Embed Size (px)
Citation preview

Xeroderma Pigmentosum Group C Protein Possesses a High AffinityBinding Site to Human Centrin 2 and Calmodulin*
Received for publication, March 12, 2003, and in revised form, July 29, 2003Published, JBC Papers in Press, July 30, 2003, DOI 10.1074/jbc.M302546200
Aurel Popescu‡, Simona Miron§, Yves Blouquit, Patricia Duchambon, Petya Christova¶,and Constantin T. Craescu�
From INSERM U350 and Institut Curie-Recherche, Centre Universitaire, Batiments 110-112, 91405 Orsay, France
Human centrin 2 (HsCen2), a member of the EF-handsuperfamily of Ca2�-binding proteins, is commonly asso-ciated with centrosome-related structures. The proteinis organized in two domains, each containing two EF-hand motifs, but only the C-terminal half exhibits Ca2�
sensor properties. A significant fraction of HsCen2 islocalized in the nucleus, where it was recently foundassociated with the xeroderma pigmentosum group Cprotein (XPC), a component of the nuclear excision re-pair pathway. Analysis of the XPC sequence (940 resi-dues), using a calmodulin target recognition software,enabled us to predict two putative binding sites. Thebinding properties of the two corresponding peptideswere investigated by isothermal titration calorimetry.Only one of the peptides (P1-XPC) interacts strongly(Ka � 2.2 � 108 M�1, stoichiometry 1:1) with HsCen2 in aCa2�-dependent manner. This peptide also binds, with asimilar affinity (Ka � 1.1 � 108 M�1) to a C-terminalconstruct of HsCen2, indicating that the interactionwith the integral protein is mainly the result of thecontribution of the C-terminal half. The second peptide(P2-XPC) failed to show any detectable binding either toHsCen2 or to its C-terminal lobe. The two peptides in-teract with different affinities and mechanisms with cal-modulin. Circular dichroism and nuclear magnetic res-onance were used to structurally characterize thecomplex formed by the C-terminal domain of HsCen2with P1-XPC.
Centrin (also called caltractin) is a Ca2�-binding proteinhighly conserved in diverse evolutionary lineages, includingalgal, higher plant, invertebrate, and mammalian cells (1, 2). Itis an acidic protein of 19.5 kDa belonging to the highly con-served EF-hand calmodulin (CaM)1 superfamily. Comparative
sequence analysis suggests that centrins consist of two struc-tural domains, each containing two putative Ca2�-binding EF-hand motifs. In humans, three centrin isoforms (HsCen1 toHsCen3) have been identified so far (3–5) with variable se-quences and different tissue and cell distributions. HsCen1 andHsCen2 are highly similar to each other (sequence identity84%) and to the algae centrin (68 and 71%, respectively),whereas HsCen3, discovered lately (5), has a more distantsequence; it shows only 54% identity with both HsCen1 andHsCen2, and is slightly closer to Cdc31, the centrin equivalentin yeast Saccharomyces cerevisiae (59% sequence identity). Alarge part of the sequence diversity among the centrins occurswithin the first 20 residues of the N-terminal domain, thathave no counterpart in the “standard” Ca2�-binding protein,CaM.
The centrins are usually found in association with the mi-crotubule organizing centers (centrosomes in animal cells, andspindle pole bodies in yeast) that are cytoplasmic organellesencountered in almost all eukaryotic cells, with an importantrole in microtubule structural and temporal organization (1, 2).HsCen2 is ubiquitously expressed but was first discovered inthe distal lumen of centrioles, where its presence is required fornormal centriole duplication during the cell cycle (6). A largefraction of the cellular centrin is not permanently associatedwith the centrosome (7), but fractionates with the cytoplasmand nuclei in human cells. The precise function of these pools isnot well understood and constitutes a subject of intense inves-tigation. For instance, the presence of HsCen2 in the nuclearfractions is thought to play a role in coordinating the nuclearand cytoplasm events during the division cycles. Recently,studies conducted in the Hanaoka group (8) shed a new light onthe possible role of nuclear HsCen2 fraction in the nucleotideexcision repair (NER) process. Nuclear excision repair is amajor pathway for recognition and removal of bulky DNA le-sions such as the UV photoproducts, or carcinogen adducts. Itsdysfunction produces severe disorders in humans such as xe-roderma pigmentosum, a hereditary disease characterized by ahigh photosensitivity and a large incidence of sunlight-inducedcancer. One of the molecular component involved in severalxeroderma pigmentosum forms is the XPC complex, a het-erodimer composed of the XPC gene product (XPC) and HR23B,the human homologue of yeast Rad23 B. XPC complex plays akey role in the initial phase of NER and is involved in therecognition of the DNA damage. NER is of great importance forthe maintenance of the genomic integrity, but the molecularmechanism of the NER pathway involving damage recognition,excision, gap-filling, and ligation steps has not been elucidated(9).
* This work was supported in part by the Institut Curie, INSERM,and CNRS. The costs of publication of this article were defrayed in partby the payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
‡ Present address: Faculty of Physics, University of Bucharest,077125 Bucharest, Romania.
§ Recipient of the European Marie Curie fellowship.¶ Present address: Inst. of Organic Chemistry, Bulgarian Academy of
Sciences, 1113 Sofia, Bulgaria.� To whom correspondence should be addressed: INSERM U350 and
Institut Curie-Recherche, Centre Universitaire, Batiments 110-112,91405 Orsay Cedex, France. Tel.: 33-1-69-86-31-63; Fax: 33-1-69-07-53-27; E-mail: [email protected].
1 The abbreviations used are: CaM, calmodulin; CD, circular dichro-ism; HsCen2, human centrin 2; SC-HsCen2, a short C-terminal frag-ment of human centrin2 (Thr94–Tyr172); ITC, isothermal titration cal-orimetry; NER, nuclear excision repair; P1-XPC, Asn847–Arg863
fragment of xeroderma pigmentosum protein; XPC, xeroderma pigmen-tosum complementing group C protein; P2-XPC, Pro703–Ala720 frag-
ment of XPC; SPB, spindle pole body; BisTris, 2-[bis(2-hydroxyethyl)-amino]-2-(hydroxymethyl)propane-1,3-diol; HSQC, heteronuclear sin-gle-quantum correlation.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 41, Issue of October 10, pp. 40252–40261, 2003© 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org40252
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from

According to the recent work of Araki et al. (8), the XPCcomplex may contain an additional component, HsCen2, which,together with HR23B, stabilizes in a cooperative manner XPC,and thus stimulates the NER activity in vitro. This observationopens a new field of investigation of the centrin cellular func-tions, with the possibility that centrin may fill the gap betweenthe DNA nuclear repair process and the functions of the mitoticspindle apparatus.
Recent biochemical and biophysical studies performed in ourand other laboratories have provided a wealth of physicochem-ical data on the structure and Ca2� binding properties of cen-trins of various origins. From this, in agreement with thesequence alignment analysis, it appears that the integral cen-trins are composed of two independent domains, each contain-ing two putative EF-hand motifs (10, 11). In contrast to theChlamydomonas centrin (11), which is able to bind four Ca2�
ions with significant affinity, HsCen2 exhibits one strong andone weak binding site, localized in the C-terminal domain (10,12). Structural NMR studies provided evidence that the C-terminal domain of HsCen2 is conformationally sensitive toCa2� binding, and folds into an open conformation with a largeexposed hydrophobic surface (10). The capacity of both proteinsto bind amphiphilic peptides, primarily mediated by the C-terminal domain (10, 11), strongly supports the hypothesis of aCa2�-dependent regulatory role of centrins. The next step inthis direction is the search for specific molecular targets andexploration of conformational, energetic, and functional as-pects of the corresponding intermolecular interactions.
In this work, we focused on a sequence-based identificationof the XPC binding site to HsCen2 and on the thermodynamicand structural characterization of the intermolecular interac-tions. ITC experiments showed that one of the two predictedbinding peptides derived from XPC binds with high affinity toHsCen2, the isolated HsCen2 C-terminal domain, and CaM. Inall cases, the peptide binding is Ca2�-dependent, but stoichi-ometry and molecular mechanism seem to be different forcentrin and CaM. CD and preliminary NMR experiments en-abled us to characterize some structural aspects of the complexbetween one XPC peptide and the C-terminal domain ofHsCen2.
EXPERIMENTAL PROCEDURES
Protein Expression and Purification—Recombinant proteins HsCen2and SC-HsCen2 (Thr94–Tyr172) were overexpressed in Escherichia coliand purified as described previously (10, 12). For 15N-labeled samples,we used a culture medium (M9) containing 15NH4Cl (1.5 g/liter) as thesole source of nitrogen, and the induction step with isopropyl-1-thio-�-D-galactopyranoside (0.1 mM) was prolonged to 18 h.
Target Peptides—Two peptides from the human XPC protein, encom-passing the sequences Asn847–Arg863 (P1-XPC) and Pro703–Ala720 (P2-XPC) were purchased from Biofidal (Vaulx en Velin, France). Puritywas greater than 95%, as assessed by high pressure liquid chromatog-raphy analysis.
CD Spectroscopy—CD experiments were performed on a Jasco 715CD spectrometer equipped with a Peltier temperature control unit.Far-UV spectra were recorded between 195 and 250 nm at 20 °C using1-mm quartz cells. Spectra were collected as an average of four scans,with a scan speed of 20 nm/min and a response time of 1 s. Samples (30�M) were dissolved in Tris-HCl buffer (10 mM) containing 100 mM NaCland 2 mM CaCl2. Temperature denaturation curves were obtained be-tween 20 and 95 °C, with a temperature increasing rate of 1 °C/min.
Isothermal Titration Calorimetry—Thermodynamic parameters ofmolecular interactions between human centrin or calmodulin and thetarget peptides at 30 °C were investigated by ITC using a MicroCalMCS instrument (MicroCal Inc., Northampton, MA). The proteins andpeptides were equilibrated in the same buffer containing 20 mM Tris (orBisTris), pH 6.5, 100 mM NaCl, and Ca2� (2 mM) or EDTA (5 mM). In astandard experiment, the protein (7–20 �M) in the 1.337-ml calorimetercell was titrated by the peptide (generally 10 times more concentrated)by �30 successive automatic injections of 7–10 �l each. The first injec-tion of 2–3 �l was ignored in the final data analysis. Integration of the
peaks corresponding to each injection and correction for the base linewere done using Origin-based software provided by the manufacturer.Fitting of the data to various interaction models results in the stoichi-ometry (n), equilibrium binding constant (Ka), and enthalpy of complexformation (�H). The reported thermodynamic parameters represent anaverage of at least two experiments. Usually, control experiments,consisting of injecting peptide solutions into the buffer, were performedto evaluate the heat of dilution.
NMR Spectroscopy—NMR samples (0.7–1.2 mM) were obtained bydissolving the lyophilized protein in deuterated Tris-HCl buffer (20 mM,pH 6.5) containing 100 mM NaCl and 5 mM CaCl2. NMR spectra wererecorded on a Varian Unity 500 NMR spectrometer equipped with atriple resonance probe and a Z-field gradient, at 308 K. Homonuclearand heteronuclear [15N-1H]HSQC two-dimensional spectra in 1H2Owere performed using standard pulse sequences (13, 14). Spectra anal-ysis was carried out using Felix software (Accelrys, San Diego, CA).
RESULTS
Prediction of the Centrin Binding Site of XPC Protein—Theavailable structural and functional data consistently suggestthat HsCen2 is a sensor protein, capable to translate a Ca2�
cellular signal into an activation/inhibition of a target molecule(10). CaM, the prototype of the Ca2� sensor proteins, has alarge and diverse number of known target proteins, analysis ofwhich revealed some molecular characteristics of the CaM-binding sites, including �-helix propensity and the basic am-phiphilic character (15–17). Recently, a web-based data basecontaining the known CaM target sequences and their proper-ties, as well as sequence analysis tools permitting to predictputative CaM binding sites within a given protein sequence,has been developed (17). Assuming that CaM and HsCen2binding sites may share common molecular features, we pro-ceeded to an analysis of the XPC sequence using these tools.Sequence comparison failed to find any region in the XPCprotein sharing significant sequence homology with knownCaM binding sites but did predict two sequences with a goodprobability as putative CaM binding sites. The two peptides(P1-XPC, Asn847–Arg863; and P2-XPC, Pro703–Ala720), situatedin the last quarter of the XPC sequence (Fig. 1A), were associ-ated with the moderate to excellent score of probability (9 and6 for P1-XPC and P2-XPC, respectively). Sequence comparisonwith classical CaM binding motifs suggests that the peptidesbelong to the 1–14 class, including the recognition sites ofmyosin light chain kinase, CaM kinase IV, calcineurin A, andhuman death-associated kinase I. P1-XPC is predicted to be thebest binding sequence with a pattern of bulky hydrophobic sidechains at positions 1, 5, 8, and 14 and a good propensity to forman amphiphilic �-helix (Fig. 1A). It is worth noting that P1-XPC together with Cdc31 binding peptide Kar1 include nega-tively charged side chains, whereas the CaM-binding peptidesare generally positively charged (16).
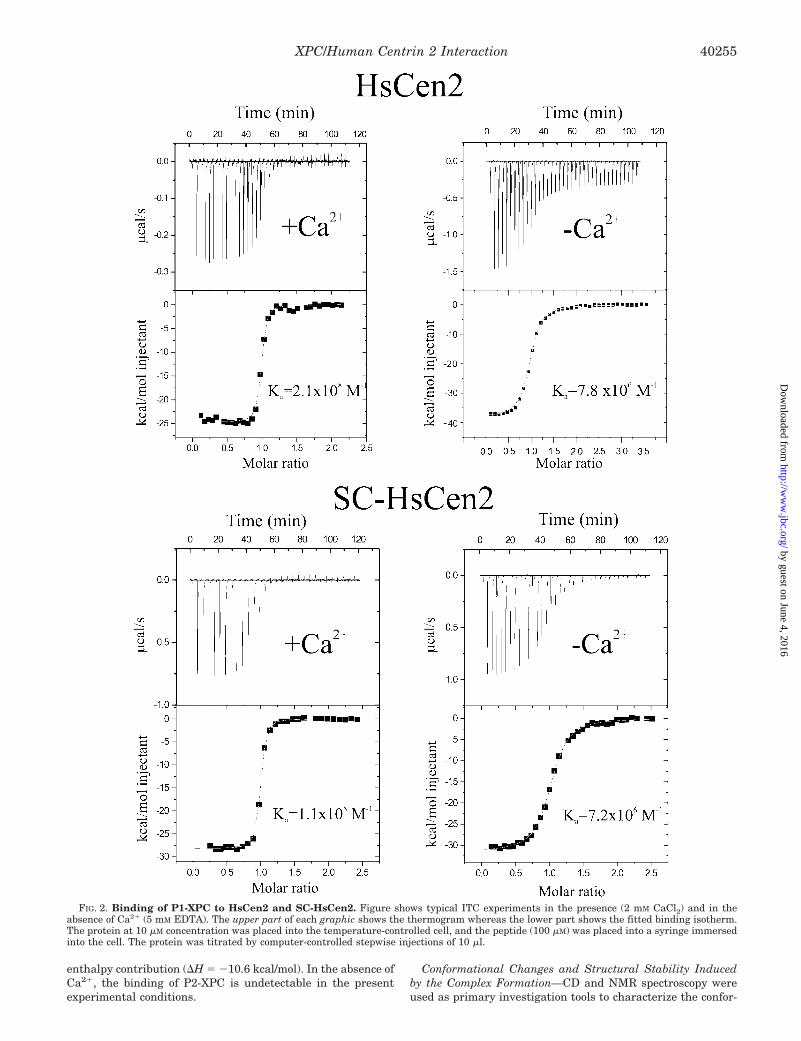
Binding to HsCen2 and Its C-terminal Domain—Binding ofthe P1-XPC and P2-XPC peptides was studied using the ITCmethod with the protein solution in the cell and the peptides inthe syringe. As illustrated in the thermograms of Fig. 2, suc-cessive injections of the peptides are accompanied by exother-mic heat pulses for which the integral decreases to a stablebase line. In the presence of saturating Ca2� concentrations,the data analysis indicates that the integral protein HsCen2binds the P1-XPC peptide with a stoichiometry of 1:1 and ahigh affinity Ka � 2.2 (�0.4) � 108 M�1 (Kd � 4.5 nM). The largereaction enthalpy of approximately �27 kcal/mol largely over-comes the entropic component to the free energy of interaction(Table I). The short C-terminal domain SC-HsCen2 binds thepeptide with a similar stoichiometry, affinity, and binding en-thalpy, strongly suggesting that it represents the major inter-acting domain. Indeed, the free energy of P1-XPC binding toSC-HsCen2 represents 97% of the binding energy of the intactprotein. In the absence of Ca2�, both the integral protein and
XPC/Human Centrin 2 Interaction 40253
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from

the SC-HsCen2 domain are still able to bind exothermally thepeptide, but with a considerably decreased affinity (28 and 17times, respectively) (Fig. 2 and Table I).
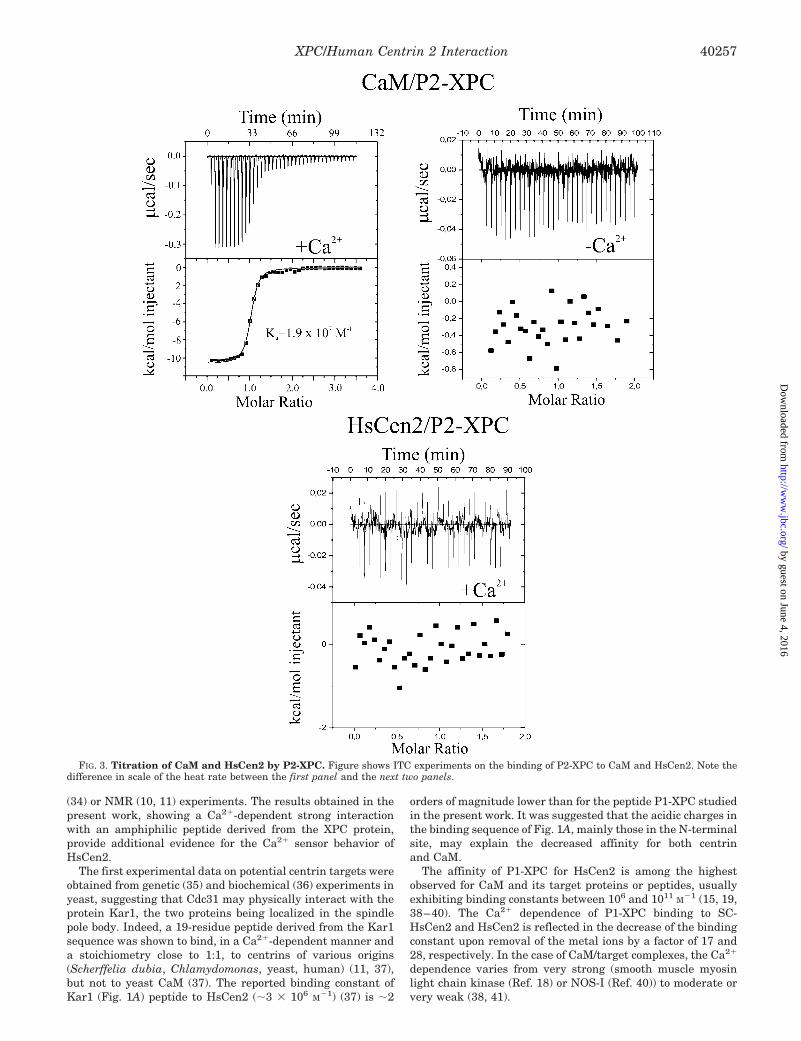
In the case of P2-XPC, the heat rate is very small andremains roughly constant during the titration (Fig. 3), failing toshow the typical transition phase observed for interacting sys-tems. This observation indicates the absence of binding, or avery weak binding (Ka � 103 M�1), that cannot be detected inthe present conditions. Therefore, in agreement with theknowledge-based prediction, HsCen2/Ca2�, mainly through itsC-terminal domain, exhibits a strong binding to one of the twoputative CaM binding sites situated in the C-terminal part ofthe XPC protein.
Analysis of the free energy components (Table I) shows thatthe decrease in binding energy in the absence of Ca2� is theresult of an increase of the unfavorable entropic contribution,only partially compensated by the increase in enthalpy. Thistrend is characteristic both for the intact protein and the shortC-terminal construct, and it may be related to the fact that, inthe absence of Ca2�, the C-terminal domain represents anensemble of highly disordered conformations (10), stabilizationof which upon complex formation gives a negative entropiccontribution.
Measurement of the temperature dependence of the bindingenthalpy allowed us to evaluate the heat capacity changes(�Cp) caused by protein/peptide interactions. For the com-
plexes of P1-XPC with HsCen2 and SC-HsCen2, the negativebinding enthalpy increases significantly when the temperaturevaries between 20 and 34 °C. The heat capacity, calculated asthe correlation coefficient between �H and temperature, haslarge negative values (�995 � 78 and �366 � 49 cal/K mol forHsCen2 and SC-HsCen2, respectively) that correspond to theupper range of values observed for CaM/target peptides (18,19). It was noted (19) that, among the CaM binding peptides,the heat capacity changes were more negative for those exhib-iting a primary interaction with the C-terminal protein domainand having a bulk aromatic side chain in the peptide N-side, asis the case for P1-XPC. Empirical observations in a number ofcases pointed to the existence of a linear correlation betweenthe interaction-induced heat capacity changes and the accom-panying decrease in the water-accessible surface area (20, 21).The corresponding dehydration results in a positive entropiccontribution, essentially caused by the released water molecules.Therefore, the negative heat capacity changes observed in thiswork should be associated with a major contribution of aliphaticand aromatic side chains to the total decrease of the accessiblesurface area upon complex formation (22). This is in agreementwith the large hydrophobic surface exposed by the C-terminaldomain of HsCen2 in the presence of Ca2� ions (10) and isconsistent with a largely apolar protein/peptide interface. Anadditional factor that may account for the large negative valuesof the heat capacity changes observed upon P1-XPC complexformation is the significant conformational changes of both part-ners, as was suggested for many protein/DNA interactions (23).
We also performed titration calorimetric experiments tomeasure the interaction between HsCen2 and melittin, a nat-ural peptide extracted from the bee venom often used as aCaM-binding peptide. In the presence of Ca2�, the binding isweaker than for P1-XPC (Ka � 1.6 � 107 M�1) but is still drivenby a negative enthalpy change (�H � �8 kcal/mol, at 30 °C).
Binding to Calmodulin—The two XPC peptides used in thepresent work have been selected based on the CaM targetconsensus sequence, and the hypothesis that proteins from theEF-hand superfamily may share similar targets. On the otherhand, CaM and centrins could be observed in the same cellularregions (24–26), where proteins possessing CaM-binding mo-tifs (27, 28) were identified. This raised the question of thepossible promiscuity in recognition and binding of the molecu-lar targets by the two EF-hand proteins, and motivated us toinvestigate the binding of P1-XPC and P2-XPC to CaM in thesame physicochemical conditions.
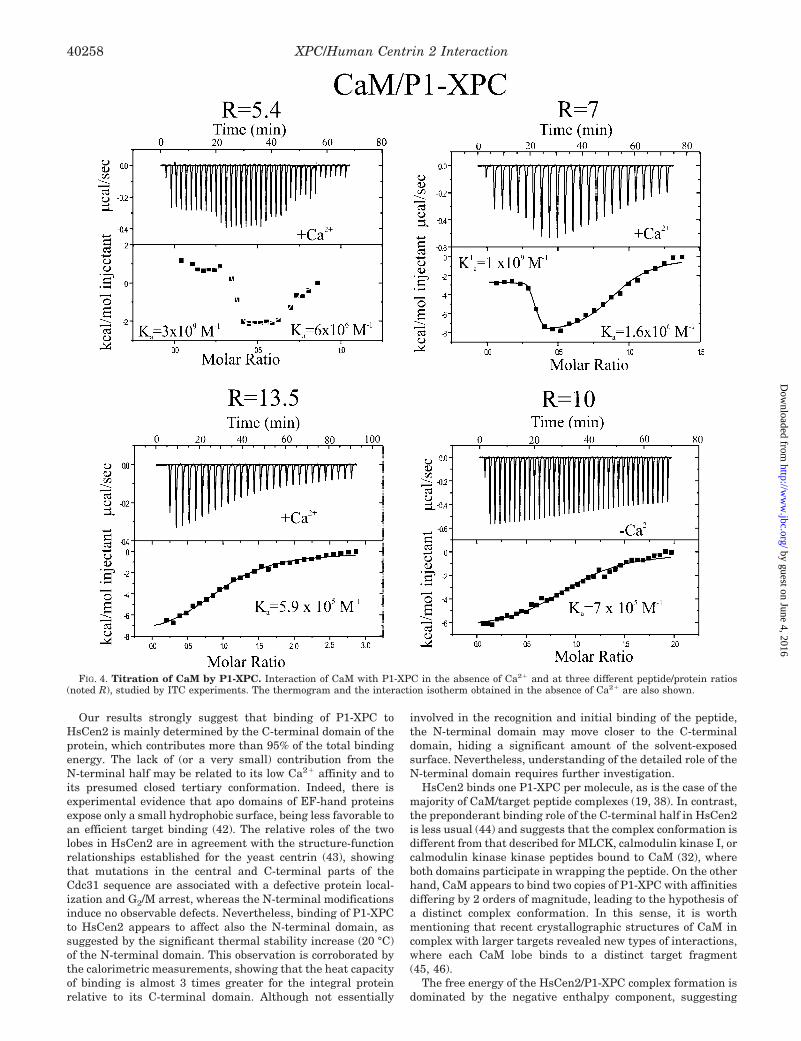
In the presence of Ca2�, the thermogram corresponding tothe titration of CaM by P1-XPC shows a complex pattern,which could not be fitted using a single-site model (Fig. 4),suggesting that CaM exhibits two binding sites with distinctaffinities. When the peptide to protein concentration ratio (R) ischanged from 5.4 to 13.5, the thermogram becomes progres-sively dominated by the low affinity binding contribution, andthe corresponding binding isotherm can be fitted confidently toa single-site model (Fig. 4). This allows us to obtain an estima-tion of the thermodynamic parameters characterizing the weakbinding site: Ka � 5.9 � 105 M�1 and �H � �8.3 kcal/mol.Although the thermograms for lower peptide to protein ratioscould not be modeled confidently, we assume that the affinity ofthe strong binding site is of the order of 108 M�1. P1-XPCbinding to CaM is also Ca2�-sensitive, because in the absenceof the divalent cations, a single low affinity binding site (Ka �7 � 105 M�1) was observed (Fig. 4).
In contrast to HsCen2, the P2-XPC peptide also shows asignificant binding to CaM. The binding isotherm could befitted to a single-binding site model (Fig. 3), with a significantaffinity (Ka � 1.9 � 107 M�1) largely accounted for by the
FIG. 1. Sequence analysis. A, schematic representation of the posi-tion of the two putative CaM-binding sites in the sequence of the XPCprotein. The approximate localization of the regions involved in bindingto HR23B, DNA, and TFIIH complex is indicated by the blue bars. Thesequence of the two studied peptides is shown in comparison with otherCaM-binding fragments and the Kar1 site, that was found to bind yeastand Chlamydomonas centrins. B, wheel representation of the N-termi-nal sequence of P1-XPC showing its amphiphilic character. The hydro-phobic and basic side chains are shown on a green and blue background,respectively. C, superposition of the P1-XPC sequence on similar se-quences from homologous proteins in various organisms (human, yeast,mouse, Drosophila melanogaster, Arabidopsis thaliana, and Caenorh-abditis elegans).
XPC/Human Centrin 2 Interaction40254
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from
enthalpy contribution (�H � �10.6 kcal/mol). In the absence ofCa2�, the binding of P2-XPC is undetectable in the presentexperimental conditions.
Conformational Changes and Structural Stability Inducedby the Complex Formation—CD and NMR spectroscopy wereused as primary investigation tools to characterize the confor-
FIG. 2. Binding of P1-XPC to HsCen2 and SC-HsCen2. Figure shows typical ITC experiments in the presence (2 mM CaCl2) and in theabsence of Ca2� (5 mM EDTA). The upper part of each graphic shows the thermogram whereas the lower part shows the fitted binding isotherm.The protein at 10 �M concentration was placed into the temperature-controlled cell, and the peptide (100 �M) was placed into a syringe immersedinto the cell. The protein was titrated by computer-controlled stepwise injections of 10 �l.
XPC/Human Centrin 2 Interaction 40255
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from
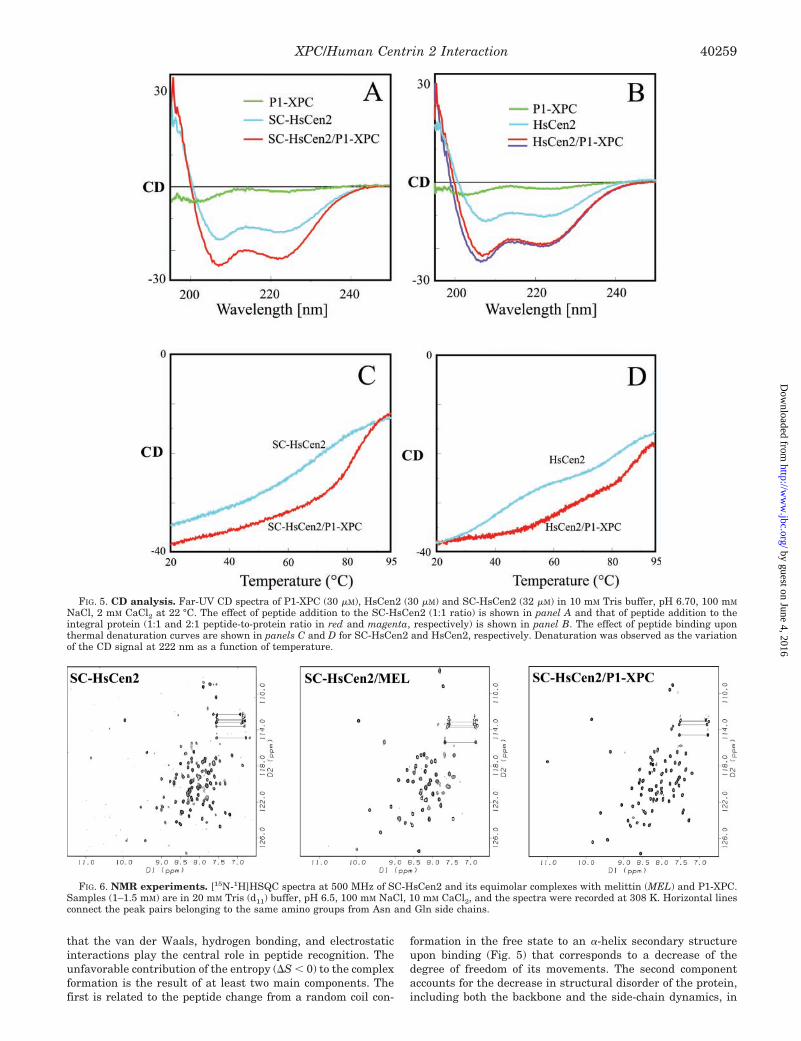
mational properties of the HsCen/P1-XPC interaction. Fig. 5illustrates the far-UV CD experiments on the complex forma-tion of HsCen2 and its C-terminal domain with the P1-XPC inthe presence of Ca2�. The peptide alone exhibits a CD spectrumcharacteristic for a highly disordered structure, as is generallythe case for linear polypeptides of this size. In contrast, theCa2�-bound integral protein and SC-HsCen2 domain exhibit aCD spectrum typical for a well folded protein with a major�-helical content. The spectral characteristics of the helicalsecondary structure are the negative bands at 222 and 207 nmand the positive band at 195 nm (29).
Adding of P1-XPC, at a 1:1 molar ratio, to SC-HsCen2/Ca2�
induces a considerable enhancement of the CD signal (by�50%) with a rough conservation of the relative intensities ofdifferent bands (Fig. 5A). A moderate increase of the negativeellipticity (on the order of 10–15%) could be eventually associ-ated to the conformational rearrangements of the protein, aswas suggested in the case of Ca2� binding to the apo N-termi-nal domain of troponin C (30, 31). However, structural studieson complexes of EF-hand domains with peptide targets showedthat the protein domains undergo only small tertiary changes(32). Therefore, the large variation of the negative band inten-sity at 222 nm, the most characteristic for the � structure,suggests that a fragment of the peptide undergoes a randomcoil-to-helix structural transition upon binding to the protein,bringing a significant contribution to the �-helix CD band. Thisconclusion is corroborated by preliminary NMR studies of theSC-HsCen2/P1-XPc complex.
In the case of the integral protein (Fig. 5B), addition of thepeptide induces a similar (but slightly larger) increase in thespectrum intensity, probably the result of a larger �-helix con-tent in the protein stabilized by the complex formation. Dou-bling the peptide-to-protein ratio (2:1) is not accompanied bysignificant CD changes, in agreement with the ITC-observed1:1 stoichiometry.
The thermal denaturation profile of the Ca2�-bound SC-HsCen2, studied by monitoring the CD signal at 222 nm, showsthat �60% of the �-helical structure unfolds progressively,starting from 20 °C (Fig. 5C). The single smooth transitionextending over more than 40 °C, with the midpoint tempera-ture of 74 °C, is characteristic for loosely organized structures(33). The temperature-induced process is totally reversible, asthe CD spectrum recorded after a rapid restoration of theinitial conditions is indistinguishable from the reference spec-trum. The complex of SC-HsCen2/Ca2� with P1-XPC shows amuch steeper temperature dependence and an increase in tran-sition temperature by 16 °C (Tm � 90 °C), reflecting a morestable structure.
The Ca2�-bound integral protein (Fig. 5D) unfolds reversiblyin two well distinguished steps (midpoint temperatures �47
and �82 °C), corroborating the results of the chemical dena-turation experiments (10). This observation supports the ideathat the two EF-hand domains behave like independent foldingunits, as was also demonstrated for Chlamydomonas centrin(11). Comparison with the denaturation curve of SC-HsCen2may suggest that the N-terminal domain, apparently unable tobind Ca2� ions, is less stable and unfolds first. Complex forma-tion keeps the modular unfolding behavior and increases sig-nificantly the thermal stability. It is interesting to note that thedenaturation mid-temperature of the first step, presumablyinvolving the N-terminal domain is up-shifted by about 20 °C.
Information on the structural and dynamic properties of thecomplex formed by SC-HsCen2/Ca2� and P1-XPC were ob-tained from NMR experiments. Fig. 6 compares the two-dimen-sional [15N-1H]HSQC spectrum of SC-HsCen2 with those of thecomplex formed with melittin and P1-XPC. With the exceptionof those connected by horizontal lines, each cross-peak repre-sents a protein backbone NH group and is situated at thecrossing point of the corresponding proton (horizontal scale)and 15N (vertical scale) resonance frequencies. The Ca2�-boundSC-HsCen2 alone gives a well dispersed spectrum that con-tains more than expected peaks of unequal intensity. Addingmelittin to the protein solution changes considerably the spec-trum and reduces the number of observable cross-peaks (�55peaks), less than the expected number (78 peaks) for this pro-tein construct. The observed signals are globally broader thanin the spectrum of the unbound protein. A careful analysis, ata lower contour level, revealed a peak subset with strong linebroadening, near the detection limit of the spectrometer. Fi-nally, the complex with P1-XPC exhibits a distinct HSQC spec-trum, containing the appropriate number of cross-peaks and amore regular line shape. Nevertheless, the peaks undergoing alarge line broadening in the complex SC-HsCen2/melittin arestill broadened in the SC-HsCen2/P1-XPC complex, albeit at asmaller scale. These findings indicate that, in agreement withthe relative binding affinities of the two peptides, the protein/peptide interaction results in well organized, unique, and sta-ble structures, which can be amenable to a more detailed struc-tural investigation.
DISCUSSION
HsCen2 as a Ca2� Sensor—The sequence similarity withcalmodulin and the ability to bind Ca2� ions have suggestedthat centrins possess a two-domain structural organization andmay play a regulatory role in one or several Ca2�-triggeredprocesses. Previous studies on Chlamydomonas centrin (11) orHsCen2 (10) confirmed the modular organization and demon-strated the functional autonomy of the two domains in Ca2�
binding. Additionally, the Ca2� sensitivity of the global confor-mation for centrins from several organisms were shown by CD
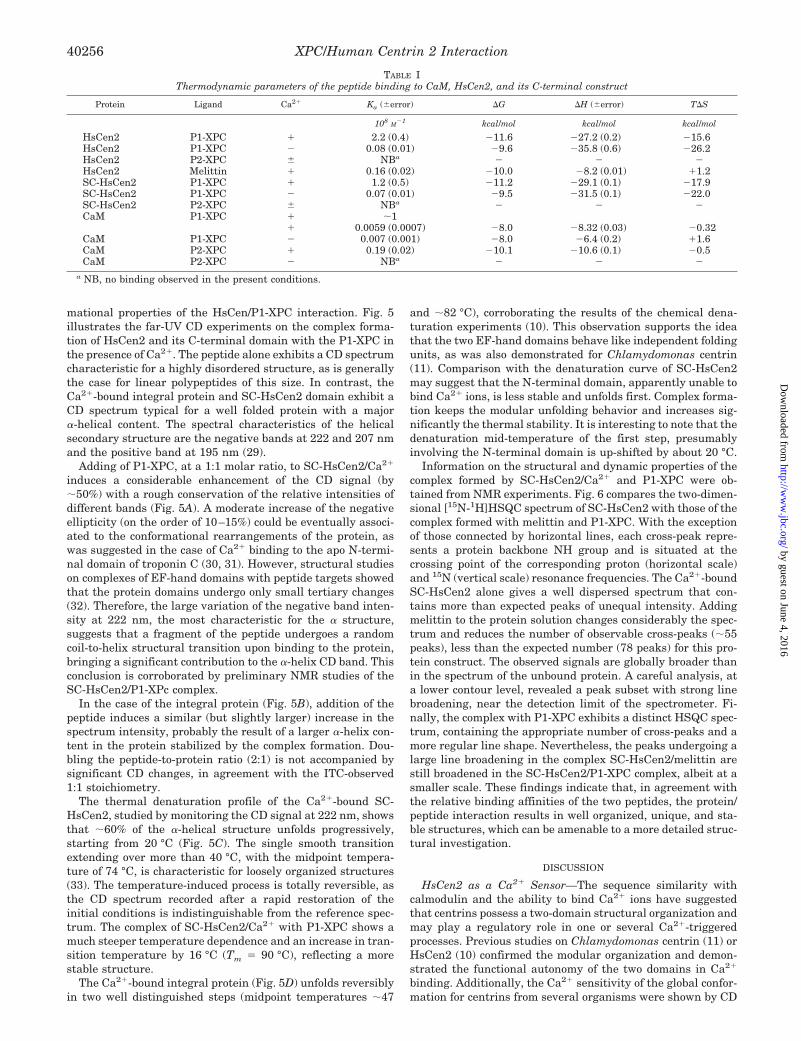
TABLE IThermodynamic parameters of the peptide binding to CaM, HsCen2, and its C-terminal construct
Protein Ligand Ca2� Ka (�error) �G �H (�error) T�S
108M
�1 kcal/mol kcal/mol kcal/mol
HsCen2 P1-XPC � 2.2 (0.4) �11.6 �27.2 (0.2) �15.6HsCen2 P1-XPC � 0.08 (0.01) �9.6 �35.8 (0.6) �26.2HsCen2 P2-XPC � NBa � � �HsCen2 Melittin � 0.16 (0.02) �10.0 �8.2 (0.01) �1.2SC-HsCen2 P1-XPC � 1.2 (0.5) �11.2 �29.1 (0.1) �17.9SC-HsCen2 P1-XPC � 0.07 (0.01) �9.5 �31.5 (0.1) �22.0SC-HsCen2 P2-XPC � NBa � � �CaM P1-XPC � �1
� 0.0059 (0.0007) �8.0 �8.32 (0.03) �0.32CaM P1-XPC � 0.007 (0.001) �8.0 �6.4 (0.2) �1.6CaM P2-XPC � 0.19 (0.02) �10.1 �10.6 (0.1) �0.5CaM P2-XPC � NBa � � �
a NB, no binding observed in the present conditions.
XPC/Human Centrin 2 Interaction40256
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from
(34) or NMR (10, 11) experiments. The results obtained in thepresent work, showing a Ca2�-dependent strong interactionwith an amphiphilic peptide derived from the XPC protein,provide additional evidence for the Ca2� sensor behavior ofHsCen2.
The first experimental data on potential centrin targets wereobtained from genetic (35) and biochemical (36) experiments inyeast, suggesting that Cdc31 may physically interact with theprotein Kar1, the two proteins being localized in the spindlepole body. Indeed, a 19-residue peptide derived from the Kar1sequence was shown to bind, in a Ca2�-dependent manner anda stoichiometry close to 1:1, to centrins of various origins(Scherffelia dubia, Chlamydomonas, yeast, human) (11, 37),but not to yeast CaM (37). The reported binding constant ofKar1 (Fig. 1A) peptide to HsCen2 (�3 � 106 M�1) (37) is �2
orders of magnitude lower than for the peptide P1-XPC studiedin the present work. It was suggested that the acidic charges inthe binding sequence of Fig. 1A, mainly those in the N-terminalsite, may explain the decreased affinity for both centrinand CaM.
The affinity of P1-XPC for HsCen2 is among the highestobserved for CaM and its target proteins or peptides, usuallyexhibiting binding constants between 106 and 1011 M�1 (15, 19,38–40). The Ca2� dependence of P1-XPC binding to SC-HsCen2 and HsCen2 is reflected in the decrease of the bindingconstant upon removal of the metal ions by a factor of 17 and28, respectively. In the case of CaM/target complexes, the Ca2�
dependence varies from very strong (smooth muscle myosinlight chain kinase (Ref. 18) or NOS-I (Ref. 40)) to moderate orvery weak (38, 41).
FIG. 3. Titration of CaM and HsCen2 by P2-XPC. Figure shows ITC experiments on the binding of P2-XPC to CaM and HsCen2. Note thedifference in scale of the heat rate between the first panel and the next two panels.
XPC/Human Centrin 2 Interaction 40257
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from
Our results strongly suggest that binding of P1-XPC toHsCen2 is mainly determined by the C-terminal domain of theprotein, which contributes more than 95% of the total bindingenergy. The lack of (or a very small) contribution from theN-terminal half may be related to its low Ca2� affinity and toits presumed closed tertiary conformation. Indeed, there isexperimental evidence that apo domains of EF-hand proteinsexpose only a small hydrophobic surface, being less favorable toan efficient target binding (42). The relative roles of the twolobes in HsCen2 are in agreement with the structure-functionrelationships established for the yeast centrin (43), showingthat mutations in the central and C-terminal parts of theCdc31 sequence are associated with a defective protein local-ization and G2/M arrest, whereas the N-terminal modificationsinduce no observable defects. Nevertheless, binding of P1-XPCto HsCen2 appears to affect also the N-terminal domain, assuggested by the significant thermal stability increase (20 °C)of the N-terminal domain. This observation is corroborated bythe calorimetric measurements, showing that the heat capacityof binding is almost 3 times greater for the integral proteinrelative to its C-terminal domain. Although not essentially
involved in the recognition and initial binding of the peptide,the N-terminal domain may move closer to the C-terminaldomain, hiding a significant amount of the solvent-exposedsurface. Nevertheless, understanding of the detailed role of theN-terminal domain requires further investigation.
HsCen2 binds one P1-XPC per molecule, as is the case of themajority of CaM/target peptide complexes (19, 38). In contrast,the preponderant binding role of the C-terminal half in HsCen2is less usual (44) and suggests that the complex conformation isdifferent from that described for MLCK, calmodulin kinase I, orcalmodulin kinase kinase peptides bound to CaM (32), whereboth domains participate in wrapping the peptide. On the otherhand, CaM appears to bind two copies of P1-XPC with affinitiesdiffering by 2 orders of magnitude, leading to the hypothesis ofa distinct complex conformation. In this sense, it is worthmentioning that recent crystallographic structures of CaM incomplex with larger targets revealed new types of interactions,where each CaM lobe binds to a distinct target fragment(45, 46).
The free energy of the HsCen2/P1-XPC complex formation isdominated by the negative enthalpy component, suggesting
FIG. 4. Titration of CaM by P1-XPC. Interaction of CaM with P1-XPC in the absence of Ca2� and at three different peptide/protein ratios(noted R), studied by ITC experiments. The thermogram and the interaction isotherm obtained in the absence of Ca2� are also shown.
XPC/Human Centrin 2 Interaction40258
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from
that the van der Waals, hydrogen bonding, and electrostaticinteractions play the central role in peptide recognition. Theunfavorable contribution of the entropy (�S � 0) to the complexformation is the result of at least two main components. Thefirst is related to the peptide change from a random coil con-
formation in the free state to an �-helix secondary structureupon binding (Fig. 5) that corresponds to a decrease of thedegree of freedom of its movements. The second componentaccounts for the decrease in structural disorder of the protein,including both the backbone and the side-chain dynamics, in
FIG. 5. CD analysis. Far-UV CD spectra of P1-XPC (30 �M), HsCen2 (30 �M) and SC-HsCen2 (32 �M) in 10 mM Tris buffer, pH 6.70, 100 mM
NaCl, 2 mM CaCl2 at 22 °C. The effect of peptide addition to the SC-HsCen2 (1:1 ratio) is shown in panel A and that of peptide addition to theintegral protein (1:1 and 2:1 peptide-to-protein ratio in red and magenta, respectively) is shown in panel B. The effect of peptide binding uponthermal denaturation curves are shown in panels C and D for SC-HsCen2 and HsCen2, respectively. Denaturation was observed as the variationof the CD signal at 222 nm as a function of temperature.
FIG. 6. NMR experiments. [15N-1H]HSQC spectra at 500 MHz of SC-HsCen2 and its equimolar complexes with melittin (MEL) and P1-XPC.Samples (1–1.5 mM) are in 20 mM Tris (d11) buffer, pH 6.5, 100 mM NaCl, 10 mM CaCl2, and the spectra were recorded at 308 K. Horizontal linesconnect the peak pairs belonging to the same amino groups from Asn and Gln side chains.
XPC/Human Centrin 2 Interaction 40259
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from

agreement with the enhanced thermal stability of the com-plexes, observed in CD experiments (Fig. 5). In the presentcase, these two negative entropy changes are larger than theusually large entropy increase caused by the partial dehydra-tion of the interacting surfaces and the associated enhancedmovement freedom of water molecules. It is expected that, inreal interacting systems, including HsCen2 and XPC, wherethe target binding site is already �-helical, an important neg-ative component is canceled and the entropy contribution maybecome more favorable for the interaction.
Nevertheless, this thermodynamic analysis is incomplete,and its predictive capacity should be taken with some caution.The published data show indeed that peptide binding to CaM orapoCaM can be both enthalpically and entropically driven pro-cesses (18). Although the majority of the studied peptides in-teract exothermically, melittin or fragments from cNOS andphosphodiesterase bind to CaM with a positive enthalpy (19,47), driven by an entropic factor. The variability in energeticbalance may also depend on the Ca2�-binding protein. Forinstance, in the case of melittin, our calorimetric experimentsrevealed a negative enthalpy change upon binding to HsCen2,in opposition with the endothermic character of the binding toCaM (19). This observation points to a distinct behavior be-tween HsCen2 and CaM in their target recognition capacitythat may be physiologically relevant for the discriminationbetween the specific signaling pathways of the two proteins.
Promiscuity in Intermolecular Interactions—The simplifyingconception of single targets for each signaling molecule and ofa high selectivity in molecular recognition must be adapted tothe increasing evidence for multiple and interfering interac-tions between proteins in a cellular network. CaM is distrib-uted throughout the cytoplasm and nucleus of mammalian cellsbut is particularly concentrated in the centrosome and thesubmembrane region around the cell (24), showing a significantparallelism with the centrin distribution. The presence of CaMwithin or close to the microtubule organizing centers fromyeasts (25) to humans (24, 26) is well documented, and its corre-lation with the key steps of the cell division was demonstrated invarious organisms (26, 48, 49). In addition, an increasing numberof centrosomal proteins were found to possess CaM-bindingmotifs (27, 28), suggesting that their participation in the con-trol of microtubule organization could be modulated by theinteraction with proteins from the CaM superfamily.
Site-directed mutagenesis studies with CaM or Cdc31 haveshown that different structural sites or particular residues playa distinct role in diverse essential functions (43, 51, 52). Theorigin of this diversity should reside in the existence of a rangeof molecular targets recognized and activated (inhibited) byEF-hand proteins. The coexistence of CaM and HsCen2 in thesame tissues, the multiplicity of centrin isoforms, and the ca-pacity to bind common peptides raise the question of the mech-anisms capable to avoid the promiscuous interactions duringthe cell regulatory processes.
The selection of the appropriate signaling pathway involvingone of the two proteins may depend on several modulatingfactors: (a) the abundance of each protein in a given tissue orcellular compartment, as well as the time-dependent expres-sion during differentiation and development (4, 5, 53); (b) thetime and space characteristics of the Ca2� signal (54); (c)the molecular properties of the EF-hand proteins, including thenumber of active binding sites, equilibrium and kinetic ionbinding properties, cooperativity, and Ca2�/Mg2� discrimina-tion; (d) the remarkable flexibility and conformational versa-tility (42, 45, 55–60) as a response to ion binding or targetcomplexation; and (e) the site-specific phosphorylation, its en-zymatic control, and temporal progress (61, 62).
Implications for the NER Process—Although the centrinswere initially identified in the microtubule organizing centers,the centrosomal fraction of human centrins represents lessthan 10% of the total cellular pool, a significant amount beinglocalized in the nucleus (7). No functional information aboutthis last fraction was available until the recent discovery (8)that HsCen2 could associate with the XPC/HR23B complexthat intervenes in the first step of the genome-wide NER proc-ess. HsCen2 did not show any measurable influence on theefficiency of the XPC-dependent NER activity in vitro, butseems to increase the thermal stability at higher temperatures(40 °C) and assembly efficiency of the XPC complex. Ourpresent results reinforce the previous observations by the iden-tification of a short XPC fragment that is able to bind stronglyboth to HsCen2 and CaM. This binding site is situated withinthe C-terminal end (constituting 10% of the sequence) of thenuclear protein, showing a high sequence conservation amongvarious homologues from yeast to humans. In particular theTrp residue, placed in the N-terminal part of the binding site,but also the other residues forming the consensus hydrophobicand basic pattern, are highly conserved (Fig. 1C). This obser-vation supports the biological relevance to the P1-XPC se-quence tested in the present work and suggests that the modeof interaction HsCen2/XPC in human cells could be extended tothe corresponding molecular pairs in other eukaryotic cells. Forinstance, Rad4, the S. cerevisiae counterpart of XPC, may in-teract with Cdc31 protein, which plays the role of the humancentrin in yeast.
The physicochemical and structural properties of XPC pro-tein are presently not known, precluding a coherent molecularexplanation of its functional mechanism. However, recent ex-perimental results (63) suggested that the last 124 residues,including the P1-XPC fragment studied here, are criticallyinvolved in the recruitment of the large transcription factorcomplex TFIIH (Fig. 1A) that is necessary to unwind the dam-aged DNA double helix. The functional importance of this C-terminal domain is further supported by the analysis of theXPC gene in xeroderma pigmentosum patients (64). In themajority of studied cases, presenting a rather homogeneousphenotype, the identified causative mutations produce a trun-cated protein of various lengths, but always lacking the TFIIHbinding region. These observations emphasize the critical func-tional role of the P1-containing structural domain of XPC andsuggest a possible interference in the binding of HsCen2 andthe transcription factor to the same structural domain.
By its implication in the nuclear DNA repair process, centrinmay constitute a functional link between the chromosome du-plication and the centrosome control of the mitotic spindles.Existence of this type of correlation was previously suggestedby genetic studies in yeast showing that deletion of Rad23 andDSK2 (very similar to Rad23) provokes a lethal phenotypeproduced by an inefficient spindle pole body duplication (65).
Elucidation of the HsCen2 role in the NER system and theeventual correlation with its centrosomal function need furtherinvestigation. Utilization of peptides as models for the protein/protein interactions has obvious limitations: other fragments ofthe target protein may participate to the contact interface orthe access binding site could be hindered in the native three-dimensional structure of the integral target. In addition, themechanism of the physiological response consecutive to theinteraction could hardly be revealed by using the simplifiedmodel (50). Expression of a larger XPC construct, includingthe P1-XPC peptide, and the investigation of its physicochem-ical, biochemical, and functional properties are currently inprogress in our laboratory.
XPC/Human Centrin 2 Interaction40260
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from

REFERENCES
1. Schiebel, E., and Bornens, M. (1995) Trends Cell Biol. 5, 197–2012. Salisbury, J. L. (1995) Curr. Opin. Cell Biol. 7, 39–453. Errabolu, R., Sanders, M. A., and Salisbury, J. L. (1994) J. Cell Sci. 107, 9–164. Lee, V. D., and Huang, B. (1993) Proc. Natl. Acad. Sci. U. S. A. 90,
11039–110435. Middendorp, S., Paoletti, A., Schiebel, E., and Bornens, M. (1997) Proc. Natl.
Acad. Sci. U. S. A. 94, 9141–91466. Salisbury, J. L., Suino, K. M., Busby, R., and Springett, M. (2002) Curr. Biol.
12, 1287–12927. Paoletti, A., Moudjou, M., Paintrand, M., Salisbury, J. L., and Bornens, M.
(1996) J. Cell Sci. 109, 3089–31028. Araki, M., Masutani, C., Takemura, M., Uchida, A., Sugasawa, K., Kondoh, J.,
Ohkuma, Y., and Hanaoka, F. (2001) J. Biol. Chem. 276, 18665–186729. Benhamou, S., and Sarasin, A. (2000) Mutat. Res. 462, 149–158
10. Matei, E., Miron, S., Blouquit, Y., Duchambon, P., Durussel, I., Cox, J. A., andCraescu, C. T. (2003) Biochemistry 42, 1439–1450
11. Veeraraghavan, S., Fagan, P. A., Hu, H., Lee, V., Harper, J. F., Bessie, H., andChazin, W. J. (2002) J. Biol. Chem. 277, 28564–28571
12. Durussel, I., Blouquit, Y., Middendorp, S., Craescu, C. T., and Cox, J. A. (2000)FEBS Lett. 472, 208–212
13. Wuthrich, K. (1986) NMR of Proteins and Nucleic Acids, John Wiley & Sons,New York
14. Cavanagh, J., Fairbrother, W. J., Palmer, A. G., III, and Skelton, N. J. (1996)Protein NMR Spectroscopy: Principles and Practice, Academic Press, SanDiego
15. O’Neil, K. T., and DeGrado, W. F. (1990) Trends Biochem. Sci. 15, 59–6416. Rhoads, A. R., and Friedberg, F. (1997) FASEB J. 11, 331–34017. Yap, K. L., Kim, J., Truong, K., Sherman, M., Youan, T., and Ikura, M. (2001)
J. Struct. Funct. Genomics 1, 8–1418. Wintrode, P. L., and Privalov, P. L. (1997) J. Mol. Biol. 266, 1050–106219. Brokx, R. D., Lopez, M. M., Vogel, H. J., and Makhatadze, G. I. (2001) J. Biol.
Chem. 276, 14083–1409120. Baker, B. M., and Murphy, K. P. (1997) J. Mol. Biol. 268, 557–56921. Luque, I., Todd, M. J., Gomez, J., Semo, N., and Ernesto, F. (1998) Biochem-
istry 37, 5791–579722. Makhatadze, G. I., and Privalov, P. L. (1995) Adv. Protein Chem. 47, 307–42523. Spolar, R. S., and Record, M. T., Jr. (1994) Science 263, 777–78424. Li, C.-J., Heim, R., Lu, P., Pu, Y., Tsien, R. Y., and Chang, D. C. (1999) J. Cell
Sci. 112, 1567–157725. Moser, M. J., Flory, M. R., and Davis, T. N. (1997) J. Cell Sci. 110, 1805–181226. Moisoi, N., Erent, M., Whyte, S., Martin, S., and Bayley, P. M. (2002) J. Cell
Sci. 115, 2367–237927. Flory, M. R., Moser, M. J., Monnat, R. J., Jr., and Davis, T. N. (2000) Proc.
Natl. Acad. Sci. U. S. A. 97, 5919–592328. Matsumoto, Y., and Maller, J. L. (2002) Science 295, 499–50229. Johnson, W. C. (1999) Proteins Struct. Funct. Genet. 35, 307–31230. Li, M. X., Gagne, S. M., Tsuda, S., Kay, C. M., Smillie, L. B., and Sykes, B. D.
(1995) Biochemistry 34, 8330–834031. Gagne, S. M., Tsuda, S., Li, M. X., Chandra, M., Smillie, L. B., and Sykes, B. D.
(1994) Protein Sci. 3, 1961–197432. Hoeflich, K. P., and Ikura, M. (2002) Cell 108, 739–74233. Tsalkova, T. N., and Privalov, P. L. (1985) J. Mol. Biol. 181, 533–544
34. Wiech, H., Geier, B. M., Paschke, T., Spang, A., Grein, K., Steinkotter, J.,Melkonian, M., and Schiebel, E. (1996) J. Biol. Chem. 271, 22453–22461
35. Vallen, E., Ho, W., Winey, M., and Rose, M. D. (1994) Genetics 137, 407–42236. Spang, A., Courtney, I., Grein, K., Matzner, M., and Schiebel, E. (1995) J. Cell
Biol. 128, 863–87737. Geier, B. M., Wiech, H., and Schiebel, E. (1996) J. Biol. Chem. 271,
28366–2837438. Crivici, A., and Ikura, M. (1995) Annu. Rev. Biophys. Biomol. Struct. 24,
85–11639. Murase, T., and Iio, T. (2002) Biochemistry 41, 1618–162940. Censarek, P., Beyermann, M., and Koch, K.-W. (2002) Biochemistry 41,
8598–860441. Bahler, M., and Rhoads, A. (2002) FEBS Lett. 513, 107–11342. Ikura, M. (1996) Trends Biochem. Sci. 21, 14–1743. Ivanovska, I., and Rose, M. D. (2001) Genetics 157, 503–51844. Elshorst, B., Hennig, M., Forsterling, H., Diener, A., Maurer, M., Schulte, P.,
Schwalbe, H., Griesinger, C., Krebs, J., Schmid, H., Vorherr, T., andCarafoli, E. (1999) Biochemistry 38, 12320–12332
45. Schumacher, M. A., Rivard, A. F., Bachinger, H., Peter, and Adelman, J. P.(2001) Nature 410, 1120–1124
46. Drum, C. L., Yan, S.-Z., Bard, J., Shen, Y.-Q., Lu, D., Soelaiman, S., Grabarek,Z., Bohm, A., and Tang, W.-J. (2002) Nature 415, 396–402
47. Moorty, A., K., Gopal, B., Satish, P. R., Bhattacharya, S., Bhattacharya, A.,Murthy, M. R. N., and Surolia, A. (1999) FEBS Lett. 461, 19–24
48. Rasmussen, C., and Means, A. (1989) EMBO J. 8, 73–8249. Sun, G., Hirata, A., Ohya, Y., and Anraku, Y. (1992) J. Cell Biol. 119,
1625–163950. Chin, D., and Means, A. R. (2000) Trends Cell Biol. 10, 322–32851. Kink, J., Maley, M., Preston, R., Ling, K., Wallen-Friedman, M., Saimi, Y., and
Kung, C. (1990) Cell 62, 165–17452. Ohya, Y., and Botstein, D. (1994) Science 263, 963–96653. Wolfrum, U., and Salisbury, J. L. (1998) Exp. Cell Res. 242, 10–1754. Berridge, M. J., Lipp, P., and Bootman, M. D. (2000) Nat. Rev. Mol. Cell. Biol.
1, 11–2155. Meador, W. E., George, S. E., Means, A. R., and Quiocho, F. A. (1995) Nat.
Struct. Biol. 2, 943–94556. Meador, W. E., Means, A. R., and Quiocho, F. A. (1993) Science 262, 1718–172157. Osawa, M., Swindells, M. B., Tanikawa, J., Tanaka, T., Mase, T., Furuya, T.,
and Ikura, M. (1998) J. Mol. Biol. 276, 165–17658. Wang, E., Zhuang, S., Kordowska, J., Grabarek, Z., and Wang, C.-L. A. (1997)
Biochemistry 36, 15026–1503459. Osawa, M., Tokumitsu, H., Swindells, M. B., Kurihara, H., Orita, M., Shi-
banuma, T., Furuya, T., and Ikura, M. (1999) Nat. Struct. Biol. 6, 819–82460. Rustandi, R. R., Baldisseri, D. M., and Weber, D. J. (2000) Nat. Struct. Biol. 7,
570–57461. Nigg, E. A. (2001) Nat. Rev. Mol. Cell. Biol. 2, 21–3262. Lutz, W. H., Lingle, W. L., McCormick, D., Greenwood, T. M., and Salisbury,
J. L. (2001) J. Biol. Chem. 276, 20774–2078063. Uchida, A., Sugasawa, K., Masutani, C., Dohmae, N., Araki, M., Yokoi, M.,
Ohkuma, Y., and Hanaoka, F. (2002) DNA Repair 1, 449–46164. Chavanne, F., Broughton, B. C., Pietra, D., Nardo, T., Browitt, A., Lehmann,
A. R., and Stefanini, M. (2001) Cancer Res. 60, 1974–198265. Biggins, S., Ivanovska, I., and Rose, M. D. (1996) J. Cell Biol. 133, 1331–1346
XPC/Human Centrin 2 Interaction 40261
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from

Constantin T. CraescuAurel Popescu, Simona Miron, Yves Blouquit, Patricia Duchambon, Petya Christova and
Human Centrin 2 and CalmodulinXeroderma Pigmentosum Group C Protein Possesses a High Affinity Binding Site to
doi: 10.1074/jbc.M302546200 originally published online July 30, 20032003, 278:40252-40261.J. Biol. Chem.
10.1074/jbc.M302546200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/278/41/40252.full.html#ref-list-1
This article cites 63 references, 24 of which can be accessed free at
by guest on June 4, 2016http://w
ww
.jbc.org/D
ownloaded from
















