Embed Size (px)
Citation preview

USE OF A MATHEMATICAL MODEL TO STLIDY THE DYNAMICS OF CTENOCEPHAL~DES FELIS POPULATIONS IN THE HOME ENVIRONMENT
AND THE IMPACT OF VARIOUS CONTROL MEASLIRES BEUGNET F.'. PORPHYRE T.". SABATIER P." & CHALVET-MONFRAY K.*'
Summary :
The biology of fleas has been studied by a nurnber of authors, as has the impact 01 various types of controi rneasures. However, there are no mathematical modes sirnuating rhe dynamics of a population of Ctenocephalides ielis felis fleas on their host (the cat/ and in their close environment (apartment). The rnodel presented in this paper allows for integration of the numerous biological and behavioural parameters of the parasites and their hosts and for the variation of these same parameters The various types of conirol rneasures can be prograrnrned so that their impoct over time can be studied The model coniirrns the key role played by adult fleas, or ernerged fleas contained in the cocoon. Oniy regular applications of persistent insecticides to the host animal will enable controi of the parasite population. A cornbination of these insecticides with an IGR [lnsect Growth Regulator) will accelerate decontarnination of the home environment and see the disappearance of the parasites altogether if they are not re- introduced. The association of additional rneasures such as vacuum cleaning w i l acceierate the process of decontamination but will have no impact if carried out in isolation. One-off treotment with insecticide will not enable a reduction in the parasite popuaiion, even i f carried out frequently. Use of insecticides on the home enviranment premises alone does not appear to be an adequate rneans of control. The preseni rnodel can be used io test various integrated control rneasures which take into account different factors such as the nurnber of host animais, the frequency of rnovement outdoors, the impact of the seosons
Résumé : MOD~LISATION DF. LA DYNkUlQLIE DE POPIJLATIONS DE CTF.\;OCEPHALIDES FEUS DANS L'ENVIRONNEMENT DOhlESTIQUE ET ÉTllrlE DF L'IMPACT DES DIFFÉRENTS MOYENS DE CONTKÔLE
10 biologie des puces a été étudiée por de nombreux aiiteurs, comme les différentes modalités de traitement. Il n'existe cependant pas de modèle mothémotique simulont la dynamique d'une populotion de puces Cteno~e~halides feis felis sur son hôte (chat) et dons l'environnement proche /de type oppartement). l e modèle ici présenté permet d'intégrer et de foire varier les nombreux paramètres biologiques et comporlementaux des parosites et de leurs hôtes. Les différents types de traiiement peuvent être programmés de façon à étudier leur impact dans le temps. l a dynamique coniirme le rôle clé joué par les puces adulbs on encore émergées et contenues dans le cocon. Seuls des traityents insecticides rémanents appliqués régulièrement sur l'hôte d'obtenir un contrôle des populations parasitaires. Une combinaison avec des /GR (lnsect Growth Regulator) accélère Io décontamination de l'habitat et peut permettre une disparition des porosites si ces derniers ne son1 pas réintroduits dons le milieu Lossociation oux mesures complémenbires de type aspiration occélère le processus de décontaminotion, mois celles-ci n'ont aucun impact si elles sont isolées. Les traitements insecticides ponctuels ne permertent pas de réduire la population parasitaire, même s'ils sont faiis lréquemment. La lutte insecticide dans l'environnement n'apparaît pas comme un moyen de Iurte adéquat. Ce modèle pourra être utilisé pour tester différents moyens de lune intégrée, selon différentes situations : nombre d'hôtes, sorties fréquentes. incidence saisonnière,
KEY WORDS : Ctenocepha/ides ieiis, mathematical rnodel, population dynamics, insecticide, GR [lnsect Growth Regulator]. MOTS CLES : Cienocephaides feis. modélisation rnathémoiiqiie, dynomique
de population, insecticide, IGR.
T tie biology of Ctenocephalides felis,felis, the cat flea, is well known today through the work of various teams, in particular those of Dryden
(Bowman, 1999; Reugnet & Bourdeau, 1999; Dryden & Rust. 1994; Kaufnlan, 1996). Domestic carnivores can be infected by several types of flea (Ctenocephalides felis. Ctenocephalides canis, Pulex iwitarîs, Archeopsylla erinacei, Spilopsyllzrs cuniculi. Xenopsylla cheopis, Cera-
* Merial. 29, avenue Tony Garnier. 69007 Lyon, France. ** Bioriiiithernarics Unir, National Veterinar) School, Lyon, 1, avenue Bourgelat, 69480 Marcy ]'Étoile, France. Correspondence: F. Beugner. E-riidil: [email protected]
toph.yl1u.ï gallinae and Lqtopsylla segnis) but, with few exceptions it is the cat flea, Ctenocephalides felis.felis, that is involved in 90 % of cases of infestation of pet cats and dogs. The fleas of rodents, small carnivores, wild insectivores or birds are obsen-ed more rarely (Beaucournu & blénier, 1998; Franc et al., 1998; Kettle, 1995). Cteizoccphalides cariis is obsen ed throughout Europe, but its frequency varies from 2 to 30 % of cases, depending on the country. This species is palearctic in origin, adapted to colder zones. C. felis is African in origin and is thought to have been irnported into Europe by returning crusaders. It then spread throu- ghout the world in the wake of human migration. The sub-species present in Europe is C. .felis felis (Beau- cournu & Ménier, 1998). The cat flea is not particu-
Parasite, 2004. 11. 387-399

larly specific and can fèed off the blood of a variety of lnammals (carnivores, rabbits, ruminants or hilinans). It is not uncommon for pet owners to be bitten by fleas (Ménier & Beaucournu, 1999). The flea is the most common ectop~irasite, infesting both rural and urban carnivores. It is adapted to 130th exterior and home environments and can be observed throughout the year, although infestation tends to he greatest from spring through to autiimn (Dryden Oi Rust, 1994). Ctenocephalides felis filis is a small, wingless. insect, 2 to 4 mm in length, reddish-brown to black in colour. The body is laterally compressed: which allows for easy movement through fur. and the hind legs are highly developed, powerful for jumping. The average distance juinped by Ctenocephalides, felis, felis is 20 cm (varying frorn 2 to 48 cm), and 30 cm (from 3 to 50 cm) for Ctenocephalides canis. ,4s to height, this is approxi- mately 15 cm, with a maximum of 25 cm for C. ca??is (Cadiergues et ul., 2000). Knowledge of the flea life cycle is essential to devi- sing effective prevention of infestation. '4 number of commonly accepted views are false, in particular that the adult flea is a transient par:isite, present on the host cat or dog only when it requires a fresh 13lood rnenl. Adult fleas tend to stay perrnanently on the same animal (Kettle, 1995; Kramer & Mencke, 2001). \Xrhen they fa11 off: they can only survive approxirnately three to five days in the home environment. A very small proportion of fleas present on a carnivore will change liost and infest another animal. The risk of contami- nation through direct contact between animals, such 21s in a vet's waiting room, although often mentioned, is low (Franc Oi Cadiergues, 1997). It is not aln:ays easy for a pet owner to see nhen an animal is infested with fleas. A number of experiments have shown that they are only seen on about one third of animals infested. The use of a fine comb increases the sensitivity of the search and seeing them on the animal in about two thirds of cases (Gregory et ul., 1995). It is much easier to find their fecal matter, in the form of little black specks 0.5 to 1 mm in size: which is evidence of the fresh blood meal. Adult fleas reproduce rapidly. approximately 48 hours after their first fresh blood meal n;hich usually occurs within 30 niinutes of arriva1 on the cat or dog host animal. Each female flea can produce 11p to 50 eggs a day for between 50 and 100 days, with an average of 20 to 30 eggs per day over a period of two months. A flea can live for a maximum of 100 days (Dryden, 1994; Kramer & Rlencke, 2001). The smooth, white, oval eggs measure ahout half a mil- limetre in length. They are not fixed to the host animal, but faIl to the ground as it iiioves about or while it rests or sleeps. Under favourable conditions of tem- perature and humidity, the eggs will hatch in three to seven days as wormlike larvae a few millimetres long.
Larvae feed on almost any organic debris, in particiilar dried adult flea fecal matter. Larvae prefer humidity and avoid light. They crin move horizontally about 20 cm, in order to take cover (under a chair, for example, or to the bottoln of carpet and mg fibres) (Robinson, 1995). After three stages of lanral development and in a period of anything from one n:eek to one month, each 6 to 7 mm stage 3 larvae will spin a cocoon in nhich the pupae will develop into the adult stage in about 10 days. If conditions are favourable, i.c. if ani- mals are present in the environment, the adults will emerge rapidly. If conditions are not favourable, however, the young adult fleas in the cocoon, called non-emerged adults, can still survive for several months (011 average 150 days) protected by the silk cocoon (Beugnet & Bourdeaii. 1999; Dryden & Rust, 1994; Kramer & Mencke. 2001). These non-emerged adults are a significant soutce of parasites im~nediately avai- lable should a host animal happen to pass in the vici- nity. In addition, theÿ are relatively protected from insecticides. Newly emerged fleas actively look for host animals and can survive about a week without a meal. Environriiental conditions play a part in flea ecology and in the chr~nology of the flea iife cycle. Ali stages are sensitive td'drying and a relative humidity rate of 85 !/o is optimal. 'Temperature accelerates or slows deve- lopment, a minimiim of 22" C seemingly being required, n:ith an optirnurii of 25-26" C. On the other hand, a tem- perature above 30°C nrill decrease the life span of adults. In winter, an outside temperature approaching O" C ndl cause the death of both larvae and pupae. If temperature in the house is 19" C, the life cycle is consi- derably slon.ed and only pre-enierged adults will reiiiain. These factors explain the presence of fleas al1 year round, and nhy, given their prolificacy and the rapi- dity of the development cycle, there is a population explosion as soon as fine weather appears. In condi- tions of sufficient humidity, Ctenocephalides , felis has a life cycle which lasts 14 days at a temperature of 29" C and 140 days at 13°C (Dryden c'i Rust, 1994). Adult fleas will eiiierge from the cocoon when sub- mitted to various stimuli. A passing shadon:, footsteps, vibrations, can al1 cause fleas to leave the cocoon, as can ninning a vacuum cleaner over the floor. So it can be of great interest to do this before, or just after: treat- ment of the home environriient with an insecticide. Host animals can be infected by moving into an infec- ted environment, whether outside, season permitting (walks, holidays), or inside (another pet owner's home environment). Often cats will "import" fleas into a home environment, starting the life cycle going, then any dog or dogs who reside there niIl also become infected. Given biological and ecological data, a mathematical mode1 of a flea population in a defined environriient, such as an apartment where a dog or cat is present, can be envisaged. If the results tally n~ith in vivo and

in vitro biological studies, this niodel would then - T2: period of nyniph stage of development IT2 = four perniit simulation of the impact of various types of daysl; control iileasures - chernical (with insecticides or hor- - T:: period during whicli latent nyrnphs do not die nlone inliibitors), or mechanical - and an appraisal of (,IVL, (t)) [T3 = 90 days]; combinations of these ~ypes of control measures. - T,: period during which latent nymphs die OVL, (t))
(T4 = 150 daysl:
MATERIAL AND METHODS - Tg: average tinie of persistence at UFA stage (UFA =
pre-emerged fleas, still in cocoon) [TT = one dayl.
iven the known hiology of the differerit stages of the flea life cycle, tlie mode of development, and a chronological sequence estahlished in
terms of the surroundings and cbnditions in the honie environment, it is possible to reproduce this cycle in diagram foriii.
GENERAL DESCRIPTION OF MODEL AND DEFINITION OF VARIABLES
We first of al1 considered the five variables represen- ting the stages of flea development: - A (t): total numher of adiilt fleas at time period t; - WT (t): total nurnher of fleas at egg and larvae stage at time period 1;
- N (t): total nun~ber of fleas at nymph stage, as a func- tion of time; - IVL (t): total numher of fleas at latent nyiiipli stage ( i . ~ . that will not ininiediately evolve into newly emerged adults), as a f~inction of time; - UR4 (t): total number of pre-adult fleas, as a func- tion of tinie.
Clironological paranieters were then assigned for the various stages in the flea life cycle: -Tl: period of embryo and kamal stages of development [Tl = from 10 to 30 days, deperiding from tlie climate];
Bio1ogic:il paranieters concerning rates of mortality, prolificacy, sex ratio, arere also assigned at various sta- ges: - pl: rilortality rate for adults; - y,: mostality rate for rggs/lan.ac/nymphs; - ;L-:: niortality rate for latent nyrriphs: - AL,: mortality rate for UFA; - x,: nurnl->er of adult fleas brought in from outdoors (introduced into tlie model); - x,: numher of I_!FA1s brought in from outdoors: - r: derisity-dependent excess mortality rate: - k: maximum reception capacity, dependent on host animal; -.fem: proportion of fernale in a d u l ~ population; - a: prolificity; - rr: pqcentage of fertile eggs; - fl péicentage of success rate for development of inimatuie stages; - y percentage of' latent nymphs surviving after tinie 13, during T,: - V: rate of passage of la~ent nymphs to UFA, depen- dent on presence of host animal; - 8 percentage of success of LIFA'S.
These parameters are sunimarized in Table 1. They enable a progressive model to huilt.

Definition 1 ~ymbol unit 1 Value
Variables 1 1 1 Nuniber o t adults
Biological parameters -
Period of embryo ,lnd 1,1n31 development
A
Pe-h development =ige time for nymphs before dying Periocl d u r i n g w h i c h m p h s die A\.eragc time of persistence at UFA stage .\duit niortality rate LJFA mortality rate Number of adults I~rought in frorii outdoors Number of UFA's broiiglit in from outdoors Excess mortality rate V t i o n capacity Female proportion in the adult flea population
Prolificity Success rate egg/larvae development Ç I i r r e s s h dedevelopnient ? O of nymph survival after T3+'T-t
indepen-resence of host animal Proportion of nymphs becoming
Number of eggz, l a m e Number of nymphs - Number of latent nymplis Numher of unformed adtilt\
l numl->er
Tl
Prohability of contact of a UFA m-itli 3 liost 1 6
presence of host animal l
-
T2
T3 T4 T i
11 1 p4 x l x2 r K
feiii h a P Y
-
Environmental parameters 1 1 1
WL N NL
LlF.4
day
1 1 ~ l * S l x ~ ~ r ~ p * Y N S n r Presence of host animal Host behaviour - going outdoors
1
numl~er number niiniber numher
30
day day &y d:iv
1 d3v 1, &ay
N 2 weeks N e e e k s
Hea -
N eggdfemale -
-
-
Proportion of nymphs l~econiing LIFA'S
Alternation of seasons 1 1 Season 1 - 0
4
90 --
150 1
0.02 1 5 5
3 0.71
300 2 3
27*(IGRto*IGRvo) (alf [ . . . l*sol~(Am*IGRe~
0.6 0.10
Modulating function of floor&\ er nig/carpet on population
v l
--
Modulating funçtion of home environment on succrss of hatching
\ - ', 1
Treatment parameters
Floor -+ alf [T OC. O h H2OI 1
Efficacy of mechanical action on environment Efficacy of insecticide on environment Efficacy of IGR o n envi roment Efficacy of non persistent insecticide Efficacy of persistent insecticide on host aninxil Efficacy of persistent insecticide in presence o f ~ i l a r shampoo treatment Effic~lcy of IGR on host animal, topical
formulation 1 1
forrriulation ~ i p to 90 days and development
Am
Efficacy of IGR on host animal, oral
1 for 30 days rhen r:ipid decrease IGRvo
Table 1. - Principal parameters of the model: symbol, unit and value
-
Parasite. 200 t . 11, 387-399
INSe lGRe
INSnr = Shamp INSr
Modulation of INSr
IGRto
60 % for 1 day -
-
-
-
-
- -
80 o/, for 1 dav 60 O ? for 60 da. then r:ipid decrease
m s 100 O o for 30 days then decrease
to O O ? at D90 Efficacy reduced to 80 O? for 30 days tlien
mpid decrease to 0 % at D40 100 O/o for 30 days then slow decrease

FOR~VULATION OF MODEL where:
Formulation of model is presented on Figure 1, in which: - WL: represents the population of eggs and larvae. They have a quick evolution, Tl (from 10 to 30 days), before becoming nyrnphs; - N: nymphs, with a mortality rate of p2 which could not be distinguish from the global moratlity of eggs and
P larvae. The nyrnph will evolve within a T2 time (four days) to hecome a NL stage which includes a future adult flea. The nymph population is subdivided in NL1 and NL2, which are the final nymph stages, including a non-emerged flea; - NL1: late nymph or cocoon which includes a pre- adult stage = non emerged flea. NL1 could evolve very quickly by releasing newly emerged fleas, especially in case of the presence of host. Nevertheless, in case of absence of the host, NL1 will become latent, which corresponds to the NL2 status; - NL2: latent pre-emerged fleas inside the cocoon, which represents a stock of future flea in the environ- ment. These stages are waiting for on host (dog,/cat) to emerge and hecome newly emerged fleas (= LTFA); - NL1 + NL2 are living during a minimum of T3 time, which is around 90 days. After this delay, the rate of mortality increase and the NL population is decreasing during a T4 time (in average 150 days); - UFA: Newly emerged fleas, which could come from NL1 (quick appearance) or from the NL2 stock (quies- cent pre-emerged fleas present in the environment and waiting for a stimulus). The newly emerged fleas have to find and jump very fast on their host as they cannot stay alive a long time without a first hlood meal. The average time of life before becoming Adult parasite stage or dying is TS (from one to three days, one in the model helow).
Given the diagram and the parameters set, the model can be formulated into an equation. The equations des- cribing the flea life cycle as a function of time are written as follows:
F ( t ) ahfemA(t) ( 2 ) where terms of mortality p are equal to:
p4= 1 - 6
T5 and where Tl is the development tiine for eggs and larvae so that:
T , = (1 - p) x Tl,,, (4) T, is the development time for nymphs. As to nymphs-in-waiting, at the end of a maximum time T, + T4, only y remain. With k dependent on the host animal, and v and 6 dependent on the presence of the host, v and 6 are ni1 in the absence of a host.
In the case where the host is absent, adult fleas disap- pear, whereas the immature stages continue their deve- lopment to the latent nyrnph stage. The numerical values are as follow. The follb,wing initial conditions have been selected for this part of the model:
We thus start with a healthy environment and the intro- duction of an animal infested with 20 adult fleas. We then consider the existence of in-flows which will interfere with the base model. These flows consist of the entry of adult fleas subsequent to the outdoor movements of the host animal (classic for the cat, for example), but also the possihility of nymphs/pupae being introduced by an owner.
a 4 ( t ) - P F ( t - q -T2 -T,)e-"" - ( V + ~ ) N ~ ( , ) - ~ F ( ~ - T , -T, -< - T , ) ~ - ~ ~ - ( ' + * ) T , Ir-
Parasite, 2004, 11. 387-399

Two in-flows, independent of the development cycle. can be distinguished: An adult flow k, originating from outdoors that the host animal brings in to the liome environment after having been outdoors. It should be noted that k, is a func- tion of time. k,(t) = x, in the case where there is a host going outdoors in the warm season (spring to auturnn), othemise k,(t) = 0. This is summarized as:
k,(t) = x, x Season x HostComp x Host (6) where Season and HostComp are such that:
1 if host goes out HostComp =
O if host does not go out
To these equations, we then added the effect of the home environment on the successful development of embq-os and larvae. The success rate for this period is indicated using the parameter P. In the home environment, two elements are necessary for the survival of individual fleas: i) that floor cove- rings be suitable for larvae developriient, i.e. mgs. car- pets and old parquet flooring, and ii,l conditions of tem- perature and humidity in the home environment. These two parameters are summarized as follows: -sol: modulating hnction of floor coverings rugs/carpet on the population; - betu [T O C , % H,Ol: modulating function of lioriie envi- ronment on the success of developnient; with, for the function sol, the following values accor- ding to the type of covering:
O % O IO O6, 20 %[ 0.4 [20 Oh, 50 %[ 0.7
[50 %, 100 %] 1 and for f~inction beta [T OC, % H,Ol, resulting froni the combined effect of temperature T OC and humidity % H,O, the values are:
[10-GO[ 160-751 175-85i a 85
O O O O.? 0.4 0.3 O
i22-30[ 0.5 0.6 0.5 O 0.7 0.8 0.7 O
40 O O O 2 0.2 O
Paranieter p thus becomes a f~inction which depends on conditions in the home environment and can then be written:
p = a x sol x beta [T OC, % H,O] (7)
We need to be able to induce the supposed effects of various types of treatment in the previously defined niodel. The types of treatment and their effects are directly derived from current knowledge concerning the control of flea populations (Carlotti 8: Jacobs, 2000).
DEFINITION OF TYPES O F TREATMENTS
Let us first of al1 consider the eight \rariables repre- senting the various types of anti-flea treatment: - 4 s (t): action of home vacuum cleaner on popula- tion dynamics; - INS ( t ) : action of an insecticide applied to the home environnient premises; - IGR (t): action of an IGR (Insect Growth Regulator) applied to the home environment premises; - INSr (t): action'of the treatment of host animal with a persistent insecticide; - ILVSnr (t): action of the treatlnent of host animal with a non-persistent insecticide; - IGRvo (t): action of the treatment of host aninial with an IGR adpnistered orally; - ZGRto (t):.action of the treatment of host animal with an IGR administered locally.
Mechanical action: use of vacuum cleaner The aspirating action of the vacuum cleaner is a tem- porary one. The eggs sucked up leave the cycle while those not taken up continue to develop without any problem. However, this is a regular action. We will consider that thereafter it is done periodically, a period t more or less long, depending on the hygiene of the host animal's owner. Action As (t), which represents the change in sumival rate at the WL stage linked to the action of the vacuum cleaner, can thus be sum- marized as:
For t = 772, As(t) = 1 - e, (8) For t # 172, As(t) = 1
where el is the efficacy of vacuuming. However, effi- cacy e l can Vary, so we have varied the values from 60 % to 40 %. In Figure 2, el is equivalent to 40 % and t is equal to 7. Which corresponds to vacuuming once a week. It is difficult to assess the rate of efficacy of vacuuming, depending on several factors rythm of application, power of the material. density :ind length of carper fibers, surface of carpet ... (Dryden, 1994). The choice of 40 5% for efficacy of vaccuming is based on publi- shed data (Dryden, 1994) as well as a recent experi- ment done in Germany and comparing various vac- curning cleaners and various type of carpets (2004, Veterinary Faculty of Münich, unpublished data).
Parasite. 2004. 11. 387-399

O O O 20 40 60 80 1 O0 120 O 50 1 O0 150 200
Time (days) Tlme (days)
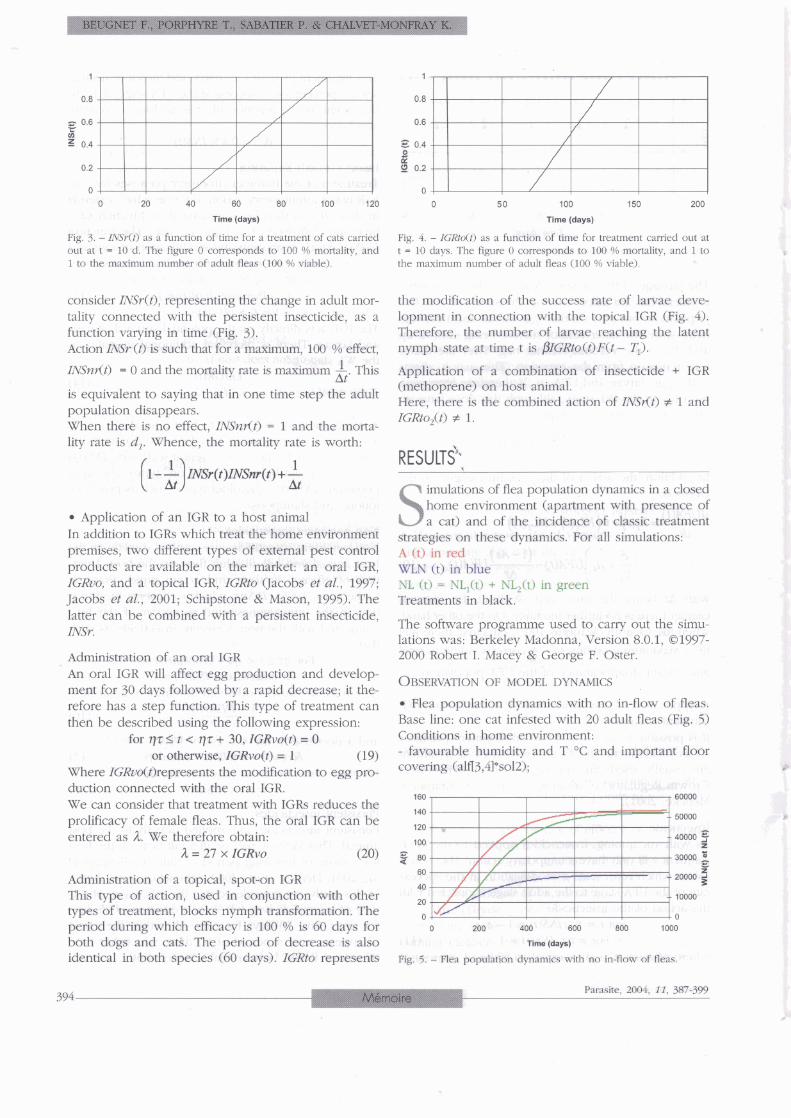
Fig. 3. - INSr(t) as a function of time for a treatment of cats carried Fig. 4. - ZGRto(t) as a function of time for treaunent carried out at out at t = 10 d. The figure O corresponds to 100 % mortality, and t - 10 days The figure O corresponds to 100 % mortality, and 1 to 1 to the maximum number of adult fleas (100 YO viable). the maximum number of adult fleas (100 Oh viable)
consider INSr(t), representing the change in adult mor- the modification of the success rate of larvae deve- tality connected with the persistent insecticide, as a lopment in connection with the topical IGR (Fig. 4). function varying in time (Fig. 3). Therefore, the number of larvae reaching the latent Action Z N 9 (t) is such that for a maximum, 100 % effect, nymph state at time t is PlGRto(t)~(t - Ti).
1 INSnKt) = O and the mortality rate is maximum -. This Application of a combination of insecticide + IGR At (methop~ene) on host animal. is equivalent to saying that in one time step the adult Here, there is the combined action of ZNSdt) # 1 and
population disappears. IGRto,(t) # 1. When there is no effect, ZNSnrft) = 1 and the morta- lity rate is dl. Whence, the mortality rate is worth:
1 (1 - h) ~ ~ s r ( t ) ~ ~ ~ n r ( t ) + -
At
Application of an IGR to a host animal In addition to IGRs which treat the home environment premises, two different types of extemal pest control products are available on the market: an oral IGR, IGRuo, and a topical IGR, IGBo (Jacobs et al., 1997; Jacobs et al., 2001; Schipstone & Mason, 1995). The
imulations of flea population dynamics in a closed home environment (apartment with presence of a cat) and of the incidence of classic treatment
strategies on these dynamics. For al1 simulations: A Ct) in red %LN (t) in Mue NL (t) = NL,(t) + NL,(t) .kt greed Treatrnents in black.
latter can be combined with a persistent insecticide, ZNSr. The software programme used to carry out the simu-
lations was: Berkeley Madonna, Version 8.0.1, 01997- Administration of an oral IGR 2000 Robert 1. Macey & George F. Oster. An oral IGR will ment for 30 days refore has a step
affect egg production and develop- followed by a rapid decrease; it the- function. This type of treatment can
then be described using the following expression: for qz 5 t c q~ + 30,ZGRvo(t) = O
or otherwise, IGRvo(t) = 1 (19) Where ZGRvo(t)represents the modification to egg pro- duction connected with the oral IGR. We can consider that treatment with IGRs reduces the prolificacy of female fleas. Thus, the oral IGR can be entered as A. We therefore obtain:
a = 27 x IGRVO (20)
OBSERVATION OF MODEL DYNAMICS
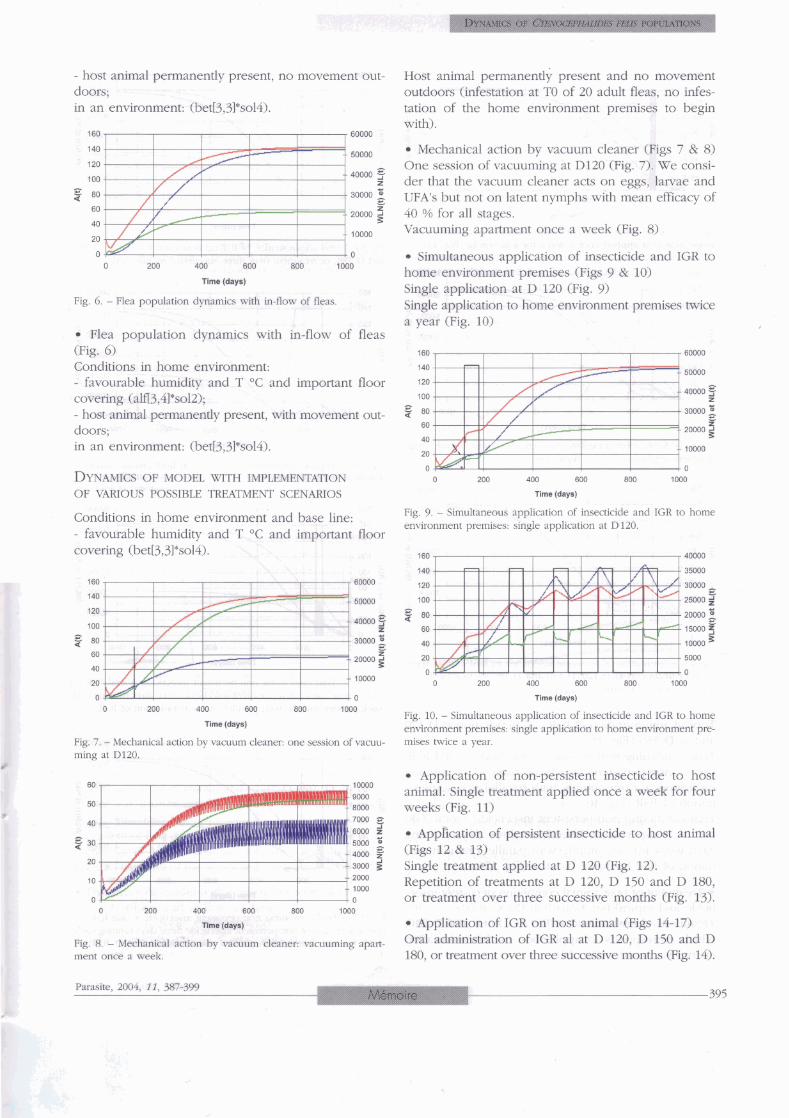
Flea population dynamics with no in-flow of fleas. Base line: one cat infested with 20 adult fleas (Fig. 5) Conditions in home environment: - favourable humidity and T OC and important floor covering (alfl3,4l*so12);
Y
60 Administration of a topical, spot-on IGR 2 0 0 ~ 0 3
40 This type of action, used in conjunction with other 10001)
types of treatment, blocks nymph tran~formation. The O O
period during which efficacy is 100 % is 60 days for O 200 400 600 800 1 000
both dogs and catS. The period of decrease is also r i (days)
identical in both species (60 days). ZGRto represents Fjg. 5 . - Fkea population dynamics with no h-flow of fleas
Parasite, 2004, 11, 387-399
- host animal permanently present, no movement out- doors; in an environment: (bet[3,31*so14).
Time (days)
Fig. 6. - Fîea population dynamics with in-flow of fleas
Flea population dynamics with in-flow of fleas (Fig. 6) Conditions in home environment: - favourable humidity and T OC and important floor covering (alfl3,41*so12); - host animal permanently present, with movement out- doors; in an environment: (bet[3,31*so14).
DYNAMICS OF MODEL WITH IMPLEMENTASION O F VARIOUS POSSIBLE TREATMENT SCENARIOS
Conditions in home environment and base line; - favourable humidity and T OC and important floor covering (betl3,31*so14).
Time (days)
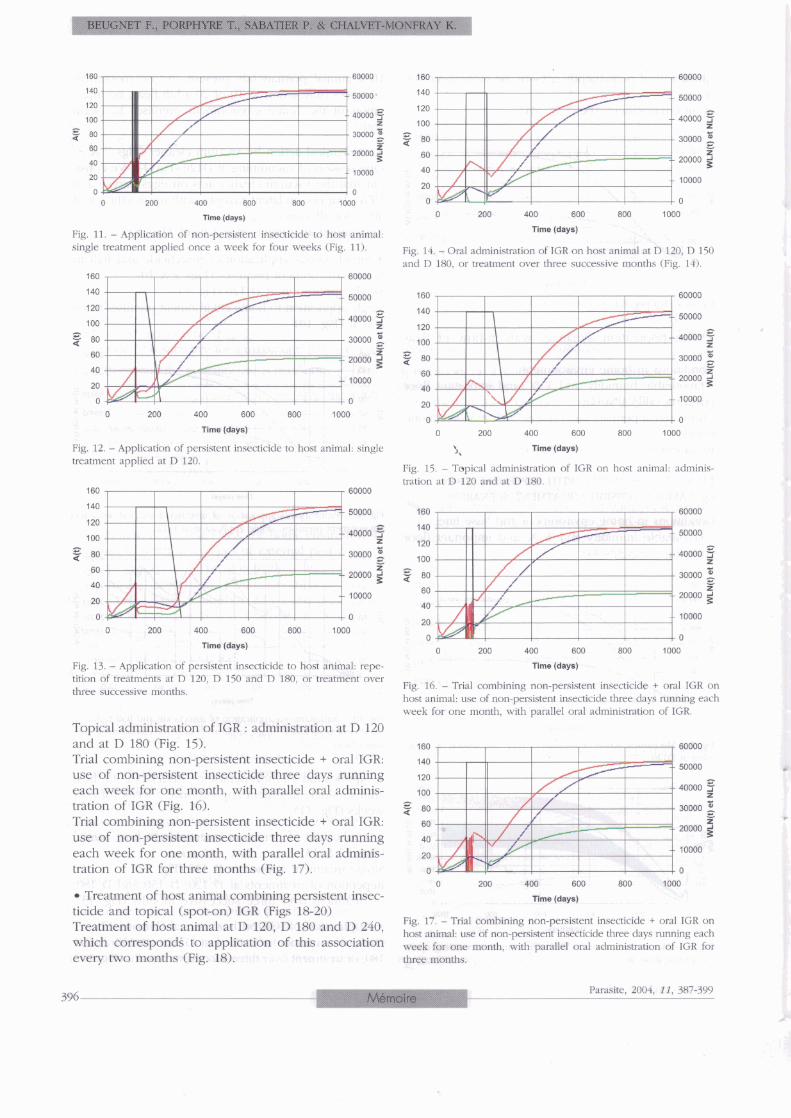
Fig. 7. - Mechanical action by vacuum cleaner. one session of vacuu- ming at D120.
Hoa animal present and no movement outdoors (infestation at TO of 20 adult fleas, no infes- tation of the home environment premises to begin with).
Mechanical action by vacuum cleaner (Figs 7 & 8) One session of vacuuming at Dl20 (Fig. 7). We consi- der that the vacuum cleaner acts on eggs, larvae and UFA's but not on latent nymphs with mean efficacy of 40 % for al1 stages. Vacuuming apartment once a week (Fig. 8)
Simultaneous application of insecticide and IGR to home environment premises (Figs 9 & 10) Single appluiation at D 120 (Fig. 9) Single application to home environment premises twice a year (Fig. 10)
Tlme (days)
Fig. 9. - Simultaneous application of insecticide and IGR to home environment premises: single application at D120.
O 200 400 600 800 1000
Tlme (days)
Fig. 10. - Simultaneous application of insecticide and IGR to home environment premises: single application to home environment pre- mises twice a year.
Application of non-persistent insecticide to host animal. Single treatment applied once a week for four weeks (Fig. 11)
~ppfication of persistent insecticide to host animal (Figs 12 & 13) Single treatment applied at D 120 (Fig. 12). Repetition of treatments at D 120, D 150 and D 180, or treatment over three successive months (Fig. 13).
O 20a 400 600 800 1 O00
~ l m o (-1 Application of IGR on host animal (Figs 14-17) Fig, 8, - Mahania] a&on by vacuum cleaner vacuuming apafi- Oral of IGR at lZ0, 150 and ment once a week. 180, or treatment over three successive months (Fig. 14).
T i m O 200 400 800 800 1000
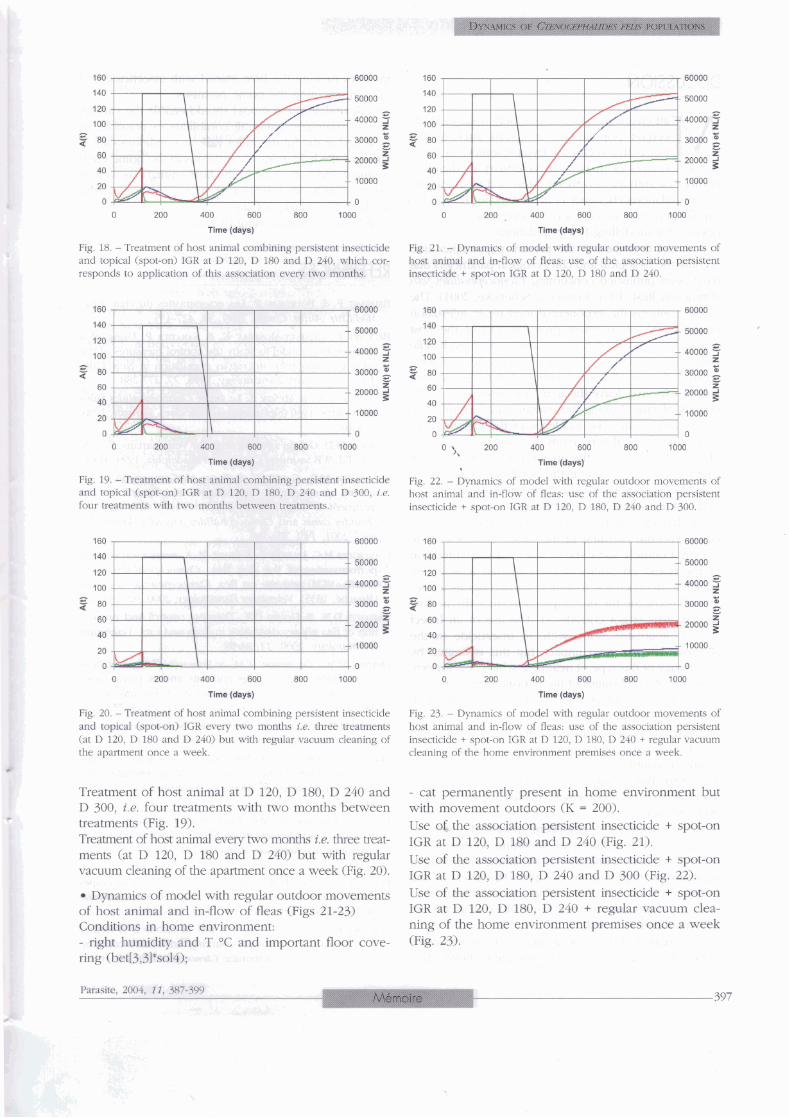
Fig. 11 - Applicritiofi of non-persistent inseaici& to hcxff animal Tifne t*W)
single treament applied once a week for £Our weeks (Fig. 11). ~ i ~ , 14, - oral of IGR on hast at D 120, D 150 and D 180, or treatment over three successive months (Fig. 14).
160 , , 60000
Fig. 12. - Application of persistent insecticide to host animal: single treatment applied at D 120.
Fig. 13. - Applihtion of persistent insecticide Co host aimal: repe- tition of treamients at D 120, D 150 and D 180, or katment over three sumernive months.
Topical admiisiration of iGR : adminismtbn at D 120 and at D 180 (Fig. 15). Trial combining non-persistent insecticide + oral IGR: use of non-persi-F>t;ent insecticide three days nurning each week fos one month, with paralle1 oral admin&- tration of IGR (Pig. 16). Trial combinmg non-persistent insecticide + oral IGR: use of of* a &&&ch &see days nuinhg each week for one month, 4th parallel oral adminis- tration of IGR fof threth months (Fig, 13.
Tm-t of hmt amftql perszïj-t inrjec- ticide and topiçal Cspor-ofw) G R ( F i e 18-20) Treaunerrt d kba animai at it la, D 18û and D 240, wkiçh c-afid5 to appksuiun cd thea assaciatkm every ma nurnrhs @ig. 18).
Fig. 15. - Tapical administration of IGR on host animal: adrninis- tration at D 120 and at D 180.
Fig 16. - Trial combiniig non-persistent insecticide + oral IGR on host animai: use of non-persistent insecticide three days ru- each week fm one mmth, wlrh parallel oral administration of IGR.
r insecticick. + oral IGR on a& three days running each
oral administration of IGR for
and aphd mipands io

DISCUSSION includes treating the host animal with insecticide and vacuum cleaning the home premises.
athemritical models are nidely used to study certain populations of insects such as the .tsetse fly, vector of trypanosomes, or mos-
quitoes, vectors of malaria. In veterinary medicine, a number of authors have been interested in inodelling tick populations (Beugnet t.t al., 1998) but no model, simple and able to be used by every-one, has been devised for modelling flea populations. The model described in the present paper takes into account the biological and chronological parameters that have been published concerning Cte~zocepbalides felis (Dryden & Rust, 1994; Kramer & Schencke, 2001). The model confirms the exponential increase in infestation of the home environment premises and of the host animal. This very flexibly constmcted rriodel can handle variations of nuinerous parameters: seasonal variations, impact of type of floor covering, behaviour of host animal who may leave the home preinises for variable periods of time, leaving the contaminated premises in an "on-hold" state. The incidence of the various types of treatment can also be easily factored in. The inodel confirms the major role of nymphs or latent cocoons containing the nenly emerged future fleas (Beugnet & Bourdeau. 1999). It is the resistance of this stage of the cycle that makes its mark on popu- lation dynamics and inakes treatment difficult. For ins- tance, vacuum cleaning of the home premises alone does not entirely solve the problem of contamination of the home environment even though it is highly effective. No chemical treatment carried out as a one- off will eliininate the flea population since the in-flow of new fleas is not considered. The use of an Insect Gromzh Regulator along with an insecticide in the preriiises has a certain impact, but this is limited by the efficacy of this type of treatment which, it is known, does not affect al1 stages of the parasite's development. There can he problems of concentration, of lawae in carpets or flooring, and so forth. The simulations do confirin the need to use persistent insecticides regularly on pet animals as being the best ineans of control (Carlotti & Jacobs, 2000; Dryden et al., 2000). But as soon as the applications stop, the reservoir of nymphs/non-emerged adults starts the whole
. .. cycle off again. The association of a persistent insecticide with an Insect Growth Regulator can, according to the model, result in ~ L I H decontamination of the home environment preinises by depletion of nymphs. However, as the sinlulation shows, it only requires the host animal to move outdoors and to then reintroduce fleas for the decontamination to fail. A combination of the various ineans of control seems to be what is required, as the simulation shows. This
The flexibility of the model should enable the various control strategies, using al1 combinations possible, to be tested in the future under various conditions (infes- tation of the host animal, conditions in the home envi- ronment and behaviour of the host animal), as well as eventual new treatments that may come on to the market. The impact of an eventual anti-flea clccine could also be tested using this model.
REFERENCES
BEITG'IET F. & BOURDEAL: P. Les ectoparasites du chat. Prut. Méd.Chir. Anim. Comp., 1999. 34, 427.435.
BELJGNET F., CHALVFT-MONFRAY K. & SABATIER P. Using of a mathematical model to study the control measures of the cattle tick Boophilus microplus population in Nen- Cale- donia. Veterinnqr Parasitolog): 1998, 77. 277-288.
B ~ ~ r r c o r r ~ ~ u JC. & MÉ'JIER K. Le genre Ctenocephalides Stiies and Collins. 1930 (Siphonaptera, Piilicidae). Parasite, 1998, 5, 3-16.
BOWMAN Q.D. Georgi's Parasitology for Veterinarians. 71h edi- tion. ~df'WE3 Saunders Company, Philadelphia: 1999, 414 p.
CADIERGUES M.c., CAIJBET C. & FRANC M. Cornparison of the activity of selamectin, imidacloprid and fipronil for the treatment of dogs infested experimentally with Ctenoce- phulides canis and Ctenocephalides ,plis felis. Veterinaiy Rec., 2001, 149, 704-706.
CADIERGCES M.C, JOURERT C. & FRANC M. A cornparison of jump performances of the dog flra, Ctenocephalides cunis (Curtis, 1826) and the cat flea, Ctenocep/~ulidesfelis,felis (Bouché, 1835). Vetet?i?nry Parasitology, 2000, 92, 239-241.
CARLOTI D.N. & JACORS D.E. Therapy, control ancl preven- tion of flea allergy dermatitis in dogs and cats. Veterina y De~w~utology; 2000, 11, 83-98.
DRYDEK M.W., DENFKHERG T.M. & BLJNCH S. Control of fleas o n naturally infested dogs and cats and in private resi- dences with topical spot applications of fipronil or imi- dacloprid. Veterinaiy Purasitology, 2000, 93, 69-75,
DRYDEK M.W. & RLIST K.M. The cat flea: biology, ecology and control. Ikterinaty Parasitology, 1994, 52. 1-19.
FRANC M. & CADIEKGLE h1.C. Efficacité d'une association Dichlo- ros-Fénitrothion contre les puces, Ctenocephalides felis, chez le chat. Revue de Médecine Vétérinaire, 1995, 146, 341-344.
FRANC M. & CADIERGL~ES MC. Mode of contamination of dogs by adult fleas (Ctenoephalides,felic) in different controlled environments. Revue de Médecine Vétérinaire, 1997, 148, 23-26.
FRANC M., CHOQLART P. & CADIERG~JES M.L. Répartition des espè- ces d e puces rencontrés chez le chien en France. Revue de Médecine I ijtérinaire, 1998, 149, 135- 140.
GREGORY L.M., ZAKSON hl., ENDRIS R.G. & SHOOP L. A further co~nparison of the thumb-counting and comb-counting techniques used to determine Ctenocephalid~~sfelis infes-
Parasite, 2004, 11, 387-399

tation levels on dogs. Iétel-inary Parasitology, 1995, 56, 349-352.
J ~ c o s s D.E., HUTCHIKSON M.J., Fox M.T. 8 KRIEGER K.J. Com- parison of flea control strategies using imidacloprid or lufe- nuron on cats in a controlled simulated home environ- ment. American Joutwal of Veteritza y Research, 1997, 58, 1260-1262.
J ~ c o s s D.E., HUTCHINSON M.J. & RYAN W.C. Control of flea populations in r i simulated home environment mode1 using lufenuron; imidacloprid or fipronil. Medical and Vetel-inary Entomolog).: 2001, 25, 73-77.
KAIIFMANN J. Parasitic infections of domestic animals. Ed. Bir- khauser. Basel, 1996, 423 p.
KETTLE D.S. Medical and Veterinary Entomology, 2nd Edition. CAB International, Wallingford, 1995, 725 p.
KR~VER F. 8r MENCKE N. Flea Biology and Control. Ed. Sprin- ger, New York, 2001, 192 p.
MÉIZIER K. & BEAUCOURNU J.C. Importance médico-vétérinaire des puces du genre Ctenocephalides Stliles et Collins. 1930. Revue de Médecine Vétérinaire, 1999, 150. 675-680
ROBINSON W.H. Distribution of cat flea larvae in the carpeted household environment. Iéteilnary Dermatology, 1995. 6 (3)) 1+5-150.
SCHENKER R.. TINEMBART O., HUMBERT-DROZ E.. CAVALIERO T. & YERLY B. Comparative speed of kill between nitenpyram. imidacloprid, selanlectin and cythioate against adult Cteno- cephalides,felis. Veterinary Parasitology, 2003, 2520, 1-6.
SHIPSTONE M.A. S; MASON K.V. The use of insect developrnerit inhibitors as an oral medication for the control of the fleas Ctenocephalides felis, Ct. catais in the dog and cat. Vete- r inay Dennatology, 1995. 6(3) , 131-137.
Reçu le 2 avril 2004 Accepté le 8 juillet 2004
Parasite. 2004, 11, 387-399 < < f i





























