Embed Size (px)
Citation preview

J . Sci. Food Agric. 1988.42, 109-1 18
Uptake of Small Neutral Peptides by Mixed Rumen Microorganisms In Vitro*
Glen A. Broderick
US Dairy Forage Research Center, Agricultural Research Service, US Department of Agriculture, Madison, WI 53706, USA
R. John Wallace and Nest McKain
Rowett Research Institute, Bucksburn, Aberdeen AB2 9SB, UK
(Received 23 March 1987; accepted 5 June 1987)
A BS T R A CT
The metabolism of a range of neutral di- and oligopeptides and peptide p-nitroanilides was investigated in strained ruminal fluid diluted with anaerobic buffer. Peptide uptake was assayed by decrease in fluorescamine-reactive material in extracellular fluid, and peptide p-nitroanilide hydrolysis by diazotisation of the released p-nitroaniline. Addition of glucose and dithiothreitol did not alter uptake of di- or trialanine. Calculated V,, for di- and trialanine uptake were 1-4 and 1.9 nmol min-I mg DM-I, corresponding K , were 0.30 and 0.14 mmol litre-'. Dipeptide uptake was 0.72 to 0.90 nmol min-I mg DM-I, except f o r glycylproline, which was taken up at 0-43 nmol min-1 mg DM-I . There was a wider range for tripeptide uptake (0-5 to 1.6 nmol min-I mg DM-I) , with trialanine being taken up most rapidly. Tetra- and pentaalanine were removed at 0.93 and 0.71 nmol min-I mg DM-I . Uptake of amino acid residues as alanine oligopeptides was two to three times more rapid than uptake as dialanine. These data suggest that peptides would accumulate in rumen fluid during hydrolysis of rapidly degraded proteins, but peptide uptake would exceed rate of release f rom more slowly degraded proteins. Rates of hydrolysis of eight peptide p-nitroanilides were 0.56 to 1.53 nmol min-I mg DM-I, and
*Mention of commercial products or sources in this paper does not constitute endorsement by the US Department of Agriculture or the Agricultural Research Service.
109
J . Sci. Food Agric. 0022-5142/88/$03.50 0 Society of Chemical Industry, 1988. Printed in Great Britain

110 G. A. Broderick, R . J . Wallace, N . McKain
were of similar magnitude to uptake of di- and tripeptides; proline-contain- ing p-nitroanilides were also more slowly metabolised.
Key words: Peptide metabolism, rumen microorganisms.
1 INTRODUCTION
Protein breakdown by rumen microorganisms leads to the production of ammonia concentrations in rumen contents which frequently exceed microbial growth requirements. The excess ammonia is absorbed through the rumen wall and eventually excreted as urea, thus decreasing the efficiency of nitrogen utilisa- tion by the animal.’.2 Peptides are intermediates in the conversion of protein to ammonia, and there is some in v i t r ~ ~ . ~ and in vivo5 evidence that peptide hydro- lysis may be the rate-limiting step in the process, at least for rapidly degraded proteins. It is possible that catabolism of protein may be effectively arrested at this step.
Rumen bacteria show a preference for peptides over free amino acids as growth substrates,”8 and amino acids in peptides are more rapidly metabolised than the corresponding free amino WrightL2 demonstrated that ruminal pep- tidase activity was cell-associated, and Bladen etal. l 3 concluded from their survey of rumen bacteria that Bacteroides ruminicola was the most significant producer of ammonia from mixed peptides. This organism can apparently assimilate pep- tides of molecular weight (MW) up to 2000.10 Other bacteria involved in produc- ing ammonia from peptides were Sefenomonas ruminantium, Megasphaera efsdenii and some Butyrivibrio is01ates.l~ The mechanisms of peptide uptake and hydrolysis are fairly well understood in many organisms. l4I7 For example, Escherichia coli is known to have three permeases for peptide transport across the cell mernbrane.l6 However, the mechanisms of peptide uptake and hydrolysis have been little studied in rumen organisms.
The purpose of the research reported here was to investigate the rates of uptake of small neutral peptides by mixed rumen microorganisms. A brief report of some of this work has been made.18
2 EXPERIMENTAL
2.1 Animal, diet and sample preparation
A mature wether sheep, fitted with a permanent rumen cannula, was fed 450 g diet each at 0800 and 1600 h. The diet consisted of 67% grass hay and 33% concentrate.lY Strained rumen fluid (SRF) was prepared about 2 h after the morning feeding by straining rumen contents through four layers of muslin. Cell- free supernatant was prepared by centrifuging SRF at 27200Xg for 15 min.
2.2 Incubation procedure
SRF was diluted with 0.33 vol of 100 mmol litre-’ potassium phosphate buffer, pH 7.0, previously made anaerobic by boiling and gassing with 0,-free CO, and at

Rumen peptide metabolkm 111
39°C. Tubes were capped, mixed by inversion, and incubated at 39°C for 15 to 20 min. Peptide incubations were terminated by centrifuging tubes (12 OOOXg, 2 min). Supernatants were stored on ice until analysed for remaining peptide; peptide disappearance was negligible under these conditions. Separate 0-h sam- ples were prepared in a similar manner except that blanks and peptide-containing tubes were held on ice; iced inoculum was added to tubes before mixing and centrifuging as described. p-Nitroanilide incubations were terminated by the addition of 0.25 ml of 250 g litre-' trichloroacetic acid and centrifuging (12000xg, 2 min). Duplicates of each blank and sample type were run; results are the means from incubations with three or four separate samples of SRF taken on different days.
2.3 Analyses
The dry matter (DM) content of inocula was estimated in quadruplicate by centrifuging (272OOxg, 4"C, 15 min) and drying the pellet at 105°C to constant weight (about 6 h). This was assumed to be DM from mixed rumen microorga- nisms. The protein content of the pellet was determined using a modified Lowry procedure with bovine serum albumin as standard.20
Peptides were assayed by a fluorimetric procedure similar to those described by Perrett et d.*l and Nisbet and Payne,22 in which fluorescamine reacts preferen- tially with peptides rather than free amino acids. The response of individual amino acids in the procedure is only 1 to 5% that of peptides. Supernatant (75 pl) was added to 2-25 ml of 0.2 mol litre-' sodium citrate buffer, pH 6.2, at room temperature. Fluorescamine solution (0.28 g litre-' in acetone) was added while this solution was vortexed. After mixing for a further 3-4 s, fluorescence was measured within 2 to 30 min using a Baird Nova spectrofluorimeter (Baird- Atomic Ltd, Braintree, UK), with excitation at 390 nm and emission at 485 nm. Although the fluorescence response differed markedly between peptides, each gave a linear response. Net peptide disappearance (i.e. blank-corrected) was determined in each experiment relative to net 0-h fluorescence.
Hydrolysis of peptide p-nitroanilides was measured from the rate of appearance of p-nitroaniline using the diazotisation procedure of A p ~ e 1 . ~ ~
2.4 Materials
Peptides and peptide p-nitroanilides were obtained from Sigma Chemical Co. Amino acids present in peptides were all of the L-configuration.
2.5 Statistical procedures
Data were analy~ed2~ by one-way analysis of variance. Where significant treat- ment effects were detected (P<0-05), mean separation was by least significant difference.
3 RESULTS
3.1 Factors influencing peptide uptake
The rates of uptake of different peptides varied from day to day in inocula prepared from SRF taken from the same sheep; however, several general proper-
112 G. A . Broderick, R . J . Wallace, N . McKain
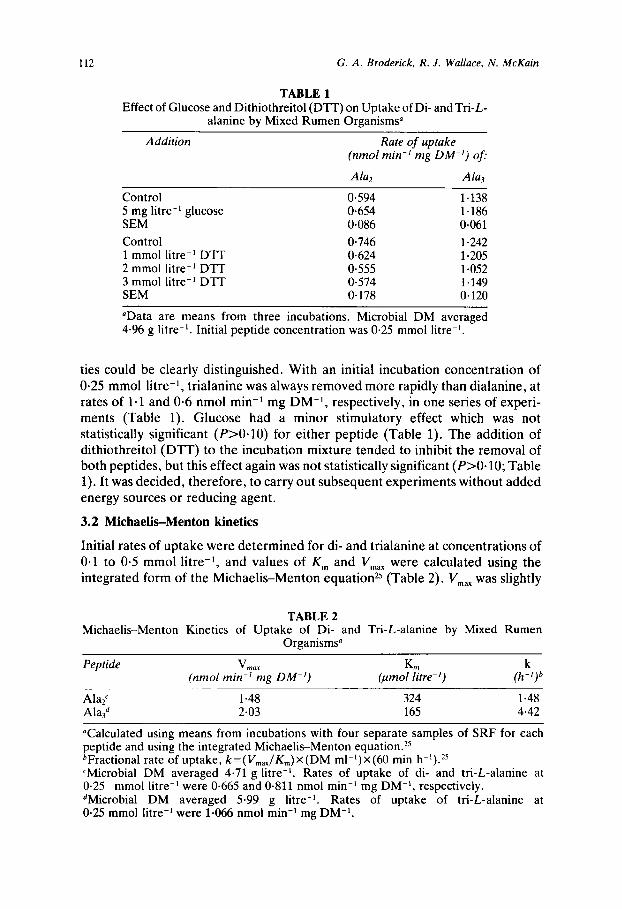
TABLE 1 Effect of Glucose and Dithiothreitol (DTT) on Uptake of Di- and Tri-L-
alanine by Mixed Rumen Organisms"
Addition Rate of uptake (nmol min-' mg DM-') of:
Control 5 mg litre-' glucose SEM Control 1 mmol litre-' DTT 2 mmol litre-' DTT 3 mmol litre-' DTT SEM
0.594 0.654 0.086 0.746 0.624 0.555 0.574 0.178
Ala3
1.138 1.186 0.061 1.242 1.205 1.052 1.149 0.120
'Data are means from three incubations. Microbial DM averaged 4.96 g litre-'. Initial peptide concentration was 0.25 mmol litre-'.
ties could be clearly distinguished. With an initial incubation concentration of 0.25 mmol litre-', trialanine was always removed more rapidly than dialanine, at rates of 1-1 and 0.6 nmol min-l mg DM-l, respectively, in one series of experi- ments (Table 1). Glucose had a minor stimulatory effect which was not statistically significant (P>O.lO) for either peptide (Table 1). The addition of dithiothreitol (DTT) to the incubation mixture tended to inhibit the removal of both peptides, but this effect again was not statistically significant (P>O. 10; Table 1). It was decided, therefore, to carry out subsequent experiments without added energy sources or reducing agent.
3.2 Michaelis-Menton kinetics
Initial rates of uptake were determined for di- and trialanine at concentrations of 0.3 to 0.5 mmol litre-', and values of K , and V,,, were calculated using the integrated form of the Michaelis-Menton equationZS (Table 2). V,,, was slightly
TABLE 2 Michaelis-Menton Kinetics of Uptake of Di- and Tri-L-alanine by Mixed Rumen
Organisms"
Peptide Vm, K m k (nmol min-I mg DM-') (pmol litre-') (h-')b
Alazc 1.48 324 1.48 Ala3d 2.03 165 4.42
"Calculated using means from incubations with four separate samples of SRF for each peptide and using the integrated Michaelis-Menton equation.2s *Fractional rate of uptake, k=(V,,,/K,,,)x(DM ml-')X(60 min h-').l5 'Microbial DM averaged 4.71 g litre-'. Rates of uptake of di- and tri-l-alanine at 0.25 mmol litre-' were 0.665 and 0.811 nmol min-' mg DM-', respectively. dMicrobial DM averaged 5.99 g litre-'. Rates of uptake of tri-l-alanine at 0.25 mmol litre-' were 1.066 nmol min-I mg DM-I.
Rumen peptide metabolism 113
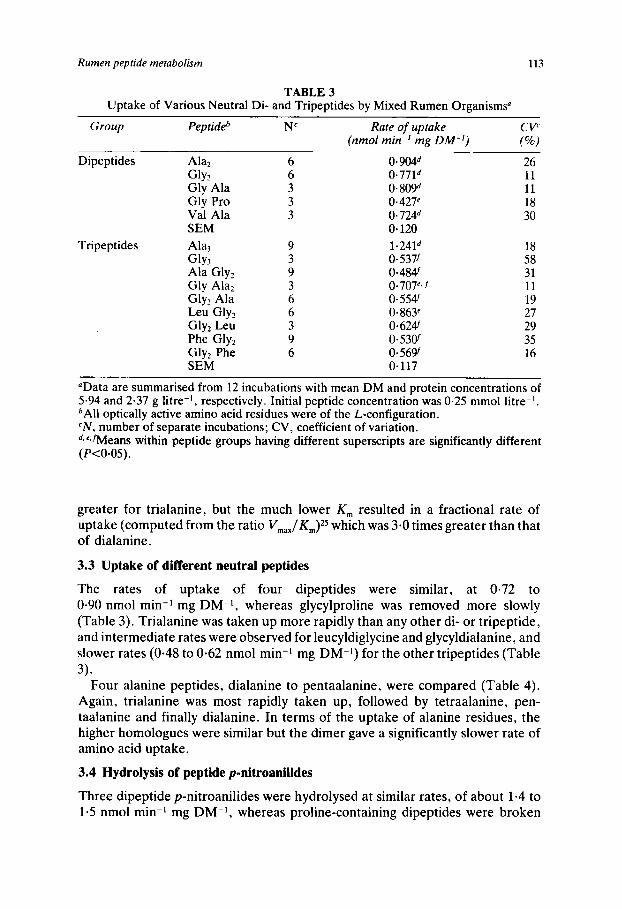
TABLE 3 Uptake of Various Neutral Di- and Tripeptides by Mixed Rumen Organisms"
Group Peptideb N' Rate of uptake CV' (nmol min-I mg DM-I) f%)
Dipeptides Alaz 6 0.904' 26 GlY, 6 0.771' 11 Gly Ala 3 0.809' 11 Gly Pro 3 0.427' 18 Val Ala 3 0.724' 30 SEM 0.120
Tripeptides A h 3 9 1.241d 18 GlY3 3 0.537f 58 Ala Gly? 9 0.484' 31
Glyz Ala 6 0.554' 19 Leu Glyz 6 0.863' 27 Gly, Leu 3 0.624 29
Gly, Phe 6 0.569f 16 SEM 0.117
Gly Alaz 3 0.707'. f 11
Phe Gly, 9 0.53W 35
nData are summarised from 12 incubations with mean DM and protein concentrations of 5.94 and 2-37 g litre-', respectively. Initial peptide concentration was 0.25 mmol litre-'. bAll optically active amino acid residues were of the L-configuration. ' N , number of separate incubations; CV, coefficient of variation. d* eJMeans within peptide groups having different superscripts are significantly different ( P t 0 . 0 5 ) .
greater for trialanine, but the much lower K , resulted in a fractional rate of uptake (computed from the ratio Vma,/K,,,)25 which was 3.0 times greater than that of dialanine.
3.3 Uptake of different neutral peptides
The rates of uptake of four dipeptides were similar, at 0.72 to 0.90 nmol min-l mg DM-I, whereas glycylproline was removed more slowly (Table 3). Trialanine was taken up more rapidly than any other di- or tripeptide, and intermediate rates were observed for leucyldiglycine and glycyldialanine, and slower rates (0.48 to 0.62 nmol min-I mg DM-I) for the other tripeptides (Table
Four alanine peptides, dialanine to pentaalanine, were compared (Table 4). Again, trialanine was most rapidly taken up, followed by tetraalanine, pen- taalanine and finally dialanine. In terms of the uptake of alanine residues, the higher homologues were similar but the dimer gave a significantly slower rate of amino acid uptake.
3.4 Hydrolysis of peptide p-nitroanilides
Three dipeptide p-nitroanilides were hydrolysed at similar rates, of about 1.4 to 1.5 nmol min-l mg DM-I, whereas proline-containing dipeptides were broken
3).
114 G. A. Broderick, R. J . Wallace, N . McKain
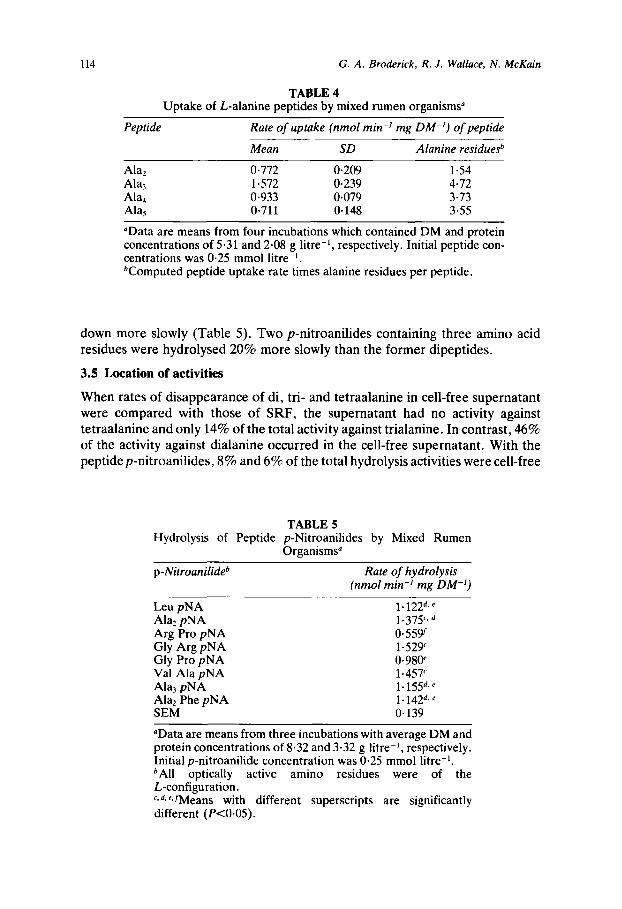
TABLE 4 Uptake of L-alanine peptides by mixed rumen organisms"
Peptide Rate of uptake (nmol min-' mg DM-I) of peptide
Mean SD Alanine residuesb
Alaz Ala3 Ala, AlaS
0.772 0.209 1.572 0.239 0.933 0.079 0.711 0.148
1.54 4.72 3.73 3.55
"Data are means from four incubations which contained DM and protein concentrations of 5.31 and 2.08 g litre-', respectively. Initial peptide con- centrations was 0.25 mmol litre-I. bComputed peptide uptake rate times alanine residues per peptide.
down more slowly (Table 5). Two p-nitroanilides containing three amino acid residues were hydrolysed 20% more slowly than the former dipeptides.
3.5 Location of activities
When rates of disappearance of di, tri- and tetraalanine in cell-free supernatant were compared with those of SRF, the supernatant had no activity against tetraalanine and only 14% of the total activity against trialanine. In contrast, 46% of the activity against dialanine occurred in the cell-free supernatant. With the peptide p-nitroanilides, 8% and 6% of the total hydrolysis activities were cell-free
TABLE 5 Hydrolysis of Peptide p-Nitroanilides by Mixed Rumen
Organisms"
p-Nitroanilideb Rate of hydrolysis (nmol min-' mg DM-I)
Leu pNA Ala2 pNA Arg Pro pNA Gly Arg pNA Gly Pro pNA Val Ala pNA Ala, pNA Alaz Phe pNA SEM
1.122d. 1.375'. 0.559 1.529' 0*98@ 1.457' 1.155" 1.142*, ' 0.139
"Data are means from three incubations with average DM and protein concentrations of 8.32 and 3.32 g Iitre-I, respectively. Initial p-nitroanilide concentration was 0.25 mmol litre-I. bAll optically active amino residues were of the L-configuration. r , d r 'JMeans with different superscripts are significantly different (PcO.05).

Rumen peptide metabolism 115
with the di- and the trialanine p-nitroanilides, respectively. The monoalanine derivative was also incubated with both preparations, and was hydrolysed at a rate of 0.74 nmol min-l mg DM-l, of which 0-01 nmol min-I mg DM-1 equivalent was cell-free.
4 DISCUSSION
Until now, only relatively poorly defined mixed peptides have been used to study peptide metabolism in rumen microorganisms. The main feature of the present paper is the description of the metabolism of single, defined peptides by the mixed rumen population, an area of metabolism which has received considerable atten- tion in other microbial systems. 14,15-17,26 Although the results have not proved to be unusual or surprising in the light of this existing knowledge, some interesting features have emerged.
As was found by Wright12 with 14C-labelled peptides from a Chlorella hydroly- sate, peptides were metabolised predominantly by cell-bound activities. The exception was dialanine, which was metabolised by cell-free rumen liquor at nearly half the rate of intact SRF. Therefore, peptidase activity appears to be more closely cell-associated than protease a c t i ~ i t y . ~ ~ . ~ ~ The measurements reported here were from net removal of fluorescamine-reactive peptide from the extracellular fluid, so they include intracellular accumulation as well as hydro- lysis. Furthermore, no distinction can be made between hydrolysis by cell wall associated enzymes, as occurs with some s t r e p t o c o ~ c i , ~ ~ ~ ~ and the more common accumulation followed by intracellular hydr01ysis.l~. 16,26 Nevertheless, the kine- tics of di- and trialanine uptake (Table 2) indicated that V,,, of the mixed rumen population for di- and trialanine uptake was 1.3% and 2.9%, respectively, of those of wild-type E. ~ 0 l i ; ~ l corresponding K, in E. c0li3I was 20% and 23% of those for mixed rumen microbes. The activities in rumen contents were more similar to those of Saccharomyces cerevisiae, which ranged from 1 to 5 nmol min-* mg DM-l for di- and tripeptides.14
In E. coli, the composition of the amino acid side chains has little influence on peptide ~ p t a k e , ~ ~ , ~ ~ whereas S. cerevisiae is more ~e1ective.l~ Here, various tripeptides were taken up at only slightly different rates (Table 3), possibly reflecting the complex mixture of organisms present. A proline-containing dipep- tide (Table 3) and two peptide p-nitroanilides (Table 5) were more slowly metabolised, presumably because of the secondary amino acid. However, all of the observed differences were not great, and it may be concluded that the amino acid content of small neutral peptides has only a minor influence on their rate of uptake by a complex mixture of rumen microorganisms. The same may not be true for other mixtures of rumen microbes or for particular species of rumen bacteria.
Peptide metabolism should not be considered solely in terms of rumen bacteria. Rumen contents from faunated sheep had a significantly higher rate of trialanine uptake than that from ciliate-free animals fed the same diet, whereas dialanine uptake was not different (R. J. Wallace, G. A. Broderick and M. L. Brammall,

116 G. A . Broderick, R . J . Wallace, N . McKain
unpublished). Dipeptide metabolism therefore appears to be distinct in character from tripeptide metabolism, and associated primarily with the rumen bacteria. Both protozoa and bacteria appear to have a role in the metabolism of oligopep- tides, and their relative importance is dependent upon relative numbers.
The lack of effect of glucose and DTT on the observed rates of peptide uptake (Table 1) should not be interpreted to mean that peptide uptake is not energy dependent or 0,-sensitive, but rather that the assay conditions were adequate without further additions. Rumen microorganisms usually have an ample supply of reserve polysaccharide to provide energy during the frequent periods of energy starvation that occur in the rumen,8 and these reserves may have been used to drive peptide transport in these incubations. Whether or not peptide uptake may be 0,-sensitive can only be speculated on, since the high cell density and the anaerobic buffer used in the assay would result in the rapid reduction of any O2 entering the incubation mixture.
In view of the re~ervations'~. '~ on the value of model compounds (such as peptide p-nitroanilides) as substrates for transport systems and peptidase assays, the correspondence of results obtained with SRF is surprising. Rates of metabol- ism were similar, at 0.6 to 1.5 nmol min-I mg DM-' for peptide p-nitroanilides (Table 5) and 0.4 to 1-5 nmol min-l mg DM-' for unmodified peptides (Tables 3, 4), and proline-containing compounds were metabolised more slowly (Tables 3, 5). Peptide p-nitroanilide hydrolysis activity was also predominantly cell-associ- ated, as was oligopeptide metabolism. The only difference was in the finding of extracellular dipeptidase activity, which may be difficult to detect using p-nitro- anilide~.~' Therefore, p-nitroanilides may be useful in the study of rumen oligo- peptide metabolism.
The assay conditions used here gave apparent rates of peptide uptake of about 4 nmol alanine residues min-I mg DM-1 for the higher homologues at a concen- tration of 0.25 mmol litre-I (Table 4), and a maximum value of 5.6 nmol alanine min-' mg DM-l for the peptide most rapidly taken up, trialanine (Table 2). The corresponding rate of casein degradation can be calcul- ated to be greater than these rates. For example, if casein was present at 0.1 mg N ml-I and its fractional rate of hydrolysis was 0-4 h-l,34 its rate of breakdown would be about 0.7 mg N min-l litre-I, or, based on the average DM content of SRF measured here at 5 g litre-I, 0.14 mg N min-l g DM-I. Since the amino acid content of casein is 54 pmol mg-I N,35 this is equivalent to a rate of release of amino acids of 7.6 nmol min-' mg DM-I. Thus, for rapidly degraded proteins like casein, the rate of proteolysis may exceed the rate at which peptides are removed, and peptides may accumulate transiently in rumen fluid. However, if protein were present at lower concentration, or if it were less rapidly degraded than casein, peptides would not accumulate. In vivo experiments tend to support this argument. Heat treatment of soya bean meal decreased its degradability and reduced peptide flow from the rumen of cattle by 80%.s In sheep fed dietary supplements of 6% casein, peptides accumulated to a concentration of 3.8 pmol trialanine equivalents ml-I in rumen fluid 1 h after feeding, whereas with the more resistant albumin only 0-3 pmol min-I was found (R. J. Wallace and G. A. Broderick, unpublished).

Rumen peptide metabolism 117
ACKNOWLEDGEMENTS
The authors thank Margaret Brammall and Nicola Watt for technical help. G. A. Broderick acknowledges the US Department of Agriculture for funding his leave at the Rowett Research Institute, where this work was performed.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
Leng, R. A.; Nolan, J. V. Nitrogen metabolism in the rumen. J . Dairy Sci. 1984, 67,
Tamminga, S. Protein degradation in the forestomachs of ruminants. J . Anim. Sci.
Russell, J. B.; Sniffen, C. J. ; Van Soest, P. J . Effect of carbohydrate limitation on degradation and utilization of casein by mixed rumen bacteria. J . Dairy Sci. 1983, 66,
Broderick, G. A.; Craig, W. M. Mechanism of protein degradation by rumen micro- bes. Fed. Proc. 1983, 42 (3), 532 (abstract). Sniffen, C. J . ; Chen, G.; Russell, J. B. Concentration and flow of peptides from the rumen of cattle fed different amounts and types of protein. In: Report on the XVllIth Conference on Rumen Function, Chicago (Allison, M. J., Ed.), 1985, 28. Pittman, K. A.; Bryant, M. P. Peptides and other nitrogen sources for growth of Bacteroides ruminicola. J. Bacteriol. 1964, 88, 401-410. Prins, R. A. Biochemical activities of gut microorganisms. In: Microbial Ecology ofthe Gut (Clarke, R. T. J . ; Bauchop. T., Eds), Academic Press, New York. 1977.73-183. Russell, J. B. Factors influencing competition and composition of the rumen bacterial flora. In: Herbivore Nutrition in the Subtropics and Tropics (Gilchrist, F. M. C.; Mackie, R. I., Eds), The Science Press, Craighall, South Africa, 1984, 313-345. Allison, M. J. Nitrogen metabolism of ruminal microorganisms. In: Physiology of Digestion and Metabolism in the Ruminant (Phillipson, A. T., Ed.), Oriel Press, Newcastle upon Tyne, 1970, 456-473. Pittman, K. A.; Lakshmanan, S.; Bryant, M. P. Oligopeptide uptake by Bacteroides ruminicola. J. Bacteriol. 1967, 93, 1499-1508. Prins, R. A.; Van Hal-Van Gestel, J. C . ; Counotte. G. H. M. Degradation of amino acids and peptides by mixed rumen microorganisms. Z . Tierphysiol. Tiererniihr. Futtermittelk. 1979, 42, 333-339. Wright, D. E. Metabolism of peptides by rumen microorganisms. A p p f . Microbiol.
Bladen, H. A.; Bryant, M. P.; Doetsch, R. N. A study of bacterial species from the
1072-1 089.
1979,49, 161.5-1630.
763-775.
1967, 15,547-550.
rumen which produce ammonia from protein hydrolysate. A p p f . Miciobiol. 1961, 9, 17.5- 180. Becker, J. M.; Naider, F. Transport and utilization of peptides by yeast, In: Micro- organisms and Nitrogen Sources (Payne. J. W., Ed.), Wiley. New York, 1980, 257- 279.
15. Payne, J. W. Transport and utilization of peptides by bacteria. In: Microorganisms and
16. Payne, J. W. Peptide transport in bacteria: methods. mutants and energy coupling.
17. Wolfinbarger, L. Transport and utilization of peptides by fungi. In: Microorganisms
18. Broderick, G. A.; Wallace, R. J . ; McKain, N. E . Uptake of neutral peptides by mixed
19. Whitelaw, F. G . ; Bruce, L. A, ; Eadie, J. M.; Shand, W. J. 2-Aminoethylphosphonic
Nitrogen Sourcess (Payne, J. W., Ed.), Wiley, New York, 1980, 211-2.56.
Biochem. SOC. Trans. 1983, 11,794-798.
and Nitrogen Sources (Payne, J. W., Ed.), Wiley, New York, 1980,281-300.
ruminal organisms in vitro. Fed. Proc. 1987, 46 (4), 1176 (abstract).

118 G. A . Broderick, R . .I. Wallace. N . McKain
acid concentrations in some rumen ciliate protozoa. Appl. Environ. Microbiol. 1983,
20. Herbert, D.; Phipps, P. J.; Strange, R. E. Chemical analysis of microbial cells. In: Methods in Microbiology, Vol. 5B (Norris, J. R.; Ribbons, D. W., Eds), Academic Press, London. 1971, 209-344.
21. Perrett, D.; Webb, J. P. W.; Silk, D. B. A.; Clark, M. L. The assay of dipeptides using fluorescarnine and its application to determining dipeptidase activity. Anal. Biochem.
22. Nisbet. T. M . ; Payne, J. W. Peptide uptake in Saccharomyces cerevisiae: charactens- tics of transport systems shared by di- and tripeptides. J . Gen. Microbiol. 1979. 115,
23. Appel, W. Peptidases. In: Methodsof Enzymatic Analysis, Vol. 2 (Bergmeyer, H. U., Ed.), Academic Press, London, 1974,2nd edn, 949-978.
24. Steel, R. G. D.; Torrie. J. H. Principles and Procedures ofStatistics. McGraw-Hill. New York, 1960.
25. Cornish-Bowden, A. J . Principles of Enzyme Kinetics, Butterworths, Boston, Mass., 1976.
26. Payne, J. W. Transport and hydrolysis of peptides by microorganisms. In: Peptide Transport and Hydrolysis, Ciba Foundation Symposium No. 50, Elsevier, Amster- dam, 1977, 305-334.
27. Brock, F. M . ; Forsberg, C. W.; Buchanan-Smith. J. G. Proteolytic activity of rumen microorganisms and effects of protease inhibitors. Appl. Environ. Microbiol. 1982.44,
28. Kopecny, J.; Wallace, R. J . Cellular location and some properties of proteolytic enzymes of rumen bacteria. Appl. Environ. Microbiol. 1982, 43, 1026-1033.
29. Exterkate, F. A. Location of peptidases outside and inside the membrane of Strep- tococcus cremoris. Appl . Environ. Microbiol. 1984, 47, 177-183.
30. Kolstad, J.; Law, B. A. Comparative peptide specificity of cell wall, membrane and intracellular peptidases of group N streptococci. J . Appl. Bacteriol. 1985,58,449-456.
31. Alves, R. A.; Cleaves, J. T.; Payne, J. W. The role of outer membrane proteins in peptide uptake by Escherichia coli. FEMS Microbiol. Lett. 1985,27, 333-338.
32. Bergmeyer. H. U. (Ed.). Enzymes 3: peptidases, proteinases and their inhibitors. In: Methods of Enzymatic Analysis, Vol. 5. Verlag Chemie, Weinheim. 3rd edn, 1984. 74-98.
33. Hermsdorf, C. L.; Simmonds, S. Role of peptidases in utilization and transport of peptides by bacteria. In: Microorganisms and Nitrogen Sources (Payne, J . W.. Ed.), Wiley, New York, 1980, 301-334.
34. Broderick, G. A. In vitro procedures for estimating rates of ruminal protein degrada- tion and proportions of protein escaping the rumen undegraded. J . Nutr. 1978, 108,
35. Block, R. J.; Weiss, K. W. The Amino Acid Handbook, Charles C. Thomas, Springfield, Ill., 1956.
46, 951-953.
1975,68, 161-166.
127-133.
56 1-569.
181-190.





























