Embed Size (px)
Citation preview

Folia Microbiol. 47 (1), 61~7 (2002) h t tp : / /www, biomod, eas . e z / r a b u / f o l i a /
Viability of Dried Filaments, Survivability and Reproduction under Water Stress, and Survivability Following Heat and UV Exposure in Lyngbya martensiana, Oscillatoria agardhii, Nostoc calcicola, Hormidium
fluitans, Spirogyra sp. and Vaucheria geminata S.C. AGRAWAL, V. SINGH
Department of Botany, University of Allahabad, Allahabad, lndia Received 4 June 2001
ABSTRACT. Dried vegetative filaments of Spirogyra sp., Vaucheria geminata and Nostoc calcicola died within �89 1 and 4 h, respectively; those ofHormidiumfluitans, Oscillatoria agardhii and Lyngbya martensiana retained under similar storage conditions viability for 3, 5 and 10 d, respectively. The viability of dried vegetative filaments ofL. martensiana, O. agardhii and H. fluitans decreased on storage at 20 ~ in the dark. L. martensiana and O. agardhii tolerated 0.8 mol/L NaCI. The resistance to desiccation in L. martensiana and O. agardhii exhibited similar dependence as that to frost, to heat and UV light. O. agardhii filaments became slightly broader and their cells developed large number of gas vacuoles when grown in 0.8 mol/L NaCl-containing medium. The water stress imposed on growing algae either on high-agar solid media or in NaCl-containing liquid media reduced hormogonium formation in L. martensiana and O. agardhii, hetero- cyst and akinete formation in N. calcicola and fragmentation in H. fluitans; it did not induce conjugation in Spirogyra sp. and formation of reproductive organs in V. geminata. In all studied algae the stress reduced at various levels the survival of vegetative parts. Generally, algal body form and composition rather than habitats seem to decide primarily the level of resistance against various stress conditions.
The impact of water stress (Cameron and Blank 1966; Whitton 1987; Tomaselli and Giovannetti 1993; Caiola et al. 1996), heat (Buzer et al. 1985; Hawes et al. 1992) and UV light (Garcia-Pichel et al. 1992; Garcia-Pichel and Castenholz 1993; Danilov and Ekelund 2000) has been studied with respect to survival and growth of different algae. Some earlier work showed that dessication can induce formation of cysts in Vau- cheria spp. (Stahl 1879) and Protosiphon spp. (Moewus 1935) and akinetes in Zygnema spp. (Fritsch 1916).
The aim of our study was to determine how long and to what extent Lyngbya martensiana MENE- GHINII, Oscillatoria agardhii GOMONT, Nostoc calcicola BREBISSON, Hormidium fluitans (GAY) HEERING, Spirogyra sp. and Vaucheria geminata (VAUCH.) DE CANDOLLE tolerate dry storage at different tempera- tures, and to what extent they survive and reproduce under physical and physiological water stress imposed by growing them on media with a high agar content and/or in NaCl-containing liquid media. The extent of subsequent exposure to heat and UV-light tolerating by them is also reported.
MATERIALS AND METHODS
Algal material was collected from Allahabad (India). L. martensiana grew firmly attached to rock surfaces exposed to running tap water; O. agardhii trichomes (vegetative filaments) formed mucilagenous sheet on moist soil surface; N. calcicola and H. fluitans were isolated from a fresh-water pond and tank, res- pectively; Spirogyra sp. and K geminata were isolated in a road side fresh-water ditch and on damp soil sur- face, respectively.
Growth conditions. Blue-green algae were grown in liquid BGII medium (Stanier et al. 1971); green algae and K geminata in liquid Bold's basal medium (Nichols and Bold 1965; pH adjusted prior to autoclaving to 7.5). Their cultures were maintained in controlled culture conditions at 22 ~ and light inten- sity of ca. 40 pmol m -2 s -1 from daylight fluorescent tubes for 16 h a day.
Viability of dried filaments. Seven-d-old, actively growing, vegetative filaments of all algae obtai- ned from liquid media were separately placed on filter paper and kept in desiccators over fused calcium chloride at 20 ~ in light (10 h at ca. 28 p.mol m -2 s -1 intensity), 20 ~ in dark, 12 ~ in dark and 0 ~ in dark for �89 h to 15 d. Filaments suspended in sterile distilled water and exposed to similar storage conditions served as controls. The viability of dried filaments was determined by counting the number of surviving filaments after 10 d of inoculation in liquid media placed in a culture chamber.

62 S.C. A G R A W A L and V. S I N G H Vol. 47
. ~ \ X .i, 2
X':i " ': / , ,"
�9 - .d /
T - _
i
J )
q.
~ ,
~ ~
i I
r
I ~ i �9 ~.;ii~ ~ \.i
o Iii --
~ ~i~/~i~;.i~.Si,/~" ~:~ ~ii~il
2002 SURVIVABILITY OF ALGAE AFTER WATER AND tlEAT STRESS, AND UV EXPOSURE 6 3
Table I. Survival (%) of dried vegetative filaments of different algae after storage at 20, 12 and 0 ~
Time d l'ightb
Dark
20 12 0
L. m a r t e n s i a n a
I 100 100 100 50 100 100 88 10
2 100 100 100 50 89 85 70 I
3 100 100 68 10 80 78 30 0
5 100 100 30 0 50 55 12 0
10 100 100 0 - 11 13 0 -
15 100 100 - - 0 1 - -
O. agardh i i
1 100 100 100 44 51 56 50 8
2 I00 100 92 20 50 51 43 0
4 100 100 55 0 39 36 13 0
5 88 80 36 - 14 15 7 --
8 79 76 20 - 0 0 0 -
Dark Time Light b
d 20 12 0
N. calc icola
I c 100 100 100 30 30 38 25 0
4 c 100 100 100 0 0 0 0 0
H. f l u i t a n s
1 100 100 100 50 69 68 31 8
2 100 100 100 0 38 36 12 0
3 100 100 85 - 14 8 0 -
5 100 100 72 - 0 0 0 -
Sp i rogyra sp.
�89 100 100 100 100 0 0 0 0
g e m i n a t a
1 c 100 I00 100 100 1 3 0 0
aln desiccators over fused calcium chloride for different time periods; measured 10 d after inoculation in liquid media; controls were maintained in sterile distilled water and kept under similar storage conditions; first lines - controls, second lines - dried stage; mean of three replicates.
bStorage at 20 ~ under 10-h illumination with fluorescent light of 28 gtmol m-2s -I. Cln h.
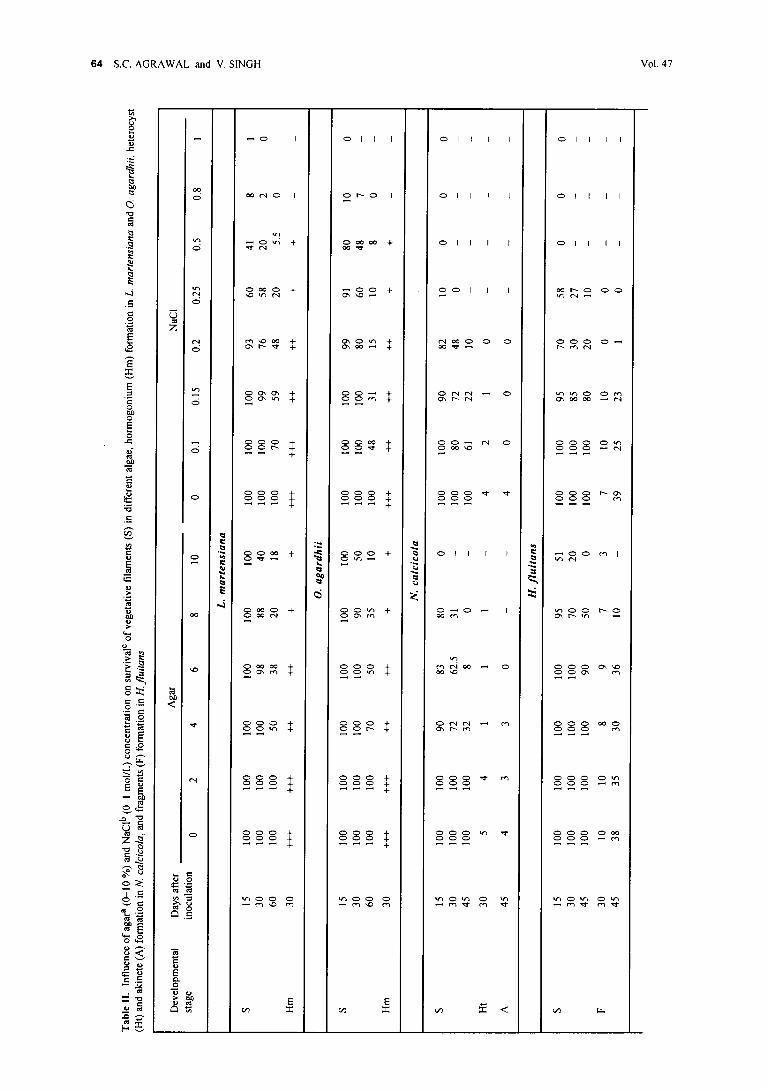
Survival and reproduction under physical and physiological water stress. Seven-d-old vegetative filaments of all algae were separately inoculated on solid media containing 2-10 % agar and in liquid media containing 0.1-0.8 mol/L NaCI and kept in culture chamber; the controls were maintained in liquid media. The percentage survival of vegetative filaments was determined by counting the number of surviving fila- ments from a total of 3000-4000 filaments.
Since L. martensiana and O. agardhii formed hormogonia profusely in mass, quantification was difficult and therefore approximate evaluation (signs + and -) had to be used to express their decreasing or increasing amounts. The formation of heterocysts and akinetes in N. calcicola was estimated by counting their percentages relative to the total number of 5000~5000 vegetative cells. The fragmentation in H. fluitans was estimated by counting the percentage of filaments showing bending and consequent fragment formation out of a total number of 2500-3000 filaments.
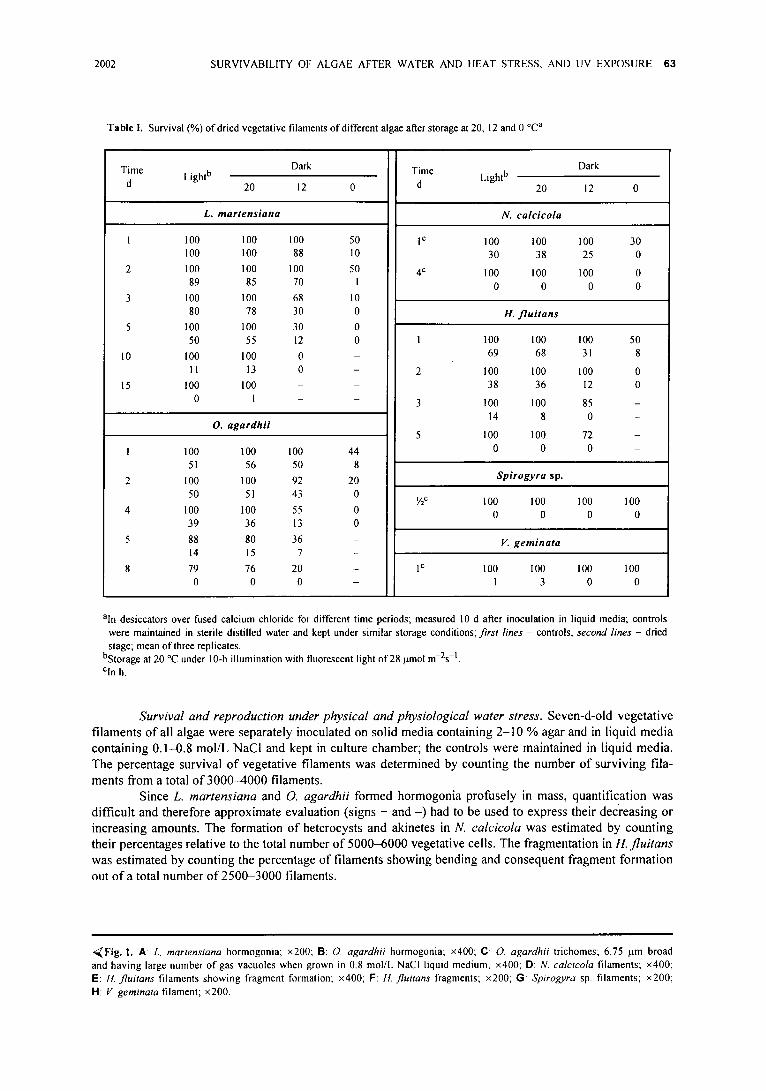
,~Fig. !. A: L. martensiana hormogonia; • B: O. agardhii hormogonia; • C: O. agardhii trichomes; 6.75 p.m broad and having large number of gas vacuoles when grown in 0.8 mol/L NaCI liquid medium: • D: N. calcicola filaments; • E: tl. f luitans filaments showing fragment formation; • F: It. fluitans fragments; • G: Spirogyra sp. filaments; • H: V. geminata filament; •
64 S.C. A G R A W A L and V. SINGH V01.47
~5
o
.=_
=,5
t ~
O
o
J
~ v
'4] ~ . ~ +
~ + ~ +
E
o r I I I
~ <
C ,
t L

2002 SURVIVABILITY OF AI.GAE AFTER WATER AND HEAT STRESS, AND UV EXPOSURE 65
it%
==
7"
+
+.5
o
c)
o
.E x
I
E
j ~
Survival of filaments following exposure to heat and UV. The vegetative filaments of all algae separately suspended in sterile distilled water (adjusted to pH 7.5), were placed at 42--48 ~ Aliquots were withdrawn after 15-60 min and then inoculated into culture media, placed in a culture cham- ber, and scored for percentage survival of vegetative fila- ments.
The vegetative filaments of all algae separately placed in 10 mL sterile distilled water, spread in open Petri dishes (90 mm diameter) were exposed to UV light from a Philips germicidal lamp (main output at 254 nm and a fluence rate of 3.2 W/m2. The energy fluence of UV light which was obtained by increasing the time of exposure from 5 to 40 min ranged from 0�9 to 7.68 kJ/m 2. After irradiation, the material was centrifuged, transferred to fresh culture media and kept in darkness for I d to avoid photoreactivation and then maintained in a culture chamber under normal 16/8 h light--dark cycle; then the percentage survival of vegetative filaments was determined. Materials not irradiated but trea- ted similarly served as controls.
R E S U L T S AND DISCUSSION
L. martensiana and O. agardhii reproduced in culture through hormogonia (Fig. I A, B), while N. calcicola through hormogonia (Fig. I D) and akinetes. In N. calcicola, about 4 % of vegetative cells existed as heterocysts in 15-d-old culture (Fig. I D), and 4-8 % of vegetative cells differenti- ated into akinetes in 45-60-d-old culture. 11. fluitans repro- duced in culture through fragmentation (Fig. I E, F). Its filaments (10-40 %) showed bending and fragment format- ion in 30-45-d-old culture. Spirogyra sp. (Fig. IG) and V. geminata (Fig. 1 H) did not form any reproductive stage or organ in culture. In Spirogyra sp., no reproductive stage was observed even in material collected from nature. V. geminata was observed to form reproductive organs and co-spores.
Viability of dried filaments�9 L. martensiana and O. agardhii tolerated desiccation much better than H. flui- tans, N. calcicola, Y. geminata and Spirogyra sp. (Table I); this might be due to their osmotically active nature of vege- tative cells since both could almost equally tolerate even 0.8 mol/L NaCI (Table II). l[owever, it is not a rule that desiccation-tolerant alga is also a salt-tolerant onei Chlorella vulgaris, a desiccation-tolerant alga was found to be salt- sensitive; it probably tolerated desiccation by its unicellular nature and/or chemical composition of the cell wall contain- ing sporopollenin (Agrawal and Singh 2001).
Although L. martensiana and O. agardhii survived nearly equally on the highly agarized media and in NaCI- containing liquid media (Table II), the L. martensiana fila- ments tolerated desiccation at 20 ~ by 5 d better than O. agardhii filaments which tolerated a similar desiccation level for only 5 d (Table I); this may be explained by L. martensiana sheath.
66 s.c AGRAWAL and V. SINGtl Vol 47
H. f lui tans tolerated desiccation at 20 ~ (to some extent only) for 3 d while Spirogvra sp., V. gemi- nata and N. calcicola did not tolerate a similar level of desiccation even for �89 1 and 4 h, respectively (Table l). L. martensiana, O. agardhii and H. fluitans tolerated desiccation almost equally whether stored in light or dark at 20 ~ but lowering of temperature from 20 to 12 and finally to 0 ~ decreased the via- bility of algae progressively (Table 1). The sensitivity to frost ran somewhat parallel with that to desiccation (Table I). Similar results were obtained with Rhizoclonium hieroglyphicum, Cladophora glomerata and Oscillatoria acuminata (Agrawal and Singh 1999a,b, 2000).
Survival and reproduction o f algae under physical and physiological water stress. Growing algae on high-agar media and in NaCl-containing liquid media decreased vegetative survival at different levels in all of them. Hormogonium formation in L. marterL, iana and O. agardhii, and heterocyst and akinete format- ion in 11. f lui tans (Table lI) was diminished. In Spirogyra sp. these growth conditions avoided induction of conjugation, in V. geminata, the formation of reproductive organs was inhibited. Hence water stress was not helpful in inducing reproduction in algae but decreased it and evoked cell death. Evans (1958) showed that survival o f pond algae after desiccation had no relation to the production of spores, Roelofs and Oglesby (1970) observed that akinete formation was not a result of unfavorable conditions, as akinetes were seen during the exponential phase of growth. Water stress also decreased vegetative survival and heterocyst form- ation in Anabaena iyengarii, Westiellopsis prolifiea and Nostochopsis lobatus, akinete formation in PRho- phora oedogonia, A. iyengarii, W. prolifiea and N. lobatus, and zoosporangia formation in Cladophora glomerata and Rhizoclonium hieroglyphicum (Agrawal and Singh 1999a, b).
The breadth o f O. agardh!i filaments increased from 4.5-6.0 lam in control to 5.3-6.8 lam when they were grown in 0.8 mol/L NaCl-containing medium; in this medium O. agardhii cells also developed large number o f gas vacuoles (Fig. 1C). The blue-green alga Microcys t i s f i rma accumulated in medium with 0.9 mol/L NaCI a large amount of polyols (Erdmann and Schiewer 1984); after growth in 0.8 mol/L NaCI-containing medium, Oscillatoria acuminata developed a thin sheath (Agrawal and Singh 1999b).
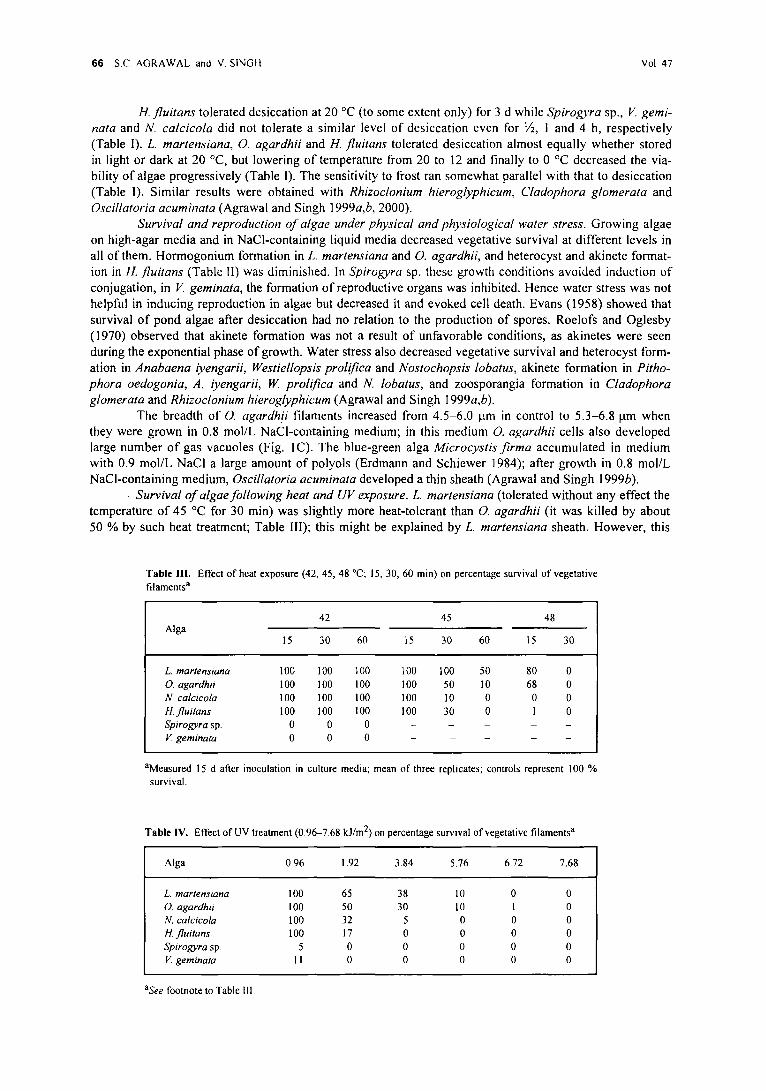
�9 Survival o f algae fol lowing heat and UV exposure. L. martensiana (tolerated without any-effect the temperature o f 45 ~ for 30 min) was slightly more heat-tolerant than O. agardhii (it was killed by about 50 % by such heat treatment; Table 1II); this might be explained by L. martensiana sheath. However, this
Table IlL Effect of heat exposure (42~ 45, 48 ~ 15, 30, 60 min) on percentage survival of vegetalive filaments a
Alga 42 45 48
15 30 60 15 30 60 15 30
L. martensiana 100 100 100 100 100 50 80 0 O. agardh# 100 100 100 100 50 10 68 0 N. calcicola I O0 1 O0 1 O0 1 O0 10 0 0 0 tl. fluitans I O0 1 O0 1 O0 1 O0 30 0 I 0 Spirogyra sp. 0 0 0 . . . . .
geminata 0 0 0 . . . . .
aMeasured 15 d after inoculation in culture media; mean of three replicates; controls represent 100 % survival.
Table IV. Effect of UV treatment (0.96-7.68 kJ/m 2) on percentage survival of vegetative filaments a
Alga 0.96 1.92 3.84 5.76 6.72 7.68
L. martensiana 100 65 38 10 0 0 O. agardhii 100 50 30 10 1 0 N. calcicola 100 32 5 0 0 0 H. fluitans 1 O0 17 0 0 0 0 Spirogyra sp. 5 0 0 0 0 0
geminata 11 0 0 0 0 0
aSee |botnote to Table [II.

2002 SURVIVABILITY OF ALGAE AFTER WATER AND HEAT STRESS, AND UV EXPOSURE 67
hyal ine sheath was not able to protect the alga against U V light, since both L. martensiana and O. agardhii
were nearly equal ly UV-to le ran t as the other algae (Table IV). The presence o f the extracel lular p igment
scy tonemin (Garcia-Pichel et al. 1992) and mycospor ine- l ike compounds (Garcia-Pichel and Cas tenho lz
1993) in terrestrial and sha l low-water blue-green algae was assumed to be important as a sun screen in U V -
light protection.
V. geminata ( isolated from damp-soi l surface) and Spirogyra sp. ( isolated from a fresh-water ditch)
were found to be a lmost equal ly sensit ive to desiccation, frost, salt, heat and UV light. Consider ing, on the
o ther hand, that N. calcicola and H. fluitans ( isolated from a fresh-water pond and tank) tolerated these
stress factors much more than above ment ioned algae (Table l - IV) , algal body form and compos i t ion rather
than habitats can be recognized as primary factors in the tolerance to var ious stress conditions.
The authors thank the Head, Department of Botany, University of Allahabad (India) for providing laboratory facilities, and the University Grants Commission (New Delhi) for financial support in the form of a research grant.
REFERENCES
AGRAWAI, S.C., SINGH V.: Viability of dried vegetative cells and the formation and germination of reproductive structures in Pitho- phora oedogonia, Cladophora glomerata and Rhizoclonium hieroglyphicum under water stress. Folia Microbiol. 44, 63-70 (1999a).
AGRAWAL S.C., SINGH V.: Viability of dried vegetative trichomes, formation of akinetes and heterocysts and akinete germination in some blue-green algae under water stress. Folia Microbiol. 44, 411--418 (1999b).
AGRAWAL S.C., SINGH V.: Vegetative survival, akinete formation and germination in three blue-green algae and one green alga in relat- ion to light intensity, temperature, heat shock and UV exposure. Folia Microbiol. 45, 439--446 (2000).
AGRAWAL S.C., SINGH V.: Viability of dried cells, and survivability and reproduction under water stress, low light, heat and UV expo- sure in Chlorella vulgaris. IsraelJ.Plant Sci. 49, 27-32 (2001).
BUZER J.S., DOHMEIER R.A., DU TOIL D.R.: The survival of algae in dry soils exposed to high temperatures for extended time periods. Phycologia 24, 249-251 (1985).
CAIOLA M.G., BILLI D., FRIEDMANN E.I.: Effect of desiccation on envelopes of the Cyanobacterium Chroococcidiopsis sp. (Chroo- coccales). Eur.J.Phycol. 31, 97-105 (1996).
CAMERON R.E., BLANK G.R.: Desert algae: soil crusts and diaphanous substrata as algal habitats, pp. 1-45 in Jet Propulsion Labora- tory Pasadena Tech.Rep. no. 32-971, Pasadena (USA) 1966.
DANILOV R.A., EKELUND N.G.A.: Effects of increasing doses of UV-B radiation on photosynthesis and motility in Chlamydomonas reinhardtiL Folia Microbiol. 45, 41-44 (2000).
ERDMANN N., SCHIEWER U.: Cell size changes as indicator of salt resistance of blue-green algae. Arch.Hydrobiol. (Suppl.) 67, 431-439 (1984).
EVANS J.H.: The survival of freshwater algae during dry periods - I. An investigation of the algae of five small ponds. J.Ecol. 46, 149- 168 (1958).
FRITSCH F.E.: The morphology and ecology of an extreme terrestrial form of Zygnema ericetorum (KUTZ.) HASS. Ann.Bot. 30, 135-149 (1916).
GARCIA-PICHEL F., CASTENHOLZ R.W.: Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl.Environ. Microbiol. 59, 163-169 (1993).
GARCIA-PICHEL F., SttERRY N.D., CASTENHOLZ R.W.: Evidence for an ultraviolet sunscreen role of the extracellular pigment scyto- nemin in the terrestrial cyanobacterium Chlorogloeopsis sp. Photochem.Photobiol. 56, 17-23 (1992).
I tAWES I., HOWARD-WILI,IAMS C., VINCENT W.I z.: Desiccation and recovery of Antarctic cyanobacterial mats. Polar Biol. 12, 587-594 (1~2).
MOEWUS F.: Die Vererbung des Geschlechts bei Verschiedenen Rassen von Protosiphon botryoides. Arch.Protistenkd. 86, 1-57 (1935).
NICHOLS H.W., BOLD H.C.: Trichosarcina polymorpha gen. et sp.nov. ,L Phycol. I, 34-38 (1965). ROELOFS T., OGLESBY R.T.: Ecological observations on the planktonic cyanophyte Gloeotrichia echinulata. Limnol.Oceanogr. 15,
224-229 (1970). STAH L E.: Uber die Ruhezustande der Vaucheria geminata. Bot. Z. 37, 129-137 (1879). STANIER R.Y., KUNISAWA R., MANDEL M., COHEN B.: Purification and properties of a unicellular blue-green alga (order Chroo-
coccales). BacterioLRev. 35, 171-205 (1971). TOMASELLI L., GIOVANNETTI L. : Survival of diazotrophic cyanobacteria in soil. World,L MicrobioL Biotechnol. 9, I 13-116 (1993). WHITTON B.A.: Survival and dormancy of blue-green algae, pp. 109-167 in Survival and Dormancy of Microorganisms (Y. ltenis,
Ed.). John Wiley & Sons, New York 1987.





























