Embed Size (px)
Citation preview

1Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
www.nature.com/scientificreports
Venous Endothelial Marker COUP-TFII Regulates the Distinct Pathologic Potentials of Adult Arteries and VeinsXiaofeng Cui1,3,*, Yao Wei Lu4,*, Vivian Lee2,3, Diana Kim2,3, Taylor Dorsey2,3, Qingjie Wang5, Young Lee3, Peter Vincent4, John Schwarz4 & Guohao Dai2,3
Arteries and veins have very different susceptibility to certain vascular diseases such as atherosclerosis and vascular calcification. The molecular mechanisms of these differences are not fully understood. In this study, we discovered that COUP-TFII, a transcription factor critical for establishing the venous identity during embryonic vascular development, also regulates the pathophysiological functions of adult blood vessels, especially those directly related to vascular diseases. Specifically, we found that suppression of COUP-TFII in venous ECs switched its phenotype toward pro-atherogenic by up-regulating the expression of inflammatory genes and down-regulating anti-thrombotic genes. ECs with COUP-TFII knockdown also readily undergo endothelial-to-mesenchymal transition (EndoMT) and subsequent osteogenic differentiation with dramatically increased osteogenic transcriptional program and calcium deposition. Consistently, over-expression of COUP-TFII led to the completely opposite effects. In vivo validation of these pro-atherogenic and osteogenic genes also demonstrates a broad consistent differential expression pattern in mouse aorta vs. vena cava ECs, which cannot be explained by the difference in hemodynamic flow. These data reveal phenotypic modulation by different levels of COUP-TFII in arterial and venous ECs, and suggest COUP-TFII may play an important role in the different susceptibilities of arteries and veins to vascular diseases such as atherosclerosis and vascular calcification.
COUP-TFII (Chicken ovalbumin upstream promoter transcription factor 2), also known as NR2F2 (nuclear receptor subfamily 2, group F, member 2), is an orphan member of the steroid receptor super-family. COUP-TFII is widely expressed in a variety of tissues in the body, and plays an important role in the function and homeostasis of many tissues and organs, such as stem cell differentiation, angiogenesis, lipid/glucose metabolism, and organ development1–5. Because of its widespread functions, COUP-TFII also has been implicated in pathological conditions such as developmental defects, tumor growth and metastasis6–8.
In vascular endothelial cells (ECs), COUP-TFII is expressed at much higher level in venous than arterial endothelium, thus serves as a robust venous EC marker9. Previously, venous cell fate is thought to be the default pathway of vascular development as a result of lacking Notch activation. The landmark discovery of COUP-TFII’s function in vein development demonstrated that it is essential for venous
1School of Chemistry, Chemical Engineering and Life Sciences, Wuhan University of Technology, Wuhan, China 430070. 2Department of Biomedical Engineering, Rensselaer Polytechnic Institute, Troy, NY 12180, USA. 3Center for Biotechnology and Interdisciplinary Studies, Rensselaer Polytechnic Institute, Troy, NY 12180, USA. 4The Center for Cardiovascular Sciences, Albany Medical College, Albany, NY 12208, USA. 5Neural Stem Cell Institute, Rensselaer, NY 12144, USA. *These authors contributed equally to this work. Correspondence and requests for materials should be addressed to G.D. (email: [email protected])
received: 22 June 2015
Accepted: 05 October 2015
Published: 05 November 2015
OPEN

www.nature.com/scientificreports/
2Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
specification via suppressing Notch signaling, suggesting that venous pathway is not a default path-way but under active control of COUP-TFII transcription factor9. Since then, there have been extensive studies on the molecular mechanisms of how COUP-TFII regulates arterial venous marker expression during embryonic vascular development in zebrafish and mouse model10,11. However, little is known about whether these developmental programs lead to the differences in pathophysiologic potential of adult arteries and veins, especially those related to vascular diseases such atherosclerosis and vascular calcification. To investigate whether COUP-TFII has a broader role in regulating adult vascular phe-notypes beyond the arterial venous specification, we performed comprehensive studies on the role of COUP-TFII in adult EC gene expression patterns, focusing on gene pathways that are directly relevant to the atherosclerosis disease and vascular calcification process. Using a model of COUP-TFII knockdown and over-expression in adult human ECs, we identified the COUP-TFII target genes by transcriptional profiling and further characterized functional phenotypes by using additional molecular and cell bio-logical approaches. These findings were subsequently validated in adult blood vessels in animal models. Besides regulating the known arterial and venous markers, we found COUP-TFII is a critical factor in determining multiple gene pathways that are preferentially expressed in adult arterial vs. venous ECs, including extracellular matrix (ECM), inflammation, lipid/metabolism, growth factors, junction proteins, vasomotor factors and thrombosis. Specifically, we found that suppression of COUP-TFII in venous ECs switched its phenotype toward pro-atherogenic by up-regulating the expression of inflammatory genes and down-regulating anti-thrombotic genes. We also discovered that COUP-TFII is a strong regulator of the TGFβ /BMP pathway. ECs with COUP-TFII knockdown readily undergo endothelial-to-mesenchymal transition (EndMT) and subsequent osteogenic differentiation with dramatically increased osteogenic transcriptional program and calcium deposition. Consistently, over-expression of COUP-TFII suppressed all these effects, leading to an anti-atherogenic phenotype and less calcium deposition. These data suggest COUP-TFII may play multifaceted roles in defining the distinctive susceptibilities of adult arteries and veins to vascular diseases such as atherosclerosis and vascular calcification.
MethodsHuman Endothelial Cell Culture. Human saphenous vein endothelial cells (HSVECs, VEC TECHNOLOGIES, INC.) were cultured at 37 °C in 5% CO2 in Endothelial Cell Growth Medium-2 (EGM-2, Lonza). The culture media was changed every two days. HSVECs were routinely passaged onto tissue culture flasks and discarded after 8 passages to ensure representation of key endothelial charac-teristics. For each experiment, EC were plated at an initial density of 50,000 cells/cm2 on 0.1% gelatin (Sigma) coated tissue culture plate. After 24 hours, the confluence of each EC monolayer was verified by phase contrast examination. To validate that COUP-TFII regulated gene patterns are also true in other endothelial cell types, human aortic endothelial cells (HAEC, Lonza) were cultured and selected genes were validated using RT-PCR (supporting Figure S4).
Lentivirus to Over-express COUP-TFII. The human Nr2f2 (COUP-TFII) cDNA was ampli-fied from pCMV6-XL5-Nr2f2 (OriGene) by PCR, then sub-cloned into pCDH-EF1-T2A-GFP lentiviral vector (System Biosciences) between EcoRI and NotI digestion sites. Sequences of the PCR primer pair are: Forward, GGAATTCGCCACCATGGCAATGGTAGTCAGCAC; Reverse, ATTTG CGGCCGCTTGAATTGCCATATACGGCCA. To package the lentivirus, the pCDH-EF1-Nr2f2-T2A-GFP or the pCDH-EF1-T2A-GFP constructs were co-transfected with pCMV-VSVG and pCMV-dvpr into 293FT cells using X-tremeGENE HP transfection reagent (Roche). Supernatant was harvested 2 and 3 days later and further concentrated by ultra-centrifugation. The virus pellet was re-suspended in PBS and aliquoted, then put into − 80 C for storage. Lentiviruses were used at 10 MOI for cell transduc-tion, and undergo puromycin (2 μ g/ml) selection to establish stable ECs that permanently over-express COUP-TFII.
Isolate Fresh RNA from Endothelium of Mouse Aorta and Vena Cava and cDNA synthesis. Endothelium RNA from mouse aorta and vena cava was isolated using protocol previously described with minor modification12. Briefly, mice were euthanized with intra-peritoneal injection of sodium pentobarbital according to protocol approved by Albany Medical College’s Institutional Animal Care and Use Committee (IACUC). Animal’s vasculature was pressure perfused with saline solution with 10 U/ml heparin for 2 minutes via left ventricle after severing inferior vena cava. Aorta and inferior vena cava were isolated, and peri-adventitial fat were carefully cleaned in ice-cold PBS. The lumen of the vessels was quickly flushed with 300 ul ice-cold TRIzol reagent (Invitrogen) using 25 G needle into a microcen-trifuge tube. Total RNA were isolated and purified from the elute using RNeasy Plus Kit (Qiagen), and subsequent cDNA synthesis using Superscript III First-Strand Synthesis system. To verify the purity of the RNA (without smooth muscle cells contamination), gene expression of alpha-smooth muscle actin (α -SMA) was assessed on the RNA sample and compared to the RNA isolated from the media layer of the blood vessel segment. The purify of the RNA was validated to have < 1% α -SMA contamination (supporting Figure S5).
En face Immunofluorescence Microscopy of Mouse Aorta and Vena Cava. En face immu-nofluorescence microscopy of mouse aortic arch was performed as described previously13. FVB/NJ

www.nature.com/scientificreports/
3Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
mice were purchased from Jackson Laboratory (Bar Harbor, ME) and maintained in accordance with Institutional Review Board approved protocols. To examine cytokine induced adhesion molecule VCAM-1 expression, TNF-a (30 ug/kg body weight, ~1 ug per mouse) was injected via tail vein of the mouse based an established protocol14. After 16 hour, mice were anesthetized and perfused with PBS and 2% paraformaldehyde, aorta and vena cava were then harvested. The vessel was then cut open and suspended in PBS followed by permeabilization in 0.1% Triton X-100/PBS. After blocking with 5.5% FBS in 0.1% Triton X-100/PBS, the aorta was incubated with either mouse anti-BMP4 antibody (1:100; Santa Cruz Biotechnology), or rabbit anti-Cx40 antibody (1:100; Alpha Diagnostic International), or FITC conjugated rat anti–mouse VCAM-1 (1:100; BD Pharmingen). Endothelium was co-stained by goat anti-VE-cadherin antibody (1:100; Santa Cruz Biotechnology) in PBS at 4 °C overnight. Secondary antibodies included Alexa Fluo 488 donkey anti-mouse (1:200; Life Technologies), Alexa Fluo 488 don-key anti-rabbit (1:200; Life Technologies) Alexa Flur 594 donkey anti mouse (1:200, Life Technologies) and Alexa Fluo 647 donkey anti-goat (1:200, Life Technologies), Alexa Fluor 488 goat anti–rat antibody (1:200 Life Technologies).
To identify nuclei, DAPI (1:10,000, Life Technologies) was added during the incubation of secondary antibodies. For the aorta, the lesser curvature of the aortic arch and a portion of thoracic aorta were cut and mounted with the endothelium facing up. Images of the en face preparation were obtained using a Zeiss LSM META 510 confocal microscope system with Zeiss Zen software, or Zeiss Axio Observer Z1 inverted microscope with the Apotome module and Zeiss AxioVision software.
Assay the Osteogenic Potential of Cultured ECs. To assay the osteogenic potential of cultured HSVECs, the culture medium was switched from EGM-2 to StemPro osteogenesis differentiation medium (Life Technologies). After 7 days of culture, RNA was isolated and Taqman RT-PCR was performed to assess the gene expression involved in osteogenic pathway: RUNX2, SP7, DLX5, ALPL, COL1A1, IBSP, BGLAP, SPP1 and BMP4. To further validate the functional consequences of the osteogenic potential, Alkaline Phosphatase (ALP) and Alizarin Red S staining were performed on cells following 7 days of culture under osteogenic differentiation medium. Briefly, for Alkaline Phosphatase staining, cells were rinsed twice by PBS and then fixed using 1 mL 10% formaldehyde in each well for 1 min (fixing cells longer than 2 min will result in the inactivation of alkaline phosphatase). After the fixation, the fixative was aspirated and the cells were rinsed again using PBS. ALP activities were detected by SigmaFast BCIP/NBT (Sigma-Aldrich) following the manufacturer protocol. For Alizarin Red S staining, cells were rinsed twice using calcium and magnesium free PBS and then fixed using 1 mL 10% formaldehyde in each well for 10 min. Cells were then washed twice using PBS and stained using an Alizarin Red S kit (American MasterTech) to evaluated the calcium deposit following the manufacturer protocol.
Additional materials and methods are described in detail in Supporting Information.
ResultsCOUP-TFII Regulates Multiple Gene Pathways Related To Atherosclerosis In Adult ECs. We first applied RNAi to knockdown COUP-TFII expression in cultured human saphenous vein endothelial cells (HSVEC), and performed transcriptional profiling of the genes that are differentially expressed. As shown in Table S1 (in Supporting Information), knockdown of COUP-TFII leads to the increase of selected arterial markers such as Ephrin-B2, Hey1,2, Dll1,4, Flt1, Notch4, Jag1, Hes4, Foxc2, which is consistent with the role of COUP-TFII in venous specification as reported before9. The regulation pattern of arterial markers (Ephrin-B2, Hey2, DLL4) by COUP-TFII in cultured adult ECs also confirmed the recent study, in which COUP-TFII negatively regulates the Ephrin-B2, Hey2 and DLL4 expression15 in human umbilical artery endothelial cells. Several genes involved in angiogenesis/vasculogenesis are also up-regulated including EPAS1/HIF-2a, Ephrin-A1, Tie-2, EphA4. The up-regulated genes also include several previously unknown targets including Sema7a (Semaphorin 7A), a molecule involved in the axon guidance, and Vash1 (Vasohibin 1), an angiogenesis inhibitor. Gap junction proteins (Connexin 37 & 40) are also strongly up-regulated. Meanwhile, knockdown of COUP-TFII reduces the Nrp2, a venous EC marker. We validated several selected arterial and venous markers by RT-PCR, shown in Fig. 1. Overall, knockdown of COUP-TFII switches on the arterial EC markers, suggesting that COUP-TFII not only plays a role during development, but also is important for maintaining the venous identity in adult ECs.
Importantly, we found large number of chemokine/inflammatory genes are up-regulated as well as several genes involved in thrombosis (supporting Table S1). To validate this finding, we performed RT-PCR analysis on several key genes known to be important for pro- or anti-atherogenic process. As shown in Fig. 1C, COUP-TFII knockdown lead to a decrease in TFPI2 (tissue factor pathway inhibitor 2), an anti-thrombotic gene, while increased the expression of VCAM-1 (vascular cell adhesion mole-cule), E-selectin, PAI-1 (plasminogen activator inhibitor 1), ECE (endothelin converting enzyme), ET-1 (endothelin 1), which are pro-atherogenic genes.
The overall pattern of pro-/anti-atherogenic gene expression suggests that the ECs with COUP-TFII knockdown switch to a more pro-atherogenic phenotype. To further validate this, we assessed inflam-matory cytokine induced VCAM-1, the adhesion molecule involved in leukocyte-endothelial interac-tion in early atherosclerosis and is essential for disease progression16. As shown in Fig. 1D, COUP-TFII knockdown increased VCAM-1 expression at the basal level and also under low dose TNF-α stimulation. These data suggests that suppression of COUP-TFII in ECs makes them more sensitive to inflammatory

www.nature.com/scientificreports/
4Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
stimuli. To further evaluate the functional significance of COUP-TFII knockdown in disease process, we performed leukocyte-endothelial adhesion assay. Shown in Fig. 1E, decreased COUP-TFII in ECs resulted in an increased THP-1 cell adhesion to EC surface under low dose of TNF-α (0.01 ng/ml) stimulation.
Figure 1. COUP-TFII not only controls molecular markers of arterial and venous identity, but also regulates EC genes of inflammation, thrombosis and vasomotor functions. (A–C) HSVECs were transfected with COUP-TFII or Control RNAi (10 nM). RNA was collected 72 hours after transfection. Gene expressions were measured by Taqman-RT PCR and normalized to RNA 18S. Data are represented as relative expression level, n = 3, *p < 0.05. (D) HSVECs were transfected with COUP-TFII or Control RNAi (10 nM) for 48 hours, and then treated with different doses of TNF-α (0–1 ng/ml) for 24 hours. VCAM-1 gene expressions were measured by Taqman-RT PCR and normalized to RNA 18S, n = 3. (E) HSVECs were treated with 0.01 ng/ml TNF-α for 24 hours, and THP-1 cells (labeled with CellTracker Green) adhesion to the EC surface was quantified by counting number of attached cells within each imaging field and averaged over 10 imaging fields. *p < 0.05.

www.nature.com/scientificreports/
5Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
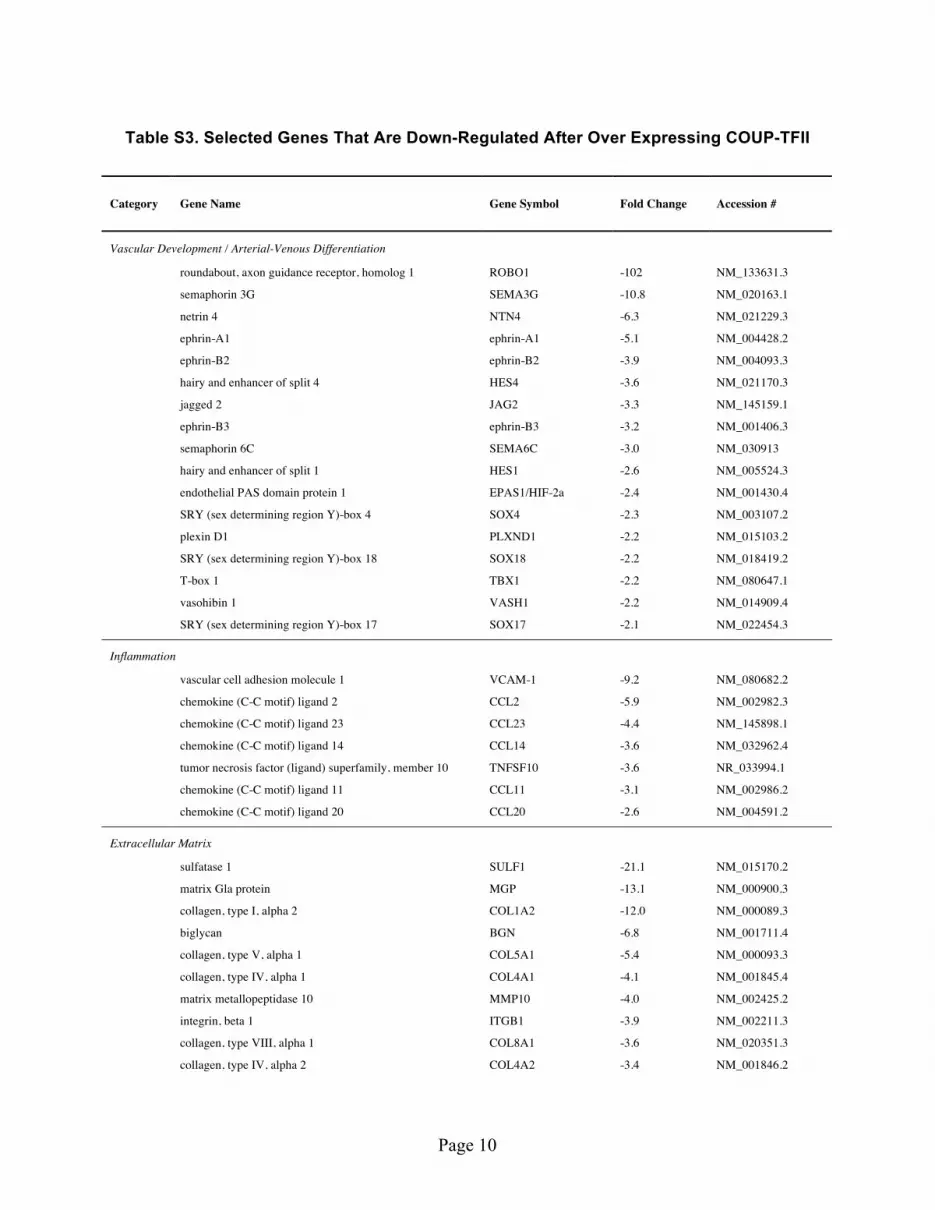
To further understand the functions of COUP-TFII in adult ECs, we made lentivirus over-expressing COUP-TFII and used it to generate confluent EC monolayer that stably expresses COUP-TFII. The transcriptional profiling analysis (supporting Table S3) revealed that COUP-TFII suppresses the known arterial marker expression, such as Ephrin-B2, Hes1/4. Multiple genes involved in angiogenesis/vascu-logenesis are also suppressed, including Ephrin-A1, Ephrin-B3, HIF-2a, Sox4/17/18, Tbx1 and Vash1. Interestingly, many molecules involved in nerve development/axon guidance were also suppressed, such as Robo1, Netrin, Semaphorin and Plexin D1, which is consistent with the notion that nerve develop-ment guides the arterial differentiation17,18 whereas COUP-TFII suppresses these signals and leads to venous specification. Meanwhile, COUP-TFII up-regulated Lyve1 (lymphatic vessel endothelial hyaluro-nan receptor 1), a known marker for lymphatic ECs, consistent with the finding that lymphatic ECs are derived from venous ECs but not arterial ECs19,20.
COUP-TFII also suppresses or promotes distinct set of genes involved in the biosynthesis and remod-eling of ECM. Besides various types of collagen, laminin, integrin and fibronectin, many ECM compo-nents and enzyme pathways are differentially regulated, including sulfatase, biglycan, versican, syndecan and matrix Gla protein (Supporting Table S3, S4). These data suggest that COUP-TFII may contribute to the distinct difference in the structural composition of arteries and veins. In agreement of the data that COUP-TFII knockdown increases inflammation and thrombosis, COUP-TFII over-expression sup-pressed inflammation and thrombotic gene profile (supporting Table S3) (e.g., VCAM-1, von Willebrand factor, thrombospondin, multimerin), and increased anti-thrombotic gene expression (supporting Table S4). We did additional validation using RT-PCR on selected genes. Shown in Fig. 2B, COUP-TFII increased tPA (tissue plasminogen activator) and TFPI2, both of which are anti-thrombotic, and sup-pressed the expression of PAI-1, VCAM-1, E-selectin, ET-1, ECE-1, PDGF-β (platelet derived growth factor beta), ACE (angiotensin converting enzyme), BMP4 (bone morphogenetic protein 4), all of which are pro-atherogenic genes.
To further validate these findings, we examined the protein expression of COUP-TFII, BMP4 and VCAM-1. As shown in Fig. 3C, COUP-TFII RNAi completely knockdown the COUP-TFII at the protein level, whereas lentivirus over-expressing COUP-TFII increased the COUP-TFII protein, indicating the efficiency of the COUP-TFII knockdown and over-expression. Cytokine induced VCAM-1 expression demonstrates an opposite pattern of COUP-TFII expression (Fig. 3D): COUP-TFII suppressed TNF-α
Figure 2. Over-expressing COUP-TFII suppresses arterial markers and promotes an athero-protective gene profile. (A,B) HSVECs were infected with lentivirus-COUP-TFII or control lentivirus (10 MOI), RNAs were isolated after confluent HSVEC monolayer is reached and all cells were permanently infected with lentivirus. Gene expressions were measured by Taqman-RT PCR and normalized to RNA 18S, n = 3, *p < 0.05.

www.nature.com/scientificreports/
6Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
stimulated VCAM-1 protein, suggesting COUP-TFII has anti-inflammatory effect in ECs. Similarly, the BMP4 expression demonstrated opposite direction to COUP-TFII level (Fig. 3C): it increased in COUP-TFII knockdown and decreased in COUP-TFII over-expression, suggesting COUP-TFII is a neg-ative regulator of BMP4. Because COUP-TFII demonstrates a strong regulation pattern on BMP4, we assessed whether COUP-TFII directly regulates BMP4 at the transcriptional level by ChIP (Chromatin Immunoprecipitation)-PCR. As shown in Fig. 3E, in both the control and COUP-TFII over-expression group, the immunoprecipitated chromatin using the COUP-TFII antibody demonstrates an enrichment of the BMP4 promoter compared to IgG control, suggesting that COUP-TFII may repress the BMP4 expression by binding directly to the BMP4 promoter region.
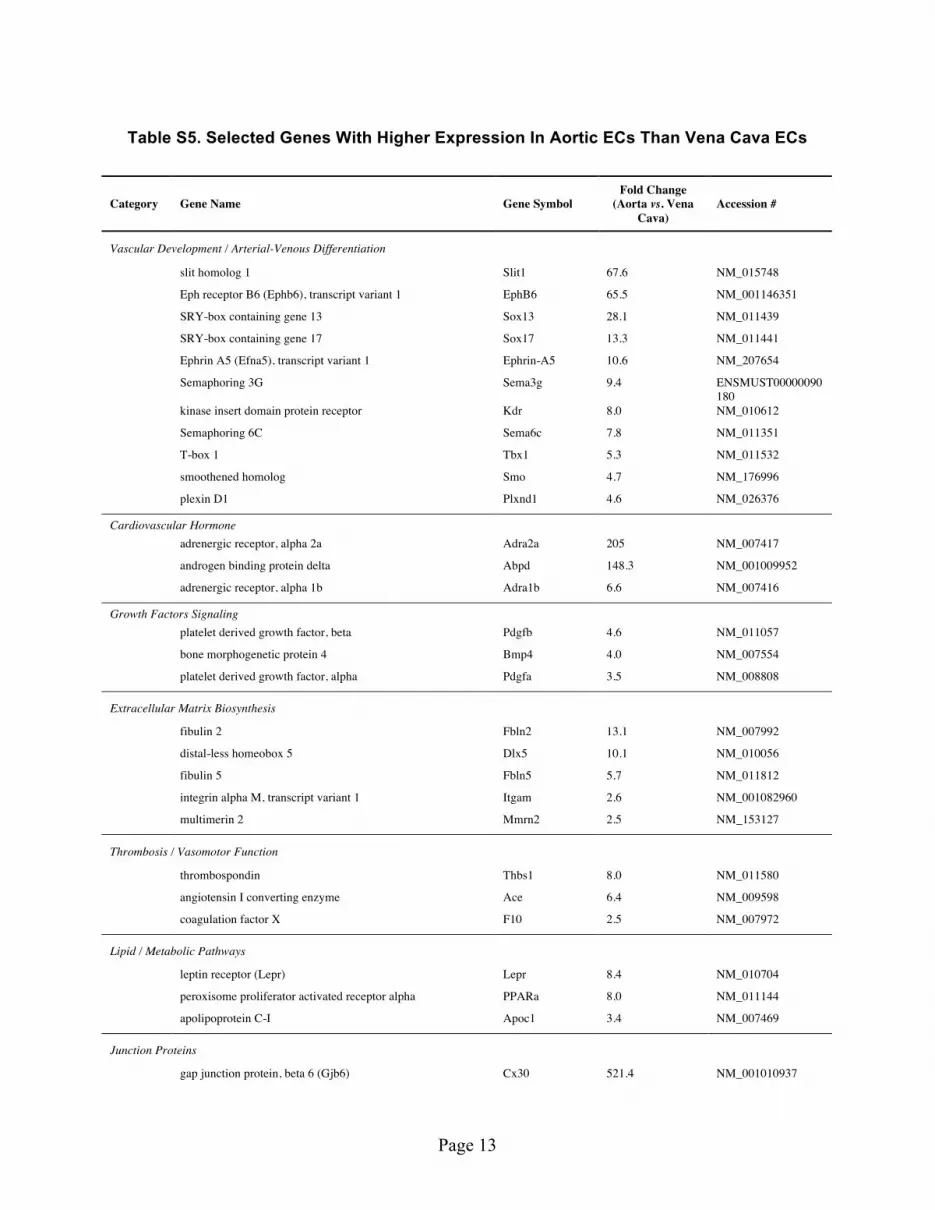
Adult Arterial and Venous ECs Demonstrate Dramatic Differences in Gene Regulation Patterns and Response to Inflammatory Stimuli. To analyze the in vivo gene expression patterns, we devel-oped the techniques to freshly isolate pure EC RNA (smooth muscle cell contamination < 1%, supporting Figure S5) directly from intact mouse aorta and vena cava according to protocol approved by Albany Medical College’s Institutional Animal Care and Use Committee (IACUC). Analysis of transcriptional profiles showed that the fresh isolated EC RNA was able to re-confirm many of the known arterial venous marker expression patterns (Supporting Table S5, S6, Fig. 4A)21. We identified many new targets that are significantly expressed in arterial or venous ECs (Supporting Table S5, S6, Supporting Figure S1). The differences are dramatic; some of the genes are expressed more than 10–100 fold in arterial or venous ECs. Noticeable targets include Cx30/40, Adra2a (adrenergic receptor, alpha 2a), Abpd (androgen
Figure 3. COUP-TFII regulates Cx40, BMP4 and VCAM-1 at protein level. (A) HSVECs were transfected with COUP-TFII or Control RNAi, and Cx40 was detected with immunofluorescent staining. (B) Fluorescent images of HSVECs transfected with lenti-Control-GFP or lenti-COUP-TFII-GFP. (C) Western blot analysis of BMP4 and COUP-TFII in HSVECs after COUP-TFII knockdown or over-expression. (D) HSVECs were treated with TNF-α (0.01 ng/ml) for 24 hours, western blot analysis of VCAM-1 expression was performed. (E) ChIP-PCR assays on cultured HSVECs using anti-COUP-TFII antibody (solid bar) or IgG (open bar). Bar graphs show enrichment of DNA fragments pulled down by COUP-TFII antibodies, *p < 0.01.

www.nature.com/scientificreports/
7Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
binding protein delta), Slit1, EphB6, Sox13/17, Fibulin 2, Ephrin-A5, Dlx5, which are highly expressed in artery, and Nppa/c (natriuretic peptide A & C), Tbx5, Claudin 11, Wnt2/9b, Apelin and COUP-TFII, which are highly expressed in vein. Importantly, we were able to confirm the pattern of COUP-TFII regulation related to thrombosis/atherogenesis: the expression of anti-thrombotic molecules tPA, TFPI2, uPA (urokinase-type plasminogen activator) is higher in vein, whereas pro-atherogenic molecules PAI-1, VCAM-1, ET-1, PDGF-α /β , BMP4 and ACE are higher in artery (Fig. 4B)
One striking finding is gap junction protein Cx40, which has ~300 fold higher expression in arterial than venous ECs (Fig. 4A). The regulation by COUP-TFII is also consistent with this finding: COUP-TFII knockdown lead to a dramatic induction (> 160 fold) of Cx40 (Fig. 1A), whereas it suppressed Cx40 when COUP-TFII is over-expressed (Fig. 2A). As shown in Fig. 3A, cultured HSVEC has very little Cx40 expression, which increased dramatically upon COUP-TFII knockdown. The en face immunofluorescent staining of mouse aorta and vena cava (Fig. 5B) further confirm this: Cx40 is completely absent in ECs of vena cava while it is abundantly located in the cell-cell junctions in both the straight and curvature portion of the aorta. Taken together, these data suggest that COUP-TFII is a strong regulator of gap junction protein Cx40 expression.
Using en face immunofluorescent staining technique, we also confirmed the BMP4 expression in artery and vein, and found BMP4 only appeared in the endothelial layer but not in the smooth muscle layer of the aorta (supporting Figure S2). As shown in Fig. 5A, BMP4 is highly expressed in the ECs of aortic curvature, and the expression is lower in the straight portion but clearly visible in some cells. In contrast, BMP4 is completely absent in the vena cava. This pattern of expression is consistent with the in vivo gene expression data and also the in vitro COUP-TFII regulation patterns, suggesting that COUP-TFII is a strong negative regulator of the BMP4 expression.
To further assess the phenotypic differences of adult arterial and venous ECs, we examined the cytokine induced adhesion molecule VCAM-1 expression, which has been demonstrated to play an essential role in the initiation and progression of atherosclerosis16. TNF-a was injected into the mice as the inflammatory stimuli, and vascular expression of VCAM-1 was assessed via en face confocal immu-nofluorescent staining. As shown in Fig. 6, VCAM-1 staining is obvious across various parts of the arter-ies. In the inner curvature of the aortic arch where blood flow is disturbed, VCAM-1 appears as multiple punctuated staining throughout most regions, while the staining is more diffused and occurs in a few isolated cells in the thoracic aorta and infra-renal aorta. In contrast, VCAM-1 staining is completely
Figure 4. In vivo arterial and venous ECs demonstrate distinct profiles of certain molecular markers (A) and atherosclerosis related genes (B). RNA was isolated from the endothelium layer of mouse aorta and vena cava. Gene expression was analyzed by Taqman RT-PCR (n = 3), *p < 0.05.

www.nature.com/scientificreports/
8Scientific RepoRts | 5:16193 | DOi: 10.1038/srep16193
absent in the thoracic vena cava and infra-renal vena cava, suggesting that adult artery and vein ECs have very different response to inflammatory stimuli.
To identify differentially expressed genes in arteries and veins that are also regulated by COUP-TFII, we compiled the list of genes that demonstrated consistency between in vitro COUP-TFII regulation
Figure 5. BMP4 and Cx40 protein is expressed in mouse arteries but not in veins. en face confocal immunofluorescent images of BMP4 (A) and Cx40 (B) expression in the endothelium layer of mouse aorta (straight portion and curvature) and vena cava.

www.nature.com/scientificreports/
9SCIENTIfIC REPORTS | 5:16193 | DOI: 10.1038/srep16193
and in vivo gene expression, and listed them in Fig. 7. Many arterial and venous genes are regulated by COUP-TFII, such as Cx37/40, Ephrin-B2, Notch4, Dll4, Tbx1, Sox17, HIF-2a, Sema3g, Vash1, Plxnd1, Nrp2 and Lyve1. Importantly, the selected anti- and pro-atherogenic genes are differentially expressed in vivo and they are also regulated by COUP-TFII: TFPI2 and tPA are higher in venous ECs while PAI-1, VCAM-1, ACE, BMP4, PDGFβ and Thrombospondin are higher in arterial ECs. Similar pattern of expression also includes genes in lipid/metabolic pathway such as PPAR-α and leptin receptor. Overall, arterial ECs demonstrated a more pro-atherogenic profile than venous ECs, and this difference is con-sistent with the in vitro COUP-TFII regulation data.
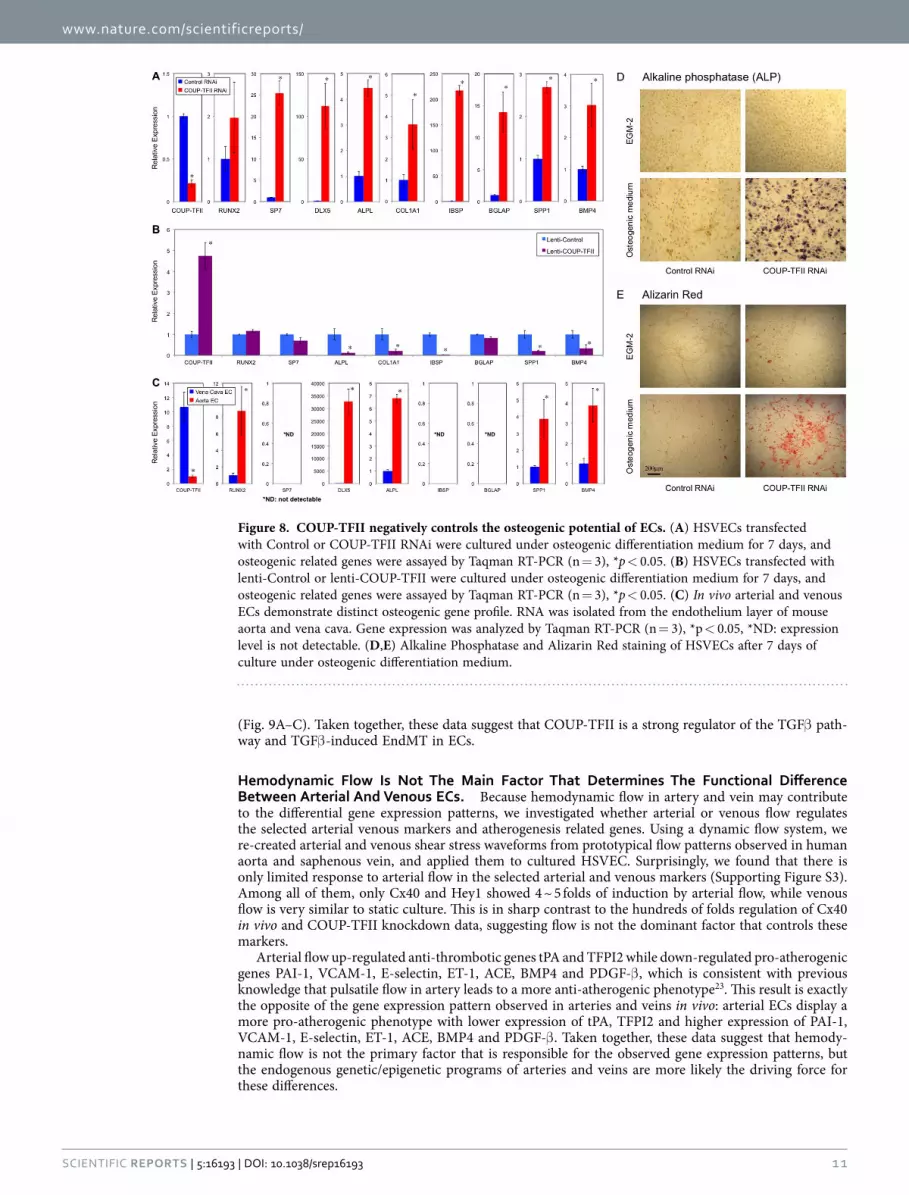
COUP-TFII Regulates Endothelial-To-Mesenchymal Transition And Osteogenic Programs. Because several genes involved in bone homeostasis were regulated by COUP-TFII or showed differential expression in artery and vein (Supporting Table S3, S5), such as BMP4, MGP (matrix Gla protein) which is involved in calcium regulation and Dlx5 (distal-less homeobox 5) which acts as the early BMP-responsive transcriptional activator needed for osteoblast differentiation, we hypothesize that COUP-TFII regulates the osteogenic potential of ECs. To investigate the functional significance of COUP-TFII in this process,
Figure 6. VCAM-1 is expressed in arteries but not veins in response to TNF-a stimulation. TNF-a was injected into mice for 16 hours. en face confocal immunofluorescent images of VCAM-1 expression in the endothelium layer of mouse arteries (inner-curvature of aortic arch, thoracic aorta, infra-renal aorta) and vena cava (thoracic vena cava and infra-renal vena cava).

www.nature.com/scientificreports/
1 0SCIENTIfIC REPORTS | 5:16193 | DOI: 10.1038/srep16193
we treated the cells with BMP4 and switched the culture conditions to osteogenic medium, and assessed the osteogenic related programs in ECs with COUP-TFII knockdown and over-expression. Shown in Fig. 8A, in COUP-TFII knockdown condition, ECs readily undergo osteogenic differentiation with dra-matically up-regulated genes involved in bone formation, such as Sp7 (osterix), Dlx5, ALPL (alkaline phosphatase), COL1A1 (collagen, type I, alpha 1), IBSP (integrin-binding sialoprotein), BGLAP (bone gamma-carboxyglutamate (gla) protein or osteocalcin), SPP1 (secreted phosphoprotein 1 or osteopontin) and BMP4. The effect is so significant that some of them have more than 10 or 100 fold of induction, suggesting that COUP-TFII in ECs acts as a strong repressor of its osteogenic potential in normal condi-tion, and upon removal, osteogenic programs are greatly increased. Consistent with this, over-expression of COUP-TFII suppressed these genes: ALPL, COL1A1, IBSP, SPP1, BMP4 (Fig. 8B). Furthermore, bio-chemical assays demonstrated significant induction of alkaline phosphatase activity and calcium deposi-tion in ECs cultured under osteogenic medium when COUP-TFII is knockdown (Fig. 8D,E). To further validate whether these osteogenic genes are also differentially expressed in vivo, we confirmed their expression pattern in ECs of mouse aorta and vena cava, and found that the expression of RUNX2, DLX5, ALPL, SPP1 and BMP4 are significantly higher in aortic ECs than vena cava ECs (Fig. 8C). Because upregulation of the osteogenic programs in ECs involves TGFβ /BMP signaling and the sub-sequent osteogenic differentiation, it is possible that TGFβ /BMP induced endothelial-to-mesenchymal transition (EndMT) is also affected. To test whether COUP-TFII regulates the EndMT, we cultured ECs under TGFβ 2 treatment, which is a known inducer of EndMT. Transcriptional profile analysis revealed that several critical genes involved in the EndMT process are significantly induced when COUP-TFII is knockdown (Supporting Table S7). These changes collectively suggest an enhanced EndMT process22. Gene ontology enrichment analysis revealed that TGFβ pathway is particularly affected by COUP-TFII, including SMAD6, FST (follistatin), INHBA (inhibin, beta A, a subunit of activin and inhibin), TGFβ 2 and TGFBI (transforming growth factor, beta-induced, 68 kDa) (Supporting Table S7). The most signif-icantly up-regulated gene is TGFBI, which plays a role in cell-collagen interactions and is implicated in the endochondrial bone formation in cartilage. Shown in Fig. 9A, TGFβ 2 or BMP4 treatment is a very weak inducer of EndMT in HSVECs. However, when COUP-TFII is knockdown by RNAi, the EndMT is significantly induced, demonstrated by increased expression of α -SMA, SM22α , FSP-1, Snail and Slug
Figure 7. COUP-TFII controls the differential expression pattern of selected genes in artery vs. vein. Patterns of selected gene expression in COUP-TFII knockdown, COUP-TFII over-expression and in vivo arterial and venous ECs are color-coded. Genes that demonstrate consistent patterns of in vitro regulation (either knockdown or over-expression) with those in vivo expression patterns are selected.
www.nature.com/scientificreports/
1 1SCIENTIfIC REPORTS | 5:16193 | DOI: 10.1038/srep16193
(Fig. 9A–C). Taken together, these data suggest that COUP-TFII is a strong regulator of the TGFβ path-way and TGFβ -induced EndMT in ECs.
Hemodynamic Flow Is Not The Main Factor That Determines The Functional Difference Between Arterial And Venous ECs. Because hemodynamic flow in artery and vein may contribute to the differential gene expression patterns, we investigated whether arterial or venous flow regulates the selected arterial venous markers and atherogenesis related genes. Using a dynamic flow system, we re-created arterial and venous shear stress waveforms from prototypical flow patterns observed in human aorta and saphenous vein, and applied them to cultured HSVEC. Surprisingly, we found that there is only limited response to arterial flow in the selected arterial and venous markers (Supporting Figure S3). Among all of them, only Cx40 and Hey1 showed 4 ~ 5 folds of induction by arterial flow, while venous flow is very similar to static culture. This is in sharp contrast to the hundreds of folds regulation of Cx40 in vivo and COUP-TFII knockdown data, suggesting flow is not the dominant factor that controls these markers.
Arterial flow up-regulated anti-thrombotic genes tPA and TFPI2 while down-regulated pro-atherogenic genes PAI-1, VCAM-1, E-selectin, ET-1, ACE, BMP4 and PDGF-β , which is consistent with previous knowledge that pulsatile flow in artery leads to a more anti-atherogenic phenotype23. This result is exactly the opposite of the gene expression pattern observed in arteries and veins in vivo: arterial ECs display a more pro-atherogenic phenotype with lower expression of tPA, TFPI2 and higher expression of PAI-1, VCAM-1, E-selectin, ET-1, ACE, BMP4 and PDGF-β . Taken together, these data suggest that hemody-namic flow is not the primary factor that is responsible for the observed gene expression patterns, but the endogenous genetic/epigenetic programs of arteries and veins are more likely the driving force for these differences.
Figure 8. COUP-TFII negatively controls the osteogenic potential of ECs. (A) HSVECs transfected with Control or COUP-TFII RNAi were cultured under osteogenic differentiation medium for 7 days, and osteogenic related genes were assayed by Taqman RT-PCR (n = 3), *p < 0.05. (B) HSVECs transfected with lenti-Control or lenti-COUP-TFII were cultured under osteogenic differentiation medium for 7 days, and osteogenic related genes were assayed by Taqman RT-PCR (n = 3), *p < 0.05. (C) In vivo arterial and venous ECs demonstrate distinct osteogenic gene profile. RNA was isolated from the endothelium layer of mouse aorta and vena cava. Gene expression was analyzed by Taqman RT-PCR (n = 3), *p < 0.05, *ND: expression level is not detectable. (D,E) Alkaline Phosphatase and Alizarin Red staining of HSVECs after 7 days of culture under osteogenic differentiation medium.

www.nature.com/scientificreports/
1 2SCIENTIfIC REPORTS | 5:16193 | DOI: 10.1038/srep16193
DiscussionIn this study we investigated the role of COUP-TFII in regulating adult EC functions, focusing on their significance in vascular diseases. We aim to examine the multifaceted roles of COUP-TFII in vascular disease by comprehensive analysis of EC gene expression and phenotypes using a model of COUP-TFII knockdown and over-expression in adult human ECs, and subsequent validation in adult blood ves-sels in animal models. We have identified differential patterns of gene expression in adult arterial and venous ECs, and confirmed that some of those genes are under the control of COUP-TFII. Many of the differentially regulated genes have pathophysiological functions in atherosclerosis initiation and/or progression, thus demonstrating the importance of COUP-TFII in regulating adult EC functions. In particular, venous ECs with COUP-TFII knockdown acquire some of the arterial gene expression patterns and a pro-inflammatory phenotype, expressing several important molecules in atherogenesis and lowering several anti-thrombotic genes. It also elicited an enhanced cytokine-inducible cell surface expression of the atherosclerosis-associated adhesion molecule VCAM-1 and leukocyte adhesion to EC surface. Importantly, we also discovered that COUP-TFII not only regulates the expression of BMP4 and TGFβ 2, but also affects multiple signaling molecules in the BMP4/TGFβ pathway. BMP signaling is known to promote vascular calcification, and others have reported that limiting vascular BMP signaling decreases both atherosclerotic lesion and diabetic medial calcification24. In addition, we have observed an enhanced EndMT and strikingly increased osteogenic potential of ECs when COUP-TFII is knockdown. Consistently, over-expression of COUP-TFII resulted in the totally opposite effects. Finally, we validated these findings in ECs of mouse aorta and vena cava in vivo. Taken together, these data strongly implicate the modulation of endothelial phenotype by venous marker COUP-TFII in the vascular system.
Specification of vascular progenitors into arterial and venous ECs is an essential process of vascular development9,25–32. Several signaling pathways are involved in the arterial-venous differentiation, among which the Notch signaling plays a central role29,30,33,34. Blood flow is another critical factor in determin-ing arterial differentiation35. Lack of Notch activation or blood flow was thought to be the reason for venous differentiation. However, the discovery of COUP-TFII demonstrated that COUP-TFII functions as a key player that confers vein identity by repressing Notch signaling9. Recent study also confirmed that over-expression of COUP-TFII suppresses Notch pathway in cultured human umbilical artery
Figure 9. COUP-TFII negatively regulates EndMT. (A) HSVECs transfected with Control or COUP-TFII RNAi were cultured under EGM-2 (Control), or endothelial medium without VEGF and with TGFβ 2 (10 ng/ml) or BMP4 (10 ng/ml) for 7 days, and gene expression was assayed by Taqman RT-PCR (n = 3), *p < 0.05. (B) Immunofluorescent staining of α -SMA. (C) Immunofluorescent staining of Slug.

www.nature.com/scientificreports/
13SCIENTIfIC REPORTS | 5:16193 | DOI: 10.1038/srep16193
endothelial cells15. Our data demonstrated that COUP-TFII not only plays a role in regulating arterial venous markers, but also control important functions in adult ECs, suggesting that these developmental programs also contribute toward the adult blood vessel functions in the pathogenesis of vascular diseases.
Although blood flow plays an important role in arterial EC specification during early vascular devel-opment, arterial markers in adult blood vessel are less responsive to hemodynamic flow after the vascu-lature is fully developed. When placed in arterial circulation, vein graft undergoes “arterialization”36,37 in which smooth muscle cells adapted to higher pressure and cyclic strain in responses to the new milieu. In ECs, venous identity (EphB4) is lost during this process, but arterial identities (Ephrin-B2, DLL4, Notch4) are not gained, suggesting an incomplete adaptation of vein graft in adult patients38,39. Our data also confirmed that the arterial-venous markers in adult ECs are less responsive to flow condition. We found that adult EC has very limited response to flow, and most arterial and venous markers do not change except Cx40 and Hey1. The relatively small change of Cx40 is also far from the hundreds fold differences observed in artery vs. vein in vivo. This result demonstrates that adult ECs lose some of their plasticity and are not able to completely adapt to arterial circulatory conditions.
We have also identified many new molecular markers that are highly expressed in adult artery or vein. Some of these markers show more than 10–100 fold of differential expression, and therefore, can be used as a more robust marker to identify artery vs. vein than those with only 2 ~ 3 folds of difference. Surprisingly, some of the well-established arterial markers (Nrp1, Hey1, Jag1, Dll1) are not differentially expressed in adult arteries vs. veins, although they have been used extensively in the past as the proof of arterial differentiation in many developmental biology studies. It is likely that they appear as early arte-rial marker during vascular development, and then the difference disappears after maturation into adult blood vessel. Interestingly, although COUP-TFII controls many arterial venous markers, EphB4 seems not influenced by COUP-TFII at all. This suggests that other additional unknown endogenous programs are involved in venous specification besides COUP-TFII.
In arteries, atherosclerosis often develops in regions of disturbed flow, which promotes a pro-inflammatory phenotype of ECs23,40,41. In contrast, laminar pulsatile flow in straight portion of arteries leads to an anti-inflammatory and anti-thrombotic phenotype, therefore, is considered athero-protective42–44. Unlike in arteries, blood flow in veins is not pulsatile but phasic with low average wall shear stress (< 1 dyn/cm2). Based on our previous extensive knowledge of flow-regulated EC func-tions, this type of flow is not considered “athero-protective” since it has very small influence on EC gene expression (compared to static control) whereas the influence of laminar pulsatile arterial shear is much larger. Despite this unfavorable hemodynamic condition, veins rarely develop certain types of vascular diseases such as atherosclerosis and vascular calcification, suggesting that endogenous genetic/epigenetic programs in veins protect them from these vascular diseases. Our data indicate arterial flow promotes an athero-protective phenotype while the ECs cultured under venous flow conditions exhibit a more athero-prone phenotype in comparison to arterial flow. However, the transcriptional profiling analysis in actual arteries and veins reveals a profile that is completely the opposite: arterial ECs demonstrate a more athero-prone profile and osteogenic program than venous ECs, which is more consistent with the pattern of COUP-TFII regulation. The en face VCAM-1 staining in blood vessels also confirmed that arterial ECs have a much stronger response to inflammatory stimuli by turn on VCAM-1 expression whereas venous ECs do not express VCAM-1 at all. VCAM-1 was reported to express at the arteries and post-capillary venules after cytokine stimulation14, but its expression in the major adult veins has not been examined in the past. Our finding that VCAM-1 is absent in large veins is very exciting. Because VCAM-1 is an essential adhesion molecule involved in atherosclerosis initiation and progress, the drastic difference in its expression in adult arteries and veins suggest that it may correlate with the low susceptibility of ath-erosclerosis in veins. Our data again confirmed that arterial ECs have a more athero-prone profile than venous ECs in response to inflammatory stimuli, which is consistent with the COUP-TFII regulated gene patterns but is totally the opposite to those predicted by hemodynamic flows. Taken together, these data suggest that hemodynamic flow is not the primary factor that determines the molecular divergences in artery and vein. Instead, the endogenous genetic/epigenetic programs (such as COUP-TFII) are stronger regulators that control these molecular and functional differences.
Vascular calcification is a frequent complication of vascular disease, such as diabetes mellitus, renal disease, and atherosclerosis. It is a regulated process with strong similarities to bone formation driven by osteo-progenitor cells in the media layer of blood vessel. Vascular EC represents a distinct cell line-age from osteo-progenitor cells. In healthy blood vessels and also in normal EC culture, the osteogenic program is usually absent and thus ECs do not contribute to the calcium deposition in healthy blood vessels. However, recent studies have demonstrated a role for the endothelium in vascular calcification45 in disease condition with high BMP activity. Under the stimulation of BMP4 (or TGFβ 2), ECs can be converted into mesenchymal stem cell (MSC)-like cells through EndMT process and undergo osteogenic differentiation under osteogenic condition46. This leads to the up-regulation of series gene pathways sim-ilar to osteogenic differentiation of MSCs. EndMT has also been reported in ECs derived from the mitral valve leaflets47 and in HAECs in vitro48, which support an important role for the endothelium in the development of cardiovascular calcification. Interestingly, in animal models, the EndMT and multipotent cells were found to be restricted to the artery wall but not in the veins in these mice45. Our data reveal mechanistic insight into this difference that COUP-TFII negatively regulates the EndMT process. It also plays a strong role in suppressing the osteogenic potential and subsequent calcium deposition of adult

www.nature.com/scientificreports/
1 4SCIENTIfIC REPORTS | 5:16193 | DOI: 10.1038/srep16193
ECs, suggesting that COUP-TFII may contribute to the different susceptibility of arteries and veins to calcification-related vascular diseases. The higher expression of RUNX2, ALPL, SPP1 and DLX5 in arte-rial ECs in vivo is also striking, indicating a stronger osteogenic potential of arteries. EndMT has been described in different pathologies, and is implicated in many disease situations such as cancer progres-sion49, cardiac fibrosis50, vascular calcification45, and intimal hyperplasia during vein graft remodeling51. Our findings of COUP-TFII as the suppressor of EndMT process suggest that it can be used as a broad therapeutic target for a variety of EndMT-related disease conditions.
In summary, COUP-TFII, a transcription factor expressed higher in venous EC, plays multifaceted roles in determining the functional difference between arteries and veins. Multiple important gene path-ways involved in vascular diseases are regulated by COUP-TFII. ECs with higher COUP-TFII expres-sion confer a more anti-atherogenic phenotype and lower osteogenic potential, while lower COUP-TFII expression elicits a more pro-atherogenic phenotype and greater sensitivity to EndMT and osteogenic stimulation. Our findings provide new insights into the mechanism of arterial diseases and suggest potential roles of COUP-TFII in determining the distinctive susceptibilities of arteries and veins to vas-cular diseases.
References1. Hu, S. et al. MicroRNA-302 increases reprogramming efficiency via repression of NR2F2. Stem Cells 31, 259–268 (2013).2. Takamoto, N. et al. COUP-TFII is essential for radial and anteroposterior patterning of the stomach. Development 132, 2179–2189
(2005).3. Xu, Z., Yu, S., Hsu, C.-H., Eguchi, J. & Rosen, E. D. The orphan nuclear receptor chicken ovalbumin upstream promoter-
transcription factor II is a critical regulator of adipogenesis. Proc Natl Acad Sci USA 105, 2421–2426 (2008).4. Okamura, M. et al. COUP-TFII acts downstream of Wnt/beta-catenin signal to silence PPARgamma gene expression and repress
adipogenesis. Proc Natl Acad Sci USA 106, 5819–5824 (2009).5. Xie, X., Qin, J., Lin, S. H., Tsai, S. Y. & Tsai, M. J. Nuclear receptor chicken ovalbumin upstream promoter-transcription factor
II (COUP-TFII) modulates mesenchymal cell commitment and differentiation. Proc Natl Acad Sci USA 108, 14843–14848 (2011).6. Qin, J., Chen, X., Xie, X., Tsai, M.-J. & Tsai, S. Y. COUP-TFII regulates tumor growth and metastasis by modulating tumor
angiogenesis. Proc Natl Acad Sci USA 107, 3687–3692 (2010).7. Qin, J., Chen, X., Yu-Lee, L.-Y., Tsai, M.-J. & Tsai, S. Y. Nuclear receptor COUP-TFII controls pancreatic islet tumor angiogenesis
by regulating vascular endothelial growth factor/vascular endothelial growth factor receptor-2 signaling. Cancer Res 70, 8812–8821 (2010).
8. Qin, J. et al. COUP-TFII inhibits TGF-β -induced growth barrier to promote prostate tumorigenesis. Nature 493, 236–240 (2013).9. You, L.-R., Lin, F.-J., Lee, C. T., DeMayo, F. J., Tsai, M.-J. & Tsai, S. Y. Suppression of Notch signalling by the COUP-TFII
transcription factor regulates vein identity. Nature 435, 98–104 (2005).10. Swift, M. R., Pham, V. N., Castranova, D., Bell, K., Poole, R. J. & Weinstein, B. M. SoxF factors and Notch regulate nr2f2 gene
expression during venous differentiation in zebrafish. Dev Biol 390, 116–125 (2014).11. Lindskog, H. et al. Molecular identification of venous progenitors in the dorsal aorta reveals an aortic origin for the cardinal vein
in mammals. Development 141, 1120–1128 (2014).12. Nam, D. et al. Partial carotid ligation is a model of acutely induced disturbed flow, leading to rapid endothelial dysfunction and
atherosclerosis. Am J Physiol Heart Circ Physiol 297, H1535–1543 (2009).13. Iiyama, K. et al. Patterns of vascular cell adhesion molecule-1 and intercellular adhesion molecule-1 expression in rabbit and
mouse atherosclerotic lesions and at sites predisposed to lesion formation. Circ Res 85, 199–207 (1999).14. Wang, C., Qin, L., Manes, T. D., Kirkiles-Smith, N. C., Tellides, G. & Pober, J. S. Rapamycin antagonizes TNF induction of
VCAM-1 on endothelial cells by inhibiting mTORC2. J Exp Med 211, 395–404 (2014).15. Korten, S. et al. Impact of Hey2 and COUP-TFII on genes involved in arteriovenous differentiation in primary human arterial
and venous endothelial cells. Basic research in cardiology 108, 362 (2013).16. Cybulsky, M. I. & Gimbrone, M. A., Jr. Endothelial expression of a mononuclear leukocyte adhesion molecule during
atherogenesis. Science 251, 788–791 (1991).17. Larrivee, B., Freitas, C., Suchting, S., Brunet, I. & Eichmann, A. Guidance of vascular development: lessons from the nervous
system. Circ Res 104, 428–441 (2009).18. Carmeliet, P. & Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature 436, 193–200 (2005).19. Lee, S. et al. Prox1 physically and functionally interacts with COUP-TFII to specify lymphatic endothelial cell fate. Blood 113,
1856–1859 (2009).20. Srinivasan, R. S. et al. The nuclear hormone receptor Coup-TFII is required for the initiation and early maintenance of Prox1
expression in lymphatic endothelial cells. Genes & Development 24, 696–707 (2010).21. Aranguren, X. L. et al. Unraveling a novel transcription factor code determining the human arterial-specific endothelial cell
signature. Blood 122, 3982–3992 (2013).22. Maddaluno, L. et al. EndMT contributes to the onset and progression of cerebral cavernous malformations. Nature 498, 492–496
(2013).23. Dai, G. et al. Distinct endothelial phenotypes evoked by arterial waveforms derived from atherosclerosis-susceptible and
-resistant regions of human vasculature. Proc Natl Acad Sci USA 101, 14871–14876 (2004).24. Chang, K. et al. Bone morphogenic protein antagonists are coexpressed with bone morphogenic protein 4 in endothelial cells
exposed to unstable flow in vitro in mouse aortas and in human coronary arteries: role of bone morphogenic protein antagonists in inflammation and atherosclerosis. Circulation 116, 1258–1266 (2007).
25. Wang, H. U., Chen, Z. F. & Anderson, D. J. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell 93, 741–753 (1998).
26. Villa, N., Walker, L., Lindsell, C. E., Gasson, J., Iruela-Arispe, M. L. & Weinmaster, G. Vascular expression of Notch pathway receptors and ligands is restricted to arterial vessels. Mech Dev 108, 161–164 (2001).
27. Krebs, L. T. et al. Notch signaling is essential for vascular morphogenesis in mice. Genes Dev 14, 1343–1352 (2000).28. Gale, N. W. et al. Haploinsufficiency of delta-like 4 ligand results in embryonic lethality due to major defects in arterial and
vascular development. Proc Natl Acad Sci USA 101, 15949–15954 (2004).29. Zhong, T. P., Childs, S., Leu, J. P. & Fishman, M. C. Gridlock signalling pathway fashions the first embryonic artery. Nature 414,
216–220 (2001).30. Fischer, A., Schumacher, N., Maier, M., Sendtner, M. & Gessler, M. The Notch target genes Hey1 and Hey2 are required for
embryonic vascular development. Genes Dev 18, 901–911 (2004).

www.nature.com/scientificreports/
1 5SCIENTIfIC REPORTS | 5:16193 | DOI: 10.1038/srep16193
31. Duarte, A. et al. Dosage-sensitive requirement for mouse Dll4 in artery development. Genes Dev 18, 2474–2478 (2004).32. Mukouyama, Y. S., Shin, D., Britsch, S., Taniguchi, M. & Anderson, D. J. Sensory nerves determine the pattern of arterial
differentiation and blood vessel branching in the skin. Cell 109, 693–705 (2002).33. Lawson, N. D. et al. Notch signaling is required for arterial-venous differentiation during embryonic vascular development.
Development 128, 3675–3683 (2001).34. Lawson, N. D. & Weinstein, B. M. Arteries and veins: making a difference with zebrafish. Nat Rev Genet 3, 674–682 (2002).35. le Noble, F. et al. Flow regulates arterial-venous differentiation in the chick embryo yolk sac. Development 131, 361–375 (2004).36. Henderson, V. J., Cohen, R. G., Mitchell, R. S., Kosek, J. C. & Miller, D. C. Biochemical (functional) adaptation of “arterialized”
vein grafts. Ann Surg 203, 339–345 (1986).37. Bush, H. L., Jr., Jakubowski, J. A., Curl, G. R., Deykin, D. & Nabseth, D. C. The natural history of endothelial structure and
function in arterialized vein grafts. J Vasc Surg 3, 204–215 (1986).38. Kudo, F. A. et al. Venous identity is lost but arterial identity is not gained during vein graft adaptation. Arteriosclerosis, thrombosis,
and vascular biology 27, 1562–1571 (2007).39. Muto, A. et al. Eph-B4 prevents venous adaptive remodeling in the adult arterial environment. J Exp Med 208, 561–575 (2011).40. Hsiai, T. K. et al. Monocyte recruitment to endothelial cells in response to oscillatory shear stress. FASEB J 17, 1648–1657 (2003).41. Chiu, J. J. & Chien, S. Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives.
Physiol Rev 91, 327–387 (2011).42. Davies, P. F. Hemodynamic shear stress and the endothelium in cardiovascular pathophysiology. Nat Clin Pract Cardiovasc Med
6, 16–26 (2009).43. Chien, S. Mechanotransduction and endothelial cell homeostasis: the wisdom of the cell. Am J Physiol Heart Circ Physiol 292,
H1209–1224 (2007).44. Gimbrone, M. A. & García-Cardeña, G. Vascular endothelium, hemodynamics, and the pathobiology of atherosclerosis.
Cardiovascular pathology: the official journal of the Society for Cardiovascular Pathology 22, 9–15 (2013).45. Yao, Y., Jumabay, M., Ly, A., Radparvar, M., Cubberly, M. R. & Bostrom, K. I. A role for the endothelium in vascular calcification.
Circ Res 113, 495–504 (2013).46. Medici, D., Shore, E. M., Lounev, V. Y., Kaplan, F. S., Kalluri, R. & Olsen, B. R. Conversion of vascular endothelial cells into
multipotent stem-like cells. Nature Medicine 16, 1400–1406 (2010).47. Wylie-Sears, J., Aikawa, E., Levine, R. A., Yang, J.-H. & Bischoff, J. Mitral valve endothelial cells with osteogenic differentiation
potential. Arteriosclerosis, thrombosis, and vascular biology 31, 598–607 (2011).48. Tang, R., Gao, M., Wu, M., Liu, H., Zhang, X. & Liu, B. High glucose mediates endothelial-to-chondrocyte transition in human
aortic endothelial cells. Cardiovascular diabetology 11, 113 (2012).49. Potenta, S., Zeisberg, E. & Kalluri, R. The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer 99,
1375–1379 (2008).50. Zeisberg, E. M. et al. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat Med 13, 952–961 (2007).51. Cooley, B. C. et al. TGF-beta signaling mediates endothelial-to-mesenchymal transition (EndMT) during vein graft remodeling.
Sci Transl Med 6, 227ra234 (2014).
AcknowledgementsWe like to thank Dr. Michael Gimbrone and Dr. Sally Temple for helpful comments on this manuscript, Leilani Cruz and Lonnie Seo for cell culture assistance. This study was supported mainly by grants from American Heart Association Scientist Development Grant (12SDG12050083 to G.D.), National Institute of Health (R21HL102773, R01HL118245 to G.D.) and National Science Foundation (CBET-1263455, CBET-1350240 to G.D.).
Author ContributionsX.C., Y.L., V.L., D.K., T.D. and Y.L. designed and performed the research. Q.W. made the lentivirus overexpressing COUP-TFII. Y.L., P.V. and J.S. performed the experiments on mouse blood vessels and contributed to scientific discussion. G.D. designed the research and wrote the paper.
Additional InformationSupplementary information accompanies this paper at http://www.nature.com/srepCompeting financial interests: The authors declare no competing financial interests.How to cite this article: Cui, X. et al. Venous Endothelial Marker COUP-TFII Regulates the Distinct Pathologic Potentials of Adult Arteries and Veins. Sci. Rep. 5, 16193; doi: 10.1038/srep16193 (2015).
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Com-
mons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/

Page 2
Supporting Information
SI Materials and Methods
Apply Arterial And Venous Flow To Cultured ECs
To apply arterial and venous flow to cultured ECs, we used the dynamic flow system previously
developed in the lab 1 to re-create the pulsatile shear stress waveform derived from typical human
abdominal aorta and saphenous vein blood flow patterns. The dynamic flow system is under the computer
control to enable accurate program of any shear stress waveform and applied it to cultured ECs. HSVECs
were plated at an initial density of 50,000 cells/cm2 on 0.1% gelatin (Sigma) coated tissue culture plate
and the cell culture plate was assembled in the dynamic flow system. Fully confluent EC monolayer was
then exposed to the arterial or venous shear stress waveforms for 72 hours. Culture medium in the shear
apparatus was replenished during the experiment at an exchange rate of 0.1 ml/min, and the enclosed
environment was maintained at 37°C in a humidified 5% CO2/95% air atmosphere. For comparison, EC
from the same passage were plated on an identical plastic plate assembly and incubated at 37°C in a 5%
CO2/95% air atmosphere under static (no flow) conditions.
RNA Isolation and Real-Time PCR
RNA was isolated using RNeasy Plus Mini Kit (Qiagen) according to the manufacturer’s
protocol. The concentration and quality of RNA was determined by NanoDrop (Thermo Scientific). Gene
expression was measured by Quantitative Real-time PCR reactions using Taqman RT-PCR assays (Life
Technologies) on StepOne Plus RT-PCR system (Life Technologies). The relative gene expression was
normalized to 18S ribosomal RNA.
Microarray Hybridization and Statistical Methods for Microarray Data Analysis

Page 3
For microarray analysis, three independent sets of experiments were performed. Microarray
analysis was performed using the Human Whole Genome OneArray® v5 (Phalanx Biotech), which
contains 30,275 oligonucleotides representing 29,187 human genome probes and 1,088 experimental
control probes. RNA quality and integrity were determined utilizing an Agilent 2100 Bioanalyzer
(Agilent Technologies) and a NanoDrop spectrophotometer (Thermo Scientific). Only high quality RNA,
having a RIN of >6.0, and absorbance ratios A260/A280 >1.8 and A260/A230 >1.5, was utilized for
further experimentation. RNA was converted to double-stranded cDNA and amplified using in vitro
transcription that included amino-allyl UTP, and the cDNA product was subsequently conjugated with
Cy5™ NHS ester (GEH Lifesciences). Fragmented cDNA was hybridized at 42 ⁰C overnight using the
HybBag mixing system with 1X OneArray Hybridization Buffer (Phalanx Biotech), 0.01 mg/ml sheared
salmon sperm DNA (Promega), at a concentration of 0.025 mg/ml labeled target. After hybridization, the
arrays were washed according to the OneArray protocol.
Raw intensity signals for each microarray were captured using a Molecular Dynamics™ Axon
4100A scanner, measured using GenePixPro™ Software, and stored in GPR format. The data from all
microarrays in each experimental set was then passed to Rosetta Resolver (Rosetta Biosoftware) for
analysis. Testing was performed by combining replicates and performing statistical analyses using
Rosetta Resolver. The Rosetta error model captures the variance-intensity relationship for various types of
microarray technologies. This error model conservatively estimates intensity error and uses this value to
stabilize the variance estimation. These technology-specific error models are designed and optimized for
different microarray technologies, such as Affymetrix® and Agilent Technologies. Data from three
independent experiments were analyzed, and the averaged ratio and p-value were obtained for each paired
comparison. To decrease the false-positive rate associated with the multiple comparisons, the Bonferroni
correction was applied to the analysis, gene regulations were considered statistically significant when p <
0.05/(total number of genes tested)=0.05/29,187.

Page 4
Chromatin Immunoprecipitation (ChIP)-PCR
Confluent ECs under various experimental conditions were cross-linked by addition of 1%
formaldehyde and subsequently harvested. Chromatin immunoprecipitation on the samples was
performed as previously described 2, using mouse monoclonal anti-human COUP-TFII antibody (R&D
Systems). Mouse IgG (Life Technologies) was used as negative control. DNA released from the
precipitation was subjected to qPCR analysis for quantification of the presence of specific loci. The
promoter region of human BMP-4 was analyzed by qPCR with the primers 5’-
ACAGCTCCATCAGAGGCAGT -3’ and 5’- GAAACAGGCTGTGTGCAGAA -3’, which spans the
1.5kb upstream of the transcription start site of BMP4 gene. The relative expression was compared with
the amplification of input chromatin not subjected to immunoprecipitation.
Transfection of siRNA Duplexes into HSVEC
siRNA to knockdown COUP-TFII and control siRNA were synthesized by Thermo Scientitic
Bio. siRNA was transfected into HSVEC with Lipofectamine RNAiMAX (Life Technologies) and Opti-
MEM (Life Technologies) at a final concentration of 10nM according to manufacturer’s instruction. RNA
was isolated 72 hours after the siRNA transfection. The efficiency of knockdown was greater than 80% as
assessed by both western blot and RT-PCR. To confirm the specificity of the siRNA knockdown, selected
experiments were also performed using a different siRNA construct (Life Technologies) and an shRNA
construct (Sigma Mission Lentivirus shRNA, Sigma).
Immunofluorescence Microscopy of Cellular Proteins
EC were rinsed in cold PBS and fixed in 4% paraformaldehyde for 10 minutes and then
permeabilized in 0.1% Triton X-100 for 1 minutes at room temperature. To reduce non-specific
immunoglobulin binding, cells were pre-incubated in 1% BSA for 1 hour. For Cx40 staining, cells were
incubated with anti-Cx40 antibody (1:200, rabbit polyclonal IgG, Alpha Diagnostic International) for 1
hour at room temperature, rinsed with 0.1% BSA in PBS, and incubated for 30 minutes with 2% goat

Page 5
serum in PBS. Cells were incubated with Alexa Fluor 488 goat anti-rabbit (1:500, Life Technologies) for
1 hour at room temperature, rinsed in PBS, and mounted using Gel-Mount (Biomeda). Immunostaining of
Slug and α-SMA was done using the same method with anti-Slug antibody (1:200, rabbit polyclonal IgG,
Cell Signaling) and anti-α-SMA antibody (1:500, mouse monoclonal IgG, Sigma).
Western Blot Analysis of Protein Expression
Cells growing on 6-well plates were rinsed with ice-cold PBS and scraped after lysis with 300ul
Laemmli buffer containing complete protease inhibitor mixture (Roche Applied Science), PhosSTOP
phosphatase inhibitor mixture (Roche Applied Science), and subsequently boil for 5 minutes. 30ul of the
cell lysate from each condition were then loaded on standard SDS-PAGE gels and transferred to
nitrocellulose membranes. Immunoblots were performed by blocking the membranes with 3% non-fat
milk in TBS and incubate at 4°C overnight with any of the following antibodies: anti-BMP4 (1:500,
mouse monoclonal IgG, Santa Cruz Biotechnology), anti-COUP-TFII (1:250, mouse monoclonal IgG,
R&D Systems), anti-VCAM-1 (1:250, mouse monoclonal IgG, BD Biosciences) and anti-β-actin
antibody (1:5000, mouse monoclonal IgG, Sigma-Aldrich). Secondary anti-mouse antibody conjugated
with HRP (Jackson ImmunoResearch Laboratories) were incubated for 2 hour at room temperature.
Membrane were developed using SuperSignal West Pico or Femto chemiluminescent substrate (Pierce)
and Fujifilm LAS-3000 imaging system.
Leukocyte Adhesion Assays
THP-1 cells (ATCC) were cultured in suspension in tissue culture flask with RPMI-1640 Medium
(ATCC) supplemented with 50uM β-mercaptoethanol and 10% fetal bovine serum, and passaged
according to manufacturer’s protocol. One million THP-1 cells were labeled with Cell Tracker Green
(Life Technologies) as instructed by the manufacturer. To evaluate the leukocyte-endothelial adhesion,
confluent HSVEC monolayer was treated with TNF-α (0.01ng/ml) for 24 hours. HSVECs were washed
twice with PBS to remove cell debris, and the 5 ml of THP-1 suspension (1 million cells total) was added

Page 6
to the cells. The dish was place on a horizontal rotator at approximately 60 rpm for 10 minutes at room
temperature to allow binding. Monolayers were then washed gently with PBS 3 times, and then visualized
under a phase-contrast microscope with a 488 filter (to visualize labeled THP-1 cells). For quantification
of bound THP-1 cells, the number of cells in each field is counted and averaged from 10 randomly chosen
field.

Page 7
Supporting Tables
* Data from three independent experiments were analyzed by statistical methods (see Supporting
Materials and Methods). To decrease the false-positive rate associated with the multiple
comparisons, the Bonferroni correction of p-value was applied. The listed genes have at least two
folds changes, and have a p< 0.05/(total number of genes tested)=0.05/29,187.
Table S1. Selected Genes That Are Up-Regulated After COUP-TFII Knockdown
Category Gene Name Gene Symbol Fold Change Accession #
Vascular Development / Arterial-Venous Differentiation
hairy/enhancer-of-split related with YRPW motif 2 HEY2 6.7 NM_012259.2
semaphorin 7A, GPI membrane anchor SEMA7a 6.0 NM_003612.3
delta-like 4 DLL4 4.1 NM_019074.3
hairy and enhancer of split 4 HES4 3.4 NM_021170.3
hairy/enhancer-of-split related with YRPW motif 1 HEY1 3.1 NM_012258.3
fms-related tyrosine kinase 1 FLT1/VEGFR-1 2.9 NM_002019.4
endothelial PAS domain protein 1 EPAS1/HIF-2a 2.8 NM_001430.4
notch 4 Notch4 2.8 NM_004557.3
forkhead box C2 FOXC2 2.3 NM_005251.2
delta-like 1 DLL1 2.2 NM_005618.3
vasohibin 1 VASH1 2.2 NM_014909.4
ephrin-A1 Ephrin-A1 2.1 NM_004428.2
TEK tyrosine kinase, endothelial TIE-2 2.1 NM_000459.3
EPH receptor A4 EphA4 2.0 NM_004438.3
ephrin-B2 Ephrin-B2 2.0 NM_004093.3
jagged 1 JAG1 2.0 NM_000214.2
Inflammation
chemokine (C-X-C motif) ligand 10 CXCL10 36 NM_001565.2
chemokine (C-X-C motif) ligand 11 CXCL11 10 NM_005409.4
tumor necrosis factor (ligand) superfamily, member 10 TNFSF10 3.9 NR_033994.1
chemokine (C-C motif) ligand 5 CCL5 3.8 NM_002985.2
interleukin 15 receptor, alpha IL15RA 3.4 NM_002189.3
chemokine (C-C motif) ligand 5 CCL5 3.3 NM_002985.2
chemokine (C-X3-C motif) ligand 1 CX3CL1 3.2 NM_002996.3

Page 8
interleukin 1, alpha IL1A 2.3 NM_000575.3
chemokine (C-X-C motif) ligand 12 CXCL12 2.2 NM_000609.5
interleukin 3 receptor, alpha (low affinity) IL3RA 2.0 NM_002183.2
tumor necrosis factor receptor superfamily, member 4 TNFRSF4 2.0 NM_003327.3
chemokine (C-X-C motif) ligand 2 CXCL2 2.0 NM_002089.3
Extracellular Matrix Biosynthesis
collagen, type IV, alpha 2 COL4A2 2.3 NM_001846.2
integrin, alpha 3 ITGA3 2.3 NM_005501.2
laminin, alpha 5 LAMA5 2.2 NM_005560.3
Thrombosis / Vasomotor Function
serpin peptidase inhibitor, clade B, member 1 SERPINB1 3.7 NM_030666.2
serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 2 SERPINE2/PAI-1 2.4 NM_006216.3
endothelin converting enzyme 1 ECE-1 2.0 NM_001397.2
Lipid / Metabolic Pathways
apolipoprotein L, 3 ApoL3 2.7 NR_027835.1
ATP-binding cassette, sub-family G, member 1 ABCG1 2.1 NM_207629.1
Junction Proteins
gap junction protein, alpha 5, 40kDa Cx40 6.2 NM_181703.2
gap junction protein, alpha 4, 37kDa Cx37 3.4 NM_002060.2

Page 9
Table S2. Selected Genes That Are Down-Regulated After COUP-TFII Knockdown
Category Gene Name Gene Symbol Fold Change Accession #
Vascular Development / Arterial-Venous Differentiation
nuclear receptor subfamily 2, group F, member 2 COUP-TFII -5.3 NM_021005.3
neuropilin 2 Nrp2 -2.0 NM_201266.1
Thrombosis / Vasomotor Function
tissue factor pathway inhibitor 2 TFPI2 -2.3 NM_006528.2
prostaglandin reductase 1 PTGR1 -2.0 NM_012212.3
Extracellular Matrix
syndecan 1 SDC1 -2.2 NM_002997.4
Lipid / Metabolic Pathways
low density lipoprotein receptor LDLR -4.3 NM_000527.4
Page 10
Table S3. Selected Genes That Are Down-Regulated After Over Expressing COUP-TFII
Category Gene Name Gene Symbol Fold Change Accession #
Vascular Development / Arterial-Venous Differentiation
roundabout, axon guidance receptor, homolog 1 ROBO1 -102 NM_133631.3
semaphorin 3G SEMA3G -10.8 NM_020163.1
netrin 4 NTN4 -6.3 NM_021229.3
ephrin-A1 ephrin-A1 -5.1 NM_004428.2
ephrin-B2 ephrin-B2 -3.9 NM_004093.3
hairy and enhancer of split 4 HES4 -3.6 NM_021170.3
jagged 2 JAG2 -3.3 NM_145159.1
ephrin-B3 ephrin-B3 -3.2 NM_001406.3
semaphorin 6C SEMA6C -3.0 NM_030913
hairy and enhancer of split 1 HES1 -2.6 NM_005524.3
endothelial PAS domain protein 1 EPAS1/HIF-2a -2.4 NM_001430.4
SRY (sex determining region Y)-box 4 SOX4 -2.3 NM_003107.2
plexin D1 PLXND1 -2.2 NM_015103.2
SRY (sex determining region Y)-box 18 SOX18 -2.2 NM_018419.2
T-box 1 TBX1 -2.2 NM_080647.1
vasohibin 1 VASH1 -2.2 NM_014909.4
SRY (sex determining region Y)-box 17 SOX17 -2.1 NM_022454.3
Inflammation
vascular cell adhesion molecule 1 VCAM-1 -9.2 NM_080682.2
chemokine (C-C motif) ligand 2 CCL2 -5.9 NM_002982.3
chemokine (C-C motif) ligand 23 CCL23 -4.4 NM_145898.1
chemokine (C-C motif) ligand 14 CCL14 -3.6 NM_032962.4
tumor necrosis factor (ligand) superfamily, member 10 TNFSF10 -3.6 NR_033994.1
chemokine (C-C motif) ligand 11 CCL11 -3.1 NM_002986.2
chemokine (C-C motif) ligand 20 CCL20 -2.6 NM_004591.2
Extracellular Matrix
sulfatase 1 SULF1 -21.1 NM_015170.2
matrix Gla protein MGP -13.1 NM_000900.3
collagen, type I, alpha 2 COL1A2 -12.0 NM_000089.3
biglycan BGN -6.8 NM_001711.4
collagen, type V, alpha 1 COL5A1 -5.4 NM_000093.3
collagen, type IV, alpha 1 COL4A1 -4.1 NM_001845.4
matrix metallopeptidase 10 MMP10 -4.0 NM_002425.2
integrin, beta 1 ITGB1 -3.9 NM_002211.3
collagen, type VIII, alpha 1 COL8A1 -3.6 NM_020351.3
collagen, type IV, alpha 2 COL4A2 -3.4 NM_001846.2

Page 11
collagen, type V, alpha 2 COL5A2 -3.1 NM_000393.3
fibronectin 1 FN1 -2.6 NM_054034.2
laminin, alpha 4 LAMA4 -2.4 NM_002290.3
integrin, alpha V ITGAV -2.3 NM_002210.3
Thrombosis / Vasomotor Function
von Willebrand factor vWF -5.5 NM_000552.3
angiotensin I converting enzyme 1 ACE -5.0 NM_001178057.1
multimerin 1 MMRN1 -3.6 NM_007351.2
endothelin 1 ET-1 -3.5 NM_001955.4
thrombospondin 1 THBS1 -2.6 NM_003246.2
endothelin converting enzyme 1 ECE-1 -2.1 ,NM_001397.2
Growth Factor Signaling
insulin-like growth factor 2 IGF2 -10.3 NM_000612.4
bone morphogenetic protein 4 BMP4 -6.1 NM_130851.2
latent transforming growth factor beta binding protein 1 LTBP1 -5.2 NM_206943.2
platelet-derived growth factor beta polypeptide PDGF-β -4.7 NM_002608.2
placental growth factor PGF -3.3 NM_001207012.1
cysteine rich transmembrane BMP regulator 1 CRIM1 -2.6 NM_016441.2
Lipid / Metabolic Pathways
peroxisome proliferator-activated receptor alpha PPARα -5.0 NM_005036.4
leptin receptor LEPR -3.1 NM_001003679.3
retinol binding protein 1 RBP1 -2.8 NM_002899.3
Junction Proteins
gap junction protein, alpha 4, 37kDa Cx37 -5.8 NM_002060.2
gap junction protein, alpha 1, 40kDa Cx40 -3.3 NM_005266.5

Page 12
Table S4. Selected Genes That Are Up-Regulated After Over Expressing COUP-TFII
Category Gene Name Gene Symbol Fold Change Accession #
Vascular Development / Arterial-Venous Differentiation
lymphatic vessel endothelial hyaluronan receptor 1 LYVE1 6.8 NM_006691.3
nuclear receptor subfamily 2, group F, member 2 COUP-TFII 4.6 NM_021005.3
Extracellular Matrix
versican VCAN 9.5 NM_004385.4
syndecan 1 SDC1 8.7 NM_002997.4
laminin, gamma 2 LAMC2 5.9 NM_018891.2
collagen, type IV, alpha 6 COL4A6 5.5 NM_033641.2
ADAM metallopeptidase with thrombospondin type 1 motif, 18
ADAMTS18 4.3 NM_199355.2
matrix metallopeptidase 1 MMP1 3.2 NM_002421.3
collagen, type XVII, alpha 1 COL17A1 2.6 NM_000494.3
ADAM metallopeptidase with thrombospondin type 1 motif, 9
ADAMTS9 2.6 NM_182920.1
Lipid / Metabolic Pathways
lipase, endothelial LIPG 3.9 NM_006033.2
Thrombosis / Vasomotor Function
serpin peptidase inhibitor, clade D (heparin cofactor), member 1
SERPIND1 Heparin cofactor II
3.7 NM_000185.3
serpin peptidase inhibitor, clade B (ovalbumin), member 8 SERPINB8 2.4 NM_198833.1
plasminogen activator, tissue tPA 2.2 NM_033011.2
tissue factor pathway inhibitor 2 TFPI2 2.1 NM_006528.2
Page 13
Table S5. Selected Genes With Higher Expression In Aortic ECs Than Vena Cava ECs
Category Gene Name Gene Symbol Fold Change
(Aorta vs. Vena Cava)
Accession #
Vascular Development / Arterial-Venous Differentiation
slit homolog 1 Slit1 67.6 NM_015748
Eph receptor B6 (Ephb6), transcript variant 1 EphB6 65.5 NM_001146351
SRY-box containing gene 13 Sox13 28.1 NM_011439
SRY-box containing gene 17 Sox17 13.3 NM_011441
Ephrin A5 (Efna5), transcript variant 1 Ephrin-A5 10.6 NM_207654
Semaphoring 3G Sema3g 9.4 ENSMUST00000090180
kinase insert domain protein receptor Kdr 8.0 NM_010612
Semaphoring 6C Sema6c 7.8 NM_011351
T-box 1 Tbx1 5.3 NM_011532
smoothened homolog Smo 4.7 NM_176996
plexin D1 Plxnd1 4.6 NM_026376
Cardiovascular Hormone
adrenergic receptor, alpha 2a Adra2a 205 NM_007417
androgen binding protein delta Abpd 148.3 NM_001009952
adrenergic receptor, alpha 1b Adra1b 6.6 NM_007416
Growth Factors Signaling
platelet derived growth factor, beta Pdgfb 4.6 NM_011057
bone morphogenetic protein 4 Bmp4 4.0 NM_007554
platelet derived growth factor, alpha Pdgfa 3.5 NM_008808
Extracellular Matrix Biosynthesis
fibulin 2 Fbln2 13.1 NM_007992
distal-less homeobox 5 Dlx5 10.1 NM_010056
fibulin 5 Fbln5 5.7 NM_011812
integrin alpha M, transcript variant 1 Itgam 2.6 NM_001082960
multimerin 2 Mmrn2 2.5 NM_153127
Thrombosis / Vasomotor Function
thrombospondin Thbs1 8.0 NM_011580
angiotensin I converting enzyme Ace 6.4 NM_009598
coagulation factor X F10 2.5 NM_007972
Lipid / Metabolic Pathways
leptin receptor (Lepr) Lepr 8.4 NM_010704
peroxisome proliferator activated receptor alpha PPARa 8.0 NM_011144
apolipoprotein C-I Apoc1 3.4 NM_007469
Junction Proteins
gap junction protein, beta 6 (Gjb6) Cx30 521.4 NM_001010937

Page 14
gap junction protein, alpha 5 (Gja5) Cx40 189.6 NM_008121
claudin 5 Cldn5 4.7 NM_013805
gap junction protein, alpha 4 (Gja4) Cx37 2.3 NM_008120

Page 15
Table S6. Selected Genes With Higher Expression In Vena Cava ECs Than Aortic ECs
Category Gene Name Gene Symbol Fold Change
(Vena Cava vs. Aorta)
Accession #
Vascular Development / Arterial-Venous Differentiation
wingless-type MMTV integration site 9B Wnt9b 91.2 NM_011719
T-box 5 Tbx5 48.1 NM_011537
apelin Apelin 27.7 NM_013912
nuclear receptor subfamily 2, group F, member 2 COUP-TFII 10.2 NM_009697
endoglin Eng 9.4 NM_001146350
activin A receptor, type 1 Acvr1 9.4 NM_001110204
semaphorin 5A Sema5a 6.8 NM_009154
lymphatic vessel endothelial hyaluronan receptor 1 Lyve1 6.2 NM_053247
neuropilin 2 Nrp2 5.6 NM_001077403
Eph receptor B4 Ephb4 3.2 NM_001159571
Extracellular Matrix Biosynthesis
matrix metallopeptidase 23 Mmp23 25.2 NM_011985
laminin, alpha 4 Lama4 15.1 NM_010681
matrix metallopeptidase 2 Mmp2 6.4 NM_008610
laminin, beta 2 Lamb2 6.2 NM_008483
collagen, type IV, alpha 2 Col4a2 5.9 NM_009932]
collagen, type XVI, alpha 1 Col16a1 5.8 NM_028266
tissue inhibitor of metalloproteinase 4 Timp4 5.8 NM_080639
collagen, type V, alpha 3 Col5a3 5.3 NM_016919
tissue inhibitor of metalloproteinase 2 Timp2 4.8 NM_011594
collagen, type III, alpha 1 Col3a1 3.7 NM_009930
a disintegrin-like and metallopeptidase (reprolysin type) with thrombospondin type 1 motif, 2
Adamts2 3.7 NM_175643
Thrombosis / Vasomotor Function
serine (or cysteine) peptidase inhibitor, clade A, member 3N
Serpina3n 68.2 NM_009252
serine (or cysteine) peptidase inhibitor, clade G, member 1 Serping1 18.2 NM_009776
plasminogen activator, tissue tPA 6.3 NM_008872
tissue factor pathway inhibitor, transcript variant 2 TFPI2 5.3 NM_001177319
serine (or cysteine) peptidase inhibitor, clade B, member 6d Serpinb6d 3.8 NM_001076790
Cardiovascular Hormone
natriuretic peptide type A Nppa 221.9 NM_008725
natriuretic peptide type C Nppc 38.0 NM_010933
Junction Proteins
claudin 11 Cldn11 57.3 NM_008770

Page 16
Table S7. Selected Differentially Regulated Genes By COUP-TFII Under TGF-β2 Treatment
Category Gene Name Gene Symbol Fold Change
(COUP-TFII vs. Control RNAi)
Accession #
Endothelial-to-Mesenchymal Transition
calponin 1, basic, smooth muscle CNN1 8.6 NM_001299.4
collagen, type I, alpha 2 COL1A2 5.9 NM_000089.3
CD44 molecule CD44 5.4 NM_000610.3
serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 2 SERPINE2/PAI-1 4.1 NM_006216.3
transgelin TAGLN/SM22a 3.9 NM_003186.3
snail homolog 2 SNAI2 2.4 NM_003068.4
TGF-β Signaling Pathway
transforming growth factor, beta-induced, 68kDa TGFBI 23.1 NM_000358.2
follistatin FST 10.6 NM_006350.3
inhibin, beta A INHBA 7.5 NM_002192.2
transforming growth factor, beta 2 TGFB2 5.5 NM_003238.3
SMAD family member 6 SMAD6 2.9 NM_005585.4

Page 17
SI References: 1. Dai G, et al. Distinct endothelial phenotypes evoked by arterial waveforms derived from
atherosclerosis-susceptible and -resistant regions of human vasculature. Proc Natl Acad Sci U S A 101, 14871-14876 (2004).
2. Lee TI, Johnstone SE, Young RA. Chromatin immunoprecipitation and microarray-based
analysis of protein location. Nat Protoc 1, 729-748 (2006). 3. Iiyama K, et al. Patterns of vascular cell adhesion molecule-1 and intercellular adhesion
molecule-1 expression in rabbit and mouse atherosclerotic lesions and at sites predisposed to lesion formation. Circ Res 85, 199-207 (1999).
4. Wang C, Qin L, Manes TD, Kirkiles-Smith NC, Tellides G, Pober JS. Rapamycin antagonizes
TNF induction of VCAM-1 on endothelial cells by inhibiting mTORC2. J Exp Med 211, 395-404 (2014).

Page 18
Supporting Figures
Figure S1. Arterial and venous ECs demonstrate distinct gene expression pattern. Some highly
differentially expressed genes in aorta or vena cava ECs are listed. All genes listed here are statistically
different in aorta vs. vena cava.

Page 19
Figure S2. BMP4 protein expresses only in the endothelium layer of mouse aorta. Side view (x-z
plane) of the en face confocal immunofluorescent images of BMP4 expression in mouse aorta and vena
cava.

Page 20
Figure S3. Arterial flow promotes athero-protective phenotype but has very small influence on
arterial venous markers. HSVECs were cultured under static (no flow), venous or arterial flow
condition for 72 hours. Gene expression was analyzed by Taqman RT-PCR, n=3, *p<0.05.

Page 21
Figure S4. (A) HAECs were transfected with COUP-TFII or Control RNAi (10nM). RNA was collected
72 hours after transfection. Gene expressions were measured by Taqman-RT PCR and normalized to
RNA 18S. Data are represented as relative expression level, n=3. (B) HAECs were infected with
lentivirus-COUP-TFII or control lentivirus (10 MOI), RNAs were isolated after confluent monolayer is
reached and all cells were permanently infected with lentivirus. Gene expressions were measured by
Taqman-RT PCR and normalized to RNA 18S, n=3.

Page 22
Figure S5. RNA was isolated from freshly harvested mouse aorta, and the purity of RNA was checked
for smooth muscle contamination by quantifying the expression of smooth muscle alpha actin (SM a-
actin) in comparison to RNA samples isolated from aorta media layer. The smooth muscle contamination
was determined to be <1%. Hypoxanthine-guanine phosphoribosyltransferase (HPRT) was used as
control to verify the equal amount of RNA loading across all samples (n=6).





























