Embed Size (px)
Citation preview

Université de Montréal
Fonctions de transport et d'hydrolyse dans le complexe glucose-6-phosphatase microsornai de foie
Par
Borhane Annabi
Département de Biochimie
FacuIté de Médecine
Thèse présentée a la Faculté des études supérieures en vue de l'obtention du grade de
Philosophiæ Doctor (Ph.D.) en Biochimie
janvier, 1997
%orhane Annabi, 1997

National Library IJll of,,, Bibliothèque nationale du Cana~a
Acquisitions and Acquisitions et Bibliog aphic Services services bibliographiques 395 Wellington Street 395. rue Wellington Ottawa ON K1A ON4 ûttawaON K1AON4 Canada Canada
your Me Vom nibmce
Our üle N m reMmta3
The author has granted a non- exclusive licence allowing the National Library of Canada to reproduce, loan, distribute or sell copies of this thesis in microfom, paper or electronic formats.
The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fkom it may be pninted or otherwise reproduced without the author's permission.
L'auteur a accordé une Licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de rnicrofiche/nlm, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.

Université de Montréal
Faculté des études supérieures
Cette thèse intitulée :
Fonctions de transport et d'hydrolyse dans le complexe glucose-6-phosphatase rnicrosomal de foie
présentée par :
Borhane Annabi
a été évaluée par un jury composé des personnes
n
suivantes :
Thèse acceptée le : J J: O % 7 9

Décembre 1996
Les CO-auteurs autorisent l'université de Montréal à microfilmer cette
thèse rédigée par article et à p r ê t e r ou à vendre d e s copies du microfilm à
d e s f i n s d'enseignement et de recherche.
Noms et Signatures des CO-auteurs:
Dr. Gérald van de Werve
Dr. Hubert Vidal
\\ Dr. Jean-François St-Denis
Dr. Mark H. Rider
2 Mme Hanane Khoury

Universi té de Montréal
Bibliothèque

La glucose-6-phosphatase @-glucose-6-phosphate phosphohydrolase; EC
3.1.3.9) (Glc-6-Pase) est l'enzyme qui catalyse la dernière étape de la production
de glucose au niveau du foie (et du cortex rénal) et donc, l'ultime étape de la
gluconéogénèse et de la glycogénolyse. Cette enzyme hydrolyse le glucose-6-
phosphate (Glc-6-P) en glucose (Glc) et en phosphate (Pi) et est donc essentielle
dans le maintien de la glycémie en période de jeûne. Chez l'homme, un certain
nombre de maladies métaboliques héréditaires sont associées à des déficits de la
Glc-6-Pase (diabète, glycogénoses de type 1). Cette enzyme est formée d'un
complexe multipeptidique dans la membrane du réticulum endoplasmique (RE). Ce
complexe serait constitué de la sous-unité catalytique et de transporteurs, encore
hypothétiques, pour le Glc-6-P (substrat de l'enzyme), le Glc et le Pi (produits de
la réaction). La validation d'une telle organisation moléculaire ne pourra être
obtenue qu'après l'identification, la purification et le clonage de ces différentes
composantes. Une fois la Glc-6-Pase purifiée et sa séquence connue, il nous sera
possible de comprendre les relations structure/fonction de cette enzyme
fondamentale du métabolisme du glucose, mais aussi approcher les pathologies du
diabète et des glycogénoses de type 1 au niveau moléculaire.
Deux théories sont proposées en ce qui a trait à l'organisation moléculaire
du système Glc-6-Pase dans la membrane du RE. D'une part, un modèle
co~fomationnel implique la reconnaissance et la liaison du substrat à la Glc-6-
Pase sur la surface externe de la membrane microsomale et mène à son hydrolyse
et à la libération des produits dans la lumière microsomale. C'est donc un système
où un ou plusieurs peptides synchronisent tant le transport que la fonction
catalytique du système. D'autre part, le modèle de tran~port $14 substrat postule la

présence d'un transporteur spécifique (Tl) du Glc-6-P, transportant le substrat du
cytosol vers la lumière microsomale où a lieu son hydrolyse par une
phosphohydrolase non spécxque. T l serait alors la protéine responsable de Fétape
ümitante de la conversion de Glc-6-P en Glc et en Pi. La Glc-6-Pase serait donc un
complexe de protéines comprenant l'unité catalytique (la phosphohydrolase), un
transporteur de Glc-6-P (Tl), un transporteur de Pi (T2), et un transporteur de Glc
(T3 ou GLUT 7).
L'étude des cinétiques de capture et d'hydrolyse du Glc-6-P par l'enzyme in
sifir dans la membrane du RE (microsomes) suggère que la présence de Tl n'est
pas essentielle pour expliquer la spécificité de l'enzyme pour ce substrat. De plus,
le traitement des rats avec un analogue des giucocorticoides (la triamcinoIone)
ainsi que l'utilisation de modifications chimiques spécifiques de certains résidus
possiblement impliqués dans le mécanisme catalytique de l'enzyme nous ont permis
de réévaluer le modèle d'organisation moléculaire de la Glc-6-Pase. Nous
postulons la présence de protéines auxiliaires contrôlant l'accès du Glc-6-P au site
catalytique. Ces protéines seraient égaiement responsables du couplage de la
fonction d'hydrolyse du Glc-6-P à celle d'accumulation du Glc dans la lumière du
RE.
Mots clés: glucose-6-phosphatase; foie; rnicrosomes; purification;
glucocorticoïdes; modifications chimiques

TABLE DES MATIÈRES
Sommaire
Table des matières
Liste des tableaux
Liste des figures
Liste des abbréviations
Remerciements
1- REVUE DE LA LITTÉRATURE
1.1 Introduction
1.2 La glucose-6-phosphatase
1.2.1 Distribution tissulaire et Iocalisation subcellulaire
1 -2.2 Homologie de la glucose-6-phosphatase avec d'autres protéines
1.3 Propriétés cinétiques du système glucose-6-phosphatase
1.3.1 Activités multiples de la glucose-6-phosphatase
1.3.2 Mécanisme catalytique de la fonction d'hydrolyse du glucose-6-
phosphate
1.3 -3 Transport et accumulation du glucose à l'intérieur des
microsornes
1.3.4 Un nouveau concept du phénomène de latence
1.4 Topologie du système glucose-6-phosphatase dans la membrane du
réticulum endoplasmique
1.4.1 Identification et caractéristiques des composantes du système
glucose-6-p hosp hatase
1.4.2 Modèle proposés pour l'organisation moléculaire du système
glucose-6-phosp hatase
iii
v
ix
ix
si
xii

1.5 Clonage du gène codant pour la glucose-6-phosphatase
1 S. 1 Clonage de la sous-unité catalytique
1.5.2 Clonage du promoteur
1.6 Déficiences génétiques du système glucose-6-phosphatase
1.6.1 Manifestations cliniques et métaboliques des glycogénoses
1.6.2 Base moléculaire des glycogénoses de type Ia
1 -6.2.1 Déficit en Glc-6-Pase, Ia
1.6.2.2 Déficit en transporteur de Glc-6-Pase microsomal, type Ib
1.6.2.3 Déficit en transporteur de PVPPi rnicrosomal, type Ic
1.6.2.4 Déficit en transporteur de Glc rnicrosomal, type Id
1.7 Contrôle nutritionnel et hormonal de la glucose-6-phosphatase
1.8 Objectifs de la thèse
2- M~THODOLOGIE
2.1 Préparation des microsomes de foie
2.2 Mesures des activités enzymatiques
2.2.1 Glucose-6-phosphatase et mannose-6-phosphatase
2.2.2 Phosphoglucomutase
2.3 Purification de la glucose-6-phosphase
2.3.1 Solubilisation des microsomes a l'aide de différents détergents
2.3.2 Effet de la force ionique sur la distribution de la Glc-6-Pase lors
de la centrifugation des microsomes solubilisés
2.3.3 Chromatographie sur colonne d'hydroxylapatite
2.3.4 Chromatographie sur colonne d'affinité
2.4 Caractérisation de la Glc-6-Pase par la formation d'un intermédiaire
phosphorylé

2.4.1 Synthèse du r2p] GIC-6-P, [ 3 2 ~ ] ~ a n - 6 - ~
2.4.2 Marquage radioactif et autoradiograp hie
2.5 Inactivation spécifique de l'activité phosphohydrolase du système
Glc-6-Pase
2.5.1 Modification des résidus Aspartate et Glutamate avec le
dicyclohexylcarbodümide
2.5.2 Modification des résidus Lysine avec le pyridoxal5'-phosphate
2.6 Mesure de transport de glucose par la membrane microsomale
2.7 Traitement des rats à la triamcinolone et à la streptozotocine
2.8 Immunobuvardage
3- ARTICLES
3.1 Sécificité du système Glc-6-Pase pour le Glc-6-P
3.1.1 Histone II-A stimulates glucose-6-phosphatase and reveals
mannose-6-phosphatase activities without permeabilization of
Iiver microsomes. St-Denis, J.-F., Annabi, B., Khoury, H., and
van de Werve, G. (1995) Biochem. J. 3 10:2 10-224
3.2 Transport de glucose associé au système Glc-6-Pase
3 -2.1 Glucose transport and glucose-6-phosphate hydrolysis in intact
rat liver microsomes. St-Denis, LF. , Berteloot, A., Vidal, H.,
Annabi B., and van de Werve, G. (1995) J. Biol. Chem. 270:
2 1092-2 1097
3.3 Résidus impliqués dans le processus catalytique de la Glc-6-Pase
3.3.1 Inactivation of rat liver rnicrosomal glucose-6-phosphatase by
dicyclohexylcarbodiirnide: Evidence for the role of carboxyl
groups in catalysis. Annabi, B., van de Werve, G., and Rider
M.H. (1 996) (soumis)
vii
47
48
48
48
50
50
52
52
55
55

3.4 Fonctions distinctes de production de glucose par le système Glc-6-Pase
3.4.1 Evidence that the transit of glucose into liver microsornes is not
required for fùnctiomal glucose-6-phosphatase. Annabi, B., and
van de Werve, G. (1996) (soumis)
4- DISCUSSION
4.1 Réévaluation du modèle de transport du substrat
4.1.1 Diagnostic des glycogénoses de type 1
4.1.2 Spécificité de la Glc-6-Pase pour le Glc-6-P: un nouveau
concept du phénomène de latence
4.2 Transport de glucose unidirectionnel associé au système Glc-6-Pase
4.3 Modulation de I'activité Glc-6-Pase
4.3.1 Effet des histones &A sur la Gk-6-Pase
4.3.2 Fonctions distinctes de production et d'accumulation de glucose
4.3.3 Etat oligomérique de la Glc-6-Pase
4.4 Résidus impliqués dans le processus catalytique de la Glc-6-Pase:
Inactivation de la Glc-6-Pase par le dicyclohexylcarbodiimide
5- CONCLUSIONS GÉNÉRALES
5.1 Le modèle conformatio~el: une alternative au modèle de transport du
substrat
5.2 Preuves d'un couplage des fonctions d'hydrolyse du Glc-6-P et
d'accumulation du glucose
5.3 Le cycle GldGlc-6-P: rôle de la Glc-6-Pase comme transporteur
secondaire de glucose

LISTE DES TABLEAUX
Tableau 1 : Enzymes déficientes et symptômes des différents types de
glycogénose 25
Tableau 2 : Purification de la Glc-6-Pase hépatique de rat 45
Tableau 3 : Caractéristiques cinétiques de différentes conformations de Ia
Glc-6-Pase microsomale. 129
LISTE DES FIGURES
Figure 1 : Glycogénolyse et néoglucogénèse
Figure 2 : Représentation schématique proposée du systeme Glc-6-Pase
dans son environnement membranaire natif 12
Fiyre 3 : Représentation schématique des deux modèles proposés pour la
topologie du système Glc-6-Pase dans les microsornes de foie 18
Figure 4 : Représentation de la séquence en acides aminés et topologie
mernbranaire de la protéine 36kDa 23
Figure 5 : Solubilisation des microsomes à Saide de différents détergents
et stabilité thermique de la Glc-6-Pase 38
Figure 6 : Effet de la force ionique sur la distribution de la Glc-6-Pase
lors de la centrifugation des microsornes solubilisés 40

Figure 7 :
Figure 8 :
Figure 9 :
Figure 10 :
Figure 11 :
Figure 12 :
Figure 13 :
Figure 14 :
Figure 15 :
Purification de la Glc-6-Pase par chromatographie sur colonne
d'hydroxylapatite
Purification de la Glc-6-Pase par chromatographie sur colonne
de Glc(NH2)-6-P-Mépharose CL 6B
Électrophorèse sur gel de polyacrylamide en condition
dénaturantes des différentes étapes de la purification de la Glc-
6-Pase
Réactions impliquées dans la modification des groupements
carboxylés des résidus aspartate et glutamate par le
dicyclohexylcarbodiirnide
Liaison du pyridoxal S'-phosphate avec son apoenzyme et
réaction avec les groupements aminés des résidus lysine
Immunobuvardage utilisant l'Ac anti-Glc-6-Pase avec des
microsornes isolés de foie de souris
Immunobuvardage utilisant l'Ac anti-Glc-6-Pase avec des
microsornes isolés de foie humain normal et atteint de
glycogénose de type Ia
Etat d'oligomérisation de la Glc-6-Pase suite à un traitement
acide, organique, ou de dénaturation thermique
Rôle de la GIc-6-Pase dans le tranmort secondaire du Glc

CP : DCCD : DIDS : EDC : EDTA : EGTA :
FSRFA :
GEE : GK : Glc : Glc- 1-P : Glc(NH2)-6-P Glc-6-P : Glc-6-Pase : GLUT : Man : Man-6-P : Man-6-Pase : NaBH4 : Pi : PGlcM : PLP : PPi : RE : RT-PCR :
acide désoxyribonucléique complémentaire acide ribonucléique messager adénosine triphosphate 3 -[(3 -Cholamido pro p y1)-dirnethylammonio] -2-hydroxy- 1 -propanesulfonate carbarnyl-phosphate MN' dicyclo hexylcarbodümide acide diisothiocyanatostilbène 2,Zdisulfonique 1 -et hyl-3-[-3 -dimethylamino propyl]carbodiimide acide éthylenediamuietétraacétique acide éthylène glycol bis@-aminoét h y1 éther)-N N.N',N'- tétraacétique (Fast-Sampling, Rapid-Filtration Apparatus) appareil à échantillonnage et à filtration rapide glycine ethyl ester gluco kinase gIucose glucose- 1 -phosphate glucosamine-6-p hosp hate glucosed-phosphate glucose-6-phosphatase transporteur de glucose facilité mannose mannose-6-phosphate mannose-6-phosphatase borohydrure de sodium phosphate inorganique phosphoglucomutase pyridoxal S'-phosphate pyrophosp hate inorganique réticulum endoplasmique (Reverse Transcn ption-Polymerase Chain Reaction) réaction en chaine de la polymérase transcriptase inverse transporteur microsomal de Glc-6-P transporteur microsomal de Pi, PPi ou de CP transporteur rnicrosomal de Glc

REMERCIEMENTS
Je remercie le Professeur Gérald van de Werve pour m'avoir accueilli au
sein de son groupe de recherche. Je lui suis particulièrement reconnaissant de
m'avoir incité à participer à de nombreux congrès, et de m'avoir encouragé à croire
en mes capacités dans les moments difficiles.
Je remercie également les Dr Hubert Vidal et Mark Rider que j'ai eu la
chance de cotoyer à l'Unité HORM de l'Institut de Pathologie Cellulaire en
Belgique et a qui je voudrais dire comme il fut agréable d'avoir pu travailler en leur
compagnie et d'avoir reçu leur aide, leurs conseils et leur amitié.
raimerais exprimer ma gratitude à tous mes collègues de laboratoire qui
m'ont appuyé pendant ces longues années : Dr Duna Massillon, Dr Jean-François
St-Denis, Angela Romanelli, Dr Thierry Brun, Hanane Khoury, Didier
Vertommen, Luc Bertrand, Guylaine Gevry et Sophie Tchu.
Qu'il me soit permis finalement de remercier l'Association du diabète du
Québec, la Faculté des Études Supérieures de l'Université de Montréal, le
Gouvernement du Québec ainsi que le Professeur Gérald van de Werve pour leur
appui financier.
Je remercie aussi de tout mon coeur mes parents et ma soeur, auprès
desquels j'ai toujours trouvé beaucoup d'encouragements et de compréhension.
Enh, une pensée toute spéciale à Dominique Mainville pour son amour, son
écoute et sa disponibilité dans le dernier droit.

1.1- Introduction
Le glucose (Glc) sanguin, dont la capture est facilittie par des transporteurs
spécifiques (GLUT1 et GLUT2 pour le foie, GLUTl et GLUT4 pour le muscle),
est rapidement phosphorylé par une hexokinase dans les cellules musculaires et
aussi par une glucokùiase (GK) spécifique du Glc dans les cellules
parenchyrnateuses hépatiques pour former du glucose-6-phosphate (Glc-6-P). Ce
dernier est ensuite métabolisé en empruntant soit la voie glycolytique menant à la
formation de pyruvate, soit la voie glycogénique pour former du glycogène. Le
glycogène est la principale forme d'emmagasinage des glucides chez les animaux et
constitue une réserve énergétique locale et une source de substrat rapidement
disponible pour la glycolyse. Les variations de concentration de glycogène, qui
permettent de maintenir le Glc sanguin à un taux remarquablement stable (* 5
mM), sont étroitement contrôlées par des mécanismes homéostatiques et
hormonaux (van de Werve et Jeanrenaud, 1987). En période de jeûne, du Glc-6-P
est donc formé à partir du glycogène par la voie giycogénolytique et/ou à partir de
substrats tri-carbonés par la voie de la néoglucogénèse. La glucose-6-phosphatase
(GIc-6-Pase) catalyse la dernière étape de la production de Glc dans les tissus qui
possèdent cette enzyme (foie et cortex rénal), en hydrolysant le Glc-6-P

2
(Nordlie et Sukdski, 1985) (voir figure 1), permettant ainsi au Glc devenu libre de
diffuser de la cellule dans le sang. La Glc-6-Pase est indispensable au maintien de
la glycémie et joue donc un rôle primordial dans le contrôle de Iboméostase du
Glc.
Les maladies de l'emmagasinage du glycogène forment un groupe de
désordres héréditaires caractérisés par une mobilisation déficiente du glycogène ou
une déposition de formes anormales de glycogène. L'étape catalysée par la
phosphorylase est l'étape lirnitante de la glycogénolyse et, combinée avec Faction
des autres enzymes débranchantes, dégrade complètement le glycogène. Le Glc- 1-
P produit par la phosphorylase peut être transformé en Glc-6-P par l'action de la
phosphoglucomutase (PGlcM), dont une des formes est microsomale (voir figure
1). Un cycle du Glc dit "futile" a été décrit dans le foie (Katz et al., 1978, Christ et
aL, 1986). Durant ce cycle, le Glc est phosphorylé en Glc-6-P par la GK et
déphosphorylé en Glc par la Glc-6-Pase avec consommation ÇI'ATP. Une
augmentation de ce cycle a été observée chez des patients diabétiques non
insulino-dépendants (Efendic et al., 1988) et chez des chiens pancréatectornisés
(Shi et a l , 1994). Des niveaux élevés dtARNm de la Glc-6-Pase hépatique ainsi
que de l'activité de t'enzyme sont observés dans le diabète insulino-dépendant (Liu
et al., 1994) et peuvent contribuer a l'augmentation de la néoglucogénèse et à
l'hyperglycémie qui caractérisent cette condition (Burchell et Cain, 1985).

.Membrane plasmique
Figure 1. GlycogCnolyse et néogiucogenèse hépatique. Le foie joue un rôle important dans la régulation des niveaux de Glc sanguin et possède 2 voies métaboliques qui lui permettent de fabriquerAu Glc, soit la néoglucogenèse et la giycogènolyse. La glycogène phosphorylase et la glycogène synthase sont controlées par un groupe de réactions complexes faisant appel à des modications covalentes @hosphorylation~déphosphorylation) et à des mécanismes a.Uostériques (+, stimulation, -, inhibition). Le Glc-6-P se trouve au carrefour de ces 2 voies métaboliques. Une seule enzyme, la Glc-6-Pase localisée dans le réticulum endoplasrnique (RE), catalyse l'étape h a i e de la production de Glc.

1.2- La glucose-6-phosphatase
1.2.1- Distribution tissulaire et localisation subcellulaire
Des niveaux d'activités élevés de la Glc-6-Pase ont été mesurés dans les
foies de nombreux mamrnifiires, oiseaux, amphibiens, reptiles, poissons et
arthropodes (Nordlie, 1 97 1, Hemnann et Nordlie, 1972, Nordlie, 1979). Bien que
plus abondante dans le foie et le rein, une activité Glc-6-Pase a aussi été détectée
dans le cerveau (homogénats du tissu et astrocytes en culture primaire; Forsyth et
al., 1993, Bell et al., 1993), la muqueuse intestinale (Barash et al., 199 1, Pears et
ai., 1992), les glandes surrénales (Hume et al., 1995), les testicules, la rate, les
poumons (Nordlie, 1979, Colilla et al., 1975), les cellules P des îlots du pancréas
(Waddell et Burchell, 1988), ainsi que dans la vésicule biliaire (Hill et al., 1989).
En plus du foie et du rein, tous deux exprimant des niveaux élevés
d'activité Glc-6-Pase, plusieurs autres tissus semblent donc aussi pouvoir exprimer
la Glc-6-Pase. Dans ces tissus, par contre, cette activité, dosée initialement dans
des homogénats (Ashmore et Weber, 1959, Nordlie, 1974) et plus tard dans des
microsomes (Mithieux et al., 1995, Mithieux et al., 1996), est au moins 10 fois
plus basse que celle retrouvée dans le foie ou le rein et pourrait être attribuée à des
phosphatases non-spécifiques. Waddell et Burchell (1988) ont, par contre,
rapporté une activité Glc-6-Pase dans des rnicrosomes isolés dTlots du pancréas
plus élevée que dans le foie. Cependant, en utilisant des techniques d'analyse des

ARNm (acide ribonucléique messager) par éléctrophorèse suivie de transfert et
d'hybridation sur filtre à I'aide de sondes d'ADNc (acide désoxyribonucléique
complémentaire) spécifiques (Northern blots) et de la RT-PCR (reverse
transcription-polymerase chah reaction), 1'ARNm de la Glc-6-Pase n'a pu être
detecté que dans le foie et le rein, où de l'activité a été mesurée (Mithieux et al.,
1996). Ces résultats suggèrent fortement que la Glc-6-Pase soit exprimée
spécifiquement dans les tissus giuconéogéniques et que I'activité mesurée dans les
autres tissus n'est qu'artefactuelle et due à des phosphatases non-spécifiques
(Essner, 1973). Des études récentes ont cependant rapporté la présence d'ARNm
de la Glc-6-Pase dans divers autres tissus exempts d'activité (Shingu et al., 1996),
dont le muscle squelettique (Gamberucci et al., 1996), l'épididyme (BurcheU et al.,
1996), ainsi que les îlots isolés du pancréas (Matschinsky, 1996). Le rôle de la Glc-
6-Pase dans ces divers tissus n'a pas été défini.
1.2.2- Homologie de la glucose-6-phosphatase avec d'autres protéines
Feldman et Butler (1969, 1972) ont démontré que la Glc-6-Pase possédait
un résidu histidine dans son site actif qui pouvait être phosphorylé lors du
processus catalytique d'hydrolyse du Glc-6-P. Cette propriété qu'a la Glc-6-Pase
de former un intermédiaire phosphorylé (détaillée à la section 1.3.2) a été exploitée
pour l'identifier dans le foie, en marquant radioactivernent la Glc-6-Pase à l'aide de
[ 3 2 ~ ] ~ l c - 6 - ~ et de [ 3 2 ~ ] ~ ~ i (Countaway et al., 1988, Mithieux et al., 1995).

6
II a été proposé par Granner et Pilkis (1990) que la présence d'une
phosphohistidine au site actif de la Glc-6-Pase la classait dans la famille des
enqmes comprenant la mictose-2,6-bisphosphatase, la phosphoglycerate mutase,
et la phosphatase acide, dont l'activité phosphohydrolase est aussi caracterisée par
la formation d'un intermediaire phosphorylé (Bazan et al., 1989). Cette Evnilie
possède une séquence commune RHG (arginine-histidine-glycine) entourant
l'histidine accepteur de phosphate, et il a été présumé qu'il en était de même pour la
Glc-6-Pase. Ce concept a été remis en question étant donné que la Glc-6-Pase est
une protéine membranaire extrêmement hydrophobique. Le fait de n'être soluble
qu'en présence de lipides et/ou de détergents, empêche la Glc-6-Pase d'avoir la
structure dB de la famille d'enzymes solubles formant un intermédiaire
phosphorylé au cours de la catalyse. L'analyse de la séquence pirnaire de la Glc-6-
Pase hépatique chez l'humain et chez le rat ne démontre d'ailleurs pas la présence
de cette séquence RHG ni d'autres séquences conservées dans la famille hctose-
2,6-bisphosphatase/phosphoglycerate mutasdphosphatase acide, confirmant la non
appartenance de la Glc-6-Pase à cette famille. La séquence nucléotidique et
peptidique de la Glc-6-Pase humaine et de souris (Lei et al., 1993, Shelly et al.,
1993) ne révèle aucune identité avec celles connues à ce jour. Soulignons,
néanmoins, que des courtes séquences d'acides aminés, correspondant à des sites
de phosphorylation par différentes kinases, ainsi que certains motifs communs à la
famille des protéines transrnembranaires ont été identifiés dans la Glc-6-Pase.

1.3 - Propriétés cinétiques du système glucose-6-phosphatase
1.3.1- Activités multiples de la glucoso6-phosp hatase
C'est en 1959 que l'équipe de Hass et Byme, ainsi que celle de Segal ont noté
que le Glc pouvait inhiber l'hydrolyse du Glc-6-P, et ce sont eux qui ont plus tard, été
les premiers à étudier l'activité Glc-6-P/GIc phosphotransférase de la Glc-6-Pase pq.
(211. La Glc-6-Pase est donc une enzyme multifonctionnelie réalisant non seulement
l'hydrolyse du Glc-6-P pq . (l)], mais aussi sa synthèse [Eq. (2)-(4)] en catalysant le
transfert d'un groupe phosphoryle sur le Glc en position 6 (Nordlie, 197 1, 1974). Il est
maintenant bien connu qu'à la diffërence de certaines phosphatases non-sp-ques ne
possédant qu'une fible capacité synthétique, ceiie de la Glc-6-Pase, eff e, peut excéder
de plus de 50% sa fonction hydrolytique maximale (Znieck et al., 1972). A titre
d'exemple, en présence de 100 mM Glc, l'activité phosphotransferase de la Glc-6-Pase
est pratiquement comparable à sa capacité d'hydrolyser le Glc-6-P. Il est à noter que
des réactions de synthèse, telle que la formation de Glc-6-P a partir de Glc et de PPi,
n'ont probablement que peu d'importance physiologique puisque le Km pour le Glc,
dans cette réaction, est supérieur à 100 mM alors que la concentration du Glc à
l'intérieur de l'hépatocyte est de l'ordre du rnillimolaire. De plus, Bontemps et al.
(1978), en étudiant le rôle de la GK dans la phosphorylation du Glc dans les
hépatocytes isolés intacts ainsi que dans les conditions où la GK est absente (nouveau-
né), démontrent que l'activité phosphotransférase de la Gk-6-Pase ne joue aucun rôle

8
dans la phosphorylation du Glc, contrairement à ce que prétend Nordlie (1 97 1, 1974,
1 976).
Glc-6-P + H20 + Glc + Pi (1)
Glc-6-P + [ 1 4 ~ ] ~ ~ ~ $ [ 1 4 ~ ] ~ ~ ~ - 6 - ~ + GIc (2)
PPi + Glc -) Glc-6-P + Pi (3
Carbamyl-P + Glc + Glc-6-P + NH3 + C a (4)
PPi + H20 + 2 Pi ( 5 )
Carbamyl-P + Hz0 + Pi + MI3 + CO2 (6)
Des études subséquentes ont, par la suite, démontré la présence de ces
multiples activités de la Glc-6-Pase associées au foie, au rein, ainsi qu'à plusieurs autres
tissus (Nordlie et Arion, 1964, Lygre et Nordlie, 1968, CoiiUa et al., 1975). L'activité
phosphotransférase de la Glc-6-Pase a, de plus, été trouvée dans le RE, les membranes
nucléaire et plasmique, ainsi que la mitochondrie du foie (Gunderson et Nordlie, 1975,
Nordlie et lorgenson, 1976). Enfin, en plus du G1c-6-P, le Carbamyl-P et le PPi, tel
qu'indiqué par les équations (1)-(6), ainsi que le phosphoénolppvate, le mannose-6-P
et certains autres nucléosides di- et triphosphatés ont été rapportés comme des esters-
phosphate reconnus par la Glc-6-Pase. Finalement, en plus du Glc, le mannose, le 3-0-
methyl-D-Gic, et le 2-deoxy-D-Glc ont été décrits comme étant de bons accepteurs
dans les réactions de transfert de phosphate (Stetten, 1965).

1.3.2- Mécanisme catalytique de la fonction d'hydrolyse du Glc-6-P
La réaction catalysée par la Gl6Pase se caractérise par la formation
transitoire d'un complexe enzyme-phosphate (E-P) avec la formation d'une liaison
covalente entre le groupe imidazole d'un résidu histidine (N-3) de l'enzyme et le
phosphate du Glc-6-P (Nordlie et Lygre, 1966, Feldman et Butler, 1972). En utilisant
cette propriété, il a été possible de marquer la Gic-6-Pase au moyen de [ 3 2 ~ ] ~ l c - 6 - ~
ainsi qu'avec du [ 3 2 ~ ] ~ ~ i et de montrer que sa masse moléculaire devait être d'environ
36,5 kDa (Countaway et al., 1988, Mithieux et al., 1995).
Le mécanisme catalytique d'hydrolyse du Glc-6-P généralement accepté est
décrit par les réactions 1 et 2, où E représente la fonction phosphohydrolase de
l'enzyme et ki-k< les constantes réactionnelles pour chacune des étapes (Segal, 1959,
Arion et Nordlie, 1964, Waiiïn et Arion, 1973, Nordlie, 1982).
kl k3
Glc-6-P + E GE-(Glc-6-P) + E-P + Glc (1) k2 k4
De la même manière, le processus catalytique d'hydrolyse du pyrophosphate
inorganique par l'enzyme peut être défini par les réactions 3 et 4 et implique aussi le
transfert du phosphate de l'ester phosphate à i'enzyme, puis à l'eau.

PPi + E+E-@Pi)-&P + Pi (3)
E-P+HzO+E+Pi (4)
A i'état stationnaire, la vitesse d'hydrolyse du Glc-6-P (v,) doit être directement
proportionnelle à la concentration de I'itermédiaire phosphorylé, [E-Pl, Le. v, =
k{[w][E-Pl, où k5' représente la valeur du taux de renouvellement du processus
catalytique de l'enzyme. En considérant la concentration de H n constante, k<[H2G]
s'exprime alors comme ks, et peut être évlué en calculant V, I[E-Pl. Comme la
formation de E-P est directement et quantitativement proportionnelle à l'activité Glc-6-
Pase (Countaway er al., 1988), ks' a pu être estimé autour de 700 sec-', ce qui est très
rapide et ne permet pas, par ailleurs, de réaliser des études cinétiques sur la formation
et l'hydrolyse de E-P. En prenant pour acquis que l'intermédiaire phosphoqlé a un M,
de 36,500 et en se plaçant à des concentrations saturantes de Glc-6-P, il est possible
d'estimer la quantité totale d'enzyme Glc-6-Pase à environ 0.07% des protéines
microsomales totales. Cette dernière observation suggère, par contre, la nécessité d'une
purification d'au-dessus de 1000 fois pour espérer obtenir une protéine pure et
homogène (Countaway et al., 1988).

11
1 3 . 3 Transport et accumulation du glucose à l'intérieur des microsomes
Récement, Waddell et aL (1991) ont identifié un nouveau transporteur de
Glc de 52 kDa dans les microsomes de foie de rat. Ce transportair, dénommé GLüT7,
a été purifié puis clôné et sa séquence déduite de I'ADNc correspondant (Waddeii et
al., 1992). Ii représente 68% d'identité avec GLUT2. L'expression de GLUT7 dans des
cellules COS 7 a permis de montrer une localisation subceiiulaire de la protéine au sein
du RE (WaddelI et al., 1992). La question qui se pose alors consiste à savoir si GLUT7
correspond au tramporteur T3 des modèles proposés plus loin (section 1.4.2).
Berteloot et al. (1 99 1 a) ont montré que le w-l4~]Gk provenant de l'hydrolyse du CU-
"CJGIC-6-P s'accumule dans les microsomes de foie de rat et atteint un plateau. La
valeur de ce plateau diminue lorsquiin excès de Glc-6-P non radioactif est ajouté,
indiquant ainsi qu'a I'équilibre #accumulation, l'hydrolyse du Glc-6-P et la production
de Glc dans la lumière des microsomes est suivie d'un efflwc de Glc des vésicules
(Waddell et al., 199 1, St-Denis et al., 199%). Aucune capture de radioactivité n'a
néanmoins pu être observée dans les rnicrosomes incubés en présence de [3H]GIc ou de
[U-'~C]GIC (Rornaneili et al., 1994). Ces résultats, indiquant que le transport de Glc est
unidirectionnel dans la membrane des microsomes, suggèrent que les propriétés du
transport de Glc associé à la Glc-6-Pase sont différentes de ceiles des transporteurs de
Glc conventionnels, qui sont réversibles. St-Denis et al. (199Sb) ont donc émis
l'hypothèse que I'accumulation de Glc, produit lors de I'hydrolyse du Glc-6-P, est liée à
i'activité de la Glc-6-Pase, alors que son efflux peut se produire de manière
indépendante. Des considérations subséquentes, tenant compte des caractéristiques des
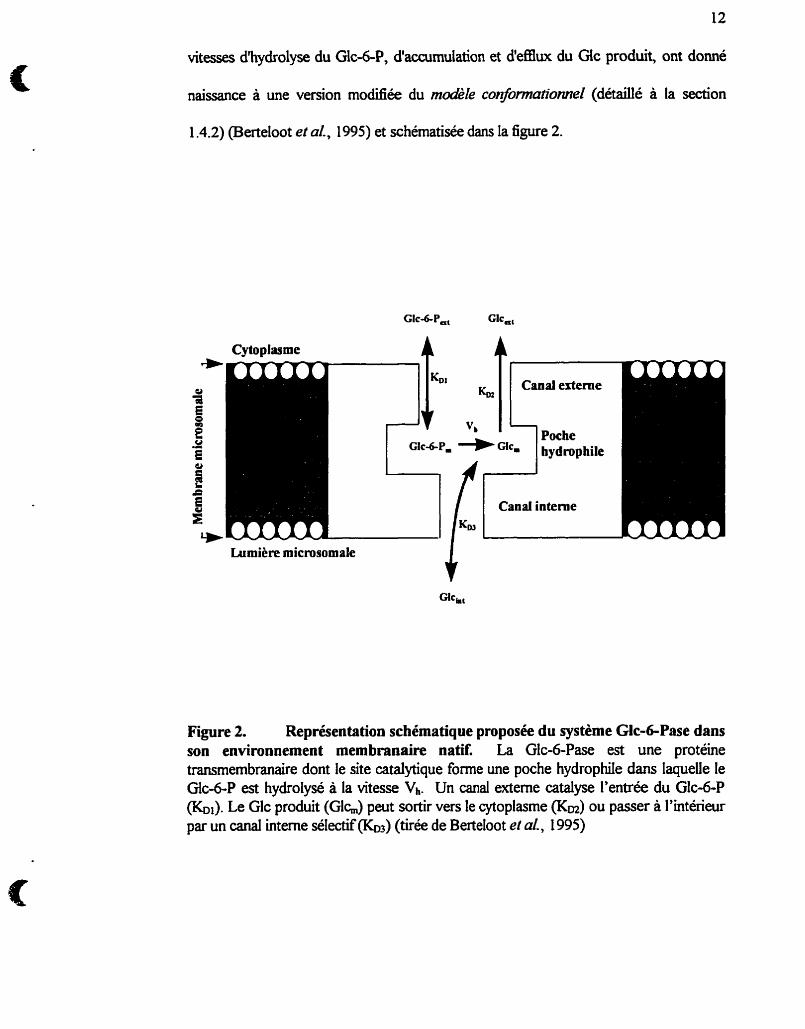
12
vitesses d'hydrolyse du Glc-6-P, d'accumulation et d'e£Eiux du Gic produit, ont donné
naissance à une version modiiée du modèle conformutionne1 (détaillé à la section
1.4.2) (Berteloot et al., 1995) et schématisée dans la figure 2.
Cytoplasme 4 4
Lumière microsomale C
Figure 2. Représentation schématique proposée du système Glc-GPase dans son environnement membranaire natit La Glc-6-Pase est une protéine transmembranaire dont le site catalytique forme une poche hydrophile dans laquele le Glc-6-P est hydrolysé à la vitesse Vh. Un canal externe catalyse l'entrée du Glc-6-P (&). Le Glc produit (Glh) peut sortir vers le cytoplasme (Km) ou passer à l'intérieur par un canal interne sélectif&) (tirée de Betteloot et a[., 1995)

13.4- Un nouveau concept du phénomène de latence
Le phénomène de latence de la Glc-6-Pase fat référence à l'augmentation
d'activité d'hydrolyse du Glc-6-P induite lors de la peméabiiisation de la barrière
membranaire. En effet, tel que prédit par le m d I e de transport du subsirat, l'étape de
transport du Glc-6-P dans les vésicules intactes cofere la spécificité pour le substrat et
restreint la vitesse d'hydrolyse du Glc-6-P par î'enzyme. Cette étape étant court-
circuit& lors de la solubilisation des microsomes, l'unité catalytique a dors libre accès a
son substrat et perd de sa spécificité en hydrolysant divers autres esters phosphates
dont le Man-6-P. L'activité Man-6-Pase a donc été utilisée comme indice de la
peméabiiité membranaire, mais est cependant remise en question par les observations
que nous rapportons dans l'article 2 (St-Denis et al., 1995a).
En utilisant un appareil d'échantillonnage et de filtration rapide, Berteloot et al.
(199 la) ont montré que les cinétiques rapides de la Glc-6-Pase sont caractérisées par
une surproduction temporaire ("burst") de Glc et de Pi par des rnicrosomes intacts
durant les 15 premières secondes de l'hydrolyse du Glc-6-P suivie d'une période où
l'activité de la Glc-6-Pase décroît dans les rnicrosomes intacts pour atteindre une vitesse
stationnaire plus fâible. La solubilisation subséquente de la membrane microsomale à
I'aide d'un détergent rétablit des propriétés cinétiques de la Glc-6-Pase semblables à
celies observées pendant les premières secondes d'hydrolyse du Glc-6-P dans les
microsornes intacts. Enfk, l'égalité des vitesses initiales d'hydrolyse du Glc-6-P entre
les microsomes intacts et les vésicules traitées au détergent (Berteloot et al., 1991a)
indique que le Glc-6-P a un accès similaire au site actif de la Glc-6-Pase dans

14
des microsornes intacts et perméabilisés pendant les premières secondes d'incubation. A
partir de ces résultats, il a été conclu: i) que la latence de la Glc-6-Pase dans les
microsornes intacts est compatible avec le concept d'une lente transition hystérétique,
ü) que la solubiIisation de la membrane microsomale à l'aide de détergent empêche
cette transition et üi) que le transport de Glc-6-P n'est pas restrictif dans les
microsornes intacts.
1.4 Topologie du système glucose6-phosphatase dans la
membrane du réticulum endoplasmique
1.4.1- Identifution et caractéristiques des composantes du système glucose-6-
phosphatase
Considérée d'abord comme une enzyme unique, la Glc-6-Pase requiert, pour
être fonctionnelle, un ensemble complexe de protéines. L'enzyme elle-même est
localisée dans la membrane du réticulum endoplasmique lisse, possiblement stabilisée
par une protéine de 2 1 kDa (Waddell et Burcheii, 199 1). Mais il faut que lui parvienne
son substrat, le Glc-6-P ou du pyrophosphate, et que soient ensuite évacués les
produits de la réaction, Pi et Glc, ce qui réclame l'action de trois protéines de transfert
ou translowes, Tl, T2 et T3 (définies plus loin), localisées dans la membrane du RE.

15
Une diEcuIté partidère liée à l'identification de la composante d y t i q u e de
la Gic-6-Pase a été son extrême instabilité en absence de phospholipides (Gariand et
Cori, 1972), qui entraînait son inactivation lors des maintes tentatives de purification
(Bickerstaff et Burcheil, 1980, Burchell et Burcheü, 1982, Rymsa et de Groot, 1988,
WaddeU et Burchell, 1991, Speth et Schulze, 1992, Kosel et al., 1993). Le poids
molécuiaire de la Glc-6-Pase de foie humain ou de rat a été évalué a 63 kDa (Kosel et
al, 1993) par migration des protéines microsomales sur un gel de polyacrylade en
conditions non-dénaturantes, et à 77 kDa dans des expériences d'inactivation par
irradiation (Collipp et al., 1974, Ness et al., 1989). Puis, diiérents protocoles de
purification ont permis d'estimer à 18,s kDa (Burcheii et BurcheU, 1982) et à 58-64
kDa (Reuek et Viee, 1982) le poids moléculaire de la Glc-6-Pase. Ce n'est que tout
récemment que, par une combinaison de marquage au 3 2 ~ et d'anticorps monoclonaux
obtenus à partu d'une préparation partieliement pur%&, on a pu la caractériser sous
forme d'une protéine de 36,5 kDa chez le rat (Countaway et al., 1988, Mthieux et al.,
1995) puis chez l'homme (Burcheil et al., 1988). Speth et Schulze (1 W2), en stabilisant
la Glc-6-Pase à i'aide de pyridoxal 5'-phosphate, ont pu obtenir une fraction purifiée
d'environ 700 fois de la Glc-6-Pase et ont évalué sont poids moléculaire à 3 5 kDa.
La présence d'un transponeur microsoma1 de Glc-6-P (Tl) a d'abord été
proposé par le groupe d'Arion lors de la présentation du modèle de tran.spori &
substrat (Arion et al., 1975). Cette protéine serait responsable du transport spécifique
du Glc-6-P dans la lumière des microsornes et cette étape serait lirnitante dans la
réaaion globale de transformation du Gb6-P en Glc et en Pi. La seule tentative de

caractérisation du transporteur putatif Tl a été rapportée par le groupe de Kamovsky
(Zoccoli et al., 1982). Ce groupe a identifié un polypeptide de 54 kDa à I'aide de
[ ~ I D S , un inhibiteur de Iliydrolyse du Gic-6-P dans des microsornes intacts, et qui
serait imptiqué dans le transport rnicrosomal du Glc-6-P. L'identité de ce polypeptide
reste, par contre, controversée étant donné que sa purification et sa reconstitution dans
des liposomes, pour tester le transport de Glc&P, n'ont pas encore été réalisées. De
plus, Ananthanarayanan et al. (1988) ont montré que la protéine de 54 kDa marquée
par le [)WDIDS était en fâit un transporteur d'anions organiques indépendant du Na'.
Des marquages au [QPLP (Speth et Schulze, 1992) ainsi qu'au FP]G~C-6-P
(Mithieux et al., 1995) ont pennis d'identifier des polypeptides de 54 D a possiblement
impliqués dans le processus de reconnaissance et/ou d'hydrolyse du Glc-6-P.
Un transporteur T2 du PPi (ou de Pi) de part et d'autre de la membrane du RE,
a pu être identitié et purifié à l'aide d'anticorps dirigés contre un antiporteur
mitochondrial de PVOK (Waddell et al., 1988). L'existence de T2, une glycoproteme
de 37 Da, est beaucoup moins contestée que celle de Tl. Des études subséquentes
par le groupe de Nordlie (1992) ont permis de conclure que T2 est formé de 2
isofomes, T 2 a pour le transport de Pi généré par i'hydrolyse du Glc-6-P, et T2B pour
le transport du PiPPVcarban~yI-P.
Pour expliquer la sortie du Glc du RE hépatique après hydrolyse du Gic-6-P,
un transporteur de Glc a été postulé (T3) (Arion et al., 1980). Tel que décrit plus haut
(section 1.3.3), un tel transporteur a été identifié dans le foie de rat (Waddeil et al.,
199 1) puis purifié, cloné et exprimé dans des cellules COS 7 (Waddeli et al., 1992).

17
Les résultats obtenus suite aux transftxtions et les caractéristiques de ce
transporteur de glucose microsornai sont controversés par le fhit que le système
d'expression (ceUules rénales COS) choisi par les auteurs powait déjà posséder
certaines composantes du système Gic-6-Pase a que ses caractéristiques pourraient
être modulées par la présence d'autres éléments du système Glc-6-Pase.
1.4.2- Modèles proposés pour l'organisation moléculaire du système
glucose-6-p hosphatase
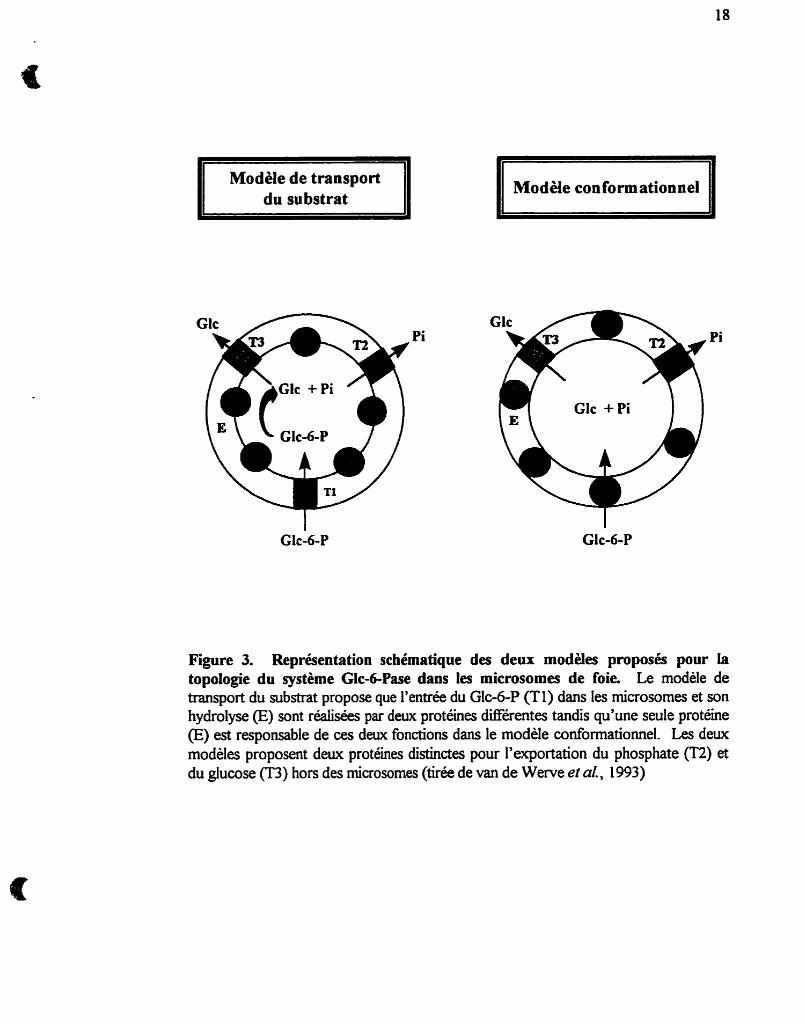
La figure 3 montre deux des modèles les plus courants de l'organisation
moléculaire du système Gic-6-Pase dans les microsomes. Les deux modèles prévoient
que l'hydrolyse du Glc-6-P mène a la production de Glc et de Pi dans l'espace
intramicrosomal (correspondant à la lumière du RE) et que leur transport vers le
cytosol (milieu extramicrosomd) soit réalisé par des transporteurs distincts pour le Pi
(T2) et pour le Glc (T3). Le modèle de banport dir substrat (Arion et al., 1975) est
basé sur les postulats suivants. (i) La sous-unité d y t i q u e de la Glc-6-Pase est une
phosphohydrolase non spécifique, localisée sur la face interne de la membrane
microsomale. Par conséquent, la membrane empêche l'accès du Glc-6-P au site actif de
l'enzyme. (ü) Pour expliquer l'hydrolyse du Glc-6-P dans les microsomes intacts, les
chercheurs ont pris pour acquis que le transporteur Tl reconnaît spécifiquement le Glc-
6-P et lui Fdt traverser la membrane pour lui permettre I'a& au site actif' et que, ci) le
transport de Glc-6-P par Tl est l'étape limitante de Iliydrolyse du Glc-6-P par la Glc-6-
Pase.
II Modèle de transport du substrat II II Modèle con form ationnel Il
Figure 3. Représentation schématique des deux modèles proposés pour la topologie du système Glc-dPase dans les microsornes de foie. Le modèle de transport du substrat propose que l'entrée du Glc-6-P (Tl) dans les microsornes et son hydrolyse (E) sont réalisées par deux protéines différentes tandis qu'une seule protéine (E) est responsable de ces deux fonctions dans le modèle conformationnel. Les deux modèles proposent deux protéines distinctes pour l'exportation du phosphate (T2) et du glucose (T3) hors des microsumes (tirée de van de Wente et al., 1993)

19
Quelques propriétés cinétiques du système giucuse-6Pase donnent
apparemment crédit au modèle de @amport du substrat. Le traitement des microsomes
avec des détergents augmente, en éliminant la barrière membranaire, l'activité de la Glc-
6-Pase (concept de la latence) en accord avec les postulats (i) et ci). On observe
d'aiüeurs une perte de spécificité de l'enzyme pour le Glc-6-P dans des vésicules
perméabiiisées, conformément aux postulats (i) et ci). La présence de Glc-6-P Libre n'a
cependant jamais été démontrée bien que la perméabilité des microsornes intacts au
Glc-6-P ait été indirectement suggérée au moyen de la technique de difiction de la
lumière (Fulcmi et ai., 1992).
Des études cinétiques récentes apportent plusieurs arguments contre
l'organisation moléculaire de la Glc-6-Pase telle que prédite par le morlée de bansport
di substrat et ont donné naissance au m d l e conf~rrn~oiineZ (Schulze et d, 1986,
Berteloot et al., 199 1 a, Speth et Schulze, 1992, Berteloot et al., 1995) qui propose que
la Glc-6-Pase puisse exister sous différents états conformationnels avec des propriétés
cinétiques distinctes. Par modification de sa conformation dans la membrane pendant la
catalyse, la Glc-6-Pase serait capable de fixer son substrat du côté cytosolique des
microsomes et de libérer les produits de la réaction dans la lumière. D'après ce modèle,
le traitement par un détergent entherait une augmentation de l'activité Glc-6-Pase en
modifiant les interactions entre i'enzyme et certains constituants lipidiques
(phospholipides) de la membrane indispensables au fonctionnement normal de la Glc-6-
Pase (Nordlie et Sukalski, 1985, Stetten et Bumett, 1967, Zakim et Dannenberg, 1990,
Schulze et al., 1986, Ness et al., 1989, Speth et Schulze, 1992).

1.5- Clonage du gène codant pour la glucos4-phosphatase
1.5.1- Clonage de la sous-unité catalytique
Le clonage de I'ADNc de la Glc-6-Pase de foie de souris n'a &é r é s é que
récemment (Shelly et al., 1993). Les auteurs ont isolé et caractérisé le gène de la Gioo-
Pase en criblant différentiellement une banque d'ADNc avec de I'ARNm total isolé du
foie de souris normales et albinos, déficientes en activité Glc-6-Pase (Glueckson-
Waelsch et al., 1979). La protéine encodée par cet ADNc, lorsqu'exprimée dans des
cellules COS- 1, possédait les propriétés attendues de la Glc-6-Pase (paramètres
cinétiques, latence, stabilité thermique, inhibition par le vanadate). Le gène couvre une
région d'environ 10 kb et est constitué de cinq exons : I (3 1 1 pb), II (1 IO pb), III (106
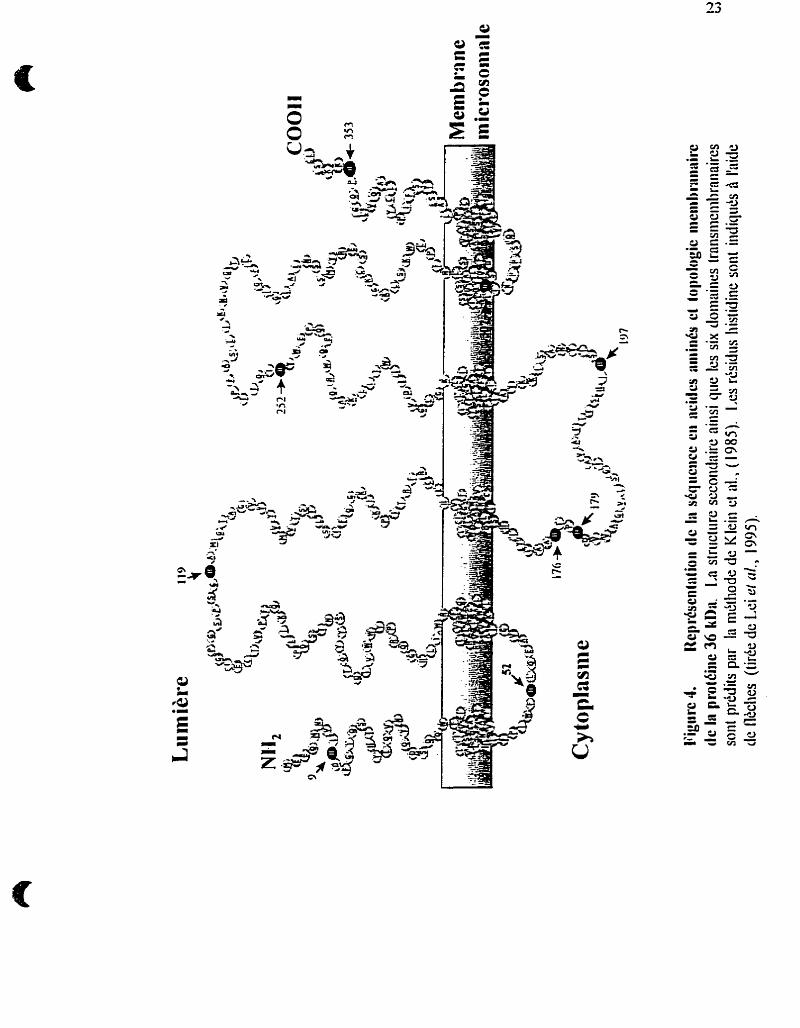
pb), IV (1 16 pb) et V (161 5 pb). Le polypeptide encodé par cet ADNc possède un
caractère hautement hydrophobe et contiendrait 6 domaines transmernbranaires, ainsi
que 2 résidus lysine (KK), en position 3 et 4 de l'extrémité C-terminale, formant la
séquence requise pour la rétention des protéines à la membrane du RE. Trois sites
potentiels de glycosylation sur les résidus asparagine (UYS) suggèrent que la Gic-6-
Pase soit une glycoproteme (voir figure 4). Le poids moléculaire prédit de la protéine
est de 40 kDa et semble compatible avec celui d'environ 37 kDa déterminé par
autoradiographie de la [ 3 2 ~ ] ~ l c d - ~ a s e (Countaway et al., 1 988, Mithieux et al., 1 995).
Ce même groupe a, par la suite, isolé à l'aide d'une paire d'amorces oligonucléotidiques
un ADNc codant pour la GlcdPase humaine et a localisé le gène sur le chromosome
17 (Lei et ai., 1994). Depuis, I'ADNc de rat a été isolé et séquencé par d'autres

21
groupes (Lange et d , 1994, Haber et al, 1995, Mithieux et al.. 1996). Enfin, il existe
9 résidus histidine cornés dans la séquence primaire de la Gic-6-Pase et qui
pourraient être impliqués dans la formation de l'intermédiaire phospho-histidine fomé
lors du processus catalytique de la GIA-Pase. Seule la mutation Hl 19A résulte en une
perte d'activité complète de l'enzyme (Lei et al., 1995). La mutation du résidu Arg-83,
situé du même côté de la membrane du réticulum endoplasmique que le résidu His- 1 19,
conduit aussi à l'inactivation totale de i'aaivité Glc-6-Pase suggérant que ce résidu soit
important pour la stabiiisation de i'iitermédiaire phosphorylé transitoire.
1.5.2- Clonage du promoteur
Ii a été rapporté récemment environ une vingtaine de mutations différentes
toutes localisées dans la région codante du gène de la Glcd-Pase chez les patients
atteints de glycogénose de type la (Lei et al., 1998. Par contre, cette même étude
rapporte que dans 17% des cas de giycogénoses de type la aucune mutation n'a pu être
détectée. L'explication la plus plausible de ces observations serait la présence de
mutations situées dans le promoteur du gène codant pour la Glc-6-Pase. Le clônage et
le séquençage des premières 1.2 kb de la région 5' du promoteur de la Glc-6-Pase n'a
cependant été réaiisé que tout récemment (Schrnoii et al., 1996). La régulation
transcriptionneile du gène a aussi été récemment étudiée dans diérents états
nutritionnels et hormonaux (Schrnoii et al., 1996, Argaud et al., 1996). Plusieurs sites
de liaisons pour diérents facteurs transcxiptionnels ont été dénombrés. Ceux-ci
incluent 3 sites où se trouvent des séquences stimulatrices de transcription CCAAT

22
(CWP), 9 sites de liaisons pour facteurs nucléaires hépatiques 0, 3 sites AP-1
fixant chacun le complexe/un/fos, 2 sites pour l'élément de réponse à I'AMPc (CRE), 1
site pour l'élément de réponse à I'insuline (RE), et un site pour l'élément de réponse
aux glucocorticoides (GRE). La présence d'autant de sites de régulation
transcriptio~eiie est donc un reflet du haut degré de régulation du gène de la Gic-6-
Pase. Les niveaux d'ARNm de la Glc-6-Pase ont donc pu être quantifiés et mis en
parallèle avec les niveaux d'activités enzymatiques. Il a été, par contre observé, que le
glucocorticoïde dexarnethasone augmentait les niveaux d'ARNm et I'activité de la Glc-
6-Pase, mais que paradoxalement, l'analogue des glucocorticoïdes triamcinolone,
pourtant réputé augmenter l'activité Glc-6-Pase (Traxinger et Nordlie, 1990),
n'induisait aucune augmentation des niveaux dARNm de l'enzyme (Argaud et al.,
1996). Dans ce dernier travail, ahsi que dans les observations de Shingu et al. (1996),
il a été rapporté une partie de la région 3'-non-traduite (0,s kb) chez le rat qui est
absente de la séquence du gène cloné par le groupe de Chou chez la souris (Shelley et
al., 1993) et par le groupe de Pilkis chez le rat (Lange et al., 1994). De ses
obsetvations, il se pourrait qu'il y ait deux formes d'ARNm provenant de deux gènes
codant pour des formes distinctes de la Glc-6-Pase.

1.6- Déficiences génétiques du système glucose-&phosphatase
1.6.1- Manifestations ciiniques et métaboiiques des giycogénoses
Les glycogénoses constituent un groupe de maladies génétiques affkctant les
voies métaboliques de mise en réseme des hydrates de carbone sous forme de
giycogène et I'utilisation du glycogène pour le maintien de la glycémie et la fourniture
d'énergie. Certaines ne sont pas associées à une augmentation du contenu en glycogène
des tissus mais plutôt à une anomalie de structure de ce dernier. Le foie normal
contient 10-50 mg glycogèndg de tissu. Dans les glycogénoses hépatiques de type I,11
et VI, on trouve une augmentation du glycogène et des lipides intrahépatiques. Les
types III et N sont associés à une anomalie de la structure du giycogene et il peut y
avoir une cirrhose du foie. 11 faut, pour porter le diagnostic, une étude biochimique et
enzymatique du tissu hépatique.
Les maladies du métabolisme du glycogène sont des états héréditaires qui sont
la conséquence d'une déficience de certaines enzymes spécifiques impliquées dans la
synthèse ou la dégradation du giycogène. Le tableau clinique et la nature des organes
impliqués dépendent du déEiut enzymatique spécifique et de sa distribution tissulaire.
Le tableau 1 résume les diffërents types de glycogénoses et leurs principales
caractéristiques.
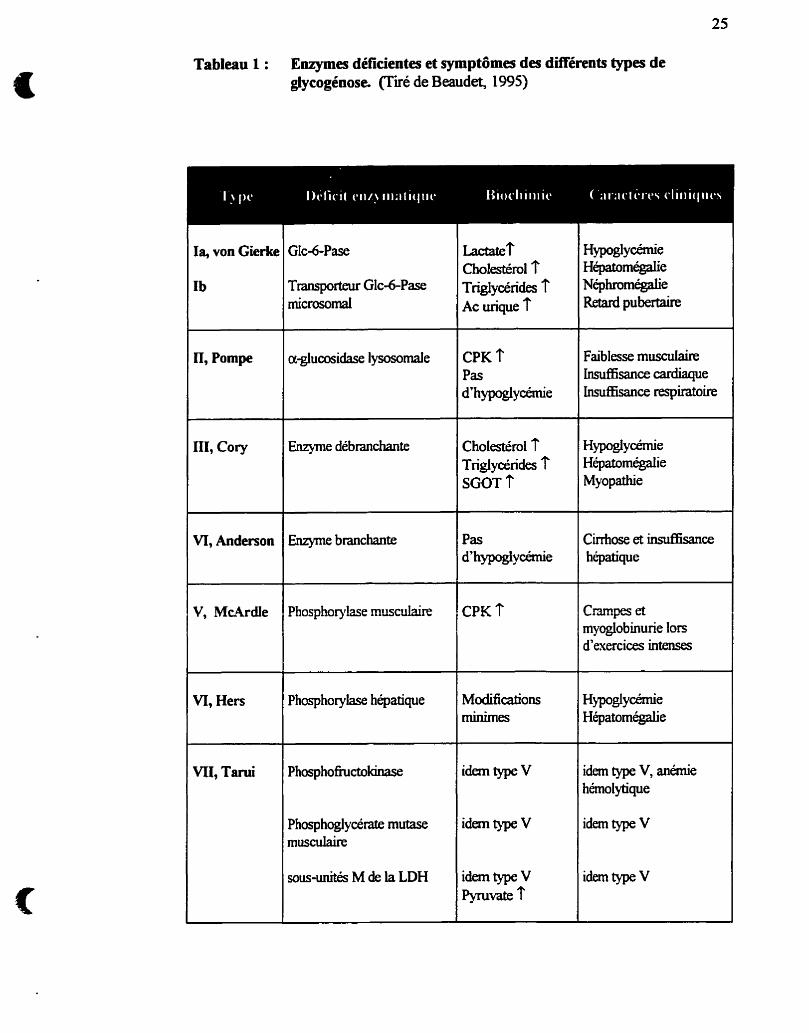
Tableau 1 : Enymes déficientes et symptômes des différents types de glyeogénose (Tiré de Beaudet, 1995)
~actater Cholestéml -r Trigiycérides f Ac urique î
Hypoglycemie Hépatomégalie Néphromégalie Retard pubertaire
Ia, von Gierkt
Ib Transporteur Glc-6-Pase rnicrosomal
Faiblesse musculaire Insufnsance cardiaque Insuf]6sancerespiratok
CPK ? Pas d'hypoglycémie
II, Pompe
Cholestérol T Triglycérides f SGOT 7
Hypoglycémie Hépatomégahe Myopathie
nr, coq Enzyme débranchante
Cirrtioseetï.rldkmce hépatique
Pas d'hypoglycémie
VI, Anderson Enzyme branchante
CPK ? Phosphoqlase musculaire C m p e s et myoglobinurie lors d'exercices intenses
Modifications minimes
Hypoglycémie Hépatomégahe
VI, Hers Phosphorylase hépatique
idem type V idem type V, anémie hémolytique
idem type V idem type V musculaire
jousiuiités M de la LDH idem type V I'yTllvate 7
idem type V

1.6.2- Bases moléculaires des glycogénoses de type 1
1.6.2.1- Déficit en Glc-&Pas% type Ia
Le déficit en Gic-6-Pase, ou maladie de von Gierke, est une anomalie génétique
transmise comme un caractère autosomique récessif dont i'incidence est de 1 cas sur
100 000 à 400 000 naissances (Hers et ai., 1989, Beaudet, 1995) et représente près de
90% des glycogénoses de type 1. La maladie se manifeste habituellement dans les 12
premiers mois de la vie par des hypoglycémies symptomatiques sévères (giycérnies
inférieures à 0.8 rnmoVl ou 15 mg/dl) ou par la découverte d'une hépatomégalie. Le
tableau clinique tend, par contre, à s'améliorer si le patient peut h c h i r les premières
années. Un renouveau d'intérêt pour le déficit en Glc-6-Pase a, de plus, été suscité en
i'associant à l'ensemble des symptômes de la mort subite du nourrisson (Emery et ai.,
1988, Burchell et al., 1989).
L'absence de Glc-6-Pase hépatique provoque un déficit de la production
hépatique de Glc tant à partir de la glycogénolyse que de la néoglucogenèse, encore
qu'environ 6-8% des résidus glycosyl liés en 1,6 aux points de branchement du
glycogène puisse être Libéré du foie par I'amylo- 1,6-glucosidase (enzyme débranchante
du glycogène). Une hypogScémie sévère est observée pendant le jeûne.
L'hypoglycémie chronique est associée à une mobEsation des lipides et à une
cétonérnie accrues. Une hyperlipidérnie est fréquente et comporte une élévation à la
fois du cholestérol et des triglycérides lors des périodes d'absorption insuEsante
d'hydrates de carbone. Il peut y avoir une acidose métabolique chronique à cause de la
production augmentée d'acide lactique et de corps cétoniques. L'hyperuricémie est

27
caractéristique et reflète un défaut d'excrétion rénale secondaire à l'augmentation des
taux d'acides lactique et @hydroxybutinque associée à la production accrue d'acide
urique. L'ensemble de ces symptômes suggère le diagnostic de glycogénose de type 1.
Des tests fonctionnels sont utiles pour établir le diagnostic adéquat. La
glycémie des malades atteints de glycogénose de type 1 n'est pas augmentée à la suite
de l'administration d'adrénaline ou de glucagon. Comme la dégradation du glycogène
est normale mais que le glucose libre ne peut être généré, la production de lactate est
augmentée par ces hormones, ce qui exagère l'acidose métabolique. L'administration de
fnictose, de galactose ou de glycirol augmente le lactate plasmatique mais pas le
glucose. Généralement, le diagnostic de glycogénose de type 1 ne peut être fait avant la
naissance et il n'existe pas, à ce moment, de traitement spécifique. Le diagnostic
biochimique exige, entin, que soit démontrée l'absence d'activité Glc-6-Pase sur des
biopsies correctement prélevées et traitées de foie, de muqueuse intestinale ou de rein.
Le dosage enzymatique doit, de plus, comporter un contrôle d'activité phosphatase
non spécifique.
D'un point de vue moléculaire, la glycogénose de type Ia est causée par la
déficience de l'unité catalytique de la Glc-6-Pase. L'activité Glc-6-Pase est absente dans
les microsornes de foie intacts et perméabilisés. Le clonage de l'unité catalytique a
permis d'identifier plusieurs mutations dans les régions codantes du gène chez les
patients soufüant de glycogénose de type Ia, résultant en l'absence totale ou en la
réduction de l'activité Glc-6-Pase. La plupart de ces patients possèdent plusieurs
mutations ponctuelles faux sens, les plus communes étant : R83C dans la population

28
juive (Parvari et al., 1999, G327A dans la population chinoise (Hwu et al , 1995), et
G727T dans la population japonaise (Kajihara et al., 1995) ou celles pemirbant le
cadre de lecture par insertioddélétion. Par contre, une étude sur 140 allèles a permis
d'en identier 17 qui ne contenaient pas de mutations dans les régions codantes du gène
(Lei et al., 1995). Des mutations dans les régions non-codantes du gène de la Glc-6-
Pase, pouvant &er les taux de transcription et de traduction ou la stabilité de
l'ARNm, ont par ailleun déjà été rapportées (Chevalier-Porst et al., 1996).
1.6.2.2- Déficit en transporteur de GIc-6-P microsomal, type Ib
Le déficit présumé en transporteur de Glc-6-P microsomal (Tl) a une incidence
de 1/80 à 1/500 de ceile du type Ia Les caractères cliniques sont similaires a ceux du
type Ia, mais comprennent, en outre, une neutropénie, des anomalies de la migration
des neutrophiles et des Uifdons récurrentes (Gitzelmann et Bosshard, 1993); en
général, le type Ib est plus sévère que le type la.
Le type Ib a été initialement distingué du type Ia par la présence d'une activité
Glc-6-Pase normale lors du dosage sur biopsie tissulaire en présence de détergent
(Narisawa el al., 1978, Schaub et Heyne, 1983, Nordlie et SuMski, 1985, Burcheil,
1990). Cependant, l'activité Glc-6-Pase est basse dans le type Ib quand les tissus fiais
sont homogénéisés et les dosages réalisés en I'absence de détergent. Ces résultats ont
été interprétés comme un déficit génétique de Tl à l'origine du défàut primaire de la
glycogénose de type Ib. La cause de la neutropénie et des anomalies de migration des

29
neutrophiles est inconnue, mais un défaut dans la capture et la phosphoqdation du Glc
a été identifié dans des leucocytes de patients atteints de glycogénose de type [b
@ashan et al., 1993).
Une façon convaincante de démontrer l'absence fonctionnelle de TI serait de
mesurer le hansport du Glc-6-P dans les microsomes. Cette démonstration n'a pas été
fate jusqu'à maintenant. St-Denis et al. (1995b) ont montré, en utilisant un inhibiteur
de la Glc-6-Pase (le vanadate) qu'il nl avait pas de transport du Glc-6-P indépendant
de l'activité Glc-6-Pase et que I'acaimulation de radioactivité à partir du [u - '~c ]G~c-~-
P dans les microsomes (essentiellement du &J-"C]GIC) est directement proportio~eile
à i'aaivité Glc-6-Pase. De plus, des microsomes préparés à partir du foie d b e patiente
diagnostiquée glycogénose de type Ia, où seule la fonction d'hydrolyse du Glc-6-P
devait être déficiente, n'accumulent pas de radioactivité en présence de [u-"CIGIC-6-P
suggérant l'absence de transport de Glc-6-P dans ces rnicrosomes (St-Denis et al.,
1994). L'explication proposée par les auteurs est que dans ce cas de glycogénose la
même protéine, transportant et hydrolysant le Glc-6-P, est déficiente. Entre temps Lei
et a/. (1995) ont montré I'absence de mutations dans la réion codante du gène de la
Glc-6-Pase chez 3 patients de type Ib. Ces résultats sont en accord avec l'hypothèse
qu'une protéine autre que l'unité catalytique de la Glc-6-Pase soit impliquée dans la
glycogénose de type Ib, mais n'excluent pas, encore une fois, l'existence de mutations
dans les régions non-codantes du gène. Alternativement, la glycogénose de type Ib
pounait ètre dûe à un défaut d'insertion de la protéine Glc-6-Pase dans la membrane
microsomale, ou à un défàut post-traductio~el flêctant son interaction avec les lipides
de la membrane du RE.

1.6.2.3- Déficit en transporteur Pi/PPi microsornai, type Ic
La glycogénose de type Ic a été proposée comme étant causée par la déficience
en T2, le transporteur de phosphate et de pyrophosphate des microsornes (Nordlie et
al., 1983). L'absence de T2 dans les microsornes de foie de patients de glycogénose de
type Ic a été mise en évidence à raide d'anticorps spécifiques pour T2 (Waddeli et al.,
1988, Nordlie et al., 1992). Lei et al., (1995) ont montré l'absence de mutations au
niveau du gène codant pour l'unité catalytique Glc-6-Pase chez un patient atteint de
glycogénose de type Ic, chez lequel il n'y avait pas d'accumulation de pyrophosphate
dans les microsornes de foie. Dans ce cas-ci, c'est I'isoforme T2P qui serait responsable
de la glycogénose.
1.6.2.4- Déficit en transporteur Glc microsomal, type Id
Aucun cas de déficience du transporteur microsomal de Glc T3, nommée
glycogénose de type Id, n'a été observé à ce jour, quoique ce déficit ait été proposé sur
une base théorique (Burchell, 1994).

1.7- Contrôle nutritionnd et hormonal de In glucosedphosphatase
L'activité de la Gic-6-Pase est géneraiement considérée comme étant régi& par
la concentration de substrat parce que le Km de la Glc6Pase pour le Glc-6-P (2-5
mM) (Arion er al., 1976b) est plus élevé que le contenu intracellulaire en Gic-6-P (70-
500 nmoVg foie). Le flux de substrat qu'emprunte la voie de la Glc-6-Pase n'est
cependant pas toujours compatible avec les concentrations de Glc&P, en particulier
lorsque le rapport insuhe/glucagon est modifié (Newgard et al., 1984, Christ et al,
1986).
La déficience en uwline obtenue à la suite d'un jeûne prolongé ou par un
diabète expérimental (administration de streptozotocine ou d'doxane) augmente
l'activité de la Glc-6-Pase (Ashmore et Weber, 1959, Nordlie, 1976) et induit
probablement la synthèse de mvo de la sous-unité catalytique de I'enzyme (Burcheil et
Cain, 1985), parce que cette augmentation est plus évidente dans des rnicrosomes
perméabilisés à l'aide d'un détergent que dans des vésicules intactes (la latence
apparente est augmentée). L'administration de glucocorticoides à des rats augmente la
Glc-6-Pase de foie dans les rnicrosomes intacts plutôt que perméabilisés (la latence
apparente est diininuée), ce qui amène i'hypothèse que, dans ce cas, c'est la transiocase
putative Tl qui est augmentée (Sukalski et Nordlie, 1989). La formation de sous-unités
catalytiques altérées par les glucocorticoides a également été proposée (Traxlliger et
Nordlie, 1 990).
Avec la disponibilité de 1'ADNc de la Glc-6-Pase, plusieurs études ont pu être
réalisées sur les variations dans l'abondance de ilARNm de la Gic-6-Pase dans

32
différentes conditions et différents tissus. La variation dans le temps de l'activité de la
Glc-6-Pase pendant le j e h e est complexe. L'activité de la Gic-6-Pase hépatique
augmente suite à un jeûne de 24 et 48 heures @fimissan et Mithieux, 1994, Minassian
et d, 1994) en accord avec la capacité néogiucogénique qu'a le foie dans cette
situation. Par contre, l'activité de la GIc-6-Pase hépatique diminuera si le jeûne est
prolongé à 72 et 96 heures en accord avec la diminution de la production hépatique de
Glc qui suMent lors diin jeûne prolongé (Owen et al., 1969). Cette situation de jeûne
prolongé fit, par contre, augmenter la néoglucogénèse dans le rein et peut compter
jusqu'à 40% de la production endogène totale de Glc. Parallélement à la participation
accrue du rein dans la production de Gic durant le jeûne prolongé, l'activité de la Glc-6-
Pase, qui augmente dans le rein lors d'un court jeûne, continue à augmenter si le jeûne
est prolongé durant 2 à 3 jours (Minassian et Mithieux, 1994). Ces observations
révèlent le rôle important qu'a la Glc-6-Pase dans la néoglucogénèse hépatique et
rénale. En particulier durant la période du jeûne, ce rôle pourrait même s'avérer plus
important que celui de la phosphoénolpyruvate carboxykinase, pourtant considérée
comme une enzyme régulatoire (le senseur) de la néogiucogénese (Pontremoli et al.,
1974). Les variations de l'activité de la Glc-6-Pase dans les différents états
nutntiomek sont corrélées avec la quantité d'ARNrn (Minassian et al., 1996). De plus,
la normalisation des niveaux d'ARNm et de l'activité Glc-6-Pase, après la renutrition de
rats préalablement mis à jeun, se faisant plus rapidement dans le foie (moins de 90
minutes) que dans le rein (plusieurs heures) (Mithieux et al., 1996), il a été suggéré que
le contrôle de la Glc-6-Pase durant la période de jeûne s'exerçait principalement à un
niveau transcriptionnel.

33
De la même manière, les niveaux d'ARNm et d'activité de la Gic-6-Pase sont
augmentés dans le foie et le rein dans différents modèles de rats diabétiques (Liu et al,
1994, Mthieux et al., 1996, Massiiion et al, 1996). De plus, le Fdt de rétablir les
niveaux d'ARNm de la Glc-6-Pase par l'insuline dans les deux tissus diabétiques
suggère un rôle suppressif de I ' i e sur l'expression de la Glc-6-Pase (Mithieux et
al., 1996). EnfirS la normalisation des niveaux d'ARNm (Massillon et cd, 1996) mais
non de l'activité (Lavoie et al., 1992) de la Glc-6-Pase a été observé lorsque
l'hyperglycémie était supprimée par un traitement à la phioridzine. Ces résultats
suggérent un rôle du Glc dans l'induction de l'expression de la Glc-6-Pase mais dont la
signification physiologique n'est pas claire à cause de l'absence d'effet sur l'activité de
l'enzyme. Ils sont aussi contradictoires avec le fiiit que les niveaux d'ARNm et d'activité
de la Glc-6-Pase soient augmentés durant le jeûne (condition de glycémie basse), puis
normalisés suite à une renutrition (condition de glycémie élevée).
Les observations, in vitro, telles que l'inhibition par FUisuline de la Glc-6-Pase
dans des microsornes isolés (Gardner et al., 1993), ou dans des hépatocytes en culture
(Chnst et al., 1986), ainsi que les observations, in vivo, de la suppression totale et
concornmitante de la production de Glc hépatique avec seulement une légère
diminution de la concentration du Glc-6-P durant un clamp euglycémique (Minassian et
al., 1995) nous amènent à postuler l'existence d'un mécanisme d'inhibition de la Glc-6-
Pase sous le contrôle direct de l'insuline. Un tel mécanisme d'inhibition ainsi que la
possibilité d'un réglage hormonale par phosphorylation/déphosphorylation de la Glc-6-
Pase (Burchell et Burchell, 1980, Begley et Craft, 198 1, Burchell et al., 1982, Singh et
al., i 983) demeurent cependant encore des concepts incertains.

1.S Objectifs de la thèse
Les résultats de notre étude visent à comprendre le fonctionnement de la Gle
6-Pase depuis le niveau fondamental jusqu'à son implication en pathologie. Pour mener
à bien ce projet, trois approches expérimentales ont été utilisées : 1) I'étude cinétique de
l'enzyme in situ dans la membrane du RE par des mesures de production totale et
d'accumulation de Glc à l'intérieur des rnicrosomes, 2) la modification chimique et
spMque de certains résidus impliqués dans le mécanisme catalytique de l'enzyme et,
3) la purification en vue du clonage et du séquençage des composantes du système
Glc-6-Pase.
La première approche nous a permis d'étudier la relation entre les
caractéristiques cinétiques de la Glc-6-Pase et la perméabilité des rnicrosomes suite à
un traitement avec les histones K A et nous amène à réévaluer le concept de la latence
(article 1). De plus, la relation entre le transport de Glc et l'activité de la Glc-6-Pase
nous a permis de caractériser un transport unidirectionnel i e GIc (article 2). Les résidus
impliqués dans le processus d'hydrolyse du GIc-6-P ainsi que l'état dtoligoménsation de
la sous-unité catalytique de la Glc-6Pase ont été approchés par l'inhibition et par le
marquage radioactif avec le DCCD/GEE (article 3) et complérnentés par des études
imrnunologiques. Enfin, la présence de Tl, le transporteur de Glc-6-P microsornai, est
remise en question dans l'étude des fonctions de production et d'accumulation de Glc
dans le système Glc-6-Pase (article 4).

2.1- Préparation des microsomes de foie
Des rats Wistar mâles pesant de 150 à 200 g sont mis à jeun 48 heures puis
sacrifiés par décapitation. Le foie est excisé rapidement et homogénéisé a i'aide diin
homogénéisateur Yamato LH-21 a 1200 rpm dans 5 volumes (v/w) d h tampon
HEPES IO mM, Sucrose 250 mM, EDTA 1 rnM, EGTA 1 mM, dithiothreitol2 mM,
Benzamidine-HCl 2 mh4, phenyhnethanesulfonyl fluoride 0.2 mM, leupeptine 0.5
mghi à pH 7,3 maintenu à 4OC pour avoir une solution finale de 20% (w/v). Deux
centrifùgations ou l'on garde le surnageant à chaque fois, l'une pendant IO minutes à
1,000 x g et l'autre 10 minutes à 12,000 x g, précèdent une troisième centrifugation à
100,000 x g pendant 90 minutes à 4OC, qui nous permet de précipiter la fiaction
microsornale contenant un mélange hétérogène de vésicules e ~ c h i e s en microsornes
tisses et en microsomes rugueux. Le dernier culot est resuspendu dans un tampon
HEPES 20 m!M, EC&POa 1 mM, EDT.4 1 mM, EGT.4 0.5 rnM, dithiothreitol 1 mM,
Benzamidine-HC12 mM, phenylmethanesulfonyl fluoride 0.1 rnM, leupeptine 1 mglml
giycerol 20% à pH 7,3 maintenu à 4OC, et peut être utilisée directement pour les
différentes mesures enzymatiques ou de transpoK ou comme matériel de départ
servant à la purification de la Glc-6-Pase. Alternativement, les microsomes peuvent être
répartis en échantillons et congelés à -80°C. La même procédure est utilisée pour la
préparation de microsomes à partir de foie de souris ou de biopsies de foie humain.
Aucune perte d'activité Glc-6-Pase ou de capacité de transport n'a été observée dans
des microsomes ayant été congelés pendant plusieurs semaines.

2.2- Mesure des activités enzymatiques
L'activité de la Glc-6-Pase est mesurée à 30°C dans un tampon Tris-HCI 50
mM à pH 7.3 par la production de [u-'~c]G~c à partir de différentes concentrations de
W-'~C]G~C-~-P. La réaction est arrêtée en ajoutant cinq volumes de suffite de zinc
0,3N suivi de cinq volumes dliydroxide de barium saturé. Le précipité zindbariurn
formé entraine avec lui les esters phosphate tels que le Glc-6-P et une fiaction du
surnageant est récoltée pour mesurer le W-~~C]GIC. Il est à noter qu'avec cette
méthode on mesure le W - ~ ~ C ] G ~ C total dans le milieu d'incubation qu'il soit présent à
l'intérieur ou à l'extérieur des microsomes au moment de l'arrêt de la réaction. L'activité
de la mannose-6-phosphatase est mesurée de la même façon que l'activité de la Glc-6-
Pase mais en remplaçant le ~ - 1 4 ~ ] ~ l c - 6 - ~ par le W - ~ ~ C ] M ~ ~ - ~ P . La synthèse du CU-
13 ClMan-6-P a été réalisée a partir de [ u - ' ~ c ] M ~ ~ (300 mCVrnrnol), dATP-Mg et
d'hexohase selon la méthode de Forsyth et al. (1993). La réaction a, cependant, été
arrêtée avec du charbon actif à 1%, suivie diine centntigation de 1 min a 14,000 rpm
dans une centrifugeuse Eppendorf. Le surnageant a ensuite été chargé sur une colonne
de Dowex AG 1x8 sous sa forme acétate. Le W - ' ~ C ] M ~ ~ qui n'a pas réagi élue dans
le volume mort de la colome, don que le [u - '~c ]M~~-~-P est élué à l'aide d'un
gradient dacide acétique (0,2-1 M). Le [u-~~c]M&P est ensuite concentré mus
vide et resuspendu dans 4 m M Man-6-P fioid (solubilisé dans du Tris-HCl50 mM, pH
7,3) à une activité spécifique hale de 2.4 nCilnmo1.

2.2.2- Phosphoglucomutase
Une des isoenzymes de la phosphoglucomutase (PGicM) de foie est associée à
la membrane du RE (Mithieux et al, 1995). La PGlcM catalyse la formation du Gic à
partir du Glc-1-P. Cette enzyme est phosphorylke et le groupement phospho- prend
part à une réaction réversible où le Glc-1,6-Pz est un intermédiaire. La PGlcM est
mesurée à 30°C dans un tampon HEPES 50 mM à pH 7.1 contenant 0.2 mM NADP,
5 rnM MgC12, 50 mM Glc- 1 ,6-P2, 1 m . Glc- 1-P. La réaction est lancée avec 10 mu
de glucose-6-phosphate deshydrogénase et la production de NADPH, lors de la
conversion du Glc-6-P en gluconolactone-6-P est mesurée au spectrophotomètre à 340
nm.
2.3- Purification de la glucose-6-phosphatase
23.1- Solubiüsation des microsornes à l'aide de différents détergents
Différents détergents ont été étudiés tant pour leur pouvoir de solubilisation de
la membrane du réticulum endoplasmique, que pour leur capacité à préserver i'activité
de la Glc-6-Pase lors de tests de dénaturation thermique. Les détergents étudiés sont de
trois classes soient :
1) Ioniques:
2) Non-ioniques:
3) Zwitterioniques:
Sodium Dodécyi Suhte (SDS), Sodium
Deoxycholate @OC)
Triton X-100, Triton N-10 1, Tween 20
CHAPS, CHAPSO, Zwittergent.
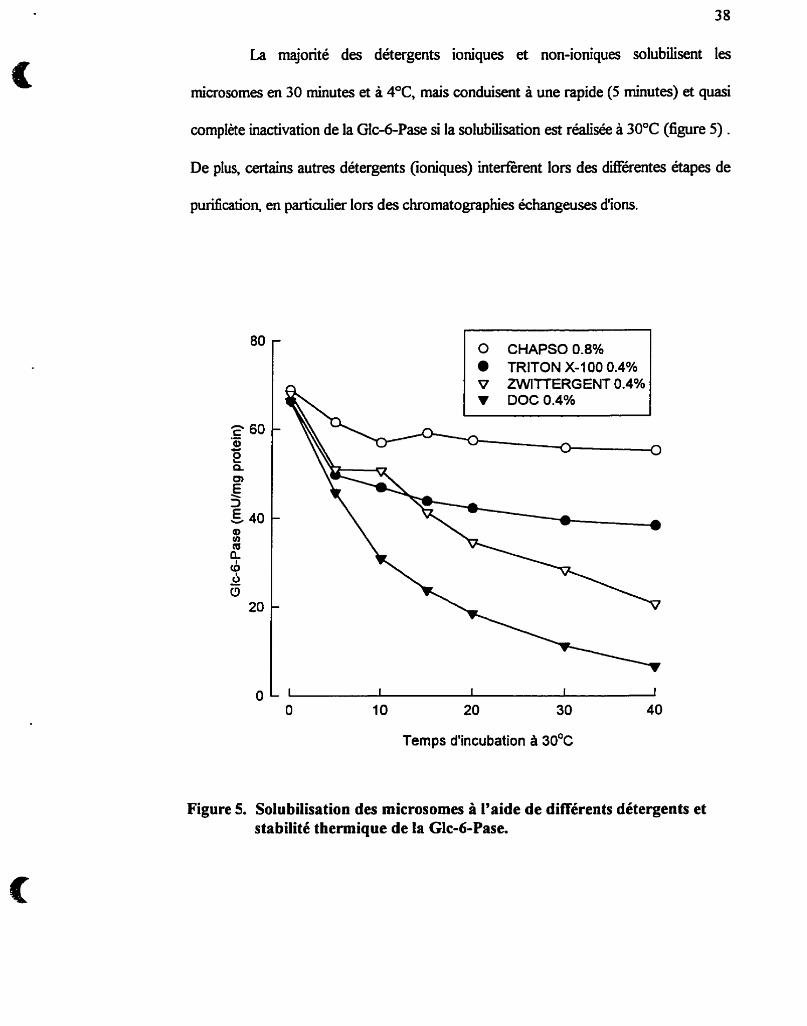
La majorité des détergents ioniques et non-ioniques solubilisent les
microsornes en 30 minutes et à 4OC, mais conduisent à une rapide (5 minutes) et quasi
complète inactivation de la Glc-6-Pase si la solubilisation est réalisée à 30°C (figure 5) .
De plus, certains autres détergents (ioniques) interferent lors des différentes étapes de
pur-cat io~ en particulier lors des chromatographies échangeuses d'ions.
O CHAPSO0.8% TRITON X-1 O0 0.4%
V ZWllTERGENT 0.4% V DOC 0.4%
Temps d'incubation A 30°C
Figure 5. Solubilisation des microsornes à l'aide de différents détergents et stabilité thermique de la Glc-6-Pase.

Les détergents zwitterioniques, quant à eux, possèdent des groupements
chargés positivement et négativement et n'interfêrent pas (ou très peu) avec les
chromatographies échangeuses d'ions. Ce sont ces détergents qui permettent de
conserver le mieux la structure native et l'activité catalytique de l'enzyme, tout en ayant
un pouvoir de solubilisation intégrale des protéines de la membrane du réticulum
endoplasrnique. La Glc-6-Pase n'est d'ailleurs inactivée que de 10 à 15% après 45
minutes d'incubation à 30°C par un traitement préaiable des microsomes avec du
CHAPSO (figure 5). C'est ce détergent que nous utiliserons tout au murs de notre
étude.
23.2- Effet de la force ionique sur la distribution de la GlcdPase lors de la
centrifugation des microsomes solubilisés
Le traitement des microsomes par le CHAPSO supplémenté de concentrations
croissantes de NaCl (O à 1M) ne diminue pas le pouvoir de solubilisation du détergent.
La Glc-6-Pase est inhibée par le NaCl (CoUa et al., 1974, Nordlie et aL, 1993), et la
force ionique semble, de plus, avou un effet sur la propriété qu'a la protéine Glcd-Pase
de se distribuer entre le d o t et le surnageant suite à une centritiigation des
microsomes solubilisés à haute vitesse (100,000 x g) (figure 6). En e f f i la proportion
de l'enzyme associée au culot d ï u e et ceUe dans le surnageant augmente avec une
force ionique croissante, jusqu'à un maximum de 0,6 M NaCI. La présence d'une
concentration minimale de NaCl est, par contre, critique pour i'étape de purification
suivante (chromatographie sur colonne d'hydroxylapatite). La présence d'une force
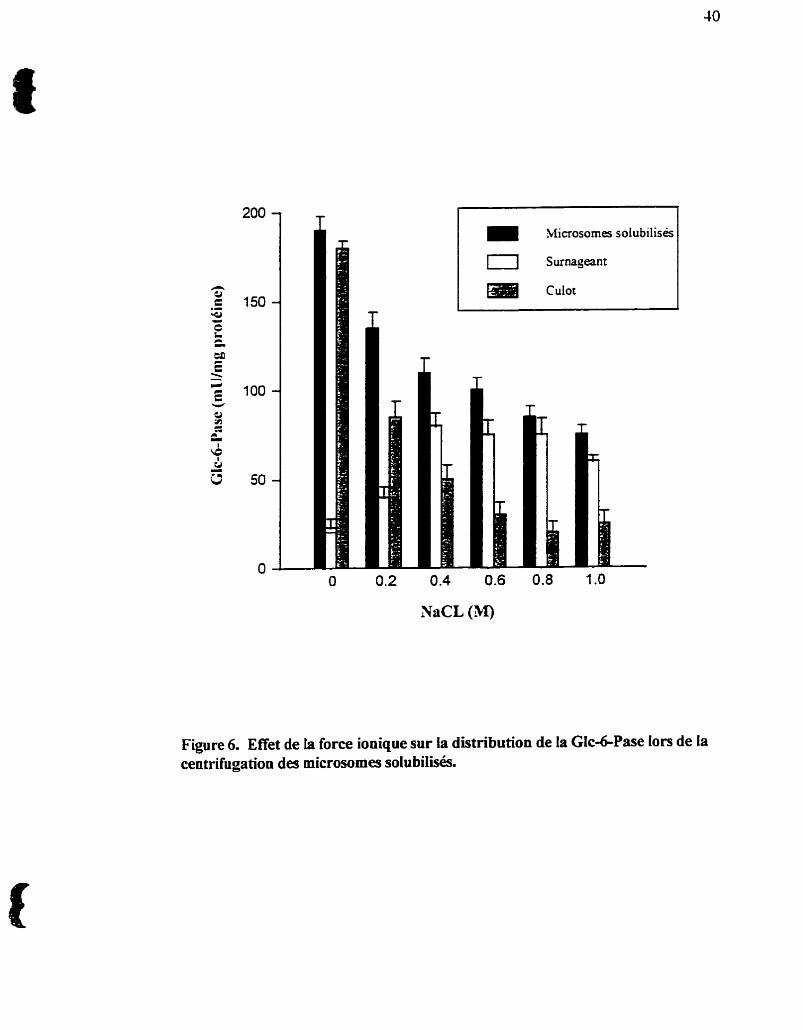
%ficrosomes solubilisés
Surnageant
Culot
Figure 6. Effet de la force ionique sur la distribution de la GIc&Pase lors de la centrifugation des microsornes solubilisés.

41
ionique élevée permet à l'enzyme d'éluer dans le volume mort de la colonne, ce qui
nt& pas le cas loque le sel est absent, la Gic-6-Pase restant alors fixée au gel (Rymsa
et de Groot, 1988, Speth et Schulze, 1992) et ne peut être élu& que par un gradient de
phosphate. Nous avons donc solubilisé les microsomes avec du CHAPSO 98% en
présence de NaCl 0,6 M (30 minutes a 4OC), puis centrifigé les microsomes solubilisés
à 100 000 x g pendant 45 minutes à 4OC, et gardé le surnageant contenant l'activité
Glc-6-Pase. Cette procédure nous a permis de minimiser toute manipulation mécanique
de renzyme Glc-6-Pase et de préserver un maximum d'activité.
2.3.3- Chromatographie sur colonne d'hydroxylapatite
L'hydroxylapatite (Ca2(P04)rhCa(OH)z (HA), la forme cristalline inorganique
du phosphate de calcium, est largement utilisée pour le fiactionnement et la purification
des protéines, enzymes, acides nucléiques, virus et autres macromolécules.
L'adsorption des protéines dûe aux interactions hydrophobes y est minime, et l'-té
avec laquelle elles se fixeront au gel sera donc, entre autre, fonction de la densité de
charge à la sufice de la protéine et du rapport chargdmasse.
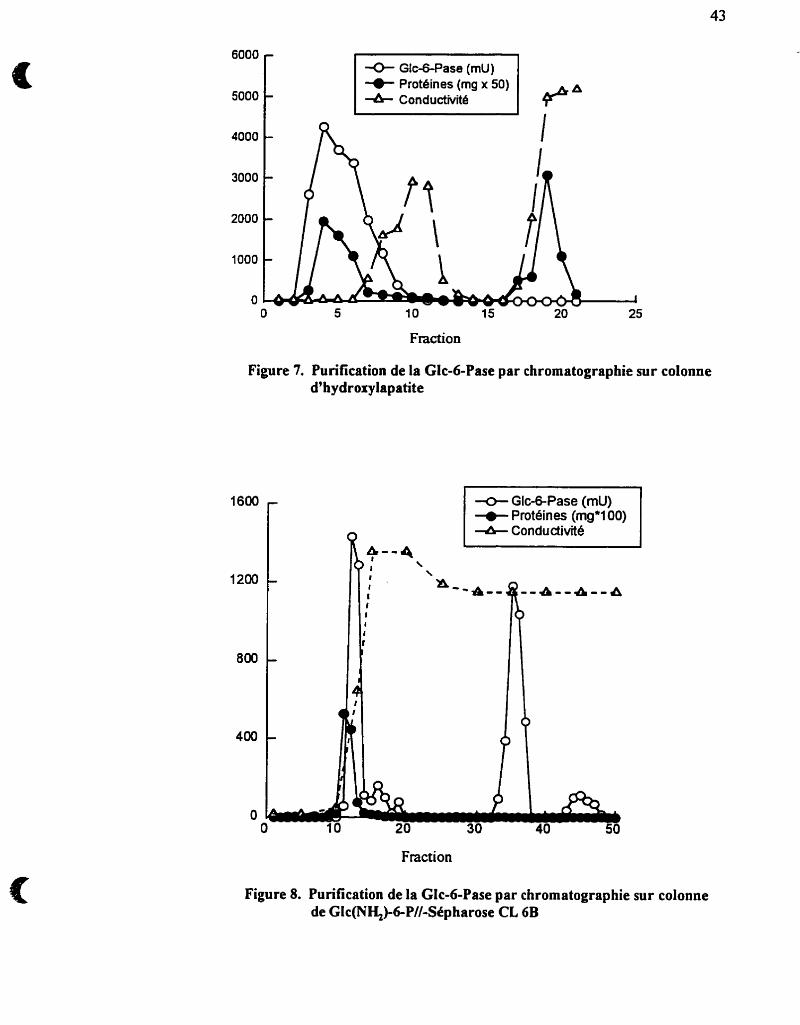
La chromatographie sur colonne d m est introduite après l'étape de
solubilisation et de centrifùgation des microsomes. L'application, puis l'élution de la
fiaction "surnageant" permet de recueiiiir une £faction jaunâtre dans le volume mort de
la colonne et à laquelle est associée pratiquement toute l'activité Glc-6-Pase (figure 7).
L'élimination d'environ 80 à 90% des protéines7 retenues par le gel, résulte en une
purification de 1 0 à 1 5 fois.

2.3.4- Chromatographie sur colonne d'aflinité Glc(NH+6-P-//-Sephamse CL6B
Une purification ultérieure a été obtenue en utilisant une chromatographie sur
colonne d ' f i t é . Nous avons donc couplé de la glucosarnine-6-phosphate (Glc(NH2)-
bP), un analogue structural du Glc-6-P, par I'intennédiaire de son groupement
fonctionnel amhé sur un gel de Sepharose CL- activé au bromure de cyanogène
(CNBr) (Sundberg et Porath, 1974).
La synthèse du gel d'afnnité se fait comme suit : Le CNBr réagit avec les
groupes hydroxylés de la Sepharose CL-6B et les aansfonne en groupes
irnidocarbonates, lesquels réagissent avec les groupements amines primaires du ligand
(dans notre cas la Glc(NH2)-6-P) pour former des liaisons isourée. Le protocole
consiste a peser environ 40 gr de Sepharose CL43 et de la resuspendre dans 40 ml
d'eau distillée, baisser la température du gel a IO-lS°C et porter le pH à 10.8 avec du
NaOH 4N, y ajouter graduellement, sous une hotte, 4 gr de CNBr en maintenant la
température et le pH constant. Une fois le CNBr complètement dissout et le pH
stabilisé, le gel est filtré à froid, et le filtrat est recueilli dans 20 gr de FeS04 pour
neutraliser le CNBr qui n'aurait pas réagi. Le gel est lavé avec 2L d'eau distillée et 2L
de NaHCO, 100 rnM à pH 10. Le gel est resuspendu dans 100 mi de NaHC03 a pH 10
contenant 3 gr de Glc(NHZ)-6-P, et est agité pendant 12 lus à 4°C. Le gel est ensuite
lavé avec 2L de NaHC03 100 mM à pH 10 et les groupements réactifs sont bloqués
pendant 12 hrs avec 100 ml de Glycine 1 M à pH 8.0 et à 4OC. Le gel est finaiement
extensivement lavé avec de i'eau distillée, resuspendu dans le tampon d'équilibration et
conservé à 4OC. L'efficacité de couplage a été évaluée à plus de 70% au moyen de [1-
' 4 ~ ] ~ l ~ 2 ) - 6 - ~ .
Fraction
Figure 7. Purification de la Glc-6-Pase par chromatographie sur colonne d'hydroxylapatite
* Glc-&Pase (mu) Protéines (mg'100)
& Conductivité
Fraction
Figure 8. Purification de la GIc-6-Pase par chromatographie sur colonne de Glc(NH&bP//-Sepharose CL 6B

La chromatographie se fat, par la suite, en équilibrant une colonne (0.8 x 90
cm) de Glc(NH&6-P-//-Sepharose CL43 avec le tampon d'équilibration contenant
0.6% C W S O à 4°C. La fiaction active éluée de la colonne d'HA contenant la Glc-6-
Pase est chargée à un débit de 10 mVhr. La colonne est ensuite lavée avec 5 volumes de
tampon d'équilibration, suivi de 3 volumes du même tampon supplementé de KC1500
mM, et de 3 volumes supplementé de KCI 400 mM. La Glc-6-Pase est enfui éluée
spécifiquement avec le dernier tampon contenant outre le KCl 400 XTM, du Gic-6-P
100 mM. Le débit est augmenté à 30 m L h et le volume des fiactions recueillies est de
5 mL. L'activité Glc-6-Pase éIuée est extrêmement instable.
Un point intéressant à noter est que nous détectons trois pics d'activité Glc-6-
Pase lors de cette chromatographie (figure 8). Le premier pic d'activité sort avec la
majorité des protéines lors du lavage de la colonne avec 500 mM KCI. Ce premier pic
représente systématiquement -50°! de l'activité Glc-6-Pase appliquée sur la colonne et
ne semble donc pas avoir une très forte affinité pour le Ligand (Glc(NHZ)-6-P). Le
second pic d'activité est celui qui sort le premier Ion de l'élution spécifique avec du
Glc-6-P 100 mM. Ce pic représente l'autre 50% de l'activité Glc-6-Pase. Un troisième
et dernier pic sort aussi lors de I'élution spécifique, et plus de 80% de cette activité est
attribuable à une activité phosphatasique non-spécifique (activité f3-
glycérophosphatase). L%ydrolyse du pglycérophosphate est négligeable pour les deux
premiers pics. Ce profil de pudication est reproductible et pourrait refléter l'existence
de deux états oligomériques de la Glc-6-Pase. Un tableau de purification (tableau 2)
ainsi qu'une éléctrophorese en condition dénaturantes (figure 9) résument notre
protocole de purification.
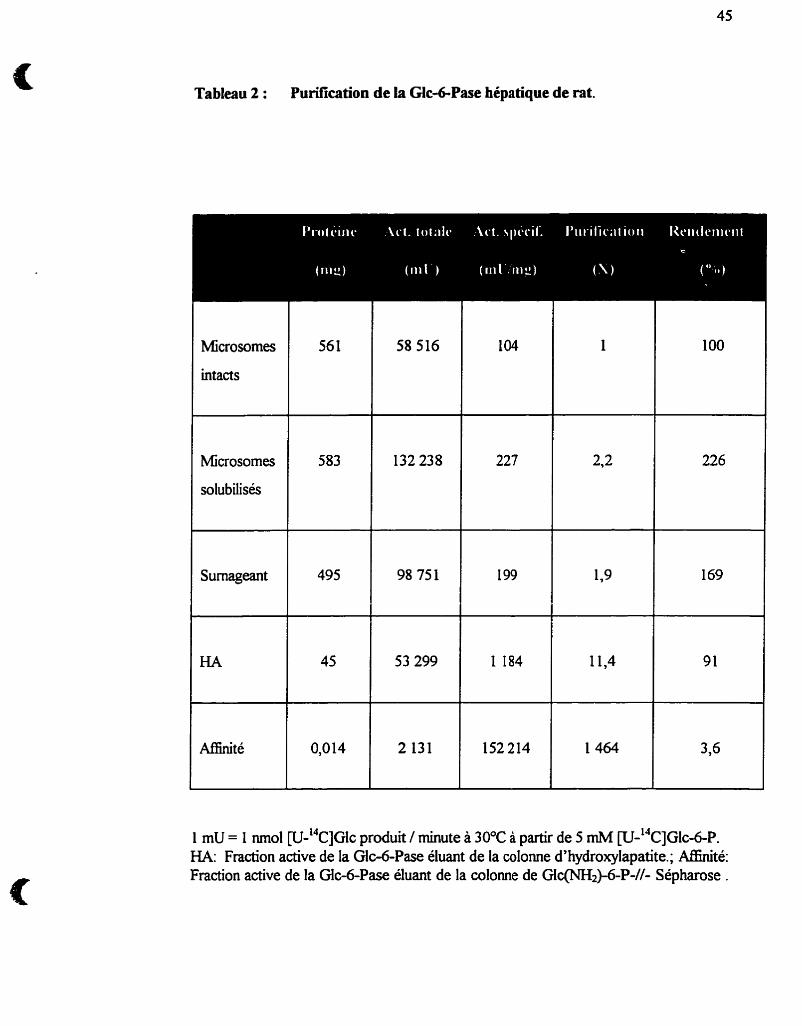
Tableau 2 : Purification de la Glc&Pase hépatique de rat.
Microsornes
intacts
1 mU = 1 mol W-'~C]G~C produit / minute à 30°C à partir de 5 rnM [U-~~C]GIC-~-P. HA: Fraction active de la Gic-6-Pase éluant de la colonne d'hydroxylapatite.; Affinité: Fraction active de la GIc-6-Pase éluant de la colonne de Glc(NH2)-6-P-//- Sépharose .
Microsornes
solubilisés
Surnageant
56 1 58 516
583
495
132 238
98 751
227
199
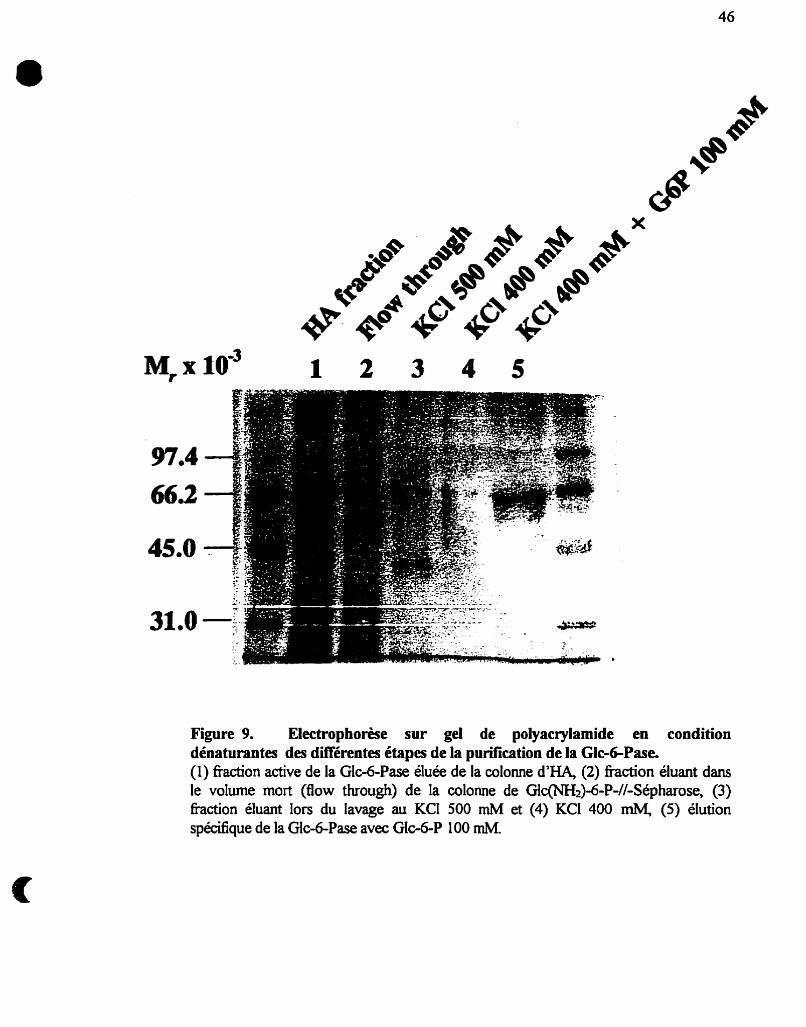
Figure 9. Electrophorèse sur gel de polyacrylamide en condition dénaturantes des différentes étapes de la purification de la Glc-6-Pase. (1) fiaction active de la Gle6Pase élu& de la colonne d'HA, (2) fiaction éluant dans le volume mort (flow through) de la colonne de Glc(NH~)-6-P-//-Sépharose, (3) £Faction éluant lors du lavage au KCI 500 mM et (4) KCl 400 mM, (5) élution spécifique de la Glc-6-Pase avec Glc-6-P 100 mU

47
2.4- Caractérisation de la glucosedphosphatase par la formation
d'un intermédiaire phosphorylé
Comme d k i t précédemment (voir section 1.3.2), la Gic-6-Pase se caractérise
par la formation, sur un résidu histidine, d'un intermédiaire phosphorylé transitoire Ion
du processus catalytique d'hydrolyse du Glc-6-P (Feldman et Butler, 1969) et peut être
utWsé pour identifier spécifiquement la Gic-6-Pase (Countaway et al, 1988, Mithieux
et aL, 1995).
2.4.1- Synthèse du [ n ~ ] ~ l c - 6 - ~ , [=P]M.~-&P
La synthèse du [ 3 2 ~ ] ~ l c - 6 - ~ de même que celle du [ 3 2 ~ ] ~ a n - 6 - ~ se fait tel que
démit par Berteloot et aL (1991a) mais avec quelques modifications. Le principe
consiste à phosphoryler du Glc ou du Man par I'hexokinase avec de l ' [ 3 2 ~ ] ~ ~ ~ - ~ g
(ICN Biomedicals, Montréal, Canada). Pour synthétiser du [ 3 2 ~ ] ~ c - 6 - ~ 2 m . il faut
que ltATP-Mg soit le réactif limitant. Le volume réactionnel est composé de 2 mM
ATP-Mg, 10 rnM Glc (ou 10 rnM Man), 125 mCi ["P]ATP, O. 1 mM EDTA, 5 mM
MgC12, et 50 mM HEPES 2 pH 7.1, et est incubé 4 hn a 30°C avec 10 mg
dbexokinase. La réaction est arrêtée en rajoutant 1% de charbon activé pour trapper
I'ATP qui n'aura pas réagi et que l'on éliminera en centrifùgeant 10 min à lq000g. Le
surnageant est par la suite chargé sur une colonne de DOWEX-Cl-. Le r2~]~ lc -6 -P est
par la suite séparé de I'[~~P]ATP contaminant en lavant la colonne avec un gradient de
NaCl dant de O à 200 mM. L'activité spécifique obtenue est de 300-400 cpdprnol.

2.4.2- Marquage radioactif et autoradiographie
L'incubation de l'enzyme avec du [ 3 2 ~ ] ~ l c - 6 - ~ nous permet donc de suivre et
de capturer l'intermédiaire phosphoryIé ( E ~ ~ P ) formé lors du processus catalytique de
l'enzyme. Le volume réactionnel contient 0.5 mM [ n ~ l ~ l c d - ~ , 0.05% b-
mercaptoethanol, 25 mM imidazole à pH 7.2. La réaction est lancée avec 5-30 mg de
proteme et incubée 10 sec à 30°C. La réaction est mêtée avec 2% SDS, 1W glycéroi,
60 mM Tris-HCI à pH 6.8. Ltéchantilion est ensuite soumis à une éléctrophorèse sur
gel de polyacrylamide 1W en conditions dénaturantes. Le gel séché sur du papier
WHATMAN (3 MM Ch) est enfin exposé contre un film KODAK O[-OMAT) dans
une cassette pour autoradiographie a -80°C pendant 2-3 jours.
2.5- Inactivation spécifique de l'activité phosphohydrolase du
système glucose-6-phosphatase
2.5.1- Modification de l'extrémité carboxyiée des résidus Aspartate et
Glutamate avec le dieydohexylcarbodiimide
Le dicyclohexylcarbodiimide (DCCD) est un carbodiimide hydrophobe
pouvant modifier irréversiblement les groupements carboxylés des protéines. Le
groupement réactif carboxyle forme dors un dérivé o-acylisourée instable pouvant 1)
par hydrolyse, regénérer le groupement carboxyle non modifié, 2) générer un dérivé O-
acylurée stable, ou 3) réagir avec un groupement aminé d'un résidu situé à proximité
dans la chaine polypeptidique (formant une liaison intramoléailaire) ou provenant d'un
agent nucléophile exogène, teUe la glycine ethyl ester (GEE) (voir figure 10).
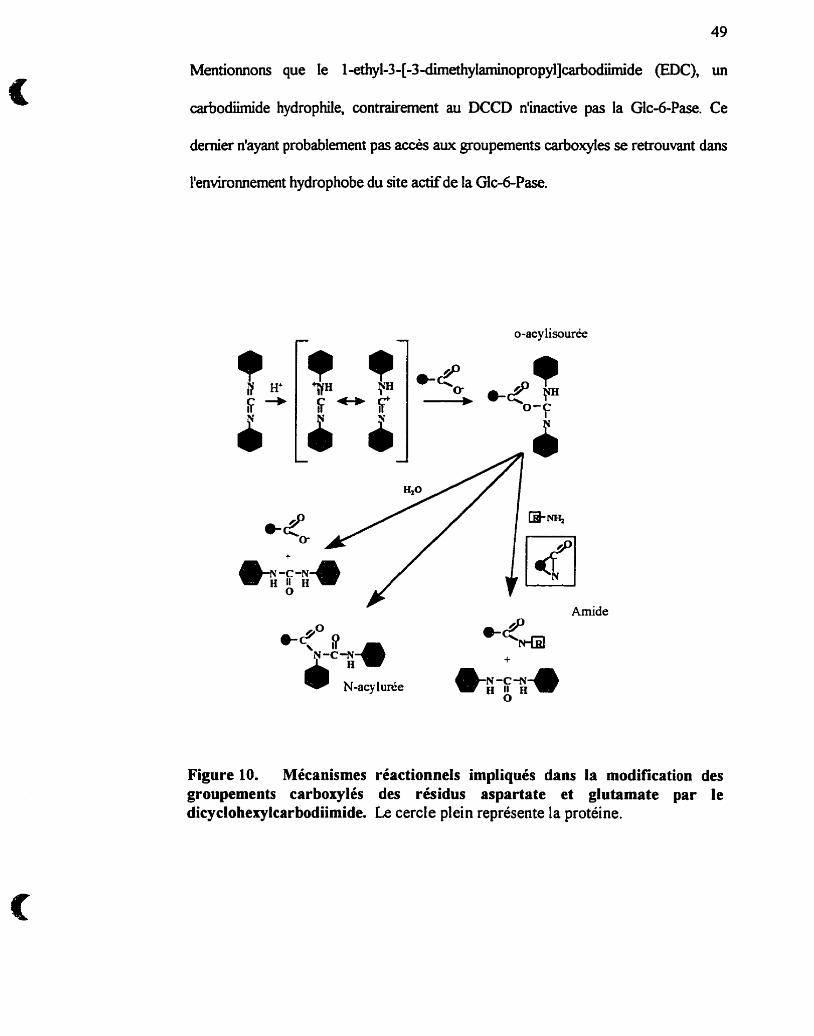
49
Mentionnons que le l-ethyl-3-[-3-dimethyIaminopropyl]Carf,odümide (EDC), un
carbodümide hydrophile, contrairement au DCCD n'inactive pas la Glc-6-Pase. Ce
dernier n'ayant probablement pas accès aux groupements carboxyles se retrouvant dans
l'environnement hydrophobe du site actif de la Gic-6-Pa.
Amide
+
N-acy lunie
Figure 10. Mécanismes réactionnels impliqués dans la modification des groupements carbosylés des résidus aspartate et glutamate par le dicyclohexylcarbodiimide. Le cercle plein représente la protéine.

2.5.2- Modification des résidus Lysine avec le pyridoral S'-phosphate
Le pyridoxal S'-phosphate (PLP) est un dérivé phosphorylé de la vitamine Bs et
sert de coenzyme dans les réactions de transamination et de décarboxylation des acides
aminés. Le coenzyme PLP se lie à son apoemyme par l'intermédiaire dime base de
SchE entre sun groupement Caldéhydique et un groupement &-aminé diin résidu de
lysine dans l'enzyme et d'une liaison ionique @ont salin) entre son groupement
phosphate et i'enzyme (voir figure 11). Cette propriété qu'a le PLP de modiier
spécifiquement les résidus lysine a donc été exploitée ainsi que la réduction
subséquente de la base de Sch8 par le borohydrure de sodium (NaBK), qui rend
covalente la liaison du PLP au résidu lysine modifié (voir figure 1 1).
2.6- Mesure de transport de glucose par la membrane microsomale
Pour mesurer les cinétiques d'accumulation de [u-'~c]G~c à partir de p-
"C]GIC-6-P, ou de [u-"c]M~ à partir de &J-14~]~an-6-p dans des vésicules
intactes, nous avons utilisé un appareil d'échantiiionage et de filtration rapide (FSRFA)
(Berteloot et al., 1991b). Les microsornes intacts sont incubés à 30°C dans une
chambre d'mcubation et des échantillons d'environ 50 mI sont récoltés
automatiquement sur des filtres de nitrocellulose. Ces filtres sont portés par un
collecteur de fraaions qui se déplace sous la chambre d'incubation permettant le
prélèvement des différents échantillons à des intervalles de temps aussi courts que 250
ms. Les fitres sont ensuite automatiquement lavés trois fois et la radioactivité est
mesurée à l'aide d'un compteur à scintillation liquide. Les analyses cinétiques ont été
réalisées à l'aide du programme de régression non-linéaire"Enzfitter"@1sevier-Bio&).
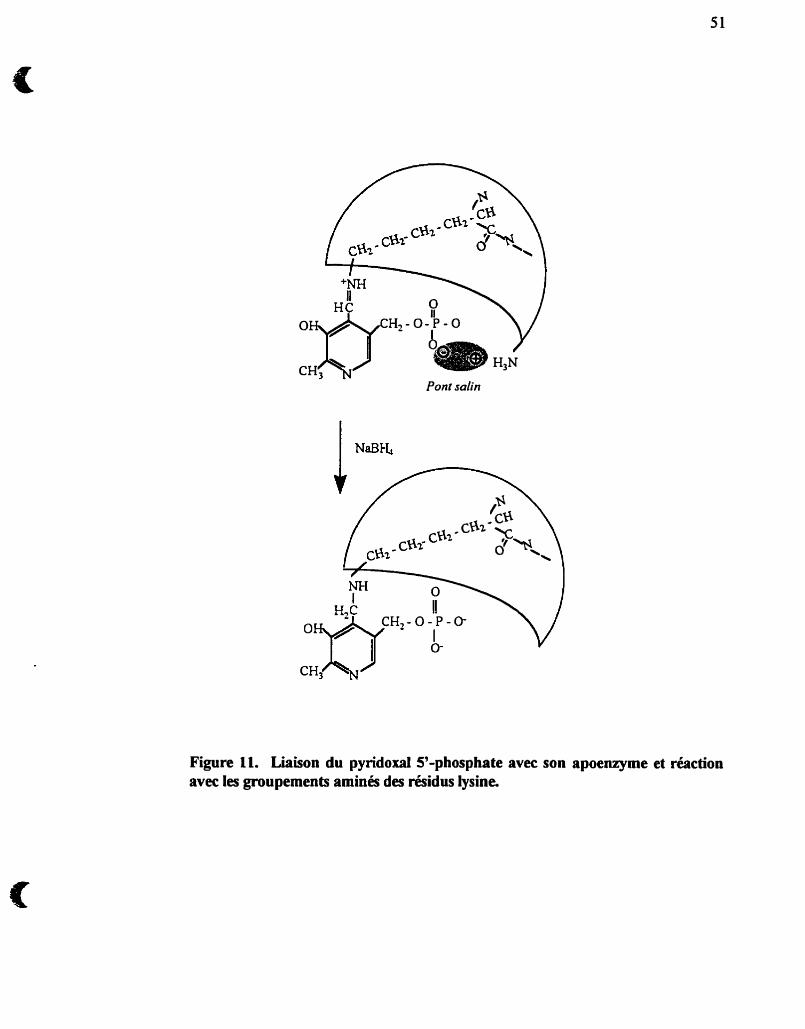
- - Pont salin
Figure 11. Liaison du pyridoxal S'-phosphate avec son apoenzyme et réaction avec les groupements aminés des résidus iysine.

2.7- Traitement des rats à la triarncinolone et à la streptozotocine
La streptozotocine est une drogue détruisant les cellules P sécrétrices d'insuline
dans le pancréas induisant ainsi un diabète insulinoprive. La streptozotocine est
préparée fiaî~hen~ent dans un tampon Na-citrate 4 mM à pH 4.5 ( H e m et Nordlie,
1972) et est administrée à une dose de 100 mg/Kg de poids corporel par une injection
intrapérïtonéaie à des rats à jeun. La sévérité du diabète est jugée par la mesure de la
glycémie à jeun (>300 mg/100 ml) et la perte de poids 8 jours suivant le traitement. Les
rats traités à la tnarncinolone, un glucocorticoide synthétique, om égaiement été mis à
jeun la veiiie, puis anesthésiés sous ether. Une légère incision sur le dos permet
l'iitrodudon sous-cutanée d'une pastille placebo ou d'une pastille de triarncinolone (50
mglpastille) (Innovative Research of America, Toledo, Ohio) avec un taux de
relachement de 50 mg/3 semaines. La dose quotidienne est donc, pour un rat de 200g
d'environ 2 mg/100 g de poids corporel (Arion et Nordlie, 1967, Traxinger et Nordlie,
1990). Les rats ont été traités pendant 6 jours et sacrifiés au septième.
2.8- Immunobuvardage
Nous avons obtenu du Dr Janice Chou des anticorps polyclonaux
générés chez le lapin et dirigés contre la sous-unité catalytique de la Glc-6-Pase (Shely
et al., 1993). L'éléctrophorèse des échantillons en conditions dénaturantes sur un gel de
polyacrylamide 1û% était suivie d'un t d e r t des protéines sur une membrane de
PVDF. Cette membrane est ensuite saturée avec 7.5% (w/v) d'albumine sérique de
boeuf pendant 2 hrs à température de la pièce, suivi de l'incubation en présence de

53
présence de l'anticorps primaire anti-Gic-6-Pase, puis de l'incubation en présence de
l'anticorps secondaire ad-IgG couplé à la peroxydase. Pour tester la spécificité de ces
anticorps, nous avons &ectué un immunobuvardage avec des microsornes isolés de
foie de souris, dom le gène codant pour la Glc-6-Pase de 36 kDa a été inactivé (figure
12), ainsi qu'avec des microsomes isolés de foie humain atteint de glycogénose de type
Ia (figure 13). Dans tous les cas de déficit de la Gic-6-Pase, aucune bande
immunoréactive n'a été observée à 36 kDa
]Rat Mouse
Figure 12. Immunobuvardage utilisant l'anticorps anti-Glc-6-Pase avec des microsornes isolés de foies de souris. (1) microsomes isolés de foie de rat, (2 - 5) microsomes isolés de foie de souris homozygotes (+/+) ou hétérozygote (+/-) pour l'allèle de la Glc&-Pase, agées de 1 ou 15 joun, (6) microsornes isolés de foie de souris dont le gène codant pour la Gic-6-Pase a été inactivé.
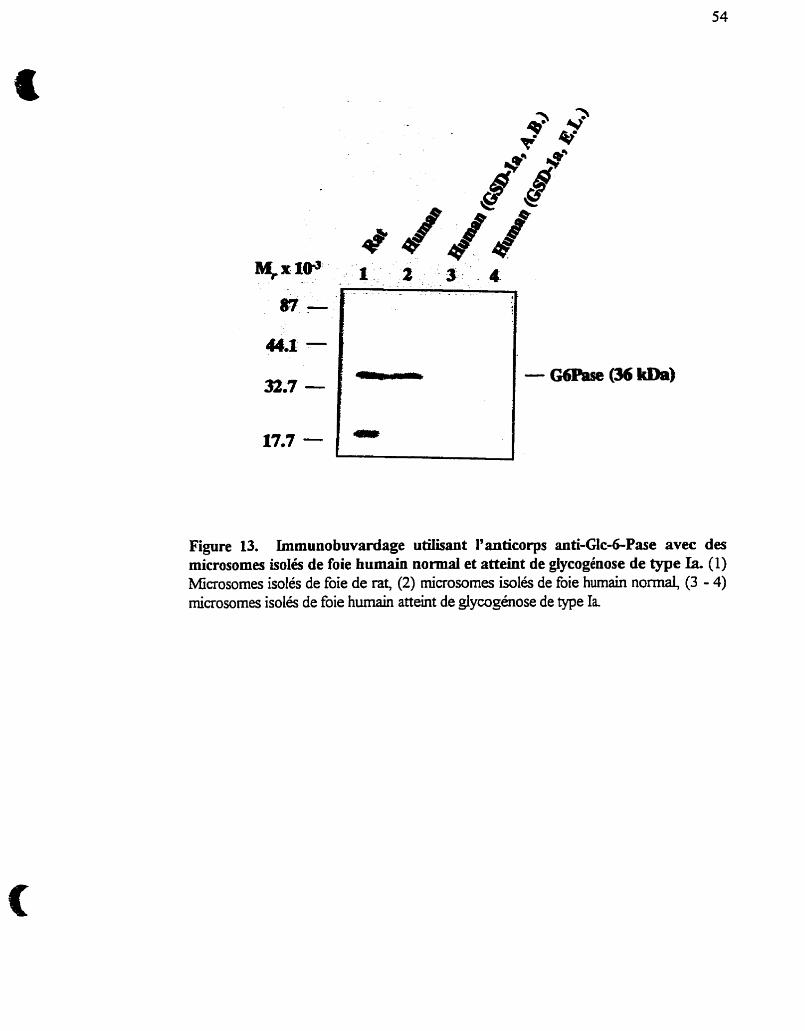
Figure 13. Irnrnunobwardage utiüsant l'anticorps anti-Gk-&Pase avec des microsomes isolés de foie humain normal et atteint de glycogénose de type Ia. (1) Microsornes isolés de foie de rat, (2) microsomes Wlés de foie humain normal, (3 - 4) microsornes isolés de foie humain atteint de glycogénose de type Ia

3.1- Spécificité du système giucos&phosphatase pour le Glcd-P :
Réévaluation du phénomène de latence.
3.1.1- Article I : Histone II-A stimulates gluucose-6-phosphatase and reveals
mannose-6-phosphatase activities without permeabiiization of
iiver microsomes. St-Denis J.-F., Annabi B., Khoury H. & van
de Werve G. (1995) Biochem. J 310,22 1-224

Article 1
Hypothèse : Les histones II-A modifient les caractéristiques cinétiques et la
spécificité de la Gica-Pase en ne peméabilisant pas les microsomes.
On a attniué à certaines protéines polycationiques teks la spermine, la
spermidine, la putrescine, ainsi que les histones (Nordlie et al., 1979, Blair et Burcheli,
1988) la propriété de perméabiliser ou de créer des pores dans les membranes
plasmiques ou dans les membranes des organelles intracellulaires (mitochondrie,
réticulum endoplasmique). Les histones II-A ont donc remplacé, dans certaines études,
l'usage classique des détergents pour solubiliser la membrane du réticulum
endoplasmique où les caractéristiques de la Glc-6-Pase sont identiques entre les deux
traitements. La perméabilité des vésicules n'a, cependant, pas été testée et la preuve
irrétùtable de l'intégrité membranaire des microsomes traités avec les histones II-A est
toujours manquante.
Ma contribution à cet article a été de démontrer que d'autres protéines
polycationiques (spermine, spermidiie, putrescine, . . .) avaient un effet stimulateur,
semblable à celui des histones II-& sur I'activité Glc-6-Pase sans perméabiiisation de la
membrane microsomale et que ce phénomène était réversible, par neutralisation de
charge, en présence de chélateurs (EGTA, EDTA). J'ai aussi préparé une hction
partiellement pufiée de la Glc-6-Pase par chromatographie sur colonne
d'hydroxylapatite et testé l'effet des histones II-A sur la Glc-6-Pase et la Man-6-Pase.

Biociiem. J. (1995) 310. 221-224 (Pnnted in Great Britain) 221
Histone Il-A stimulates glucose-6-phosphatase and reveals mannose-6- phosphatase activities without permeabilization of liver microsomes Jean-François Sr-DENIS, Bofiane ANNABI, Hanane KHOURY and Géraid VAN DE WERVE' Laboratoire d'Endocrinologie Métabalique, Departrnents of Nutrition and Biochemistry. Université de Montréal. Mon:réal. Québec H3C 3J7. Canada
b
The effect of histone II-A on glucose-6-phosphatase and mannose-6-phosphatase activities was investigated in relation to microsornai membrane permeability. It was found that glucose- &phosphatase activity in histone II-A-pretreated liver micro- s o m a was stimulated to the same extent as in detergent- permeabilized microsomes. and that the substrate specificity of the enzyme for glucose 6-phosphate was lost in histone II-A- pretreated microsomes, as [U-14C)gIucose 6-phosphate hydrolysis was inhibited by mannose 6-phosphate and [U-"CJrnannose 6-phosphate hydrolysis was increased. ï h e accumulation of [U-14C]gIucose from [U-'4qglucose 6-phosphate into untreated
microsomes was completely abolished in detergent-treated ves- ides. but was increased in histone II-A-treated microsomes. accounting for the increased glucose-6-phosphatase activity. and demonstrating that the microsomal membrane was stili intact. The stimulation of glucose-6-phosphatase and mannose-6-phos- phatase activities by histone II-A was found to be reverssd by EGTA. I t is concluded that the effects of histone II-A on glucose- 6-phosphatase and mannose-6-phosphatase are not caused by the permeabilization of the microsomal membrane. The measure- ment of mannose-6-phosphatase Iatency to evaluate the intact- ness of the vesicles is therefore inappropriate.
INTRODUCTION from NEN-DuPont (Mississauga. Canada). ,411 reagents were of analytical grade.
Glucose-6-phosphatase (GoPase) (EC 3. 1 .3.9) catalyses the final step of hepatic glucosc production and is located at the endo- Preparation of llver microsomes plasmic-reticulum membrane [ I l . In intact microsomes G6Pase hydrolyses only glucose 6-phosphate (G6P). whereas in detergent- Liver microsomes were isolated from overnight-fasted male
treated rnicrosomes an increased rate of G6P hydrolysis as well Wistar rats (Charles River. St-Constant. Canada) (200-250 g) as
as the hydrolysis of a variety of phosphorylated sugars are described previously (51. and were resuspended in 0.75 M su-
observed [Il. crose/50 mM Tris. pH 7.3. at a protein concentration of approx.
The measurement of mannose-6-phospha tase activi ty 15 rng/ml. Microsomal vesicles wcre either used the same day or
(MbPase) is widely uscd for the determination of microsomal kept at -80 OC for later use. Freezing of microsomcs. if thawed
membrane intactness [2]. This activity is latent in untreated once at 4 O C . did not affect the kinetic propcrties of G6Pase [6].
microsornes and is revealed when rnicrosomes are treated with a as also documented by others [7]. Microsomal protein con- centration was measured by the method of Bradford. tvith BS.4 detergent. Histone Il-A has k e n reported to stimulate GoPase
standard [3.4] rinu M6Püscr activities [JI. and was therefore proposed as an alternative agent to detcrgent for the pcrmeabilization of the microsornes [JI. In keeping with these findings. we found that Partial purification of G6Pase G6Pase and M6Pase activities are stimulated in histone II-A- A partially purified G6Pase from rat liver microsomcs \vas treated microsomes to the same extent as in detergent-treated prepared by a procedure f9] rnodified from that of Rymsa and Dc. microsomes. but. unlike the effect ofdetergents, histone II-A was Groot [[O]. Briefly, microsomes were solubiiized with the non- not found to permeabilize the membrane barrier. denatunng zwitterionic detergent CHAPSO (0.8 O , . w/v) in thc
presence of 0.6 M NaCl and centrifuged at IO5 000 g for 30 min. The resulting supernatant was applied on a hydroxyapatite (HA)
' WPERIMENTAL column and G6Pase activity was collected in the void volume. The specific activity (with 0.2 rnM [U-''qG6P as substrate)
Materials of the pooled fractions (HA fraction) ranged from 33 to
6 3-[(3-CholamidopropyI)dimethyIammonio]-2-hydroxy- 1 -pro- 213 nmol/min per mg of prorein for diferent preparations.
panesulphonate (CHAPSO). EGTA. G6P (monosodium salt). histone II-A and mannose 6-phosphate (M6P) (monosodium Enzyme assays
sait) were al1 purchased from Sigma Chemical Co. (St. Louis. G6Pase activity was rneasured at 30 OC in untreated microsomes MO). Tris base was from Boehringer Mannheim Corp. (Laval. and in the HA fraction by the production of [U-"C]glucose from Canada). Sucrose (ultra-pure) and [U-'4C]G6P were from ICN 0.2 mM [U-14C]G6P (0.4 nCi/nmol), as described previously [SI. Biomedicals (Montréal. Canada). [U-lJC]Mannose was obtained M6Pase activity was measured likewise by using 0.2 mM [U-
Abbreviations used: CHAPSO. 3-~(3-cholamrdopropyI)dimethylammonio)-2-hydroxy-l-propanesulphonate: FSRFA. fast-sampling rapid-filtration apparatus; G6P. glucose 6-phosphate; G6Pase. glucose-6-phosphatase: M6P. mannose 6-phosphate; M6Pase. mannose-&phosphatase. HA. hydroxyapatite.
'TO whom correspondence should be addressed. at: Department of Nutrltion. université de Montréal. C. P. 6128. Succursale centre-ville. Montréal. Ouebec H3C 37. Canada.

222 J.-F. St-Denis and others
14C]M6P (0.2 nCiJnmol). G6Pase and M6Pase activities were also measured in microsomes previously treated with CHAPSO (0.8 for 30 min at 4 OC (0.08 ?b CHAPSO in the assay medium) as described previously [6] and in microsomes previously mixed with 10 mg/ml histone II-A at 4 OC (1 rng/ml in the assay medium).
[U-"C]MGP synthesis
[U-laC)M6P was prepared by the phosphorylation of w- laC]mannose (300 mCi/mmol) with ATP by using hexokinase as previously described [Il], with the following modifications. The reaction was stopped by the addition of 1 O. activated charcoal, followed by centrifugation (1 min, 16000 g in an Eppendorf centrifuge). The supernatant was then applied on a Dowex AG 1x8 column (acetate form). [UJ4CJMannose that had not reacted came out in the Elow-through, whereas [U-'"CIM6P was eluted with an acetic acid gradient (0.2-1 M). [U-"CJM6P was then pooled, freezedried (with a Savant SpeedVac concen- trator) and resuspended in 4 m M unlabelled M6P (in 50 mM Tris. pH 7.3) to a final specific radioactivity of 2.4 nCi/nmol. The yieid of [U-'T]M6P synthesis was 99 O,, .
Uptake measurements [U-'T]Glucose uptake in microsomes from 0.2 mM [U-IJC]G6P (20 nCi/nmol) was deterrnined by using a fast-sampling rapid- filtration apparatus (FSRFA) as describd and vaIidated else- where [6.lt. 131. Uptake measurements were also performed in microsomes previously treated with IO mg/ml histone II-A for 2 min rit 4 OC (0.3 mg/ml in the uptake medium) and in vesicles previously treated with CHAPSO (0.8 O,) for 30 min at 4 O C
(0.03",, CHAPSO in the uptake medium).
RESULTS
TübIe 1 shows that G6Pase activity in liver microsomes is stimulated by about 2-fold and M6Pase by about 6-fdd by either histone II-A or CHAPSO. The increased rates of [U-LJC]M6P and [CI-laC]GfiP hydrolysis in histone II-.4-treated microsornes were not due to the stimulation of non-specific phosphatases. as we verified that the hydrolysis of fl-glycerophosphate (20 mM)
Table 1 Stimulation by histone Il-A of G6Pase and M6Pase acüviües from rat Iiver microsomes
[u-"CjGlucose production from 0.2 mM [u-"C]G~P was measured in liver microsomes and !n a partiatly purilied G6Pase preparalion (HA fraction) under dilferent conditions. [U-"CIMannose produclion from 0.2 mM [u-"C]M~P was rnasured in liver microsomes. Oata are the means +S.EM. for R experimrnts: ' P c 0.05 compared with the respeclive untreated values by an unpaired Sludent's I test. and " P c 0.01 and "'P < 0 0 5 cornpared with the untreated value by a paired Sludent's t test
Liver microsomes HA traction
G6Pase M6Pase G6Pase M6Pas (nfnOl/rnin per mg) (nmol/min per mg) fi) ($1 (n = 5) (n = 4) (0 = 9) (n = 51
C Untreated 12.8 + 1 2 3.3 + 0.6 100 100 Histone Il-A- 22.1 23.7' 1 7 2 I 3 1 ' 123+5" 119-Ç_5"'
lrealed (1 mgml)
CHAPSO-treated 23 5 1 1 4' 21.9 I 2 . 2 ' - - (O 08%)
Figure 1 Inhibiüon by M6P of ihe production of [U-14C]glucose from 0.2 m l (U-14C]G6P
[u-"C]G6P i0.2 mM) hydrolysis was measured in Ihe presence of the indimted concentrations of unlabelled M6P in untreated microsomes (O). histone Il-A-treated microsornes (1 mg/rnl in the assay medium) (a). CHAPSO-treated rnicrosomes (a). and in microsomes previously treated with both CHAPSO and histone Il-A (A). The results are expressed as the percentage of the enzymic activities measured In the absence of M6P (100% represents 9.6. 24 9. 31 3 and 31 6 nmol/mtn per mg for unlreated. CHAPSO-. histone Il-A- and CHAPSO+ histcne Il-A-treated microsornes respectively).
was unchanged. The eKect of other polycationic proteins (1 m M spermine. spermidine and putrescine) wos tested on G6Pase activity, and we observed results (not shown) similar to those reported by Nordlie et al. [14]. We have also tested the effect of histone II-A on G6Pase and M6Pase activities in a partially purified preparation (HA fraction). Since the purification factor difkred from one p repa~ t ion to another. the specific activities of G6Pase and M6Pase in the HA fraction were expressed as percentages of the control value for each experiment. A slight but signifiant stimulation of GoPase and M6Pase activities was observed when the partially purificd enzyme was treated with histone II-A (Table 1). The results shown in Figures 1-3 are iIIustrative of a series of
rxperimrnrs on aifirent microsomai preparations which gave consistentIy the same type of rcsults. The spccificity of G6Pase for [U-'TJG6P was further studied by competition with un- labelled M6P. Figure 1 shows that M6P had M e effect on G6Pase activity in untreated microsomes, but almost complctely inhibited G6Pase activity either in detergent-treated microsomes. as reported previously [15,16]. or in microsomes treated with histone II-A or with CHAPSO and histone II-A together. The K, values of M6P towards [U-Y]G6P hydrolysis are 0.85.0.89 and 0.90 m M in CHAPSO-, histone II-A- and CHAPSO + histone II-
I A-treated rnicrosomes rcspectively.
The stimulation of G6Pase activity by histone II-A herein reponed was observed previously by other investigators, who suggested that this effect was due to the disruption of the '
microsomal membrane [4]. To verify this possibility, we compared the effect of the detergent CHAPSO and histone II-A on [U- L4C]G6P uptake and hydrolysis (Figure 2). As expected, the accumulation of [U-14C]glucose was precluded by the treatrnent of vesicles with detergent, but, in contrast, was increased by histone II-A pretreatment, thus indicating that histones do not permeabilize microsomes in these experimental conditions. The same plateau of [U-laC]glucose accumulation was observed whether histone II-A was added together with [U-14C]G6P o r 2 min after preincubation of microsomes with the Iatter. The higher accumulation of [U-14Cjglucose in histone II-A-treated microsornes compared with control microsomes is proportional
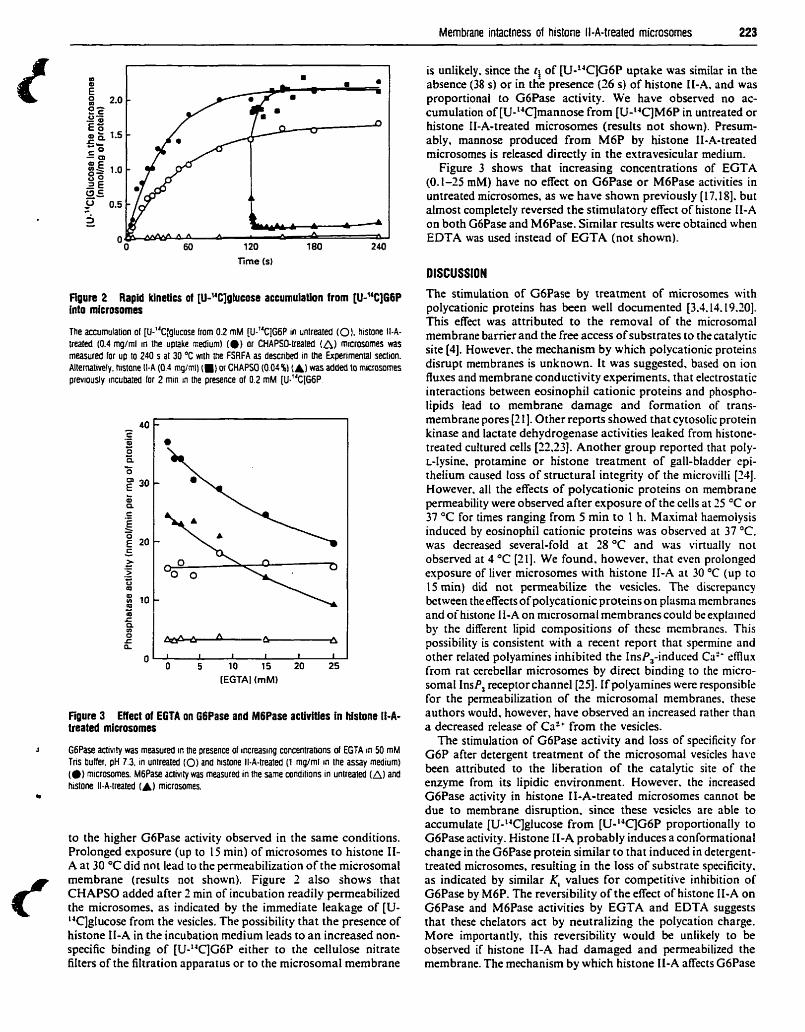
Membrane intactness of histone Il-A-treated microsornes 223
is unlikely. since the ri of [U-TIG6P uptake was similar in the absence (38 s) or in the presence (26 s) of histone II-A. and was proportional to G6Pase activity. We have obsenred no ac- cumulation of [UJ4C]mannose from [U-14C]M6P in untreated or histone II-A-treated microsomes (results not shown). Presum- ably, mannose produced from M6P by histone II-A-treated microsomes is released directly in the extravesicular medium.
Figure 3 shows that increasing concentrations of EGTA (0.1-25 mM) have no effect on G6Pase or M6Pase activities in untreated microsomes. as we have shown previously [17.18]. but alrnost completely reversed the stimulatory effect of histone II-A on both G6Pase and M6Pase. Sirnilar results were obtained when EDTA was used instead of EGTA (not shown).
DISCUSSION
Rgure 2 Rapid kinetIcs of [U-14C]glucase accumulation from [U-14C]G6P into microsomes
The accumulation of [u-"CIglucose from 0.2 mM [u-"C]GW in untreated (0). histone 114- trealed (0.4 mg/ml in the uptake medium1 (0) or CHAPSO-treated (A) microsomes was measured for up IO 240 s at 30 O C wilh the FSRFA as d ~ ~ n b e d in the Expenrnental section. Alternativeiy. histone Il-A (0.4 mgtrnl) (m) or CHAPSO (O 04%) (A) was added to microsomes previously incubated for 2 min in the presence of 0.2 mM [u."C]G~P
Figure 3 Eflect of EGTA on G6Pase and M6Pase activities in histone If-A- treated microsomes
a G6Pase acttvity was measured in the presence of increasing crincrotratrons of EGTA in 50 mM Tns buffer. pH 7 3. in untreated (0) and hisione Il-A-treated (1 mglm1 in the assay medium) (0) micrasomes. M6Pase actniify was measured in lhe same conditions in unlreated (A) and histone Il-A-lreated (A) rnicrosomes.
to the higher G6Pase activity observed in the same conditions. Prolonged exposure (up to 15 min) of microsomes to histone II- A at 30 OC did not lead to the permeabilization of the microsomal membrane (results not shown). Figure 2 also shows that CHAPSO added after 2 min of incubation readily permeabilized the microsomes, as indicated by the immediate leakage of [U- 'aC]glucose from the vesicles. The possibility that the presence of histone II-A in the incubation medium leads to an increased non- specific binding of [U-laC]G6P either to the cellulose nitrate filtes of the filtration apparatus or to the rnicrosomal membrane
The stimulation of G6Pase by treatment of microsomes with polycationic proteins has k e n well documented [3.4.14.19.20]. This effect was attributed to the removal of the microsomal membrane barrier and the free access of substrates to the catalytic site (41. However. the mechanism by which polycationic proteins disrupt membranes is unknown. lt was suggested, based on ion fluxes and membrane conductivity experiments. that electrostatic interactions between eosinophil cationic proteins and phospho- lipids lead to membrane damage and formation of trans- membrane pores [2 11. Other reports showed that cytosolic protein kinase and lactate dehydrogenase activities leaked from histone- treated cultured cells [22.23]. Another group reported that poly- r-tysine. protamine or histone treatment of gall-bladder epi- thelium caused loss of structural integrity of the microvilli [24]. However. al1 the effects of polycationic proteins on membrane permeability were observed after exposure of the cells at 25 OC or 37 "C for times ranging from 5 min to 1 h. Maximal haemoiysis induced by eosinophil cationic proteins was obsened at 37 OC. was decreased several-fold at 28 OC and was virtually not observed at 4 O C [21]. We found. however. that even prolonged exposure of liver microsomes with histone II-A at 30 O C (up to 15 min) did not permeabilize the vesicles. The discrepancy between theeffects of polycationic proteins on plasma membranes and of histone II-A on rnicrosomal membranes could be explained by the diîTcrent lipid compositions of thcsc mcmbrancs. This possibility is consistent with a recent report that spermine and other related polyamines inhibited the InsP,-induced Ca2- smux from rat cerebellar microsomes by direct binding to the micro- soma1 InsP, receptor channel[25]. If polyamines were responsible for the permeabilization of the microsornal membranes. these authors would, however, have observed an increased rather than a decreased release of Ca2+ from the vesicles.
The stimulation of GoPase activity and loss of specificity for G6P after detergent treatment of the microsornal vesicles havc k e n attributed to the liberation of the catalytic site of the enzyme from its lipidic environment. However, the increased G6Pase activity in histone II-A-treated microsomes cannot be due to membrane disruption. since these vesicles are able to accumulate [U-NC]glucose from [U-laC]G6P proportionally to GoPase activity. Histone II-A probably induces a conformational change in the G6Pase protein sirnilar to that induced in detergent- treated microsomes, resuiting in the loss of substrate specificity. as indicated by similar K, values for competitive inhibition of G6Pase by M6P. The reversibility of the effect of histone II-A on GoPase and M6Pase activities by EGTA and EDTA suggests that these chelators act by neutralizing the polycation charge. More importantly, this reversibility would be unlikely to be observed if histone II-A had damaged and penneabilized the membrane. The mechanism by which histone II-A affects G6Pase

224 J.-F. St-Denis and others
and M6Pase is unknown and beyond the scope of this study. Among the possible explanations are an interaction of histone II- A with membrane phospholipids, affecting the enzyme. or a direct interaction with the protein itself, as a slight stimulation is still retained in purified preparations.
In the present paper, we report that (1) G6Pase and M6Pase activities are both increased upon treatrnent of the microsomes with histone II-A without permeabilization of the vesicles, (2 ) the vesicles (treated or not with histone II-A) never accumulate mannose, and (3) therefore, mannose produced from M6P by histone II-A-treated microsomes is presumably released directly into the extramicrosomal medium. Taken together. these three observations indicate that the topology of G6Pase would not be that proposed by the substrate-transport model.
In summary, histone II-A cannot be used as a permeabilizing agent of microsomes, since histones do not affect the property of these vesicles to accumulate products. Also. the measurement of M6Pase latency cannot be used to evatuate the microsornal membrane intactness. since M6Pase can be revealed to the sarne extent by detergent and histone II-A. but by the latter without permeabilization of the vesicles. It is likely that M6Pase activity in untreated microsomes is the reffection of a small amount of G6Pase in a conformation capable of M6P hydrolysis.
This work was supported by Grants ME-10783 and MT40804 from the Medical Research Council of Canada.
REFERENCES 1 Nordlie, R. C. and Sukalski. K. A (1985) in The Enzymes of Bioiogical Membranes.
2nd eân. (Manonosr. A N.. ed.). MI. 2. p. 349-398. Plenum Publishing Corp.. New York
-
Anon. W. J. (1989) Methais Enzymol. 174. 5867 Benedetto. J. P. and Got R. (1980) Biochim. Biophys. Acta 614. 400-206 Blair. J. N. R. and Burchell. B. (1988) Biochirn. Biophys. Acta 964. t61-167 m de Wem. G. (1989) J. Biol. Chem. 261. 60336036 St-Oenis. J . 6 Comte. B.. Nguyen. D. K. Seidman. E. Paradis. K.. levy. E and an de Werve. G. (1994) J. Clin. Endocrinol. Metab. 75. 955-959 Lucius. R. W.. Waddell. 1. O.. Burcfidl. A. and Nordlie. R. C. (1993) Biachem. J 290. 90741 1 Bradford. M. M. (1976) Anal. Biochem. 72 248-254 AnnaBi. 0. and van de Werve. G. (1 993) Diabetes 42. 125A Ryrnsa. B. and De Grwt. H. :1988) Biol. Chem. Hoppe-Seyler 369. 175-121 hnyth. R. J.. Battletr. K. Burcheti. A.. San. H. M. and Eyre. J. A. (1993) Biûhern. J. 294. 145151 Berteioot, A. Vidal. H. and Mn de Werve. G. (1991) J. Biol. Chem. 266. 5497-5507 RomaneIli. A St-Denis. J.-F.. Vidal. H.. Tchu. S. and van de Weme. G. (1994) Biochem. Biophys. Res. Commun. 200. 1491-1497 Nordlie. R. C.. Johnson. W. 1.. Corwfzer. W E and Tweueit. G. W. (1979) Biçcfilrn Biophys. Acta 585. 12-23 Anon. W. J.. Wallin. B. K.. Carlson. P. W. and Lange. A J. (1972) J. Bioi. Chem. 247. 2558-2565 Vidal. H.. Beneloot A. Lame. M.-J.. St-Denls. J.-F. and M n de Werve. G. (19921 FEBS Len 302. 197-200 van de Werve. G. and Vidal. H. 11990) Biacnem. J. 270. 837438 Vidal. H.. St-Denis. J.-F.. Painchaud. E. Lame. M.-J. and van de Wene. G. (1991) Anal. Biochem. 193. 135-141 Vergnes. O.. Manet. M. B. and Got. R. (1981) Int. J. Biochem. 13. i26Si268 Xiao. 0.. Jaspers. 1.. Manhew. E and Lea. M. A il9931 Moi. Celi. Biochem 122. 17-24 Young. J. O.&. Peterson. C. G. B . Venge. P. and Cohn. 2. A. (1986) Naiure (London1 321. 613616 Rahmdorf. H. J.. Maicnow. D. and Gerish. G. (1979) Ce11 Biol. Int. Rep. 3. 237-245 Kubler. D . Pyerin. W. and Kinzel. V. (19821 Eur. J. Cell Biol 26. 306-309 Quinton. P M. and Philpoli. C. W (1973) J. Cell Biol. 56. 787-706 Sayen. L G. and Michelangeli. F. (19%) Biochem. Soc. Trans. 22. 152s
Rece:ved 9 January 199924 March 1995; accepled 24 April 1995

3.2- Transport de glucose associé au système glucose-6-
phosphatase
3 .S. 1 - Article 2 : Glucose transport and glucose 6-phosphate hydrolysis in intact
rat liver microsomes. St-Denis J.-F., Berteloot A, %da1 H-,
Annabi B. & van de Werve G. (1995) J. Biol. c h . 270,
21092-21097

Article 2
Hypothèse: Le transport de glucose et i'activité de la Gk-6-Pase dans les
microsomes de foie sont Liés.
Malgré le clonage récent d b transporteur microsoma1 de glucose (GLUT 7)
par WaddeU et al (1992), aucune capture directe de glucose n'a pu être observée dans
des microsomes de foie dans une variété de conditions expérimentales (Romanelii et
al., 1994). Seule l'hydrolyse du [u-'~c]G~c-~-P par la Glc-6-Pase semble permettre
llaccumulation rapide de w-'~c]G~c dans les microsomes de foie de rat. Les
caractéristiques cinétiques du transport de glucose ainsi que sa relation avec l'hydrolyse
du Glc-6-P ont donc été étudiées.
Ma contribution à cet article a été de démontrer que l'accumulation de @J-
14 C]Glc dans les vésicules est directement proportionnelle à l'activité Glc-6-Pase en
incubant des microsomes en présence de 0.2 m . W-~~C]GIC-~-P et de différentes
doses d'orthovanadate, un inhibiteur de l'enzyme.

THe JOL~YU OP BIOU)<;ICU. CHUIISIRY O 1995 by The Amenam Saaety for Biochemisuy and Molecuiar Biology. Inc.
VOL 270. ho. 36. issue of Septemkr 8. pp. 21092-21097.1995 P n n d 1n U S A
Glucose Transport and Glucose 6-Phosphate Hydrolysis in Intact ( Rat Liver Microsomes*
[Received for publication. Februsry 6. 1995, and în revised form. July 12, 1995)
Jean-François St-Denis*, Alfred BertelootM, H u b e r t Vidalal, Borhane AnnabiS, and GéraId v a n de Werve$** From the $La boni taire d'Endocrinologie Métabolique, Departments of Nutrition and Biochemistry, the §Groupe de Recherche en Transport Mernbranaire, and the Wepartment af Physiology, Uriiversité de Montréal, Montréal, Québec H3C 3J7, Canada
Glucose transport was investigated in r a t liver micro- somes i n relation to glucose 6-phosphatase (Glu-ô-Pase) activity using a fast çampiing, rapid filtration appara- tus. 1) The rapid phase in t racer uptake a n d t h e burs t phase in glucose 6-phosphate (Glu-6-P) hydrolysis ap- p e a r synchronous, while the slow phase of glucose accu- mulation occurs during the steady-state phase of glu- cose production. 2) [lJC]Glucose efflux from preloaded microsornes can be obsemed upon addition of e i the r cold Glu-6-P o r Glu-6-Pase inhibitors, b u t not cold glu- cose. 3) Similar steady-state Ievels of intramicrosomal glucose are observed under symmetrical conditions of Glu-6-P o r vanadate concentrations dur ing influx and efflux experiments, and those levels are directly propor- tional to Glu-6-Pase activity. 4) The rates of both glucose influx and effïux are characterized by t t h values that are independent of Glu-6-P concentrations. 5) Glucose efflux i n t h e presence of s a t u r a t h g concentrations of vana- d a t e was not blocked by 1 ntn phloretin, a n d t h e initial ra tes of e m u x appear directly proportionai to intrave- s icular glucose concentrations, 6) It is concluded that glucose influx into microsomes is tightly Iinked t o Glu- 6-Pase activity, while glucose enlux may occur inde- pendent of hydrolysis, so that microsornal glucose trans- port appears unidirectionai even though i t can be accounted for by diffusion only over t h e accessible range of sugar concentrations.
Given the peculiar localization of glucose 6-phosphatase ( Glu-6-Pase i L (EC 3.1.3.9) into the endoplasmic reticulum. the membrane topology of this enzyme has been controversia1 for many years. While most authors agree that the enzyme is an integral protein tightly associated with the hydrophobic part of the native membrane. as confirmed recently by the amino acid sequence inferred from the isolated cDNA encoding human ( 1)
' This work was supported by Grants ME-10783 and MT-10804 ito G. v. d. W.) from the Medical Resenrch Council of Canada. The costs of publication of this article wcre defrayed in part by the payment of page charges. This article must therefore be hereby marked %dcertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
:I Present address: Institut National de la Santé et de la Recherche Médicale U197. Faculté de Médecine LUexis-Carrel. 69008 Lyon. France.
** To whom correspondence should be addressed: Dept. of Nutrition LTniversité de Montréal, C.P. 6128. Succursale "centre-ville" Montréal 1 QC 1 H3C 357. Canada. Tel.: 5 14-343-7548; Fax: 5 14-343-6 103; E-mail: vandeweg8ere.urnontreal.ca.
C ' The abbreriations used are: Glu-6-Pase. glucose 6-phosphatase; Glu-6-P. glucose 6-phosphate; Tl. liver micmsomal glucose 6-phosphate translocase; T2. liver microsornal phosphate translocase; T3, Iiver rni- crosomal glucose translocase: GLLT. facilitative-diffusion glucose transporter: FSRF-4. fast sampling. rapid filtration apparatu tu. S. patent 07/697.769,.
and mouse (2) Glua-Pases, there is no consensus to date as to whether the catalytic site is oriented toward the lumen of the endoplasmic reticulum or freely accessible from the cytoplaçm. Two major but opposite models have thus emerged during the last 20 years.
In the substrate transport model (3-8). it is thought that glucose 6-phosphate (Glu-6-PI is hydrolyzed to glucose and phosphate in the lumen of the endoplasmic reticulum. The enzyme is thus viewed as a mdticomponent system involving a t Ieast five polypeptides subunits (4) among wbich T l , T2, and T3 would represent specific transporters for Glu-6-P, phos- phate. and glucose, respectively. In support of the substrate- transport model are reports on the successfd purification of T3 ( 6 ) as as cloning of a cDNA encoding a 52-kDa glucose- transport protein (GLUT.7) that has been assimilated to T3 (7). Since GLUT7 has been expressed in COS7 cells (71, a ce11 line that does not normally contain significant amount of GIu-6- Pase activity (81, those studies do not resolve the'functional relationships between glucose transport and Glu-6-P hydroly- sis. In the cornbined conformational fl exibility-substrate trans- port model (9-111, Glu-6-Pase traverses the microsomal mem- brane a s a channel-forming protein embedded within the hydrophobic matrix of the bilayer, a concept that is indeed supported by the cloned sequences of the hunian (1) and mu- rine (2) genes that predict up to SLX transmembrane segments in the secondary structure of the enzyme. In that model, the c=ttaIytic site is assurneci to be located within a water-filled proteinaceous pore accessible to the substrate from the cyto- plasrnic surface of the membrane (11).
In agreement with the conformational model, our recent kinetic studies using a fast sampling, rapid filtration appara- tus (FSRFA, U. S. patent 07/697,769) have brought evidence for a tight-coupIing between Glu-6-P transport and Glu-6-Pase activity (12) but failed to demonstrate any significant transport of D-glucose inta microsomes under zero-trans uptake condi- tions (131.~ In the presence of [14C]Glu-6-P in the incubation medium, however, we aiso showed that tracer glucose accumu- lates within the intramicrosomal space against a concentration gradient (12). In the present studies, we thus report on the kinetics of glucose transport into microsomes in cornparison with those of Glu-6-P hydrolysis by Glu-6-Pase. Our results demonstrate that intramicrosomal glucose accumulation is di- rectly proportional to Glu-6-Pase activity, whik giucose e m u ~
-
* Corresponding to one of the four principal experimental situations that tend themsehes well to kinetic analysis of transport processes as described for example in Ref. 25. Accordingly, in the experiments de- scribed, there was no glucose inside of the rnicrosomes, the membrane side opposite (the trans face) to the uptake medium where the radio- tracer was intmduced (and defined as the cis face). Clearly, then. zero-trans experiments can be performed under influx and emw conditions.

Glucose Transport in Liuer Microsornes 21093
may occur independent of hydrolysis, so that rnicrosomal glu- cose transport appears unidirectional.
EXPEREtIEiVTAL PROCEDURES Microsorne Purification-Liver microsornes were isolated from over-
night-fasted male Wistar rats (200-250 gJ as described previously (12) and resuspended a t a pmtein concentration of 10-16 mg/ml in a 50 a u TRIS-HCI buffer (pK 7.3) containhg 0.25 r sumse. Glucose dehydro- genase activity was used as an index of mimsome integrity since k i n g fully latent in intact mimçornes (13, 14). and our microsomal prepara- tions mutinely displayed 9Sl00ck latency.
Uptake and Ellflu Experiments-Tracer uptake into mimsomes was determined using a FSRFA as described previously (12) except for the omission of EGTA in the incubation media, which are usually adjusted to 1 mi final. Briefly, for zem-tram uptake experiments. 40 pl of rnicrosomes were injected into the incubation chamber of the FSRFA and incubated a t 30 OC in the presence of 0.2-10 a.1 concentrations of Glu-6-P while keeping the concentration of W-L'CIGlu-6-P (specific activity, 200 mCi/mmol. ICN Biomedicais. Montreai, Canada) constant a t 3-4 &ilml incubation medium. For uptake experiments in the presence of vanadate, mimsomes were preincubated or not for 1 min a t 30 "C in the presence of the vanadate concentrations shown in the legend to Fig. 3. For efïïux experiments. ["Clglucose-loaded mi- somes were first prepared upon preincubation for 2 min a t 30 OC in the incubation chamber of the FSRFA using tracer and cold Glu-6-P con- centrations as above before injection of various effectors (50 dl into the uptake medium. In both cases. samples were nutomatically collected a t different time intervals and woshed on nitrocelIulose filters before radioactivity determination in a scintillation counter. Modifications fmm this basic protocol will be indicated in the legends to figures or in the text.
rlssays-Total LU-'JClglucose production from [U-L'C]Glu-6-P hy- drolysis by Glu-ô-Pase was performed with the FSRFA as described previously t 12) except for the omission of EGTA from the incubation media. Protein was assayed according to Bradford ( 15 i using bovine serum albumin as standard.
Presentation of Data and Kinetic Analyses-Intramicrosomal con- tents are expressed as nmoVmg microsomal protein using the specific radioactivity of 0.2 mv IU-"CIGlu-6-P as the reference for al1 concen- tmtions of Glu-&P. Therefore, the apparent microsomai content de- creases with increasing concentrations of cold Glu-6-P due to isotopic dilution in the incubation medium. Intramimsomal concentrations have been estimated by assuming an intravesicular volume of 0.8 z 0.1 ~ Y m g of protein ( 12).
As shown preriously i 12. 161, total glucose production (Pl is charac- terized by a burst phase over an initial period of about 10 s followed by a steady-state phase that proceeds linearly for up to 120 S. Such data car. be fitted to Equation 1
in which V, and V, are the steady-state and initial burst rates of glucose formation. respectively. a is the inverse of the time constant for the hysteretic transition. and Pa represents the background in radioac- tivity. As validated in a previous report (12). V, can be determined independently in parallel wperiments using microsomes permeabilized with 0.4% deoxycholate. and its value was fixed into Equation 1 during the fitting process in order to get more d i a b l e estimations of a a t varying Glu-6-P concentrations.
As discussed and justified in a previous report (121, tracer accumu- lation (A) into mimsomes is best described by the double exponentid Equation 2
in which A, represents the background in radioactivity while A, and A, stand for the steady-state levels of tracer accumulation reached with first-order rate constants k , and k,, respectively. Since k, is 1 order of magnitude larger than k, (121, the uptake time courses were first fitted over the 10-'240-9 time range using Equation 3
A = (A2 -Ai)(l - e-tf3 +Ai (Eq. 3)
in order to determine the parameters A,. A,, and k,. Those values were then used as prompted constants into Equation 2 when anaiyzing the whole time range. Amplitudes and initial rates of accumulation dunng the fast (AhPl. V,,) and slow (AMI',, V,) phases were calculateci as shown in Equations 4 and 5.
The kinetic parameters pertaining to the rate and amplitude data Cv) were determined h m the analysis of the respective displacement curves of tracer (T. 0.2 m\f tracer CIu-6-P taken as a reference) by cold substrate (S) according to Equation 6
which applies to a saturable phenomenon working in parallel with a nonspecific componentisi (17. 18). In that equation. C represents the background vnlue (undisplaceable tracer), while Y,, and Km stand for the maximum rate (V-) or amplitude (AMP-J and the Michaelis- Menten constant of the saturable process. respectively. The inhibition constants rK,) for vanadate (1) were estimoted using
Equation 7
which assumes competitive inhibition relative to Glu-ô-P (S. 0.2 m\f in those experirnents) (19). and where y and y, stand for the inhibited and uninhibited rates or amplitudes, respectivety. while Km has the same meaning as above and can be fixed to the values determined in the corresponding data r 20).
Emux ( E ) data were analyzed according to Equations 8 and 9, which assume either single or double exponential decay, respectively,
and where E, represents the starting level while E , and E, stand for the steady-state levels of intramicrosomal tracer reached with first-order rate constants k, and k,, respectively. The initial efflw rates (V.1 shown in Fig. SB were calcdated from Equation 8 using Equation IO.
In this paper. we have chosen the parameter th, which represents the time at which 50% of a process has k e n completed. to characterize the rates of influx into, and e m ~ from microsomes. This parameter was calculated using Equation 11.
Ln2 'L' = first order rate constants (a. kl. k2) (Eq. 11)
Ail kinetic analyses were performed using the Enzfitter soRware t Elsevier-Biosoft) and nonlinear regression analyses to pertinent equa- tions above were performed using the robust weighting routine in conjunction with the explicit weighting mutine when appropriate. Ac- cordingly, the e m r s associated with the kinetic parameters as given in the text and figures represent the standard e m r s of regression (S.E.R.1 on these parameters.
RESULTS
Cornpardue Kinetics of Intramicrosomal Tracer Uptake and Glu-6-P Hydrolysis-Fig. 1 (closed symbols) extends to Glu-6-P concentrations varying up to 10 m~ Our former conclusion that tracer accumulation into microsomes is a biphasic process thot achieves a steady-state level of uptake over a 2-min period of incubation (12). In contrast, total glucose production h m Glu- 6-P hydrolysis (results not shown), which is also a biphasic process characterized by an initial burst phase, reaches a steady-state rate by 10 s of incubation under similar conditions (12, 16). As shown in Fig. 2 4 1) the rapid phase in tracer uptake (open triangles) and the burst phase in total glucose production (closed squares) appear synchronous and independ- ent of GIu-6-P concentrations with mean th values of 3.5 2 0.2 and 3.2 z 0.4 s, respectively, and 2) the slow phase in tracer uptake (closed circles) occurs during the steady-state phase in glucose production and is also insensitive to Glu-6-P concen- trations (mean t h = 46.4 2 5.3 s).
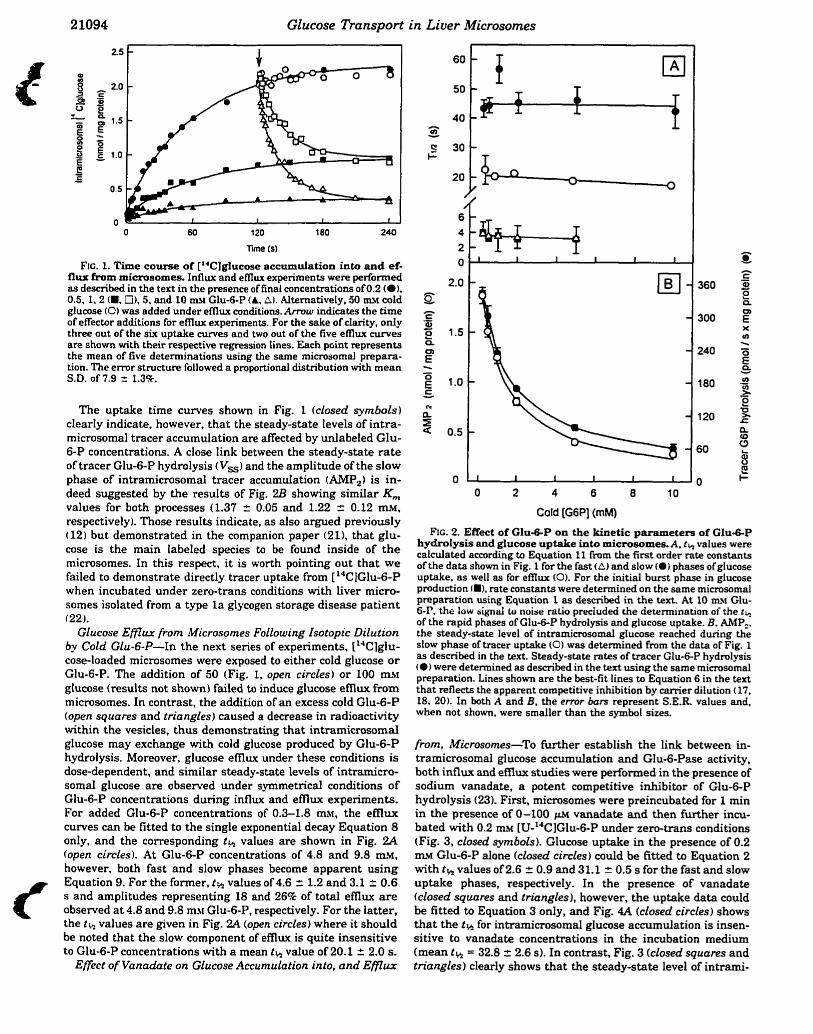
22094 Glucose Transport in Liuer Microsornes
Frc. 1. T i e course of ["Clglucose accumulation into and ef- flux h m microsomes. Influx and e m w experiments were performed as described in the text in the presenceof final concentrations o f 0 2 (0). 0.5.1.2 (M. Cl), 5. and 10 mf Glu-6-P (A, A). Alternatively, 50 UIM cold glucose (0) was added under e f h x conditions. Armw indicates the time of effector additions for emux expenments. For the sake of clarity, only three out of the six uptake m e s and two out of the five e fhx curves are s h o w with their respective regression lines. Each point represents the mean of five determinations using the same microsornal prepara- tion. The e m r structure followed a proportional distribution with mean S.D. of 7.9 = 1.3%.
The uptake time curves shown in Fig. 1 (closed syrnbols) clearly indicate, however, that the steady-state levels of intra- microsomal tracer accumulation are affected by uniabeled Glu- 6-P concentrations. A close link between the steady-state rate of tracer Glu-6-P hydrolysis Wss) and the amplitude of the slow phase of intramicrosomal tracer accumulation ( A m ) is in- deed suggested by the results of Fig. 2B showing similar Km values for both processes (1.37 2 0.05 and 1.22 i 0.12 mf,
respectively). Those results indicate, as also argued previously (12) but demonstrated in the companion paper (21). that glu- cose is the main labeled species to be found inside of the microsomes. In this respect, it is worth pointing out that we failed to demonstrate directly tracer uptake from ["CIGlu-6-P when incubated under zero-trans conditions with liver micro- somes isolated from a type la glycogen storage disease patient (22).
Glucose Ef72ux from Microsomes Following lsotopic Dilution by Cold Glu-6-P-In the next series of experiments, [l4C1glu- cose-loaded microsomes were exposed to either cold glucose o r Glu-6-P. The addition of 50 (Fig, 1, open circles) or 100 a~ glucose (results not shown) failed to induce glucose efflw from microsomes. In contrast, the addition of an excess cold Glu-6-P (open squares and triangles) caused a decrease in radioactivity within the vesicles, thus demonstrating that intramicrosomal glucose may exchange with cold glucose produced by Glu-6-P hydrolysis. Moreover, glucose emux under these conditions is dose-dependent, and similar steady-state levels of intramicro- soma1 glucose are obsewed under symmetrical conditions of Glu-6-P concentrations during influx and emux expenments. For added Glu-6-P concentrations of 0.3-1.8 mr, the efflux curves cari be fitted to the single exponential decay Equation 8 only, and the corresponding t h values are shown in Fig. 2A (open circles). At Glu-6-P concentrations of 4.8 and 9.8 tu!!, however, both fast and slow phases become apparent using Equation 9. For the former, t h values of 4.6 2 1.2 and 3.1 2 0.6 s and amplitudes representing 18 and 26% of total emux a r e observed a t 4.8 and 9.8 mv Glu-6-P, respectively. For the latter, the t S, values are given in Fig. 2A (open circles) where it shouid be noted that the slow cornponent of efflw is quite insensitive to Glu-6-P concentrations with a mean t h value of 20.1 2 2.0 S.
O O 2 4 6 8 1 O
Cold [G6P] (mM)
Frc. 2. Effect of Glu-6-P on the kinetic parameters of Glu&-P hydrofysis and glucose uptake into mictoaomes. A. cl, values were calculated according ta Equation 11 fmm the first order rate constants of the data shown in Fig. 1 for the fast (A) and slow (a) phases ofglucose uptake, as well as for e f n w (O). For the initial burst phase in glucose production (Il. rate constants were determined on the same microsamal preparation using Equation 1 as describeci in the text. At IO m~ Glu- 6-I', the low ziignd il noise ratio preciuded che determination of the tk of the rapid phases of Clu-û-P hydrolysis and glucose uptake. B. AMP,. the steady-state level of intrarnimsornal glucose reached during the slow phase of tracer uptake (0) was detelnined h m the data of Fig. 1 as descrïbed in the kxt. Steady-state rates of tracer Glu-6-P hydrolysis
were determined as described in the text using the same microsoma1 preparation. Lines shown are the best-fit lines to Equation 6 in the text that reflects the apparent competitive inhibition by carrier dilution (17, 18, 20). In both A and B. the e m r bars represent S.E.R. values and. when not shown, were smaller than the symbol sizes.
fiorn, ~Uicrosornes-To further establish the link between in- tramicrosomal glucose accumulation and Glu-6-Pase activity, both influx and efflwc studies were performed in the presence of sodium vanadate, a potent competitive inhibitor of Glu-6-P hydrolysis (23). First, microsornes were preincubated for 1 min in the presence of 0-100 vanadate and then further incu- bated with 0.2 m~ [U-'4ClGlu-6-P under zero-tram conditions (Fig. 3, closed symbols). Glucose uptake in the presence of 0.2 m w Glu-6-P alone (closed circles) could be fitted to Equation 2 with th values of2.6 2 0.9 and 31.1 2 0.5 s for the fast and slow uptake phases, respectively. In the presence of vanadate (closed squares and triangles), however, the uptake data could be fitted to Equation 3 only, and Fig. 4A (closed circles) shows that the t% for intramicrosomd glucose accumulation is insen- sitive to vanadate concentrations in the incubation medium (mean th = 32.8 2 2.6 s). In contrast, Fig. 3 (closed squares and
Effet of Vanadate on Glucose ~ccumu&otîon into. and Ef lux triangies) clearly shows that the steady-state level of intrami-
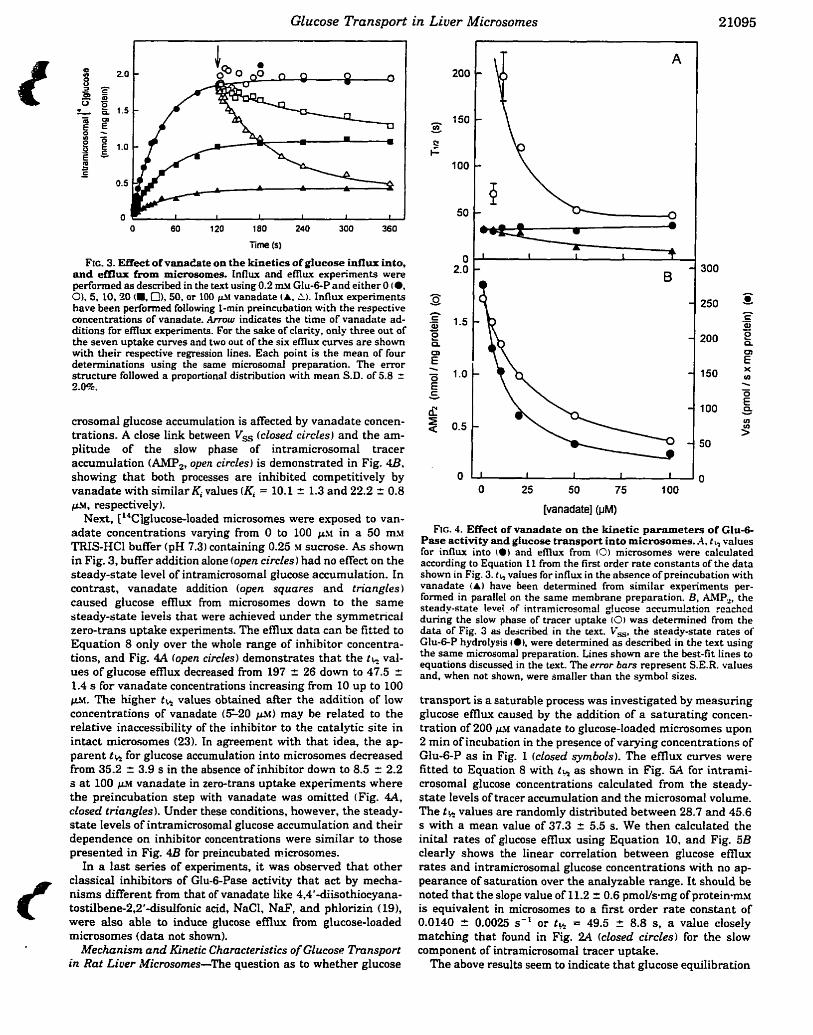
Glucose Transport in Liver Microsornes
Frc. 3. Enect ofvanadate on the kirietics of glucose intlux into, and emur from mimaomes. influx and emux expenments were performed as described in the text u i n g 0.2 m.! Glu-6-P and either O (0, O), 5, 10, '20 (W. 0). 50. or 100 +\r vanadate (A. A). Influx experiments have been performed folIowing 1-min preincubation with the respective concentrations of vanadate. Armw indicates the time of vanadate ad- ditions for eflïwt experiments. For the sake of ctarity, o d y three out of the seven uptake cuves and two out of the six emw m e s are shown with their respective regression lines. Each point is the mean of four determinations using the same microsomal preparation. The error structure followed a proportionai distribution with mean S.D. of 5.8 = 2.0%.
crosomal glucose accumuiation is affected by vanadate concen- trations. A close lirik between VS (closed circles) and the am- plitude of the slow phase of intramicrosomal tracer accumulation (AM.,, open circles) is demonstrated in Fig. 4B. showing that both processes are inhibited cornpetitively by vanadate with similar Ki values (Ki = 10.1 2 1.3 and 22.2 + 0.8 p.!!, respectively).
Next, [14C]glucose-loaded microsomes were exposed to van- adate concentrations varying From O to 100 p~ in a 50 m.1 TRIS-HCl buffer (pH 7.3) containing 0.25 M sucrose. As shown in Fig. 3, b a e r addition alone (open circles) had no effect on the steady-state level of intramicrosomal glucose accumulation. In contra t , vanadate addition (open squares and triangles ) caused glucose e m w from microsomes down to the same steady-state ieveis that were achieved under the symmetncd zem-trans uptake experiments. The e m ~ data c m be frtted to Equation 8 only over the whote range of inhibitor concentra- tions, and Fig. 4.4 (open circles) demonstrates that the th val- ues of glucose emux decreased fimm 197 = 26 down to 47.5 2 1.4 s for vanadate concentrations increasing from 10 up to 100 FH. The higher tr i values obtained &r the addition of Iow concentrations of vanadate (S20 pst) may be related to the relative inaccessibility of the inhibitor to the catalytic site in intact microsomes (23). In agreement with that idea, the ap- parent t k for glucose accumulation into microsomes decreased from 35.2 2 3.9 s in the absence of inhibitor down to 8.5 z 2.2 s a t 100 p~ vanadate in zero-trans uptake experiments where the preincubation step with vanadate was omitted (Fig. 4 4 closed triangles). Under these conditions, however, the steady- state levels of intramicrosomal glucose accumulation and their dependence on inhibitor concentrations were similar to those presented in Fig. 4B for preincubated microsomes.
In a last series of expeiiments, it was observed that other classical inhibitors of Glu-ô-Pase activity that act by mecha- nisms different from that of vanadate like 4.4'-diisothiocyana- tostilbene-2.2'-disulfonic acid, NaCI, NaF', and phlorizin (191, were also able to induce glucose efflux from glucose-Ioaded microsomes (data not shown).
Mechanism and Kinetic Characteristics of Glucosc Transport in Rat Liver ~Wcmsomes-The question as to whether glucose
[vanadate] (FM)
FE 4. Effect of vanadate on the kinetic parnmeters of Glu-6- Pase activity and glucose transport into microsomes.A. t h values for influx into (a) and emux from (0) microsornes were cnlculated according to Equotion I I from the first order rate constants of the data shown in Fig. 3. th vatues for influx in the absence of preincubation with vanadate (A) have been determined from similar experiments per- forrned in parallel on the same membrane preparation. B. AMP-, the steady-state Iwei of intramicmsornal glucose sccumuhtion rcachcd during the slow phase of tracer uptake (01 was determined from the data of Fig. 3 as dejcribed in the text. V,, the steady-state rates of Glu-ô-P hydmiysis (a), were determined as described in the text using the same microsomnl preparation. Lines shown are the bat-fi t lines to equotions discussed in the text. The error bars represent S.E.R. vaiues and, when not shown, were smaller than the symbot sizes.
transport is a saturable process was investigated by measuring glucose emux caused by the addition of a saturating concen- tration of 200 p.v vanadate to glucose-loaded microsornes upon 2 min of incubation in the presence of varying concentrations of Glu-6-P as in Fig. 1 (closed symbols). The e m w curves were fitted to Equation 8 with th as shown in Fig. 5A for intrami- crosomal glucose concentrations calculated from the steady- state levels of tracer accumulation and the microsomal volume. The t k values are randornly distributed between 28.7 and 45.6 s with a mean value of 37.3 r 5.5 S. We then calcutated the inital rates of glucose emux using Equation 10, and Fig. 5B clearly shows the linear correlation between glucose e m w rates and intramicrosomal glucose concentrations with no ap- pearance of saturation over the anaiyzable range. It should be noted that the dope value of 11.2 = 0.6 pmoi/s.mg of protein-rnsf is equivaient in microsomes to a first order rate constant of 0.0140 2 0.0025 s-' or t h = 49.5 i 8.8 s, a value closely matching that found in Fig. 2A (closed circles) for the slow component of intramicrosomal tracer uptake.
The above results seem to indicate that glucose equilibration
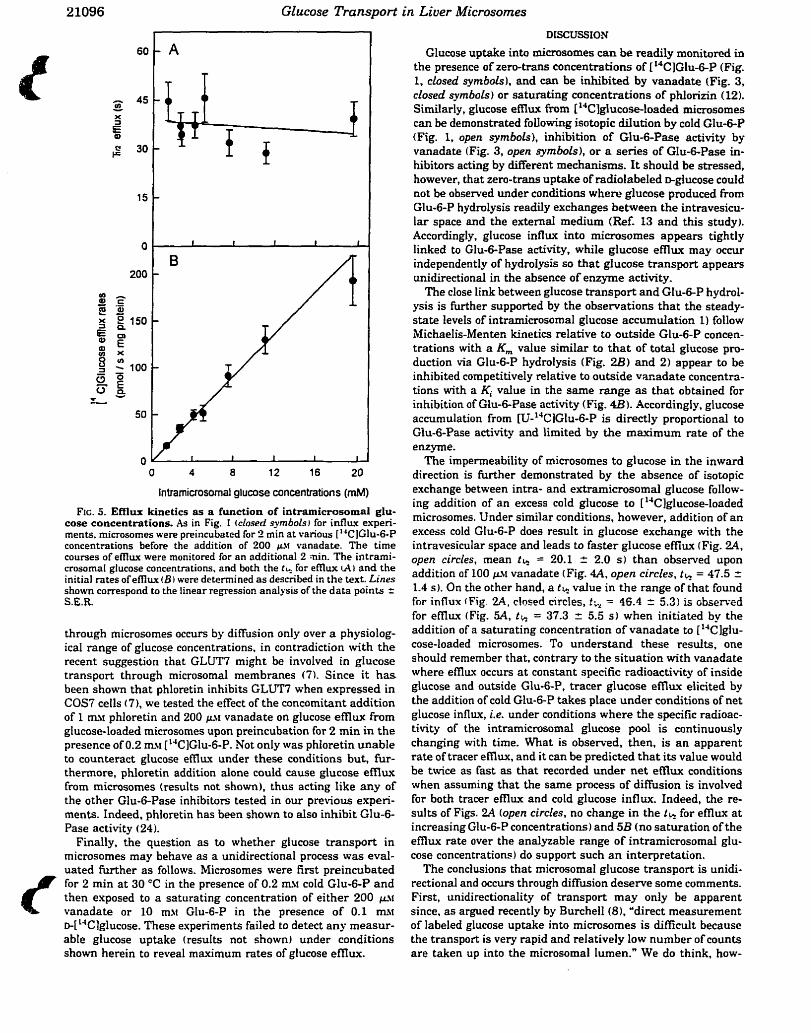
Glucose Transport in Liuer Microsomes
Intramicrosornal glucose concentrations (mM)
FIG. 5. Efflux kinetics as a function of intramicrosomd glu- cose concentrations. As in Fig. 1 iclosed syrnbols) for influx experi- ments. microsomes were preincubated for 2 min at various ['"ClGlu-6-P concentrations before the addition of 200 g.\t vanadate. The time courses of ef l lw were monitored for an odditional 2 ain. The intrami- crosomal glucose concentrations, and both the f- for emux U I and the initial rates of emux ( B I were determined as described in the text. Lines shown correspond to the linear regression analysis of the data points r S.E.R.
through microsomes occurs by diffusion only over a physiolog- ical range of gIucose concentrations. in contradiction with the recent suggestion that GLUT7 might be involved in glucose transport through microsomal membranes (7). Since it h a s been shown that phloretin inhibits GLUT7 when expressed in COS7 ceils (7), we tested the effect of the concomitant addition of 1 n~ phloretin and 200 vanadate on glucose efflux from glucose-loaded microsomes upon preincubation for 2 min in the presence of 0.2 msr ['"C]Glu-6-P. Not only was phloretin unable to counteract glucose emux under these conditions but, fùr- thennore, phloretin addition alone could cause glucose emux from microsomes (results not shown), thus acting like any of the other Glu-6-Pase inhibitors tested in our previous experi- ments. Indeed, phloretin has been shown to aiso inhibit Glu-6- Pase activity (24).
Finally, the question as to whether glucose transport in microsomes may behave as a unidirectional process was eval- uated fiuther as follows. Microsomes were k i t preincubated for 2 min a t 30 "C in the presence of 0.2 m!r cold Glu-6-P and then exposed to a saturating concentration of either 200 vanadate or 10 mv Glu-6-P in the presence of 0.1 mtr D-[ 'JClglucose. These experiments failed to detect any measur- able glucose uptake (results not shownl under conditions shown herein to reveal maximum rates of glucose e f l h .
DISCUSSION
Glucose uptake into microsornes can be readily monitored in the presence of zero-trans concentrations of 1'4C]Glu-6-P (Fig. 1, closed syrnbols), and can be inhibited by vanadate (Fig. 3, closed syrnbols) or satutating concentrations of phlorizin ( 12). Similarly, glucose e m w From [lJClglucose-loaded microsomes c m be demonstrated foUowing isotopic dilution by cold Glu-6-P (Fig. 1, open syrnbols), inhibition of Glu-6-Pase activity by vanadate (Fig. 3, open symbols), or a series of Glu-6-Pase in- hibitors acting by different mechanisms. It should be stressed, however, that zero-transi uptake of radiolabeled glucose could not be observed under conditions where glucose produced h m Glu-6-P hydrolysis readily exchanges between the intravesicu- lar space and the external medium (Ref. 13 and this studyi. Accordingly, glucose influx into microsornes appears tightly linked to Glu-6-Pase activity, while glucose efflux rnay occur independently of hydrolysis so that glucose transport appears unidirectional in the absence of enzyme activity.
The close link between glucose transport and Glu-6-P hydrol- ysis is further supported by the observations that the steady- state levels of intramicrosomal glucose accumulation 1) follow Michaelis-Menten kinetics relative to outside Glu-6-P concen- trations with a Km value similar tn t ha t of total glucose pro- duction via Glu-6-P hydrolysis (Fig. 223) and 2) appear to be inhibited competitively relative to outside vacadate concentra- tions with a Ki value in the same range a s that obtained for inhibition of Glu-6-Pase activity (Fig. 4B). Accordingly, glucose accumulation from II_I-'"CIGlu-6-P is directly proportional to Glu-6-Pase activity and limited by the maximum rate of the enzyme.
The impermeability of microsornes to glucose in the inward direction is further demonstrated by the absence of isotopic exchange between intra- and extramicrosomal glucose follow- ing addition of an excess cold glucose to [lJC]giucose-loaded microsomes. Under similar conditions, however, addition of a n excess cold Glu-6-P does resuIt in glucose exchange with the intravesicular space and leads to faster glucose efflux (Fig. 2 4 open circles, mean t k = 20.1 2 2.0 s) than observed upon addition of 100 &M vanadate (Fig. 4A, open circles, t k = 47.5 = 1.4 SI. On the other hand, a tb value in the range of that found for influx {Fig. 2A, closed circIes, th = 46.4 = 5.3) is çbserved for emw (Fig. 5A, th = 37.3 2 5.5 s ) when initiated by the addition of a saturnting concentration of vanadate to ['"C ]glu- cose-loaded microsomes. To understand these results, one shouid remember that, contrary to the situation with vanadate where eilïwt occurs a t constant specific radioactivity of inside glucose and outside Glud-P, tracer glucose efflux elicited by the addition of cold Glu-6-P takes place under conditions of net glucose influx, i.e. under conditions where the specific radioac- tivity of the intramicrosomal glucose pool is continuously changing with time. What is observed, then, is an apparent rate of tracer emux, and it can be predicted tha t its value would be twice as fast as that recorded under net emux conditions when assuming that the same process of diffusion is involved for both tracer efllux and cold glucose influx. Indeed, the re- sults of Figs. 24 (open circles, no change in the 1% for efl iw a t increasing Glu-6-P concentrations) and 5B (no saturation of the e f h x rate over the analyzable range of intramicrosomai glu- cose concentrations) do support such an interpretation.
The conclusions that microsomal glucose transport is unidi- rectional and occurs through diffusion deserve some comments. First, unidirectionality of transport rnay only be apparent since. a s argued recently by Burcheil(8). "direct measurement of labeled glucose uptake into microsomes is difficult because the transport is very rapid and relatively low number of counts are taken up into the microsomal Iumen." We do think, how-

Glucose Transport in Liver Microsornes 21097
ever. that zero-trans glucose transport, if occurring, should be rneasurable in that preparation; 1) glucose uptake can be readily meas~red~ in other vesicle systems with sirnilarly small intravesicular volumes (17); 2) the problem of low number of counts can be partly solved by using pure tracer concentrations of gIucose with high specific activity ( 18) ( the sensitivity of the tnuisport assay in t e m s of measurable cpms would be maxi- mum under those conditions, see Equation 6 when S = 0); and 3) the rapidity of the uptake process is readily overcome in our laboratory when using the FSRFA. In agreement with those statements, it is worth emphasizing that zero-tram uptake of inorganic phosphate could be successfully measured in both rat and human Iiver mimsomes (22). Moreover, since phosphate was taken up into an equivalent microsomal space of 1.2 pl/mg of protein (221, thus closely rnatching that of 0.8 pVmg of protein determinad in previous studies using 'H,O (121, it can be concluded W h e r that the use of the FSRFA does not dam- age a major fiaction of the vesicles such a s to preclude signif- icant zero-trans uptake measurements. Accordingly, our fail- ure to detect glucose uptake under similar and appropriate experimental conditions can thus be taken as evidence that glucose does not enter into a measurable intravesicular space a t any significant rate. We do not rule out, however, the pos- sibility that glucose may permeate the endoplasmic reticulum membrane through passive diffusion with extremely Iow rate constant. Next, diffusion may only be apparent since, in the low range of substrate concentrations, a low affinity carrier or channel may not show saturation kinetics (the apparent diffi- sion constant in that case is equal to the V,JK,,, ratio of the transport protein. see Equation 6 with S and T CC Km). Unfor- tunately, due to their low glucose permeability, it is not possi- ble to passively load microsomes in a reliable way a t fixed glucose concentrations, and thuç to extend the range of sugar concentrations over which saturation kinetics (or the lack thereon might be observed in plots like those shown in Fig. 5B. Accordingly, the upper range of analyzsble intravesicuIar glu- cose concentrations is fixed to 15-20 rn%t when active loading from [1JClGlu-6-P is used, a value dictated by the V,,, of Glu-6-P hydrolysis by Glu-6-Pase. In that respect, the slight deviation from linearity observed a t 20 m>r inside glucose in the plot nf Fig. 5B should not be taken as evidence for the begin- ning of the development of a hyperbolic relationship because the low signai to noise ratio that is achieved a t close to complete saturation of the enzyme does not allow for a precise estimate of the intramicrosomal glucose concentration. Moreover, if one assumes that a transport protein like GLUT7 is present within the microsomal membrane, we may expect l i inhibition of glucose d 'f ' l~ fmm microsomes by phloretin (7). 2) accelerated efflux of glucose from ['4Clglucose-loaded microsomes following addition of an excess cold glucose to the incubation medium, 3 ) zero-trans uptake of tracer glucose with th in the range of
30-50 s since the same V,,JK,,, ratios shouid be observed under influx and emuu conditions independent of whether the transporter is symmetrical or not due to the law of microscopic reversibility (251, and 4) zero-trans uptake of tracer glucose with overshoot phenomenon (25) in microsomes loaded with cold glucose. Since none of these expectations could be demon- strated in our experiments, we must conclude that glucose transport is indeed unidirectional and may not involve GLUTI. Shouid a specific glucose transport protein (T3) be present in native liver microsomes, however. we do state that its kinetic properties are different from those demonstrated for GLUTT in COS7 cells (7). maybe due to a close association with a multi- component enzyme cornplex (4, 6, 8). In any case. it should be stressed that the mode1 proposed in the companion paper (21) requires the presence of neither GLUT7 nor any other glucose transport protein but for Glu-6-Pase in order to explain the steady-state kinetics of Glu-6-P hydrolysis and glucose ex- change through the microsomal membrane.
REFERENCES Lei. KJ. . Shelly, L L. Pan, C . J . . Sidbury. J. B.. and Chou. J. Y. I 19931
Science 262,580 -083 Shelly, L. L. Lei. KJ.. Pan. C.J.. Saknta. S. F.. Ruppert. S.. Schuu. Ci.. and
Chou, J. Y. 119931 J. Biol. Chem. 268,21482-21485 Arion, W. J.. Lange. A J.. Walls. H. E.. and Bollas. L. M. t 19801 d. Biuf. Chrm.
W. 10396-10406 Burchell. A. I 1990) F S E B ./. 4.2978-2988 Waddell. 1. D.. Lindsay. J. G.. and Burchell. h I 1988, FEBS Lut[. 229. 179-1*:! WnddeIl. 1. D.. Scott. H.. Grant. A. and Burchell. A. 119911 Btocheni. J. 275.
363367 Wnddell. I. D.. Zomerschoe, A G., Voice. SI. W.. and Burchell, h r 19931
Brochem. J. 286, 173-177 Burchell. A. I 1994 I Biochem. Soc. Tmns. 22.658 -663 Nordlie. R C.. and Sukntki. K. A 119851 in The Enqmrs of Biol«grcal
Memhmnes 1 Martonosi. h N.. edi Vol. 3. pp. 349-398. Plenum Publishing Corp.. Xew York
Sehulze. H.-CI.. and Speth. hl. I 1980, Eur. ./. Biochem. 106. 50.S514 Schulze. H . X . . Nolte. B., and b n n l e r . R. ( 19861 ./. Bid. Chrm. 261.
16571-16578 Berteloot. A. Vidal. H.. nnd van de Werve. C. i 1991, J- Brui. Chem. 266.
5497-5507 Romanelli. A. St-Denis. J.-F.. Vidai. H.. Tchu, S., and vnn de Werve. C. 1994 1
Biochem. Biophys. Rrs. Commun. 200, 1491-1497 Bublitz. C.. und Stenvenson. S. I 19881 Biochrm. Biophys. Acta 965, 90-9- Brndford. M. ( 1976) Anal. Biochem. 72.2.18-25.1 Vidal. H.. Berteloot. A. Lame. 3I.J.. St-Denis, J.-F.. and vnn de Werve. G .
I 19921 FEBS Ln. 302. 197-200 Benelwt. A. and Semenzn. G. I 1990 i .\luth& En.zymoC. 192. 409 -437 Chenu. C.. and Berteloot. A 119931 J. .\-irrnbr. Bioi. 13% 9.5-1 13 Nordlie, R. CI ( 1982, iCfrthds Enzymol. 87, 319-7.5.7 Segel. 1. K. (1975) Enzyme Kinetrcs. pp. 100-160. Wiley-Intersciencr
Publication. New York BerteIoor. A.. St-Denis. J.-F.. and van de Werve. C. I 1995) J. Brol. Chrm. 270.
21098-21 102 St-Denis. J.-F.. Comte. B.. Nguyen. D. K. Seidman. E.. Pandis. K. Lew. E..
and van de Werve. G. 11994) J. Cfin. Endocnnol. Metab. 79, 955-959 Singh. J.. Nordlie. R C.. and Jorgenson. R h (1981) Biochtm. Bioph-vs. .Wu
678,477-182 Zerr. C.. and Novoa. W. B. 11968) Biochem. Biophp. Res. Commun. 32.
129-133 Stein W. D. (19811 in ,lfernbmne Transport ~Bonting, S. L.. and De Pont. J. J..
edsI pp. 123-157. EtsevierNorth Hollnnd Biomedical Press. Amsterdam

3.3- Résidus impliqués dans le processus catalytique de la
glucose6-phosphatase
3.3.1- Article 3 : Inactivation of rat liver microsomal giucose-6-phosphatase by
dicyclohexylcarbodiunide : evidence for the role of carboql
groups in catalysis. Annabi B., van de Werve G. & Rider M.H.

Article 3
Hypothèse: Le mécanisme catalytique de la Gic-6-Pase nécessite une triade
catalytique His-Asp/Glu-His semblable à celui de la famille des histidine
phosphatases.
Des études initiées par Feldrnan et Butler (1969) ont suggéré la présence de
résidus histidine impliqués dans le site actif de la Glc-6-Pase. De plus, le
diéthy lpyrocarbonate, un agent modifiant spécifiquement les résidus histidine, inhibe la
GIc-6-Pase (Arion et a(.. 1984) suggérant que le mécanisme catalytique de la Glc-6-
Pase implique une triade catalytique His-AspIGlu-His. L'absence de la séquence
consensus RHG exclut, par contre, la Gic-6-Pase des protéines faisant partie de la
famille des histidine phosphatases (Lange et al., 1994). Dans cette étude nous avons
utilisé le dicyclohexylcarbodümide comme agent chimique pouvant modifier
spécifiquement les groupements carboxyles des résidus aspartate et glutamate pour
étudier leur implication dans le mécanisme de remmaissace etlou de liaison du
substrat.

71
INACTIVATION OF RAT IWVER MICROSOMAL GLUCOSE-6-
PHOSPHATASE BY DICYCLOHEXYLCARBODIIMIDE
evidence for the role of carboxyl groups in catalysis
Borhane ~nnabi', Gérald van de wervel' and Mark H. Ride?
1 Laboratoire d'Endocrinologie Métabolique, Departments of Biochemistry and Nutrition, University of Montreal, C. P. 6 128, Succursale 4 Montréal, Québec
H3C 3 J7, Canada
'~ormone and Metabolic Research Unit, International Institute of Cellular and Molecular Pathology and University of Louvain Medicai School, Avenue
Hippocrate, 75, B- 1200 Bmssels, Belgium.
*Address for correspondence: Department of Nutrition, University of Montreal, C .P. 6 1 28 Succursale centre-ville, Montreai, Quebec H3C 337, Canada. Tel: (5 14) 343-7546 Fax: (514) 343-6103 E-mail : vandeweg@ere. umontreal. ca
Keywords : giucose-6-phosphatase; carboxyl groups; catalysis; dicyclohexylcarbodümide
CHAPSO, (3-[(3-Cholamidopropyl)-dimethy1ammonio]-2-hydro- 1 -propanesulfonate); DCCD, N,N dicyclo hexylcarbodiimide; Glc-6-Pase, glucose-6-phosp hatase (EC 3.1.3 -9); Gic-6-P, glucose 6-phosphate; SDS-PAGE, sodium dodecylsulphate polyacrylamide gel electrophoresis.

Incubation of partially purifieci glucosed-phosphatase (Glc-6-Pase) with
dicyclohexylcarbodümide (DCCD) and glycine ethyl ester Ied to its t h e - and
dose-dependent inactivation. Glucose &phosphate, mannose &phosphate, carbamyi
phosphate, inorganic pyrop hosphate and inorganic phosphate protected against the
DCCD-induced inactivation of Glc-&Pax, whereas sn-glycerol 3-phosphate,
glucosamine or glucose offered M e protection. Incubation with DCCD and [1-"CI
glycine ethyl ester labetied a band ofM, 68000, which was decreased in the presence of
glucose 6-p hosphate, mannose 6-p hosp hate and carbarnyl phosphate, but not by sn-
glycerol 3-phosphate or ATP. The findings are dimssed in relation to the rote of
carboxyl groups ir~ catalysis or substrate binding.
Introduction
Glucose-6-phosphatase (Glc-6-Pase) is a microsomai membrane protein,
which catalyses the hydrolysis of glucose 6-phosphate (Glc-6-P) to glucose and
inorganic phosphate. This reaction represents the last step in the gluconeogenic
pathway [Il . The enzyme is expressed in liver, pancreatic B-cells and kidney. The
hepatic microsomal Glc-6-Pase has been proposed to be a multicomponent system
consisting of a catalytic subunit with its active site facing the lumen of the
endoplasmic reticulum, together with transport proteins for Glc-6-P, glucose and
inorganic phosphate, which shuttle the substrate/products between the cytosol and
the endoplasmic reticulum lumen [Z]. Genetic deficiencies in any of these
components leads to different types of glycogen storage diseases, emphasizing the

73
importance of the Glc-6-Pase system in the control glucose and glycogen
metabolism [3]. This concept of the organkation of rnicrosomal Glcd-Pase has
been referred to as the substrate transport model [4].
However, kinetic studies [5,6] and accessibility to chernical modifkation
reagents [7] and proteases [8] suggest a codonnational model, in which
Glc-6-Pase is an integrated system with catalytic and transport functions residing
on a single transmembrane protein [9]. The latter rnodel postdates a hysteretic
transition of Glc-6-Pase from a fast non-specific form, present in the 6rst few
seconds, to a slower enzyme, which is specific for Glc-6-P [ 5 ] .
To idenfiSr the catalytic subunit of Glc-6-Pase, a mouse Glc-6-Pase cDNA has
ben isolated by diierentidy screening a mouse liver cDNA library with mRNAs f?om
normal and albino mutant rnice, the latter expressing rnarkedly reduced Glc-6-Pase
actMty [IO]. The deduced arnino acid sequence predicts a protein with a molecular
m a s of 405 13 Da. Oligonucleotide primers derived from the mouse cDNA were used
to isolate the rat [I l ] and human [I2] Glc-6-Pase cDNAs. Patients with glycogen
storage disease type la (defect in Glc-6-Pase catalysis) were found to have point
mutations in the Glc-6-Pase gene [12,13], whereas the Glc-6-Pase gene was normal in
patients with glycogen storage disease types 1 b and 1 c (defects in Glc-6-P translocase
and the phosphate/pyrophosphate transporter, respectively) [ 131. In FA0 hepatoma
cells, the Glc-6-Pase mRNA was induced by dexamethazone and supresseci by insulin
and, paradoxidy, also by cyclic AMP [Il]. ln livers f?om diabetic rats, Glc-6-Pase
mRNA was increased [14].

74
A major problern encountered in the study of Glc-6-Pase is its instabiiity once
extracted fiorn the microsornal membrane. Although the enzyme has never been
pur8ed to homogeneity, partial pudications of Glc-6-Pase have been reported [IS-
171. It has been suggested that Glc-6-Pase is a member of the histidine phosphatase
family [18], which includes members of the acid phosphatases, the phosphogiycerate
mutases, the bisphosphoglycerate mutases and fructose-2,6-bisphosphatase. Ste-
directed mutagenesis studies on the histidine phosphatases, fhctose-2,6-bisphosp hatase
[19], E. coli acid phosphatase [20] and rat prostatic acid phosphatase 12 11 have led to
the identification of carboxyl groups, which are involveci in d y s i s . In these enzymes,
carboxyl groups act as acid catalysts by protonating the leaving group or as base
catalysts by activating water during phosphorylenzyme breakdown.
This paper describes the inactivation of partially purified Glc-6-Pase by
N,N'dicyclohexylcarbodiimide @CCD), which chemically modifies carboxyl
groups in proteins. The results show that carboxyl groups are important for
cataiysis or substrate binding in rat liver Glc-6-Pase. During the course of these
studies a 68000-M, protein was labelled.
2. Materials and methods
2.1. Materials
Hydroqlapatite-ultragel was from IBF. [14c]~cc~, w-''c] L-glycerol 3-
phosphate and w-14c] glucose-6-phosphate were Eom Amenham International. [ 1 - 14 Cl glycine ethyl ester was from New England Nuclear. Non-radioactive DCCD and
glycine ethyl ester were from Janssen, Berse, Belgium. Rabbit phosphogiucomutase

was from Sigma AU other biochemicals were from Sigma or Boehringer.
2.2. Pun~cation of rat INer microsomol Glc-&Pare
Livers fiom 10 male Wstar rats (200-250g), starved for 48h, were rapidly
excised and homogenized as desaibed previously [SI in 5 volumes (v/w) of a b a e r
containing 250 m M sucrose, 1 m . EDTA, L m . EGTA, 2 mM dithiothreitoi, 2 mM
Benzamidine-HCi, 0.2 mM phenylmethanesulfonyl fiuoride, 0.5 &mL leupeptin, and
10 mM HEPES pH 7.0 at 4OC. The resulting homogenate was centrifùged at 12,000g
for 10 min at 4OC to remove nuclei and ceil debris. Microsornes were isolated by
centrifiiging the supernatant at 100,00Og, for 60 min at 4°C.
The microsomal hction was washed once by resuspending in a buf5er
containing 1 mM KHZPO4 1 mM EDTA, 0.5 mM EGTA 1 mM dithiothreitol, 2 mM
Benzarnidine-HCI, 0.1 rnM phenylmethanesulfony1 fluoride, 1 pg/mL Ieupeptin, 20%
(vh) Glycerol, and 20 mM HEPES pH 7.0 at 4OC @uffer A) supplemented with 0.6 M
NaCl. Mer centrifugation, microsornes were resuspended in b d e r A containuig 0.6 M
NaCl at a protein concentration of 20 mglml and solubilized for 30 min at 4°C with
0.8% (wh) CHAPSO. Foilowing centrifugation at 100 ,000~, the resulting
supernatant was applied to a column of hydroxylapatite (2.5 x 30 cm) equiiibrated in
buffer A supplemented with 0.04% (w/v) CHAPSO. Protein was eluted fiom the
column at a flow rate of 30 mVhr. Fractions containing Glc-6-Pase activity, which came
in the flow through, were pooled and stored frozen at -80°C.
2.3. Innrbatiom with DCCD/glycine ethyf ester
The hydroxylapatite fiaction (0.2-0.7 mghi) was incubateci in buffer

76
containhg 50 mM MES/6.25% (v/v) glycerol pH 6.5 and 10 rnM glycine ethyl ester at
30°C with the concentrations of DCCD indicated in the Iegends to the figures and
table. At the indicated times, aliquots were rernoved for Glc-6-Pase assay. Solutions of
DCCD were prepared fksh in DMSO. The f i r d concentration of DMSO in the
incubations was not more than 1% (vlv), a concentration which had no effkct on
Glc-6Pase activity. For the labelhg studies, the concentration of [ 1 4 ~ ] ~ ~ ~ ~ was 0.3
mM (speciiïc radioactivity 120 cpm per pmol). In some experirnents, incubations were
carried out in the presence of 0.3 mM DCCD in combination with [L14c] glycine ethyl
ester, which was present at 1.25 mM (specific radioactivity 95 cpm per pmol). Mer 15
min, protein was precipitated with 1 mi 10% (wlv) trichloroacetic acid and collecteci by
centrifùgation (Eppendorfrnicrofuge, 14000 rpm x Smin). The precipitated protein was
washed once with 1 ml of acetone and the pellet was dried in a Speed Vac
concentrator. The pellet was dissolved in 25 mi of 1 % (wh) SDS, 10% (vlv) glycerol,
50 mM dithiothreitol, 12 mM Tris-HCI, pH 6.8 (SDS-PAGE sarnple buffer). Following
SDS-P AGE, staining/destaining and gel drying, IabeUed proteins were visualized by
autoradiogaphy.
2.4. Glc-6-Pase asrqy
Glc-6-Pase was assayed at 30°C in a h a i volume of 50 ml of 50 m M HEPES
pH 7.0 containhg 5 mM [U-~'C]G~C-~-P (specific radioactivity 100-200 cpm per
nmol). At the end of the incubation, the reaction was stopped by addig O. 1 ml of 0.3
M ZnSOd. Unhydrolysed Glc-6-P was precipitated by adcihg 0.1 mi of saturateci
Ba(O& [22]. FolIowhg centrifbgation (Eppendorf microfùge, 14000 rprn x Smin),

77
glucose production was measured by wunting 0.1-0.15 mi of supernatant. One unit of
enzyme activity is the amount which catalyses the formation of 1 mmol of CU-
14~]glucose/min under the conditions of the assay.
2.5. Other methodr
Protein concentration was mea~u~ed after trichloroacetic acid precipitation of
the method of Lowry [23] using bovine serum albumin as a standard. SDS-PAGE [24]
analysis was carrieci out in 0.8 mm mini slab gels (10 x 8 cm).
3. Results and discussion
A partial purification of rat fiver rnicrosornal Glcd-Pase was achieved by
detergent extraction with CHAPSO and chrornatography on hydroqlapatite. Glc-6-
Pase was purified 15 to 20-fold by this produre and had a specific activity of 1-2
units per mg of protein (table 1). Moreover, ail the Glc-6-Pase activity present in the
rnicrosomes was recovered in the hydroxylapatite fraction (table 1) and the activity of
non-specific phosphatase ( W - ~ ~ C ] sn-glycerol 3-phosphatase) present in the
hydroxylapatite fiaction was negîigible. In addition we were unable to detect any
phosphoglucomutase activity in the hydroxylapatite fiaction, whereas activity was
detected in intact microsornes enzyme has been suggested to be labelleci by
[ . 'Z~]~ lc -6 -~ in intact and solubilized rat liver rnicrosomes [25]. Glc-6-Pase in the
hydroxylapatite hction had a K, for Glc-6-P of 1.7 mM (not shown) and was stable
for at least 6 months at -80°C.

3.2. Inactivcrtiot~ of GZc-&-Pm in the h y a h y w t e f action by DCCD
Dicyclohexylcarbodümide (DCCD) is a hydrophobic carbodide, which
partitions into the hydrophobic are of proteins and chemically modifies carboxyl
groups. The reactive carboxyl group forms an unstable o-acylisuurea derivative [26],
which bas three fates. It can 1) hydrolyse to regenerate the unmodified carboxyl group,
2) breakdown to a stable O-acylurea derivative, 3) react with an arnino group either
fiom a nearby arnino residue in the polypeptide chah (forming an iritramolecular
cross-link) or ffom an exogenously added nucleophile, such as glycine ethyl ester [27].
Incubation of the hydroxylapatite Faction in MES buEer at pH 6.5 and 30°C
with DCCD led to the irreversible loss of GIc-6-Pase activity, which was
dose-dependent and which was completely protected by Glc-6-P (Fig. 1). Inclusion of
10 mM glycine ethyl ester in the incubations, to act as a nucleophile in the chernical
modification reaction, did not af5ect the sensitivity of Glc-6-Pase towards inactivation
by DCCD (not shown). DCCD also inactivateci Glc-6-Pase in intact microsomes. The
concentration requued for half-maximai inactivation of Glc-6-Pase in intact
microsomes was about 0.4 mM (not shown), a concentration similar to that required
for haif-maximal inhibition of Glc-6-Pase by DCCD in the hydroxylapatite fi-action (0.2
mM, Fig. 1). However, the water-soluble carbodiimide, 1-ethyl-3-[-3-
diiethylaminopropyl] carbodümide, at concentrations up to 3 mM, did not lead to Gic-
6-Pase inactivation either in intact rnicrosomes or in rnicrosomes solubilized with
CHAPSO (not shown). Presumably, the water-soluble carbodiùnide is unable to access
the hydrophobic active site of Glc-6-Pase to modfy carboxyl groups.
Semi-logarithmic plots of the percentage of residuai activity of Glc-6-Pase in

the hydroqdapatite fhction versus t h e for dierent DCCD concentrations were linear
(Fig. 2). Pseudo nrst order rate constants &) were caidated f?om dopes of the
negative logarithrn of residual activity versus the . The reciprocals of k, were plotted
against the reciprocds of DCCD concentration (Fig. 2, Uwt). The secondary plot was
not hear, suggesthg that DCCD modifies more than one carboql goup. Therefore,
we were unable to calculate a Ki and first order rate constant for chernical modification
fTom such a plot. Kowever, the data are consistent with the revenible fornation of an
enzyme-inhibitor cornplex p60r to chernical modification.
3.3. Protection by vananous agenb agaizst DCCD-induced Glc-6Pme inactivation
The inactivation of Glc-6-Pase in the hydroxylapatite fiaction was almost
completely protected by Glc-6-P (Fig. 1). Therefore, it was of interest to see if other
substrates and products of the Glc-6-Pase reaction had sidar effects. When extracted
from the microsomal membrane, Glc-6-Pase loses its specificity and will also hydrolyse
mannose 6- phosphate, inorganic pyrop hosphate and carbarnyl phosphate. Indeed, these
substrates were as e E i v e as GIc-6-P in protecting against the DCCD-induced
inactivation of Glc-6-Pase (table 2). The reaction product, glucose, or the glucose
analogue, glucosamine, offered no protection against Glc-6-Fase inactivation, whereas
the reaction product, inorganic phosphate, offered complete protection (table 2).
Although sn-glycerol 3-phosphate offered little protection agakt Glc-6-Pase
inactivation, other phosphoesters, narnely glucosamine 6-phosphate, glucose- 1 - hosphate and ATP, partially protected against inactivation.

80
3.4. LubeIIing of proteim in the hpkoxyIiputite Puction &y ["'cJDCCD and
DC~~/[-'*~lglycine ethyl ester.
SDS-PAGE and Coomassie Blue staining of the hydroxylapatite hction
revealed the presence of many bands. In order to try to identify the catalytic component
of Glc-6-Pase, the hydroxylapatite hction was incubated with [ 1 4 ~ ] ~ ~ ~ ~ . This led to
the labeling of 4-5 bands with MJ between 36000 and 77000 (Fig. 3). However, the
labeling pattern was unaffied by Glc&P, suggesting that none of the labelled bands
represents the catalytic subunit of Glc-6-Pase. Presumably, the DCCD-labelled adduct
of Glc-6-Pase is unstable, or an intra-molecular cross-luik is formed in the reaction.
Therefore, to try and trap the modified carboxyl group and label the Glc-6-Pase
protein, the reaction was carrieci out with non-radioactive DCCD and [I-14c] glycine
ethyl ester. Under these conditions, a band with a Mr of 68000 was labellecl (fig. 3).
Moreover, the Iabelling of the 68000-M, band was decreased by Glc-6-P, mannose 6-
phosphate and carbarnyl phosphate, but not by sn-glycerol 3-phosphate or ATP, in
accordance with the protection data (table 2). The fact that there was no protection of
labelling by ATP indicates that the 68000-M, band is not an ATPase, which could have
been modined by DCCD [28]. There was no labehg of the hydroxylapatite hction
with [l-14c] glycine ethyl ester in the absence of DCCD. These findings suggest that
the catalytic component of the Glc-6-Pase system might reside on a 68000-Mr protein.
We exclude the possibility that the 68000-U band corresponds to
phosphoglucomutase [25], since the hydroxylapatite hction was devoid of
phosphoglucomutase activity and because cornmerciai preparations of
phosphoglucomutase were not labelled or inactivated by DCCD/glycine ethyl ester
under conditions which led to Glc-6-Pase inactivation. The labeiiing of the 68000-h&

8 1
band is surprisiig in view of the fàct that the rat Gic-6-Pase sequence predicts a
molecuiar mass of about 40000. The aberrant Mgration in SDS-gels rnight be due to
covalent modication, such as glycosylation [ 1 O, 121, or to an unusual conformation of
the protein Glc-6-Pase catalyses the hydrolysis of Glo6-P via a classical ping-pong
mechanism and incubation of microsomes with [ 3 2 ~ ] ~ l c - 6 - ~ labelleci a band with a A&
of 36500 [29]. Also, the phosphorylated amino acid was identifid as 3-
phosphohistidine [30]. However, this labeling in crude rnicrosomes could be due to
contaminating non-spdc phosphatase, eg. acid phosphatase. Unfortunately, the
relative rnolecular mass of Gic-6-Pase expressed in microsomes of COS ceUs by SDS-
PAGE has not been determineci and labelling studies with [ 3 2 ~ ] ~ ~ c - 6 - ~ in such
preparations do not appear to have been performed.
4. Conclusions
In this paper, we have used chernical modication with the Asp/Glu-specific
reagent, DCCD, to show that carboxyl groups are important for catalysis in GIc-6-
Pase. Previous studies showed that Glcd-Pase is inhibited by the histidine-specific
reagent diethylpyrocarbonate [3 11, and is phosphorylated f?om [ 3 2 ~ ] ~ l c - 6 - ~ on a His
residue [29]. Furthemore, site-directed mutagenesis of His-119 in human Glc-6-Pase
suggests that this residue acts as the phosphoacceptor [3 21. Glc-6-Pase is not related to
histidine phosphatases, because the active site His residue is not in the Ar& His Gly
(RHG) motif characteristic of this f d y of enzymes. However, the catalytic
mechanism is propably similar. The mechanism of the histidine phosphatases involves a
His-Asp/Glu-His catalytic tnad. The phosphory lenzyme intermediate is formed on one

82
of the fis residues, while the second His residue interacts with the transferred
phosphate group. The Asp/Glu residue acts as an acid catalyst in protonating the
leaving group or as a base caîaiyst in phosphoqdenzyme breakdown. By analogy, the
carboxyl group modifieci by DCCD in Glc-6-Pase could act as an acid or base catalyst.
Alternatively, it couid be involved in substrate binding.
This work was supported by the Belgian State Prime MUu'stei's Office Science
Policy Incentive Program in Life Sciences 99/93-122 (Grant 20), by gants from the
Belgian Fund for Medicai Scientific Research and by Grant MT-10804 (to G. v.d.W.)
nom the Medicd Research Council of Canada M.H.R is Chercheur Qualitié of the
Fonds National de la Recherche Scientifique (Belgium).

References
Nordlie, RC. (1971) The Enzymes, 3rd Ed., 4,543-561
Arion, W.J., Lange, AJ., Wds, H.E. & Ballas, L. (1980) J. Biol. Chem. 255, 1039640406
Burchell, A (1990) FASEB J. 4,2978-2988
Arion, W.J., W a h , B.K., Lange, AJ. & Bailas, L.M. (1975) Mol. CeU. Biochem. 6,7583
Berteloot, A, Vidal, H. & van de Werve, G. (1991) 1. Biol. Chem. 266, 5497-5507
Vidai, H., Berteioot, A, Lanie, M.-J., St-Denis, J.-F. & van de Werve, G. (1992) FEBS Lm. 302, 197-200
Schulze, H.-U., Nolte, B. & Kannier, R (1986) J. Biol. Chem. 261, 16571- 16578
Schulze, Fi.-U. & Speth, M. (1980) Eur. J. Biochem. 106,505-5 14
Ness, G. C., Sukdski, K. A, Sample, C. E., Pendleton, L. C., McCreery, M. J. & Nordlie, R C. (1989) J. Biol. Chem. 264,71 11-71 14
Sheliy, L.L., Lei, KA., Pan, C.-J., Sakata, S.F., Ruppert, S., Schutz, G. & Chou, I.Y. (1993) J. Biol. Chem. 268,21482-21485
Lange, AJ., Argaud, D., El-Maghrabi, M.R, Pan, W., Maitrq S.R & Pilkis, S. J. (1 994) Biochem. Biophys. Res. Commun. 20 1,302-309
Lei, K.-J., SheUy, L.L., Pan, C.-J., Sidbury, J.B. & Chou, J.Y. (1993) Science 262, 580-583
Lei, K.-J., Sheliy, L.L., Lin, B., Sidbuqr, J.B., Cen, Y.-T., Nordlie, RC. & Chou, J.Y. (1995) J. C h . Investigation 95,234-220
Liu, Z., Barrett, E.J., Daikin, AC., Zwart, AD. & Chou, J.Y. (1994) Biochem. Biophys. Res. Commun. 205,680-686
Reczek, P. R & Viee, C. A (1982) Biochem. Biophys. Res. Commun. 107, 1158-1 165
Speth, M. & Schuize, H.-U. (1986) W S Lett. 202,32-36

Speth, M. & Schuize, K-U. (1992) Eur. J. Biochem. 208,643-650
Pilkis, S.J., El-Maghrabi, M.R & Claus, T.H. (199 1) Diabetes Care 13, 582-599
Lin, K., Li L., Correia, J.J. & Pilkis, S.I. (1992) J. Biol. Chem. 267,6556- 6562
Ostaniq K. & van Etten, R (1993) J. Biol. Chem. 268,20778-20874
Poniari, K.S., Herraia, AM., Kurkela, RM., Taavitsainen, P.A, Lindqvist, Y., Schneider, G. & V i o , P.T. (1994) 1. Biol. Chem. 269,22642-22646
van de Werve, G. (1989) J. Bol. Chem. 264,6033-6036
Lowry, O.H., Rosenbrough, N. J., Farr, AL. & Randdi, J. (1 95 1) J. BioI. Chem. 193,265-275
Laemmli, U.K. (1970) Nature 227,680-685
Mithieux, G., Ajzannay, A & Minassian, C. (1995) 1. Biochem. 1 17, 908-914
Toner-Webb, J. & Taylor, S.S. (1987) Biochemistry 26,7371-7378
Buechier, J.A & Taylor, S.S. (1 988) Biochemistry 27,7356-736 1
Esch, F.S., Bohlen, P., Otsuka, AS., Yoshida, M. & Allison, W.S. (1981) 1. Biol. Chem. 256,9084-9089
Countaway, J. L., Waddeii, 1 D., Burcheii, k & Arion, W. J. (1988) J. Biol. Chem. 263,2673-2678
Feldman. F. & Buttler, L.G. (1 972) Biochem. Biophys. Acta 268,698-7 10
Arion, W.J., Burcheil, B. & Burcheli, A (1984) Biochem. J. 220,83 5-842
Lei, K.-J., Pan, C.-J., Liu, J.-L., Shelly, L.L. & Chou, J.Y. (1995) J. Biol. Chem. 270, 1 1882- 1 1886


Table 2 Protection bv various agents aeainst DCCD-induced Gic4Pase
inactivation.
The hydroitylapatite hction was inaibated with 0.5 mM DCCD, lOmM
glycine ethyl ester and 10 mM of each protectant as descriid in the Methods d o n .
The protectants were added prior to s t h g the reaction by the addition of the
hydroxylapatite fraction. The control incubation without DCCD contained 1% (vh)
DMSO. M e r 15min at 30°C, aliquots were removed for Glc-6-Pase assay. The results
are the means S.EM of three separate experiments. 100% of Gic-6-Pase activity
was 1.43 0.15 Unitdmg of protein. * Indicates a significant Werence (P < 0.05
paired Student's i test) compared with the control (no additions).
Additions
None
Glc-GPase activity (%)
100
DCCD 28*3 *
DCCD + G1~6-P 99k I l
DCCD + mannose 6-phosphate 101 =t 11
DCCD + glucosamine 6-phosphate 68&8*
DCCD + glucosamine 4%6*
DCCD + glucose 32*4*
DCCD + glucose- 1 -phosphate 73&9*
DCCD + sodium pyrophosphate 91&5
DCCD + potassium phosphate 101 =t 15
DCCD + cafbamyl phosphate 99*4
DCCD + sn-glycerol3-phosphate
DCCD + ATP

Figure legends
Fig.1 Dose-dependence of the inactivation of Glc-6-Pase by DCCD. Hydroxylapatite h a i o n (1Omg) was incubated in a finai vohne of 50 ml as described in the Methods section with the indicated concentrations of DCCD and 10 rnM glycine ethyl ester in the presence (e) or the absence (O) of 10 mM Glc-6-P. Mer 15 min of incubation at 30°C, sarnples were removed for Glc-6- Pase assay, 1000/o of Gic-6-Pase activity corresponds to 1 -5 1 * 0.23 Unitdmg of protein. The results are the mean * S.E.M. of three separate experirnents.
Fig.2 Tirnecourse of the inactivation of Gic-6-Pase by WCD. Hydroxylapatite hction (IOrng) was incubated in a ha1 volume of 50 ml as describecl in the Methods section with the indicated concentrations of DCCD and 10 mM giycine ethyl ester. At various times, samples were removed for Glc-6-Pase assay; 1 00% of Glc-6-Pase activity corresponds to 1 -3 1 k 0.1 2 Unitdmg of protein. Rate constants (K,) were calculated fiom plots of the negative natural logarithrn of residual activity versus the . A secondary plot of the reciprocal of Kw against the reciprocal of the concentration of DCCD is shown in the inset.
Fig.3 Labeliing of the hydroxylapatite fraction by [ " ~ D C C D and DCCD/[l- 14 q glycine ethyl ester. The hydroxylapatite hction (40mg) was incubated at 30°C with or non-radioactive DCCD and [1-I4c] glycine ethyl ester in the presence or absence of 10 rnM of each protectant in a final volume of 60 ml as described in the Methods section. Mer 15 min, the samples were precipitated with 10% (w/v) trichloroacetic acid for SDS-PAGE foliowed by Coomassie Blue staining, gel dqing and autoradiography. Lanes 1-3; hydroxylapatite fiaction incubated with [ 1 4 ~ ] ~ ~ ~ ~ (1) and [ 1 4 ~ ] ~ ~ ~ ~ + Glc- 6-P (2) or ATP (3). Lanes 4-9; hydroxylapatite fkiction incubated with non- radioactive DCCD and [1 -14~] glycine ethyl ester (4) and DCCD/[I-'~C] giycine ethyl ester + Glc-6-P (S), sn-glycerol 3-phosphate (6), mannose 6- phosphate (7), carbarnyl phosphate (8) or ATP (9).
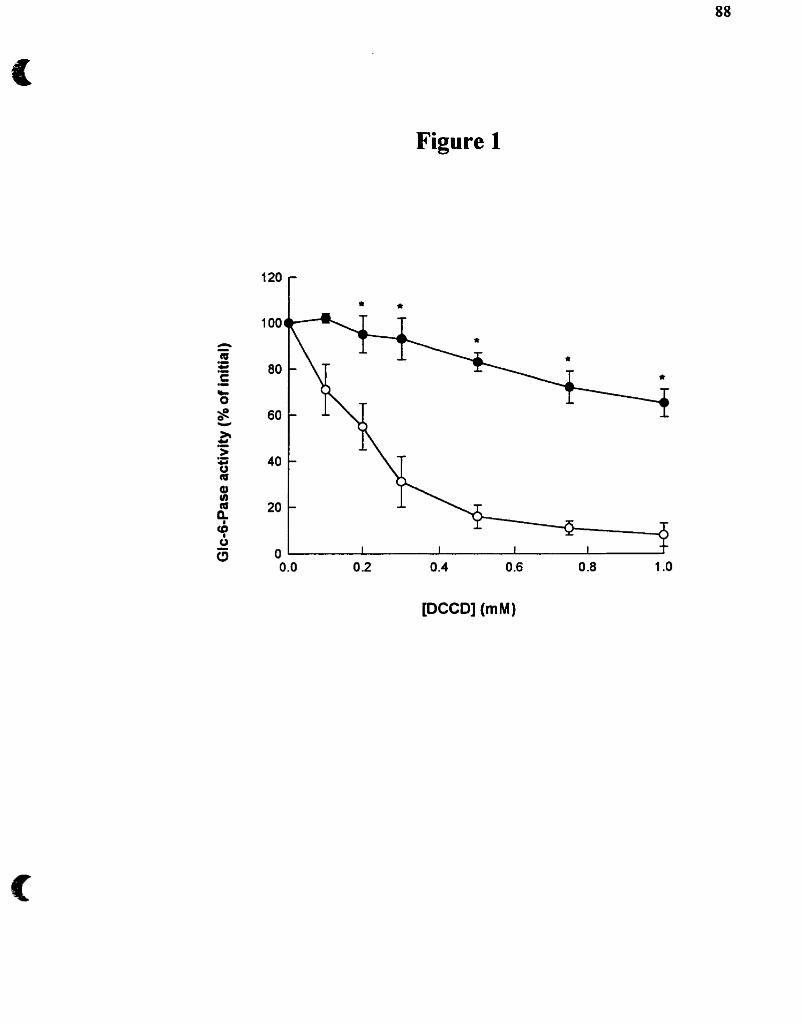
Figure 1
0 .O 0 -2 0.4 0.6 0.8 1 .O
[DCCD] (mM)
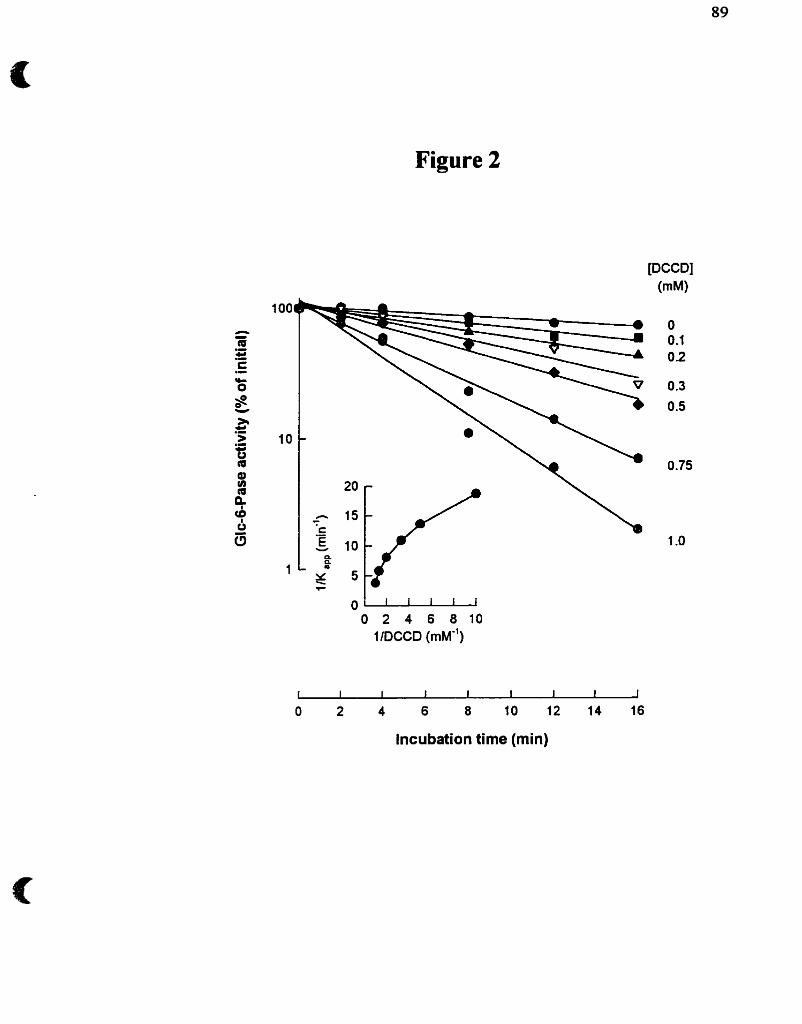
Figure 2
[DCCD]
0 2 4 6 8 1 0 1 IDCCD ( m ~ " )
O 2 4 6 8 10 12 14 16
Incubation time (min)
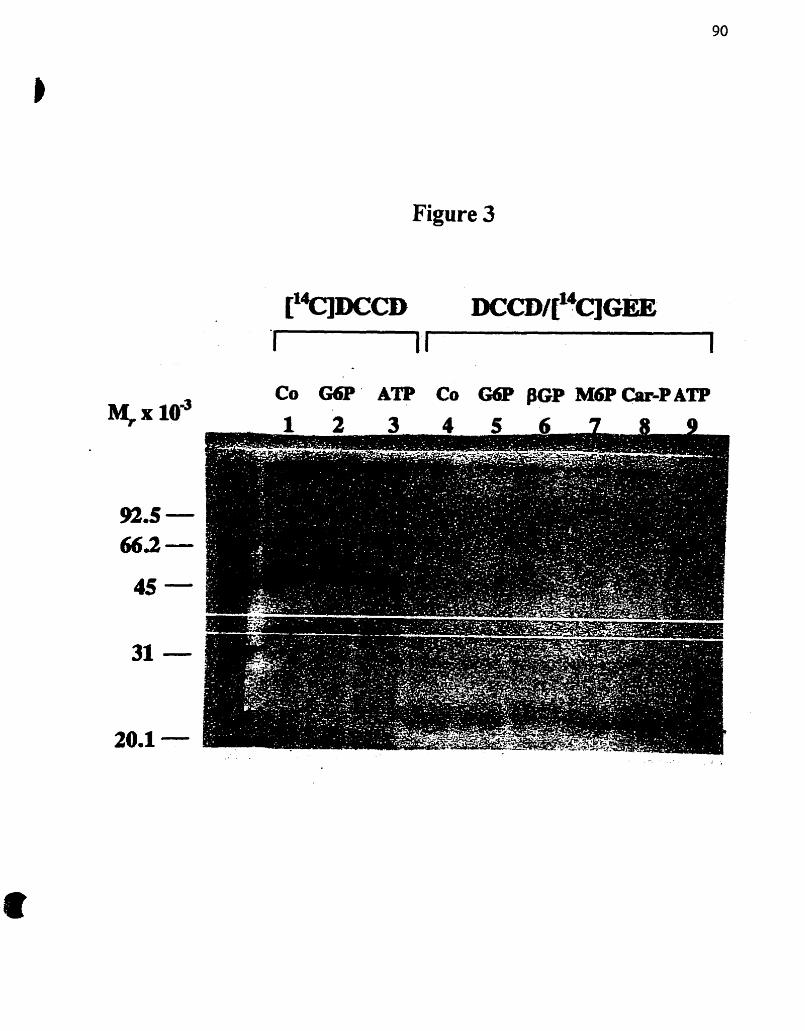
Figure 3
- CO G6P ATP CO G6P BGP M6P Car-PATP

3.4- Effet des glucocorticoides sur I'activité gIucose6-phosphatase :
Evidences contre t'existence de Tl
3 -4.1 - Article 4 : Evidence that the transit of glucose into liver microsornes is not
required for fiinctionnal glucose-6-phosphatase. Annabi B. &
van de Werve G.

Article 4
Hypothèse : L'augmentation de l'hydrolyse du Gic-6-P par la Gloo-Pase dans les
microsomes intacts isolés de foie des rats traités à la tnamcinolone n'est
pas dûe à une activité accrue du transporteur du Glc-6-P UI) tel que
proposé par le madèle de fiaruport a( s~bsfrat.
Une plus grande V,, (Burchell et Cain, 1985) et quantité d'ARNm (Lange el
al., 1994) de la Glc-6-Pase sont observés dans des microsomes perméabilisés de foie de
rats diabétiques et sont expliqués par une augmentation de la sous-unité catalytique.
L'administration de l'analogue des glucocortiwides triamcinolone augmente par contre
la V, de la Glc-6-Pase dans les microsomes intacts. Cette propriété a été attniuée à
une augmentation de la capacité de transport du Glc-6-P vers l'intérieur des
microsomes (Nordlie, 1971). Nous avons donc étudié cette fonction d'accumulation du
Glc a l'intérieur des microsomes et sa relation avec la présence d'un transporteur putatif
de Glc-6-P.

EVIDENCE TaAT TEE TRANSIT OF GLUCOSE INTO LIVER
MICROSOMES IS NOT REQUIRED FOR FUNCTLONAL GLUCOSE-&
PHOSPEATASE
by
Borhane Annabi1 and Gérald van de wervel*''
Departments of '~iochernistr~ and 2~utntion, Laboratoire d'Endocnno1ogie Métabolique, Université de Montréal, Montréal, Québec, H3C 3 J7, Canada
* Address for corres~ondence :
Subdivision : Ewology
Department of Nutrition, Université de Montréal, C.P. 6 1 28 Succursale centre-ville, Montréai, Québec H3C 3 57, Canada Tel : (5 14) 343-7546 Fax : (5 14) 343-6103 E-mail : [email protected]. M
Kewords : Maosornes, glucosed-phosphatase, glucocorticoid, pyridoxal5'- phosphate

CHAPSO, -[(3-Cholamidopropy1~unethylammonio]-2-hyo-I-propanesuffo-
nate); endoplasmic reticulum; Glc-6-Pase, glucose-6-phosphatase (EC 3.1.3.9);
Glc4-P, glucose 6-phosphate; Man-6-Pase, mannosed-phosphatase; Man-6-P,
mannose 6-phosp hate; PLP, pyridoxal 5'-phosphate; SDS-PAGE, sodium
dodecylsulphate polyanylarnide gel electrophoresis; Tl, Liver microsorna1 glucose 6-
phosphate translocase

We previously found, when cornparhg glucose-6-phosphatase (Glc-6-Pase)
activity and intramicrosomal glucose (Gic) production, that most of the Gic dex-ived
from Gfc-6-P hydrolysis is released directly in the extramiaosomal medium and only a
small portion transits through the lumen of the microsomes [Berteloot, A., St-Denis, J.-
F. and van de Werve, G. (1 995) J BioL C h . 270, 2 1098-2 1 1021. In this study, we
present evidence that the production of Glc Eom Glc-6-P hydrolysis at the outside of
the microsomes is a function of Gk-6-Pase that is independent of its property to form
Glc inside the microsomes. Indeed, d u ~ g development (before 1 day of age), mouse
liver microsomes possessed solely the property to produce Glc from Glc-6-P hydrolysis
outside the rnicrosomes without the formation of Glc inside the vesicles. Furthemore,
in vivo treatment of rats with the glucocorticoid analogue triarncinolone resulted in a
twofold increase in Glc-6-Pase activity outside but not inside microsomes. This
enhanced activity was independent Eom an increase in the catalytic subunit 36 kDa
Glc-6-Pase mRNA abundance or protein level, indicating that other h o r s induced by
triamcinolone (e.g. dtered membrane tipid environment andl or a regdatory protein)
were responsible for the activity change. TriamcinoIone treatment also Iessened the
inhibition of liver rnicrosomal Gic-6-Pase by pyridoxal S'-phosphate (PLP), but this
effect was not due to an interaction of PLP with the active site of Glc-6-Pase.
Accordingly, reversai of the inhibition was observeci after permeabilization of the
microsomes. Our results offer a rationale to previous fïndings that the latency of Glc-6-
Pase was markedly reduced by glucocorticoid treatment. The two distinct orientations

of liver microsoma1 Glc-6-P phosphohydrolase suggest dEerent physiological roles
played by this enzyme in the endoplasmic reticulum membrane.
INTRODUCTION
Microsoma1 glucose-6-phosphatase (Glc-6-Pase) CD-glucose-6-P
phosphohydrolase; EC 3.1 -3 -91 catdyzes the terminal step of glycogenolysis and
gluconeogenesis by hepatocytes and rend cortex, and is located at the endoplasmic
reticulum (ER) membrane [Il. The membrane topology of Glc-6-Pase in the ER is
however stili undefined. Most authon agree that the enzyme is an integral protein
tightly associated with the hydrophobie part of the membrane, as confirmeci recentiy by
the amino acid sequence inferreci frorn the isolated cDNAs encoding human 121, mouse
[3] and rat [4] Glc-6-Pase of approximately M, 36 kDa.
We reported before that the latency and speci£icity of this enzyme in intact
microsomes are properties that can be explained by a change in the conformation of
Glc-6-Pase [5,6]. Furthemore, we showed that changes in kinetic properties (loss of
latency and specificity) identical to those induced by permeabilization of rnicrosomes
can be induced by histones type II-4 but without permeabilkation of the vesicles [7].
These properties and the demonstration that Glc alone is not taken up by microsomes
[8,9] as weU as the observation that the Glc derived fiom Glc-6-P hydrolysis is fonned
both directly in the extramcrosomal medium and into the lumen of the rnicrosomes
[IO], led us to constmct a new model of Glc-6-Pase [IO], derived f?om the combined
flexibility-substrate transport model proposed by the group of Schul-e [11,12].

In support of this modei, in a case of glycogenosis type 1 a (Glc-6-Pase deficiency) [13]
and in 36 kDa Glc-6-Pase gene knockout mice [14 and unpublished experiments from
this laboratory], no uptake of radioacbvity was found into microsomes exposed to ILI-
"c]G~c-~-P. The recent demonstration that the gene encoding the Glc-6-Pase 36 kDa
protein is normal in glycogen storage disease type lb and lc patients [15], indicates
however that a mutation of (an)other protein(s) is responsiile for the fùnctiond loss of
Glc-6-Pase activity in vivo in these diseases.
The administration of glucocorticoids in vivo [16-191 was proposed to affect
the glucose-6-phosphatase system either through activation of a putative Glc-6-P
transporter or through alterations in the amount and kinetic properties of the enzyme's
catalytic unit [20]. Since there is much evidence against such a transporter (see above),
we reinvestigated the e f f i of tiarncinolone. We found that in Iiver rnicrosomes
isolated £?om rats treated with this glucocorticoid analogue, the increase in Glc-6-Pase
activity could entirely be accounted for by a faster rate of ~ - ' % ] ~ l c production fiom
[u-'~c]GIc-~-P hydrolysis without increased formation of [U-'~C]G~C inside the
rnicrosomal vesicles and that this property was not related to an increase in 36 kDa
Glc-6-Pase protein. Inhibition kinetics of Glc-6-Pase by pyridoxal 5'-phosphate (PLP)
and NaB& suggests that another protein, induced by triamcinolone, is responsible for
the activation of Glc-6-Pase. The dinished latency of hepatic microsoma1 Glc-6-Pase
activity, docurnented before in tiarncinolone-treated rats [21], c m therefore be
int erpreted as a specifk increase in [u-'~c]G~c production outside the rnicrosomes.
Furthemore, we also show that W-~~C]GIC-~-P hydrolysis can independently take
place in a system where ~ - 1 4 ~ ] ~ l c accumulation into microsomes is absent (1 day-old

mouse liver microsomes), thus where V ' ~ C ] G ~ C production occurred exclusively
outside the microsornes Taken together these results demonstrate that the peailiar
location of Gic-6-Pase in the rnicrosomal membrane ailows this enzyme to produce Glc
denved from Glc-6-P d i d y outside the vesicles, independently f?om a transit of Glc
through the microsoma1 lumen.

3-[(3-Cholamidopropyl)-dimethylamm0nio]-2-hyo- 1 -propaneSulfoonate
(CHAPSO), Gic-6-P, Mannose-6-phosphate, pyridoxal S'-phosphate (PLP), sodium
borohyciride (NaBK) and streptozotoch were purchased fiom Sigma Chernical Co.
(St-Louis, MO). [u-'~c]G~c-~-P was f?om ICN Biomedicals (Montreai., Canada). CU- 14 ClMannose was obtauied f?om NEN-DuPont (Mississauga, Canada). CU- 14c]~an-6-
P was prepared by the phosphorylation of annose nos no se (300 mCi/ mrnol) with
ATP by using hexokinase as previously descnbed [7] Tnamcinolone and placebo
pellets were fiom hovative Research of Arnerica (Toledo, OH). Ail reagents were of
analyticai grade.
Animais
Male W~star rats, averaging 200-250 g body weight, were purchased fkom
Charles River (St-Constant, Canada). The animals were housed under a 12 h dark : 12
h tight cycle and were given standard laboratory chow and water, Qd libitum. Fasted
rats had food, but not water, withheld for 24 h prior to use. Rats were made diabetic
with a single inhaperitoneal injection of sûeptozotocin (100 mgkg body weight)
fieshly dissolved in 4 m M sodium citrate, pH 4.5 [22]. The diabetic state was assessed
by the presence of hypergiycemia (>300 mg/100 mi) afler a 24 h fasting period, and

loss of weight gain 8 days aller treatment. Rats were used on day 9 der streptozotocin
administration. Glucocurticoid-treated rats were anesthetid with ether and had a
pellet of triamcinoIone (50 mgpellet) or the rnatching placebo peUet implanteci
subcutaneously on their back with a relative rate of release of 50 mg / 3 weeks for 6
days and were used on day 7 d e r pellet insertion. Administratecf doses of
triamcinolone were quivalent to those used by Arion and Nordlie [17] and Traxinger
and Nordiie [20].
Preparation of liver microsomes
Livers f?om overnight-fasted control, triamcinolone- or streptozotocin-treatd
rats were rapidly excised and hornogenized as describeci previously [23] in 4 volumes
(v/w) of a bae r contaking 250 rnM sucrose, 1 mM EDTq 1 mM EGTA, 2 mM
dithiothreitol, 2 mM Berilamidine-HCl 0.2 mM phenylmethanesulfonyl fluoride, 0.5
pg/mL leupeptin, and 10 mM HEPES pH 7.0 at 4OC (bufer A). Maosornes were
prepared from the homogenates by dserential centrihgation and h d y resuspended in
a buffer containing 1 rnM =POI. 1 mM EDTA, 0.5 rnM EGTA, 1 mM
dithiot hreitol, 2 mM Benzamidine-HCl, 0.1 mM phenyhnethanesulfonyl fluoride, 1
pg/rnL leupeptin, 20% (v/v) Glycerol, and 20 mM HEPES pH 7.0 at 4°C (buffer B), at
a protein concentration of 20 mg/ml. Microsoma1 vesicles were either used the same
day or kept at -80°C for later use. Freezing of microsomes, ifthawed once at 4OC, did
not affkct the kinetic propertiw of Glc-6-Pase [13, 241. Suckling C57BU6 rnice were
provideci by Dr Janice Chou, subsequent liver microsomal isolation was performed by
Dr Jean-François St-Denis and resulted in a comparable yield as obtained before [13].

Treatment of microsomes with pyrîdoxai S'-phosphate and sodium borohydride
Microsornes (-1 5 mg of protein) were suspended in 1.5 ml of 50 rnM HEPES,
pH 7.0 and appropriate concentrations of fieshly prepared pyridoxal S'-phosphate. The
reaction mixture was incubated at 30°C and the reduaion was performed by the
addition of 3 mi of 100 rnM NaBh in ice-cold double-distiiied water [25, 261 and left
on ice for 15 min. The sarnple was then centrifugai at 105,000 g for 30 min at 4°C to
separate the PLP/NaB&-treated microsomes fiom unbound ragent. Microsomes
were resuspended in buffer B at a protein concentration of 5 mghl and were used as
starting matenal for subsequent treatment. Because NaBb alone does not significantly
inhibit Glc-6-Pase (not shown), the possibility that substantial amounts of intraprotein
SchitPs bases are present in the membrane can be ruled out.
Enzyme assays and uptake measurements
Glc-6-Pase and mannose-6-phosphatase (Man-6-Pase) activities were
measured at 30°C as described previously [7] in untreated and CHAPSO-trated
(0.8%, for 30 min at 4°C) microsomes. W-'~C]G~C uptake in rnicrosomes ffom 0.2
mM [u-'~c]GIc-~-P (20 nCi/nrnol) was determined essentially as described [5], except
that a single weli filtration apparatus was used to stop the reaction afler a 2 min
incubation at 30°C, a tirne at which a steady-state of accumulation was reached [5,9].

~Glycerophosphate (20 to 100 m.) was used to assay non-spdc phosphatase
activity, and [%Ilmannitol was used to correct for non-specific binding to cellulose
nitrate filters in uptake measurements [5] .
Immunoblot and Northern Blot anaiysis
SDS-PAGE was performed on the various preparations descnbed in the legend
to Fig. 1. Proteins were electroblotted ont0 Immobilon polyvinylidine diiuonde
membranes and detected with a rabbit polyclonal anti-Glc-6-Pase antibody with a
1:1000 dilution (provided by Dr Janice Chou, NM). Bound antibodies were
recognized by a peroxidase-antiperoxidase complex (PAP f?om Prosan) at 15000
dilution and revealed by the ECL Western blotting system (Amersham) or by ushg
alkaline phosphatase (AP)-linked goat anti-rabbit IgG (Amersham). Total RNA was
isolated fi-om quickly fiozen liver tissues and assessed for punty by the 260/280 ratio
absorbante. Twenty mg of total RNA were subrnitted to electrophoresis, transferred,
and hybndized with a 1.25-kilobase pair EcoRI-HindIII hgrnent of the Glc-6-Pase
cDNA as described by Massillon et al. [27].
Other rnethods
Protein concentration was rneasured by the method of Lowry et al. [28] using
bovine semm albumin as a standard. SDS-PAGE analysis was canied out in 0,8 mm
mini slab gels (10 x 8 cm).

ER't of streptozotocin-induc diabetes and triamcinolone treatment on kinetic
parameters and expression of glucost6-phosphatase
Table 1 confirms previous data that Glc-6-Pase activity (Vmax) assayed in
intact tiver microsornes isolateci korn triamcinoIone-treated rats is already fiily active
and is not fbrther activated by detergent treatment and that Km values are unchangeci
[lq. This contrasts with diabetic rat liver microsomes, where Vmax was increased by
about fourfold and Km was reduced by haif in detergent-treated rnicrosomes 1291,
consistent with the reported increase in rnRNA abundance [4] and protein level of 36
kDa GIc-6-Pase in diabetes [30].
The immunoblot represented in Fig. 1 shows that whereas streptozotocin-
induced diabetes indeed increased liver microsomal 36 kDa Glc-6-Pase protein,
triamcinolone-treatment was without effect on this parameter despite the twofold
increase in Glc-6-Pase activity (Table 1). Accordingiy, the 36 kDa Glc-6-Pase rnRNA
abundance meanired by Northern blotting was not increased in triamcinolone-treated
rat liver (not show).
Intramicrosomd ~ ' ' ' ç j ~ l u c o s e uptake and total w - ' " ~ j ~ l u c o s e production
In our mode1 of Glc-6-Pase (Fig. 2, adapted from [IO]), Glc-6-P uptake and
hydrolysis are tightly linked [9]. Glc-6-Pase traverses the microsomal membrane as a

channel-forming protein within the hydrophobie matrk of the bilayer, where the
cataiytic site is proposed to be located within a hydrophilic pocket accessible to the
substrate fiom the cytoplasrnic swfiice of the membrane. An outer channel controls
both Glcd-P uptake and unidirectional efflux of Glc formed f?om Glc-6-P, hydrolysis
at steady state rate Vh. An inner channel dows Glh to be transported into the lumen
of the microsorne, where at equilibrium, a plateau Glci is reached.
The majority of the previous studies have examined the o v e d hydrolysis of
Glc-6-P to Glc, but without discrimination of Glc produced inside (Glq) or outside
(GlcJ microsomes. When total [u-~~c]G~c production (Glc, + Glci + Gl%) fiom [U-
1 4 ~ ] ~ l c - 6 - ~ hydrolysis was compared to W-'~C]G~C fomed inside the rnicrosomes
(Glq) (Table 2), the higher steady-state level of [U-'~C]GIC accumulation (Giq)
reached in rnicrosomes isolated f?om streptozotocin-treated rats p d e l e d the total
Glc-6-Pase activity (Glc, + Glci + Glc,,). In contra we found no significant increase
in the plateau of W-'~C]GIC accumulation (Glc) into microsomes isolated fiom
triamcinolone-treated rats, despite an increase in the total Glc-6-Pase activity.
Tnamcinolone treatment therefore affects specifidy the outer channel in Fig. 2.
This latter observation was of particular interest, siice we discovered that CU-
14 C ] Glc fomed fiom ~- '*c]G~c-~-P hydrolysis in microsomes isolated fiom livers of 1
day-old rnice (but not older animds) was not produced inside (Glî not detectable) but
exclusively outside the rnicrosomes (Table 3). This means that [U-'~C]G~C-~-P
hydrolysis is possible by the sole operation of the outer chwe1.

Effect of pyridoxai S'-phosphate and sodium borohydride treatment on total
' 4q~ lucose production
In order to m e r understand these hivo fùnctions of Glc-6-Pase, we used
pyridoxai S'-phosphate (PLP), a linear noncornpetitive inhibitor of Glc-6-Pase in intact
microsomes [25]. PLP specifically modifies lysine residues by reacthg with their side-
chah e-amino group to generate the revenible formation of a Schiff's base (hine). It
has also been show to react with Iysiie residues at the phosphate binding sites of
several enzymes [3 11. The hrther reduction of the imine by NaBK to a stable
secondary amine stabilizes the intemediate by covdently Linking the PLP to the
modified residues, which can then be identified by classical Edman sequencing of
peptides. Since Glc-6-Pase inhibition by PLP was ody shown in intact microsomes
[25,3 1,321, it was assumed that PLP modified essential lysine residues implicated in the
transport mechanism of the abstrate Gic-6-P [3 1,321.
Fig. 3 shows a dosedependent inhibition of Glc-6-Pase activity in microsomes
treated with PLP and NaBh. The PLP concentration required for half-maximal
inactivation of Glc-6-Pase was 0.52M.07 mM for control and 2.4150.4 m . for
microsomes isolated &om triamcinolone-treated rats. With an optimal dose (100 mM),
the tm for PLP inhibition was 1.18 0.13 min and 3.85 * 0.14 min in control and
triamcinolone-treated rats and complete inhibition was reached d e r 4 and 16 min
respectively (Fig. 4). Microsornes isolated from streptozotoch-treated rats showed
essentiaüy the sarne inhibition of Glc-6-Pase by PLPMaB& as in vesicles isolated fkom
control rats (not shown). Dinerent kinetics of inhibition are thus observed between

controi- and triamcinolone-treated rats, even though total Glc-6-Pase activity revealed
by detergent permeabilization (Table 1) was the sarne.
Gic-6-P was reported to prevent Gic-6-Pase inhibition caused by PLP [3 1, 321.
We show however, by covalently linking PLP to the modified residues with N a B a
that neither Glc-6-P, Man-6-P, pyrophosphate, carbamyl-P and glucose (ail ligands to
the active site of Glc-6-Pase), nor b-giycerophosphate prevent Glc-6-Pase inhibition in
microsomes isolated fiom control rats (Fig. 5). Moreover, subsequent permeabilization
of the PLP/NaB&-treated microsomes (in the presence or absence of ligands) with the
detergent CHAPSO restores a fÙUy active Glc-6-Pase (Fig. 5). These results confirm
that the SchitPs base is formed at a site distinct from the site of Glc-6-P hydrolysis, as
suggested by Gold and Widnel 1251. Omission of the last reaction step involving the
reduction of the SchitPs base by NaBK probably explains the Glc-6-P protection
against PLP inhibition of Glc-6-Pase reported previously [3 1,321. Treatment of the
microsomes with NaBK aione foiiowed by removal of the unreacted ragent, as
described in the expenrnental procedures, did not affect Glc-6-P hydrolysis (not
shown).

DISCUSSION
Mechanisms invohred in triamcinolone stimulation of Glc-6-Pase activity
The e f f i of glucocorticoids on Glc-dPase activation in cultureci Liver ceUs
and in vivo is weli documented. Dexamethasone inmeases 36 kDa Gic-6-Pase mRNA
levels in FA0 hepatoma ceils [4] and Glo6-Pase activiîy was shown to increase with
corticosterone, cortisone, and cortisol [33]. The mechanism involveci in glucocorticoid
action is however not clear. In this study, the mRNA levels and the amount of cataiytic
subunit protein were found to be unchanged after triarncinolone treatment in vivo,
indicating that the change in activity is not directly related to the 36 kDa Glcd-Pase.
Similar observations were very recently made but with adrenalectornized rats treated
with triarncinolone [34]. The administration of glucocorticoids in vivo is thought to
affect the liver microsomal Glc-6-Pase system either through activation of a putative
Glc-6-P transporter protein [21] or through aiterations in the properties of the Glc-6-
Pase catalytic unit [20]. Our observation that the accumulation of W-*~C]G~C nom CU- 14 C]Glc-6-P hydrolysis reached the sarne steady-state level in microsornes isolated from
control or triamcinolone-treated rats, mla out an increase of a putative translocase
activity to explain the enhanced Glc-6-Pase activity. The dinished latency of hepatic
microsomal Glc-6-Pase activity, documented before in tiamcinolone-treated rats [2 11,
c m therefore be explaineci by the sole increase in Glc production from Glc-6-P at the
outside of intact microsomes.

Since the amount of 36 kDa GIc-6-Pase protein is not increased by
triamcinolone (Fig. 1), that PLP inhibition is reversible by detergent treatment (Fig. 5)
and that the half-mawnal inhibition of Glcd-Pase by PLP requires higher
concentrations of PLP (Fig. 3) and has a higher tw of inactivation in microsornes
derived from triamcinolone-treated rats (Fig. 4), these results suggest that
triamcinolone induces a stimulatory protein for Gic-6-Pase. Another explanation would
be that triarncinolone administration modifies the intimate microenvironment of the
Iiver microsomal membrane [35, 361 which could modify the native confornation of
the enzyme, exposing less lysine residues for direct PLP modification at the recognition
site of Glc-6-P. This recognition site could then be locaiized on the outer channel
leading to the hydrophilic pocket embedded in the microsomal membrane (Fg. 2).
Both explanations (stimulatory protein or recognition site) for the effect of PLP are
consistent with the fuidings of Speth and Schulze (1992), who showed that the
exposure of intact microsomes with [WPLP labelled several proteins, including a 36
kDa species with Glc-6-Pase activity.
In the mode1 represented in Fig. 2, the effi of triamcinoIone treatment on a
putative stiiulatory protein could specifically enhance the outer channel activity, i-e.
both the rates of Glc-6-P transport (I&) into the hydrophilic pocket and the efflux of
Glc (Km), without increasing Glc uptake into the vesicles (Km) at the inner channel.

The transit of glucose into l ~ e r microsornes iP not required for functional Glc-6-
Pase
In suppon for the notion that the inner channel (Fig. 2) is not required for Gio
6-Pase activity, we show that ( U - ' 4 ~ ] ~ ~ - 6 - ~ hydrolysis can exclusively take place
outside 1 day-old mouse liver rnicrosomes (mer channel is not operative). The tact
that Gic-6-Pase activity at that age was only haifthat of older mice, suggests however
that the two orientations of Glc production are necessaxy for fui expression of Glc-6-
Pase activity. Glc-6-Pase protein at 1 day of age might still need post-translationai
modifications (or the microsomal membrane be of a diierent lipid composition) to
allow an insertion into the endoplasrnic reticulum membrane that would confer its
property to produce Glc both outside and inside the rnicrosomes.
The molecular topoiogy of Glc-6-Pase w i t h the microsomal membrane we
proposed [IO], (Fig. 2), does not exclude that the cataiytic subunit be associated with
one or several proteins (subunits). The intnguing property of Glc-6-Pase to be able to
produce Glc both outside and into the lumen of the rnicrosomes and the evidence for
one function to operate independently 6om the other suggests that the association of
Glc-6-Pase with the ER membrane allows the enzyme to fùnction either way in vivo.
Whether there are s p d c physiologie roles for the formation of Glc from Glc-6-P
inside and outside the ER remains to be determined.

We thank Dr Janice Chou for providing us with the anti-Glc-6-Pase antiiody and the
36 kDa Gicd-Pase gene knock-out mice, Dr Duna Massillon for the Northem blot
andysis, and Dr Jean-François St-Denis for the preparation of mouse liver m*crosomes.
This work was supportai by Grant MT-10804 fiom the Medid Research Council of
Canada.

1. Nordlie, RC. and SuMski., KA (1985) Multifunctional giucose-6-phosphatase : a criticai review, in The Enzyme of Biological Membranes 2"6 ed. (ed Marionosi, AN.), 2,349-398 (Plenum Publishing Corp., New York)
2. L&, K-J., Sheiiy, L.L., Pan, C.-J., Sidbury, J.B., and Choy J.Y. (1993) Mutations in the glucose-6-phosp hatase gene that cause glycogen storage disease type 1 a Science 262,580-583
3. Sheily, L.L., LA& K.-J., Pan, C.-J., Sakatii, S.F., Ruppert, S., Schutz, G., and Choy I.Y. (1993) Isolation of the gene for glucose-6-phosphatase, the enzyme deficient in glycogen storage disease type la. 1 Biol. Chern. 268,2 1482-2 1485
4. Lange, AI., Argaud, D., El-Maghrabi, M.R, Pan, W., Maitra, S R , and Pilkis, S.J. (1994) Isolation of a cDNA for the catalytic subunit of rat Liver glucose-6-phosphatase : Regulation of gene expression in FA0 hepatoma ceils by Uwlin, dexamethasone and CAMP. Biochem. Biophys. Res. Cornmim. 201,302-309
5. Berteloot, A, Vidal, H & van de Werve, G. (1991) Rapid kinetics of liver microsomal glucose-6-phosphatase. J. Bid. Chem. 266, 5497-5 5 07
6. Vidal, H., Berteloot, A, Lame, M . 4 , St-Denis, J.-F. and van de Werve, G. (1992) Interaction of mannose-6-phosphate with the hysteretic transition in glucose-6- phosphate hydrolysis in intact liver microsomes. FEBS Le#. 302, 1 97-200
7. St-Denis, J.-F., Annabi, B., Khoury, H., and van de Werve, G. (1995) Histone II-A stimulates glucose-6-phosphatase and reveals mannose-6-phosphatase activities without permeabhtion of liver rnicrosomes. Biochern. J. 310,221-224
8. RomaneIli, A, St-Denis, J.-F., Vidal, H., Tchu, S. and van de Werve, G. (1994) Absence of glucose uptake by iiver microsomes: An explanation for the complete latency of glucose dehydrogenase. Biochent. Biophys. Res. Commzin. 200, 149 1 - 1 497.
9. St-Denis, LF., Berteloot, A Vidal, R, Annabi., B., and van de Werve, G. (1 995) Glucose transport and glucose 6-phosphate hydrolysis in intact rat iiver rnicrosomes. J Biol. Chern. 270,2 1092-2 1097
10. Berteloot, A, St-Denis, J.-F., and van de Werve, G. (1995) Evidence for a membrane exchangeable glucose pool in the fùntioning of rat liver glucose-6- phosphatase. J. Biol. Chern. 270,2 1098-2 1 102

1 1. Schulze, K-U., and Speth, M. (1980) Investigations on the possible involvement of phosphoiipids in the glucose-6-phosphate transport system of rat-liver rnicrosornal glucose-6-phosphatase. Eur. 1 BBiochem. 106,505-5 14
12. Schulze, H.-U., Nolte, B., and Kannler, R (1986) Evidence for changes in the conformational status of rat liver microsomal glucose-6-phosphate : phosphohydrolase during detergentdependent membrane modification. Effect of pmercunienzoate and organomercurial agarose gel on glucosed-phosphatase of native and detergent- modifled rnicrosornes. J. Biol. C h . 261, 1657 1- 16578
13. St-Denis, J.-F., Comte, B., Nguyen, D.K., Seidman, E., Paradis, K., Lévy, E. and van de Werve, G. (1994) A conformational mode1 for the human liver microsomal glucose-6-phosphatase system : Evidence for rapid kinetics and def&s in glycogen storage disease type 1 . 1 Clin. fitdocrinolnoL Metab. 79,955-959
14. Lei, K.-J., Chen, H., Pan, C.-J., Ward, J.M., Mosinger, B., Lee, E.J., Westphal, H., Mansfield, B.C., and Chou, J.Y. (1 996) Glucose-6-phosphatase dependent substrate transport in the giycogen storage disease type- la mouse. Nature Genetics 13,203-209
15. Lei, K.-J., SheUy, L.L., Lin, B., Sidbuiy, J.B., Chen, Y.-T., Nordlie, RC., and Chou, J.Y. (1 995) Mutations in the glucose-6-phosphatase gene are associateci with glycogen storage disease types la and laSP but not lb and lc. J. Clin. Imesi. 95,234- 240
16. Nordlie, RC., Arion, W.J., and Glende, E.A. (1965) Liver microsomal glucose-6- phosphatase, inorganic pyrop hosp hatase, and pyrophosphate-glucose phosphotransferase. Effêcts of adrenalectomy and cortisone administration on activities assayed in the absence and presence of deoxycholate. J. Biol. C h . 240,3479-3484
17. Arion, W.J., and Nordlie, RC. (1967) Biological replation of inorganic pyrophosphate-glucose phosphotransferase and glucose-6-phosphatase. Activation by triarncinolone, in vivo, in the presence of actinomycin D. J. Biol. Chem. 242, 2207- 2210
18. Nordlie, RC., and Snoke, RE. (1967) Regdation of liver rnicrosomd inorganic pyrophosp hate-glucose p hosp hotmuferase, glucose-6-p hosphatase, and inorganic pyrophosphatase. Biochim. Biophys. Acta 148,222-232
19. Proctor, E.B., and Nordlie, RC. (1972) Responses of synthetic and hydrolytic activities of rat liver glucose-6-phosphatase to concurrent cortisone administration and experimental diabetes. Arch. Int. Physiol. Biochim 75,461-467
20. Traxinger, RR, and Nordiie, RC. (1990) Hormonal responses of glucose& phosphatase catalytic unit studied by stopped-flow analysis. Biochem. Cell Biol. 68, 454-45 8

21. Arion, W.J., Lange, AJ., and Baiias, L.M (1976) Quantitative aspects of relationship between glucose-6-phosphate transport and hydrolysis for liver microsomal glucose-6-phosphatase system. J. BioL Chem. 251,67846790
22. Hemnan, J.L., and Nordlie, RC. (1972) The catalytic potential of cart>amyl phosphate : glucose phosphotransferase : A species distribution study. Arch Biochem. Biophys 152,180-186
23. van de Werve, G. (1989) Liver glucose-6-phosphatase is modulated by physiologid intracelular Ca* concentrations. J. BioL Chem. 264,603 3 -603 6
24. Lucius, RW., Waddeü, I.D., Burcheii, A, and Nordlie, RC. (1993) The hepatic glucosed-phosphatase system in Ehrlich-ascites-himour-bearhg mice. Biochem. J 290,907-9 1 1
25. Gold, G., and Widnell, CC. (1 976) Relationship between microsomal membrane pemeability and t!e inhibition of hepatic glucose-6-phosphatase by pyridoxal phosphate. J. BioL Chem. 254, 1035- 1041
26. Speth, M., and Schuize, K.-U. (1992) The purification of a detergent-soluble glucose-6-phosphatase fiom rat liver. Eur. J. Biochem. 208,643-650
27. Massillon, D., Barzilai, N., Chen, W., Hu, M., and Rossetti, L. (1996) Glucose regulates in vivo glucose-&phosphatase gene expression in the iiver of diabetic rats. 1 Biol, C h . 271,9871-9874
28. Lowry, O.K., Rosebrough, N.J., Fan; AL. and Randali, RJ. (1951) Protein measurement with the f u h phenol reagent. J. Biol, C'hem. 193,265-275
29. Hanson, T.L., and Nordlie, RC. (1970) Liver microsomal inorganic pyrop hosphate-glucose phosphotransferase and glucose-6-phosphatase. E f f i s of diabetes and insului administration on kinetic parameten. Biochim. Biophy. Acla 198, 66-75
30. Burcheli, A, and Cain, D. 1. (1 985) Rat hepatic microsomal glucose&-phosphatase protein levels are increased in streptozotocin-induced diabetes. Diabetdogiu 28, 852- 856
31. Maddaiah, V.T., Chen, S.Y., Rezvani, I., Sharma, R, and Coiiipp, P.J. (1971) Inactivation of microsomal giucose-6-phosphatase by pyridoxal-S'-phosphate. Biochem. Biophys. Res Comm. 43, 1 14-1 18
32. Carvo, M., Maddaiah, V.T., Coliipp, P.J., Chen, S.Y. (1974) Hepatic rnicrosomd membrane : Activation of glucose-6-phosphatase. F m Lett. 39, L 02- 104

33. Speth, M., and Schulze, H J . (198 1) Hormone-induced effects on the rat liver microsorna1 glucose-6-phosphatase system in vitro. Biochem. Biophys. Res. Commun. 99, 134-141
34. Argaud, D., Zhang, Q., Pan, W., Maitra, S., Pilkis, S., and Lange, A (1996) Regulation of rat liver glucose-6-phosphatase gene expression in different nutritional and hormonal States. Diabetes 45, 1563- 157 1
35. Nordlie, RC. (1 974) Metabolic regdation by multininctional glucose-6- phosphatase, In Cz~rrent Topics in CettziIar Reptation (B.L. Horecker and E.R Stadtman, eds) 8,33- 1 17 (Academic Press, New York)
36. Band, J-C., and Jones, C.T. (1980) Functional activation by glucagon of glucose-6- phosphatase and gluconeogenesis in the perfised liver of the fetal guinea pig. FEBS Lett. 119, 190-194

Figun 1 : Western blotting with poiyclonal antibody raised against the cataiytic subunit of rat h e r Glc-bPase. Twenty mg of liver rnicrosomai protein, isolated ftom control (lane l), diabetic (lane 2) and triamcinolonetreated rats (lane 3) were loaded per track.
Figure 2 : Combined conformationai flexibiiity-substrate transport mode1 (adapted from Berteioot et ui!, 1995). Gic-6-Pase is represented as a protein spanning the entire membrane, with the rntalytic site located in a hydrophilic pocket (Vh: rate of Glc-6-P hydrolysis). Exchange of abstrate and products is realized by two channek with specïfic properties. An outer channel aiiows Glc-6-P uptake hi) and Glc production (km) outside the microsome. An inner channel hansports Glc into the lumen of the microsome (km). Glc and Glc-6-P represent their respective concentrations, the indexes O, m and i refer to the extemal medium, the pocket and the lumen of the microsome respectively.
Figure 3 : Glucosed-phosphatase inhibition by pyridoxai 5'-phosp hate in microsomes isoiated from control and triamcinolone-treated rats. htact microsomes isoiated fiom controi (O) or triarncinoione- (@) treated rats were incubated at 30°C for 16 min and in the presence of the indicated concentrations of PLP. Reaction was stopped with NaB& as described under "Experimental Procedures". Data are the means * S.E.M. using three dserent rnicrosomal preparations, and the enzymatic activities were measured with 5.0 mM [U-~~C]GIC-~-P.
Figure 4 : Tirnecourse of pyridoxal 5'-phosphate-induced Glc-dPase inhibition. Intact liver microsornes isolated fiom control (O) or triamcinoIone- (a) treated rats were incubated at 30°C in the presence of 100 rnM PLP. Data are the means k S.E.M. using three diierent rnicrosomal preparations.
Figure 5 : Effect of ligands of Glc-GPase on pyridoxal5'-phosphate-induced inactivation. Microsornes isolated fkom control rats were preinaibated in the presence of 20 mM of the indicated ligands, then incubated for 4 min at 30°C in the presence or the absence of 10 mM PLP and the reaction stopped with NaB& as described under "Experimental Procedures". The PLP/NaB&-treated microsomes were fùrther nibjected to detergent peneabilization with CHAPSO 0.8% or not. Data are the means * S.E.M. using three dserent microsoma1 preparations.
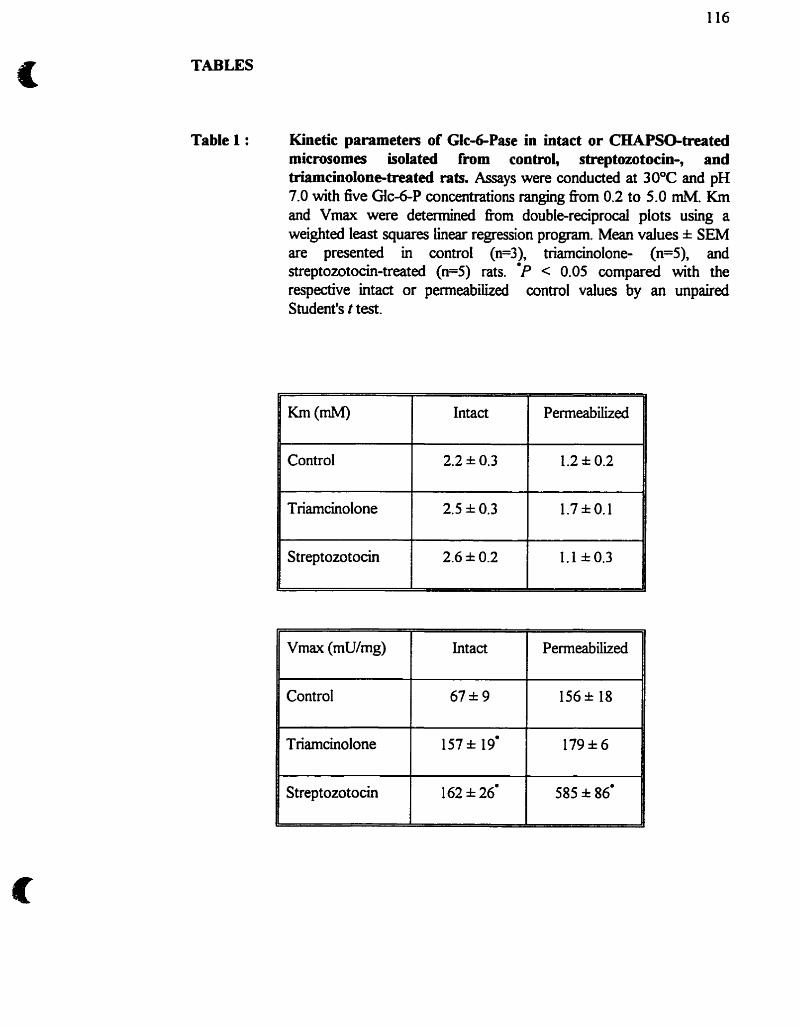
TABLES
Table 1 : Kinetic parameters of Glc-6-Pase in intact or CHAPSO-treated microsornes isolated fmm control, streptozotocin-, and hinmcinolone-treated rats. Assays were conducteci at 30°C and pH 7.0 with five Glc-6-P concentrations ranghg fiom 0.2 to 5.0 mM. Km and Vmax were determineci fkom doubie-reciprocal plots using a weighted least squares linear regrasion prograrn. Mean values SEM are presented in control (n=3), triarncinolone- (n=5), and streptozotocin-treated ( ~ 5 ) rats. 'P < 0.05 compared with the respective intact or pemeabilwd control values by an unpaireci Student's f test.
Intact
Control 1 2.2*0.3 1 1.2A0.2
Triamcinolone
Streptozotocin
Vmax (mU/mg)
Control
Triamcinolone
Streptozotocin 162 26' 585 k 86'
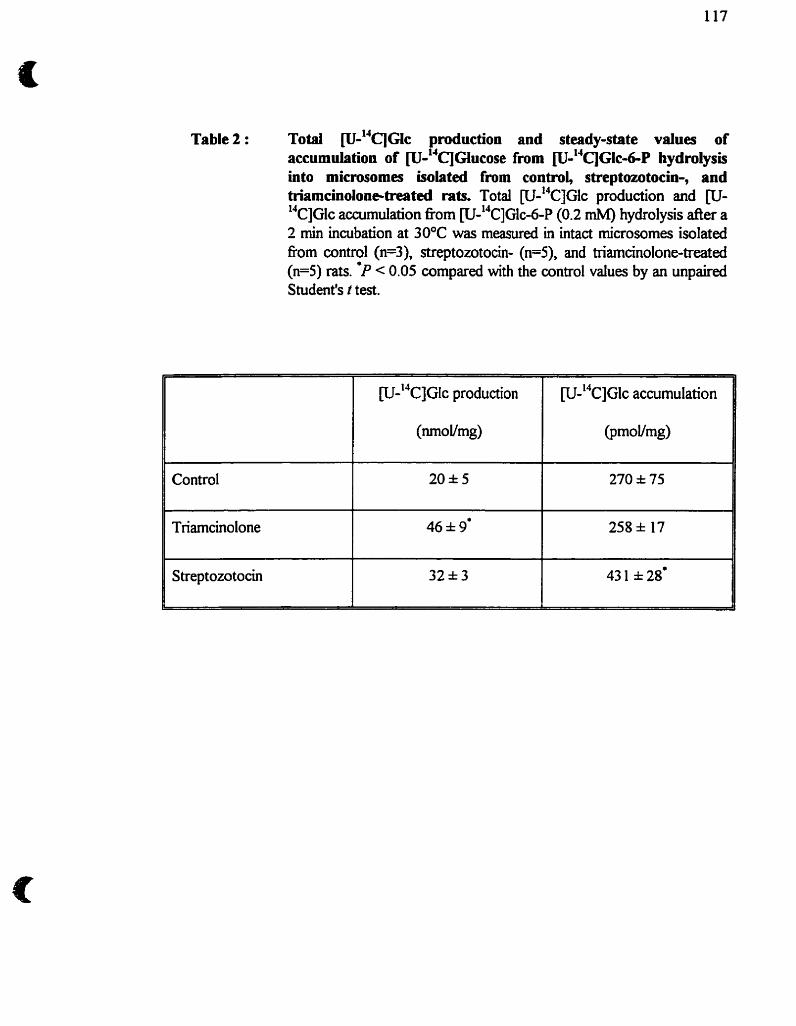
Table 2 : Total ~ 1 4 q ~ l c production and stesdy-state values of accumuiation of ~ ' 4 ~ ~ l u c o s e from W ' ~ ~ G I C ~ P hydrolysis into microsornes isolated from controi, streptaotocin-, and hiamcinolone-trrrited rats. Total [u-"CIGIC production and CU- 14~]GIc accumulation nom W-'~C]G~C-~-P (0.2 rnM) hydrolysis after a 2 min incubation at 30°C was measured in intact microsornes isolated ftom control (n=3), streptozotocin- (n=5), and triamcinolone-treated ( ~ 5 ) rats. 'P < 0.05 compared with the control values by an unpaireci Student's r test.
Control
S treptozotocin
W-'~C]GIC accumulation
@moVmg)
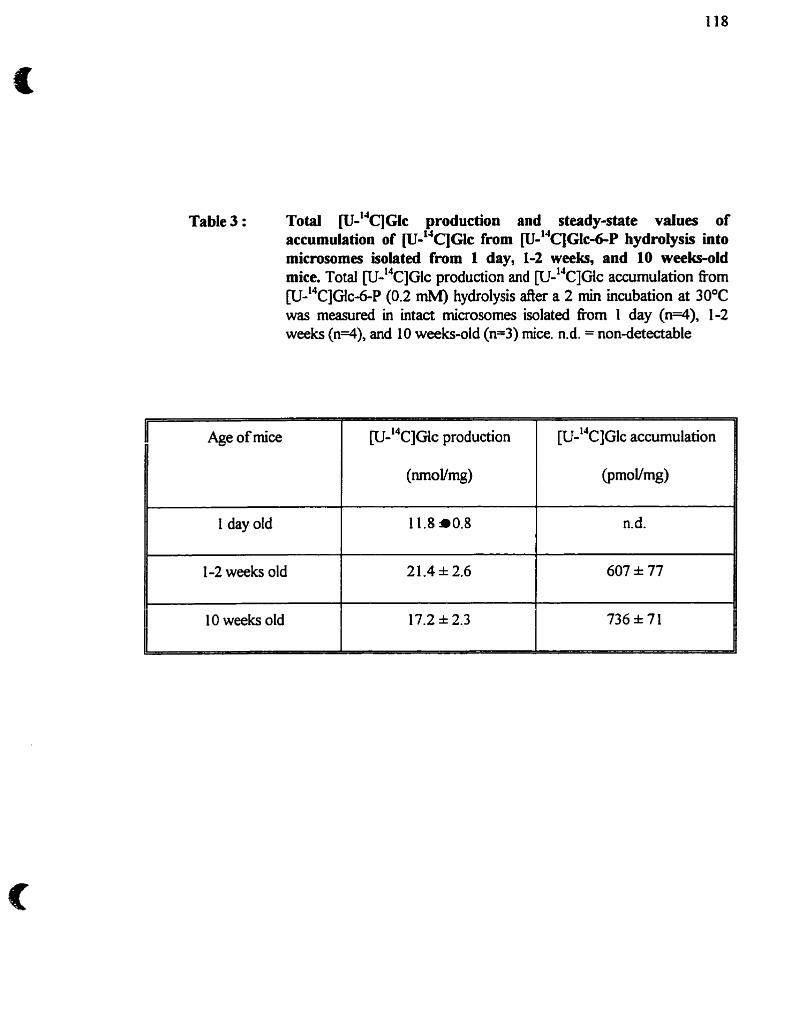
Table 3 : Total ~-"'C'JGIC production and steady-state values of accumulation of ~ - " q ~ l c from ~ " ' q ~ l c 4 - P hydrolysk into microsornes isolated from 1 day, 1-2 weeks, and 10 weeks-old mice. Total [u-'~c]G~c production and w-"~]Glc accumulation fiom P-"C]G~C-~-P (0.2 mM) hydrolysis der a 2 min incubation at 30°C was rneasured in intact microsornes isolated fiom 1 day (n=4), 1-2 weeks (n=4), and 10 weeks-oid (n=3) mice. n.d. = nondetectable
Age of mice W-"C]G~C production [U-~~CIGIC accumulation
(nl-novmg) @moVmg)
1 day old 1 1.8 a0.8 n,d.
1-2 weeks old 21.4 2.6 607 * 77
10 weeks old 17.2 * 2.3 736 * 71
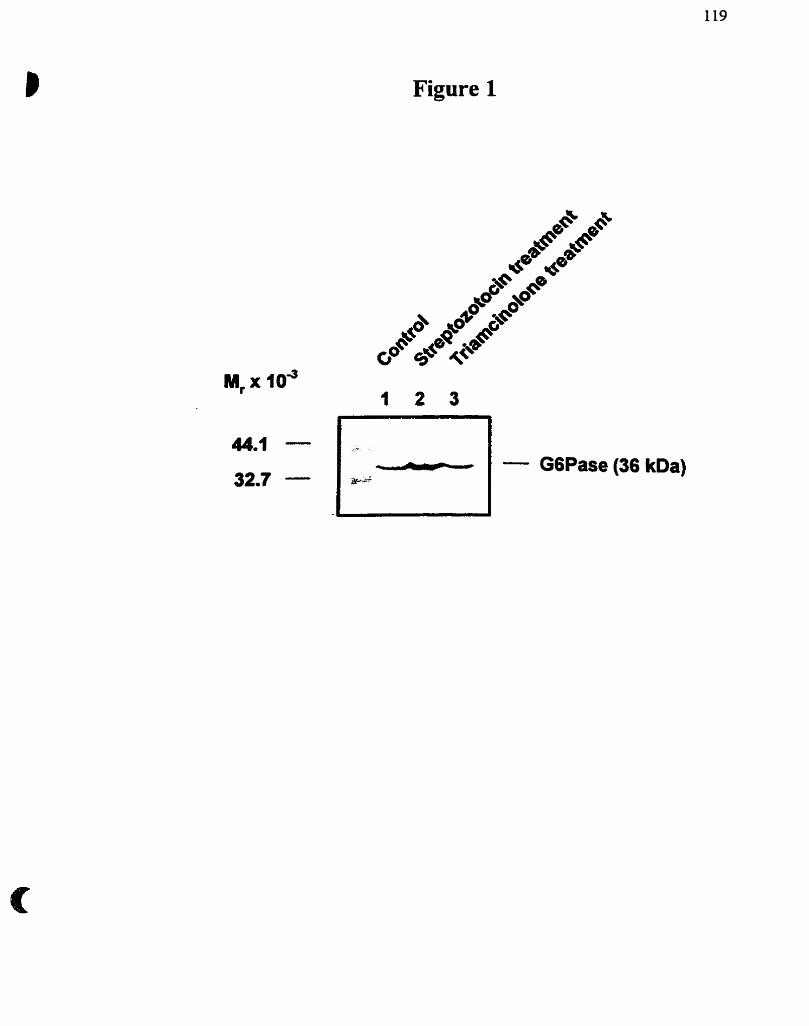
Figure 1
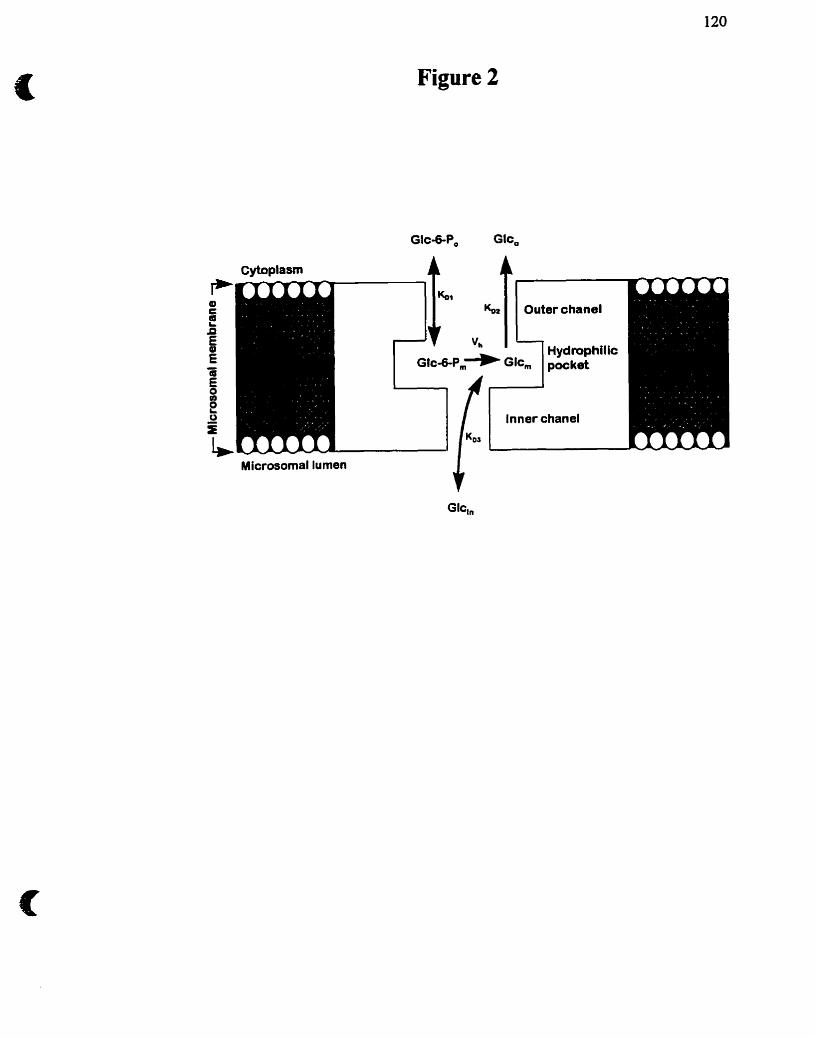
Figure 2
Glc+P, Glc
Cytoplasm A 4
Microsomal lumen
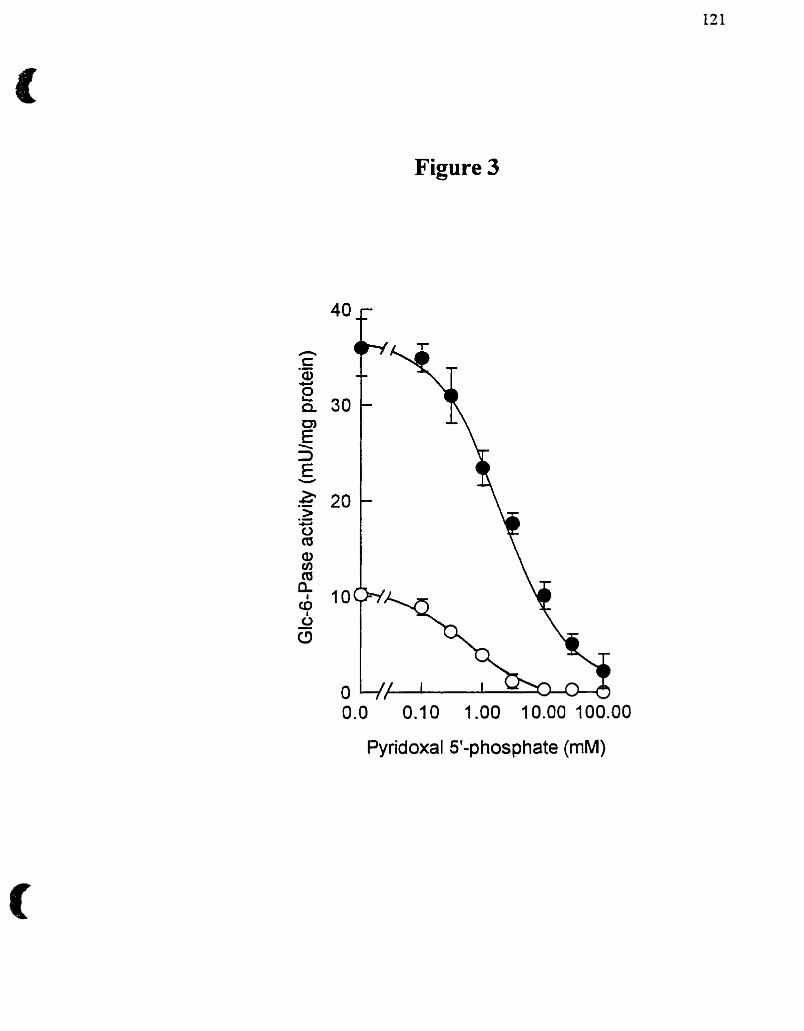
Figure 3
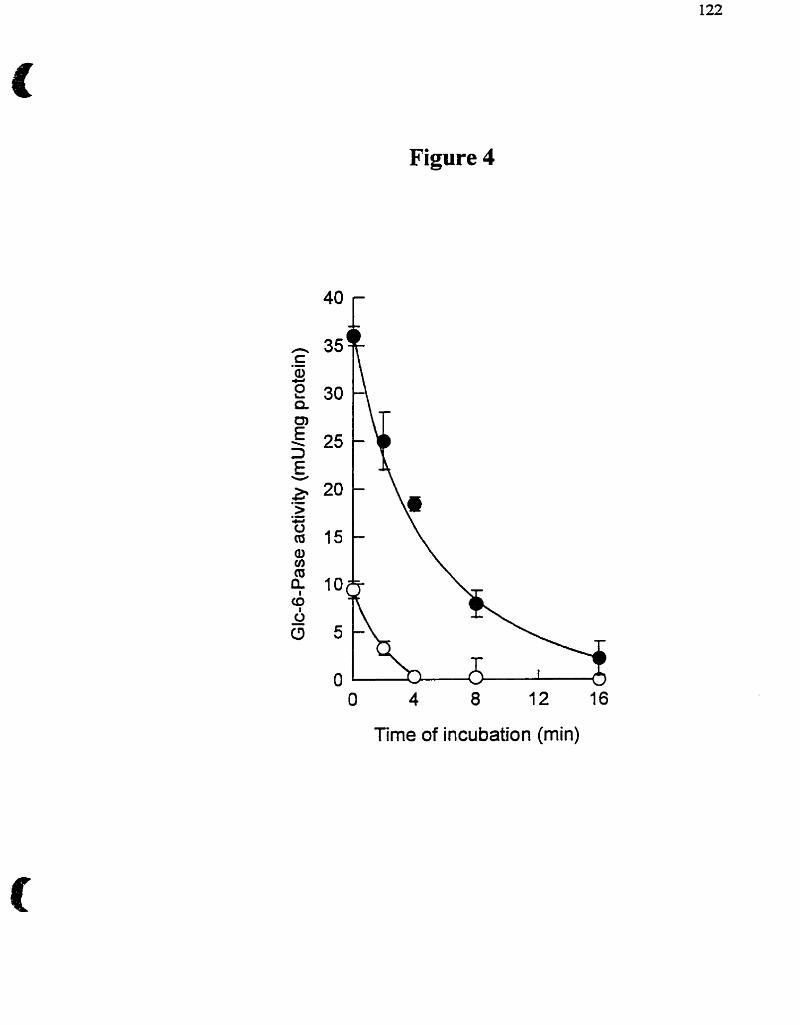
Figure 4
O 4 8 12 16
Time of incubation (min)
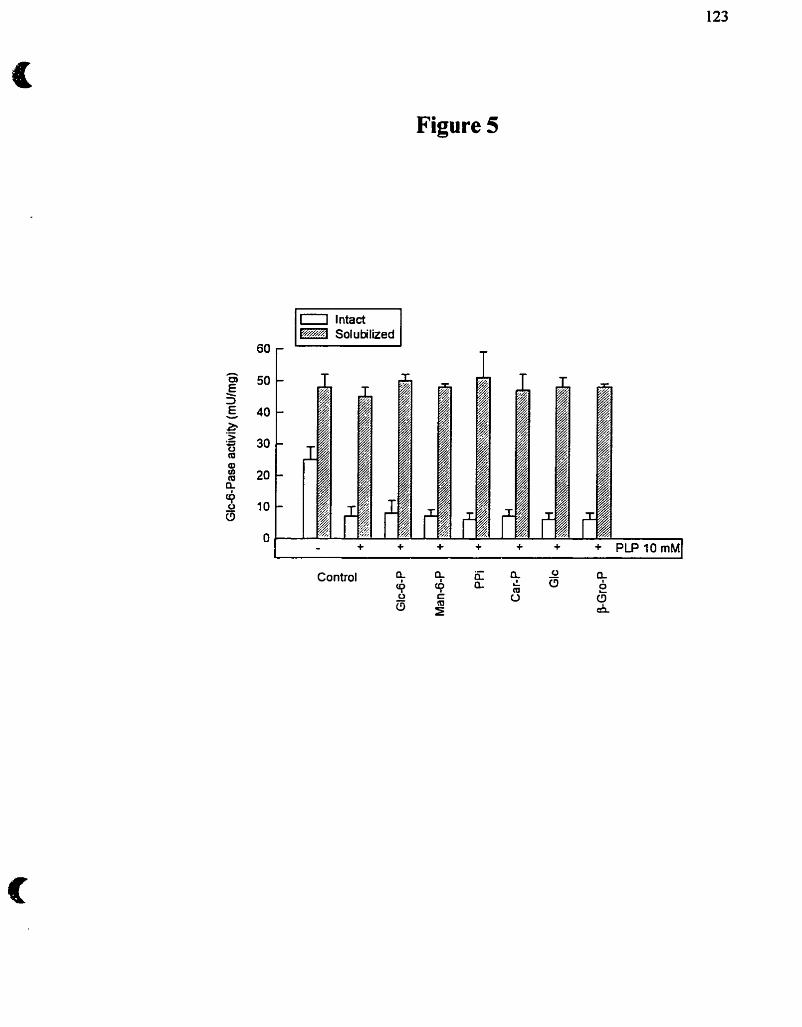
Figure 5
i intact Solubilized

4.1- Réévaluation du mdèle de transport du subsbat
4.1.1- Diagnostic des glycogénoses de type I
En examinant plus en détail les données publiées dans la littérature, ii nous
apparait que les conclusions tirées des données enzymologiques et/ou irnmunologiques,
et basées sur le mdèle ct? ~ ~ r t du substrat, sont loin de fàire I'unanimite, en
particulier dans le diagnostic des différents types de glycogénoses de type 1. En voici
quelques exemples : 1) Nordlie et al. (1983) ont caractérisé le premier w de
giycogénose de type Ic (déficience en T2) où curieusement le Pi ne pouvait s'accumuler
à l'intérieur des microsornes lorsque le substrat était du Glc-6-P. Associant ce cas à une
glycogénose de type Ib, les auteurs reconnaissent non seulement que le modèle de
tramport du substrat ne pouvait prédire une teile observation mais ont émis hypothèse
d'un système de transport de Pi plus complexe et composé d'au moins deux autres
protéines (Nordlie et al., 1992). 2) Le seul patient, à ce jour, diagnostiqué comme
glycogénose de type IaSP sur la base de données Unmunologiques (Burcheil et
Waddeil, 1990) a par la suite été démontré par Lei et al. (1995) comme un cas
classique de glycogénose de type Ia lorsque le gène de la Glc-6-Pase a été analysé. 3)
Rappelons qu'aucun cas de glycogénose de type Id (déficience en T3), n'a encore été
rapporté. 4) Enfin, plusieurs cas de déficience apparente du système Glc-6-Pase dans

les microsornes isolés de biopsies de foie humain, où une perméabilisation des vésicules
à I'aide de détergent révélait une activité Glc&Pase, pouvaient être renversés par un
lavage extensif des microsornes intacts ou par la présence d'albumine sérique de boeuf
(Pears et al., 1991, Burcheli et Gibb, 1991). Ces observations indiquent que les
glycogénoses de type 1 peuvent être dues à la présence de différents types d'inhibiteurs.
Ii a été rapponé récemment que les esters d'acyl-CoA pouvaient appartenir à une telle
classe d'ibiteurs (Mithieux et Zitoun, 1996).
4.1.2- Spécificité de la Glc-6-Pase pour le Glc-6-P : Un nouveau concept du
phénomène de latence
Les arguinenrs majeurs ailant contre I'organisation moléculaire du système Glc-
6-Pase dans la membrane du RE tel que prédit par le modèle de transport dir substrat
sont apportés par de nouvelles approches cinétiques. La validation cinétique d'un tel
modèle implique que soient vérifiés au moins 4 postulats qui, par ailleurs, ont tous été
contredits.
1) L'hydrolyse du Glc-6-P par la Glc-6-Pase dans les microsornes intacts
devrait être caractérisée par une latence due à I'équilibration préalable du Glc-6-P de
part et d'autre de la -membrane du RE par un transporteur putatif Tl. Deux groupes
indépendants (Berteloot el al., 199 1 a, Ajzannay et Mithieux, 1996) ont non seulement
démontré l'absence d'un tel phénomène, mais plutôt la présence d'une surproduction
("burst") de Glc dans les 15 premières secondes de l'hydrolyse du Glc-6-P.

2) Le nançpon du Glc-6-P, étape limitante dans le processus cataiytique de la
Glc-6-Pase, à travers la membrane intact des microsomes se doit d'être spécifique au
Glc-6-P et aucun autre hexose-phosphate. Cette propriété a été contredite d'abord par
%dai et al. (1992) en démontrant que le Man-6-P était, tout comme le Glc-6-P,
reconnu et hydrolysé pendant les premières secondes de la réaction, puis par St-Denis
et al. (1995a) en démontrant que le traitement des microsornes avec les histones II-A
permettait de révéler une activité Man-6-Pase sans en affecter i'intégrité rnembranaire.
3) Les produits d'hydrolyse du Gic-6-P doivent être formés à l'intérieur des
microsomes ou se trouve la sous-unité catalytique de la Glc-6-Pase. Au contraire,
seulement une hction (moins de 10%) da Glc produit par l'hydrolyse du Glc-6-P
transite dans la lumière des vésicules, alors que la majorité du Glc est relâchée dans le
milieu extramicrosomai (Berteloot et al., 199 1 a, Berteloot et al., 1995). De plus, le
Man produit lors de l'hydrolyse du Man-6-P, par les microsomes préalablement traités
aux histones II-4 l'est directement à l'extérieur des microsomes (St-Denis et al.,
1995a).
4) La spécificité de la Gk-6-Pase pour le Glc-6-P ne devrait être présente que
dans les microsoines intacts et non perméabilisés. L'absence de membrane permet
l'accès libre de différents hexoses-phosphate au site catalytique, mais il semblerait, que
l'enzyme soit capable de distinguer le Glc-6-P des autres hexoses-phosphate dans ces
conditions (Ajzannay et al., 1993, Ajzannay et al., 1994).
Les fài'bIesses du modèle de trmwport th substrat sont flagrantes. Il est donc
nécessaire de reconsidérer ce modèle en fonction d'un modèle plus simple basé sur les

nouveîîes données cinétiques nous permettant d'intégrer aussi les observations issues
des différentes glycogénoses de type 1. La Glc-6-Pase pourrait très bien n'être qu'une
seule protéine transmembranaire capable d'adopter trois types de conformations
cinétiques. La première serait présente dans la membrane intacte du RE dunuit la
première phase d'hydrolyse (état pré-stationnaire) du Gic-6-P. Cette première
conformation passerait d'une forme catalytique "rapide" mais peu spécifique dans les
premières secondes de cataIyse, à une seconde conformation "moins rapide" mais plus
spécifique pour le Glc-6-P (état stationnaire). Cette observation appuie notre
conclusion précédente qui énonce qu'une forme cinétique de l'enzyme? similaire à d e
des vésicules traitées au détergent, est présente dans les microsomes intacts au début de
lthydrolyse du Glc-6-P et que le transport spécifique du substrat ne peut être l'étape
lirnitante de l'hydrolyse du Glc-6-P @erteioot et a!., 199 1 a). Ce concept est compatible
avec une lente transition hystérétique de l'enzyme documentée par plusieurs groupes
(Foster et al., 199 1, Berteloot et al., 199 la, Jonges et al., 1992, Vidal et al., 1992,
Nelson-Rossow et al., 1993), ou la présence de la membrane joue un rôle essentiel
pour qu'une teUe transition ait lieu (Berteloot et al., 199 la, Ajzannay et Mithieux,
1996), et où la solubiiisation de la membrane microsornale à l'aide de détergent
empêche cette transition. La latence apparente de la Glc-6-Pase dans des microsomes
intacts ne peut donc s'expliquer par un transport limitant de Glc-6-P dans le
compartiment vésiculaire, mais dépend d'une phase transitoire et réversible du
mécanisme moléculaire d'hydrolyse du Glc-6-P. Notons, enfin, que l'absence de la
membrane microsomale (traitement au détergent) induit une troisième conformation de
la Glc-6-Pase où la transition vers une forme plus spécifique pour le Glc-6P est

empêchée et est semblable à celle induite suite au traitement des microsornes avec les
histones II-A (St-Denis et d, 1995a). Les caractéristiques de la Glc-6-Pase dans ces
trois conformations cinétiques sont résumées dans le tableau 3.
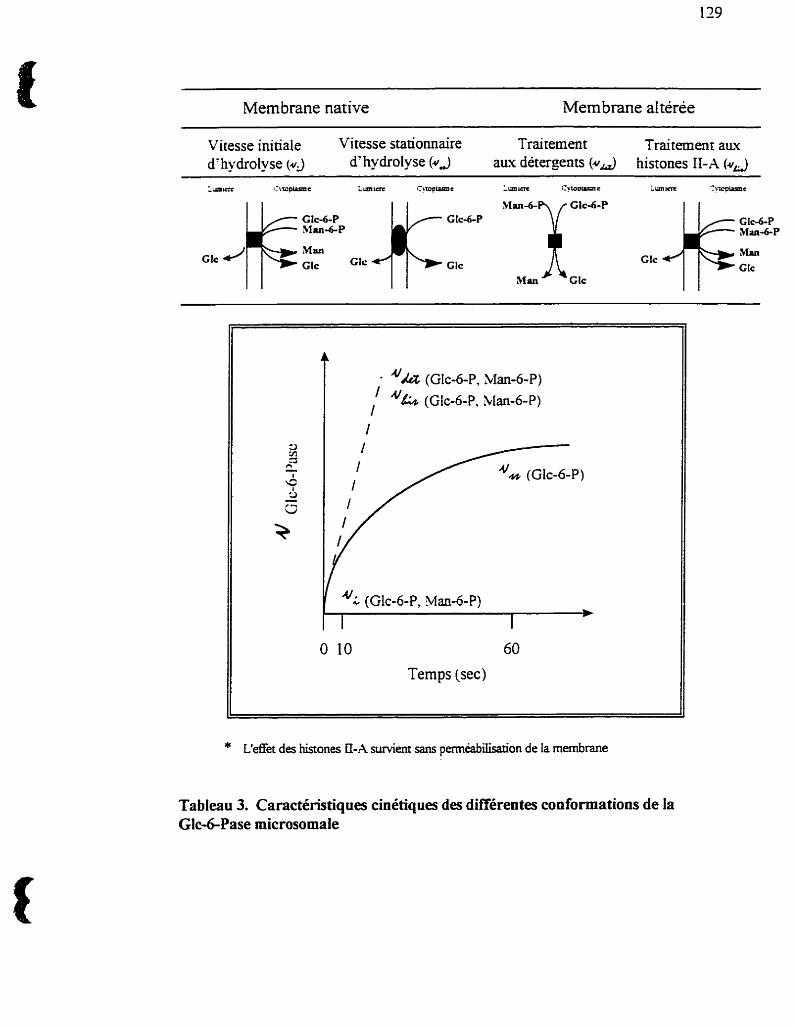
- - -- - - - - - -
Membrane native Membrane al téree
Vitesse initiale Vitesse stationnaire Traitement Traitement aux d'hydrolyse (4 d'hydrolyse (#A aux détergents (43 histones II-A ( u a
Temps (sec)
* t'effet des histones [I-A s u ~ e r n sans perméabilisation de la membrane
Tableau 3. Caractéristiques cinétiques des différentes conformations de la Glc-6-Pase microsomaie

4.2- Transport de glucose unidirectionnel associé au système Glc-6-
Pase
Le transporteur de glucose (GLUT 7), qui a été récemment cloné, a été
proposé comme étant le transporteur T3 du système Glc-6-Pase (Waddell et al., 1992).
Par contre, il a été démontré que les microsomes de foie ne captaient pas directement le
Glc (Rornanelli et al., 1994) contrairement au Pi qui s'accumule à Fintérieur des
microsornes (St-Denis et al., 1994). Cette absence d'entrée directe de Glc dans les
microsornes est intéressante puisqu'eiie pose la question de sa sortie des vésicules
intactes après que le Glc y soit formé suite à l'hydrolyse du Glc-6-P. Nous avons donc
étudié le transport de Glc dans les microsomes de foie pour établir ses caractéristiques
cinétiques ainsi que sa relation avec l'hydrolyse du Glc-6-P. Essentiellement, les
conclusions majeures sont que l'accumulation de [u- '~c]G~~ à partir de [u-"c]G~c-~-P
dans les vésicules est directement proportionnelie à l'activité Glc-6-Pase, mais que sa
sortie est indépendante de l'activité de l'enzyme. Nous montrons donc que le transport
de Glc est unidiiectiomei (de la lumière microsornale vers le cytosol) et avons en outre
observé que le Glc transitant a l'intérieur des microsomes n'est qu'une hction de d u i
produit majoritairement (plus de 90%) dans le cytosol. Enfin, l'identité de GLUT 7
n'ayant pas été confirmée par d'autres, il nous est permis de douter de l'existence de ce
type de transporteur de Glc microsomal. Alternativement, les propriétés du transport
de Glc telies que nous les décrivons dans la membrane native du RE sont assujetties a
la présence d'autres composantes du système Glc-6-Pue.

4.3- Modulation de l'activité glucose-6-phosphatase
4.3.1- Effet des histones II-A sur la glucose-6-phosphatase
On a attribué a certaines protemes polycationiques teiles la spermine, la
spermidine, la putrescine, ainsi que les histones (Nordlie et al., 1979, Blair et Burcheii,
1988), la propriété de perméabiliser ou de créer des pores dans les membranes
plasmiques ou dans les membranes des organelles intracellulaires (mitochondrie, RE).
Les histones II-A ont donc remplacé, dans certaines éîudes, l'usage classique des
détergents pour solubiliser la membrane du RE ou les caractéristiques de la Glc-6-Pase
sont identiques entre les deux traitements. La perméabilité des vésicules n'a, cependant,
pas été testée et la preuve irréfùtable de I'intégrité rnembranaire des microsomes traités
avec les histones II-A est toujours manquante.
Le mécanisme d'action des histones, en particulier celui des histones II-A, est
pour le moment inconnu. Des interactions électrostatiques entre les charges positives
des histones et les phospholipides membranaires chargés négativements sont postulées
(Johnson et Nordlie, 1980, St-Denis er al., 1995a). Rappelons que le traitement des
microsomes avec un détergent induit une activité Glc-6-Pase accrue (phénomène de
latence) et une perte de spécificité de la Glc-6-Pase envers le Glc-6-P. La Glc-6-Pase
est d o n capable d'hydrolyser différents ester-phosphates, dont le Man-6-P.
L'augmentation de i'activité Man-6-Pase a dès lors été utilisée comme indice de
l'intégrité membranaire des microsomes. Nos résultats montrent que le traitement des

microsomes intacts avec les histones Il-A résulte effectivement en une activité accrue
de la Glc-6-Pase comparable à celle induite par un traitement avec le détergent
CHAPSO. De plus, le Mand-P étant reconnu et hydrolysé au même taux que le Gk-6-
P, les activités Glc-6-Pase et Man-6-Pase sont donc en tout point identiques entre un
traitement au détergent ou aux histones II-A Mais qu'en est4 de I'intégrité
membranaire des microsornes ?
Une expérience simple a donc consisté à mesurer l'accumulation du Glc, à
partir du Glc-6-P, à l'intérieur des microsomes. Pour ce faire, un appareil à filtration et
à échantillonnage rapide a été utilisé. Le résultat est sans équivoque : le traitement des
microsomes avec les histones II-A préserve l'intégrité membranaire des vésiailes,
contrastant avec i'effet du détergent, où aucune accumulation n'est observée.
L'augmentation du plateau d'accumulation de Glc est, de plus, proportionnelle à
i'augmentation de l'activité Glc-6-Pase. Enfin, aucune accumulation de Man, résultant
de l'hydrolyse du Man-6-P, n'a pu être détectée, malgré la révélation d'une activité
Man-6-Pase. Finalement, une interaction de type éléctrostatique a été postulée et mise
en évidence avec la réversibilité par IEGTA et I'EDTA de i'effet des histones II-&
suggérant que ces chélateun agissent par neutralisation de charges. Plus important
encore, cette réversibilité ne semit pas observée si la membrane avait été peméabilisée.
De plus, une interaction de type protéine-protéine sur une préparation partiellement
purifiée de la Glc-6-Pa est suggérée étant donné que la Glc-6-Pase y est toujours
stimulée par les histones 11-A

La stimulation de l'activité Glc-6-Pase ainsi que la perte de spécificité de
l'enzyme envers le Glc-6-P, après traitement des microsornes avec un détergent ou avec
des protéines polycationiques, sont attribuées à la libéraîion du site catalytique de son
environnement lipidique. L'augmentation d'activité Gic-6-Pase dans les microsomes
traités avec les histones II-A ne peut, par contre, être attniuée à une perméaûiiisation
des vésicules, ces derniers pouvant toujours accumuler du Glc proportionnellement à
l'augmentation de l'activité Glc&Pase. Les histones II-A induisent probablement un
changement conformatio~el de la protéine Glc-6-Pase similaire à celui induit par le
traitement au détergent. Les histones II-A ne peuvent donc être utilisés comme agent
de perméabilisation des microsomes. La mesure de la Man-6-Pase ne peut être
considérée comme un indice de l'intégrité des microsomes étant donné que l'activité
Man-6-Pase est identique entre un traitement au détergent ou aux histones 11-4 mais
ce dernier sans perméabiliser les vésicules. De plus, le Mm, ne s'accumulant pas dans
les microsomes traités aux histones 11-4 est produit directement dans le milieu
extramicrosomal. Enfin, la tàible activité Man-6-Pase retrouvée dans les microsomes
intacts est le reflet d'une proportion de la Glc-6-Pase qui est dans une conformation
capable d'hydrolyser le Man-6-P. Nos données indiquent donc que la topologie de la
Glc-6-Pase n'est pas d e prédite par le modèle de transport di srrbstrat.

4.3.2 Fonctions distinctes de production et d'accumulation de glucose
Les changements dans les propriétés cinétiques de la Glc-6-Pase hépatique
induits par l'administration de doses pharmacologiques de streptozotoche, d'afloxane,
ou de glucocorticoides ont bien été documentés (Nordlie et al., 1965, Nordlie et al.,
1968, Hanson et Nordlie, 1970, Proctor et Nordlie, 1972, Arion et al., 1976a). En
réponse aux glumcorticoides, i'augmentation de la Vrnax ainsi que la diminution de la
latence de l'activité Glc-6-Pase a été interprétée, en se basant sur le modèle de
transporf dtc substrat, comme une augmentation de l'activité de Tl (Arion et al., 1975,
Arion et al., 1976% Traxinger et Nordlie, 1990). La déficience en insuline, quant à eile,
résulte en une augmentation de la Vrnax de 2 à 3 fois et en une latence augmentée. Ces
observations ont été attribuées à la seule augmentation de la quantité de protéine de la
sous-unité catalytique de la Glc-6-Pase (Arion et al., 1975, Burchel et Cain, 1985).
Toutes ces observations ont été faites en examinant la production totale de Glc
résultant de l'hydrolyse du Glc-6-P et ce sans discrimination quant au Glc produit à
I'extérieur ou transitant à I'itérieur des microsornes.
Nous confirmons, dans un premier temps, que la production de Glc se faisait
majoritairement vers l'extérieur des rnicrosomes dans une proportion d'environ 90% à
I'extérieur et 10% du Glc qui transite à l'intérieur des rnicrosomes (St-Denis et al.,
1995b, Berteloot et al., 1995). Nous montrons, pour la première fois, que ces deux
fonctions de production de Glc (l'une vers l'extérieur et l'autre vers i'intérieur des
microsornes) sont indépendantes l'une de l'autre (article 4). L'observation que les

microsornes isolés de souris âgées de un jour puissent hydrolyser du Glc-6-P sans pour
autant en accumuler le prouve et va à l'encontre d'un des postulats du rnodgle de
tramport dh substrat qui est que le Glc-6-P soit d'abord transporté à l'intérieur des
microsomes afin d'être hydrolysé et que les produits d'hydrolyse, le Glc et le Pi, soient
ensuite transportés vers l'extérieur des microsomes. L'hypothèse d'une forme de la Glc-
6-Pase nécessitant certaines modifications post-traductionnelies supplémentaires pour
acquérir une fonction nomale ou une insertion optimale dans la membrane du RE n'est
pas exclue. Nous démontrons aussi que le traitement des rats à la triamcinolone
induisait une augmentation d'activité Glc-6-Pase dans les microsomes intacts.
Cependant, cette activité se traduit exclusivement par l'augmentation de la fonction de
production de Glc vers i'extérieur des microsornes sans production accrue de Glc vers
l'intérieur des microsomes. La latence diminuée de la Glc-6-Pase observée lors de
traitement aux glucocorticaides tels la corticostérone, la cortisone, et le cortisol (Speth
et Schube, 198 1) pourrait dorénavant s'expliquer par l'augmentation spécifique de cette
fonction de production de GIc vers l'extérieur des microsomes.
L'effet du traitement in vivo des rats avec l'analogue des glucocorticoides
triamcinolone semble donc moduler spécifiquement la Glc-6-Pase dans sa fonction de
production de Glc vers l'extérieur des microsomes. Cet effet de la triamcinolone est
indépendant des niveaux dARNm ainsi que de la quantité de protéine Glc-6-Pase. Ces
résultats sont différents de ceux de la dexarnéthasone dans des celu1es en cultures FA0
(Lange et al., 1994) où les niveaux d'ARNm de la Glcd-Pase sont augmentés. Pour
expliquer l'effet de la triamcinolone, nous postulons un changement dans

l'environnement membranaire, dans la composition en phospholipides et/ou dans la
fluidité de la membrane (Band et Jones, 1980), ou l'induction diine protéine régulatrice
q" aurait une fonction de couplage entre les fonctions de recomaissance/hydrolyse du
Glc-6-P et de production de Glc vers I'intérieur des rnicrosomes. Dans le but de tester
l'hypothèse de la protéine régulatrice, nous avons utilisé le pyridoxal 5'-phosphate
(PLP) comme outil dans des expériences de modifications chimiques, et observons des
cinétiques d ' i i i t ion différentes des rats controles (valeurs de Ki et tx d'inhibition par
le PLP plus grandes) nous indiquant la possibilité d'une induction diine protéine
régulatrice par la triamcinolone. L'hypothèse alternative serait que le PLP modifie un
site de reconnaissance du Glc-6-P dans les microsomes intacts mais que la contrainte
membranaûe induite par la tnamcinolone empêche le PLP dv avoir accès. Ces
observations sont compatibles avec celles de Speth et Schuize (1992). qui ont pu
marquer plusieurs polypeptides avec du L~H]PLP. Dans une préparation partiellement
purifiée, un de ces polypeptides de 36 kDa a été identifié comme la sous-unité
catalytique de la Glc-6-Pase. La disponibilité d'anticorps spécifiques dirigés contre
l'unité catalytique de la Glc-6-Pase devrait permettre d'irnmunoprécipiter et de
confimer l'identité de cette protéine marquée radioactivement.

4.3.3- État oligoménque de la glucosebphosphatase
On retrouve dans la littérature plusieurs estimations du poids moléculaire de la
Glc-6-Pase, que ce soit à partir de préparations partieliement purifiées de foie humain
(Reczek et Viee, 1982), ou déduites à l'aide de techniques telles hactivation par
irradiation (Ness et al., 1989). Ces études ont toutes révélé des formes de Glc-6-Pase
de poids moléculaire plus élevé allant, la plupart, de 58 a 75 D a . Nos résultats
indiquent l'existence diine teiie protéine rnernbranaire de poids moléculaire d'environ 64
kDa et ayant les propriétés aîtendues de la Glc-6-Pase. De plus, l'observation récente
diine éventuelle forme longue, encodée par un gène de la Glc-6-Pase avec une
extrémité 3'-non-traduite supplémentaire (Lange ei al., 1994, Argaud ei al., 1 996),
suggère la présence dhne telie sous-unité catalytique de plus haut poids moléculaire
(64 kDa) d'où proviendrait une forme native instable et de poids moléculaire plus fable
(36,5 kDa) (Countaway et aL, 1988, Speth et Schulze, 1992).
Les premières indications ont été apportées lors de la mise au point du
protocole de purification de la Glc-6-Pase. Tel que décrit plus haut (section 2.3) et
iilustré par la figure 9, nous arrivons à purifier une protéine de 64 kDa sur une colonne
d'affinité et possédant toujours une activité Glc-GPase, quoique très instable. Cette
protéine, à caractère fortement hydrophobe, n'a maiheureusement pu être
microséquencée et sa séquence primaire reste à être déterminée. Rappelons que Kosel
et al. (1993) ont également pu& une protéine de 63 kDa et, à l'aide d'anticorps
générés contre la Glc-6-Pase, ont démontré que son irnrnunoréactivité suite à une

éléctrophorèse en conditions dénaturantes disparaissait dans certains des microsornes
isolés de patients atteints de glycogénose de type l a Cette immunoréactivité était
absente à 36 kDa et apparaissait à 260 kDa lorsque I'éléctrophorèse était effechée en
conditions non-dénaturantes (Kosel et al., 1993, Mithieux et al., 1995). Ces
obsenmtions nous amènent à postuler une capacité d'oligomérisation de la sous-unité
catalytique de la Glc-6-Pase.
La possibilité que la Glc-6-Pase de 36 kDa puisse être retrouvée sous forme
d'homodimère, d'hétérodimère, ou même de multirnère dans des conditions non-
dénaturantes suggère qu'elle puisse soit se lier à d'autres polypeptides par des ponts
disuhres, soit s aggréger. Cette possibilité a été étudiée en traitant une hction
partieHement purifiée de la Glcd-Pase sur colonne d'hydroxylapatite dans des
conditions favorables à la formation d'aggregats de protéines hydrophobes résistants au
SDS (Sagné et al., 1996). Ces conditions peuvent être le traitement à pH acide avec de
l'acide trichloroacétique (TCA), avec un solvant organique tel que l'acétone, ou le
chauffage de i'échantillon. L'immunobuvardage à I'aide d'un anticorps dirigé contre la
sous-unité catalytique de 36 kDa de la Glc-6-Pase (figure 14) nous révèle le résultat de
tels traitements. L'extrème instabilité et susceptibilité de la protéine à la protéolyse est
démontrée par la disparition de la bande immunoréactive à 36 kDa lors du chauffage.
De plus, le traitement au TCA ou à l'acétone fàvorise I'apparition d'une protéine
irnmunoréactive à 64 kDa qui est moins susceptible à la protéolyse. Notons que ces
expériences ont été Eiites en conditions dénaturantes et que la possibilité de générer des
formes de plus haut poids moléculaire (260 kDa) n'est pas exclue. Enfin, ce phénomène

d'aggrégation pourrait très bien avoir lieu en présence d'une autre protéine intimement
liée au processus catalytique de la Glc-6-Pase dans les microsornes intacts. Rappelons
simplement que dans certains cas de giycogénoses de type Ib, caractérisées comme
étant dus à un défaut dans la fonction de transport du Glc-6-P, aucune mutation n'a été
observée dans le gène codant pour la Glc-6-Pase (Lei et al., 1995). Ce type de
glycogénose pourrait donc être causé par i'absence ou la déficience d'une éventuelle
proteme régulatrice.
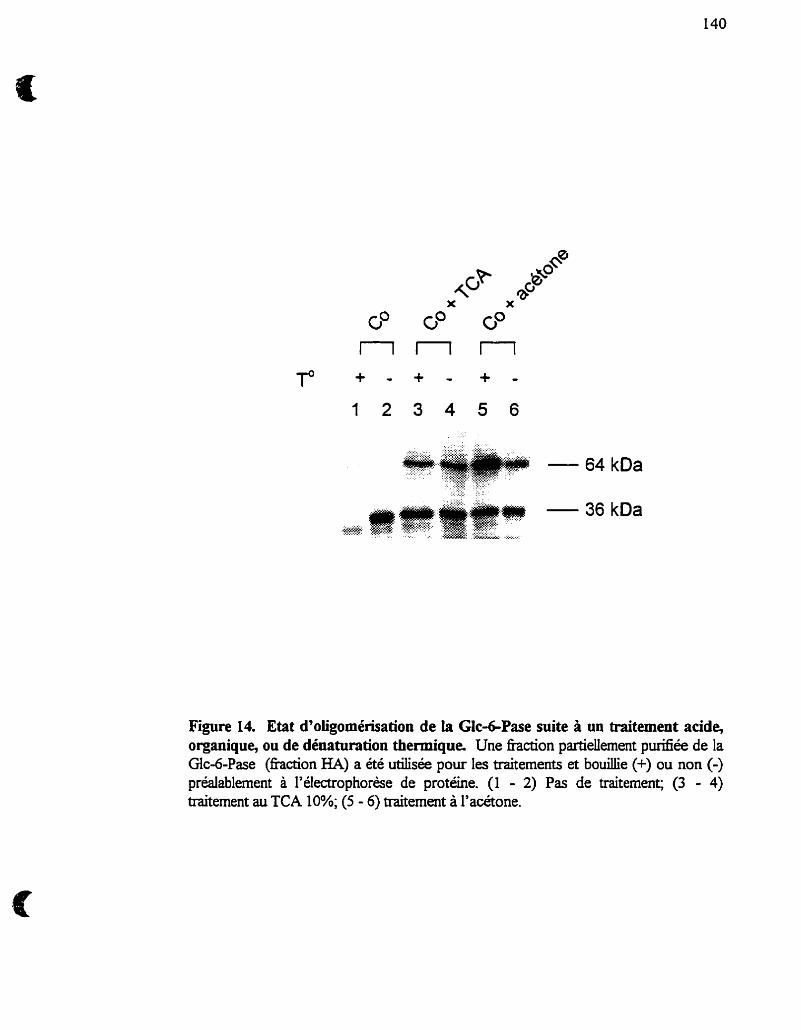
Figure 14. Etat d'oiigomérisation de la Glc4Pase suite à un traitement acide, organique, ou de dénaturation thermique. Une fiaction partiellement purifiée de la Glc-6Pase (fraction HA) a été utilisée pour les traitements et bouillie (+) ou non (-) préaiablement à l'électrophorèse de protéine. (1 - 2) Pas de traitement; (3 - 4) traitement au TCA 10%; (5 - 6) traitement à l'acétone.

4.4 Résidus impliqués dans le catalytique de la Glcd- Pase:
Inactivation de la Glc-GPase par le dicyclohexylcarbodümide
Dans cette étude nous avons utilisé le DCCD comme agent chimique pouvant
modifier spécifiquement les groupements carboxyles des résidus Aspartate et
Glutamate. Des études initiées par Feldman et Butler (1969) ont suggéré la présence de
résidus histidines impliqués dans le site actif de la Glc-6-Pase. De plus, le
diéthylpyrocarbonate, un agent modifiant spécifiquement les résidus histidine, inhie la
Glc-6-Pase (Arion et al., 1984). Le mécanisme catalytique des protéines ftisant partie
de la famille des histidine phosphatases implique une triade catalytique His-Asp/Glu-
His. L'intermédiaire E-P serait formé sur un des résidus His, dors que le second résidu
His serait impliqué dans le transfert du groupement phosphate. Le résidu Asp/Glu
servirait alors de donneur de proton acide, ou de base lors de l'hydrolyse du phosphate
de l'intermédiaire E-P. Par analogie, les groupements carboxyles modifiés par le DCCD
auraient le même rôle lors du processus catalytique de la Glc-6-Pase. Alternativement.,
les résidus (modifiés par le DCCD) pourraient être impliqués dans le mécanisme de
reconnaissace et/ou de liaison du substrat.
L'incubation d'une hction partieuement purifiée de la Glc-6-Pase (sur colonne
d'HA) en présence de DCCD conduit à une perte d'activité dose-dépendante (article 3).
La concentration de DCCD requise pour inactiver de 50% la Mc-6-Pase est de 0.2-0.4
rnM, et l'ajout de GEE, en tant qu'agent nucléophile Ion de la réaction de modification

chimique, n'affecte pas l'inactivation de la Glc-6-Pase par le DCCD. La représentation
serni-logarithmique de l'activité résiduelle de la Glc-6-Pase en fonction du temps pour
différentes concentrations de DCCD résulte en des tracés linéaires. La pente des tracés
(ELPP) en fonction de la concentration de DCCD nous prédit la modifiication de plus
d'un groupement carboyle. De plus, l'inactivation de la Glc-6-Pase par le DCCD est
compléternent protégée par la présence de Glc-6-P et nous avons donc tenté de voir si
d'autres substrats, ou produits des réactions catalysées par la Glc-6-Pase, pouvaient
avoir le même effet protecteur. La Glc-6-Pase, lorsqdextraite de la membrane
microsomale, perd de sa spécificité envers le Glc-6-P et peut hydrolyser le Man-6-P, le
PPi et le carbamyl-phosphate. Ces substrats ont tous eu un effet protecteur envers
l'inactivation de la Glc-6-Pase par le DCCD, dors que le sn-glycerol 3-phosphate, la
Glc(NHt)-6-P. le Glc- 1-P, le Glc, et I'ATP n'offiraient que peu de protection.
Dans le but d'identifier la composante catalytique de la Gic-6-Pase, la hction
hydroxylapatite a été incubée en présence de [ 1 4 ~ ] ~ ~ ~ ~ . L'autoradiographie
subséquente résulte en un marquage de 4 a 5 bandes avec des M, 36,000-77,000. Par
contre, le patron de marquage n'est pas affecté par la présence de Glc-6-P, suggérant
qu'aucune des protéines marquées ne représentait la sous-unité catalytique de la Glc-6-
Pase (article 3). À l'inverse, en utilisant du DCCD non-radioactif en présence de [I-
1 4 ~ ] ~ ~ ~ nous parvenons à marquer une seule protéine de 68 KDa. De plus, le patron
de marquage est alors afExte par le Glc-6-P, le Man-6-P, et le carbamylP, mais pas
par le sn-glycerol3-P ou I'ATP en accord avec les données de protection de l'activité

Glc-6-Pase. Enfin, notons qu'aucun marquage n'a été observé en présence de [l-
1 4 ~ ] ~ ~ ~ seul, et que la PGlcM commerciale n'est ni affectée, ni marquée par le
DCCD/[ 1 - ' 4 ~ ] ~ ~ ~ . Ces résultats suggèrent que la composante catalytique du système
Glc-6-Pase est une protéine de 68 kDa
Plusieurs questions restent à être élucidées quant a l'identité et au rôle de
cette protéine de 65-68 kDa. La protéine marquée avec le DCCD/[I-14c]~EE est-
elle la même que celle reconnue par l'anticorps généré contre la sous-unité
catalytique de la Glc-6-Pase de 36 kDa et que celle que nous purifions sur une
colonne de Glc(NH2)-6-P-Sepharose? Ces questions ne trouveront de réponses
que lorsqu'on aura la séquence primaire de la protéine purifiée que l'on pourra,
alors, comparer à la séquence de la Glc-6-Pase clonée. Le couplage de l'anticorps à
une matrice de Sépharose pourrait nous servir subséquemment d'outil pour purifier
la protéine de 65-68 D a .

5- CONCLUSIONS GENERALES
Le modèle confotmafionne : une alternative au modéle de
b.aqott du substrat
L'organisation moléculaire du système Glc-6-Pase a fàit l'objet de nombreuses
études, et a donne naissance aux deux modèles discutés plus haut. Les résultats obtenus
au cours des dernières années, en particulier à l'aide de la technique d'échantuomage et
de filtration rapide, ont grandement aidé à élucider le fonctionnement de la Glc-6-Pase
dans son environnement membranaire et renforcent le modèle confomationnel. Que ce
soit 1) i'absence détape ümitante de transport du Glc-6-P lors de la réaction globale
d'hydrolyse; 2) la reconnaissance aussi bien du Glc-6-P que du Man-6-P par la Glc-6-
Pase dans des microsomes intacts; 3) la proportion de Glc majoritairement produit vers
I'extérieur des microsornes (plutôt que dans la lumière du RE), ou 4) la perie de
spécificité de la Glc-6-Pase envers le Glc-6-P sans perméabilisation de la membrane
microsomaie, tous ces effets peuvent être expliqués en terme de changement de
conformation de la Glc-6-Pase et amènent une alternative intéressante au concept de la
latence de la Glc-6-Pase décrit par le ntcxièle de t r ~ l z ~ p o r i ? dr< substrd.
Enfin, nous proposons un mécanisme possible expliquant le phénomène de la
latence en démontrant que la fonction de production de Glc vers l'extérieur peut être
indépendante de celie vers l'intérieur des microsornes (article 4). Alternativement, le Pi

produit lors de l'hydrolyse du Glc-6-P dans les microsomes intacts, et connu pour
inhiber la Glc-6-Pase, pourrait s'accumuler dans la poche hydrophylique de la Glc-6-
Pase et être responsable de la latence. La caractérisation cinétique d'un tel phénomène
et la relation entre lhydrolyse du Glc-6-P et le transport de Pi restent à être
démontrées. La régulation hormonale de l'activité latente de la Glc-6-Pase in situ peut
donc avoir lieu par un tel mécanisme fisant appel soit à la régulation des fonctions
distinctes de production de Glc, soit au mécanisme d'inhibition par le Pi.
5.2- Preuves d'un couplage des fonctions d'hydrolyse du Glc-6-P et
d'accumulation du glucose
Plusieurs arguments favorisent donc un modèle d'organisation de la Glc-6-Pase
autre que celui prédit par le modèle de tramport du substrat, et soulèvent plusieurs
questions quant à la caractérisation fonctioimelle et organisationnelle des composantes
du système. Des questions telles que 1) la sigrilfication de la fable proportion (lû??) du
Glc produit suite à l'hydrolyse du Glc-6-P qui transite à l'intérieur des microsomes
(Berteloot et al., 199 la, St-Denis et al., 1995b), 2) la signification physiologique du
concept redéfini de la "latence" (Vidal et al., 1992, St-Denis et al., 1995a, article 4), 3)
La présence de la sous-unité de 36 kDa de la Glc-6-Pase dans différentes lignées
cellulaires exempt es d'activité d'hydrolyse du Glc-6-P, ainsi que dans plusieurs tissus
non-gluconéogéniques (Gamberuai et al., 1996, Shingu et al., 1996), 4) les effets
contraires des giucocorticoides dexamethasone et triamcinolone sur les niveaux
dlARNm et de protéine Glc-6-Pase (Lange et al., 1994, Argaud et al., 1996, article 4).

Toutes ces questions nous ramènent au concept du couplage des fonctions
d'hydrolyse du Glc-6-P et d'accumulation du Glc. Ce couplage fonctionnel ne peut être
réalisé par des entités distinctes et indépendantes lhne de l'autre, mais pourrait
nécessiter la présence d'une ou de plusieures protéines auxiliaires. La preuve la plus
flagrante est apportée par les microsomes isolés de souris où le gène de la sous-unité
catalytique de la GIc-6-Pase est rendu non-fonctionnel Pei et al., 1996). Dans ce
modèle, non seulement l'hydrolyse du Gic-6-P a été annulée, mais on n'observe pas
d'accumulation de Glc non plus à l'intérieur des vésicules (Lei et al., 1996). Ce résultat
nous indique que l'hydrolyse du Glc-6-P est requise pour voir s'accumuler le Glc à
l'intérieur des rnicrosomes, conclusions déjà tirées à i'aide d'études cinétiques (Berteloot
et al., 1991a, St-Denis et ai., 1995b, Berteloot et al., 1995), ou alors que le
transporteur putatif de Glc-6-P (Tl) n'existe pas.
5.3- Le cycle GldGlc-6-P : rôle de la Glc-dPase comme
transporteur secondaire de glucose
L'hypothèse que la Glc-6-Pase puisse servir de transporteur de Glc secondaire
est basée sur trois observations majeures : 1) En présence de 0.2 mM ~- ' 'c ]G~c-~-P
dans le milieu d'incubation, Berteloot et al. (1991a) montrent que le W-~~C]GIC
s'accumule à l'intérieur de I'espace intramicrosomal contre un gradient de concentration
de Glc pouvant représenter au-dessus de 200 fois la concentration de Glc dans le milieu
extravésiculaire (10-16 mM), 2) nous démontrons, de plus, que l'accumulation de Glc à
l'intérieur des microsomes était directement proportionnelie à l'activité Glcd-Pase,

suggérant déjà que la Glc-6-Pase puisse in Mvo semir de transporteur secondaire de
Glc, et que 3) I'efflux de Glc était indépendant de l'hydrolyse du Glc-6-P, et que ce
transport était unidirectionnel (St-Denis et al., 1995b). A la lumière de ces
observations, il est tentant de suggérer que le Glc produit suite à l'hydrolyse du GIc-6-P
puisse être spécinquement orienté soit vers I'itérieur des microsornes ou déversé
directement dans le cytosol et que ce phénomène puisse avoir une implication
physiologique encore à déterminer.
Ce rôle de transporteur actif de Gic secondaire peut donc être attribue à la Glc-
6-Pase et son fonctionnement se ferait via le cycle GldGlc-6-P. Rappelons que la
production de Glc-6-P dans les cellules hépatiques requiert de I'ATP comme source
d'énergie pour l'activité de la GK, et que son hydrolyse subséquente par la Glc-6-Pase
résulterait en un transport de Glc vers l'intérieur du RE ou en sa production
directement dans le cytosol (voir figure 15). La rephosphorylation ATP-dépendante du
Glc assurerait le maintien d'une concentration de Glc-6-P et permettrait à plus de Gic
de s'accumuler à l'intérieur du RE. Un modèle de souris où le gène de GLUT 2 serait
inactivé, et où il serait possible de mesurer la production de Glc à partir de précurseurs
néoglucogéniques tels le lactate et le pynivate, aiderait à élucider le rôle que pourrait
avoir cette voie de production de Glc hépatique à travers la membrane du RE.
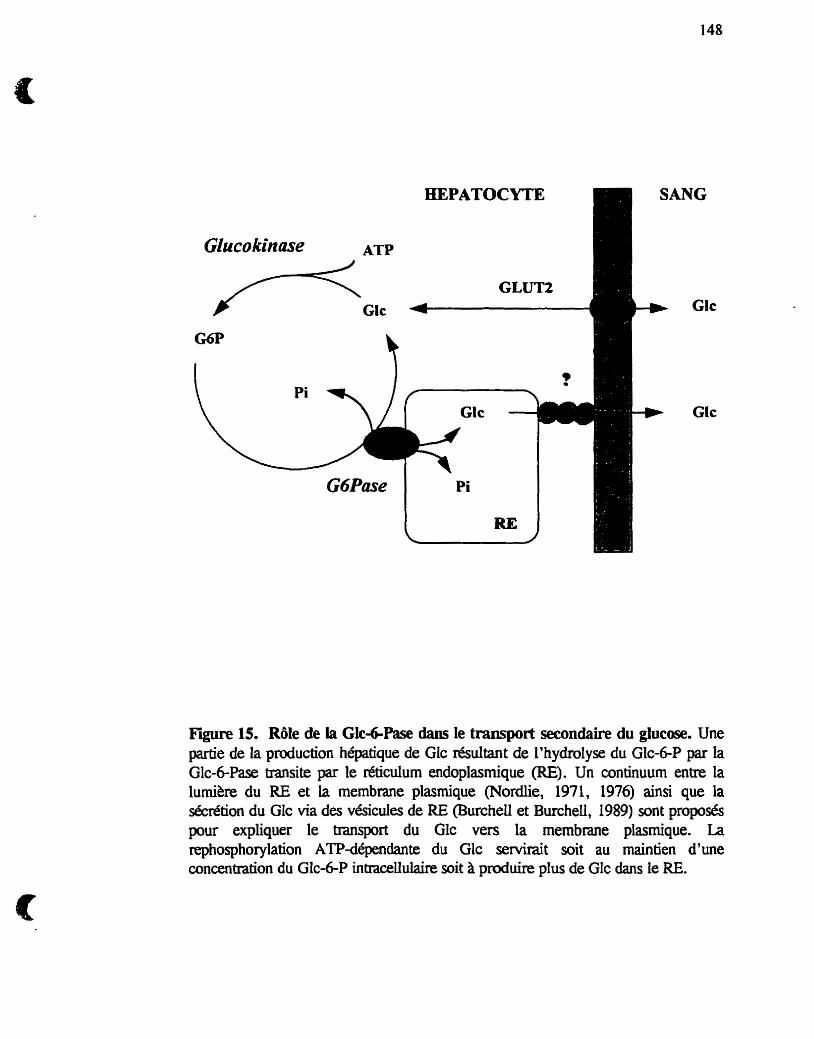
r Glc q-1
NG
Glc
Gtc
Fîre 15. Rôle de la Glc+Pase dans le transport secondaire du glucose. Une partie de la production hépatique de Glc résultant de l'hydrolyse du Glc-6P par la Glc-6-Pase transite par le réticulum endoplasrnique (RE). Un continuum entre la lurnikre du RE et la membrane plasmique (Nordlie, 1971, 1976) ainsi que la sécrétion du Glc via des vésicules de RE (Burchell et BurcheLl, 1989) sont proposés pour expliquer le transport du Glc vers la membrane plasmique. La rephosphorylation ATPdépendante du Glc servirait soit au maintien d'une concentration du Glc-6-P intraceliulaire soit A produire plus de Glc dans le RE.

Ajzannay, A, Minassian, C., Riou, J.P., and Mithieu, G. (1993) GIucose-6-phosphate phosphohydrolase of detergent-treated liver microsomal membranes exhibits a s p d c kinetic behaviour towards glucose-6-phosphate. Eur. J. Biochem. 212:335-338
Ajzannay, A, Minassian, C., Rioy J.P., and Mithie- G. (1994) Glucose-6- phosphatase specüicity after membrane solubiiization by detergent-treatment. J. Biochem. 1 16:1336-l34O
Ajzannay, A, and Mithieu, G. (1996) Glucose-6-phosphate and mannose-6- phosphate are equaiiy and more actively hydrolysed by glucose-6-phosphatase during hysteretic transition within intact microsomal membrane than after detergent-treamient. Arch. Biochem. Biophys. 326:238-242
Ananthanarayanan, M., von Dippe, P., and Levy, D. (1 988) Identification of the Na+- dependent bile acid transport using monoclonal antibodies. J. BioL Chem. 239:8388- 8343
Argaud, d., Zhang, Q., Pan, W., Maitra, S., Pilkis, S., and Lange, A.. (1996) Regulation of rat liver glucose-6-phosphatase gene expression in different nutritional and hormonal States. Diabetes 45: 1563- 1571
Arion, W. J., and Nordlie, RC. (1 964) Liver microsomal glucose-6-phosphatase, ino rganic pyrop hosp hatase and pyro p hosp hate-glucose p hosp hotramferase. J. BioL Chem. 239:2752-2757
Anon, W.J., and Nordlie, RC. (1967) Biological reylation of inorganic pyrophosphate-glucose phosphotransferase and glucose-6-phosphatase. Activation by tiamcinolone, in vivo, in the presence of actinomycin D. J. Biol. Chem. 242:2207- 2210
Arion, W.J., Wallin, B.K., Lange, AJ., and Baiias, L.M. (1975) On the involvement of a glucose-6-phosphate transport system in the fùnction of microsomal glucose-6- phosphatase. Mol. CeU. Biochem. 6: 75-83
Arion, W.J., Lange, AJ., and Bailas, L.M. (1976a) Quantitative aspects of relationship between glucose-6-phosphate transport and hydrolysis for üver rnicrosomal glucose-6- phosphatase system. J. Biol. Chem. 25 :6784-6790

Arion, W.J., Baiias, L.M., Lange, AJ., and W a h , B K (1976b) Mcrosornal membrane permeabiIity and the hepatic glucose-6-phosphatase systern. J. Biol. Chem. 25 1 :490 1-4907
Arion, W.J., Lange, AJ., Wds, H.E., and Bdas, L.M. (1980) Evidence for the participation of independent tmnslocases for phosphate and glucose &phosphate in the microsomal glucose-6-phosphatase system. Interactions of the system with orthophosphate, inorganic pyrophosphate, and cahamyl phosphate. JO BioL Chem. 255: IO396-lO406
Arion, W.J., Burchell, B., and Burchell, A (1984) Specific inactivation of the phosphihydrolase component of the hepatic microsumal gluwse-6-phosphatase system by diethyl pyrocarbonate. Biochem. L 220: 83 5-842
Ashmore, J., and Weber, G. (1959) The role of hepatic glucose-6-phosphatase in the replation of carbohydrate metabolism. Vit. H o m 17:9 1 - 132
Band, K., and Jones, C.T. (1980) Functionnal activation by glucagon of glucose-6- phosphatase and gluconeogenesis in the perfùsed tiver of the fetal guinea pig. FEBS Lett. 119: 190-194
Barash, V., Riskh, A, Shafiir, E., Waddeil, ID., and Burcheli, k (1991) Kinetic and imrnunologic evidence for the absence of glucose-6-phosphatase in early human chorionic viili and term placenta Biochem. Biophys. Acta 1073 : 16 1 - 167
Bashan, N., Potashnilg R, Peist, A, Peleg, N., Moran, A, and Moses, S.W. (1993) Deficient glucose phosphoqlation as a possible common denominator and its relation to abnormal leucocyte fùnctioq in glycogen storage disease type I b patients. Eur. J. Pediatr. lS2:S44-S48
Bazan, J.F., Retterick, R, and Piikis, S.J. (1989) Evolution o f a bifùnctional enzyme : 6-phosphofnicto-2-kinase/hctose-2,6-biphosphat. Proc Natl Acad. Sc. USA. 86:9642-9646
Beaudet, A L. (1 995) Harrison's P~ciples of Intemal Medicine, 13th Ed., New York Mc Graw-HU, 2099-2 105
Begley, P.J., and Cd?, J.A. (1981) Evidence for protein phosphorylation as a regulatory mechanism for hepatic microsomal glucose-6-phosphatase. Biochem. Biophys. Res. Commun. 103: 1029- 1034
Be& J.E.. Hume, R, Busutili, A., and Burchell, k (1993) Immunocytochemical detection of microsomal glucose-6-phosphatase in human brain astrocytes. Neuropathology and applied Neurobiology. 19:429-43 5

Berteloot, A, Vidal, H & van de Werve, G. (1991a) Rapid kinetics of liver rnimsornal gIucose-6-phosphatase. Evidence for tight couphg between giucose-6- phosphate transport and phosphohydrolase activity. J. BioL Chem. 2665497-5507
Berteloot, A, Malo, C., Breton, S., and Brunette, M. (199 1b) Fast sarnpling, rapid filtration apparatus : principal characteristics and validation from stuclies of Dglucose transport in human jujenal bnish border membrane vesicles. J. Membrane Biol. 12211 11-125.
Berteloot, A, St-Denis, J.-F., and van de Werve, G. (1995) Evidence for a membrane exchangeable glucose pool in the fùnctioning of rat liver glucose-6-phosphatase. J. Biol. Chem. 27O:2 1098-2 1 102
Bickerstaff, G.F., and BurcheU, B. (1980) Studies on the purification of glucose-6- phosphatase frorn rabbit liver rnicrosomal hction. Biochem. Soc Trans. 8339-390
Blair, J.N.R and Burchell, B. (1988) The mechanimi of histone activation of the hepatic rnicrosomal glucose-6-phosphatase system : a novel method to assay glucose- 6-phosphatase activity. Biochem. Biophys. Acta 964: 16 1- 167
Bontemps, F., Hue, L, and Hers H.G. (1978) Phosphorylation of glucose in isolated rat hepatocytes. Sigrnoidal kinetics explaineci by the activity of glucokinase aione. Biochem. J. 1 74:603-6 1 1
Burchell, A , Watkins, S.L., Hume, R (1996) Human fetal testis endoplasrnic reticulum glucose-6-phosphatase enzyme proteh. BioL Reprod. 55298-303
Burcheli, A (1994) Hepatic microsomal glucose transport. Biochem. Soc. Trans. 22:658-663
Burchell, A, and BurcheU, B. (1980) Stabilmtion of partially-punfied glucose-6- phosphatase by fluoride. 1s enzyme inactivation caused by dephosphorylation ? FEBS Lett 118: 180-184
Burcheil, A, and Burcheil, B. (1982) Identification and purification of a liver microsomal glucose-6-phosphatase. Biochem. J. 205 567-573
Burcheil, A, Burcheü, B., Arion, W.J., and Walls, H.E. (1982) A critical evaluation of the possible modulation of hepatic microsomal glucose-6-phosphatase activity by protein phosphorylation. Biochem. Biophys. Res. Commun. 107: 10464052
Burchell, A., and Cain, D.I. (1 985) Rat hepatic microsomal glucose-6-phosphatase protein Ievels are increased in streptozotocin-induced diabetes. Diabetologia 28352- 856

Burcheli, A, Waddell, I.D., Countaway, J.L., Arion, W.J., and Hume, R (1988) Identification of the human hepatic microsoma1 glucose-6-phosphatase enzyme. FEBS Lêtt. 242: 153- 156
BurcheU, A, Bell, J.E., Hume, R, and Buaittil, A (1989) Hepatic microsoma1 glucose-6-phosphatase system and sudden death syndrome. Lancet 2:29 1-294
BurcheU, A, and Burchel, B. (1989) Molecular pathologies of the hepatic endoplasmic reticulum. Cun: Opinion in Ceil BioL 1 :7 12-7 17
Burcheli, A (1990) Molecular pathology of glucose-6-phosphatase. FASEB J. 4:2978-2988
Burcheii, A, and Waddell, I.D. (1 990) Diagnosis of a novel glycogen storage disease : type laSP. J. Inher. Metab. Dis. 13 :247-249
Burcheli, A., and Gibb, L. (1991) Diagnosis of type lb and lc glycogen storage disease. J. Inher. Metab. Dis. 14:305-307
Chevalier-Porst, F., Bozon, D., Bonardot, AM., Bruni, N., Mithieux, G., Mathieu, M., and Maire, 1. (1996) Mutation analysis in 24 french patients with glycogen storage disease type l a J. Med. Genet. 33 :358-360
Christ, B., Probst, L, and Jungemann, K. (1986) Antagonistic regulation of the glucoselglucose Gphosphate cycle by insulin and glucagon in cultured hepatocytes. Biochem. J. 238: 185-191
Colliia, W., Johnson, W.T., and Nordlie, RC. (1974) The nature of modifications by various anions of synthetic and hydrolyric activities of multi-fùnctional glucose-6- phosphatase. Biochim. Biop hys. Acta. 3 64%-87
ColWa, W., Jorgenson, RA, and Nordlie, RC. (1975) Marnrnalian carbamyl phosphate:glucose phosphotransferase and glucose-6-phosphate phosphohydrolase : Extended tissue distribution. Biochern. Biophys. Acta. 377: 1 17-125
Collipp, P.J., Carsten, A, Chen, S.Y., Thomas, J., and Maddaiah, V.T. (1974) Molecular weight of microsoma1 glucose-6-phosphatase of rat and human iiver. Biochemical Medicine 1 O:3 12-3 19
Countaway, J.L., Waddeil, LD., Burcheü, A, and Arion, W.J. (1988) The p hosp ho hydrolase component of the hepatic rnicrosomal glucosed-phosp hatase system is a 36.5-kilodaiton polypeptide. J. Biot. Chem. 263:2673-2678

Emery, J.I., Howat, AC., Variend, S., and Vawter, G.F. (1988) Investigations of inbom errors of metabolism in unexpected infant deaths. Lancet 2:29-30
Efendic, S., Karlander, S.G., and Vranic, M. (1988) Mild type II diabetes markedly increases glucose cycling in the postabsorptive aate and during glucose infusion hespective of obesity. J. Clin. Invest. 8 1 : 1953- 1 96 1
Essner, E. (1973) Phosphatases. In Electron Microscopy of Enzymes : Principles and Methods, M.A Hayaî, ed. (Van Nostrand Reinhold, New York) 1:44-76
Feldman, F., and Butler, L.G. (1969) Detection and characterization of the phosphorylated fom of microsornai glucose6-phosphatase. Biochem. Biophys. Res. Commun. 3 6: 1 19- 125
Feldrnan, F., and Butler, L.G. (1972) Protein-bound phosphoryl histidine : a probable intermediate in the microsomal glucose-6-p hosp hata~~norganic pyrop hosphatase reaction. Biocbem. Biophys. Acta. 268:698-7 10
Foster, ID., Nelson, K.L., Sukalski, KA, Lucius,RW., and Nordlie, RC. (1991) Hysteretic behavior of the hepatic microsomal glucose-6-phosphatase system. Biochim. Biophys. Acta. 1 1 l8:g 1-98
Fonyth, RJ., Bartlett, K., Burchell, A., Scott, H.M. and Eyre, J.A. (1993) Astrocytic glucose-6-p hosp hatase and the pemeability of brain microsornes to glucose 6- phosphate. Biochem. J. 294: 145- 1 5 1
Fulceri, R, Bellomo, G., Gamberucci, A, Scott, KM., Burchell, A, and Benedetti, A (1992) Permeability of rat iiver microsomal membrane to &;ose 6-phosphate. Biochem. J. 286:8 13-8 17
Gambenicci, A., Marcolongo, P., Fulceri, R, Giunti, R, W a t h , S.L., Waddell., LD., Burchell, A., and Benedetti, A (1996) Low levels of giucose-6-phosphate hydrolysis in the sarcoplasmic reticulum of skeletal muscle : involvement of glucose-6-phosphatase. Molec. Membr. Biol. 13 : 103- 1 O 8
Gardner, L.B., Liu, Z., and Barrett, E.J. (1993) The role of glucose-6-phosphatase in the action of insulin on hepatic glucose production in the rat. Diabetes 42: 16 14- 1620
Garland, RC., and Cori, C.F. (1972) Separation of phospholipids fiom glucose-6- phosphatase by gel chromatography. Specificity of phospholipid reactivation. Biochernistry 1 1 :47 12-47 18
Gitzelrnann, R, and Bosshard, N.U. (1993) Defective neutrophil and monocyte fùnctions in glycogen storage disease type lb : a litterature review. Eur. J. Pediatr. 1522333438

Glueckson-Waelsch, S. (1979) Genetic control of morphogenetic and biochemical daerentiation : lethal albino deletions in the mouse. Cek 16:225-23 7
Gold, G., and W~dnell, C.C. (1976) Relationship between microsomal membrane perrneability and the inhiiition of hepatic gIucose-6-phosphatase by pyridoxal phosphate. J. BioL Chem. 254: 103 5- 104 1
Granner, D., and Pilkis, S. (1990) The genes of hepatic glucose metabolism. J. Biol. Chem. 265:10173-10176
Gunderson, H.M., and Nordlie, RC. (1975) CarbamyI phosphate:glucose phosphotransferase and glucose-&phosphate of nuclear membrane. uiterrelationships between membrane integrity, enzyme latenq, and cataiytic behavior. JO BioL Chem. 250:3552-3559
Haber, B.A, Chin, S., Chuang, E., Buikhuisen, W., Naji, A, and Taub, R (1995) High levels of glucuse-6-phosphatase gene and protein expression reflect an adaptive response in proliferating liver and diabetes. L Clin. Invest. 953332-84 1
Hanson, T.L., and Nordlie, RC. (1970) Liver microsoma1 inorganic pyrophosphate glucose phosphotderase and glucose-6-phosphatase. Effects of diabetes and Uisulin administration on kinetic parameters. Biochim. Biophys. Acta. 198:66-75
Hass, L.F., and Byme, W.L. (1 960) The mechanism of glucose-6-p hosphatase. J. Am. Chem. Soc. 821947-952
Hemnan, J.L., and Nordlie, RC. (1972) The catalytic potential of carbamyl phosphate: glucose phosphotransferase : A species distribution study. Arch. Biochem. Biophys. 152: 180- 186
W, A, Waddell, I.D., Hopwood, D., and BurcheU, A. (1989) The rnicrosomal glucose-6-phosphatase enzyme of human gall-bladder. JO Pathol. 158: 53-56
Hume, R, Voice, M., Pazouk. S., Giuntii R, Benedetti, 4 and Burchell, A (1995) The human adrenal microsornai glucose-6-phosphatase system. J. Clin. Endocr. Metab. 80: 1960-1966
Hwu, W.L., Chuang, S.C., Tsai, L.P., Chang, M.H., Chuang S.M., and Wang, T.R (1995) Glucose-6-phosphatase gene G327A mutation is cornmon in Chinese patients with glycogen storage disease type 1 a Hum Mol. Genet. 4: 10%- 1090
Johnson, W.T., and Nordlie, RC. (1980) Stimulation of gluwse-6-phosphatase by polyamines is a membrane-mediated eveb. Life Sciences 26:297-302

Jonges, G.N., van Noorden, C. J.F., and Laners, W.K (1 992) In situ kinetic parameten of glucose-6-phosphatase in the rat liver lobdus. J. BioL Chem. 267:4878488 1
Kajihara, S., Matsuhadi, S., Yamamoto, K., Kido, K., Tsuji, K., and Tanae, A (1995) Exon redehition by a point mutation within exon 5 of the glucose-6-phosphatase gene is the major cause of glycogen storage disease type la in Japan. Am. J. Hum. Geneti 57549-555
Katq J., Wals, P.A, and Rognstad, R (1978) Glucose phosphorylation, glucose& phosphatase and recycling in rat hepatocytes. J. BioL Chem. 253:453M546
Kosel, S., Matern, D., and Shiq Y.S. (1993) Purification of huinan rnicrosomal liver glucose-6-phosphatase system by afEnity chromatography and immunodetection. Eur. J. Pediatr. lS2:S22-SX
Lange, AJ., Argaud, D., El-Maghrabi, M.R, Pan, W., Maitra, SR, and Piikis, S.J. (1994) Isolation of a cDNA for the catalytic subunit of rat liver glucose-6-phosphatase : Regulation of gene expression in FA0 hepatoma cells by insuiin, dexarnethasone and CAMP. Biochem. Biophys. Res. Commun. 201 :302-309
Lavoie, L., Dimitrakoudis, D., Marette, A., A ~ a b i , B., Klip, A, Vranic, M., and van de Werve, G. (1993) Opposite effects of hyperglycemia and insulin deficiency on liver glycogen synthase phosphatase aaivity in the diabetic rat. Diabetes 42:363-366
Lei, K.-J., Shelly, L.L., Pan, C . 4 , Sidbury, J.B., and Chou, J.Y. (1993) Mutations in the glucose-6-phosphatase gene that cause glycogen storage disease type la. Science 2621580-583
Lei, K.-J., Pan, C.-J., Shelly, L.L., Liu, J.-L., and Chou, J.Y. (1994) Identification of mutations in the gene for giucose-6-phosphatase, the enzyme deficient in giycogen storage disease type la. J. Clin. Invest 93 : 1994- 1999
Lei, K.J., SheUy, L.L., Lin, B., Sidbury, J.B., Chen, Y.-T., Nordlie, RC., and Chou, J.Y. (1995) Mutations in the glucose-6-phosphatase gene are associated with glycogen storage disease types la and l aSP but not 1 b and 1 c. J. Clin. Invest. 95:234-240
Lei, K.-J., Chen, H., Pan, C.-J., Ward, J.M., Mosinger, B., Lee, E.J., Westphai., H., Mansfield, B.C., and Chou, J.Y. (1996) Glucose-6-phosphatase dependent substrate transport in the glycogen storage disease type- 1 a rnouse. Nature Genetics 13 :203-209
Liu, Z., Barrett, E.J., Dalkin, AC., Zwart, AD., and Chou, J.Y. (1994) Effect of acute diabetes on rat hepatic glucose-6-phosphatase activity and its messenger RNA level. Biochem. Biophys. Res. Commun. 205680-686
Lowe, G., and Potter, B.V.L. (1982) The stereochernical course of phosphoryl transfer cataiysed by glucose-6-phosphatase. Biochem. J. 20 1 :665-668

Lucius, RW., Waddeli, LD., Burcheii, A, and Nordlie, RC. (1993) The hepatic glucose-6-p hosphatase system in EMich-ascites-tumow-bea~g rnice. Biochem. J. 290:907-911
Lueck, J.D., Herman, J.L., and Nordlie, RC. (1972) General kinetic mechanism of microsod carbamyl phosphate:glucose phosphoaansferase, giucose-6-phosphatase, and other associateci activities. Biochemistry 1 1 :2792-2799
Lygre, DG., and Nordlie, RC. (1 968) Phosphohydrolase and phosphotrannerase activities of intestinal glucose-6-phosphatase. Biochemisûy 7:32 19-3226
Maddaiah, V.T., Chen, S.Y., Rezvani, I., Sharrna, R, and Coliipp, P.J. (1971) Inactivation of microsorna1 glucose-6-phosphatase by pyridoxal-S'-phosphate. Biochem. Biop hys. Res. Comm. 43 : 1 14- 1 18
Massillon, D., Barziiai, N., Chen, W., Hu, M., and Rossetti, L. (1996) Glucose regulates in vivo glucod-phosphatase gene expression in the iiver of diabetic rats. J Biol. Chem. 27 1:9871-9874
Matschinsky, F.M. (1996) A lesson in metabolic regulation inspired by the glucokinase glucose sensor paradigm. Diabetes 45:223-24 1
Minassian, C., and Mithieux, G. (1994) Differential tirne-course of liver and kidney glucose-6-phosphatase activity during f h g in rats. Corn p. Biochem. Physioi. 109B199-104
Minassian, C., A j m a y , A., Riou, J.P., and Mithieux, G. (1994) Investigation of the mechanism of glycogen rebound in the b e r of 72 h-fàsted rats. J. BioL Chem. 265: 16585-16588
Minassian, C., Daniele, N., Bordet, J.C., Zitoun, C., and Mithieu, G. (1995) Liver glucose-6-phosphatase activity is inhibited by refeeding in rats. J. Nutr. 125:2727- 273 2
Minassian, C., Zitoun, C., and Mithieux, G. (1996) DEerential the-course of liver and kidney glucose-6-phosphatase activity during f h g in rats correlates with differentiai time-course of messenger RNA level. MOL CeU. Biochem. 155:37-4 1
Mithiey G., A j m a y , A, and Minassian, C. (1995) Identification of membrane- bound phosphogiucomutase and glucose-6-phosphatase by 32~-labeling of rat liver microsoma1 membrane proteins with n~-glucose-6-phosphate. J. Biochem. 1 17:908- 914

Mithieuq G., and Zitoun, C. (1 9%) Mechanisms by which Eitty acyl-CoA esters inhibit or activate glucose+6-phosphatase in intact and detergent-treated rat liver microsomes. Eur. J. Biochem. 2351799-803
Mthiewq G., Vidal, H., Zitoun, C., Bruni, N., Daniele, N., and Minassian, C. (1996) Glucose-6-phosphatase mRNA and activity are increased to the sarne extent in kidney and liver of diabetic rats. Diabetes 45:89 1-896
Narisawa, K., Igarashi, Y., ûtomo, H., and Tada, K. (1978) Anew variant of glycogen storage disease type 1 probably due to a defect in the glucose-6-phosphate transport systern. Biochem. Biophys. Res. Commun. 83 : 1360- 1364
Nelson-Rossow, K.L., Sukalsii, KA, and Nordlie, RC. (1993) Hysteresis at near- physiologie substrate concentrations underlies apparent sigmoid kinetics of the glucose- 6-phosphatase systern. Biochim. Biophys. Acta. 1 163 :297-302
Ness, G.C., Sukaisla, K.A, Sample, C.E., Pendleton, L.C., McCreery, MJ., and Nordlie, RC. (1989) Radiation inactivation analysis of rat iiver rnicrosomai glucose& phosphatase. J. Biol. Chem. 26471 1 1-71 14
Newgard, C.B., Foster, W., and McGarry, J.D. (1984) Evidence for suppression of hepatic glucose-6-phosphatase with carbohydrate feeding. Dia betes 3 3 : 1 92- 1 95
Nordlie, RC., and Arion, W. J. (1 964) Evidence for the cornmon identity of glucose-6- phosphatase, inorganic pyrophosphatase, and pyrophosphate-glucose phosphotradérase. J. Biol. Chem. 239: 1680-1685
Nordlie, RC., Arion, W.J., and Glends, E.A. (1965) Liver microsomal glucose-6- p hosp hatase, inorganic pyro phosphatase, and pyrophosp hate-glucose phosphotransferase. Effects of adrenalectomy and cortisone administration on activities assayed in the absence and presence of deoxycholate. J. Biol. Chem. 240:3479-3484
Nordlie, RC., and Lygre, D.G. (1966) The inhibition by citrate of inorganic pyrop hosphate-glucose p hosphotransferase and glucose-6-p hosphatase. J. Biol. Chem. Xl:3 136-3 141
Nordlie, RC., and Snoke, RE. (1967) Regulation of liver microsomal inorganic pyrophosphate-glucose phosphotransferase, glucose-6-phosphatase, and inorganic pyrophosphatase. Biochirn. Biophys Acta. 148:222-232
Nordlie, RC., Arion, W.J., Hanson, T.L., Gilsdorf; J.R, and Home, R.N. (1968) Biological regulation of liver microsoma1 inorganic pyrophosphate-glucose phosphotransferase, glucose-6-phosphatase, and inorganic pyrophosphatase. Differential effects of fâsting on synthetic and hydrolytic activities. J. Biol. Chem. 243:lI40-1146

Nordlie, RC. (1971) Gluwse-6-phosphatase, hydrolytic and synthetic activities. In The Enzymes: Boyer P.D., Ed. ,3rd ed. 4543609
Nordlie, RC. (1974) Metabolic regulation by multifunctional glucose-6-phosphatase. Curr. Top. CeU ReguL 8:33-117
Nordlie, RC. (1976) Gluwse-6-phosphatase phosphotransferase : roles and regulation in relation to gluconeogenesis. In Giuconeogenesis : Its regulation in mamrnaiian species. RW. Hanson, ed. (John Wiley and Sons, New York) 93- 152
Nordlie, RC., and Jorgenson, RA (1976) Glucose-6-phosphatase. In The Enzymes of Biologicai Membranes. Martonosi eù. (Plenum Press, New York) 2:465-49 1
Nordlie, RC. (1979) Multifùnctionai giucose-6-phosphatase. In Cellular Biology (Life Sciences) 24: 23 97-2404
Nordlie, RC., Johnson, W.T., Comatzer, W.E. and Twedel, G. W. (1 979) Stimulation by polyamines of carbamyl p hosphate : glucose phosphotderase and glucose-6- phosphate phosphohydrolase activities of multifunctional glucose-6-phosphatase. Biochem. Biophys. Acta. 585: 12-23
Nordlie, RC. (1982) Kinetic examination of enzyme mechanisms involving branched reaaion pathways-A detailed consideration of multifunctïonal glucose-6-phosphatase. Methods in Enzymology 87:3 19-3 53
Nordlie, RC., Sukaiski, K A , Munoz, J.M., and Baldwin, J.J. (1983) Type Ic, a novel glycogenosis. Underlying mechanism. J. BioL Chem. 258:9739-9744
Nordlie, RC. and Sukalski, K. A (1 985) Multifùnctional ducose-6-phosphatase : a critical review, in The Enzyme of Biologieal Membranes 2"6 ed. (ed Martonosi, AN.), 2:349-398 (Plenum Publishing Corp., New York)
Nordlie, RC., Scott, H.M., Waddell, I.D., Hume, R, and Burcheli, A (1 992) Analysis of human hepatic microsomal glucose-6-phosphatase in conditions where the T2 pyrophosphatdphosphate transport protein is absent. Biochem. J. 28 1 2359-863
Nordlie, M A , Pederson, B.A., Foster, J.D., and Nordlie, R.C. (1 993) Effkcts of ionic strength on hydrolytic and synthetic activities of glucose-6-phosphatase of intact and disrupted rat liver microsomes. FASEB S. 7:A849
Owen, D.E., Felig, P., Morgan, AP., Wahren, J., and Cahill, G.F. (1969) Liver and kidney metabolism during prolonged starvation. J. Clin. Invest. 48574583

Parmi, R, Moses, S., Henhkovia Es, Carmi, R, and Bashan, N. (1995) Characterizabon of the mutations in the giucose-6-phosphatase gene in Israeli patients with glycogen storage disease type la : R83C in six Jews and a novel V166G mutation in a Muslim Arab. Je Inher. Metab. Dis. 18:Z 1-27
Pears, J., Jung, RT., Browning, MC-K., Taylor, R, Burchell, A (1991) Reactive hypoglycemia in association with disordered islet hction and abnomal hepatic gluco&phosphatase activity : response to diazoxide. Diabetic M d 8:268-27 1
Pears, J., Jung, RT., Jankowski, J., Waddeil, LD., and Burcheü, A (1 992) Glucose-6- phosphatase in normal adult human intestinal mucosa. Clinid Science 83:683-687
Pontremoli, S., MeUoni, E., Salamino, F., De Fiora, A, and Horecker, B.L. (1974) Changes in activity and molecular properties of hctose-1,6-bisphosphatase d u ~ g fasting and refeedig. Pmc Nat . Acad. S c i USA 7 1 : 1776- 1779
Proctor, E.B., and Nordlie, RC. (1972) Responses of synthetic and hydrolytic activities of rat liver glucose-6-phosphatase to concurrent cortisone administration and experirnental diabetes. Arch. Int. PhysioL Biochim. 75:46 1-467
Reczek P.R, and Viee, C.A. (1982) A pudication of microsomal glucose-6- phosphatase ftom human tissue. Biochem. Biophys. Res. Commun. 107: 1 158- 1 165
Romanelli, A, St-Denis, J.-F., Vidai, H., Tchu, S., and van de Werve, G. (1994) Absence of glucose uptake by liver microsornes : an explmation for the complete latency of glucose dehydrogenase. Biochem. Biophys. Res. Commun. 200: 149 1- 1497
R m B., and de Groot, H. (1988) Partial purification of rat liver microsomal glucose-6-phosphatase on hydroxylapatite. BioL Chem. 369: 1 15- 12 1
Sagné, C., IsambeIt, M.-F., Henry, J.-P., and Gasnier, B. (1996) SDS-resistant aggregation of membrane proteins : application to the purification of the vesicular monoamine transporter. Biochem. J. 3 16:825-83 1
Schaub, J., and Heyne, K. (1983) Glycogen storage disease type Ib. Eur. J. Pediatr. 140:283-288
Schrnoü, D., Man, B.B., and Burchell, A (1996) Cloning and sequencing of the 5' region of the human glucose-6-phosphatase gene : transcriptional regdation by CAMP, insulin and glucocorticoids in the H4IIE hepatoma cells. FEBS Letters 383 :63-66
Schuize, H.-W., and Speth, M. (1980) Investigations on the possible involvement of phospholipids in the glucose-&phosphate transport system of rat-liver microsomal glucose-6-phosphatase. Eur. J. Biochem. 1 O6:505-5 14

Schulze, H-U., Nolte, B., and Kannler, R (1986) Evidence for changes in the confornational status of rat Liver microsomal glucose-6-phosphate : phosphohydrolase during detergent-dependent membrane modification. Eff& of prnerainbenzoate and organomercurïal agarose gel on glucose-6-phosphatase of native and detergent- modified microsomes. J. BioL Chem. 26 1 : 1657 1 - 16578
Segai, HL. (1959) Some consequence of the non-competitive inhibition by glucose of rat liver glucose-6-phosphatase. J. Am. Chem. Soc 8 1 :40474050
Sheily, L.L., Lei, K.-J., Pan, C.-J., Sakata, S.F., Ruppert, S., Schua G., and Chou, J.Y. (1 993) Isolation of the gene for glucosed-phosphatase, the enzyme deficient in glycogen storage disease type l a Je BioL Chem. 268:2 1482-2 1485
Shi, Z.Q., Giacca, A, Fisher, S., Vidal, H, van de Werve, G., and Vranic, M. (1994) importance of substrate changes in the decrease of hepatic glucose cyciing during k l i n infision and declining glycemia in the depancreatized dog. Diabetes 43: 1284- 1290
Shingu, R, Nakajima, H., Horikawa, Y., Harnaguchi, T., Yarna* T., Myagawa, J.- L, Narnba, M., Hanatiisa, T., and Matsuzawa, Y. (1993) Expression and distribution of glucose-&phosphatase catalytic subunit messager RNA and its changes in the diabetic state. Moleç Pathol. Pharm. 93 : 13-24
Singh, J., Martin, RE., and Nordlie, RC. (1983) Glucose-6-phosphatase : is activity regulated by phosphorylation-dephosphorytation ? Can. J. Biochem. CeU. Biol. 61: 1085-1089
St-Denis, J.-F., Comte, B., Nguyen, D.K., Seidman, E., Paradis, K., Uvy, E. and van de Werve, G. (1994) A confornational mode1 for the human iiver microsomal glucose- 6-phosphatase system : Evidence for rapid hetics and defects in giycogen storage disease type 1. J. Clin. Endocrinol. Metab. 79:955-959
St-Denis, J.-F., Annabi, B., Khoury, H., and van de Werve, G. (199Sa) Histone II-A stimulates glucose-6-phosphatase and reveals mannose-6-phosphatase adivities without permeabiiiition of liver microsomes. Biochem. J. 3 10:22 1-224
St-Denis, J.-F., Berteloot, A Vidal, H., Annabi, B., and van de Werve, G. (1995b) Glucose transport and glucose &phosphate hydrolysis in intact rat iiver microsomes. J. Biol. Chem. 270:2 1092-2 1097
Speth, M., and Schuize, HAJ. (1981) Hormone-induced effects on the rat liver microsoma1 glucose-&phosphatase system in vitro. Biochem. Biophys. Res. Commun. 99: 134-141
Speth, M., and Schulze, H.-U. (1992) The purification of a detergent-soluble glucose- 6-phosphatase fiom rat iiver. Eur. J. Biochem. 208:643-650

Stetten, M.R (1965) Metabotism of inorganic pyrophosphate. J. BioL Chem. 240 Z48-2253
Stetten, M.R, and Bumett, F.F. (1967) Some properties of variously activateci microsomal g l u c o ~ p h o s p hatase, inorganic pyrophosphatase and inorganic pyrophosphate-glucose phosphotransferase. Shift in pH optimum. Biochem. Biophys. Acta 139: 138-147
Sukalski, K.A, and Nordlie, RC. (1989) Glucose-6-phosphatase : Two concepts of structure-funaion in relationships. In : Meister 4 Ed. Advances in Eruymology and Related Topics 62:93-117
Sundberg, L., and Porath, J. (1974) Preparation of adsorbents for biospecific attinity chromatography. Attachent of groupcontaining ligands to insoluble polymers by means of bfinctional oxiranes. J. Chromatogr. 9037-98
Traxinger, RR, and Nordlie, RC. (1990) Hormonal responses of glucose-& phosphatase catalytic unit studied by stopped-flow anaiysis. Biochem. CeU. BioL 681454458
van de Werve, G., and Jeanrenaud, B. (1987) Glycogen metabolism : an o v e ~ e w . In : Diabetes/MetaboIism Reviews (RA De Fronzo, ed.), John Wiley and Sons, New York, 3:47-78
van de Werve, G., Annabi, B., Berteloot, A, St-Denis, J.-F., and Vidal, H. (1993) Le système glucose-6-phosphatase hépatique : composantes, propriétés cinétiques, régulation et déficit. Méâecine/Sciences 9: 577-582
Vidai, H., Berteloot, A, Lame, M.-J., St-Denis, J.-F. and van de Werve, G. (1992) Interaction of mannose-6-phosphate with the hysteretic transition in glucose-6- phosphate hydrolysis in intact liver microsomes. FEBS Lett. 302: 197-200
Waddell, I.D., and Burcheli, A (1988) The microsoma1 glucose-6-phosphatase enzyme of pancreatic islets. Biochem. J. 2SS:47 1-476
Waddeii, I.D., Lindsay, J.G., and Burcheii, A (1988) The identification of Tz; the phosphatdpyrophosphate transport protein of the hepatic microsomai glucose-6- phosphatase system. FEBS Letters 229: 179- 182
Waddeli, I.D., and Burchell, A (1991) Transverse topology of glucose-6-phosphatase in rat hepatic endoplasmic reticulum. Biochem. 5. 275: 133-1 37
Waddeli, LD., Scott, H., Grant, A, and Burcheil, A (1991) Identification and characterization of a hepatic rniaosomal glucose transport protein. T3 of the glucose- 6-phosphatase system ? Biochem. J. 275:363-3 67

Waddeii, I.D., Zomerschoe, AG., Voice, M.W., Burcheli, A (1992) Cloning and expression of a hepatic microsornai glucose transport protein. Cornparison with iiver plasmamembrane glucose-transport protein GLUT2. Biochem. JO 286: 173- 177
W a h , B.K. and Arion, W.J. (1973) Evaiuation of the rate-detereg steps and the relative magnitude of the individuai rate constants for the hydrolytic and synthetic activities of the catalytic components of Liver microsorna1 glucose-6-phosphatase. JO Biol. Chem. 248:Z 80-23 86
Zakim, D., and Dannenberg, A (1 990) Thermal stability of microsoma1 glucose-6- phosphatase. J. BioL Chem. 265:2O 1-208
Zoccoli. M.A, Hoopes, RR, and Karnovsky, M.L. (1982) Identification of a rat Iiver microsomd polypeptide involved in the transport of glucose &phosphate. J. Biol. Chem. 257:39 19-3924
TEST TARGET-(QA-CS)
APPLIED IMAGE. lnc - = 1653 Easî Main Street - -. - - Rochester, NY 14609 USA -- -- - - Phone: 71ff482-0300 -- -- - - Fa: 71 6î288-5989
O 1993, App(ied Image. lnc., Nt Rtglrts Reseffed





























