Kerbel RS. A role of
survivin in chemoresistance of endothelial cells mediated byVEGF. PNAS 2002; 99: 4345- 9.
[10] Wall NR, O,Connor DS, Plescia J, Pommier Y, Altieri DC.Suppression of survivin phosphorylation on Thr34 by flavopyridol
enhances tumor cell apoptosis. Cancer Res 2003; 63: 230-5.[11] Tetsuhisa Y, Nobuhiko T. The role of survivin as a new target of
diagnosis and treatment in human cancer. Med Electron Microc2001; 34: 12-20.
[12] Altieri DC. The molecular basis and potential role of survivin incancer diagnosis and therapy. Trends Mol Med 2001; 12: 542-7.
[13] Reed JC. The survivin saga goes in vivo. J Clin Invest 2001; 108:965-9.
[14] Mahotka C, Liebman J, Wenzel M, et al. Differential subcellularlocalization of functionally divergent survivin splice variants. Cell
Death Differ 2002; 9: 1334-42.[15] Conway EM, Pollefeyt S, Cornelissen J, et al. Three differentially
expressed survivin cDNA variants encode proteins with distinctanti-apoptotic functions. Blood 2000; 1435-42.
[16] Okada E, Murai Ymatsui K, Isizawa S, et al. Survivin expression intumor cell nuclei is predictive of favorable prognosis in gastric
cancer patients. Cancer Lett 2001; 163: 109-16.
[17] Tamm I, Wang Y, Sausville E, et al. IAP family protein survivin
inhibits caspase activity and apoptosis induced by Fas (CD95),Bax, Caspases, and anti-cancer drugs. Cancer Res 1998; 58: 5315-
20.[18] Altieri DC. Validating survivin as a cancer therapeutic target. Nat
Rev Cancer 2003; 3: 46-54.[19] Song Z, Yao X, Wu M. Direct interaction between Smac/DIABLO
is essential for anti-apoptotic activity of survivin during taxol-induced apoptosis. J Biol Chem 2003; 278: 23130-40.
[20] Skaufias DA, Mollinari C, Lacroix FB, et al. Human Survivin isKinatochore-associated Passenger protein. J cell Biol 2000; 151:
1575-82.[21] Uren AG, Wong L, Pakusch M, et al. Survivin and the inner
centromere protein INCENP show similar cell cycle localizationand gene knockout phenotype. Curr Biology 2000; 10: 1319-28.
[22] Reiko Honda, Ronan K, Erich AN. Exploring the functionalinteractions between Aurora B, INCENP and Survivin in Mitosis.
MCB 2003; 14: 3325-41.[23] Konno R, Yamakawa H, Ito K, Sato S, Yajima A. Expression of
survivin and bcl2 in the normal human endometrium. Mol HumReported 2000; 6: 529-34.
[24] Endoh A, Asanuma K, Moriai R, et al. Expression of survivinmRNA in CD34+ cell. Clin Chim Acta 2001; 306: 149-51.
[25] Hoffman WH, Biade S, Zilfou JT, Chen J, Murphy M.Transcriptional repression of the anti-apoptotic survivin gene wild
type p53. J Biol Chem 2002; 277: 3247-57.[26] Mirza A, McGuirk M, Hockenberry TN, et al. Human survivin is
negatively regulated by wild type p53 and participate in p53-dependent apoptotic pathway. Oncogene 2002; 2: 2613-22.
[27] Lu C-D, Altieri DC, Tanugawa N. Expression of novel anti-apoptotic gene, survivin, correlated with tumor cell apoptosis and
p53 accumulation in gastric carcinomas. Cancer Res 1998; 58:1808-12.
[28] Sarela AI, Verveke CS, Ramsdale J, Davis CI, Markham AF,Guillou PJ. Expression of survivin, a novel inhibitor of apoptosis
and cell cycle regulatory protein, in pancreatic adenocarcinoma. BrJ Cancer 2002; 86: 886-92.
[29] Olie RA, Simoes-Wust AP, Baumann B, et al. A novel anti-senseoligonucleotides targeting survivin expression induces apoptosis
and sensitizes lung cancer cells to chemotherapy. Cancer Res 2000;60: 2805-09.
[30] Chuunyao Xia, Zhidong Xu, Xiaocheng Yuan, et al. Induction ofapoptosis in Mesothelioma cells by antisurvivin oligonucleotides.
Mol Cancer Ther 1, 687-694.[31] Ansel SM, Arendt BK, Grote DM, et al. Inhibition of survivin
expression suppresses the growth of aggressive non-Hodgkin’slymphoma. Leukamia 2004, 18: 616-23.
[32] Yamamoto T, Manome Y, Miyamoto A, Tanigava N. Developmentof novel gene therapy using survivin anti-sense expressing
adenoviral vectors. Gan To Kagaku Ryoho 2003; 30: 1805-8.[33] Mara F, Jonet P, Sophie C, et al. Fibronectin Protects Prostate
Cancer Cells from Tumor necrosis factor-alpha-induced Apoptosisvia the AKT/survivin pathway. J Biol Chem 2003; 278: 50402-11.
[34] Guan J, Chen J, Luo Y, Gao J, Qui H. Effects of Antisense bcl2 orsurvivin on the growth of neuroblastoma cell line SK-N-MC.
Zhonghua Yi Xue Za Zhi 2002; 82: 1536-40.[35] Zhao LJ, Jian H, Zhu H. Specific gene inhibition by adenovirus-
mediated expression of small interfering RNA. Gene 2003; 16:137-41.
[36] Ling X, Li F. Silencing of anti-apoptotic survivin gene by multipleapproaches of RNA interference technology. Biotechniques 2004;
36: 450-4.[37] Uchida H, Tanaka T, Sasaki K, et al. Adenivirus-Mediated
Transfer of siRNA against Survivin Induced Apoptosis andAttenuated Tumor Cell Growth in Vitro and in Vivo. Mol Ther
2004; 10: 162-71.[38] Yang G, Huang AL, Tang N, et al. Inhibition of survivin
expression in liver cancer cells by shRNA. Zhonghua Gan ZangBing Za Zhi 2003; 11: 712-5.
[39] Mesri M, Wall NR, Li J, Kim RW, Altieri DC. Cancer genetherapy using a survivin mutant adenovirus. J Clin Invest 2001;
108: 981-90.[40] Zhu HX, Zhou CQ, Zang G, et al. Survivin mutants reverse the
malignancy of Hela cells. Ai Zheng 2003; 22: 467-70.
Not For Distribtion

Human Anti-Apoptotic Survivin Gene Current Cancer Therapy Reviews, 2006, Vol. 2, No. 1 79
[41] Shui Ping Tu, Xiao Hua Jiang, Marie CM Lin, et al. Suppression of
Survivin Expression Inhibits in Vivo Tumorigenicity andAngiogenesis in Gasteric Cancer. Cancer Res 2003; 7724-32.
[42] Grunstein M. Histone acetylation in chromatin structure andtranscription. Nature 1997; 389: 349–52.
[43] Hassing CA, Schreiber SL. Nuclear histone acetylases anddeacetylases and transcriptional regulation: HATs off to HDACs.
Curr Opin Chem Biol 1999; 300–8.[44] Kouzarides T. Histone acetylases and deacetylases in cell
proliferation. Curr Opin Genet Dev 1999; 9: 40–8.[45] Jenuwein T, Allis CD. Translating the histone code. Science 2001;
293: 1074–80.[46] De Schepper S, Bruwiere H, Verhult T, et al. Inhibition of histone
deacetylases by clamydocin induces apoptosis and proteasome-mediated degradation of survivin. JPET 2003; 304: 881-8.
[47] Fei Guo, Celia Sigua, Jianguo Tao, et al. Cotreatment with HistoneDeacetylase Inhibitor LAQ824 Enhances Apo-2L/Tumor Necrosis
Factor-Related Apoptosis Inducing Ligand-Induced Death InducingSignaling Complex Activity and Apoptosis of Human Acute
Leukemia Cells. Cancer Res 2004; 64: 2580-9.[48] Choi KS, Lee TH, Jung MH. Ribozyme mediated cleavage of the
human survivin mRNA and inhibition of anti-apoptotic function ofsurvivin in MCF-7 cells. Cancer gene Ther 2003 Feb, 10, 87-95.
[49] Pennati M, Binda M, Colella G, et al. Ribozyme mediated
inhibition of survivin expression increases spontaneous and druginduced apoptosis and decreases the tumirigenic potential of human
prostate cancer cells. Oncogene 2004, 23, 386-94.[50] Pennati M, Binda M, Colella G, et al. Radiosesitisation of human
melanoma cells by ribozyme mediated inhibition of survivinexpression. J Invest Dermatol 2003; 120: 648-54.
[51] Pennati M, Binda M, Colella G, et al. Ribozyme-mediated down-regulation of survivin expression sensitizes human melanoma cells
to topotecan in vitro and in vivo. Carcinogenesis 2004; 25: 1129-36.
[52] Reker S, Becker JC, Svane IM, Ralfkiaer E, Straten PT, AndersenMH. HLA-B35-restricted immune responses against survivin in
cancer patients. Int J Cancer 2004; 108: 937-41.[53] Pisarev V, Yu B, Salup R, Sherman S, Altieri DC, Gobrilovich DI.
Full-length dominant-negative survivin for cancer immunotherapy.Clin Cancer Res 2003; 9 : 6523-33.
[54] Tetsuhiro T, Fumitake H, Toshihiko F, et al. Phase I clinical studyof anti-apoptosis protein, survivin derived peptide vaccine therapy
for patients with advanced or recurrent colorectal cancer. Journal ofTranslational Medicine 2004; 2-19.
Received: July 22, 2005 Revised: October 14, 2005 Accepted: October 19, 2005
Not For Distribtion





























