Embed Size (px)
Citation preview

J. Zool., Lond. (1976) 178, 395-412
Underwater predatory behaviour of the American mink (Mustela vison)
TREVOR B. POOLE A N D NIGEL DUNSTONE* Department of Zoology, University College of Wales, Aberystwyth, Dyfed
(Accepted 14 October 1975)
(With 1 plate and 8 figures in the text)
Fishing behaviour of the American mink (Mustela vison Schreber) was investigated in the laboratory. Data were recorded using cine film and tape recorded commentaries. Three species of prey were presented to mink, namely, carp (Cyprinus curpio), goldfish (Curussius uurutus) and minnows (Phoxinus phoxinus).
It was found to be necessary to train ranch-bred mink to enter water and catch fish; young mink appeared to be easier to train than adults. Mink spent 5-20 sec under water when fishing; prey had usually previously been located from an aerial vantage point. Predatory behaviour was highly organized sequentially whereas fish were more prone to indulge in unpredictable stratagems; the behaviour of mink and fish were highly correlated.
The mink’s efficiency in catching fish was related to prey size (smaller individuals being more vulnerable to capture) and shoaling. Minnows, which form highly organized shoals, were less easily caught when present in large numbers; this was not trueof a loosely shoaling species, the carp.
Of the three species of prey presented, vulnerability to capture took the form goldfish> carp> minnow; these differences, however, may have been influenced by the fish’s previous experience of underwater predators.
Contents
Introduction.. . . . . . . . . . . Materials and methods . . . . . . . . Results . . . . . . . . . . . . Discussion . . . . . . . . . . . . Summary .. . . . . . . . . . . References . . . . . . . . . . . . Appendix A . . . . . . . . . .
Page . . . . . . . . . . 395 . . . . . . . . . . 396 . . . . . . . . . . 398 . . . . . . . . . . 409 . . . . . . . . . . 411 . . . . . . . . . . 411 . . . . . . . . . . 412
Introduction Underwater hunting behaviour of amphibious animals is a subject which has been
neglected; the only previous account which has been published is that of Erlinge (1968) on the otter, Lutra lutra. Erlinge gave an outline description of the otter’s hunting behaviour ; he compared rates of capture of different prey species and the effects of impaired motility of the prey on the otters’ hunting success.
Benzie (1965) and Neil1 & Cullen (1974) have investigated the responses of fish to some aquatic predators but the present study appears to be the first to consider the behaviour of fish when hunted by an amphibious predator.
* Present address: Department of Zoology. University College of North Wales, Bangor, Gwynedd, U.K. 395

396 T. B . POOLE AND N . DUNSTONE
There have been many descriptions of the predatory behaviour of terrestrial mustelids (e.g. Eibl-Eibesfeldt, 1956; Wustehube, 1960; Apfelbach, 1973) but the detection and capture of prey underwater creates special problems, for example, olfaction involving the inhalation of air is not possible, the frequency response of the ear is changed and, unless a special mechanism has been evolved, a land mammal’s vision under water becomes strongly hypermetropic. Compensatory mechanisms allowing good underwater vision have been shown to exist in a number of mammals including the otter (Walls, 1942). The mink, however, possesses an eye whose visual acuity in water (31 min of arc) is inferior to that in air (15 min of arc) (see Sinclair, Dunstone & Poole, 1974).
Mink appear to show no special adaptations to an aquatic life other than behavioural ones, but fish remains are found in 30-70 % of the faeces of wild mink (Gerell, 1967, 1968 ; Erlinge, 1969; Day & Linn, 1972) so that aquatic hunting clearly plays an important role in the life of this species. Gerell(l968) has shown that, in the winter months, the propor- tion of fish in the mink’s diet increases; he puts forward the hypothesis that the homo- iothermic mink has an advantage over its poikilothermic prey under cold environmental conditions. The ability to dive and swim under water is its main behavioural adaptation to predation on live fish. In its exploratory behaviour, the mink treats water as a three dimensional environment whereas the closely related polecat M. putorius, for example, simply treats water as a surface over which it can swim (Poole, personal observation).
The aim of the experiments to be described was to analyse the hunting behaviour of the mink and the responses shown by various species of fish on which it preys.
Materials and methods The subjects of the experiments were 6 American mink (Mustela vison Schreber) of the standard
colour variety, which possesses normally pigmented eyes. One of the mink was wild-caught as an adult on the River Nevern, Pembrokeshire; the other 5 were offspring of ranch-bred parents and were hand reared from the age of 6 weeks. Mink were housed outdoors in galvanized wire mesh cages 1 x 0.5 x 0.5 m fitted with a wooden nestbox 0.3 x 0.3 x 0.3 m ; they were fed daily on a mixture of 50 g “Pelsifood” artificial mink diet and 50 g “Spratts ZF6”; water was provided ad libit urn.
Minnows (Phoxinus phoxinus), carp (Cyprinus carpio) and goldfish (Curassius aurutus) were used as prey; the minnows and carp were caught locally whilst the goldfish were obtained from a dealer. The fish were kept in the laboratory in running water at 17°C for a minimum of 2 weeks prior to an experiment. All the minnows and goldfish used were between 8 and 10 cm in length.
Experiments were carried out in an indoor arena 5 x 3 m within a windowless room under an ambient illumination level of 40 mL.
Two methods were used to record the animals’ behaviour, either tape-recorded commentaries or 16 mm cine film. When making a tape recorded commentary the observer viewed the animals from an adjacent room through a one-way glass window 2 x 1 m. When filming the mink, how- ever, the experimenter was in the same room as the mink but separated from them by a 1.5 m high wooden partition. Arriflex ST16 and Bolex HI6 cine cameras, with variable speed motors, were used and run at either 24 f.p.s. or 64 f.p.s.; both colour (Ektachrome 7242) and mono- chrome (Kodak Tri-X) cind film were used.
Frame by frame analysis of cine film was carried out using an LW224 16 mm analyser pro- jector; for each frame of cint film the behaviour of the animals (mink and fish) was ascribed to a category which was given a code number. In this way the behaviour of the mink was represented by an array M (1 : number of frames) and that of the fish by another array F (1 : number of frames) where the number of frames was equal to the number of the last frame of cine film in

U N D E R W A T E R P R E D A T I O N B Y M I N K 391
which the animals interacted. These arrays were analysed on an ICL 4130 computer (see Poole, 1973). Cine film exposed at 64 f.p.s. was used to estimate the relative speeds of the predator and prey and their manoeuvrability.
Preliminary training Since ranch-bred mink had had no opportunity to swim and catch prey the subjects of the
experiment were trained to catch fish. The first stage of training involved handling the 8-week old animals and allowing them to become familiar with the arena situation.
The mink were then trained to retrieve objects, such as corks and cotton reels. Initially, these were thrown across the arena floor, later, they were placed on the surface of the water and finally by weighting the objects the animals were trained to retrieve them from the bottom of the tank. In most cases no reinforcement was given, the stimulus for retrieval being provided by the moving object itself; one individual (Argo), however, initially retrieved only objects which had been lightly smeared with food (ZF6). Subsequently the animals were encouraged to enter a polythene tank 1.2 x 0.9 x 0.3 m filled to a depth of 10 cm with water to retrieve the same objects. Ramps were provided so that the mink could enter the water gradually since previous experience had shown the animals to be reluctant to enter the tank because of its steep sides. The water level was gradually increased to a depth of 0.3 m. Three littermates, Neptune, Nautilus and Argo, were trained as a group and frequently became involved in boisterous social play in the water.
The final stage in training involved the use of a large transparent “Perspex” tank 1.8 m long x 0.45 m deep x 0.38 m wide. The tank was positioned on a controlled contrast green base for photographic purposes. Ramps were provided at both ends and a 6 cm wide shelf bounded the tank edge. Goldfish were released into the tank and the mink entered the water to catch the prey,
The mink was released into the arena for a maximum of 1 h per day. At least 12 h prior to the introduction of the mink a goldfish had been placed in the tank; after the mink had captured the fish it was allowed to eat it and the experiment then terminated.
The animals were tested prior to their normal daily feeding time but had not otherwise been deprived of food. Detailed records of the behaviour of 2 mink (Argo and Neptune) were made.
The adult wild caught mink, Acis, was induced to enter the water to catch fish only after partial food deprivation, a method which had been previously used for adult ranch-bred mink which had been difficult to train. The method involved giving the animals 50 % of the maintenance diet for 2 days prior to their release into the arena with a shallow tank containing fish. The mink was left in the arena for up to 8 h, after which time, if no fish had been removed, the animal was returned to its home cage and for 2 days was fed the full diet. The process was continued until the mink captured a fish, after which it was allowed to gain experience in the larger tank.
Fish-catching experiments Two mink experienced in fish-catching (Nautilus and Neptune) were released singly into the
arena; underwater predation during 15 encounters each of 15 min duration was filmed for each of the 2 animals over 2 consecutive days. Single goldfish were used as the prey. Encounters took place in the “Perspex” tank; filming began when the mink dived and stopped after the mink had removed the prey or left the tank; out of tank activities were also recorded.
Nautilus was also filmed in a further 10 encounters with minnows. The behaviour of the wild-caught mink was also observed. This individual was too easily disturbed to allow filming but a tape-recorded commentary was made of its behaviour.
Fish-catching efficiency Two mink, Sven and Siren, were tested in a large polythene lined tank 2 m long x 1.5 m wide x
1.5 m deep containing 1,5,10,15 or 20 minnows or carp 8-10 cm in length. The number of fish to

398 T. B. POOLE AND N. DUNSTONE
be presented was chosen at random for every trial. One trial was carried out per day with each prey species but in all cases the fish were allowed approximately 12 h to settle down in the tank; naive fish were used for each trial. A mink was released in the arena and the experiment was terminated when the mink had caught one fish; each experiment was replicated 5 times.
To investigate possible effects of size of prey on the mink's predatory behaviour a further series of experiments was carried out using a size range of carp from 19-30 cm (weighing 105- 368 g). Under this paradigm the carp were released singly into the tank after being measured and weighed. The carp were allowed to settle down for 2 h prior to a trial. Captured fish were removed by the experimenter, killed and fed to the mink after the day's experiments. Two trials per animal per day were carried out.
The water temperature remained almost constant throughout the experiments (range 15"-18"C) and the water was changed daily. The tank was filled with tap water but there was no evidence that the degree of chlorination was sufficient to affect the behaviour of either mink or fish. The ambient illumination at the water surface was 40 mL.
In both of the above experimental situations data were recorded using a tape recorder.
Results Description of behaviour patterns of mink
Non-predatory behaviour Mink spent a considerable amount of time in all experimental sessions in exploratory
behaviour' investigating the arena, experimental tank and other equipment. Objects were scent marked and occasionally re-marked after swimming. Autogrooming and eating were observed, the latter being dependent upon the mink's success in catching fish.
On tank activities Peering:
Tank walking :
Head dipping :
crouching on the edge of the tank and looking into the water (see Plate I(a)). walking round the edge of the tank whilst following the prey's movements. immersing head and shoulders in the water whilst maintaining a grip on the tank with the hind feet; often accompanied by scanning movements of the head.
In tank activities Diving: launching the entire body head first into the water at a steep angle
(see Plate I(b)) ; three categories of diving were distinguished namely, near fish dive, remote dive and pursuit dive. Remote dives were not orientated to the prey, near fish dives were in the vicinity of a station- ary fish whereas pursuit dives were orientated towards fleeing fish. traversing the tank whilst systematically exploring the bottom and corners of the tank accompanied by sideways movements of the head. The eyes are open and vibrissae extended; speed of swimming is slower than when in pursuit of moving prey.
Search :
PLATE I. (a) Mink peering into the tank. (b) Mink diving in the vicinity of the fish. (c) Mink advances in pursuit of the fish which has just made a sharp turn. (d) Mink captures the fish. (e) Mink retrieving the fish which, in this instance, is held by the tail.
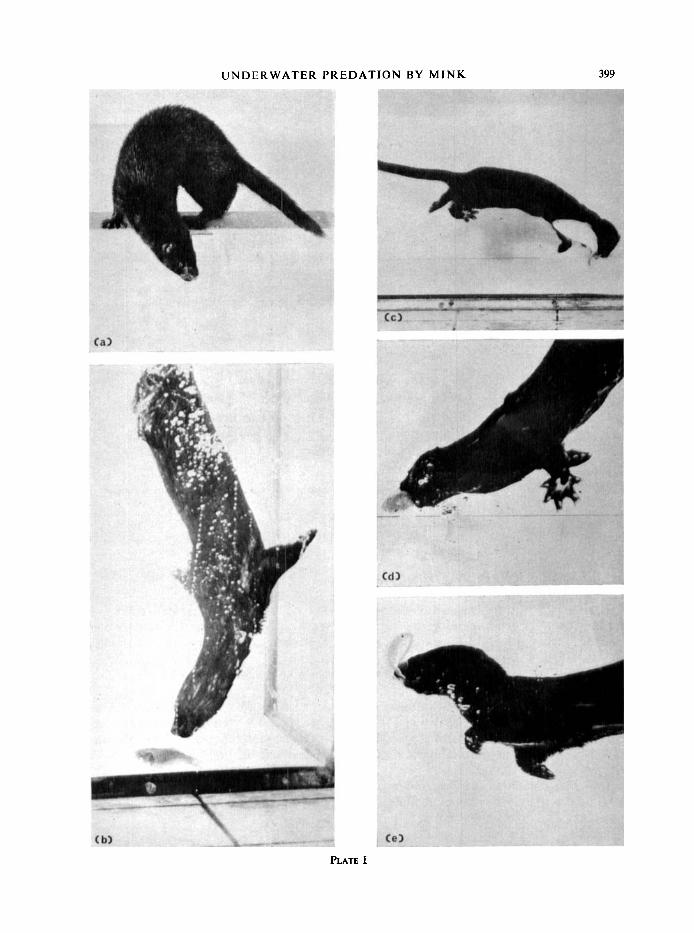
U N D E R W A T E R P R E D A T I O N B Y M I N K 399
PLATE I

400
Advance :
Orientation:
Snap :
Lose fish: Capture:
T. B . POOLE A N D N . DUNSTONE
approaching and pursuing the prey (see Plate I(c)), identifiable on cine film because the mink's behaviour is constantly orientated to its Prey. the point at which search ceased and advance commenced. Recogniz- able on cine film because the animal ceases scanning movements and increases its speed. A duration of one frame of cinC film (1/24 sec) is ascribed to this behaviour pattern. an unsuccessful attempt to capture a fish. The jaws of the mink open and shut, but the fish is not secured. the mink loses sensory contact with its prey. seizing prey in the mouth (see Plate I(d)). The fish are usually bitten behind the head; sometimes however they are grasped by head or tail (see Plate I(e)). Occasionally the mink uses its paws to manipulate fish. 45 % of captures were in tank corners. The fish is removed from the water by the mink. fish eaten on land immediately after capture. Devour:
Description of behaviour patterns of $sh Inactive: goldfish were more often inactive than minnows. Level flight: rapid horizontal swimming in short bursts. Zig-zag flight: alternating low amplitude turns to left and right with an overall
forward direction. Sharp turn : left or right lateral turn or complete 180" turn by the fish. Vertical flight or jump: upwards or downwards movement of the fish carrying it above or
below the predator. Detection: the point at which a fish was observed to be responding to the
presence of the predator; the two criteria utilized were visual fixation and/or an increased rate of fin flickering (Benzie, 1965).
Organization of predatory behaviour There was considerable variation in the length of time taken to train mink to catch fish,
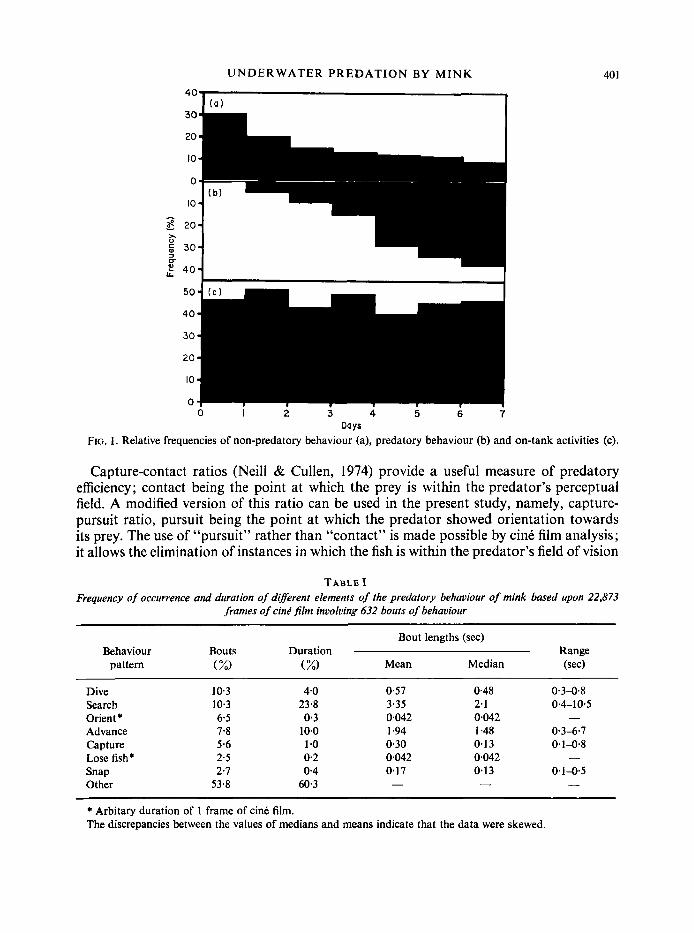
ranging from 2-94 h experience in the arena (see Appendix A). The whole course of training was recorded for 2 mink (Neptune and Argo) and analysed in detail; both mink had previously had some experience of swimming (see Fig. 1). During training a n increase in predatory activity accompanied by a decrease in non-predatory behaviour took place whereas on-tank activity remained approximately constant.
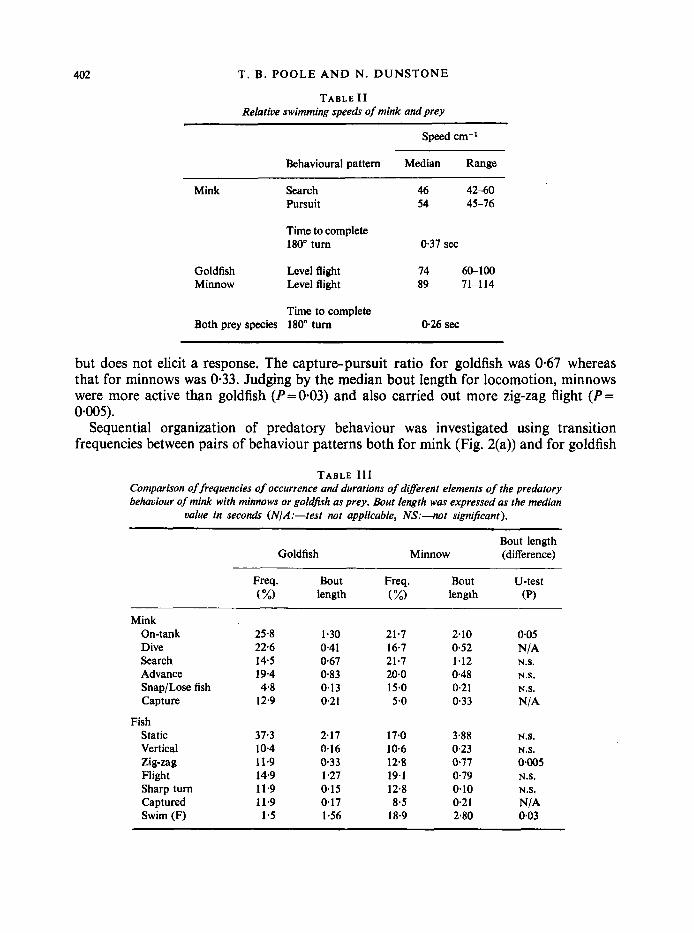
The frequency of occurrence (number of bouts) and duration of the different elements of predatory behaviour are shown in Table I; relative speeds of the predator and fish prey are given in Table 11. The results are based on 30 filmed encounters between mink (Neptune and Nautilus) and goldfish. Depending upon the behaviour of the prey, mink remained under water for 5-20 sec.
Table I11 compares the behaviours of one mink (Nautilus) with two different prey species; the predator spent more time carrying out on-tank activities with minnows as prey than with goldfish (P = 0-05).
UNDERWATER P R E D A T I O N BY M I N K 40 1
0 I 2 3 4 5 6 7 Days
FIG. I. Relative frequencies of non-predatory behaviour (a), predatory behaviour (b) and on-tank activities (c).
Capture-contact ratios (Neil1 & Cullen, 1974) provide a useful measure of predatory efficiency; contact being the point at which the prey is within the predator’s perceptual field. A modified version of this ratio can be used in the present study, namely, capture- pursuit ratio, pursuit being the point at which the predator showed orientation towards its prey. The use of “pursuit” rather than “contact” is made possible by cinC film analysis; it allows the elimination of instances in which the fish is within the predator’s field of vision
TABLE I Frequency of occurrence and duration of different elements of the predatory behaviour of mink based upon 22,873
frames of cint!film involving 632 bouts of behaviour
Bout lengths (sec) Behaviour Bouts Duration Range
pattern ( %) ( %) Mean Median (set)
Dive 10.3 4.0 0.57 0.48 0 , 3 4 8 Search 10.3 23.8 3.35 2.1 0’4-10.5 Orient * 6.5 0.3 0.042 0.042 - Advance 7.8 10.0 1 *94 1.48 0.3-6.7 Capture 5.6 1 *o 0.30 0.1 3 0 .14.8 Lose fish* 2.5 0.2 0.042 0.042 - Snap 2.7 0.4 0.17 0.1 3 0.1-0.5 Other 53.8 60.3
* Arbitary duration of 1 frame of cine film. The discrepancies between the values of medians and means indicate that the data were skewed.
- - -
402 T . B. POOLE A N D N. D U N S T O N E
TABLE I1 Relative swimming speeds of mink and prey
Speed cm-I
Behavioural pattern Median Range
Mink Search Pursuit
46 42-60 54 45-76
Time to complete 180" turn 0.37 sec
Goldfish Level flight 74 6&100 Minnow Level flight 89 71-1 14
Time to complete Both prey species 180" turn 0.26 sec
but does not elicit a response. The capturepursuit ratio for goldfish was 0.67 whereas that for minnows was 0.33. Judging by the median bout length for locomotion, minnows were more active than goldfish (P=0.03) and also carried out more zig-zag flight (P= 0.005).
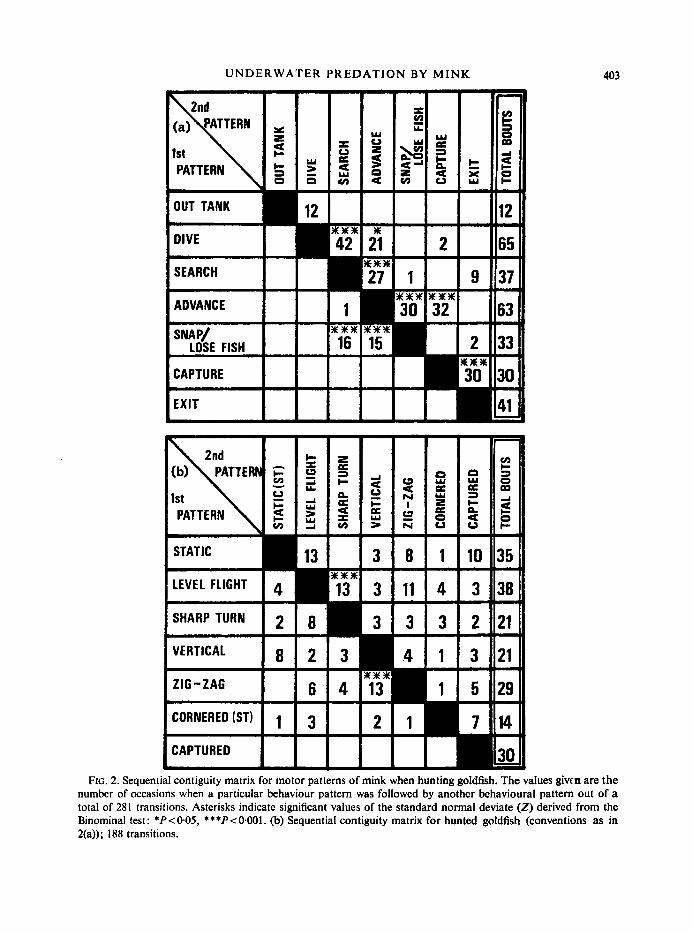
Sequential organization of predatory behaviour was investigated using transition frequencies between pairs of behaviour patterns both for mink (Fig. 2(a)) and for goldfish
TABLE 111 Comparison of frequencies of occurrence and durations of different elements of the predatory behaviour of mink with minnows or goldfish as prey. Bout length was expressed as the median
value in seconds (N/A:-test not applicable, NS:-not significant).
Bout length Goldfish Minnow (difference)
Freq. Bout Freq. (%) length ( %)
Mink On-tank 25.8 1.30 21.7 Dive 22.6 0.41 16.7 Search 14.5 0.67 21.7 Advance 19.4 0.83 20.0 Snap/Lose fish 4 8 0.13 15.0 Capture 12.9 0.21 5.0
Bout length
U-test (PI
2.10 0.52 1.12 0.48 0.21 0.33
0.05 N/A N.S. N.S. N.S. N/A
Fish Static 37.3 2.17 17.0 3.88 N.S.
Zig-zag 11.9 0.33 12.8 0.77 0.005 Vertical 10.4 0.16 10.6 0.23 N.S.
Flight 14.9 1 a27 19.1 0.79 N.S. Sharp turn 11.9 0.15 12.8 0.10 N.S. Captured 11.9 0.17 8.5 0.21 N /A Swim (F) 1.5 1.56 18.9 2-80 0.03
UNDERWATER PREDATION B Y MINK
SHARP TURN I 2 I 8 VERTICAL 1 8 I 2
Z I G - Z A G I 1 6 1 I
CAPTURED I I
$ 3 O N * I a W E > N
3 8
3 11
3 3
2 1
403
FIG. 2. Sequential contiguity matrix for motor patterns of mink when hunting goldfish. The values givcn are the number of occasions when a particular behaviour pattern was followed by another behavioural pattern out of a total of 281 transitions. Asterisks indicate significant values of the standard normal deviate (Z) derived from the Binominal test: *P< 0.05, ***P<O.OOl. (b) Sequential contiguity matrix for hunted goldfish (conventions as in 2(a)) ; 188 transitions.
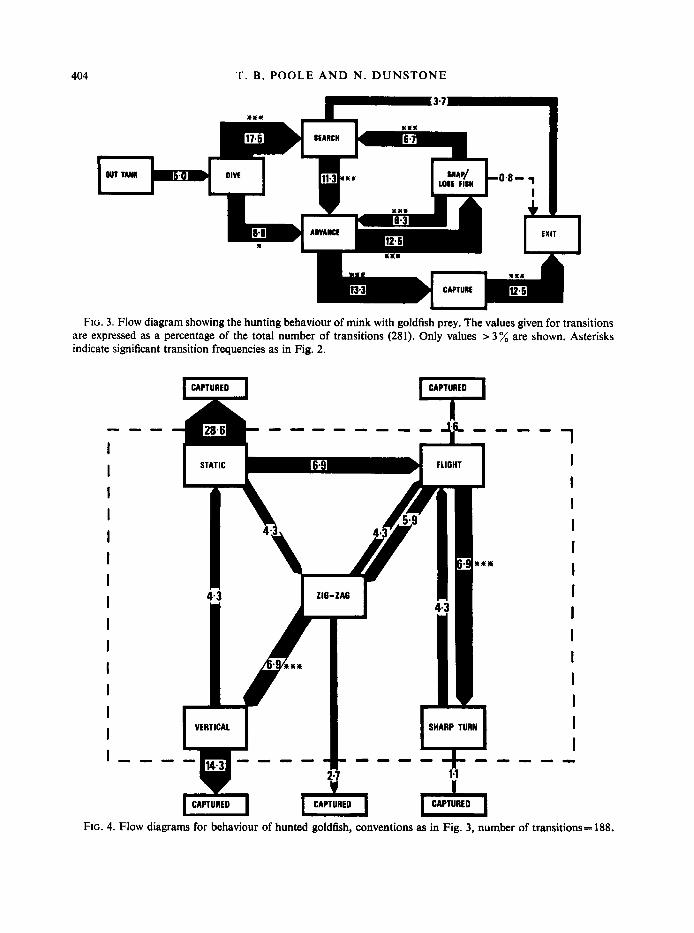
404 T. B. POOLE A N D N. D U N S T O N E
- b 12 6 I I
CAPTURE -L FIG. 3. Flow diagram showing the hunting behaviour of mink with goldfish prey. The values given for transitions
are expressed as a percentage of the total number of transitions (281). Only values > 3 % are shown. Asterisks indicate significant transition frequencies as in Fig. 2.
I STATIC 6.9 FLIGHT I I I
I I I I I I I I
' z'G-zAG I
I VERTICAL I I I - , , ,
CAPTURED
d
SHARP TURN u
I I I I I I I I I I I I I
FIG. 4. Flow diagrams for behaviour of hunted goldfish, conventions as in Fig. 3, number of transitions= 188.
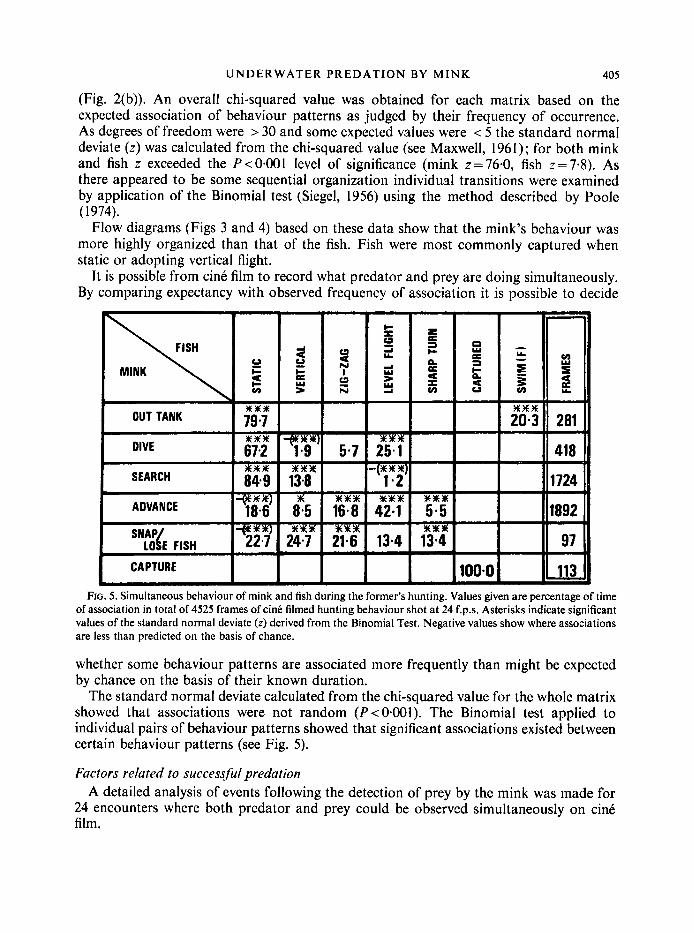
UNDERWATER PREDATION B Y MINK 405
(Fig. 2(b)). An overall chi-squared value was obtained for each matrix based on the expected association of behaviour patterns as judged by their frequency of occurrence. As degrees of freedom were > 30 and some expected values were < 5 the standard normal deviate (z) was calculated from the chi-squared value (see Maxwell, 1961); for both mink and fish z exceeded the P<O.OOl level of significance (mink z=76.0, fish z=7.8). As there appeared to be some sequential organization individual transitions were examined by application of the Binomial test (Siege], 1956) using the method described by Poole (1 974).
Flow diagrams (Figs 3 and 4) based on these data show that the mink’s behaviour was more highly organized than that of the fish. Fish were most commonly captured when static or adopting vertical flight.
It is possible from cinC film to record what predator and prey are doing simultaneously. By comparing expectancy with observed frequency of association it is possible to decide
FIG. 5. Simultaneous behaviour of mink and fish during the former’s hunting. Values given are percentage of time of association in total of 4525 frames of cine filmed hunting behaviour shot at 24 f.p.s. Asterisks indicate significant values of the standard normal deviate (z) derived from the Binomial Test. Negative values show where associations are less than predicted on the basis of chance.
whether some behaviour patterns are associated more frequently than might be expected by chance on the basis of their known duration.
The standard normal deviate calculated from the chi-squared value for the whole matrix showed that associations were not random (P<O.OOl). The Binomial test applied to individual pairs of behaviour patterns showed that significant associations existed between certain behaviour patterns (see Fig. 5).
Factors related to successful predation A detailed analysis of events following the detection of prey by the mink was made for
24 encounters where both predator and prey could be observed simultaneously on cinC film.
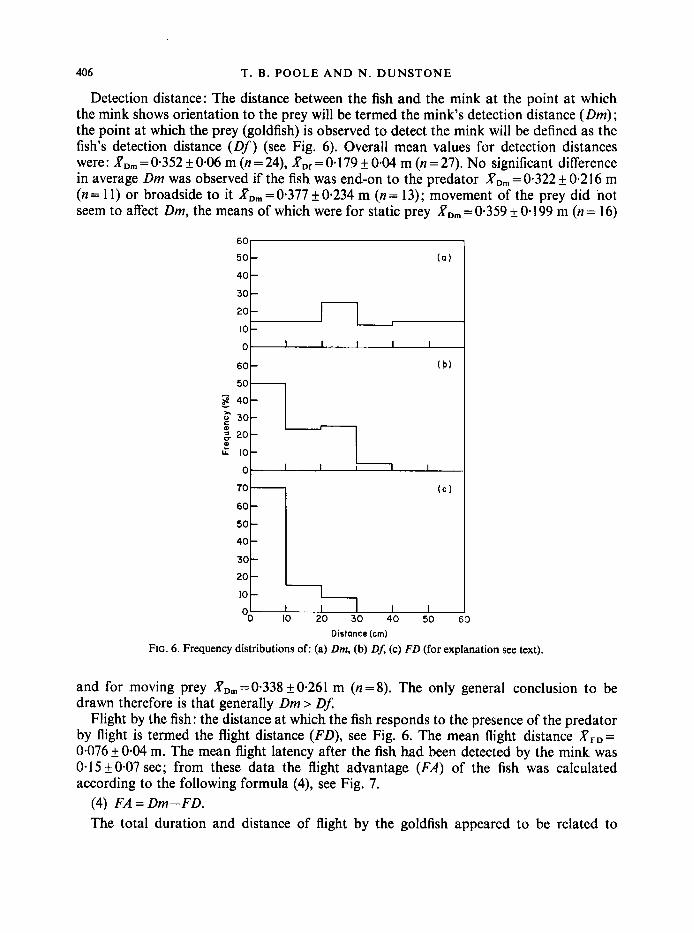
406 T. B . POOLE A N D N . DUNSTONE
Detection distance: The distance between the fish and the mink at the point at which the mink shows orientation to the prey will be termed the mink’s detection distance ( D m ) ; the point at which the prey (goldfish) is observed to detect the mink will be defined as the fish’s detection distance ( O f ) (see Fig. 6). Overall mean values for detection distances were : R,, = 0.352 & 0.06 m (n = 24), X,, = 0.1 79 f 0.04 m (n = 27). No significant difference in average Dm was observed if the fish was end-on to the predator XD, = 0.322 & 0.2 16 m (n = 1 1) or broadside to it .PD, = 0.377 f 0.234 m (n = 13); movement of the prey did not seem to affect Dm, the means of which were for static prey RD, = 0.359 f 0.1 99 m (n = 16)
60
50 40
30
20
10
30
- - - - - - I I
50 I
FIG. 6 . Frequency distributions of: (a) Dm, (b) DA (c) FD (for explanation see text).
and for moving prey RD,=0.338&0*261 m (n=8). The only general conclusion to be drawn therefore is that generally Dm > DJ
Flight by the fish: the distance at which the fish responds to the presence of the predator by flight is termed the flight distance (FD), see Fig. 6. The mean flight distance RpD= 0.076 f 0.04 m. The mean flight latency after the fish had been detected by the mink was 0.15 f 0.07 sec; from these data the flight advantage (FA) of the fish was calculated according to the following formula (4), see Fig. 7.
(4) FA= Dm-FD. The total duration and distance of flight by the goldfish appeared to be related to
UNDERWATER PREDATION B Y MINK 407
success in evading the predator. The longer the distance or duration of pursuit the greater the likelihood of capture (one sample runs test P=O.O5).
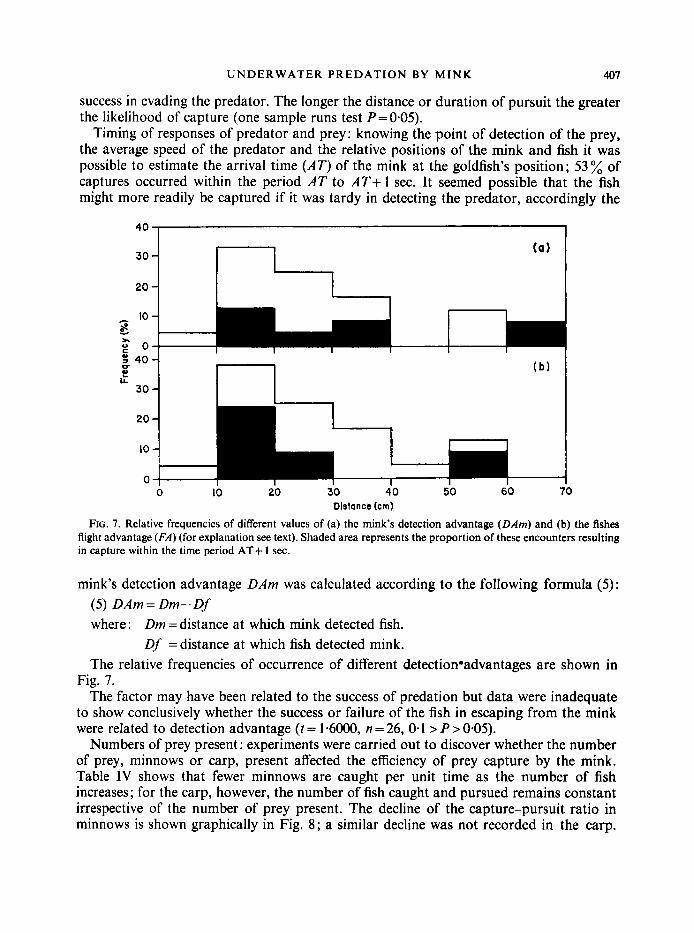
Timing of responses of predator and prey: knowing the point of detection of the prey, the average speed of the predator and the relative positions of the mink and fish it was possible to estimate the arrival time (AT) of the mink at the goldfish’s position; 53% of captures occurred within the period AT to AT+ I sec. It seemed possible that the fish might more readily be captured if it was tardy in detecting the predator, accordingly the
30-1 n
2oj 10 K T 0
0 I 0 20 30 40 50 60 70 Distance (cm)
FIG. 7. Relative frequencies of different values of (a) the mink’s detection advantage (DAm) and (b) the fishes flight advantage (FA) (for explanation see text). Shaded area represents the proportion of these encounters resulting in capture within the time period AT+ 1 sec.
mink’s detection advantage DAm was calculated according to the following formula ( 5 ) : (5) DAm = Dm- Df where : Dm = distance at which mink detected fish.
Df =distance at which fish detected mink. The relative frequencies of occurrence of different detection*advantages are shown in
Fig. 7. The factor may have been related to the success of predation but data were inadequate
to show conclusively whether the success or failure of the fish in escaping from the mink were related to detection advantage ( t = 1-6OO0, n = 26, 0.1 > P > 0.05).
Numbers of prey present : experiments were carried out to discover whether the number of prey, minnows or carp, present affected the efficiency of prey capture by the mink. Table 1V shows that fewer minnows are caught per unit time as the number of fish increases; for the carp, however, the number of fish caught and pursued remains constant irrespective of the number of prey present. The decline of the capture-pursuit ratio in minnows is shown graphically in Fig. 8 ; a similar decline was not recorded in the carp.
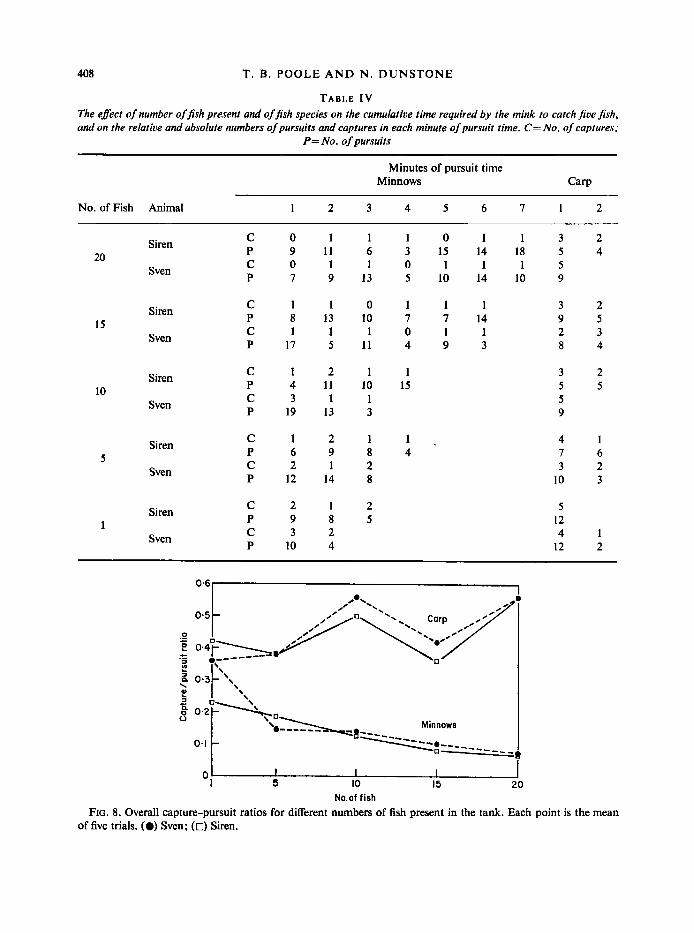
408 T. B . POOLE A N D N . DUNSTONE
TABLE I V The effect of number offish present and offish species on the cumulative time required by the mink to catch Jive fish, and on the relative and absolute numbers of pursuits and captures in each minute of pursuit time. C= No. of captures;
P= No. of pursuits
Minutes of pursuit time Minnows Carp
No. of Fish Animal 1 2 3 4 5 6 7 1 2
C P C P
C P C P
C P C P
C P C P
C P C P
Siren
Sven 20
Siren
Sven 15
Siren
Sven 10
Siren
Sven 5
Siren
Sven 1
0 9 0 7
1 8 1
17
1 4 3
19
1 6 2
12
2 9 3
10
1 11 1 9
1 13 1 5
2 11 1
13
2 9 1
14
1 8 2 4
1 6 1
13
0 10 1
11
1 10 1 3
1 8 2 8
2 5
1 3 0 5
1 7 0 4
1 15
1 4
0 1 1 3 15 14 18 5 1 1 1 5
10 14 10 9
1 1 3 7 14 9 1 1 2 9 3 8
4 7 3
10
5 12 4
12
2 4
2 5 3 4
2 5
1 6 2 3
1 2
1 1 I 5 10 15 20
No.of fish FIG. 8. Overall capture-pursuit ratios for different numbers of fish present in the tank. Each point is the mean
of five trials. (0) Sven; (0) Siren.

UNDERWATER PREDATION B Y M I N K 409
Size of prey: fish catching efficiency of the mink was also found to be affected by the size of prey presented. Two mink were used in the experiments and the size of fish and length of pursuit were found in both instances to be positively correlated: Spearman Rank Correlation Coefficient rs for mink Sven = 0.76, P < 0.01 ; for mink Siren = 0.83, P < 0.001.
Discussion The experiments which have been described show that ranch-bred mink are capable of
actively hunting and catching fish underwater. The necessity for training them and the fact that this took longer if they were adults suggest that mink learn to catch fish when young, possibly by accompanying their mothers on fishing expeditions.
Fish were generally detected by the mink before it entered the water; they have also been observed to use this stratagem in the wild by Herrick (1892) who watched a wild female mink remaining motionless on a boulder in mid-stream, diving into the water only when fish approached. Underwater searching occurred in the laboratory tank during the experiments but this frequently took place after prey had already been observed by the mink from outside the tank. Mink entered the water to search for fish only when they were familiar with the tank and had had previous experience of catching fish in it.
If a mink lost sensory contact with the fish during pursuit it very frequently returned to the surface and re-located the prey from out of the water. The otter, by contrast, usually continues underwater searching (Erlinge, 1968); this difference in behaviour may be related to the otter’s superior vision under water (Walls, 1942; Balliet & Schusterman, 1971; Schusterman & Barrett, 1973). In our experiments the maximum length of time spent underwater by the mink was 20sec whereas Erlinge (1968) has recorded 30sec sub- mergence by the otter; unlike the otter, which expires at the end of a dive (Harris, 1968), the mink expires underwater during swimming; this may be advantageous in reducing buoyancy but must reduce the amount of time that the mink can spend underwater.
Movement of the prey has been shown to be an important stimulus to predatory behaviour in other carnivores such as polecats (Eibl-Eibesfeldt, 1956; Apfelbach, 1973), mongooses (Rasa, 1972) and cats (Leyhausen, 1960). When the mink is outside the tank, movement of the fish appears to be an important stimulus to elicit diving. However, underwater movement of the fish had no measurable influence upon the distance at which mink detected their prey. Furthermore, mink retrieved both inanimate objects and dead fish from the tank spontaneously. This is rather surprising in view of the fact that, in general, moving targets are more readily detected by the mammalian eye than stationary ones (see Walls, 1942).
Having once detected the fish from a terrestrial vantage point, the mink’s optimum stratagem is to dive directly on to the victim using gravity to provide extra velocity. If, however, the fish is at some distance from the mink, the predator dives into the water and then searches until it relocates its prey. Mink, like otters (Erlinge, 1968), do not stalk or ambush prey and, in general, behave like the perch (Pevcufluviutilis) where the main stratagem is to rush in upon the prey (Benzie, 1965; Neil1 & Cullen, 1974).
Sequences of behavioural elements were shown to be highly organized in predatory behaviour and, in contrast, the behaviour of the prey was less highly organized and therefore less predictable ; unpredictability was increased by protean stratagems such as zigzag and sharp turn. The efficiency of the predator in capturing its prey is probably

410 T. B . POOLE A N D N . DUNSTONE
over-estimated in these experiments because the fish were in a confined space. The high incidence of vertical flight, for example, is probably related to the predator’s ability to corner the fish in the tank. Vertical flight was normally in an upwards direction so that the fish swam directly into the mink’s forwardly directed visual field (see Johnson, 1901). Goldfish were most commonly captured when static (59%) or adopting vertical flight (29%); they were rarely caught when performing zig-zag, sharp turn or horizontal flight. The duration of pursuit was positively related to probability of capture of the prey (P=0-05, one sample runs test) so that although fish were, on average, faster swimmers than mink it seems that mink were capable of fatiguing their prey by lengthy pursuit. Larger fish swim more rapidly (Bainbridge, 1960) and in the present experiments, as would be expected, the mink was seen to be more efficient in capturing smaller fish. In the case of larger fish longer pursuits by the mink were necessary to tire the prey. Ryder (1955) has suggested that otters catch fish in proportion to their abundance and speed of swimming.
The experiments with three different species of fish raise the problems of alertness of the prey and its shoaling behaviour. The most difficult species to catch (the minnow) was also the most alert ; the goldfish was least alert. It is not known whether alertness is a specific characteristic of these fish or results from their previous experience of predators (see Seghers, 1974). Although, as far as is known, none of the prey had any previous exposure to mink, the minnows came from a lake which contained an underwater predator (the trout), the carp from a pond which was frequented by herons and the goldfish from a dealer where predation was less likely.
Experiments with different numbers of prey presented simultaneously, clearly demon- strated that in the minnow, a species which forms highly organized shoals (see Pitcher, 1973), increasing numbers of prey decreased the predator’s efficiency although there was no evidence that bouts of pursuit were shorter when more fish were present (see Deelder, 1951 ; Hoogland, Morris & Tinbergen, 1956; Benzie, 1965). When hunting the shoal of minnows the mink dived into the shoal, dispersing it and then attempted to capture stragglers or to isolate one fish from the group. It would seem probable that shoals of fish are more conspicuous when viewed from above the water than single fish if they are within the detection range of the predator.
Detailed analysis of the predatory behaviour of mink on goldfish makes it possible to identify factors which contribute to the mink’s successful predation and those which favour the escape of its prey.
The mink is favoured by an aerial vantage point from which it can observe its prey; even underwater the mink generally detects the goldfish before being detected by the prey. The mink, although slower than the fish, has greater stamina in pursuit and its sinuous neck enables it to make a grab at a fleeing fish if it has succeeded in cornering it.
The goldfish has the advantages of greater speed and manoeuvrability which enable it to confuse the predator; it is also favoured by the mink’s reduction in visual acuity under water and its need to return to the surface for air and to relocate its prey. It may also be that the greater predictability of the mink’s behaviour favours the fish, by enabling it to anticipate the predator’s actions.
It is clear that the advantages to fish and mink are closely balanced; although the mink appears to be a typical terrestrial carnivore possessing only behavioural adaptations to an amphibious mode of life it is none the less a highly effective predator on small fish.

UNDERWATER PREDATION BY M I N K 41 1
summary Underwater predatory behaviour of American mink (Mustela uison) was investigated
in the laboratory using cinC film. The behaviour of mink and responses of three species of fish are described. It was
necessary to train ranch-bred mink to enter water and catch fish; young mink appeared to be easier to train than adults. Mink spent 5-20 sec under water when fishing; prey had usually previously been located from an aerial vantage point. Predatory behaviour was highly organized sequentially whereas fish were more prone to indulge in unpredictable strategems; the behaviour of mink and fish were highly correlated.
The mink’s efficiency in catching fish was related to prey size (smaller individuals being more vulnerable to capture) and shoaling. Minnows which form highly organized shoals were less easily caught when present in larger numbers; this was not true of a non-shoaling species, the carp.
Of the three species of prey presented, vulnerability to capture took the form goldfish > carp > minnow; these differences, however, may have been influenced by the fish’s previous experience of underwater predators.
We would like to record our gratitude to the S.R.C. for providing financial assistance in the form of a research studentship to Mr N. Dunstone. We are also grateful to Mr R. A. Moore for taking the cine film.
R E F E R E N C E S Apfelbach, R. (1973). Woran erkennt ein Raubtier seine Beute? Umschau 73 (15): 453457. Bainbridge, R. (1960). Speed and stamina in three fish. J. exp. Biol. 37: 129-153. Balliet, R. F. & Schusterman, R. J. (1971). Underwater and aerial visual acuity in the Asian “clawless” otter
Benzie, V. L. (1965). Some aspects of the anti-predator response of two species of stickleback. D.Phil. Thesis. Oxford
Day, M. G . & Linn, I. (1972). Notes on the food of feral mink Mustela vison in England and Wales. J. Zool., hnd.
Deelder, C. L. (1951). A contribution to the knowledge of the stunted growth of perch(Percafluviafi1is) in Holland.
Eibl-Eibesfeldt, I. (1956). Angeborenes und Erworbenes in der Technik des Beutetotens. Z. Siiugetierk. 21: 135-137. Erlinge, S. (1968). Food studies on captive otters Lutra lutra L. Oikos 19: 259-270. Erlinge, S. (1969). Food habits of the otter Lutra lutra L. and the mink Mustela uison Schreber in a trout water in
Gerell, R. (1967). Food selection in relation to habitat in mink (Mustela vison Schreber) in Sweden. Oikos 18:
Gerell, R. (1968). Food habits of the mink, Mustela vison Schreber, in Sweden. Viltrevy 5 : 119-21 I . Harris, C. J. (1968). Otters. A study of recent Lutrinae. London: Weidenfeld & Nicholson. Herrick, C. L. (1892). The mammals of Minnesota. Bull. Minn. geol. Surv. No. 7 : 1-301. Hoogland, R., Morris, D. & Tinbergen, N. (1956). The spines of sticklebacks (Gasterosteus and Pygosteus) as a
means of defence against predators (Perca and Esox). Behaviour 10: 205-236. Johnson, L. (1901). Contributions to the comparative anatomy of the mammalian eye, chiefly based on ophthal-
moscopic examination. Phil. Trans. R. SOC. (B.) 194: 1-82. Leyhausen, P. (1960). Verhaltensstudien an Katzen. Z. Tierpsychol. Beiheft 2: 1-120. Maxwell, A. E. (1961). Analysing quantitative data. London: Methuen. Neill, S. R. St. J. & Cullen, J. M. (1974). Experiments on whether schooling by their prey affects the hunting
Pitcher, T. J. (1973). The three dimensional structure of schools in the minnow, Phoxinus phoxinus (L.). Anim.
(Ambionyx cineria cineria). Nature, Lond. 234: 305-306.
University.
167: 463473.
Hydrobiologica 3: 357-378.
southern Sweden. Oikos 20: 1-7.
233-246.
behaviour of cephalopods and fish predators. J. Zool., Lond. 172: 549-569.
Behav. 21 : 673-686.
412 T. B. POOLE A N D N. D U N S T O N E
Poole, T. B. (1973). Cink film and computer analysis of rapid sequences of behaviour. Med. Biol. Zllustr. 23: 170-175. Poole, T. B. (1974). Detailed analysis of fighting in polecats (Mustelidae) using cint film. J. Zool., Lond. 173:
Rasa, 0. A. E. (1972). Prey capture, feeding techniques and their ontogeny in the African dwarf mongoose,
Ryder, R. A. (1955). Fish predation by the otter in Michigan. J. Wildl. Mgmt 19: 497-498. Schusterman, R. J. & Barrett, B. (1973). Amphibious nature of visual acuity in the Asian “clawless” otter.
Seghers, B. H. (1974). Geographical variation in the responses of guppies (Poecilia reticulata) to aerial predators.
Siegel, S. (1956). Non-parametric statisticsfor the behavioral sciences. New York: McGraw Hill. Sinclair, W., Dunstone, N. & Poole, T. B. (1974). Aerial and underwater visual acuity in the mink (Mustela vison
Walls, G . L. (1942). The vertebrate eye and its adaptive radiation. New York: Cranbrook Inst. Sci. Wiistehube, C. (1960). Beitrage zur Kenntnis besonders des Spiel-und Beuteverhaltens einheimischer Musteliden.
369-393.
Helogale unduletarupa. Z . Tierpsychol. 32 : 449-488.
Nature, Lond. 244: 5 18-5 19.
Oecologia 14: 93-98.
Schreber). Anim. Eehao. 22: 965-974.
Z . Tierpsychol. 17: 579-613.
Appendix A Origins, experience and latency to first capture by mink used in the experiments
Mink
Neptune Argo Nautilus Sven Rover Siren Cicely Nasteg Acis *Snowy
Origin
Ranch Ranch Ranch Ranch Ranch Ranch Ranch Ranch Wild Ranch
Age (months)
2 2 2
15 9 9
10 9
c 15 5
Sex
d d 6 ? 6 0 P d 6 6
- Previous experience Training method
Latency to 1st capture (hours arena exper.)
Swimming only Swimming only Swimming only None None None None None ? None
Full diet Full diet Reduced diet Reduced diet Reduced diet Reduced diet Reduced diet Reduced diet Reduced diet Reduced diet
2 4 4
22 46 14 26 22 34 95
* Albino mink with unpigmented eyes, others dark.





























