Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attached
copy is furnished to the author for internal non-commercial research
and education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling or
licensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of the
article (e.g. in Word or Tex form) to their personal website or
institutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies are
encouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Type I interferon (IFN-a/b) rescues B-lymphocytes from apoptosisvia PI3Kd/Akt, Rho-A, NFjB and Bcl-2/BclXL
Gamal Badr a,b,*, Heba Saad c, Hanan Waly a, Khadega Hassan a, Hanem Abdel-Tawab a,Ibrahim M. Alhazza b, Emad A. Ahmed a
a Zoology Department, Faculty of Science, Assiut University, Egyptb Zoology Department, Faculty of Science, King Saud University, Saudi Arabiac Faculty of Medicine, Assiut University, Egypt
a r t i c l e i n f o
Article history:
Received 5 October 2009Accepted 19 February 2010Available online 24 February 2010
Keywords:
ApoptosisB-lymphocytesInterferonPhosphatidylinositol 3-kinasePKB/AktNFjB
a b s t r a c t
Although IFN-awas reported to promote the survival of peripheral B-lymphocytes via the PI3-kinase-Aktpathway, the triggered signalling pathways involved in the protection of B cell from apoptosis need to beclarified. Using flow cytometry and western blot analysis, we have found that type 1 IFNs (IFN-a/b) pro-tect human B cells in culture from spontaneous apoptosis and from apoptosis mediated by anti-CD95agonist, in a dose- and time-dependant manner. IFN-a/b-mediated anti-apoptotic effect on human B cellswas totally abrogated by blockade of IFNR1 chain. Our data indicate that PI3Kd, Rho-A, NFjB and Bcl-2/BclXL are active downstream of IFN receptors and are the major effectors of IFN-a/b-rescued B cells fromapoptosis. Furthermore, immunohistochemical results show marked reduction in numbers of CD20 posi-tive B cell in both spleen and Peyer’s patches from mice treated with anti-IFNR1 blocking antibody com-pared with control group. Moreover, ultrastructural observations of these organs show an obviousincrease in apoptotic cells frommice treated with anti-IFNR1 blocking antibody. Our results provide moredetails about the triggered signalling pathways and the phosphorylation cascade which are involved inthe protection of B cell from apoptosis after treatment with IFN-a/b.
Ó 2010 Elsevier Inc. All rights reserved.
1. Introduction
B cells are lymphocytes that play a large role in the humoralimmune response. The principal function of B cells is to makeantibodies against antigens. Human tonsils are a rich source ofB-lymphocytes exhibiting a variety of phenotypes and activationstates. Tonsillar B-lymphocytes (TBL) died gradually when culturedin vitro and approximately 60% of TBL died within 4 days of culture,and only 2–3% of cells survived after 8 days [1]. In vivo, cytokineswere found to play an important role in regulating the developmentand homeostasis of B cells through controlling their viability [2]. Ofthese cytokines, particular attention has been given to the regula-tory effect of IL-4, IL-13 and interferon alpha (IFN-a) [3–5]). IFN-a
belongs to type I IFNs which are pleiotropic antiviral cytokines pro-duced in response to viral infection [6]. Mammalian type I IFNs con-stitute a multigene family with at least eight subclasses, of whichIFN-a, IFN-b, IFN-x are present in human [7]. Type I IFNs were re-ported to induce apro-apoptotic state inuninfected cells and tohaveadual role in the control of apoptosis through inductionor inhibitionof apoptosis [8,9]. IFN-bwas found to up-regulate the expression ofCD95 (cell death receptor) on B lymphoma cells and to play a role inthe induction of apoptosis [10]. However, the first identified anti-apoptotic proteins, Bcl-2 and BclXL, were discovered in B-cell lym-phoma [11].
The biological effect of IFN-a and IFN-b is initiated upon itsbinding to the IFN type I receptor (IFNAR), which results in the acti-vation of several downstream effector molecules. Recently, T and Bcells were considered to be direct targets for type I IFNs, as theantibody response was greatly impaired in mice with selectivedeletion of IFNAR in T or B cells [12]. Previous experiments haveshown that PI3-kinase is physically associated with IFNAR[13,14]. In both hematopoietic cells and fibroblasts, PI3-kinasewas reported to initiate a phosphorylation cascade involving theserine/threonine kinase Akt leading to both survival and prolifera-tion signals (reviewed in [15,16]). Although IFN-a was reportedto promote the survival of peripheral B-lymphocytes via the
0008-8749/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved.doi:10.1016/j.cellimm.2010.02.012
Abbreviations: AKT/PKB, protein kinase B; Bcl-2, B-cell leukemia 2; DMSO,dimethylsulfoxide; ECL, enhanced chemiluminescence; ERK, extracellular signal-regulated protein Kinase; IFN, interferon; IjB, inhibitor of jB; MEK, mitogen-activated kinase; NFjB, nuclear factor kappa B; PDC, plasmacytoid dendritic cells;PDK1, phosphoinositide-dependent kinase 1; PI3 K, phosphatidylinositol 3-kinase;PIP3, phosphatidylinositol-3,4,5-bisphosphate; PKB, protein kinase B; STAT, signaltransducer and activator of transcription; WM, wortmannin.* Corresponding author. Address: Zoology Department, Faculty of Science, Assiut
University, Egypt.E-mail address: [email protected] (G. Badr).
Cellular Immunology 263 (2010) 31–40
Contents lists available at ScienceDirect
Cellular Immunology
journal homepage: www.elsevier .com/locate /yc imm

Author's personal copy
PI3-kinase-Akt pathway, the triggered signalling pathways in-volved in the protection of B cell from apoptosis need to beclarified.
To the best of the present author’s knowledge, partial informa-tion is available about which class of PI3 K is the activated down-stream IFN-a/b receptor (IFNAR) after IFN-a/b treatment. Thepresent work is an attempt to fulfill this gap using inhibitors of dif-ferent classes of PI3 K and other cytoplasmic effectors and flowcytometry analysis. Then, the present study aims to investigatethe triggered signalling pathways that are involved in the protec-tion of B cell from apoptosis after being treated with IFN-a/b andalso to comprehensively investigate the in vivo effect of IFN-a/bon B cells after the inhibition of IFN-a/b receptor (IFNAR) in micemodel.
2. Materials and methods
2.1. B-cell preparation and culture
B cells were obtained from palatine tonsils of 10 donors as previ-ously described [17]. Briefly, after one cycle of rosette formation,residual T cells and monocytes were depleted with CD2- andCD14-coated magnetic beads (Dynabeads M-450, Dynal AS, Oslo,Norway). The total B-cell population was depleted from CD38 + GCB cells by Percoll gradient separation according to Badr et al. [18].The resulting B-cell population, referred to hereafter as B cells, was98 ± 5.4%CD19; 59 ± 4.9% sIgD + B cells and41 ± 6.3%CD27 + B cells.Viability was tested with Aqua Live/Dead fixable dead cell Stain Kit(BD Biosciences, France) andmore than 98% of cells were viable. Forsome experiments, total tonsillar B cellswere separated into surface(s)IgD+ (naïve B cells) and sIgDÿ populations by being incubated for30 min with saturated amounts of anti-IgD MoAb (TA4-1) and sub-sequent removal of IgDhigh cells from the cell suspension using goatanti-mouse IgG-conjugated magnetic beads (Dynal). Surface IgDÿ Bcells were further separated into CD44high (memory B cells) andCD44low/ÿ (Germinal center) B cells using a similar protocol and sat-urating amounts of anti-CD44 MoAb (BF24, Diaclone). All of thepurification procedures were carried out at 4 °C to prevent sponta-neous apoptosis. Theviability of these cell fractionswas consistentlyhigher than 93%. For in vitro culture assays, purified B cells (2 �
106 cells/ml) were cultured in culture medium (500 ml RPMI 1640supplemented with 5 ml penistreptomycine, 5 ml sodium pirovate,10 ml Hepes and 5 ml non-essential amino acids). These cells wereincubated with or without 100–10 000 IU/ml IFN-a and IFN-b fordifferent incubation hours (0–48 h). In some experiments, B cellswere incubated with different inhibitors including; 100 nM or1 lM wortmannin (wortmannin (WN), PI3 K/PI4 K inhibitor),10 lM IC8711 (PI3 K delta inhibitor), 10 lM PD98059 (PD, mito-gen-activated protein kinase kinase 1/2 (MEK1/2) inhibitor),10 lMSB203580 (P38 inhibitor), 100 nM Y27632 (Rho-A inhibitor),1 lM SN50 (inhibitor of NFjB nuclear translocation), 10 lM SH5(phosphoinositide-dependent protein kinase 1 (PDK1) inhibitor,all fromAlexis, Coger, France or dimethylsulfoxide (DMSO) as a con-trol, for 1 h before being subjected to the IFN treatment.
2.2. Apoptosis detection
Apoptosis was analyzed by Annexin V/propidium iodide. Stain-ing was performed according to the manufacturer’s instructions.Stained cells were analyzed by FACScan flow cytometer.
2.3. Flow cytometry
For flow cytometric analysis, 10 000 viable cells per samplewere analyzed. IFN-a/b receptor expression was analyzed using
PE-conjugated anti-interferon receptor monoclonal antibody(R&D systems, FAB245P, Abingdon, UK) PE-conjugated CD19(IgG1, BD Biosciences, France) and CD5 (IgG1, BD Biosciences,France) monoclonal antibody and FITC- and PE-conjugated mouseisotype-matched control IgG (BD Biosciences, France). For apopto-sis detection, B cells were labeled for surface antigens using PerCP-conjugated CD3 and APC-conjugated CD19 all from BD Biosciences.Then cells were permeabilized using BD permeabilized kits andstained for intracellular proteins using PE-conjugated BclXL (fromGene Tex, CN: GTX46035) and FITC-conjugated Bcl-2 (from BD Bio-sciences). Staining was done according to the manufacturer’s pro-tocol. ‘‘Fluorescence minus one” control stains were used todetermine background levels of staining. FACScan and FACSCaliberflow cytometry were used for data acquisition and CellQuest soft-ware (BD Biosciences) and FlowJo software V8 were used for dataanalysis. For each marker, the threshold of positivity was definedby the non-specific binding observed in the presence of the rele-vant control IgG.
2.4. Western blot analysis
B cells (2 � 106/ml) were incubated in culture medium for 1 h at37 °C, and then were suspended at a density of 1 � 107 cells/ml inpre-warmed RPMI 1640 without FCS and stimulated for 5 min at37 °C with medium, 3000 IU/ml IFN-a and 6000 IU/ml IFN-b. Ly-sates were prepared as previously described [19]. Equal amountsof total cellular protein were subjected to SDS-PAGE and were blot-ted onto nitrocellulose membrane (MilliPore, Bedford, MA, USA).The primary antibodies recognizing phospho-AKT (S473), AKT,phospho-ERK1/2 (T202/Y204), phospho-IjB (S32/36), IjB, phos-pho-P38MAPK (T180/Y182), P38MAPK (all from New England Bio-labs, Beverly, MA, USA) or ERK1/2 (Santa Cruz Biotechnology, SantaCruz, CA, USA) were diluted 1:1000. After incubation with second-ary antibodies conjugated HRP, rabbit anti-mouse (DAKO) or goatanti-rabbit (Santa Cruz Biotechnology), levels of active Rho-A weremeasured by Rho-A activity assay using the Rhotekin Rho BindingDomain protein (Upstate, NY, USA). Lysates were incubated for45 min at 4 °C with 30 ll of Rhotekin (Rho)-binding domain-aga-rose beads under agitation. Beads were washed three times withlysis buffer, re-suspended in Laemmli buffer and analyzed by wes-tern blotting with anti-Rho-A monoclonal antibody (Santa CruzBiotechnology, CA, USA). In some experiments, B cells were incu-bated with different inhibitors including; 100 nM or 1 lM wort-mannin (wortmannin (WN), PI3 K/PI4 K inhibitor), 10 lM IC8711(PI3 K delta inhibitor), 10 lM PD98059 (PD, mitogen-activatedprotein kinase kinase 1/2 (MEK1/2) inhibitor), 10 lM SB203580(P38 inhibitor), 100 nM Y27632 (Rho-A inhibitor), 1 lM SN50(inhibitor of NFjB nuclear translocation), 10 lM SH5 (phosphoin-ositide-dependent protein kinase 1 (PDK1) inhibitor, all fromAlexis, Coger, France or dimethylsulfoxide (DMSO) as a control,for 1 h before being subjected to the IFN stimulation. The antigenswere visualized using chemiluminescence (ECL, SupersignalWestpico chemiluminescent substrate, Perbio, Bezons, France)and exposure to X-ray film (Amersham Biosciences, France). TheECL signal was recorded on ECL hyperfilm. To quantify band inten-sities, films were scanned, saved as TIFF files and analyzed withNIH Image software.
2.5. Immunohistochemistry
Paraffin sections of spleen and Peyer’s patches were preparedfrom control mice and mice injected with mAb antagonist accord-ing to NIH protocol to block type I IFN receptor signaling. Slideswere defrosted and dried at 37 °C for 30 min, dewaxed in Histo-clear (twice for 5 min), and rehydrated in 100% ethanol (twice for5 min), 75% ethanol (5 min) and tapwater (5 min). Endogenous
32 G. Badr et al. / Cellular Immunology 263 (2010) 31–40
Author's personal copy
peroxidase was blocked with 0.3% H2O2 in PBS for 15 min at RT.After washing in PBS, paraffin sections were digested with0.0025% trypsin (Difco; Detroit, MI) in 0.l% calcium chloride, pH7.8, for 5 min, followed by washing in three changes of PBS. Block-ing serum, secondary antibodies, and ABC reagents were obtainedfrom Vector Labs (Peterborough, UK). Non-specific binding siteswere blocked with 1.5% normal rabbit serum in PBS for 30 min atRT. Excess serum was removed, the sections circled with a water-proof pen (Dako; Glostrup, Denmark), and incubated with primaryCD20 MAb at 4 °C overnight. After washing in PBS the slides wereincubated with biotinylated secondary antibody, followed by avi-din-biotinylated horseradish peroxidase complex (ABC) and diam-inobenzidine (Sigma) according to the Vectastain protocol (Vector).Slides were counterstained with hematoxylin solution (DakoCyto-mation), dehydrated, cleared and mounted in synthetic resin (Poly-Mount; Poly Scientific).
2.6. Statistical analysis
Differences between groups were assessed using analysis ofvariance (ANOVA) and SPSS software version 16.
3. Results
3.1. IFN-a/b rescues human B cells from apoptosis in a dose- and time-
dependent manner
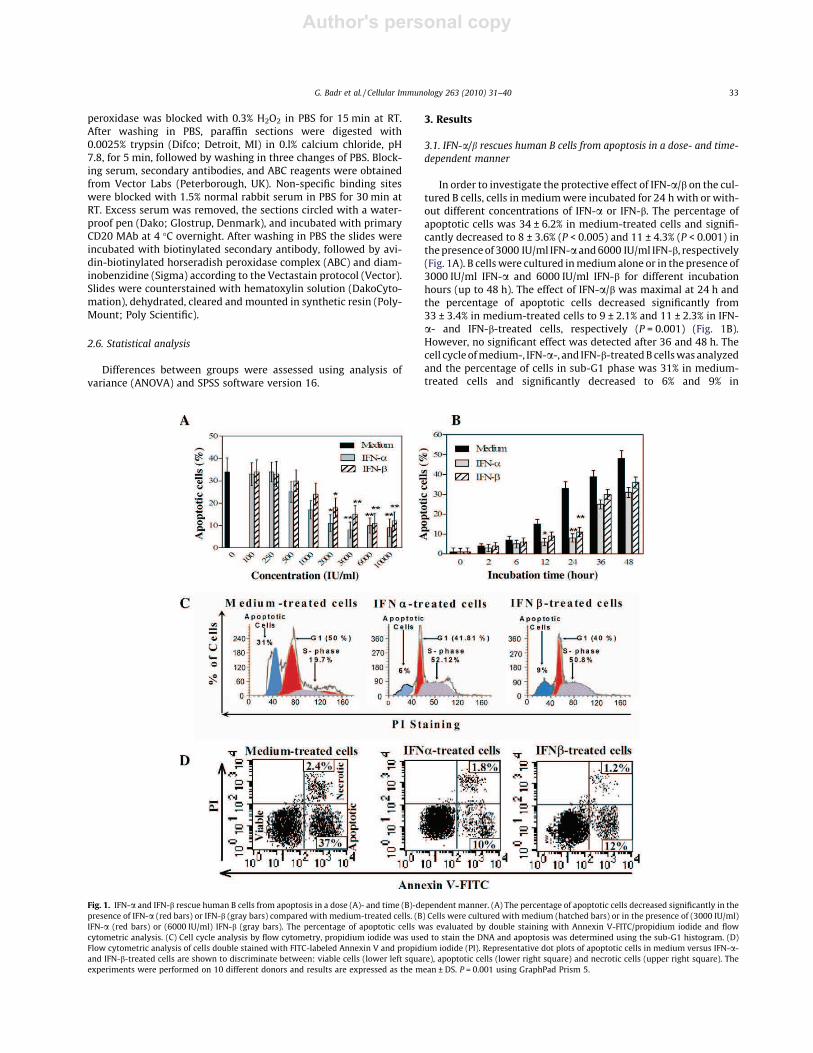
In order to investigate the protective effect of IFN-a/b on the cul-tured B cells, cells in mediumwere incubated for 24 h with or with-out different concentrations of IFN-a or IFN-b. The percentage ofapoptotic cells was 34 ± 6.2% in medium-treated cells and signifi-cantly decreased to 8 ± 3.6% (P < 0.005) and 11 ± 4.3% (P < 0.001) inthepresenceof 3000 IU/ml IFN-a and6000 IU/ml IFN-b, respectively(Fig. 1A). B cellswere cultured inmediumalone or in the presence of3000 IU/ml IFN-a and 6000 IU/ml IFN-b for different incubationhours (up to 48 h). The effect of IFN-a/b was maximal at 24 h andthe percentage of apoptotic cells decreased significantly from33 ± 3.4% in medium-treated cells to 9 ± 2.1% and 11 ± 2.3% in IFN-a- and IFN-b-treated cells, respectively (P = 0.001) (Fig. 1B).However, no significant effect was detected after 36 and 48 h. Thecell cycleofmedium-, IFN-a-, and IFN-b-treatedB cellswasanalyzedand the percentage of cells in sub-G1 phase was 31% in medium-treated cells and significantly decreased to 6% and 9% in
Fig. 1. IFN-a and IFN-b rescue human B cells from apoptosis in a dose (A)- and time (B)-dependent manner. (A) The percentage of apoptotic cells decreased significantly in thepresence of IFN-a (red bars) or IFN-b (gray bars) compared with medium-treated cells. (B) Cells were cultured with medium (hatched bars) or in the presence of (3000 IU/ml)IFN-a (red bars) or (6000 IU/ml) IFN-b (gray bars). The percentage of apoptotic cells was evaluated by double staining with Annexin V-FITC/propidium iodide and flowcytometric analysis. (C) Cell cycle analysis by flow cytometry, propidium iodide was used to stain the DNA and apoptosis was determined using the sub-G1 histogram. (D)Flow cytometric analysis of cells double stained with FITC-labeled Annexin V and propidium iodide (PI). Representative dot plots of apoptotic cells in medium versus IFN-a-and IFN-b-treated cells are shown to discriminate between: viable cells (lower left square), apoptotic cells (lower right square) and necrotic cells (upper right square). Theexperiments were performed on 10 different donors and results are expressed as the mean ± DS. P = 0.001 using GraphPad Prism 5.
G. Badr et al. / Cellular Immunology 263 (2010) 31–40 33
Author's personal copy
IFN-a- and IFN-b-treated cells, respectively (P = 0.0199); however,the percentage of cells in S-phase significantly increased in IFN-a-and IFN-b-treated B cells (Fig. 1C). In dot plots, the percentage ofapoptotic B cells was 37% in medium-treated cells and strongly de-creased to 10% and 12% in IFN-a- and IFN-b-treated cells, respec-tively (P = 0.0099) (Fig. 1D). These results indicate an inhibitoryeffect of IFN-a and IFN-b on spontaneous apoptosis induction in B-lymphocytes.
3.2. IFN-a/b, via IFNAR, protects B cell from spontaneous apoptosis and
from apoptosis mediated by anti-CD95 agonist
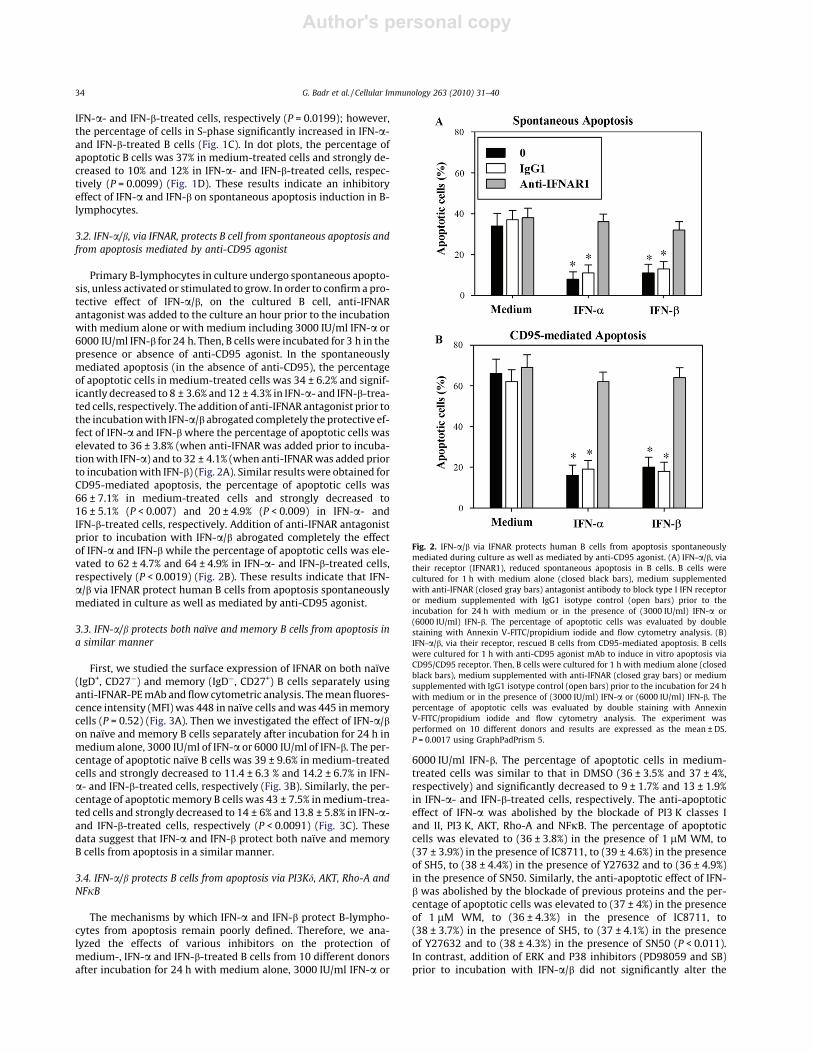
Primary B-lymphocytes in culture undergo spontaneous apopto-sis, unless activated or stimulated to grow. In order to confirma pro-tective effect of IFN-a/b, on the cultured B cell, anti-IFNARantagonist was added to the culture an hour prior to the incubationwith medium alone or with medium including 3000 IU/ml IFN-a or6000 IU/ml IFN-b for 24 h. Then, B cellswere incubated for 3 h in thepresence or absence of anti-CD95 agonist. In the spontaneouslymediated apoptosis (in the absence of anti-CD95), the percentageof apoptotic cells in medium-treated cells was 34 ± 6.2% and signif-icantly decreased to 8 ± 3.6% and 12 ± 4.3% in IFN-a- and IFN-b-trea-ted cells, respectively. The addition of anti-IFNAR antagonist prior tothe incubationwith IFN-a/b abrogated completely the protective ef-fect of IFN-a and IFN-bwhere the percentage of apoptotic cells waselevated to 36 ± 3.8% (when anti-IFNAR was added prior to incuba-tionwith IFN-a) and to 32 ± 4.1% (when anti-IFNARwas added priorto incubationwith IFN-b) (Fig. 2A). Similar results were obtained forCD95-mediated apoptosis, the percentage of apoptotic cells was66 ± 7.1% in medium-treated cells and strongly decreased to16 ± 5.1% (P < 0.007) and 20 ± 4.9% (P < 0.009) in IFN-a- andIFN-b-treated cells, respectively. Addition of anti-IFNAR antagonistprior to incubation with IFN-a/b abrogated completely the effectof IFN-a and IFN-b while the percentage of apoptotic cells was ele-vated to 62 ± 4.7% and 64 ± 4.9% in IFN-a- and IFN-b-treated cells,respectively (P < 0.0019) (Fig. 2B). These results indicate that IFN-a/b via IFNAR protect human B cells from apoptosis spontaneouslymediated in culture as well as mediated by anti-CD95 agonist.
3.3. IFN-a/b protects both naïve and memory B cells from apoptosis in
a similar manner
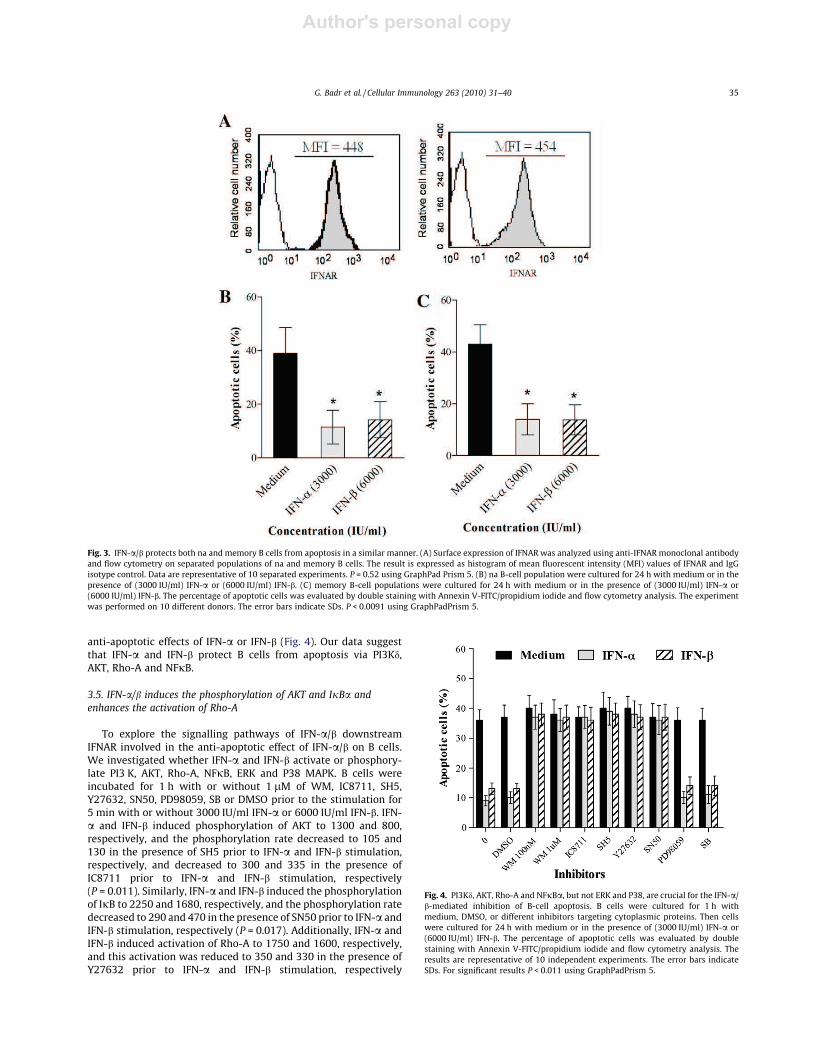
First, we studied the surface expression of IFNAR on both naïve(IgD+, CD27ÿ) and memory (IgDÿ, CD27+) B cells separately usinganti-IFNAR-PEmAb and flow cytometric analysis. Themean fluores-cence intensity (MFI) was 448 in naïve cells andwas 445 inmemorycells (P = 0.52) (Fig. 3A). Then we investigated the effect of IFN-a/bon naïve and memory B cells separately after incubation for 24 h inmedium alone, 3000 IU/ml of IFN-a or 6000 IU/ml of IFN-b. The per-centage of apoptotic naïve B cells was 39 ± 9.6% in medium-treatedcells and strongly decreased to 11.4 ± 6.3 % and 14.2 ± 6.7% in IFN-a- and IFN-b-treated cells, respectively (Fig. 3B). Similarly, the per-centage of apoptoticmemory B cells was 43 ± 7.5% inmedium-trea-ted cells and strongly decreased to 14 ± 6% and 13.8 ± 5.8% in IFN-a-and IFN-b-treated cells, respectively (P < 0.0091) (Fig. 3C). Thesedata suggest that IFN-a and IFN-b protect both naïve and memoryB cells from apoptosis in a similar manner.
3.4. IFN-a/b protects B cells from apoptosis via PI3Kd, AKT, Rho-A and
NFjB
The mechanisms by which IFN-a and IFN-b protect B-lympho-cytes from apoptosis remain poorly defined. Therefore, we ana-lyzed the effects of various inhibitors on the protection ofmedium-, IFN-a and IFN-b-treated B cells from 10 different donorsafter incubation for 24 h with medium alone, 3000 IU/ml IFN-a or
6000 IU/ml IFN-b. The percentage of apoptotic cells in medium-treated cells was similar to that in DMSO (36 ± 3.5% and 37 ± 4%,respectively) and significantly decreased to 9 ± 1.7% and 13 ± 1.9%in IFN-a- and IFN-b-treated cells, respectively. The anti-apoptoticeffect of IFN-a was abolished by the blockade of PI3 K classes Iand II, PI3 K, AKT, Rho-A and NFjB. The percentage of apoptoticcells was elevated to (36 ± 3.8%) in the presence of 1 lM WM, to(37 ± 3.9%) in the presence of IC8711, to (39 ± 4.6%) in the presenceof SH5, to (38 ± 4.4%) in the presence of Y27632 and to (36 ± 4.9%)in the presence of SN50. Similarly, the anti-apoptotic effect of IFN-b was abolished by the blockade of previous proteins and the per-centage of apoptotic cells was elevated to (37 ± 4%) in the presenceof 1 lM WM, to (36 ± 4.3%) in the presence of IC8711, to(38 ± 3.7%) in the presence of SH5, to (37 ± 4.1%) in the presenceof Y27632 and to (38 ± 4.3%) in the presence of SN50 (P < 0.011).In contrast, addition of ERK and P38 inhibitors (PD98059 and SB)prior to incubation with IFN-a/b did not significantly alter the
Fig. 2. IFN-a/b via IFNAR protects human B cells from apoptosis spontaneouslymediated during culture as well as mediated by anti-CD95 agonist. (A) IFN-a/b, viatheir receptor (IFNAR1), reduced spontaneous apoptosis in B cells. B cells werecultured for 1 h with medium alone (closed black bars), medium supplementedwith anti-IFNAR (closed gray bars) antagonist antibody to block type I IFN receptoror medium supplemented with IgG1 isotype control (open bars) prior to theincubation for 24 h with medium or in the presence of (3000 IU/ml) IFN-a or(6000 IU/ml) IFN-b. The percentage of apoptotic cells was evaluated by doublestaining with Annexin V-FITC/propidium iodide and flow cytometry analysis. (B)IFN-a/b, via their receptor, rescued B cells from CD95-mediated apoptosis. B cellswere cultured for 1 h with anti-CD95 agonist mAb to induce in vitro apoptosis viaCD95/CD95 receptor. Then, B cells were cultured for 1 h with medium alone (closedblack bars), medium supplemented with anti-IFNAR (closed gray bars) or mediumsupplemented with IgG1 isotype control (open bars) prior to the incubation for 24 hwith medium or in the presence of (3000 IU/ml) IFN-a or (6000 IU/ml) IFN-b. Thepercentage of apoptotic cells was evaluated by double staining with AnnexinV-FITC/propidium iodide and flow cytometry analysis. The experiment wasperformed on 10 different donors and results are expressed as the mean ± DS.P = 0.0017 using GraphPadPrism 5.
34 G. Badr et al. / Cellular Immunology 263 (2010) 31–40
Author's personal copy
anti-apoptotic effects of IFN-a or IFN-b (Fig. 4). Our data suggestthat IFN-a and IFN-b protect B cells from apoptosis via PI3Kd,AKT, Rho-A and NFjB.
3.5. IFN-a/b induces the phosphorylation of AKT and IjBa and
enhances the activation of Rho-A
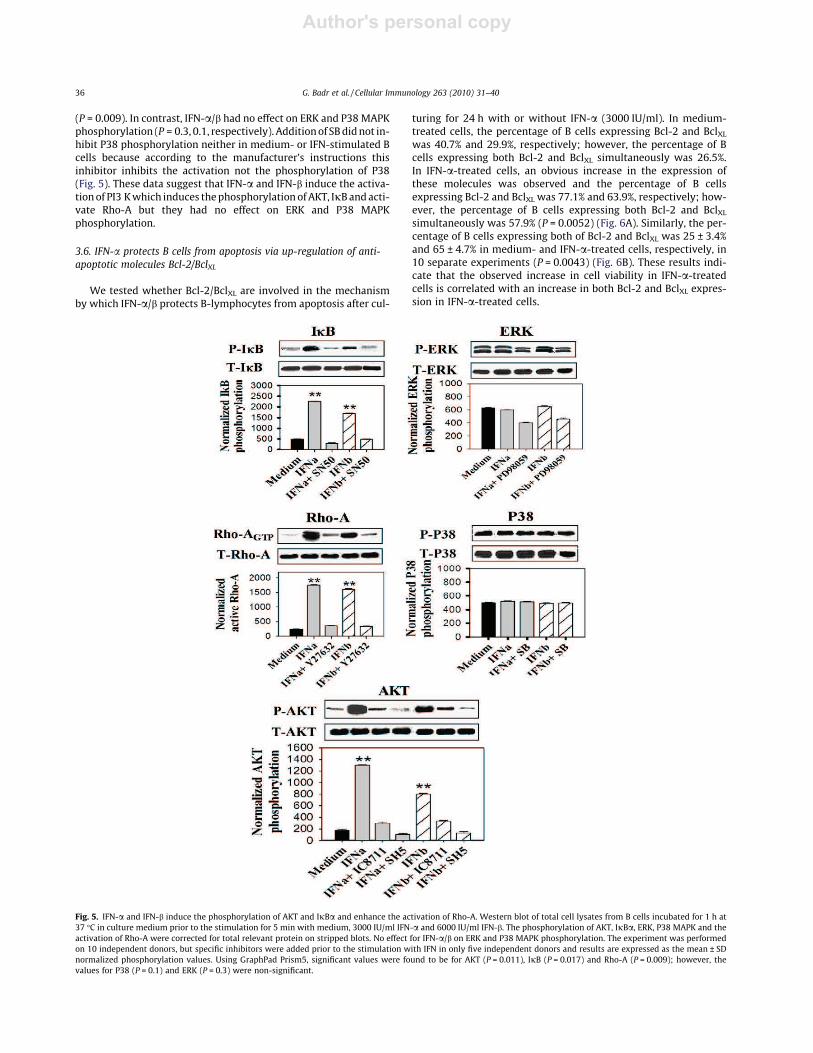
To explore the signalling pathways of IFN-a/b downstreamIFNAR involved in the anti-apoptotic effect of IFN-a/b on B cells.We investigated whether IFN-a and IFN-b activate or phosphory-late PI3 K, AKT, Rho-A, NFjB, ERK and P38 MAPK. B cells wereincubated for 1 h with or without 1 lM of WM, IC8711, SH5,Y27632, SN50, PD98059, SB or DMSO prior to the stimulation for5 min with or without 3000 IU/ml IFN-a or 6000 IU/ml IFN-b. IFN-a and IFN-b induced phosphorylation of AKT to 1300 and 800,respectively, and the phosphorylation rate decreased to 105 and130 in the presence of SH5 prior to IFN-a and IFN-b stimulation,respectively, and decreased to 300 and 335 in the presence ofIC8711 prior to IFN-a and IFN-b stimulation, respectively(P = 0.011). Similarly, IFN-a and IFN-b induced the phosphorylationof IjB to 2250 and 1680, respectively, and the phosphorylation ratedecreased to 290 and 470 in the presence of SN50 prior to IFN-a andIFN-b stimulation, respectively (P = 0.017). Additionally, IFN-a andIFN-b induced activation of Rho-A to 1750 and 1600, respectively,and this activation was reduced to 350 and 330 in the presence ofY27632 prior to IFN-a and IFN-b stimulation, respectively
Fig. 3. IFN-a/b protects both na and memory B cells from apoptosis in a similar manner. (A) Surface expression of IFNAR was analyzed using anti-IFNAR monoclonal antibodyand flow cytometry on separated populations of na and memory B cells. The result is expressed as histogram of mean fluorescent intensity (MFI) values of IFNAR and IgGisotype control. Data are representative of 10 separated experiments. P = 0.52 using GraphPad Prism 5. (B) na B-cell population were cultured for 24 h with medium or in thepresence of (3000 IU/ml) IFN-a or (6000 IU/ml) IFN-b. (C) memory B-cell populations were cultured for 24 h with medium or in the presence of (3000 IU/ml) IFN-a or(6000 IU/ml) IFN-b. The percentage of apoptotic cells was evaluated by double staining with Annexin V-FITC/propidium iodide and flow cytometry analysis. The experimentwas performed on 10 different donors. The error bars indicate SDs. P < 0.0091 using GraphPadPrism 5.
Fig. 4. PI3Kd, AKT, Rho-A and NFjBa, but not ERK and P38, are crucial for the IFN-a/b-mediated inhibition of B-cell apoptosis. B cells were cultured for 1 h withmedium, DMSO, or different inhibitors targeting cytoplasmic proteins. Then cellswere cultured for 24 h with medium or in the presence of (3000 IU/ml) IFN-a or(6000 IU/ml) IFN-b. The percentage of apoptotic cells was evaluated by doublestaining with Annexin V-FITC/propidium iodide and flow cytometry analysis. Theresults are representative of 10 independent experiments. The error bars indicateSDs. For significant results P < 0.011 using GraphPadPrism 5.
G. Badr et al. / Cellular Immunology 263 (2010) 31–40 35
Author's personal copy
(P = 0.009). In contrast, IFN-a/b had no effect on ERK and P38MAPKphosphorylation (P = 0.3, 0.1, respectively). Additionof SBdidnot in-hibit P38 phosphorylation neither in medium- or IFN-stimulated Bcells because according to the manufacturer’s instructions thisinhibitor inhibits the activation not the phosphorylation of P38(Fig. 5). These data suggest that IFN-a and IFN-b induce the activa-tionof PI3 Kwhich induces thephosphorylation ofAKT, IjBandacti-vate Rho-A but they had no effect on ERK and P38 MAPKphosphorylation.
3.6. IFN-a protects B cells from apoptosis via up-regulation of anti-
apoptotic molecules Bcl-2/BclXL
We tested whether Bcl-2/BclXL are involved in the mechanismby which IFN-a/b protects B-lymphocytes from apoptosis after cul-
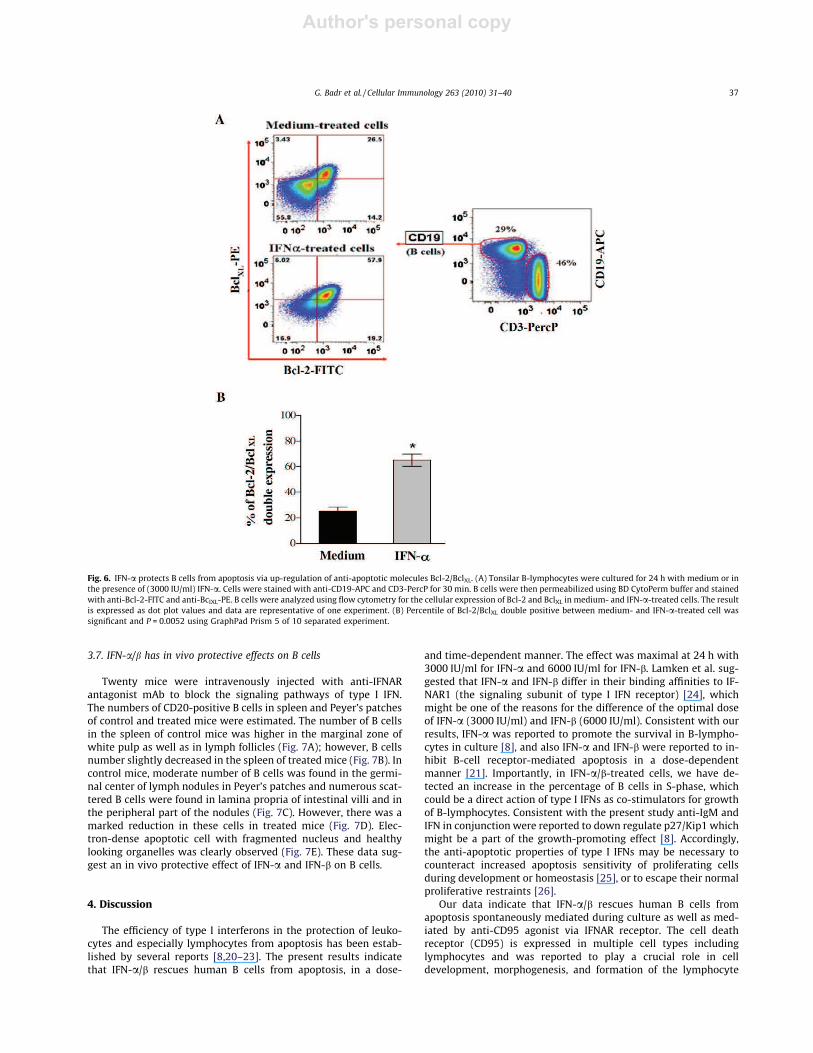
turing for 24 h with or without IFN-a (3000 IU/ml). In medium-treated cells, the percentage of B cells expressing Bcl-2 and BclXLwas 40.7% and 29.9%, respectively; however, the percentage of Bcells expressing both Bcl-2 and BclXL simultaneously was 26.5%.In IFN-a-treated cells, an obvious increase in the expression ofthese molecules was observed and the percentage of B cellsexpressing Bcl-2 and BclXL was 77.1% and 63.9%, respectively; how-ever, the percentage of B cells expressing both Bcl-2 and BclXLsimultaneously was 57.9% (P = 0.0052) (Fig. 6A). Similarly, the per-centage of B cells expressing both of Bcl-2 and BclXL was 25 ± 3.4%and 65 ± 4.7% in medium- and IFN-a-treated cells, respectively, in10 separate experiments (P = 0.0043) (Fig. 6B). These results indi-cate that the observed increase in cell viability in IFN-a-treatedcells is correlated with an increase in both Bcl-2 and BclXL expres-sion in IFN-a-treated cells.
Fig. 5. IFN-a and IFN-b induce the phosphorylation of AKT and IjBa and enhance the activation of Rho-A. Western blot of total cell lysates from B cells incubated for 1 h at37 °C in culture medium prior to the stimulation for 5 min with medium, 3000 IU/ml IFN-a and 6000 IU/ml IFN-b. The phosphorylation of AKT, IjBa, ERK, P38 MAPK and theactivation of Rho-A were corrected for total relevant protein on stripped blots. No effect for IFN-a/b on ERK and P38 MAPK phosphorylation. The experiment was performedon 10 independent donors, but specific inhibitors were added prior to the stimulation with IFN in only five independent donors and results are expressed as the mean ± SDnormalized phosphorylation values. Using GraphPad Prism5, significant values were found to be for AKT (P = 0.011), IjB (P = 0.017) and Rho-A (P = 0.009); however, thevalues for P38 (P = 0.1) and ERK (P = 0.3) were non-significant.
36 G. Badr et al. / Cellular Immunology 263 (2010) 31–40
Author's personal copy
3.7. IFN-a/b has in vivo protective effects on B cells
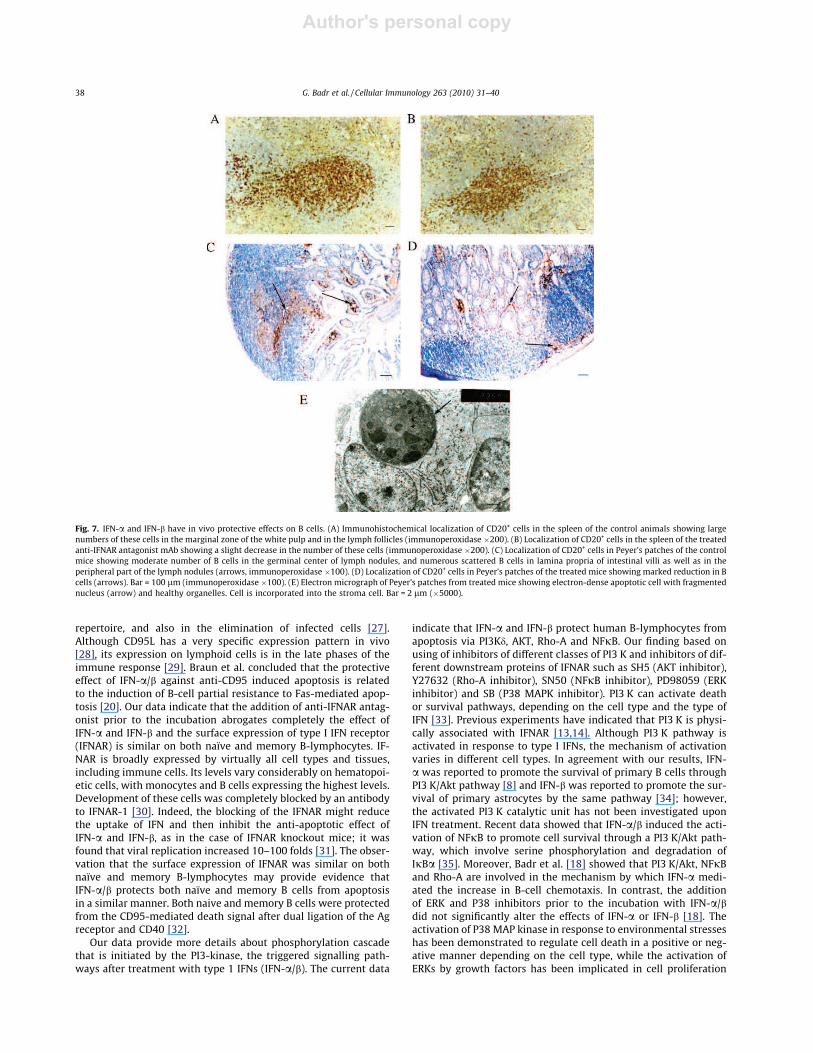
Twenty mice were intravenously injected with anti-IFNARantagonist mAb to block the signaling pathways of type I IFN.The numbers of CD20-positive B cells in spleen and Peyer’s patchesof control and treated mice were estimated. The number of B cellsin the spleen of control mice was higher in the marginal zone ofwhite pulp as well as in lymph follicles (Fig. 7A); however, B cellsnumber slightly decreased in the spleen of treated mice (Fig. 7B). Incontrol mice, moderate number of B cells was found in the germi-nal center of lymph nodules in Peyer’s patches and numerous scat-tered B cells were found in lamina propria of intestinal villi and inthe peripheral part of the nodules (Fig. 7C). However, there was amarked reduction in these cells in treated mice (Fig. 7D). Elec-tron-dense apoptotic cell with fragmented nucleus and healthylooking organelles was clearly observed (Fig. 7E). These data sug-gest an in vivo protective effect of IFN-a and IFN-b on B cells.
4. Discussion
The efficiency of type I interferons in the protection of leuko-cytes and especially lymphocytes from apoptosis has been estab-lished by several reports [8,20–23]. The present results indicatethat IFN-a/b rescues human B cells from apoptosis, in a dose-
and time-dependent manner. The effect was maximal at 24 h with3000 IU/ml for IFN-a and 6000 IU/ml for IFN-b. Lamken et al. sug-gested that IFN-a and IFN-b differ in their binding affinities to IF-NAR1 (the signaling subunit of type I IFN receptor) [24], whichmight be one of the reasons for the difference of the optimal doseof IFN-a (3000 IU/ml) and IFN-b (6000 IU/ml). Consistent with ourresults, IFN-a was reported to promote the survival in B-lympho-cytes in culture [8], and also IFN-a and IFN-b were reported to in-hibit B-cell receptor-mediated apoptosis in a dose-dependentmanner [21]. Importantly, in IFN-a/b-treated cells, we have de-tected an increase in the percentage of B cells in S-phase, whichcould be a direct action of type I IFNs as co-stimulators for growthof B-lymphocytes. Consistent with the present study anti-IgM andIFN in conjunction were reported to down regulate p27/Kip1 whichmight be a part of the growth-promoting effect [8]. Accordingly,the anti-apoptotic properties of type I IFNs may be necessary tocounteract increased apoptosis sensitivity of proliferating cellsduring development or homeostasis [25], or to escape their normalproliferative restraints [26].
Our data indicate that IFN-a/b rescues human B cells fromapoptosis spontaneously mediated during culture as well as med-iated by anti-CD95 agonist via IFNAR receptor. The cell deathreceptor (CD95) is expressed in multiple cell types includinglymphocytes and was reported to play a crucial role in celldevelopment, morphogenesis, and formation of the lymphocyte
Fig. 6. IFN-a protects B cells from apoptosis via up-regulation of anti-apoptotic molecules Bcl-2/BclXL. (A) Tonsilar B-lymphocytes were cultured for 24 h with medium or inthe presence of (3000 IU/ml) IFN-a. Cells were stained with anti-CD19-APC and CD3-PercP for 30 min. B cells were then permeabilized using BD CytoPerm buffer and stainedwith anti-Bcl-2-FITC and anti-BclXL-PE. B cells were analyzed using flow cytometry for the cellular expression of Bcl-2 and BclXL in medium- and IFN-a-treated cells. The resultis expressed as dot plot values and data are representative of one experiment. (B) Percentile of Bcl-2/BclXL double positive between medium- and IFN-a-treated cell wassignificant and P = 0.0052 using GraphPad Prism 5 of 10 separated experiment.
G. Badr et al. / Cellular Immunology 263 (2010) 31–40 37
Author's personal copy
repertoire, and also in the elimination of infected cells [27].Although CD95L has a very specific expression pattern in vivo[28], its expression on lymphoid cells is in the late phases of theimmune response [29]. Braun et al. concluded that the protectiveeffect of IFN-a/b against anti-CD95 induced apoptosis is relatedto the induction of B-cell partial resistance to Fas-mediated apop-tosis [20]. Our data indicate that the addition of anti-IFNAR antag-onist prior to the incubation abrogates completely the effect ofIFN-a and IFN-b and the surface expression of type I IFN receptor(IFNAR) is similar on both naïve and memory B-lymphocytes. IF-NAR is broadly expressed by virtually all cell types and tissues,including immune cells. Its levels vary considerably on hematopoi-etic cells, with monocytes and B cells expressing the highest levels.Development of these cells was completely blocked by an antibodyto IFNAR-1 [30]. Indeed, the blocking of the IFNAR might reducethe uptake of IFN and then inhibit the anti-apoptotic effect ofIFN-a and IFN-b, as in the case of IFNAR knockout mice; it wasfound that viral replication increased 10–100 folds [31]. The obser-vation that the surface expression of IFNAR was similar on bothnaïve and memory B-lymphocytes may provide evidence thatIFN-a/b protects both naïve and memory B cells from apoptosisin a similar manner. Both naive and memory B cells were protectedfrom the CD95-mediated death signal after dual ligation of the Agreceptor and CD40 [32].
Our data provide more details about phosphorylation cascadethat is initiated by the PI3-kinase, the triggered signalling path-ways after treatment with type 1 IFNs (IFN-a/b). The current data
indicate that IFN-a and IFN-b protect human B-lymphocytes fromapoptosis via PI3Kd, AKT, Rho-A and NFjB. Our finding based onusing of inhibitors of different classes of PI3 K and inhibitors of dif-ferent downstream proteins of IFNAR such as SH5 (AKT inhibitor),Y27632 (Rho-A inhibitor), SN50 (NFjB inhibitor), PD98059 (ERKinhibitor) and SB (P38 MAPK inhibitor). PI3 K can activate deathor survival pathways, depending on the cell type and the type ofIFN [33]. Previous experiments have indicated that PI3 K is physi-cally associated with IFNAR [13,14]. Although PI3 K pathway isactivated in response to type I IFNs, the mechanism of activationvaries in different cell types. In agreement with our results, IFN-a was reported to promote the survival of primary B cells throughPI3 K/Akt pathway [8] and IFN-b was reported to promote the sur-vival of primary astrocytes by the same pathway [34]; however,the activated PI3 K catalytic unit has not been investigated uponIFN treatment. Recent data showed that IFN-a/b induced the acti-vation of NFjB to promote cell survival through a PI3 K/Akt path-way, which involve serine phosphorylation and degradation ofIjBa [35]. Moreover, Badr et al. [18] showed that PI3 K/Akt, NFjBand Rho-A are involved in the mechanism by which IFN-a medi-ated the increase in B-cell chemotaxis. In contrast, the additionof ERK and P38 inhibitors prior to the incubation with IFN-a/bdid not significantly alter the effects of IFN-a or IFN-b [18]. Theactivation of P38 MAP kinase in response to environmental stresseshas been demonstrated to regulate cell death in a positive or neg-ative manner depending on the cell type, while the activation ofERKs by growth factors has been implicated in cell proliferation
Fig. 7. IFN-a and IFN-b have in vivo protective effects on B cells. (A) Immunohistochemical localization of CD20+ cells in the spleen of the control animals showing largenumbers of these cells in the marginal zone of the white pulp and in the lymph follicles (immunoperoxidase �200). (B) Localization of CD20+ cells in the spleen of the treatedanti-IFNAR antagonist mAb showing a slight decrease in the number of these cells (immunoperoxidase �200). (C) Localization of CD20+ cells in Peyer’s patches of the controlmice showing moderate number of B cells in the germinal center of lymph nodules, and numerous scattered B cells in lamina propria of intestinal villi as well as in theperipheral part of the lymph nodules (arrows, immunoperoxidase �100). (D) Localization of CD20+ cells in Peyer’s patches of the treated mice showing marked reduction in Bcells (arrows). Bar = 100 lm (immunoperoxidase �100). (E) Electron micrograph of Peyer’s patches from treated mice showing electron-dense apoptotic cell with fragmentednucleus (arrow) and healthy organelles. Cell is incorporated into the stroma cell. Bar = 2 lm (�5000).
38 G. Badr et al. / Cellular Immunology 263 (2010) 31–40

Author's personal copy
[36,37]. The present result is inconsistent with the finding of Ver-ma et al. who provided evidence that IFN-a and IFN-b inducedphosphorylation of the P38 mitogen-activated protein (Map) ki-nase in CD34+-derived primitive human hematopoietic progenitors[38]. In response to viral infection ERK2 has been reported to beactivated by type I IFNs and [39]. However, consistent with ourfinding, it has been shown that IFN-a/b does not induce ERK1/2activation to decrease ligand-induced chemokine receptor inter-nalization [18]. We also suggest that IFN-a protects human B-lym-phocytes from apoptosis through up-regulation of anti-apoptoticproteins expression, such as Bcl-2 and BclXL. In Ramos cells, IFN-a/b was found to inhibit BCR-mediated apoptosis correlated withan increase in the expression of the anti-apoptotic Bcl-2 and BclXLgenes [21]. Moreover, it has been demonstrated that IFN-a up-reg-ulates Bcl-2 expression and protects B-cell chronic lymphocyticleukemia (B-CLL) cells from apoptosis in vitro and in vivo [40].Inconsistent with the present result, Marrack et al. indicated thattype I IFNs keep activated T cell alive without increasing the levelsof Bcl-2 or BclXL [41]. On the other hand, IFNs were suggested to re-press gene expression of anti-apoptotic proteins such as Bcl-2 andBclXL [42]. In order to better understand the role of type I IFN andtheir receptor in the biological development of B-lymphocytes andarchitecture of the immune organs, we have used mAb antagonistto block type I IFN receptor in a mice model according to NIH pro-tocol. Immunohistochemical study showed reduction in numbersof CD20-positive B cells in spleen and Peyer’s patches of treatedanimals. Furthermore, electron microscope investigations con-firmed the increase of apoptosis in B cells in spleen and Peyer’spatches.
The current data indicate that PI3Kd is the downstream IFNAR.When PI3Kd is inhibited by its specific inhibitor (IC8711), the anti-apoptotic effect of IFN-a and IFN-b is completely abolished.Accordingly, IFN-a/b protects both naïve and memory B cells fromapoptosis by similar manner via PI3Kd, AKT, Rho-A and NFjB. Inaddition, IFN-a protect human B cells from apoptosis via increasingthe expression of Bcl-2/BclXL in these cells. To conclude IFN-a andIFN-b have also protective effects on B-lymphocytes in vivo.
Acknowledgments
This research is supported by Faculty of Science, Assiut Univer-sity, Egypt, and King Saud University, Saudi Arabia, through theNobel Laureate collaboration project, number (NLCP-1/2009).Therefore, we thank the Nobel laureate, Prof. Gunter Blobel (Labo-ratory of Cell Biology, Howard Hughes Medical Institute, Rockefel-ler University, New York), the consultant of this project.
References
[1] E. Rosati, R. Sabatini, E. Ayroldi, A. Tabilio, A. Bartoli, S. Bruscoli, C. Simoncelli,R. Rossi, P.J. Marconi, Apoptosis of human primary B lymphocytes is inhibitedby N-acetyl-L-cysteine, Leukoc. Biol. 76 (2004) 152–161.
[2] D.S. Mehta, A.L. Wurster, M.J. Whitters, D.A. Young, M. Collins, M.J. Grusby, IL-21 induces the apoptosis of resting and activated primary B cells, J. Immunol.170 (2003) 4111–4118.
[3] K. Ruuth, L. Carlsson, B. Hallberg, E. Lundgren, Interferon-promotes survival ofhuman primary B-lymphocytes via phosphatidylinositol3-kinase, Biochem.Biophys. Res. Commun. 284 (2001) 583–586.
[4] E.F. Wagner, N. Hanna, L.D. Fast, N. Kouttab, P.R. Shank, A. Vasquez, S. Sharma,Novel diversity in IL-4-mediated responses in resting human naive B cellsversus germinal center/memory B cells, J. Immunol. 165 (2000) 5573–5579.
[5] J. Lømo, H.K. Blomhoff, S.E. Jacobsen, S. Krajewski, J.C. Reed, E.B. Smeland,Interleukin-13 in combination with CD40 ligand potently inhibits apoptosis inhuman B lymphocytes: upregulation of Bcl-xL and Mcl-1, Blood 89 (1997)4415–4424.
[6] A. Battistini, E. Affabris, G. Fiorucci, E.M. Coccia, G. Romeo, G. Marziali, G.B.Rossi, Spectrum of biological activity of interferons, Ann. Ist. Super. Sanita. 26(1990) 227.
[7] S. Pestka, C.D. Krause, M.R. Walter, Interferons, interferon-like cytokines, andtheir receptors, J. Immunol. 202 (2004) 8–32.
[8] K. Ruuth, L. Carlsson, B. Hallberg, E. Lundgren, Interferon-a promotes survivalof human primary B-lymphocytes via phosphatidylinositol 3-kinase, Biochem.Biophys. Res. Commun. 284 (2001) 583–586.
[9] C. Schindler, STATs as activators of apoptosis, Trends Cell Biol. 8 (1998) 97–98.[10] E. Takada, K. Shimo, K. Hata, M. Abiake, Y. Mukai, M. Moriyama, L. Heasley, J.
Mizuguchi, Interferon-induced activation of c-Jun NH2-terminal kinasemediates apoptosis through up-regulation of CD95 in CH31 B lymphomacells, Exp. Cell Res. 304 (2005) 518–530.
[11] S. Luvia, R. Andrea, A. Maribel, F. Luis, Mouse splenic CD4+ and CD8+ T cellsundergo extensive apoptosis during a Plasmodium chabaudi chabaudi asinfection, Parasite Immunol. 23 (2001) 617–626.
[12] A. Le Bon, C. Thompson, E. Kamphuis, V. Durand, C. Rossmann, U. Kalinke, D.F.Tough, Cutting edge: enhancement of antibody responses through directstimulation of B and T cells by type I IFN, J. Immunol. 176 (2006) 2074–2078.
[13] M.H. Heim, The Jak-STAT pathway: cytokine signaling from the receptor to thenucleus, J. Recept. Signal Transduct. Res. 19 (1999) 75–120.
[14] M.F. Van den Broek, U. Müller, S. Huang, R.M. Zinkernagel, M. Aguet, Immunedefense in mice lacking type I and/or type II interferon receptors, Immunol.Rev. 148 (1995) 5–18.
[15] L.G. Guidotti, F.V. Chisari, Noncytolytic control of viral infections by theinnate and adaptive immune response, Annu. Rev. Immunol. 19 (2001)65–91.
[16] C.M. Snapper, T.M. Mclntyre, R. Mandler, L.M. Pecanha, F.D. Finkelman, A. Lees,J.J. Mond, Induction of IgG3 secretion by interferon gamma: a model for T cell-independent class switching in response to T cell-independent type 2 antigens,J. Exp. Med. 175 (1992) 1367–1371.
[17] E.A. Krzysiek Lefèvre, W. Zou, A. Foussat, J. Bernard, A. Portier, P. Galanaud, Y.Richard, Antigen receptor engagement selectively induces macrophageinflammatory protein-1 alpha (MIP-1 alpha) and MIP-1 beta chemokineproduction in human B cells, J. Immunol. 162 (1999) 4455–4463.
[18] G. Badr, G. Borhis, D. Treton, Y. Richard, IFN-alpha enhances human B-cellchemotaxis by modulating ligand-induced chemokine receptor signaling andinternalization, Int. Immunol. 17 (2005) 459–467.
[19] J. Wang, Q. Lin, H. Langston, M.D. Cooper, Resident bone marrow macrophagesproduce type 1 interferons that can selectively inhibit interleukin-7-drivengrowth of B lineage cells, Immunity 3 (1995) 475–484.
[20] D. Braun, I. Caramalho, J. Demengeot, IFN-a/b enhances BCR-dependent B cellresponses, Int. Immunol. 14 (2002) 411–419.
[21] L. Su, M. David, Inhibition of B cell receptor-mediated apoptosis by IFN, J.Immunol. 162 (1999) 6317–6321.
[22] C.H. Yang, A. Murti, S.R. Pfeffer, L. Basu, J.G. Kim, L.M. Pfeffer, IFN-alpha/betapromotes cell survival by activating NF-kappa B, Proc. Natl. Acad. Sci. U. S. A.97 (2000) 13631–13636.
[23] B.D. Stetson, R. Medzhitov, Type I interferons in host defense, Immunity 25(2006) 373–381.
[24] P. Lamken, S. Lata, M. Gavutis, J. Piehler, Ligand-induced assembling of thetype I interferon receptor on supported lipid bilayers, J. Mol. Biol. 341 (2004)303–318.
[25] K. Kobayashi, M. Hatano, M. Otaki, T. Ogasawara, T. Tokuhisa, Expression of amurine homologue of the inhibitor of apoptosis protein is related to cellproliferation, Proc. Natl. Acad. Sci. U. S. A. 96 (1999) 1457–1462.
[26] A.G. Uren, L. Wong, M. Pakusch, K.J. Fowler, F.J. Burrows, D.L. Vaux,K.H. Choo, Survivin and the inner centromere protein INCENP showsimilar cell-cycle localization and gene knockout phenotype, Curr. Biol.10 (2000) 1319–1328.
[27] T. Timmer, E.G. de Vries, S. de Jong, Fas receptor-mediated apoptosis: a clinicalapplication? J. Pathol. 196 (2002) 125–134.
[28] N. Özören, W.S. El-Deiry, Cell surface death receptor signaling in normal andcancer cells, Cancer Biol. 13 (2003) 135–147.
[29] L.P. Van Parijsand, A.K. Abbas, Homeostasis, self-tolerance in the immunesystem: turning lymphocytes off, Science 280 (1998) 243–248.
[30] S.L. Pogue, B.T. Preston, J. Stalder, C.R. Bebbington, P.M. Cardarelli, The receptorfor type I IFNs is highly expressed on peripheral blood B cells and monocytesand mediates a distinct profile of differentiation and activation of these cells, J.Interferon Cytokine Res. 24 (2004) 131–139.
[31] U. Muller, U. Steinhoff, L.F. Reis, S. Hemmi, J. Pavlovic, M.R. Zinkernagel, M.Aguet, Functional role of type I and type II interferons in antiviral defense,Science 264 (1994) 1918–1921.
[32] C. Lagresle, P. Mondière, C. Bella, P.H. Krammer, T. Defrance, Concurrentengagement of CD40 and the antigen receptor protects naive and memoryhuman B cells from APO-1/Fas-mediated apoptosis, J. Exp. Med. 183 (1996)1377–1388.
[33] A.H. van Boxel-Dezaire, M.R. Rani, G.R. Stark, Complex modulation of cell type-specific signaling in response to type I interferons, Immunity 25 (2006) 361–372.
[34] O. Barca, S. Ferre, M. Seoane, J.M. Prieto, M. Lema, R. Senaris, V.M. Arce,Interferon beta promotes survival in primary astrocytes throughphosphatidylinositol 3-kinase, J. Neuroimmunol. 139 (2003) 155–159.
[35] C.H. Yang, A. Murti, L.M. Pfeffer, Interferon induces NF-kappa B-inducingkinase/tumor necrosis factor receptor-associated factor-dependent NF-kappa B activation to promote cell survival, J. Biol. Chem. 280 (2005)31530–31536.
[36] R.J. Davis, Signal transduction by the JNK group of MAP kinases, Cell 103(2000) 239–252.
[37] G.L. Johnson, R. Lapadat, Mitogen-activated protein kinase pathways mediatedby ERK, JNK, and p38 protein kinases, Science 298 (2002) 1911–1912.
G. Badr et al. / Cellular Immunology 263 (2010) 31–40 39

Author's personal copy
[38] A. Verma, D.K. Deb, A. Sassano, S. Uddin, J. Varga, A. Wickrema, L.C.Platanias, Activation of the p38 mitogen-activated protein kinasemediates the suppressive effects of type I interferons and transforminggrowth factor-beta on normal hematopoiesis, J. Boil. Chem. 277 (2002)7726–7735.
[39] M. David, E. Petricoin, C. Benjamin, R. Pine, M.J. Weber, A.C. Larner,Requirement for MAP kinase (ERK2) activity in interferon a- and interferon-stimulated gene expression through STAT proteins, Science 269 (1995) 1721–1723.
[40] A.P. Jewell, C.P. Worman, P.M. Lydyard, K.L. Yong, F.J. Giles, A.H. Goldstone,Interferon-alpha up-regulates bcl-2 expression and protects B-CLL cells fromapoptosis in vitro and in vivo, J. Hematol. 88 (1994) 268–274.
[41] P. Marrack, J. Kappler, T. Mitchell, Type I interferons keep activated T cellsalive, J. Exp. Med. 189 (1999) 521–529.
[42] K.S. Spanaus, R. Schlapbach, A. Fontana, TNF-alpha and IFN-gamma rendermicroglia sensitive to Fas ligand-induced apoptosis by induction of Fasexpression and down-regulation of Bcl-2 and BclxL, Eur. J. Immunol. (1998)4398–4408.
40 G. Badr et al. / Cellular Immunology 263 (2010) 31–40





























