Embed Size (px)
Citation preview

Jourrial qf Neurochemr,rry Raven Press, Ltd., New York 0 199 1 International Society for Neurochemistry
Tumor Necrosis Factor-Induced Proliferation of Astrocytes from Mature Brain Is Associated with Down-Regulation
of Glial Fibrillary Acidic Protein rnRNA
*Krzysztof Selmaj, *Bridget Shafit-Zagardo, ?Dennis A. Aquino, ?Muhammad Farooq, *$Cedric S. Raine, ?$William T. Norton, and *$Celia F. Brosnan
Departments of *Pathology (Neuropathology), ?Neurology, and $Neuroscience, Albert Einstein College of Medicine, Bronx, New York, U.S.A.
Abstract: Previous results from this laboratory have shown that tumor necrosis factor (TNF) is mitogenic for bovine astrocytes in chemically defined (CD) medium. The maxi- mum mitogenic response was detected with 200 U/ml at 48 h. We have now extended these studies to assess the effect of TNF on message levels for the intermediate filament proteins glial fibrillary acidic protein (GFAP) and vimentin. The results have shown that, whereas TNF had only a slight effect on vimentin mRNA, TNF induced a marked decrease to 4.3 k 2.0% of controls in GFAP mRNA which was both time and dose dependent. The lowest effective dose was 50 U/ml and the maximal effective dose was 200 U/ml. Kinetic anal- ysis of this response demonstrated that a marked decrease in GFAP mRNA was present at 12 h and continued to decrease through 72 h. To determine the reversibility of the TNF effect, astrocyte cultures were exposed to 200 U/ml TNF for varying periods of time and then cultured in fresh CD medium. A 1-h pulse with TNF was sufficient to reduce GFAP mRNA levels when measured 24 h later. However, cultures incubated
with 200 U/ml TNF for 48 h followed by incubation in CD medium without TNF for 7 days showed that GFAP mRNA levels had returned to 60% of the control values. Nuclear runoff assays showed that the effect of TNF on GFAP mRNA was at the posttranscriptional level. Polyacrylamide gel elec- trophoretic analysis of astrocyte cytoskeletal proteins dem- onstrated that GFAP levels were reduced after a 5-day in- cubation with 200 U/ml TNF whereas protein levels of vi- mentin and actin were not significantly changed. Western blots confirmed that GFAP levels were reduced to 36% of the control values. Thus the effect of TNF on GFAP mRNA expression was not due to a generalized effect on intermediate filament metabolism. Key Words: Tumor necrosis factor- Glial fibrillary acidic protein-RNA-Gene regulation- Cytokines-CNS. Selmaj K. et al. Tumor necrosis factor- induced proliferation of astrocytes from mature brain is as- sociated With down-regulation of glial fibrillary acidic protein mRNA. J. Neurochem. 57,823-830 (1991).
Interactions between cells of the immune system and the CNS are well established. Included in these inter- actions are the observations that astrocytes both pro- duce and respond to a variety of cytokines. For ex- ample, it has been shown that astrocytes express major histocompatibility complex class I and class I1 antigens on stimulation with interferon-y (IFN,) (Hirsch et al., 1983; Wong et al., 1984; Fierz et al., 1985) and tumor necrosis factor (TNF) (Mauerhoff et al., 1988; Cannella and Raine, 1989). These properties implicate astrocytes in an array of immunological processes within the CNS.
Furthermore, astrocyte hypertrophy and hyperplasia are frequently observed at the site of immunologic in- jury in the CNS (Raine, 1983) and several studies have shown that cytokines produced by macrophages and lymphocytes can induce astrocyte proliferation in vivo (Giulian et al., 1988) and in vitro (Memll et al., 1984; Giulian and Lachman, 1985). In our previous study (Selmaj et al., 1990), we assessed the effects of several recombinant cytokines on astrocytes and demonstrated that TNF induced astrocyte proliferation in a time- and dose-dependent manner. In chemically defined
Received August 17, 1990; revised manuscript received January 3, 1991; accepted February 5 , 1991.
Address correspondence and reprint requests to Dr. K. Selmaj at Department of Pathology, Albert Einstein College of Medicine, 1300 Moms Park Avenue, Bronx, NY 10461, U.S.A.
Abbreviations used: CD, chemically defined medium; dpi, days
postinoculation, EU, endotoxin units; GFAP, glial fibrillary acidic protein; IFN,, interferon-y; IL- I@, interleukin-ID; LT, lymphotoxin; PKC, protein kinase C; PLL, poly-L-lysine; SDS, sodium dodecyl sulfate; SSC, saline-sodium citrate; TNF, tumor necrosis factor; UTR, untranslated region.
823
824 K. SELMAJ ET AL.
(CD) medium, maximum proliferative responses were obtained with 200 U of TNF following incubation for 48 h. To a lesser degree, lymphotoxin (LT) and inter- leukin-6 (IL-6) were also mitogenic for astrocytes in CD medium. In contrast, interleukin-lp (IL-1p) did not induce astrocyte proliferation in CD medium but showed some synergistic effect with serum after 72 h. A similar proliferative effect of TNF for adult human astrocytes has recently been documented by Ransohoff and colleagues (Barna et al., 1990). These observations raise the question of whether TNF, in parallel with its proliferative effect, affects the differentiation and mat- uration of astrocytes. The major specific marker for mature astrocytes is glial fibrillary acidic protein (GFAP) (Eng, 1985). In this report, we have demon- strated by northern and western blot analyses that TNF inhibited GFAP gene expression in astrocyte cultures in a time- and dose-dependent manner.
MATERIALS AND METHODS
Astrocyte cell cultures Glial cells were isolated from fresh calf white matter by
the method of Norton et al. (1988). In brief, white matter was minced, trypsinized (0.1% wt/vol), and disrupted through a 145-pm nylon screen and a 74-pm stainless steel screen. The cell suspension was fractionated by centrifugation on a 48%/44% sucrose density gradient. After centrifugation at 3,600 g for 15 min, the cells were collected as a pellet. The glial cells were cultured for 4 days in 60-mm poly-L-lysine (PLL)-coated plastic dishes at a density of 4 X lo5 cells/cm2 in medium which consisted of minimum essential medium with Earle’s salts, 15% heat-inactivated calf serum, penicillin- streptomycin, fungizone, glutamine, and 0.5% glucose (all from GIBCO, Grand Island, NY, U.S.A.). The cells were then switched to CD 0 1-serum-free medium (Bottenstein, 1986) and maintained until 21 days in vitro. At this time, the cultures comprised a flat sheet of large astrocytes with thick processes. Clusters of oligodendroglia were present on the surface of the astrocyte layer. The cells were trypsinized (0.1% wt/vol), plated in T-25 flasks coated with PLL, and incubated for 70 min to allow for astrocyte adherence. They were then shaken at 200 rpm for 20 min at 37°C. The non- adherent oligodendrocytes were removed. Adherent astro- cytes were cultured in serum-containing medium for 3 days and then trypsinized (0.1% wt/vol) and replated into T-75 flasks. Astrocytes were cultured for 5 days in serum-medium and then switched to CD medium. They were used for ex- periments 2 days later when they had formed subconfluent monolayers (3.8 X lo6 cells). At the end ofthe culture period, the maximum number of cells was 9.3 X lo6 in cultures exposed to 200 U/ml TNF for 72 h. In cultures used for this study, -98% of cells were positive for the astrocyte-specific marker GFAP and -2% were positive for the microglial cell marker EMBl1 (Esiri and McGeer, 1986) prior to experi- mentation. No fibronectin staining was detected. No altera- tion in this staining pattern was detected in cultures exposed to 200 U/ml TNF for 72 h.
RNA isolation, electrophoresis, and blotting Total RNA was isolated from astrocytes using the hot phe-
nol method (Soeiro and Darnell, 1969) and applied to 1% agarose gels containing 6% formaldehyde. Blotting onto Gene
Screen (New England Nuclear, Boston, MA, U.S.A.), and hybridization were according to Thomas (1 980). Filters were hybridized with 5 X 107-10s cpm/pg of human GFAP cDNA probe, labeled with 32P by random priming. Hybridization was camed out overnight at 65°C in 5X saline-sodium citrate (SSC), 5X Denhardt’s ( 1 X = 0.02% bovine serum albumin, 0.02% Ficoll 400, 0.02% polyvinylpyrrolidone), 50 m M Na phosphate (pH 6.8),0.1% sodium dodecyl sulfate (SDS), and 50 pg/ml of salmon sperm DNA. Filters were washed four times in 2X SSC, 0.1 % SDS; once in 1 X SSC, 0.1% SDS; and once in 0.lX SSC, 0.1% SDS, for 15 min per wash, 65°C. Autoradiography was camed out using Kodak XAR film and an intensifying screen at -70°C (Shafit-Zagardo et al., 1988).
To remove the 32P-labeled GFAP probe, filters were washed at 100°C with 0.1X SSC, 0.1% SDS, 1% sodium pyrophos- phate and exposed to film overnight to verify that no signal was emitted. Filters were subsequently hybridized with a vi- mentin cDNA probe and finally with an 18s ribosomal cDNA probe. Following exposures of the x-ray film, the densities of the hybridization signals for GFAP, vimentin, and ribosomal transcripts were determined by densitometry. Scans of hy- bridization signals on multiple autoradiograms were highly reproducible. Other probes were hybridized exactly as de- scribed for GFAP.
Probes The human GFAP cDNA clone has been described pre-
viously by Shafit-Zagardo et al. (1988). The mouse GFAP
TNF DOSE RESPONSE -
7
0-OVlMENTlN 0 .--.GFM
0 10 50 100 200
DOSE UNITS/ML
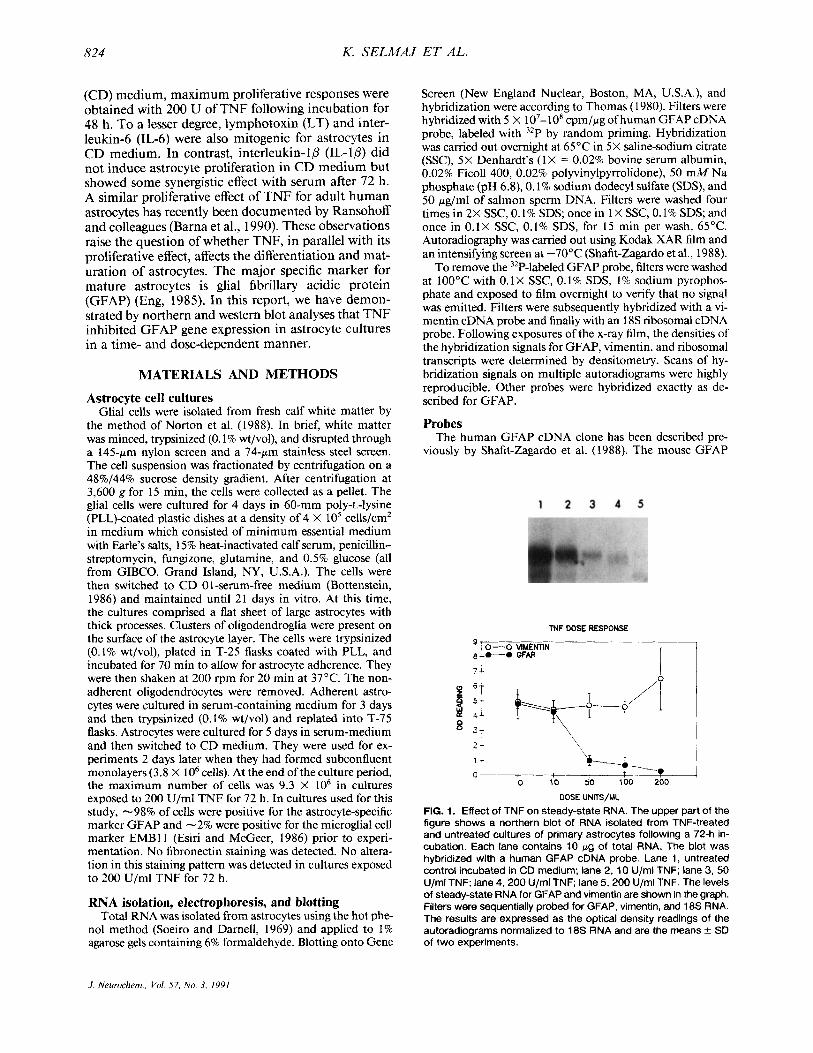
FIG. 1. Effect of TNF on steady-state RNA. The upper part of the figure shows a northern blot of RNA isolated from TNF-treated and untreated cultures of primary astrocytes following a 72-h in- cubation. Each lane contains 10 pg of total RNA. The blot was hybridized with a human GFAP cDNA probe. Lane 1, untreated control incubated in CD medium; lane 2, 10 U/ml TNF; lane 3, 50 U/ml TNF; lane 4,200 U/ml TNF; lane 5,200 U/ml TNF. The levels of steady-state RNA for GFAP and vimentin are shown in the graph. Filters were sequentially probed for GFAP, vimentin, and 18s RNA. The results are expressed as the optical density readings of the autoradiograms normalized to 18s RNA and are the means & SD of two experiments.
J. Neunshem.. Val. 57. No. 3, 1991
TNF DOWN-REGULATES GFAP mRNA 825
cDNA clone (SulI-Hind111 fragment) was utilized for the nuclear runoff assay and the full GFAP clone was provided by Dr. N. Cowan (Lewis et al., 1984). A human 18s ribosomal cDNA clone, pRM 17, was provided by Dr. Norman Arnheim (Arnheim et al., 1980). A human vimentin cDNA probe was made available by Dr. R. Baserga (Ferrari et al., 1986).
Nuclear runoff experiments and slot blot hybridizations
The isolation of nuclei, nuclear runoff experiments, and hybridization experiments were performed as described pre- viously by Shafit-Zagardo et al. (1983). 32P-labeled RNA was hybridized at 65°C overnight with cDNAs (GFAP, vimentin,
10 UNITS TNF
0- 0 VlMENnN 0-OGFAP
6
0 12h 24h 48h 72h
TIME POST-CHALLENGE
200 UNrrS TNF
5 -1 0- 0 WMEMlN 0-OGFAP
z + \
12h 24h 48h 72h TIME POST-CHALLENGE
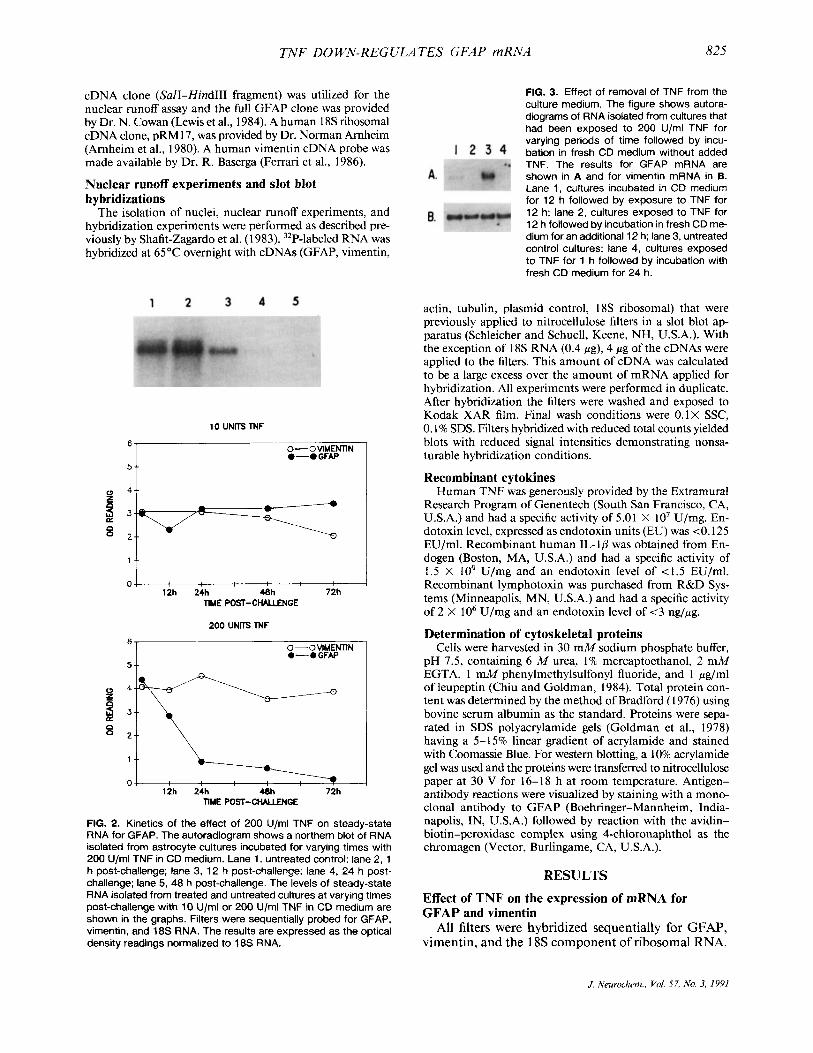
FIG. 2. Kinetics of the effect of 200 U/ml TNF on steady-state RNA for GFAP. The autoradiogram shows a northern blot of RNA isolated from astrocyte cultures incubated for varying times with 200 U/ml TNF in CD medium. Lane 1, untreated control: lane 2, 1 h post-challenge; lane 3, 12 h post-challenge; lane 4, 24 h post- challenge; lane 5, 48 h post-challenge. The levels of steady-state RNA isolated from treated and untreated cultures at varying times post-challenge with 10 U/ml or 200 U/ml TNF in CD medium are shown in the graphs. Filters were sequentially probed for GFAP, vimentin, and 18s RNA. The results are expressed as the optical density readings normalized to 18s RNA.
FIG. 3. Effect of removal of TNF from the culture medium. The figure shows autora- diograms of RNA isolated from cultures that had been exposed to 200 U/ml TNF for varying periods of time followed by incu- bation in fresh CD medium without added TNF. The results for GFAP mRNA are shown in A and for vimentin mRNA in B. Lane 1, cultures incubated in CD medium for 12 h followed by exposure to TNF for 12 h; lane 2, cultures exposed to TNF for 12 h followed by incubation in fresh CD me- dium for an additional 12 h: lane 3, untreated control cultures: lane 4, cultures exposed to TNF for 1 h followed by incubation with fresh CD medium for 24 h.
actin, tubulin, plasmid control, 18s ribosomal) that were previously applied to nitrocellulose filters in a slot blot ap- paratus (Schleicher and Schuell, Keene, NH, U.S.A.). With the exception of 18s RNA (0.4 pg), 4 pg of the cDNAs were applied to the filters. This amount of cDNA was calculated to be a large excess over the amount of mRNA applied for hybridization. All experiments were performed in duplicate. After hybridization the filters were washed and exposed to Kodak XAR film. Final wash conditions were 0 . lX SSC, 0.1% SDS. Filters hybridized with reduced total counts yielded blots with reduced signal intensities demonstrating nonsa- turable hybridization conditions.
Recombinant cytokines Human TNF was generously provided by the Extramural
Research Program of Genentech (South San Francisco, CA, U.S.A.) and had a specific activity of 5.01 X lo7 U/mg. En- dotoxin level, expressed as endotoxin units (EU) was <O. I25 EU/ml. Recombinant human IL-10 was obtained from En- dogen (Boston, MA, U.S.A.) and had a specific activity of 1.5 X lo9 U/mg and an endotoxin level of <1.5 EU/ml. Recombinant lymphotoxin was purchased from R&D Sys- tems (Minneapolis, MN, U.S.A.) and had a specific activity of 2 X lo6 U/mg and an endotoxin level of 1 3 ng/wg.
Determination of cytoskeletal proteins Cells were harvested in 30 mM sodium phosphate buffer,
pH 7.5, containing 6 M urea, 1% mercaptoethanol, 2 mM EGTA, 1 m M phenylmethylsulfonyl fluoride, and 1 pg/ml of leupeptin (Chiu and Goldman, 1984). Total protein con- tent was determined by the method of Bradford (1976) using bovine serum albumin as the standard. Proteins were sepa- rated in SDS polyacrylamide gels (Goldman et al., 1978) having a 5- 15% linear gradient of acrylamide and stained with Coomassie Blue. For western blotting, a 10% acrylamide gel was used and the proteins were transferred to nitrocellulose paper at 30 V for 16- 18 h at room temperature. Antigen- antibody reactions were visualized by staining with a mono- clonal antibody to GFAP (Boehringer-Mannheim, India- napolis, IN, U.S.A.) followed by reaction with the avidin- biotin-peroxidase complex using 4-chloronaphthol as the chromagen (Vector, Burlingame, CA, U.S.A.).
RESULTS
Effect of TNF on the expression of mRNA for GFAP and vimentin
All filters were hybridized sequentially for GFAP, vimentin, and the 18s component of ribosomal RNA.
J. Neurochem., Vol. 57, No. 3, I991

826 K. SELMAJ ET AL.
As shown in Fig. I , there is a gradual decrease in the GFAP mRNA level following incubation of astrocytes for 72 h with increasing doses of TNF. Cumulative results for two experiments are shown in the graph and indicate the densities of the hybridization signals for GFAP mRNA and vimentin mRNA, each normalized to 18s RNA. The lowest effective dose observed for GFAP mRNA was 50 U/ml and the maximal effect was noted at 200 U/ml. Higher doses of TNF did not induce further decreases in GFAP mRNA levels (data not shown). In contrast to the results obtained with GFAP, vimentin mRNA did not decrease at the same time points, indicating that the effect of TNF was not due to a generalized effect on intermediate filament gene expression.
The dose-response of the results for GFAP mRNA was consistent with the effect of TNF on astrocyte pro- liferation where a peak proliferative response occurred at 200 U/ml and no proliferative response was detected with doses of TNF lower than 50 U/ml (Selmaj et al., 1990).
Kinetics of TNF effect on GFAP mRNA. T o deter- mine how rapidly TNF induced a decrease in GFAP mRNA expression, astrocyte cultures were incubated with 10 U/ml or 200 U/ml TNF for 1-72 h. The results are shown in Fig. 2. They demonstrate that, whereas only minor fluctuations in GFAP mRNA were ob- tained with 10 U/ml, a significant decrease in GFAP mRNA was obtained in response to 200 U/ml TNF. This decrease was first detected after a 12-h incubation period and continued to decrease through 72 h. As noted previously, the levels of mRNA for vimentin were not significantly altered in response to TNF.
To test the effect of removal of TNF from the culture medium on GFAP mRNA, astrocyte cultures were ex- posed to 200 U/ml TNF for varying periods of time, then the TNF medium was removed and the cultures incubated in fresh CD medium. Companion cultures were exposed to 200 U/ml TNF for the same period of time at the end of the culture period. In Fig. 3 the control cultures are shown in lane 3. The results show that when astrocytes were incubated with TNF for 12 h followed by incubation in CD medium alone for a further 12 h, less GFAP mRNA was present (Fig. 3A, lane 2) than in cultures exposed to TNF for the final
FIG. 4. Determination of the revers- ibility of the effect of TNF on GFAP mRNA. The figure shows an auto- radiogram of RNA isolated from con- trol cultures and cultures that had been exposed to 200 U/ml TNF for 48 h. Lane 1, control untreated cells; lane 2, cultures exposed to TNF for 48 h followed by 7 days of incubation in CD medium; lane 3, cultures ex- posed to TNF for the final 48-h in- cubation.
12 h of in vitro culture (lane 1). This experiment also shows that as short a pulse with TNF as 1 h followed by a subsequent incubation with fresh CD medium for 24 h was sufficient to decrease GFAP mRNA expres- sion (lane 4) when compared to control cultures (lane 3). The results for vimentin are shown in Fig. 3B. Thus removal of TNF from the culture medium did not re- verse the effect of TNF on GFAP gene expression, at least within the initial 24 h.
Reversibility of the TNF effect on GFAP mRNA. To determine whether the effect of TNF on GFAP mRNA levels was reversible over longer periods of time, the cells were stimulated with TNF for 48 h and then in- cubated in fresh CD medium for 7 days. In these ex- periments, the level of GFAP mRNA returned to 60% of the control value (Fig. 4). Companion cultures ex- posed to TNF for the final 48 h showed the same loss of GFAP mRNA as that described previously (3.4% of the control value).
Nuclear runof assay. To determine whether the ef- fect of TNF was at the level of gene transcription, as- trocyte cultures were exposed to 200 U/ml TNF for 12 and 48 h, nuclei isolated, and nuclear runoff assays performed. The incorporation of 32P-UTP into actively synthesized RNA transcripts for GFAP, vimentin, ac- tin, tubulin, and 18s RNA was determined from iso- lated nuclei. In both the 12-h samples (Fig. 5) and the 48-h samples (data not shown) no difference in the newly synthesized message for GFAP was apparent. The possibility that these data could result from the use of saturated hybridization conditions was ruled out by the observation that when reduced total counts of labeled nuclear transcripts were added, reduced signals were obtained. The high stringency conditions used and the lack of cross-reactivity of the probe on the
FIG. 5. Nuclear runoff assays to determine whether the effect of TNF on GFAP mRNA is tran- scriptional. Panel T (TNF-treated 200 U/ml) and panel C (untreated control) illustrate autoradiograms of filter-bound cDNAs, each hy- bridized with 1 X 10' counts Of duplicate samples of total 32P-la- beled RNA. Final stringency was
vimentin; G, GFAP; A, actin; T, tubulin; 18S, 18s ribosomal RNA. Blank lanes between T and 18s RNA are the plasmid control.
0.1X SSC, 0.1% SDS, 65°C. V,
J Neurochem., Vol. 57, No. 3, 1991

TNF DOWN-REGULATES GFAP mRNA 82 7
northern blots also rule out the possibility that the nu- clear runoff experiments reflect cross-hybridization with other radiolabeled mRNAs. These results suggest, therefore, that the regulation of GFAP mRNA in as- trocytes by TNF is primarily at the posttranscriptional level.
Effects of other cytokines on intermediate filament mRNA
The cytokines IL- 1 p and LT were also tested at a concentration shown previously to be optimal for the stimulation of a proliferative effect on astrocyte cultures (Selmaj et al., 1990). Both IL- 1 /3 and LT reduced GFAP mRNA levels but the level of reduction was less than that detected with TNF. When exposed to IL-lp (200 U/ml), LT (100 U/ml), or TNF (100 U/ml) for a 72- h period, the results, expressed as a percent of control value, for GFAP mRNA were IL-lp, 32%; LT, 28%; or TNF, 3.4%; and for vimentin they were IL-ID, 96%; LT, 90%; or TNF, 90%.
Effect of TNF on cytoskeletal proteins in astrocytes When equal amounts of total astrocyte protein were
electrophoresed in a slab gel and stained with Coo- massie Blue, there was no change in the intensity of the bands for vimentin and actin in any of the samples. However, in cultures exposed to 60 U/ml or 200 U/ ml for 5 days, the intensity of the GFAP band appeared to be reduced (data not shown). A western blot (Fig. 6) of the same samples confirmed the loss GFAP im- munoreactivity after incubation with TNF for 5 days. Densitometric scans of the western blot showed that there was no change from the control value for 20 U of TNF (lane 2), a 3 1 % reduction in GFAP with 60 U of TNF (lane 3), and a 64% reduction in GFAP for 200 U of TNF (lane 4).
DISCUSSION
The results of the studies reported here show that the proliferative effect of TNF for bovine astrocytes
FIG. 6. Effect of varying doses of TNF on GFAP. The levels of GFAP were determined by western blotting of total protein isolated from cultures treated with varying doses of TNF following a 5-day incubation. Two micrograms of total protein were analyzed in each case. A minor breakdown product was observed. Lane 1, untreated culture; lane 2,20 U/ml TNF; lane 3,60 Ufrnl TNF lane 4,200 U/ ml TNF; lane 5, standard GFAP purified from rat spinal cord.
was associated with a marked decrease in the levels of mRNA for GFAP and an associated but less severe decrease in the levels of GFAP. For six separate ex- periments, the cumulative data for GFAP mRNA fol- lowing exposure to 200 U of TNF for 72 h were 4.3 k 2.0% of the control value. In contrast, the levels of vimentin mRNA, the other major intermediate fila- ment protein present in these astrocytes, was not af- fected by TNF. Analysis of the nuclear runoff assays suggests that the TNF-induced alteration in gene expression of GFAP occurred posttranscriptionally, possibly at the level of mRNA stability. Analysis of GFAP showed that following exposure to the same level of TNF for 5 days, 36% of the control value remained. This persistence of the protein is to be expected because the metabolic half-life of the largest pool of this protein in cultured rat astrocytes is approximately 8 days (Chiu and Goldman, 1984). Our data for GFAP after 5 days of TNF treatment are in excellent agreement with the measured decay of radioactivity in GFAP over the same period in pulse-labeled rat astrocytes (Chiu and Gold- man, 1984). This slow decrease in protein relative to mRNA explains why the immunocytochemical stain- ing for GFAP was not significantly altered after ex- posure to TNF for 72 h. As expected from the mRNA studies, the levels of vimentin were not altered.
The expression of GFAP by astrocytes is known to be developmentally regulated and altered in various pathologic states (Chiu and Goldman, 1985). Precursor glial cells have been shown both in vivo and in vitro to express initially only vimentin and then to express GFAP as they develop into astrocytic forms (Dahl et al., 1981; Norton et al., 1988). However, the time at which the expression of GFAP is first evident differs among species and also may differ among astrocytes in different regions of the CNS. That the expression does not always coincide with major developmental stages has led to the suggestion that GFAP may be regulated by hormones or by growth factors (Momson et al., 1985). Following injury to the brain, GFAP expression may increase rapidly and high GFAP levels are characteristic of astrocytic scars in various patho- logic conditions (Bignami et al., 1980). In acute ex- perimental allergic encephalomyelitis in the Lewis rat, widespread enhanced immunostaining for GFAP in hypertrophic astrocytes in the spinal cord has been ob- served beginning 10-1 1 days postinoculation (dpi), a time well before any increase in GFAP could be de- tected (Smith et al., 1983; Aquino et al., 1988). At l l dpi, the levels of GFAP mRNA rose to sevenfold con- trol values, but the levels of GFAP rose only slowly thereafter to 1.5- to 2-fold control values by 35 dpi (Aquino et al., 1990). This astrocyte reaction appeared not to involve any significant amount of astrocyte rep- lication (Smith et al., 1987). In other models, an in- crease in total GFAP was associated with an increase in the number of astrocytes (Topp et al., 1989).
Most of the studies that have addressed the regulation
J. Neurochem., Vol. 57, No. 3, 1991

828 K. SELMAJ ET AL.
of astrocyte GFAP expression have been performed at the protein level. Less is known about the molecular mechanisms that regulate GFAP. Shafit-Zagardo et al. (1 988) have reported the involvement of both cyclic AMP-dependent protein kinase and protein kinase C (PKC) in the regulation of GFAP gene expression. The results have shown that all cyclic AMP-stimulatory agents increase steady-state GFAP mRNA levels. In contrast, activation of protein kinase C with phorbol 12-myristate 13-acetate results in an initial increase followed by a significant decrease in GFAP mRNA levels. These results suggest that PKC activity may also be positively correlated with GFAP mRNA levels but operating through a different pathway than cyclic AMP-dependent kinase. In both second messenger systems alterations in GFAP mRNA levels occurred primarily posttranscriptionally. Analysis of the nuclear runoff assays also suggests that the TNF-induced al- teration in gene expression of GFAP was posttran- scriptional. As we have probed total RNA, potential sequestration of RNA cannot explain the observed re- sults and thus we feel that the most likely explanation is that TNF is operating through a mechanism that alters GFAP mRNA stability. Several mechanisms have been proposed for the control of posttranscrip- tional gene expression including regulatory elements in the 3' untranslated region (UTR), the sequence of bases in the 3' UTR, and cytosolic levels of soluble protein (Caput et al., 1986; Shaw and Kamen, 1986; Kruys et al., 1987; Pachter et al., 1987). GFAP mRNA has a 1.4-kb 3' UTR (Lewis et al., 1984) that may be involved in its stability. Further experiments will be designed to explore the effect of TNF on potential reg- ulatory elements in the genome. Regulation of gene expression by TNF has also been noted in other sys- tems. In cultured human marrow stromal cells, TNF increased the expression of the protooncogene c-abl (Andrews et al., 1989); the gene for HLA class I antigens was enhanced by TNF in endothelial cells and dermal fibroblasts (Collins et al., 1986); and TNF induced granulocyte-monocyte colony-stimulating factor gene expression in human lung fibroblasts, endothelial cells, and several malignant tissues (Munker et al., 1986). However, TNF inhibits adipocyte gene expression (Torti et al., 1985), and c-myc expression in HeLa cells (Yarden and Kimchi, 1986). TNF in transfected mice lowered albumin mRNA by 90% (Brenner et al., 1990). The regulation of the adipocyte-gene expression and of c-myc was shown to be at the transcriptional level.
The extent to which changes in GFAP mRNA or in GFAP are linked to cell division is not clear at the present time. In primary cultures of rodent astrocytes, GFAP content increases during the period of rapid proliferation and continues to increase more slowly after the cells are confluent (e.g., Sensenbrenner et al., 1980; Chamire-Bertrand et al., 1989). GFAP mRNA also increases during the proliferative period, but then declines as proliferation slows (Chamkre-Betrand et
al., 1989; Tardy et al., 1989). A brain-derived astroglial growth factor, which is mitogenic for primary astro- cytes, had no effect on GFAP levels (Weibel et al., 1985), whereas astrocyte replication induced by a cy- tokine involved in B-cell stimulation was associated with increases in GFAP mRNA levels (Benveniste et al., 1989). Goldman and Chiu (1 984) have shown that elevation in GFAP mRNA following exposure of rat primary astrocyte cultures to dibutyryl cyclic AMP is not solely related to cessation of mitotic activity. None of these studies have shown that GFAP or GFAP mRNA is depressed during astrocyte proliferation.
TNF was originally described as a product of acti- vated macrophages. However, more recent experiments have shown that other cells, including astrocytes, may secrete TNF following stimulation, although this latter finding is controversial. For example, Lieberman et al. ( 1989) have demonstrated that stimulation of astrocytes with lipopolysaccharide or a neurotropic paramyxo- virus induces TNF production, and Chung and Ben- veniste (1 990) have also shown production of TNF by astrocytes after induction by lipopolysaccharide, IFN., , and IL- lp. Another recent study, however, claims that TNF is synthesized only by microglia in these cultures, not by astrocytes (Hetier et al., 1990). Further support for the view that astrocytes can produce TNF comes from the immunohistochemical studies of pathologic tissue. Both the studies of Hofman et al. (1989) and ourselves (Selmaj et al., 1991) have demonstrated the presence of TNF in astrocytes in the CNS of autopsy tissue from patients with multiple sclerosis. If TNF is both secreted by and mitogenic for astrocytes it may have a role as an autocrine growth factor for these cells, similar to that of interleukin-2 for T-cells.
Acknowledgment: This work was supported by National Multiple Sclerosis Society (NMSS) grants RG 1089, RG 100 1- G-7, and, to K.S., FG 795A1, and by the National Institutes of Health grants NS 11920, NS 23705, NS 08952, and AG 07677. K.S. is a recipient of the Advanced Fellowship of NMSS and is on leave of absence from the Department of Neurology, Medical Academy of Lodz, Lodz, Poland.
REFERENCES Andrews D. F., Nemunaitis J. J., and Siner J. W. (1989) Recombinant
tumor necrosis factor alpha and interleukin- I alpha increase expression of c-abl protooncogene mRNA in cultured human marrow stroma cells. Proc. Natl. Acad. Sci. USA 86,6788-6792.
Aquino D. A,, Chiu F.-C., Brosnan C. F., and Norton W. T. (1988) Glial fibrillary acidic protein increases in the spinal cord of Lewis rats with acute experimental autoimmune encephalomyelitis. J. Neurochem. 51, 1085-1096.
Aquino D. A., Shafit-Zagardo B., Brosnan C. F., and Norton W. T. (1990) Expression of dial fibrillary acidic protein and neurofil- ament mRNA in gliosis induced by experimental autoimmune encephalomyelitis. J. Neurochem. 54, 1398- 1404.
Arnheim N., Krystal M., Schmickel R., Wilson G., Ryder O., and Zimmer E. ( 1 980) Molecular evidence for genetic exchanges among ribosomal genes on non-homologous chromosomes in man and apes. Proc. Natl. Acad. Sci. USA 77,5201-5205.
Barna B. P., Estes M. L., Jacobs B. S., Hudson S. , and Ransohoff
J Neurochem., Vol 57, No. 3, 1991

TNF DO WN-REGULATES GFAP mRNA 829
R. M. (1990) Human astrocytes proliferate in response to tumor necrosis factor alpha. J. Neuroimrnunol. 30, 239-243.
Benveniste E. N., Whitaker J. N., Gibbs D. A,, Sparacio G. M., and Butler J. L. (1989) Human B cell growth factor enhances pro- liferation and glial fibrillary acidic protein gene expression in rat astrocytes. Int. Irnmunol. 1, 219-228.
Bignami A,, Dahl D., and Rueger D. C. (1980) Glial fibrillary acidic protein in normal cells and in pathological conditions. Adv. Cell. Neurobiol. 1,285-3 10.
Bottenstein J. E. (1986) Growth requirements in vitro of oligoden- drocyte cell lines and neonatal rat brain oligodendrocytes. Proc. Natl. Acad. Sci. USA 83, 1955-1959.
Bradford M. M. (1976) A rapid and sensitive method for the quan- titation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254.
Brenner D. A., Buck M., Feitelberg S. P., and Chojkier M. (1990) Tumor necrosis factor-alpha inhibits albumin gene expression in murine model of cachexia. J. Clin. Invest. 85, 239-248.
Cannella B. and Raine C. S. (1989) Cytokine up-regulates Ia expres- sion in organotypic cultures of central nervous system tissue. J. Neuroimmunol. 24,239-248.
Caput D., Beutler B., Hartog K., Thayer R., Bronw-Shimer S., and Cerami A. ( I 986) Identification of a common nucleotide se- quence in the 3' untranslated region of mRNA molecules spec- ifying inflammatory mediators. Proc. Natl. Acad. Sci. USA 83, 1670-1674.
CharriZre-Bertrand C., LePrince G., Bouchier C., Fages C., Tardy M., and Nunez J. (1989) Expression oftubulin, GFAP and their encoding mRNAs during the proliferation and differentiation of cultured astrocytes. Neurochern. Int. IS, 2 15-222.
Chiu F.-C. and Goldman J. E. ( 1 984) Synthesis and turnover of cyto- skeletal proteins in cultured astrocytes. J. Neurochem. 42, 166- 174.
Chiu F.-C. and Goldman J. E. (1985) Regulation of glial fibrillary acidic protein expression in CNS development and in patho- logical states. J. Neuroimmunol. 8, 283-292.
Chung I. Y. and Benveniste E. N. (1990) Tumor necrosis factor alpha production by astrocytes: induction by lipopolysaccharide, in- terferon-gamma, and 1 beta. J. Immunol. 144, 2999-3007.
Collins T., Lapierre L. A., Fiers W., Strominger J. L., and Pober J. S. (1986) Recombinant human necrosis factor increases mRNA levels and surface expression of HLA-A, B antigens in vascular endothelial cells and dermal fibroblasts in vitro. Proc. Natl. Acad. Sci. USA 83,446-450.
Dahl D., Rueger D. C., Bignami A., Weber K., and Osborne M. (1981) Vimentin, the 57,000 dalton protein of fibroblast fila- ments, is the major cytoskeletal component in immature glia. Eur. J . Cell Biol. 24, 191-196.
Eng L. F. (1985) Glial fibrillary acidic protein (GFAP): the major protein of glia intermediate filaments in differentiated astrocytes. J. Neuroimmunol. 8, 203-2 14.
Esiri M. M. and McGeer J. 0. (1986) Monoclonal antibody to mac- rophages and microglial cells in human brain. J. Clin. Pathol.
Ferrari S., Battini R., Kacmarek L., Rittling S., Calabretta B., DeRiel J. K., Philiponis V., Wei J. F., and Baserga R. (1986) Coding sequence and growth regulation of the human vimentin gene. Mol. Cell. Biol. 6, 36 14-3620.
Fierz W., Endler B., Reske K., Wekerle H., and Fontana A. (1 985) Astrocytes as antigen presenting cells. 1. Induction of Ia antigen expression on astrocytes by T-cells via immune-interferon and its effect on antigen presentation. J. Irnmunol. 134,3785-3793.
Giulian D. and Lachman L. B. (1985) Interleukin-l stimulation of astroglial proliferation after brain injury. Science 228,495-499.
Giulian D., Woodward J., Young D. G., Krebs J. F., and Lachman L. B. (1 988) Interleukin-I injected into mammalian brain stim- ulates astrogliosis and neovascularization. J. Neurosci. 8,2485- 2490.
Goldman J. E. and Chiu F.-C. (1984) Growth kinetics, cell shape and the cytosleton of primary astrocyte cultures. J. Neurochern.
39,6 15-62 1.
42, 175-184.
Goldman J. E., Schaumburg H. H., and Norton W. T. (1978) Isolation and characterization of glial filaments from human brain. J. Cell Biol. 78,426-440.
Hetier E., Ayala J., Bousseau A., Denefle P., and Prochaintz A. (1990) Amoeboid microglial cells and not astrocytes synthesize TNF- alpha in Swiss mouse brain cell cultures. Eur. J. Neurosci. 2, 762-768.
Hirsch M. R., Wietzerbin J., Pierres M., and Goridis C. (1983) Expression of la antigens by cultured astrocytes treated with gamma-interferon. Neurosci. Lett. 41, 199-204.
Hofman F. M., Hinton D. R., Johnson K., and Menill J. E. (1989) Tumor necrosis factor identified in multiple sclerosis brain. J. Exp. Med. 170, 307.
Kruys V., Wathelet M., Poupart P., Contreras R., Fiers W. Content J., and Huez G. (1987) The 3'untranslated region of the human interferon-beta mRNA has an inhibitory effect on translation. Proc. Natl. Acad. Sci. USA 84, 6030-6034.
Lewis S. A,, Balcerek J. M., Krek V., Shelanski M., and Cowan N. J. ( 1984) Sequence of a cDNA encoding mouse glial fibrillary acidic protein: structural conservation of intermediate filaments. Proc. Natl. Acad. Sci. USA 81, 2743.
Lieberman A. P., Pitha P. M., Shin H. S., and Shin M. L. (1989) Production of tumor necrosis factor and other cytokines by as- trocytes stimulated with lipopolysaccharide or a neurotropic vi- rus. Proc. Natl. Acad. Sci. USA 86, 6348-6352.
Mauerhoff T., Pugol-Borrell R., Mirakian R., and Bottazzo J.-F. (1988) Differential expression and regulation of major histo- compatibility complex (MHC) products in neural and glial of human fetal brain. J. Neuroimmunol. 18, 271-289.
Merrill J. E., Kutsunai S., Mohlstrom C., Hofman F. Groopman J., and Golde D. W. ( 1 984) Proliferation of astroglia and oligoden- droglia in response to human T cell-derived factors. Science 224, 1428-1430.
Momson R. S., DeVellis J., Bradshaw R. A., and Eng L. F. (1985) Hormones and growth factors induce the synthesis of glial fi- brillary acidic protein in rat brain astrocytes. J. Neurosci. Res.
Munker R., Gasson J., Ogawa M., and Koeffler H. P. (1986) Recom- binant human TNF induces production of granulocyte-monocyte colony-stimulating factor. Nalure 323, 79-82.
Norton W. T. and Farooq M. (1989) Astrocytes cultured from mature brain derive from glial precursor cells. J. Neurosci. 9, 769-775.
Norton W. T., Farooq M., Chiu F.-C., and Bottenstein J. E. (1988) Pure astrocyte cultures derived from cells isolated from mature brain. Glia 1, 403-414.
Pachter J. S., Yen T. J., and Cleveland D. W. (1987) Autoregulation of tubulin expression is achieved through specific degradation of polysomal tubulin mRNAs. Cell 51, 283-292.
Raine C. S. (1983) Multiple sclerosis and chronic relapsing EAE: comparative ultrastructural pathology, in Multiple Sclerosis (Hallpike J. F., Adam C. W. A., and Tourtellotte W. N., eds), pp. 413-460. Williams & Wilkins, Baltimore.
Selmaj K. W., Farooq M., Norton W. T., Raine C. S., and Brosnan C. F. ( I 990) Proliferation of astrocytes in vitro in response to cytokines. A primary role for tumor necrosis factor. J. Imrnunol.
Selmaj K., Raine C. S., Cannella B., and Brosnan C. F. (1991) Iden- tification of lymphotoxin and tumor necrosis factor in multiple sclerosis lesions. J. Clin. Invesl. 87, 949-954.
Sensenbrenner M., Devilhers G., Boch E., and Porte A. (1980) Bio- chemical and ultrastructural studies of cultured rat astroglial cells. Differentiation 17, 5 1-6 I .
Shafit-Zagardo B., Brown F. L., Zavodny P. J., and Maio J. J. (1983) Transcription of the KpnI famiIies of Iong interspersed DNAs in human cells. Nature 304, 277-280.
Shafit-Zagardo B., Kuome-Iwaki A., and Goldman J. E. (1988) As- trocyte regulate GFAP mRNA levels by cyclic AMP and protein kinase C-dependent mechanisms. Glia 1, 346-354.
Shaw G. and Kamen R. (1986) A conserved AUU sequence from the 3' untranslated region of GM-CSF mRNA mediates selective mRNA degradation. Cell 46,659-667.
14, 167-176.
144, 129-135.
J. Neurochem.. Val. 57. No. 3, 1991

830 K. SELMAJ ET AL.
Smith M. E., Somera F. P., and Eng L. F. (1983) Immunocytochem- ical staining for glial fibrillary acidic protein and the metabolism of cytoskeletal proteins in experimental autoimmune enceph- alomyelitis. Bruin Rex 264, 241-253.
Smith M. E., Gibbs M. A., Forno L. S., and Eng L. F. (1987) ['HIThymidine labeling of astrocytes in experimental allergic encephalomyelitis. J. Neuroimmunol. 15, 309-32 1.
Soeiro R. and Darnell J . E. (1969) Competition hybridization by presaturation of HeLa cell DNA. J. Mol Biol. 44, 551-562.
Tardy M., Fages C., Riol H., LePrince G., Rataboul P., Chamkre- Bertrand C. , and Nunez J. (1989) Developmental expression of the dial fibrillary acidic protein mRNA in the central nervous system and in cultured astrocytes. J . Neurochem. 52, 162- 167.
Thomas P. S. (1980) Hybridization of denatured RNA and small DNA fragments transferred to nitrocellulose. Proc. Nutl. Acud. Sci. USA 11, 5201-5205.
Topp K. S., Faddis B. T., and Vijayan V. K. (1989) Trauma-induced
proliferation of astrocytes in the brains of young and aged rats. Glia 2, 20 1-2 1 1.
Torti M. T., Dieckmann B., Beutler B., Cerami A,, and Ringold G. M. (1985) A macrophage factor inhibits adipocyte gene expression: an in vitro model of cachexia. Science 229, 867- 869.
Weibel M., Pettman B., Labourdette G., Miehe M., Bock E., and Sensenbrenner M. (1985) Morphological and biochemical mat- uration of rat astroglial cells grown in a chemically defined me- dium: influence of an astroglial growth factor. Int. J. Dev. Neu- rosci. 3, 6 17-630.
Wong G. H. W., Bartlett P. F., Clark-Lewis I. , Battye F., and Schrader J . W. (1984) Inducible expression of H2 and la antigens on brain cells. Nuiure 310, 688-691.
Yarden A. and Kimchi A. (1986) Tumor necrosis factor reduces c- myc expression and cooperates with interferon-gamma in HeLa cells. Science 234, 14 19- 142 I .
J. Neurochem., Vol. 57, No. 3, 1991





























