Embed Size (px)
Citation preview

ilable at ScienceDirect
Toxicon 58 (2011) 112–122
Contents lists ava
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
Oral toxicity of extracts of the microcystin-containing cyanobacteriumPlanktothrix agardhii to the medaka fish (Oryzias latipes)
Chakib Djediat a, David Moyenga b, Mélodie Malécot b, Katia Comte b, Claude Yéprémian b,Cécile Bernard b, Simone Puiseux-Dao b, Marc Edery b,*
a Plateforme de microscopie électronique, Muséum National d’Histoire Naturelle, 12, rue Buffon, F-75231 Paris cedex 05, FrancebUMR 7245 CNRS-MNHN Molécules de communication et adaptation des micro-organismes, Muséum National d’Histoire Naturelle, 12, rue Buffon, F-75231 Pariscedex 05, France
a r t i c l e i n f o
Article history:Received 10 January 2011Received in revised form 10 May 2011Accepted 17 May 2011Available online 25 May 2011
Keywords:Microcystin-containing cyanobacteriumMedaka fishToxicityToxin immunolocalization
* Corresponding author. Tel.: þ33 140793126; faxE-mail address: [email protected] (M. Edery).
0041-0101/$ – see front matter � 2011 Elsevier Ltddoi:10.1016/j.toxicon.2011.05.011
a b s t r a c t
As previously demonstrated the medaka fish appears to offer a good model for studies ofmicrocystins (MCs) effects. Since cyanobacterial toxins are released with other moleculesin the aquatic environment when the producers are dying, in this study, we performedadditional experiments in order to compare the described effects obtained with the puretoxin microcystin-LR (MC-LR), among the most toxic MCs, to those induced by complexextracts of an MCs-producer Planktothrix agardhii, strain PMC 75.02 and a natural bloomcontaining the MCs-producer P. agardhii. The toxicity of these extracts containing severalvariants of MC was determined in adult medaka treated by gavage. Extracts of an MCs-freestrain of P. agardhii (PMC 87.02) were assayed for comparison. Extracts effects were ana-lysed on two tissues, liver and intestine by means of photon and transmission electronmicroscopy. MC was localized in these tissues by immunocytochemistry. No effect wasdetectable with extracts of the MCs-free P. agardhii strain. The two MCs-P. agardhii extracts(strain and natural bloom) were able to induce harmful effects in the liver and intestine ofthe medaka fish in acute intoxication by gavage. In these target organs as shown by toxinimmunolocalization, reactions leading to cell disjunction and lysis were observed appar-ently associated with an immune reaction implying MC containing macrophages. Theseeffects are similar to those previously described with photonic microscopy in medakatreated with pure MC-LR with additional results obtained under the electron microscope.Since no significant effect was detected with the MCs-free (PMC 87.02) extract, we thenconclude that MCs, even in complex association with other cyanobacterial components,should be responsible for the toxic effects observed in treated fish.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction produced by several cyanobacterial taxa, including species
Cyanobacterial blooms, which more or less frequentlyappear in ground water used for drinking or recreationalactivities, can impair water quality and poses risks toanimal and human health (Codd et al., 2005). Onefrequently investigated group of cyanotoxins consists of thecyclic heptapeptide microcystins (MCs). MC toxins are
: þ33140793594.
. All rights reserved.
belonging to generaMicrocystis, Anabaena and Planktothrix.These cyclic heptapeptides contain both L- and D-aminoacids, and an unusual hydrophobic C20 D-amino acid,commonly known as ADDA (3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4,6-dienoic acid). Microcystinsare named according to their variable 2-amino acids; forexample, microcystin-LR (MW: 995) contains leucine (L)and arginine (R). Over 80 MCs have been successfully iso-lated and identified, the most commonly occurring andmost studied of which is MC-LR (Codd et al., 1999, 2005).This toxin is also the one with the greatest capacity

C. Djediat et al. / Toxicon 58 (2011) 112–122 113
of inhibiting protein phosphatases (PP-1 and PP-2A;Malbrouck et al., 2004 a,b). The acute toxicity of MCs inmammals is characterized by fulminant intrahepatic hae-morrhage, followed by liver failure and death. Analogous,although not identical, pathological changes to those re-ported for mammals have also been described in fishtreated with purified MC or cyanobacterial material(Råbergh et al., 1991; Tencalla and Dietrich, 1997; Atencioet al., 2008, 2009).
During the last decade, several studies have assessed thebioaccumulation and effects of cyanobacterial toxins inaquatic organisms (Fischer and Dietrich, 2000a,b; Freitas deMagalhães et al., 2001; Malbrouck et al., 2003; Ernst et al.,2006; Atencio et al., 2009). There are clear indications thatcyanobacterial toxins may have adverse effects on fishunder field conditions (Freitas de Magalhães et al., 2001).Several studies have also investigated the effects of cya-nobacterial toxins on embryo development in aquaticanimals (Oberemm et al., 1999; Jacquet et al., 2004;Palikova et al., 2007). Moreover they have a tumorpromoting effect in the rodent liver (Ohta et al., 1992; Itoet al., 1997). Microcystins, especially MC-LR, causeadverse effects in mammals, birds, fish and molluscs, andare recognized as constituting a potent stress factor andhealth hazard in aquatic ecosystems when heavy blooms oftoxin-producing cyanobacteria appear.
The low concentrations of MC-LR found even incontaminated fish tissues, make it necessary to devisea selective and sensitive histochemical method for theidentification and localization of MCs in target organs. Inaddition to immunohistochemistry, several analyticalmethods are used, however, immunohistochemistry canprovide essential data, such as the relatively precise in situlocalization of MCs.
The effects of subacute exposure to MCs have beenextensively studied following exposure both to pure vari-ants (MC-LR: Billam et al., 2008; Prieto et al., 2009; MC-RR:Gupta et al., 2003; MC-YR: Gupta et al., 2003), and also tocyanobacterial strains (Microcystis aeruginosa: Tencalla andDietrich, 1997; Fischer and Dietrich, 2000a; Fischer et al.,2000; Planktothrix rubescens: Ernst et al., 2006) or cyano-bacterial blooms (Li et al., 2005; Hao et al., 2008; Atencioet al., 2008, 2009). In studies of strains of cyanobacteriaor of blooms, fish were exposed either to extracts (Haoet al., 2008) or directly to the cyanobacteria themselves(Ernst et al., 2006), in various ways (gavage: Fischer et al.,2000; IP injection: Li et al., 2005; in the diet: Atencioet al., 2009). As described in these experiments the puretoxins and the strain or bloom extracts seem usually toproduce similar effects. The liver was the main target,showing haemorrhage, dissociation of the hepatocytes,apoptosis and necrosis (Tencalla and Dietrich, 1997; Fischerand Dietrich, 2000a; Fischer et al., 2000). These effectsincreased with the level and duration of exposure (Fischeret al., 2000; Li et al., 2005), and appeared after briefexposures to high concentrations. The intestine and thekidneys were also targeted, resulting in disruption and celldeath (Fischer and Dietrich. 2000a; Ernst et al., 2006;Atencio et al., 2009), and the heart and gills could alsobe moderately affected (Ernst et al., 2006; Atencio et al.,2009). Immunohistological studies have pinpointed the
localization of microcystins, indicating their co-localizationwith the affected areas in the liver (Fischer and Dietrich,2000a; Fischer et al., 2000; Ernst et al., 2006), in theintestine (Ernst et al., 2006) and in the kidneys (Fischer andDietrich, 2000a; Ernst et al., 2006).
The medaka fish appeared to offer a good model forstudies of microcystins since it had been shown to besensitive to these toxins (Mezhoud et al., 2008; Deng et al.,2010; Djediat et al., 2010). In the present study, experi-ments were conducted with three extracts of the cyano-bacterium Planktothrix agardhii. Two extracts were fromcultured strains (PMC 75.02 and PMC 87.02) isolated fromthe lake of Viry-Châtillon (Ile de France, France) (Yéprémianet al., 2007). P. agardhii strain PMC 75.02 is able tosynthesizemicrocystins and PMC 87.02 is microcystins-free(Yéprémian et al., 2007). The third extract was froma natural P. agardhii bloom collected from the GrandeParoisse Lake (Ile de France, France) (Catherine et al., 2008).The toxicity of these three extracts that could be related tomicrocystins as well as other cyanobacterial metaboliteswas determined in adult medaka treated by gavage andobserving tissues (liver and intestine) bymeans of photonicand transmission electron microscopy. Immunocytochem-istry was also used to investigate the presence of MCs andsome toxic related events in contaminated fish tissues.
2. Materials and methods
2.1. Cyanobacterial strains and growing conditions
This study was carried out on two strains of P. agardhii,PMC 75.02 and PMC 87.02, collected from the lake of Viry-Châtillon (Ile de France: 2123004.2100E, 48140003.3300N)and maintained in the PMC (Paris Museum Collection).Both strains were monoclonal cultures, as they had beenisolated from a single filament of a field sample. PMC 75.02was selected as a microcystin-producer and PMC 87.02 as anon-MC producer as shown by LC/MSmethods (Yéprémianet al., 2007).
Large volumes of cultures (8 L for each strain inNalgene� polycarbonate carboys) were grown at 20 �C in Z8aerated medium and subjected to constant mechanicalshaking. They were exposed to a 16/8h light/dark cycle ina cooled Incubator Sanyo MIR 553, the light being providedby Osram FM 11W/730 fluorescent tubes, had an12 � 5 mmol s�1 m�2 (PAR) average irradiance measuredwith a LICOR LI-250 quantum-meter, equipped with an LI-190SB quantum sensor.
2.2. Preparation of crude extracts
After incubating for 9 weeks at 20 �C, cells in the lateexponential growth phase (OD750nm ¼ 0.6) were collectedby centrifugation at 6000g for 20 min at 4 �C, and thenlyophilised (Labconco apparatus). 500 mg of lyophilisedcells were used for toxin extraction, and placed in 4 tubes(15 mL Falcon tubes) to increase the efficiency of sonication(3 � 1 min, 100% high power, Elma Transsonic T 490 DH in80% methanol). After centrifugation at 4000g for 10 min at4 �C, the supernatant was filtered (GF/C 1.2 mm) andevaporated in a Rotavap (Buchi R210). The solid phase

C. Djediat et al. / Toxicon 58 (2011) 112–122114
extract was dissolved in 50% ethanol (vol/vol) and theethanol was completely evaporated. A supplement of20 mg of lyophilised cells was used for the protein phos-phatase 2A inhibition assay (pp2A test).
The natural bloom was collected from the GrandeParoisse Lake (Ile de France, France) in October 2007(further named ID 10026-O7; Catherine et al., 2008). Thewater samples were filtered and the residues were freeze-dried. 500 mg of lyophilised cells were sonicated in 80%methanol, then centrifuged and filtered as describedpreviously. The solid phase extract was dissolved in 50%ethanol (vol/vol) and the ethanol was completelyevaporated.
2.3. Measurements of the MC content
The MC content was assessed using the protein phos-phatase 2A inhibition assay (PP2A) described by Yéprémianet al. (2007), and confirmed by mass spectrometry (LC/MS)methods (data not shown). The pp2A assay was carried outin triplicate for each sample. The microcystin content ofPMC 75.02 was about 0.6 mg MC-LR eq. per mg lyophilisedmaterial. As expected, no microcystins were detected inPMC 87.02. The microcystin content of the natural bloom(ID 10026-O7) was about 0.25 mg MC-LR eq. per mglyophilised bloom. The results were confirmed by massspectrometry (LC/MS) methods (Bodo et al. personalcommunication).
2.4. Fish and treatments
Adult medaka (Oryzias latipes) were placed in 20L glassaquaria with a 12/12h light/dark cycle. The fish were fed 3times per day. The residues of PMC 75.02, PMC 87.02 and P.agardhii bloom ID 10026-O7 were dissolved at a finalconcentration of 1 mg MC-LR eq./mL extract. The fish wereanesthetised with tricaine (100 mg/L), and gavaged witharound 5 mg MC-LR Eq in 5 mL water per g bodyweight. Forboth histological observations and proteomics studies, fivesamples of 6–9 fish (total: 38) were treated with 0.44 mgMC-LR Eq per mL water per g bodyweight for strain PMC75.02. In parallel 5 samples of 2–6 medaka (total: 26) weretreated with equivalent ponderal MCs-free strain PMC87.02 extract. Two series were performed with the MCs-bloom extract (0.53 mg MC-LR Eq per mL water per gbodyweight; total: 28 fish). Several series of negativecontrols treated with distilled water were made (total: 15fish). The fish were fed, and then killed in icy water afteroral exposure for 2 h. For histology and electronmicroscopyanalyses, the liver and intestine of 2–6 medaka per samplewere collected, and immediately fixed. Samples used forphoton microscopy were fixed with a mixture of para-formaldehyde (2%), glutaraldehyde, 0.5%, picric acid 0.5%,and sucrose (0.18M) in 0.1M pH 7.4 Sørensen buffer. For theultrastructural observations the same mixture was appliedexcept for glutaraldehyde (1%), and the fixation was fol-lowed by a postfixation with osmium tetroxide (1%). Inboth cases samples were washed three times in Sørensenphosphate buffer (0.1 M, pH 7,4) in three 10 min successivebaths and were dehydrated in ethanol (50�, 70�, 90� and100�) by three baths at each step of dehydration and then
embedded in a epoxy mixture (Spurr’s resin). Medium andultrathin sections were sliced with diamond knives (Dia-tome) on a Reichert-Jung Ultracut microtome. The semi-thin sections were stained with toluidine blue 1% andsodium borate (1%) in ethanol (70%) or with methyleneblue-basic fuchsin staining and examined under a Nikonphoton microscope. Ultrathin sections were stained witha saturated solution of Uranyle acetate in 50% alcohol andthen observed by transmission electronic microscope(Hitachi H-7100).
The pictures were taken using Hamamatsu CCD camera,and further processed using Adobe Illustrator 10 software.
2.5. Immunocytochemical analysis (photon and transmissionelectron microscopy)
For immunolocalization, tissues of adult medaka werefixed after dissection in a mixture of paraformaldehyde(2%), glutaraldehyde, 0.5%, picric acid 0.5%, and sucrose(0.18M) in 0.1M pH 7.4 Sørensen buffer. Two anti-MCantibodies were used for immunolocalization: a mono-clonal antibody to MCs (Adda specific, AD4G2, Alexis) thatrecognizes all MCs and a monoclonal antibody to MC-LR(MC10E7, Alexis) that recognizes all MCs with Arg in posi-tion 4. We used a dilution 1:250 for both antibodies. Forphoton microscopy, before dehydration (pre-embedding)tissues were immerged in the diluted antibodies solution;a secondary antibody (goat anti-IgG of mouse) coupledwith a horseradish peroxidase allowed the revelation withan immunoperoxydase reaction using a DakoCytomationEnVision þ System, Peroxidase-HRP kit�. Then the sampleswere dehydrated in a graded ethanol series from 50% to100%, before being embedded in Spurr’s resin. Sections andstaining were performed as described above. For trans-mission electron microscopy (TEM), samples were fixed,dehydrated (see above) and then embedded in a non-antigenic Unicryl resin. Ultrathin sections were made andincubated with the same primary antibodies and then witha secondary antibody coupled to gold nanoballs (6 nm ø).The ultrathin sections were treated as above.
3. Results
3.1. Liver
3.1.1. Photon microscopyThe livers of the control animals gavaged with water
showed the normal hepatic structure on semi-thin sections(Fig. 1A–B and Fig. 2A–B). The tissue was compact, with thehepatocytes organized into Remak spans. The cells wereconnected, and contained glycoprotein and glycogen stores(arrow, Fig. 2B). The livers of the animals gavaged with theextract of MCs-free strain PMC 87.02 were similar (Fig. 1Cand D).
In contrast, the livers of the fish exposed to the extract ofMCs-strain PMC 75.02 included areas of cellular lysis (star,Fig. 1E). At higher magnification, cells have lost the char-acteristic shape of hepatocytes and become round (arrow,Fig. 1F); they are no longer connected and some are lysed(star, Fig. 1F).
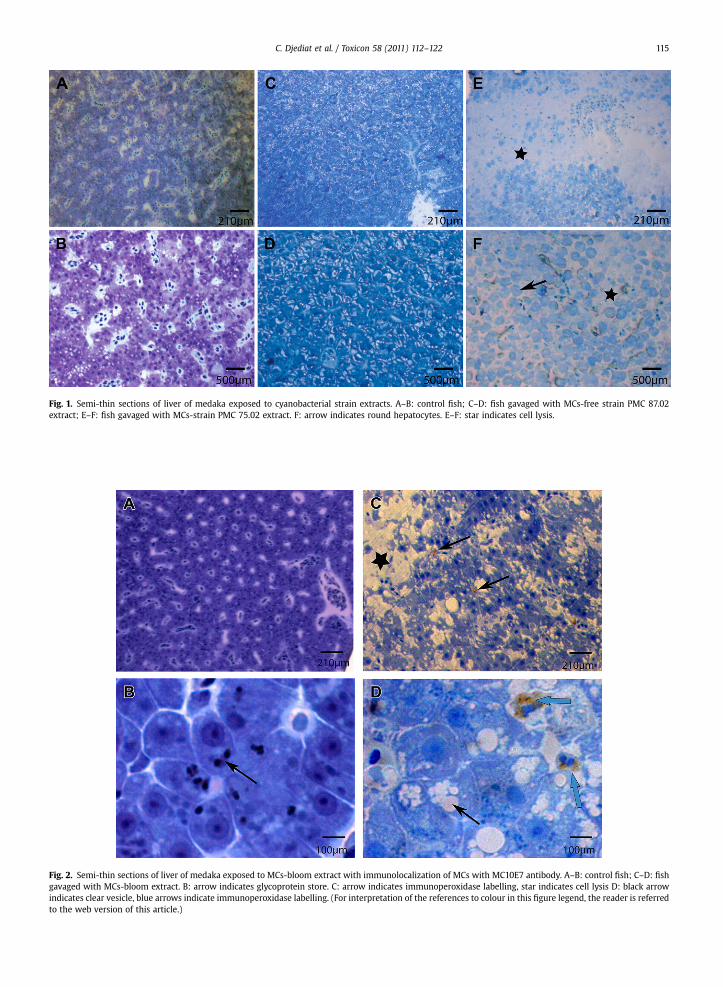
Fig. 1. Semi-thin sections of liver of medaka exposed to cyanobacterial strain extracts. A–B: control fish; C–D: fish gavaged with MCs-free strain PMC 87.02extract; E–F: fish gavaged with MCs-strain PMC 75.02 extract. F: arrow indicates round hepatocytes. E–F: star indicates cell lysis.
Fig. 2. Semi-thin sections of liver of medaka exposed to MCs-bloom extract with immunolocalization of MCs with MC10E7 antibody. A–B: control fish; C–D: fishgavaged with MCs-bloom extract. B: arrow indicates glycoprotein store. C: arrow indicates immunoperoxidase labelling, star indicates cell lysis D: black arrowindicates clear vesicle, blue arrows indicate immunoperoxidase labelling. (For interpretation of the references to colour in this figure legend, the reader is referredto the web version of this article.)
C. Djediat et al. / Toxicon 58 (2011) 112–122 115
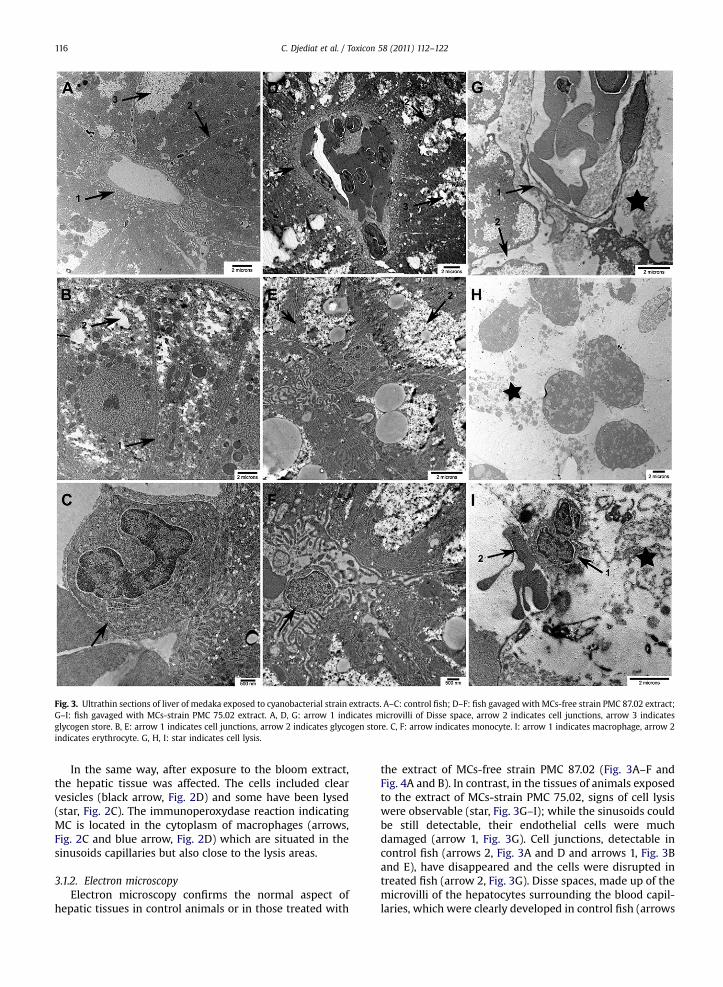
Fig. 3. Ultrathin sections of liver of medaka exposed to cyanobacterial strain extracts. A–C: control fish; D–F: fish gavaged with MCs-free strain PMC 87.02 extract;G–I: fish gavaged with MCs-strain PMC 75.02 extract. A, D, G: arrow 1 indicates microvilli of Disse space, arrow 2 indicates cell junctions, arrow 3 indicatesglycogen store. B, E: arrow 1 indicates cell junctions, arrow 2 indicates glycogen store. C, F: arrow indicates monocyte. I: arrow 1 indicates macrophage, arrow 2indicates erythrocyte. G, H, I: star indicates cell lysis.
C. Djediat et al. / Toxicon 58 (2011) 112–122116
In the same way, after exposure to the bloom extract,the hepatic tissue was affected. The cells included clearvesicles (black arrow, Fig. 2D) and some have been lysed(star, Fig. 2C). The immunoperoxydase reaction indicatingMC is located in the cytoplasm of macrophages (arrows,Fig. 2C and blue arrow, Fig. 2D) which are situated in thesinusoids capillaries but also close to the lysis areas.
3.1.2. Electron microscopyElectron microscopy confirms the normal aspect of
hepatic tissues in control animals or in those treated with
the extract of MCs-free strain PMC 87.02 (Fig. 3A–F andFig. 4A and B). In contrast, in the tissues of animals exposedto the extract of MCs-strain PMC 75.02, signs of cell lysiswere observable (star, Fig. 3G–I); while the sinusoids couldbe still detectable, their endothelial cells were muchdamaged (arrow 1, Fig. 3G). Cell junctions, detectable incontrol fish (arrows 2, Fig. 3A and D and arrows 1, Fig. 3Band E), have disappeared and the cells were disrupted intreated fish (arrow 2, Fig. 3G). Disse spaces, made up of themicrovilli of the hepatocytes surrounding the blood capil-laries, which were clearly developed in control fish (arrows
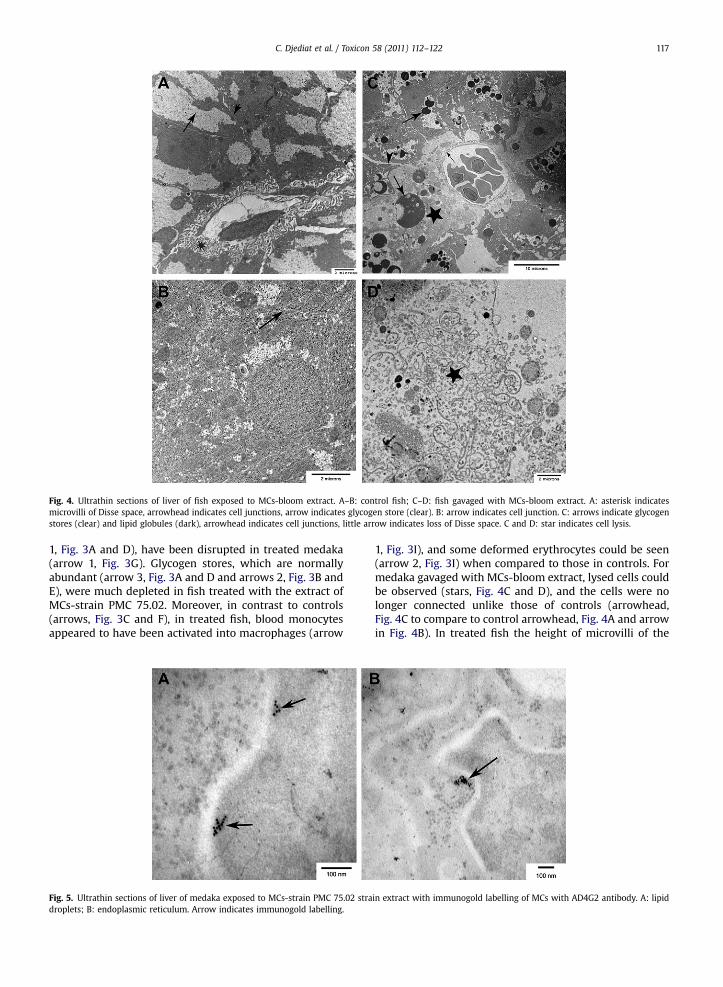
Fig. 4. Ultrathin sections of liver of fish exposed to MCs-bloom extract. A–B: control fish; C–D: fish gavaged with MCs-bloom extract. A: asterisk indicatesmicrovilli of Disse space, arrowhead indicates cell junctions, arrow indicates glycogen store (clear). B: arrow indicates cell junction. C: arrows indicate glycogenstores (clear) and lipid globules (dark), arrowhead indicates cell junctions, little arrow indicates loss of Disse space. C and D: star indicates cell lysis.
C. Djediat et al. / Toxicon 58 (2011) 112–122 117
1, Fig. 3A and D), have been disrupted in treated medaka(arrow 1, Fig. 3G). Glycogen stores, which are normallyabundant (arrow 3, Fig. 3A and D and arrows 2, Fig. 3B andE), were much depleted in fish treated with the extract ofMCs-strain PMC 75.02. Moreover, in contrast to controls(arrows, Fig. 3C and F), in treated fish, blood monocytesappeared to have been activated into macrophages (arrow
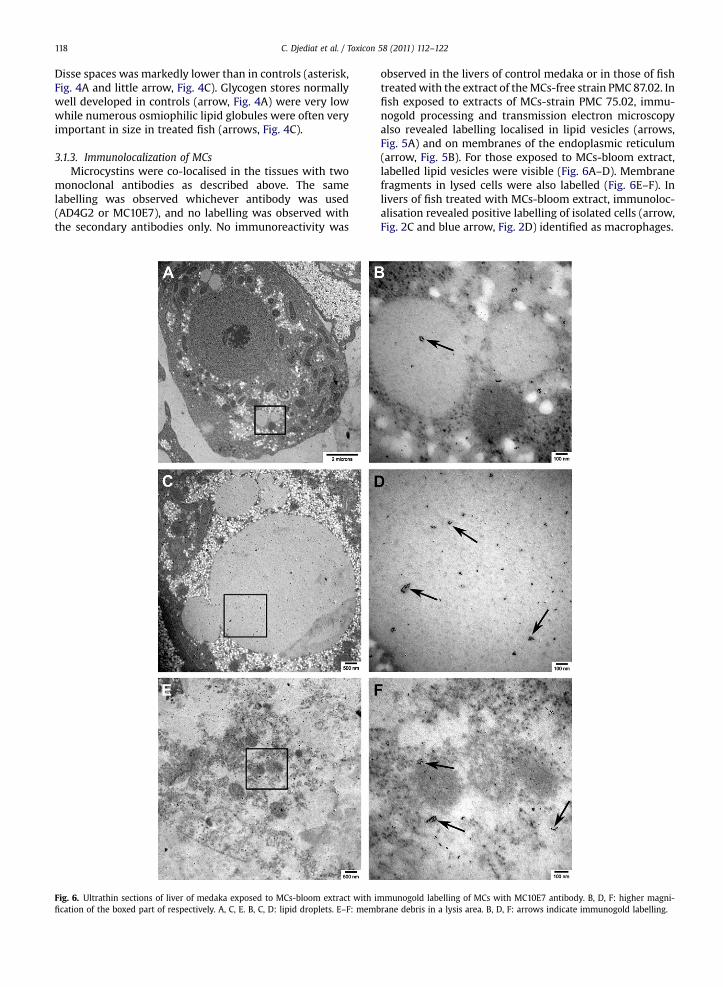
Fig. 5. Ultrathin sections of liver of medaka exposed to MCs-strain PMC 75.02 stradroplets; B: endoplasmic reticulum. Arrow indicates immunogold labelling.
1, Fig. 3I), and some deformed erythrocytes could be seen(arrow 2, Fig. 3I) when compared to those in controls. Formedaka gavaged with MCs-bloom extract, lysed cells couldbe observed (stars, Fig. 4C and D), and the cells were nolonger connected unlike those of controls (arrowhead,Fig. 4C to compare to control arrowhead, Fig. 4A and arrowin Fig. 4B). In treated fish the height of microvilli of the
in extract with immunogold labelling of MCs with AD4G2 antibody. A: lipid
C. Djediat et al. / Toxicon 58 (2011) 112–122118
Disse spaces was markedly lower than in controls (asterisk,Fig. 4A and little arrow, Fig. 4C). Glycogen stores normallywell developed in controls (arrow, Fig. 4A) were very lowwhile numerous osmiophilic lipid globules were often veryimportant in size in treated fish (arrows, Fig. 4C).
3.1.3. Immunolocalization of MCsMicrocystins were co-localised in the tissues with two
monoclonal antibodies as described above. The samelabelling was observed whichever antibody was used(AD4G2 or MC10E7), and no labelling was observed withthe secondary antibodies only. No immunoreactivity was
Fig. 6. Ultrathin sections of liver of medaka exposed to MCs-bloom extract with ification of the boxed part of respectively. A, C, E. B, C, D: lipid droplets. E–F: memb
observed in the livers of control medaka or in those of fishtreatedwith the extract of theMCs-free strain PMC 87.02. Infish exposed to extracts of MCs-strain PMC 75.02, immu-nogold processing and transmission electron microscopyalso revealed labelling localised in lipid vesicles (arrows,Fig. 5A) and on membranes of the endoplasmic reticulum(arrow, Fig. 5B). For those exposed to MCs-bloom extract,labelled lipid vesicles were visible (Fig. 6A–D). Membranefragments in lysed cells were also labelled (Fig. 6E–F). Inlivers of fish treated with MCs-bloom extract, immunoloc-alisation revealed positive labelling of isolated cells (arrow,Fig. 2C and blue arrow, Fig. 2D) identified as macrophages.
mmunogold labelling of MCs with MC10E7 antibody. B, D, F: higher magni-rane debris in a lysis area. B, D, F: arrows indicate immunogold labelling.
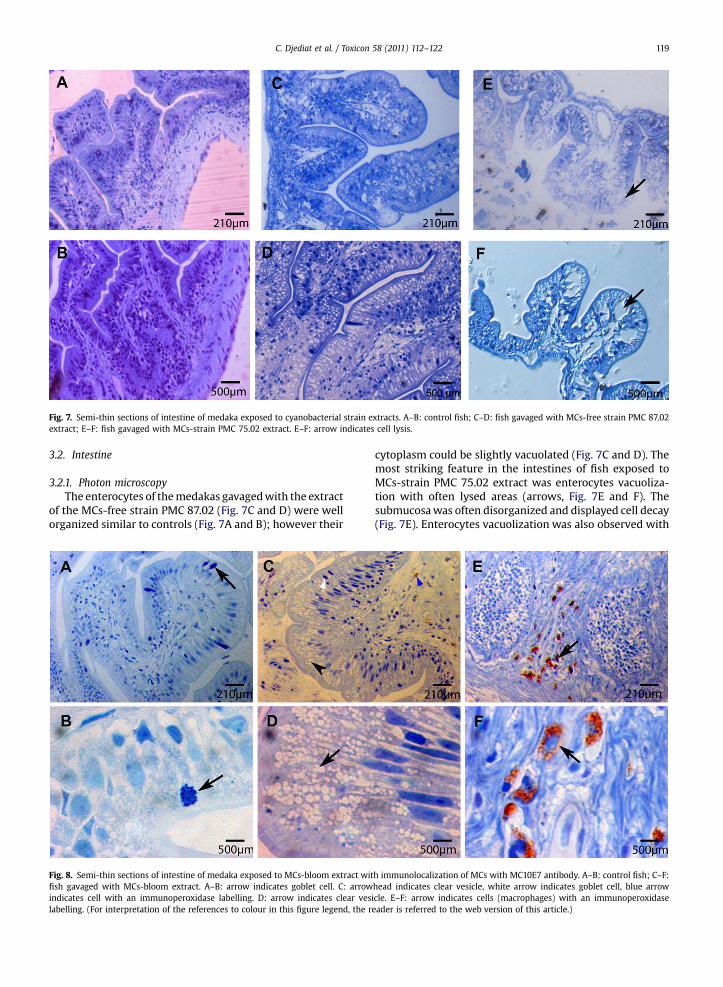
Fig. 7. Semi-thin sections of intestine of medaka exposed to cyanobacterial strain extracts. A–B: control fish; C–D: fish gavaged with MCs-free strain PMC 87.02extract; E–F: fish gavaged with MCs-strain PMC 75.02 extract. E–F: arrow indicates cell lysis.
C. Djediat et al. / Toxicon 58 (2011) 112–122 119
3.2. Intestine
3.2.1. Photon microscopyThe enterocytes of themedakas gavagedwith the extract
of the MCs-free strain PMC 87.02 (Fig. 7C and D) were wellorganized similar to controls (Fig. 7A and B); however their
Fig. 8. Semi-thin sections of intestine of medaka exposed to MCs-bloom extract witfish gavaged with MCs-bloom extract. A–B: arrow indicates goblet cell. C: arrowindicates cell with an immunoperoxidase labelling. D: arrow indicates clear veslabelling. (For interpretation of the references to colour in this figure legend, the r
cytoplasm could be slightly vacuolated (Fig. 7C and D). Themost striking feature in the intestines of fish exposed toMCs-strain PMC 75.02 extract was enterocytes vacuoliza-tion with often lysed areas (arrows, Fig. 7E and F). Thesubmucosawas often disorganized and displayed cell decay(Fig. 7E). Enterocytes vacuolization was also observed with
h immunolocalization of MCs with MC10E7 antibody. A–B: control fish; C–F:head indicates clear vesicle, white arrow indicates goblet cell, blue arrowicle. E–F: arrow indicates cells (macrophages) with an immunoperoxidaseeader is referred to the web version of this article.)
C. Djediat et al. / Toxicon 58 (2011) 112–122120
the MCs-bloom extract (arrowhead, Fig. 8C and arrow,Fig. 8D) which was never seen in controls gavaged withwater (Fig. 8A and B). There seem also to be fewer gobletcells (white arrow, Fig. 8C) compared to controls (arrows,Fig. 8 A and B).
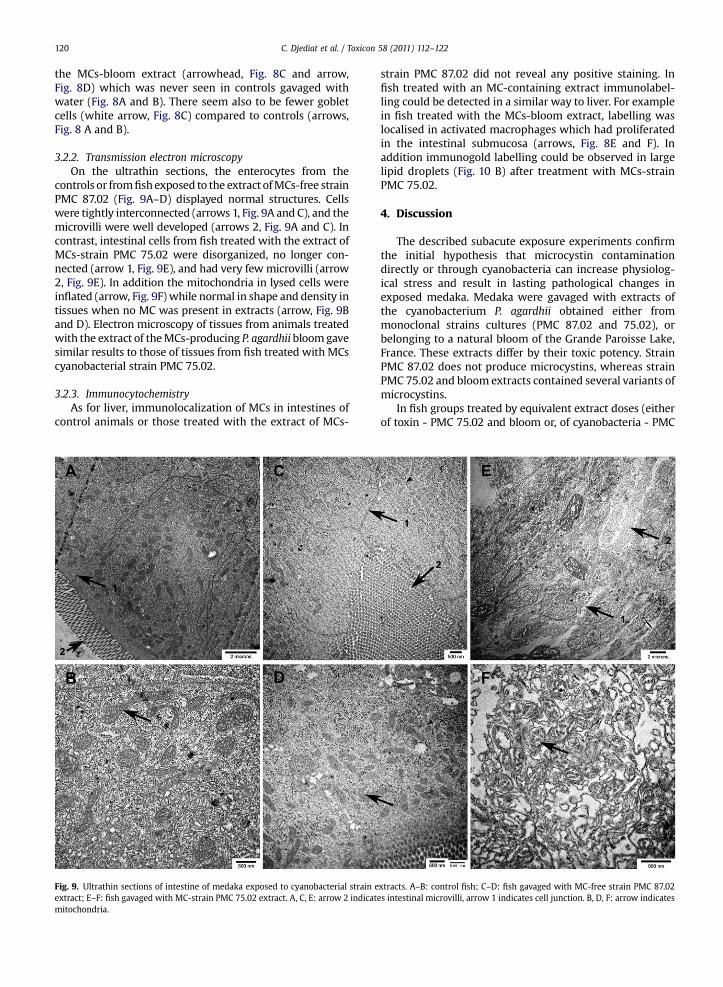
3.2.2. Transmission electron microscopyOn the ultrathin sections, the enterocytes from the
controls or fromfishexposed to the extract ofMCs-free strainPMC 87.02 (Fig. 9A–D) displayed normal structures. Cellswere tightly interconnected (arrows 1, Fig. 9A and C), and themicrovilli were well developed (arrows 2, Fig. 9A and C). Incontrast, intestinal cells from fish treated with the extract ofMCs-strain PMC 75.02 were disorganized, no longer con-nected (arrow 1, Fig. 9E), and had very few microvilli (arrow2, Fig. 9E). In addition the mitochondria in lysed cells wereinflated (arrow, Fig. 9F) while normal in shape and density intissues when no MC was present in extracts (arrow, Fig. 9Band D). Electron microscopy of tissues from animals treatedwith the extract of theMCs-producing P. agardhii bloomgavesimilar results to those of tissues from fish treated with MCscyanobacterial strain PMC 75.02.
3.2.3. ImmunocytochemistryAs for liver, immunolocalization of MCs in intestines of
control animals or those treated with the extract of MCs-
Fig. 9. Ultrathin sections of intestine of medaka exposed to cyanobacterial strain eextract; E–F: fish gavaged with MC-strain PMC 75.02 extract. A, C, E: arrow 2 indicatmitochondria.
strain PMC 87.02 did not reveal any positive staining. Infish treated with an MC-containing extract immunolabel-ling could be detected in a similar way to liver. For examplein fish treated with the MCs-bloom extract, labelling waslocalised in activated macrophages which had proliferatedin the intestinal submucosa (arrows, Fig. 8E and F). Inaddition immunogold labelling could be observed in largelipid droplets (Fig. 10 B) after treatment with MCs-strainPMC 75.02.
4. Discussion
The described subacute exposure experiments confirmthe initial hypothesis that microcystin contaminationdirectly or through cyanobacteria can increase physiolog-ical stress and result in lasting pathological changes inexposed medaka. Medaka were gavaged with extracts ofthe cyanobacterium P. agardhii obtained either frommonoclonal strains cultures (PMC 87.02 and 75.02), orbelonging to a natural bloom of the Grande Paroisse Lake,France. These extracts differ by their toxic potency. StrainPMC 87.02 does not produce microcystins, whereas strainPMC 75.02 and bloom extracts contained several variants ofmicrocystins.
In fish groups treated by equivalent extract doses (eitherof toxin - PMC 75.02 and bloom or, of cyanobacteria - PMC
xtracts. A–B: control fish; C–D: fish gavaged with MC-free strain PMC 87.02es intestinal microvilli, arrow 1 indicates cell junction. B, D, F: arrow indicates

Fig. 10. Ultrathin sections of intestine of medaka exposed to MC-strain PMC 75.02 strain extract with immunogold labelling of MCs with MC10E7 antibody. A:asterisks indicate lipid droplets. B: high magnification of A, lipid droplet, arrow indicates immunogold labelling.
C. Djediat et al. / Toxicon 58 (2011) 112–122 121
75.02 at the preceding concentration and PMC 87.02), onenotable difference was detected: a mortality rate of 45.5%was observedwith the extract of theMCs-strainwhereas themortality induced by the MCs-free strain (3.3%) was insig-nificant. However the extract of MCs-strain PMC 75.02appeared more toxic than the pure MC-LR, which produced18.7% mortality. This is possibly associated with the MCvariants content which can greatly vary depending on the P.agardhii strains (Montfort et al., 2005) and do not respondexactly in the sameway to the pp2A assay as well as they donot induce strictly the same effects in organisms theyintoxicate. But maybe other toxic metabolite(s) (Welker andvon Döhren, 2006) could interfere in a cooperative orantagonistic way.
Microscopy showed that the extract of MCs-free strainPMC 87.02 did not produce any effect on the two tissuesexamined, the liver and the intestine. In this case, thesetissues were similar to those of the control fish, whichhad been gavaged with water. On the contrary in the fishgavaged with the extract of MCs-strain PMC 75.02 orwith the extract of the MCs-P. agardhii bloom, bothtissues were severely damaged. Moreover the effectsproduced by the extract of MCs-strain PMC 75.02 and theMCs-P. agardhii bloom extract were similar to thosedescribed with the pure microcystin-LR (Djediat et al.,2010). The liver included areas of lysed cells, the hepa-tocytes were no longer connected, glycogen and glyco-protein storage were depleted and lipid vesicles hadappeared in the cells. The osmiophilic inclusionsobserved under the photon microscope were in factidentified as lipid globules by electron microscopy. Allare classical responses to toxicological stress, particularlyaccumulation of fat globules that are known to permitsequestration of lipophilic xenobiotic molecules in fish(Biagianti-Risbourg and Bastide, 1995). The intestineshowed similar damages, with isolated enterocytes andlysed cells.
All those effects have been described in fish exposed topure MC-LR (Kotak et al., 1996; Prieto et al., 2009; Djediatet al., 2010) and also to different extracts of cyanobacteriastrains or blooms (Tencalla and Dietrich, 1997; Fischer and
Dietrich, 2000a; Fischer et al., 2000; Li et al., 2005; Ernstet al., 2006; Atencio et al., 2009). They were also similarto those reported in rodents exposed to pure MC-LR (Itoet al., 1997; Hooser, 2000; Guzman et al., 2003; Guptaet al., 2003; Billam et al., 2008).
Immunocytochemistry detection permitted to localizemicrocystins inside macrophages suggesting they hadprobably phagocyted remains of lysed cells containingmicrocystins as suggested by labelling on endoplasmicreticulum membranes of cell lysed areas. Moreover label-lingwas also observed in lipid vesicles which could indicatethat lipophilic MCs while inducing toxic damages, weresequestered inside the lipid compartment to neutralizethem as has been found for pesticides (Biagianti-Risbourgand Bastide, 1995). A few immunohistochemistry studiesof extracts of cyanobacterial strains or blooms (Fischer andDietrich, 2000a; Fischer et al., 2000; Ernst et al., 2006) havealso shown that the microcystins could be located in celllysed areas.
In conclusion, the extracts of the cyanobacteriumP. agardhii for both strain PMC 75.02 and the natural bloomproducing microcystins, are able to induce harmful effectsin the liver and intestine of the medaka fish in subacuteintoxication by gavage. The liver and intestine are targetorgans with reactions leading to classical toxic injuries: celldisjunction and lysis with frequent lipid accumulation.Moreover the animal response seems to include animmune reaction implying macrophages. All these data aresimilar to those observed with pure MC-LR (Djediat et al.,2010). Since no effect was detected with the P. agardhiiclone that does not synthesize MC, this suggests that thetoxicity in all studied cases was related to microcystins.
Conflict of interest
None.
Acknowledgments
This work was supported by grants from the DGS Bio-accessMC, the ANR 07 SEST CYANOTOX 005, the AFSSET

C. Djediat et al. / Toxicon 58 (2011) 112–122122
APR EST 2007 10 and ANSES EST 2010/2/002 to Dr. MarcEdery. The authors are grateful to Dr. Arnaud Catherine forfield sampling in the Lake Grande Paroisse.
References
Atencio, L., Moreno, I., Jos, A., Pichardo, S., Moyano, R., Blanco, A.,Cameán, A.M., 2008. Dose-dependent antioxidant responses andpathological changes in tenca (Tinca tinca) after acute oral exposureto Microcystis under laboratory conditions. Toxicon 52, 1–12.
Atencio, L., Moreno, I., Jos, A., Prito, A., Moyano, R., Blanco, A., Cameán, A.M., 2009. Effects of dietary selenium on the oxidative stress andpathological changes in tilapia (Oreochromis niloticus) exposed toa microcystin-producing cyanobacterial water bloom. Toxicon 53,269–282.
Biagianti-Risbourg, S., Bastide, J., 1995. Hepatic perturbation induced byan herbicide (atrazine) in juveniles grey mullet Liza ramada (Mugi-lidae, teleostei): an ultrastructure study. Aquatic Toxicol. 31, 217–229.
Billam, M., Mukhi, S., Tang, L., Gao, W., Wang, J.S., 2008. Toxic responseindicators of microcystin-LR in F344 rats following a single-dosetreatment. Toxicon 51, 1068–1080.
Catherine, A., Quiblier, C., Yepremian, Y., Got, P., Groleau, A., Vinçon-Leite, B., Bernard, C., Troussellier, M., 2008. Collapse of Plankthotrixagardhii perennial bloom and microcystin dynamics in response toreduced phosphate concentrations in a temperate lake. FEMSMicrobiol. Ecol. 65, 61–73.
Codd, G.A., Bell, S., Kaya, K., Ward, C., Beattie, K., Metcalf, J., 1999. Cya-nobacterial toxins, exposures routes and human health. Eur. J.Phycology 34, 415–418.
Codd, G.A., Morrison, L.F., Metcalf, J.S., 2005. Cyanobacterial toxins: riskmanagement for health protection. Toxicol. Appl. Pharmacol. 203,264–272.
Deng, D.F., Zheng, K., The, F.C., Lehman, P.W., The, S.J., 2010. Toxicthreshold of dietary microcystin -LR for quart medaka. Toxicon 55,787–794.
Djediat, C., Malécot, M., de Luze, A., Bernard, C., Puiseux Dao, S., Edery, M.,2010. Localization of microcystin-LR in medaka fish tissues aftercyanotoxin gavage. Toxicon 55, 531–535.
Ernst, B., Hoeger, S.J., O’Brien, E., Dietrich, D.R., 2006. Oral toxicity of themicrocystin-containing cyanobacterium Planktothrix rubescens inEuropean whitefish (Coregonus lavaretus). Aquat. Toxicol. 79, 31–40.
Fischer, W.J., Dietrich, D.R., 2000a. Pathological and Biochemical charac-terization of microcystin-induced Hepatopancreas and kidneydamage in carp (Cyprinus carpio). Toxicol. Appl. Pharmacol. 164,73–81.
Fischer, W.J., Dietrich, D.R., 2000b. Toxicity of the cyanobacterial cyclicheptapeptide toxins microcystin-LR and -RR in early life-stages of theAfrican clawed frog (Xenopus laevis). Aquat. Toxicol. 49, 189–198.
Fischer, W., Hitzfeld, B.C., Tencalla, F., Eriksson, J.E., Mikhailov, A.,Dietrich, D.R., 2000. Microcystin-LR toxicodynamics, inducedpathology, and immunhistochemical localisation in livers of blue-green algae exposed rainbow trout (Oncorhynchus mykiss). Toxicol.Sci. 54, 365–373.
Freitas de Magalhães, V., Moares Soares, R., Azevedo, S.M.F.O., 2001.Microcystin contamination in fish from the Jacarepagua lagoon (Riode Janeiro, Brazil): ecological implication and human health risk.Toxicon 39, 1077–1085.
Gupta, N., Pant, S.C., Vijayaraghavan, R., Rao, P.V., 2003. Comparativetoxicity evaluation of cyanobacterial cyclic peptide toxin microcystinvariants (LR, RR, YR) in mice. Toxicology 188, 285–296.
Guzman, R.E., Solter, P.F., Runnegar, M.T., 2003. Inhibition of nuclearprotein phosphatase activity in mouse hepatocytes by the cyano-bacterial toxin microcystin-LR. Toxicon 41, 773–781.
Hao, L., Xie, P., Fu, J., Li, G., Xiong, Q., Li, H., 2008. The effect of cyano-bacterial crude extract on the transcription of GST mu, GST kappa andGST rho in different organs of goldfish (Carassus auratius)Aquat.Toxicol 90, 1–7.
Hooser, S.B., 2000. Fulminant hepatocyte apoptosis in vivo followingmicrocystin-LR administration to rats. Toxicol. Pathol. 28, 726–733.
Ito, E., Kondo, F., Terao, K., Harada, K.I., 1997. Neoplastic nodular formationin mouse liver induced by repeated intraperitoneal injections ofmicrocystin-LR. Toxicon 35, 1453–1457.
Jacquet, C., Thermes, V., de Luze, A., Puiseux-Dao, S., Bernard, C., Joly, J.S.,Bourrat, F., Edery, M., 2004. Effects of microcystin-LR on developmentof medaka fish embryos (Oryzias latipes). Toxicon 43, 141–147.
Kotak, B.G., Semalulu, S., Fritz, D.L., Prepas, E.E., Hrudey, S.E., Coppock, R.W., 1996. Hepatic and renal pathology of intraperitoneally adminis-tered microcystin-LR in rainbow trout (Oncorhynchus mykiss). Toxicon34, 517–525.
Li, L., Xie, P., Chen, J., 2005. In vivo studies on toxin accumulation in liverand ultrastructural changes of hepatocytes of the phytoplanktivorousbighead carp i.p.-injected with extracted microcystins. Toxicon 46,533–545.
Malbrouck, C., Trausch, G., Devos, P., Kestemont, P., 2003. Hepatic accu-mulation and effects of microcystin-LR on juvenile goldfish Carassiusauratus L. Comparative Biochemistry and Physiology Part C. Toxicol.Pharmacol. 135, 39–48.
Malbrouck, C., Trausch, G., Devos, P., Kestemont, P., 2004a. Effect ofmicrocystin-LR on protein phosphatase activity in fed and fastedjuvenile goldfish Carassius auratus L. Toxicon 43, 295–301.
Malbrouck, C., Trausch, G., Devos, P., Kestemont, P., 2004b. Effect ofmicrocystin-LR on protein phosphatase activity and glycogen contentin isolated hepatocytes of fed and fasted juvenile goldfish Carassiusauratus L. Toxicon 44, 927–932.
Mezhoud, K., Bauchet, A.-L., Château-Joubert, S., Praseuth, D., Marie, A.,François, J.C., Fontaine, J.J., Jaeg, J.P., Cravedi, J.P., Puiseux-Dao, S.,Edery, M., 2008. Proteomic and phosphoproteomic analysis of cellularresponses in medaka fish (Oryzias latipes) following oral gavage withmicrocystin-LR. Toxicon 51, 1431–1439.
Montfort, D.O., Holland, P., Sprosen, J., 2005. Method for detecting classesof microcystins by combination of protein phosphatase inhibitionassay and ELISA: comparison with LC-MS. Toxicon 45, 199–206.
Oberemm, A., Becker, J., Codd, G.A., Steinberg, C., 1999. Effects of cyano-bacterial toxins and aqueous crude extracts of cyanobacteria on thedevelopment of fish and amphibians. Environ. Toxicol. 14, 77–88.
Ohta, T., Nishiwaki, R., Yatsunami, J., Komori, A., Suganuma, M., Fujiki, H.,1992. Hyperphosphorylation of cytokeratins 8 and 18 by microcystin-LR, a new liver tumor promoter, in primary cultured rat hepatocytes.Carcinogenesis 13, 2443–2447.
Palikova, M., Krejci, R., Hilscherova, K., Babica, P., Navratil, S., Kopp, R.,Blaha, L., 2007. Effects of different cyanobacterial biomasses and theirfractions with variable microcystin content on embryonal develop-ment of carp (Cyprinus carpio L.). Aquat. Toxicol. 8, 312–318.
Prieto, A.I., Jos, A., Pichardo, S., Moreno, I., de Sotomayor, M.A., Moyano, R.,Blanco, A., Cameãn, A.M., 2009. Time-dependent protective efficacityof Trolox (vitamin E analog) against microcystin-induced toxicity intilapia (Oreochromis niloticus). Environ. Toxicol. 24, 563–579.
Råbergh, C.M.I., Bylund, G., Eriksson, J.E., 1991. Histopathological effects ofmicrocystin-LR, a cyclic peptide toxin from the cyanobacterium (blue-green alga) Microcystis aeruginosa, on common carp (Cyprinus carpioL). Aquat. Toxicol. 20, 131–146.
Tencalla, F., Dietrich, D., 1997. Biochemical characterization of microcystintoxicity in rainbow trout (Oncorhynchus mykiss). Toxicon 35, 583–595.
Welker, M., von Döhren, H., 2006. Cyanobacterial peptides-nature's owncombinatorial biosynthesis. FEMS Microbiol. Rev. 30, 530–563.
Yéprémian, C., Gugger, M.F., Briand, E., Catherine, A., Berger, C., Quiblier, C.,Bernard, C., 2007. Microcystin ecotypes in a perennial Planktothrixagardhii bloom. Water Res. 41, 4446–4456.




















