Embed Size (px)
Citation preview

Chapter 14
Molecular Identification of AnaerobicRumen Fungi
Martin Eckart, Katerina Fliegerova, Kerstin Hoffmann, and Kerstin Voigt
Abstract Anaerobic fungi are phylogenetically unique and form a separate group,
the Neocallimastigomycota, among the chitinous fungi. Until now six genera are
described within that phylum, namely the monocentric genera Neocallimastix,Caecomyces and Piromyces as well as the polycentric genera Anaeromyces,Cyllamyces and Orpinomyces. This chapter gives a brief survey of the fascinating
world of anaerobic rumen fungi, their phylogeny, and identification. The golden
standards of molecular identification as well as promising alternatives will be
discussed.
14.1 Introduction
The physiology of the microbial community is fundamental for understanding the
processes of anaerobic decomposition of plant material, and has an economic
relevance for mankind. The distribution of organisms within the rumen is essential
for our understanding of the biochemistry of cellulose degradation (Hungate 1966).
A major part of organisms within the rumen fluid encompasses bacteria and
flagellates, but fresh and undigested plant material is rapidly colonised by anaerobic
fungi. It is now generally known that the degradation of herbal carbohydrates by
rumen fungi accelerates the digestion by downsizing the plant tissue particles.
Those particles are subsequently more easily decomposed by bacteria and protozoa.
The effectiveness of digestion is an important contributor to the health of animals in
husbandry (Wulff 2001).
M. Eckart, K. Hoffmann, and K. Voigt
Institute of Microbiology, School of Biology and Pharmacy, University of Jena, Neugasse 25,
07743 Jena, Germany
e-mail: [email protected]
K. Fliegerova
Department of Biological Basis of Food Quality and Safety, Institute of Animal Physiology and
Genetics, Czech Academy of Sciences, v.v.i., Vıdenska 1083, 14220 Prague 4, Czech Republic
Y. Gherbawy and K. Voigt (eds.), Molecular Identification of Fungi,DOI 10.1007/978-3-642-05042-8_14, # Springer-Verlag Berlin Heidelberg 2010
297

Because of the economic and scientific interest in this topic, it is not surprising
that the first description of “flagellated organisms” living within the rumen was
given at the beginning of the twentieth century. But, astonishingly it needed more
than 60 years to discover these organisms to be fungi living without any oxygen.
The anaerobic environment is mandatory for the ecosystem rumen. It determines the
mode of life of microorganisms residing there. Besides being well-known prokar-
yotes, anaerobic fungi are important producers of short-chain fatty acids, which are
an essential source of nutrition for herbivores. Such a unique occupation of a special
ecologic niche by a group of heterotrophic, hyphal, and chitin containing eukaryotes
inevitably raises the question about the relationships of these fungi. Today, this
group is well supported by morphological and molecular data and accepted as the
Neocallimastigales (Li et al. 1993). Although the final position within the kingdom
Fungi is still unclear, it turned out to be a monophyletic group, as a basal lineage
besides or within the phylum Chytridiomycota, and is now recognised as phylum
(James et al. 2006). While a flagellated phase through the life cycle of chytridio-
mycetes is a case sui generis proved for Chytridiales, Rhizophydiales, Spizellomy-
cetales, Blastocladiales, and Neocallimastigales, the rumen fungi are characterised
by another unique attribute inside the kingdom Fungi: they live in anaerobiosis.
Until now, only several species of gut fungi have been described, probably because
of the problematic cultivation and maintenance of these organisms and high mor-
phological variability depending on growth conditions. Extensive studies of a broad
range of ruminants and application of modern methods in molecular biology will
probably bring deeper insights in microbial communities and species relationships.
This survey gives a brief overview of the historical background and of modern
trends in species recognition of this interesting fungal group.
14.2 Historical Background and the Discoveryof Rumen Fungi
Decisive for the terminology of anaerobic gut fungi was a flagellated organism
observed within the rumen of herbivores by Liebetanz (1910). This organism was
named Callimastix frontalis (Braune 1913) because of its high morphological
similarity to Callimastix cyclopis (order Blastocladiales) (Weissenberg 1912), a
flagellated parasite of Cyclops. Braune first described the multi-flagellated zoo-
spores seen in Fig. 14.1, but did not recognise them as a stage of a fungal life cycle
and misclassified this organism as parasite. The given name C. frontalis led to a
number of mis-assignments of parasitic flagellates within this genus. Ultrastructural
examinations of C. frontalis by Vavra and Joyon (1966) resulted in the establish-
ment of the new genus Neocallimastix. But unfortunately, the authors did not
recognise this organism as a fungus and still considered it as zooflagellate. Eight
years later, Whisler et al. (1974) assumed that organisms of the genus Neocalli-mastix are actually motile spores of an alternate life cycle of Coelomomycespsorophorae – a blastocladialean fungus – and declared the herbivores as
298 M. Eckart et al.

alternative hosts along with mosquitoes. Orpin (1977) first suggested that these
anaerobic organisms living in the rumen actually might be fungi and his assumption
was based on the recognition of chitin in the cell walls and on the morphological
description of the thallus of different Neocallimastix species (Orpin 1974, 1975,
1976). Orpin’s findings were in contrast to the general belief of microbiologists
that no obligate anaerobic fungi can exist and therefore fungal colonies growing
in anoxic tubes were discarded as oxygen contaminations (van der Giezen
2002). However, none of these scientists provided a taxonomic definition for
Neocallimastix. It was Heath et al. (1983) who linked Neocallimastix to the
chytridiomycetes by setting up the new family Neocallimastigaceae within the
Fig. 14.1 First description of Callimastix as a flagellate parasite. Front view, side view, and
cleavage with aequitorial layer (Braune 1913)
14 Molecular Identification of Anaerobic Rumen Fungi 299

order Spizellomycetales (phylum Chytridiomycota). The lack of multiple morpho-
logical characters has always been and still is a handicap for identifying these
organisms within the gut fungi.
14.3 Traditional and Current Systematics
Their incapability of locomotion and their appearance resulted in the erroneous
classification of fungi as plants before the twentieth century. An own kingdom
Fungi was recommended only in 1969 by Whittaker (1969). The Chytridiomycetes,
besides the Oomycetes andHyphochytriomycetes, were the only group of flagellated
organisms that shared the class-characteristic cell-wall polymers (Bartnicki-Garcia
1970) and lysine synthetic pathway (Vogel 1964) of the Eumycota, comprising
Zygo-, Asco- and Basidiomycetes at that time. In the 1980s, taxonomy and phylo-
geny of Chytridiomycetes were based on the thallus development, discharge of
zoospores, the size, ultrastructural complexity, and organisation of zoospores, as
well as number and length of flagella. Furthermore, characteristics like mono- and
polycentric development as well as the release of zoospores via diffusion or via
papillae affected the taxonomy and phylogeny of these basal fungi (Barr 1978).
The anaerobic gut fungi, as a special group of the flagellated fungi, often
changed their taxonomic position within the Chytridiomycetes. Because of their
late discovery and the unusual physiological character, especially the obligate
anaerobiosis, the rumen fungi were placed into different taxonomic groups over
time, first into the subdivision Mastigomycotina (Ainsworth 1966), and later into
the division Mastigomycota (Alexopoulos and Mims 1979). Within the Mastigo-
mycotina the following zoosporic fungi were accepted: Chytridiomycetes, Hypho-
chytridiomycetes, Plasmodiophoromycetes, and Oomycetes. The basis for this
classification built the zoospore with one or two flagellae as an asexual propagative
spore. The class Chytridiomycetes traditionally contained the four orders Chytri-
diales, Harpochytriales, Blastocladiales, and Monoblepharidales. Some studies
mentioned the order Harpochytriales, now known as a synonym of the group
Chytridiales (Kirk et al. 2008). Development of molecular genetic methods such
as polymerase chain reaction (PCR), cloning, and automated sequencing enabled to
generate data for diverse analyses. Traditional phylogeny based on the short-handed
phenotypic markers such as morphology, physiology, and biochemistry is now
complemented by statistically supported evolutionary analyses, which allowed
re-evaluation and re-classification of the whole kingdom Fungi including also the
young taxonomic group covering anaerobic fungi. Molecular-biological analysis of
gut fungi performed by Li et al. (1993) resulted in the establishment of an
own order, the Neocallimastigales, with only one family: Neocallimastigaceae.
Recently, Hibbett et al. (2007) postulated a separate phylum for this group, the
Neocallimastigomycota, adapted from the paraphyletic origin of the chytridiomy-
cete fungi concluded by James et al. (2006). An informal supertree based on several
analyses showed a close relationship to the chytridiomycetes (Hibbett et al. 2007).
300 M. Eckart et al.

However, new phylogenetic approaches display the chytridiomycetes again as
monophyletic group (Ebersberger et al. 2010). Therefore, the separate phylum
Neocallimastigomycota seems to be redundant. A comparison of both classical
taxonomy, based on morphology and physiology, and modern systematic methods,
based on up-to-date molecular-genetic techniques, is shown in Table 14.1.
At present, the family Neocallimastigaceae comprises six genera1 (Adl et al.
2005): Anaeromyces (Breton et al. 1990), Caecomyces (Gold et al. 1988), Cylla-myces (Ozkose et al. 2001), Neocallimastix (Vavra and Joyon ex Heath 1983),
Orpinomyces (Barr et al. 1989), and Piromyces (Gold et al. 1988). An overview
of the six taxonomic groups within the family Neocallimastigaceae is shown in
Table 14.2.
14.4 Phylogeny
Traditional phylogenetic results supported by molecular-genetic data can redraw
evolutionary hypothesis and consequently the affinity of organisms to taxonomic
groups. Like phenotypic characterisations, molecular phylogenetics should never
be based just on one character. Comparisons or combinations of morphological and
genetic characters lead to stable and well supported evolutionary hypotheses and
with this strengths and weaknesses of genetic markers become obvious. A marker
of high diagnostic value has to be unique to a species or even to a strain and at the
Table 14.1 The systematics of the chytridiomycetes based on traditional and modern classifica-
tion schemes
Traditional system Modern system
domain Eukaryota domain Eukaryota
kingdom Fungi (Linnaeus 1753) Nees 1817 kingdom Fungi (Linnaeus 1753) Nees 1817
phylum Chytridiomycota von Arx 1967
class Chytridiomycetes (de Bary 1863)
Sparrow 1958
order Chytridiales Cohn 1879
order Spizellomycetales Barr 1980
order Blastocladiales Fitzpatrick 1930
order Monoblepharidales Sparrow 1942
phylum Chytridiomycota von Arx 1967
class Chytridiomycetes (de Bary 1863) Sparrow
1958
order Chytridiales Cohn 1879
order Spizellomycetales Barr 1980
order Rhizophydiales Letcher 2006
order Neocallimastigales Li et al. 1993
class Monoblepharidomycetes Powell 2007
order Monoblepharidales Sparrow 1942
phylum Neocallimastigomycota Powell 2007
class Neocallimastigomycetes Powell 2007
order Neocallimastigales Li et al. 1993
phylum Blastocladiomycota James et al. 2006
class Blastocladiomycetes James et al. 2006
order Blastocladiales Fitzpatrick 1930
1http://indexfungorum.org/Names/familyrecord.asp?strRecordID=81063
14 Molecular Identification of Anaerobic Rumen Fungi 301

same time ubiquitous for all taxa. The more various the set taxonomic groups is, the
more conserved the marker has to be. On the other hand, clustering on lower level,
starting with the family, requires more variable data to distinguish between species
or even strains (outlined in Fig. 14.2).
Clustering methods use differences between partitions of given data to rebuild
cladistic relationships. To get quality estimation such as bootstrap proportions (BTs
or BP), checking the robustness of a set of data (Felsenstein 1985) is required.
Highly conserved data lead to stable reconstructions in early branches caused
by low clade stability supports. An example is given in Fig. 14.2. There is no
possibility to distinguish between taxon2 and taxon3 based on identical sequence
data. This exemplary marker is not adequate for molecular diagnostics on a lower
taxonomic level such as genus or species. We demonstrate these problems with an
analysis based on real data in Fig. 14.3. The gene encoding actin is highly con-
served in eukaryotes. The coding sequence divergence between plant and non-plant
actin genes shows only 15% or less variability (Hightower and Meagher 1986).
Therefore, this marker demonstrates perfectly the relationship between anaerobic
and aerobic chytridiomycetes, with the zygomycetous order Mucorales as outgroup
(Fig. 14.3). Varieties of species level cannot be determined with actin data, as this
marker lacks molecular diagnostics possibilities.
Table 14.2 Survey of the species from the anaerobic chytrids
Genus Species Author
Neocallimastix frontalis (RA Braune) Vavra and Joyon 1966 ex IB Heath et al.
(1983)
hurleyensis Theodorou and Webb (1991)
joyonii Breton, Gaillard, Bernalier, Bonnemoy and Fonty (1988)
patriciarum Orpin and Munn (1986)
variabilis Ho and Barr (1993)
Anaeromyces elegans Ho (1993)
mucronatus Breton et al. (1990)
Caecomyces communis Gold et al. (1988)
equi Gold (1988)
sympodialis Chen, Tsai and Chien (2007)
Cyllamyces aberensis Ozkose et al. (2001)
Orpinomyces bovis Barr et al. (1989)
intercalaris Ho et al. (1994)
joyonii1 (Breton, Bernalier, Bonnemoy, Fonty, Gaillard and Gouet)
Li, Heath and Cheng (1990))
Piromyces citronii Gaillard, Breton, Dusser and Julliand (1995)
communis Gold, Heath, and Bauchop (1988)
dumbonicus Li (1990)
mae Li (1990)
minutus Ho (1993)
polycephalus Chen, Chien and Hseu (2002)
rhizinflatus Breton, Dusser, Gaillard, Guillot, Millet and Prensier (1991)
spiralis Ho (1993)1Basionym, current name: Neocallimastix joyonii Breton, Bernalier, Bonnemoy, Fonty, B. Gaillard
& Gouet 1989
302 M. Eckart et al.

Nevertheless, highly variable data could result in an unstable reconstruction of
early branches caused by “long-branch-attraction” (Bergsten 2005). Although high
variable data could help measure distances between the closest neighbours and
other taxa on lower taxonomic levels, the variability of the data could lead to false
positive congruence, such as analogy instead of homology. An example is shown in
Fig. 14.4. The high variability of the internal transcribed spacer (ITS) sequences of
chytridiomycetes allow to distinguish even between strains, but alternative ways to
cluster these data decrease the robustness of the data set. One problem is the
differentiation of homologous and paralogous markers. Homology is not a problem
if orthologous genes are involved, but paralogous genes can lead to misinterpreted
results, similar to the comparison of “apples and oranges”. One example would be
the eukaryotic translation elongation factor 1-a (EF-1a) with more than one copy
within the genomes of fungi (see fungal genomes published by the JGI at http://
genome.jgi-psf.org/). False positive results in phylogenetic analysis based on align-
ments of paralogous genes are not always obvious as shown in Fig. 14.5.
14.5 Predicted Impact of Molecular Markers on FutureIdentification and Phylogeny
Based on the theory of evolution, highly conserved genes of diverse taxonomic
groups could be amplified by the combination of PCR techniques and universal
oligonucleotides (primers). The most commonly used DNA region for molecular-
genetic phylogeny is the highly repetitive cluster of the nuclear ribosomal DNA
(rDNA). The nucleotide sequences of the nuclear small (SSU) and large (LSU)
Fig. 14.2 Schematic illustration of problems occurring during the application of single phyloge-
netic markers. The master sequence should be ATTGCTAAGCGA; the (recent) taxa show
modified sequences. Changes compared to the consensus sequences are colour-coded in red.
Occurring problems are obvious: a stable backbone with statistical support is only possible with
data that are not highly diverse. However, differentiation at higher branches requires variable
sequences. To combine these datasets, several approaches like supermatrix or supertree methods
can be applied
14 Molecular Identification of Anaerobic Rumen Fungi 303

Fig.14.3
PhylogenyoftheNeocallim
astigomycota
andother
chytridiomycetesbased
onaMaxim
um
Likelihoodanalysisofactinsequencesfrom
24taxa
withatotalof903aligned
characters(unpublished
sequences).(m
ethodical
inform
ations:GAMMA
þP-Invar
model
withRAxML7.0.4
GTR-CAT(rapid
hill-clim
bingbootstrappingmethod(Stamatakis
2006,2008)),10,000rapid
bootstrap
inferencesbefore
athoroughML
search;final
ML
Optimization
Likelihood:–4676.139464)
304 M. Eckart et al.

Fig.14.4
PhylogenyoftheNeocallim
astigomycotaandother
chytridiomycetesbased
onaMaxim
umLikelihoodanalysisofITSsequencesfrom
49taxawith
atotalof1,161
aligned
characters
(unpublished
sequences).(m
ethodical
inform
ations:
GAMMA
þP-Invar
model
with
RAxML
7.0.4
GTR-CAT,
(Stamatakis2006,2008),10,000rapid
bootstrap
inferencesbefore
athoroughMLsearch.Final
MLOptimizationLikelihood:–17766.020153)
14 Molecular Identification of Anaerobic Rumen Fungi 305
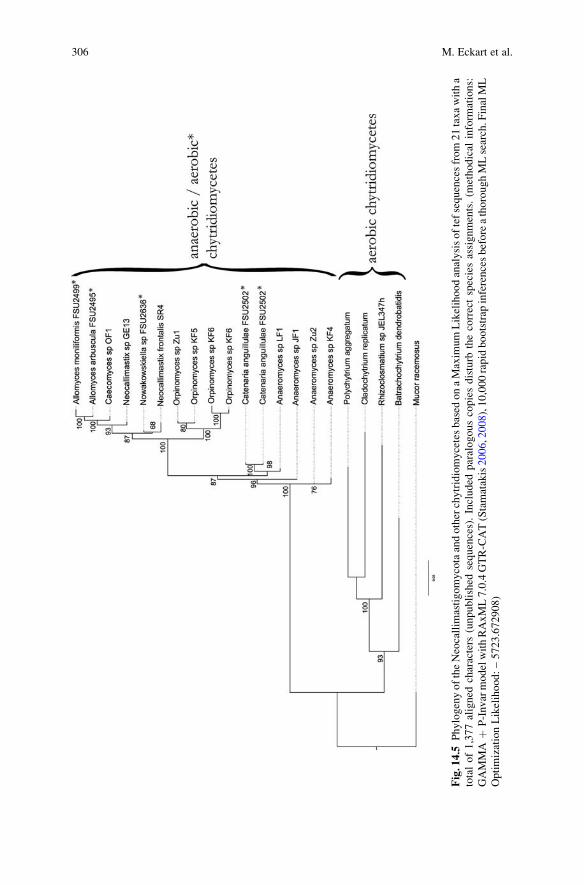
Fig.14.5
PhylogenyoftheNeocallim
astigomycotaandotherchytridiomycetesbased
onaMaxim
umLikelihoodanalysisoftefsequencesfrom21taxawitha
totalof1,377aligned
characters
(unpublished
sequences).Included
paralogouscopiesdisturb
thecorrectspeciesassignments.(m
ethodical
inform
ations:
GAMMA
þP-InvarmodelwithRAxML7.0.4GTR-CAT(Stamatakis2006,2008),10,000rapidbootstrap
inferencesbeforeathoroughMLsearch.F
inalML
OptimizationLikelihood:–5723.672908)
306 M. Eckart et al.

subunits are separated by the non-coding DNA sequences of the internal transcribed
spacer (ITS) 1 and 2 and the non-transcribed intergenic spacer (IGS). Lacking a
sufficient evolutionary pressure, the non-coding regions allow the separation of
organisms down to the levels of species and strains. Using the flanking conserved
sequences of 18S (SSU) and 28S (LSU) rDNA, these regions can easily be
amplified with universal primers. Unfortunately, the ITS regions are not single
copy regions. Although the ribosomal DNA cluster follows concerted evolution
(Arnheim 1983), the intra-specific variability among organisms cannot be denied.
Usage of this region as molecular barcode marker is therefore questionable, espe-
cially if there is no reliable and supporting approach for species identification based
on e.g. morphology (Nilsson et al. 2008). Moreover, the variability of the ITS
region is sometimes not high enough to separate at the level of species as shown
for the fungal genus Penicillium (Skouboe et al. 1996, 1999). This experience
enforced the search and establishment of alternative genetic markers like the intron-
containing protein coding genes actin (act), eukaryotic translation elongation factor
1-a (tef), or beta-tubulin (btub). A profound base for this approach requires reference
strains, which need to bemorphologically and genetically well characterised, and also
the subsequent completion of the published results and sequence submissions.
First efforts to identify the anaerobic gut fungi by molecular genetic methods
were done by Dore and Stahl (1991) and Bowman et al. (1992). Their approaches
relied on partial 18S rDNA sequences for including the anaerobic fungi into the
chytridiomycetes, but the authors did not separate the species within the genera
(Dore and Stahl 1991; Bowman et al. 1992).
Trying to clarify the phylogenetic relationships within the order Neocalli-
mastigales using sequence analysis (ITS1) combined with morphological features,
ultrastructures and mitotic characters have led to seperation of the order Neocalli-
mastigales (Li et al. 1993). Isozyme analyses or DNA hybridisation has also been
used with the aim to clarify identification of anaerobic gut fungi and to increase the
level of specificity (Ho et al. 1994).
A fast and easy method for the differentiation of polycentric anaerobic fungi is
available by (RFLP) analysis of ITS spacer and/or fragments of ribosomal large
subunit (28S rDNA) digested by proper endonucleases. However, the ribosomal
small subunit (18S rDNA) turned out to be too conservative to get a well resolved
DNA polymorphism, and therefor is not very suitable for this type of analysis
(Fliegerova et al. 2006).
Methods of molecular biology are very promising, but “old-fashioned” taxon-
omy is still substantiated despite many discrepancies. The classical approach of
Neocallimastigales identification is based on their morphological characters. Thal-
lus shape (filamentous or bulbous), zoosporangial development (monocentric or
polycentric), and number of flagella per zoospore (uni- or polyflagellated) are
decisive for genus differentiations, while the ultrastructure of the zoospore is
determinative for species. (Heath et al. 1983; Orpin and Munn 1986; Munn et al.
1988; Webb and Theodorou 1991). Unfortunately, characters observable by light
microscopy vary with culture conditions and are highly pleomorphic (Brookman
et al. 2000). Moreover, the cultures often fail to produce important structures
14 Molecular Identification of Anaerobic Rumen Fungi 307

(sporangia and zoospores) making identification even more problematic. Also the
differentiation of species using ultrastructural features of the zoospores is question-
able, because ultrastructure depends not only on the age of microorganisms but also
on the method and quality of their preparation (Ho and Barr 1995).
14.6 Molecular Identification and DNA Barcoding
One of the important characteristics of anaerobic fungi is their flagellated stage in life
cycle. However, flagellated zoospores, can be found also in other aquatic fungi, like
the Blastocladiomycota and the Chytridiomycota sensu stricto and also in some
protists, e.g., the stramenopiles and among those the oomycetes, which are derived
brown algae. The flagellae of these organisms caused the mis-applications of taxo-
nomic and phylogenetic assignments as it happened to Braune with Neocallimastix(1913). The elucidation of morphological characters is valuable and indispensable,
but has to be supported by techniques of molecular genetics because the pleomorphic
shape of fungi leads to complications in their identification. Therefore, molecular
information becomes more and more important as a primary source for species
recognition. Now, 90 years after the discovery of the anaerobic rumen fungi, molec-
ular phylogenetic studies confirmed their relationship to the kingdom Fungi (Forster
et al. 1990; Bowman et al. 1992). The choice of molecular genetic markers in the
kingdom Fungi, respectively the phylum Chytridiomycota, is clearly arranged.
Today, state of the art comprises seven markers for fungal phylogeny that provides
data over the complete spectrum of the kingdom: 18S rDNA, 28S rDNA, ITS1 and
ITS2 including the 5.8S rDNA, rpb1, rpb2, tef, and beta-tubulin. In special cases
like pathogenic species or organisms of industrial importance, some additional
markers exhibiting a higher specificity were developed. Such markers encompass
not only genes encoding calmodulin, Mcm7 (MS456), and Tsr1 (MS277) (Aguileta
et al. 2008; Schmitt et al. 2009) but also physiological properties such as toxins or
extrolite profiles, which are well established, for example, the ascomycetous genus
Penicillium (Samson and Frisvad 2004).
To find the most useful marker for “tagging” all forms of life is the aim of many
current projects involved in “DNA barcoding”. DNA barcoding is an approach to
identify any organism based on sequence analysis of selected genomic regions.
Access to these regions should be as universal as possible, comparable, reproduc-
ible, and relatively easy to accomplish. Barcoding is thought to serve not only the
identification or verification of known specimens but also to contribute in the
discovery of new, undescribed species. Although DNA barcoding already proved
to be a very useful tool for the discovery of cryptic species, which are by definition
not differentiable by morphological features (Hebert et al. 2004), barcoding is
nevertheless error-prone. Depending on the method used, DNA barcoding turned
out to be not always sufficient for species recognition (Brower 2006; DeSalle
et al. 2005; Whitworth et al. 2007). One of the major problems in all barcoding
308 M. Eckart et al.

approaches is still the question which molecular tool should be used, since every
further step in species identification is based on it.
In animal systems, the mitochondrial cox1 is widely applied (Hebert et al. 2003),
although its sufficiency is already questioned (Goetze 2003). With a slower evolu-
tionary rate of this cytochrome c oxidase, this marker is not applicable for flowering
plants (Kress et al. 2005).
One of the most discussed marker in fungal taxonomy and phylogenetics is still
the ITS regionwith all its aforementioned advantages and disadvantages (see Sect. 5).
Phylum CoSAscomycota 1,124,132
Basidiomycota 197,974environmental samples 41,955Glomeromycota 11,698Microsporidia 5,676unclassified fungi 5,107Fungi incertae sedis 5,092
Zoopagomycotina 32Entomophthoromycotina 439Kickxellomycotina 276Mucoromycotina 3,948unkown 397
Chytridiomycota 1,683Neocallimastigomycota 465Blastocladiomycota 134
a
b
Fig. 14.6 Schematic illustration and number of nucleotide sequences provided by the Interna-
tional Nucleotide Sequence Database Collaboration. (e.g., Genbank). (a) Percentages of the
number of fungal sequences provided in GenBank. The graph shows the total number of submitted
sequences within the kingdom Fungi. The subparts describe the single phyla based on the
taxonomy provided by the TaxBrowser at NCBI. The group Dikarya is represented by approxi-
mately 96% of all available sequences. The number of sequences that were generated of environ-
mental samples is higher than that of all other phyla with the exception of the Dikarya. The
Neocallimastigomycota represent with 465 sequences the second smallest group of fungal organ-
isms represented as nucleotide sequences within GenBank. (b) Survey of the nucleotide sequencesprovided by the International Nucleotide Sequence Database Collaboration (as of May 1st, 2009)
14 Molecular Identification of Anaerobic Rumen Fungi 309

Nevertheless, provided sequence data usable for species identification for anaerobic
gut fungi are restricted, e.g., only 163 “ITS” tagged sequences are assigned to the
order Neocallimastigales (465 nucleotide sequences in summary, compare Fig. 14.6),
with only 17 sequences assigned to full taxon names. Because of missing mitochon-
dria in anaerobic fungi (hydrogenosomes instead), mitochondrial based barcode
markers are out of question (Bullerwell and Lang 2005). The need for a complete bar-
coding database, as always demanded (Ekrem 2007), is obvious. But another major
drawback is the data deposited in such a barcode database. An adequate number
of well-defined reference specimens are a prerequisite for species identification
and especially for species discovery. Such references should encompass all possi-
ble variances within defined species boundaries, e.g., geographically based varia-
tions (DeSalle et al. 2005; Meyer and Paulay 2005).
Originally thought to be a fast, cheap, and easy-to-access method for the
assignment of “unknown” to “known” specimens, molecular barcoding should be
used with caution. On the one hand, supplementing a barcode marker with addi-
tional information about e.g., morphology, biogeography, or even more molecular
data will miss the aim of a single easy-to-use marker for species assignment. But on
the other hand, supplementing data is necessary as a specimen cannot be identified
or described with certainty by one molecular attribute (Brower 2006; Will et al.
2005). Storing new data in a database is always tied with responsibility of the
submitter. Open-access to such databases is necessary but at the same time prone to
errors and losing its value as shown by GenBank at the NCBI (Bridge et al. 2003).
14.7 Conclusion and Future Line of Research
According to the efforts of Aguileta et al. (2008) and Schmitt et al. (2009) more
alterantive barcoding markers need to be established and validated in order to get a
reliable identification which is in concordance with morphological and ultrastructural
characters. The increase of the complexity of research on anaerobic rumen fungi in
their composite ecosystems requires a common platform for strain and data shared
among the scientific community. It is necessary to gain a certain homogeneity and
common use of reference and type strains including reference sequences of barcode
markers and other characters suitable for a reliable identification of anaerobic rumen
fungi. This is a fundamental for cultivation-independent detection in the natural
ecosystems and habitats of anaerobic fungi as performed by Fliegerova et al. (2010).
Acknowledgments The authors express their gratitude to L. Jay Yanke (Agriculture and Agri-
Food Canada, Lethbridge Research Centre, Lethbridge, AB, Canada) for providing Neocallimastixfrontalis strain SR4. This project was a component of the institutional research plan (AV OZ 5045
0515) of the Institute of Animal Physiology and Genetics, Academy of Sciences of the CzechRepublic in Prague. The czech team was supported by The National Agency for AgricultureResearch (project no. QI92A286/2008), The german team was supported by the Deutsche For-schungsgemeinschaft (project no. Vo 772/7-1), which is part of a bilateral grant between the CzechScience Foundation and the Deutsche Forschungsgemeinschaft.
310 M. Eckart et al.

References
Adl SM, Simpson AGB, Farmer MA, Andersen RA, Anderson OR, Barta JR, Bowser SS,
Brugerolle G, Fensome RA, Fredericq S, James TY, Karpov S, Kugrens P, Krug J, Lane CE,
Lewis LA, Lodge J, Lynn DH, Mann DG, McCourt RM, Mendoza L, Moestrup O, Mozley-
Standridge SE, Nerad TA, Shearer CA, Smirnov AV, Spiegel FW, Taylor MFJR (2005)
The new higher level classification of eukaryotes with emphasis on the taxonomy of protists.
J Eukaryot Microbiol 52:399–451
Aguileta G, Marthey S, Chiapello H, Lebrun MH, Rodolphe F, Fournier E, Gendrault-Jacquemard A,
Giraud T (2008) Assessing the performance of single-copy genes for recovering robust phylo-
genies. Syst Biol 57:613–627
Ainsworth GC (1966) A general purpose classification of the fungi. Bibl System Mycol 4:1–4
Alexopoulos CJ, Mims CW (1979) Introductory Mycology 38:1–613
Arnheim N (1983) Concerted evolution of multigene families. In: Nei M, Koehn RK (eds)
Evolution of genes and proteins. Sinauer Associated, Sunderland, MA, pp 38–61
Barr DJS (1978) Taxonomy and phylogeny of chytrids. Biosystems 10:153–165
Barr DJS, Kudo H, Jakober KD, Cheng KJ (1989) Morphology and development of rumen fungi:
Neocallimastix sp., Piromyces communis, and Orpinomyces bovis gen.nov., sp.nov. Can J Bot
67:2815–2824
Bartnicki-Garcia S (1970) Cell wall composition and other biochemical markers in fungal
phylogeny. In: Harborne JB (ed) Phytochemical phylogeny. Academic Press, New York,
pp 81–103
Bergsten J (2005) A review of long-branch attraction. Cladistics 21(2):163–193
Bowman BH, Taylor JW, Brownlee AG, Lee J, Lu S, White TJ (1992) Molecular evolution of the
fungi: relationship of the basidiomycetes, ascomycetes, and chytridiomycetes. Mol Biol Evol
9:285–296
Braune R (1913) Untersuchungen uber die im Wiederkauermagen vorkommenden Protozoen.
Arch Protistenk 32:111–170
Breton A, Bernalier A, Dusser M, Fonty G, Gaillard-Martinie B, Guillot J (1990) Anaeromycesmucronatus nov. gen., nov. sp. A new strictly anaerobic rumen fungus with polycentric thallus.
FEMS Microbiol Lett 70:177–182
Bridge PD, Roberts PJ, Spooner BM, Panchal G (2003) On the unreliability of published DNA
sequences. New Phytol 160:43–48
Brookman JL, Mennim G, Trinci AP, Theodorou MK, Tuckwell DS (2000) Identification and
characterization of anaerobic gut fungi using molecular methodologies based on ribosomal
ITS1 and 18S rRNA. Microbiology 146:393–403
Brower AVZ (2006) Problems with DNA barcodes for species delimitation: ‘ten species’ of
Astraptes fulgerator reassessed (Lepidoptera: Hesperiidae). Syst Biodivers 4:127–132
Bullerwell CE, Lang BF (2005) Fungal evolution: the case of the vanishing mitochondrion. Curr
Opin Microbiol 8:362–369
DeSalle R, Egan MG, Siddall M (2005) The unholy trinity: taxonomy, species delimtiation and
DNA barcoding. Philos Trans R Soc Lond B Biol Sci 360:1905–1916
Dore J, Stahl DA (1991) Phylogeny of anaerobic rumen Chytridiomycetes inferred from small
subunit ribosomal RNA sequence comparisons. Can J Bot 69:1964–1971
Ebersberger I, Strauss S, Kupczok A, Kothe E, von Haeseler A, Gube M, Eckart M, Voigt K (2010)
A stable backbone for the fungi. in revision
Ekrem T, Willassen E, Stur E (2007) A comprehensive DNA sequence library is essential for
identification with DNA barcodes. Mol Phylogenet Evol 43:530–542
Felsenstein J (1985) Confidence limits of phylogenies: an approach using the bootstrap. Evolution
39:783–791
Fliegerova K, Mrazek J, Voigt K (2006) Differentiation of anaerobic polycentric fungi by rDNA
PCR-RFLP. Folia Microbiol (Praha) 51:273–277
14 Molecular Identification of Anaerobic Rumen Fungi 311

Fliegerova K, Mrazek J, Hoffmann K, Zabranska J, Voigt K (2010) Diversity of anaerobic fungi
within cow manure determined by ITS1 analysis. Folia Microbiol (Praha): in press
Forster H, Coffey MD, Elwood H, Sogin ML (1990) Sequence analysis of the small subunit
ribosomal RNAs of three zoosporic fungi and implications for fungal evolution. Mycologia
82:306–312
Goetze E (2003) Cryptic speciation on the high seas; global phylogenetics of the copepod family
Eucalanidae. Proc R Soc B 270:2321–2331
Gold JJ, Heath IB, Bauchop T (1988) Ultrastructural description of a new chytrid genus of caecum
anaerobe, Caecomyces equi gen. nov., sp. nov., assigned to the Neocallimasticaceae. BioSys-
tems 21:403–415
Heath IB, Bauchop T, Skipp RA (1983) Assignment of the rumen anaerobe Neocallimastixfrontalis to the Spizellomycetales (Chytridiomycetes) on the basis of its polyflagellate zoo-
spore ultrastructure. Can J Bot 61:295–307
Hebert PDN, Ratnasingham S, deWaard JR (2003) Barcoding animal life: cytochrome c oxidase
subunit 1 divergences among closely related species. Proc R Soc B 270(Suppl):96–99
Hebert PD, Penton EH, Burns JM, Janzen DH, Hallwachs W (2004) Ten species in one: DNA
barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. ProcNatl Acad Sci USA 101:14812–14817
Hibbett DS, Binder M, Bischoff JF, Blackwell M, Cannon PF, Eriksson OE, Huhndorf S, James T,
Kirk PM, Lucking R, Lumbs HT, Lutzoni F, Matheny PB, McLaughlin DJ, Powell MJ,
Redhead S, Schoch CL, Spatafora JW, Stalpers JA, Vilgalys R, Aime MC, Aptroot A,
Bauer R, Begerow D, Benny GL, Castlebury LA, Crous PW, Dai YC, Gams W, Geiser DM,
Griffith GW, Gueidan C, Hawksworth DL, Hestmark G, Hosaka K, Humber RA, Hyde KD,
Ironside JE, Koljalg U, Kurtzman CP, Larsson KH, Lichtwardt R, Longcore J, Miadlikowska J,
Miller A, Moncalvo JM, Mozley-Standridge S, Oberwinkler F, Parmasto E, Reeb V,
Rogers JD, Roux C, Ryvarden L, Sampaio JP, Schussler A, Sugiyama J, Thorn RG, Tibell L,
Untereiner WA,Walker C, Wang Z,Weir A,Weiss M,White MM,Winka K, Yao YJ, Zhang N
(2007) A higher-level phylogenetic classification of the fungi. Mycol Res 111:509–547
Hightower RC, Meagher RB (1986) The molecular evolution of actin. Genetics 114:315–332
Ho Y, Barr D (1995) Classification of anaerobic gut fungi from herbivores with emphasis on
rumen fungi from malaysia. Mycologia 87:655–677
Ho YW, Khoo IY, Tan SG, Abdullah N, Jalaludin S, Kudo H (1994) Isozyme analysis of anaerobic
rumen fungi and their relationship to aerobic chytrids. Microbiology 140:1495–1504
Hungate RE (1966) The rumen and its microbes. Academic Press, New York
James TY, Letcher PM, Longcore JE, Mozley-Standridge SE, Porter D, Powell MJ, Griffith GW,
Vilgalys R (2006) A molecular phylogeny of the flagellated fungi (Chytridiomycota) and a
proposal for a new phylum (Blastocladiomycota). Mycologia 98:860–871
Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008) Dictionary of the fungi, 10th edn. CABI
Publishing, Wallingford, UK
Kress WJ, Wurdack KJ, Zimmer EA, Weigt LA, Janzen DH (2005) Use of DNA barcodes to
identify flowering plants. Proc Natl Acad Sci USA 102:8369–8374
Li J, Heath IB, Packer L (1993) The phylogenetic relationships of the anaerobic chytridiomycetous
gut fungi (Neocallimasticaceae) and the Chytridiomycota II. Cladistic analysis of structural
data and description of Neocallimasticales ord. nov. Can J Bot 71:393–407
Liebetanz E (1910) Die parasitischen Protozoen des Wiederkauermagens. Arch Protistenk
19:19–90
Meyer CP, Paulay G (2005) DNA barcoding: error rates based on comprehensive sampling. PLoS
Biol 3:e422
Munn EA, Orpin CG, Greenwood CA (1988) The ultrastructure and possible relationships of four
obligate anaerobic chytridiomycete fungi from the rumen of sheep. Biosystems 22:67–81
Nilsson RH, Kristiansson E, Ryberg M, Hallenberg N, Larsson K (2008) Intraspecific ITS
variability in the kingdom fungi as expressed in the international sequence databases and its
implications for molecular species identification. Evol Bioinform 4:193–201
312 M. Eckart et al.

Orpin CG (1974) The rumen flagellate Callimastix frontalis: does sequestration occur? J Gen
Microbiol 84:395–398
Orpin CG (1975) Studies on the rumen flagellate Neocallimastix frontalis. J Gen Microbiol
91:249–262
Orpin CG (1976) Studies on the rumen flagellate Sphaeromonas communis. J Gen Microbiol.
94:270–280
Orpin CG (1977) Invasion of plant tissue in the rumen by the flagellate Neocallimastix frontalis.J Gen Microbiol 98:423–430
Orpin C, Munn E (1986) Neocallimastix patriciarum: a new member of the Neocallimasticaceae
inhabiting the sheep rumen. Trans Br Mycol Soc 86:178–181
Ozkose E, Thomas BJ, Davies DR, Griffith GW, Theodorou MK (2001) Cyllamyces aberensis gen.nov sp.nov., a new anaerobic gut fungus with branched sporangiophores isolated from cattle.
Can J Bot 79:666–673
Samson RA, Frisvad JC (eds) (2004) Penicillium subgenus Penicillium: new taxonomic schemes,
mycotoxins and other extrolites. Stud Mycol 49:1–251
Schmitt I, Crespo A, Divakar PK, Fankhauser JD, Herman-Sackett E, Kalb K, Nelsen MP,
Nelson NA, Rivas-Plata E, Shimp AD, Widhelm T, Lumbsch HT (2009) New primers for
promising single copy genes in fungal phylogenetics and systematics. Persoonia 23:35–40
Skouboe P, Boysen M, Pedersen LH, Frisvad JC, Rossen L (1996) Identification of Penicillium
species using the internal transcribed spacer (ITS) regions. In: Rossen L, Rubio V, DawsonMT,
Frisvad JC (eds) Fungal identification techniques. European Commission, Brussels, Belgium,
pp 160–164
Skouboe P, Frisvad JC, Taylor JW, Lauritsen D, Boysen M, Rossen L (1999) Phylogenetic
analysis of nucleotide sequences from the ITS region of terverticillate Penicillium species.
Mycol Res 103:873–881
Stamatakis A (2006) RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with
thousands of taxa and mixed models. Bioinformatics 22(21):2688–2690
Stamatakis A, Hoover P, Rougemont J (2008) A rapid bootstrap algorithm for the RAxML web-
servers. Systematic Biology 75(5):758–771
van der Giezen M (2002) Strange fungi with even stranger insides. Mycologist 16:129–131
Vavra J, Joyon L (1966) Etude sur la morphologie, le cycle evolutif et la position systematique de
Callimastix cyclopis Weissenberg, 1912. Protistologica 2:5–16
Vogel HJ (1964) Distribution of lysine pathways among fungi: evolutionary implications. Am Nat
98:435
Webb J, Theodorou MK (1991) Neocallimastix hurleyensis sp.nov., an anaerobic fungus from the
ovine rumen. Can J Bot 69:1220–1224
Weissenberg S (1912) Sitzungsbericht der Gesellschaft naturfoschender Freunde zu Berlin 5:299
Whisler HC, Zebold SL, Shemanchuk JA (1974) Alternate host for mosquito parasite Coelomo-myces. Nature 251:715–716
Whittaker RH (1969) New concepts of kingdoms or organisms. Evolutionary relations are better
represented by new classifications than by the traditional two kingdoms. Science 163:150–160
Whitworth TL, Dawson RD, Magalon H, Baudry E (2007) DNA barcoding cannot reliably
identify species of the blowfly genus Protocalliphora (Diptera: Calliphoridae) Proc Bio Sci
274:1731–1739
Will KW, Mishler BD, Wheeler QD (2005) The perils of DNA barcoding and the need for
integrative taxonomy. Syst Biol 54:844–851
Wulff C (2001) Untersuchungen zum Thiamin- und Thiaminderivatgehalt im Pansensaft des
Rindes nach Verfutterung von mit Mucor racemosus Fresenius verpilztem Heu (in vitro),
Thesis, Tierarztliche Hochschule Hannover, Hannover
14 Molecular Identification of Anaerobic Rumen Fungi 313





























