Embed Size (px)
Citation preview

ҚАЗАҚ ҰЛТТЫҚ АГРАРЛЫҚ УНИВЕРСИТЕТІ
ӘОЖ 631.4/8+576.8 Қолжазба құқығында
СЕЙТКАЛИ НУРЗИХАН
Жетісу және Іле Алатауларының жабайы жемісті орман
топырақтарындағы энтомопатогенді Bacillus thuringiensis бактерия
штаммдарының қабыршақ қанатты зиянкестерге тиімділігі
6D080800- Топырақтану және агрохимия
Ph.D докторы акедемиялық дәрежесіне ізденуге
арналған диссертация
Ғылыми жетекшілері:
а.ш.ғ.к. доцент Кубенкулов.Қ
б.ғ.д., профессор ҚР ҰҒА академигі Сагитов А.О
ҚХР, Шинжаң ауылшаруашылығы университетінің
доценті, Еркін Әлімбекұлы
Қазақстан Республикасы
Алматы, 2015

2
МАЗМҦНЫ
НОРМАТИВТІК СІЛТЕМЕЛЕР 4
АНЫҚТАМАЛАР 5
БЕЛГІЛЕУЛЕР МЕН ҚЫСҚАРТУЛАР 6
КІРІСПЕ 7
1 ӘДЕБИЕТКЕ ШОЛУ 10
1.1 Энтомопатогенді Bacillus thuringiensis бактериялары бӛлініп
алынған топырақ аймақтарына шолу
10
1.2 Топырақ жәндіктері мен микроорганизмдері 14
1.3 Кристал түзуші бактерияларды зерттеудің тарихы 18
1.4 Энтомопатогенді бактериялар негізінде алынған биопрепараттар 21
1.5 Bacillus thuringiensis бактерияларымен жасалынып
шығарылатын токсиндер
23
1.6 Қазақстандағы зиянды жәндіктерге қарсы микробиологиялық
әдіс бойынша Bt қолдана отырып ғылыми зерттеулер жүргізу
28
2 ЗЕРТТЕУ МАТЕРИАЛДАРЫ МЕН ТӘСІЛДЕРІ 30
2.1 Зерттеу нысандары 30
2.2 Далалық зерттеу жүргізу әдістемесі 31
2.2.1 Зерттеулер жүргізілген аймақтар топырақтарының морфо-
гентикалық сипаттары, құрамдары мен қасиеттерін анықтау
31
2.3 Зертханалық тәжірибе жүргізу әдістемесі 33
2.3.1 Зерттеулер жүргізілген аймақтардың топырақтарының жалпы
микрофлорасын зерттеу
33
2.3.2 Энтомопатогенді микроорганизмдерді топырақтан және
бунақденелілерден бӛліп алу
39
2.3.3 Bacillus thuringiensis энтомопатогенді бактерияларының
дақылдық-морфологиялық, биохимиялық қасиеттерін анықтау
40
2.3.4 Bacillus thuringiensis бактерия штаммдарының биологиялық
белсенділігін анықтау
44
3 ЗЕРТТЕУ ЖҤРГІЗІЛЕТІН АЙМАҚТАРДЫҢ ТАБИҒИ-
КЛИМАТ ЖАҒДАЙЛАРЫ
46
3.1 Климат 46
3.2 Жер бедері және топыраққұраушы жыныстар 50
3.3 Ӛсімдіктері 54
4 ЗЕРТТЕУ НӘТИЖЕЛЕРІ 55
4.1 Bacillus thuringiensis бактериялары қалыптасқан Іле Алатауы
топырақтарының морфо-гентикалық сипаты және құрамдары
мен қасиеттері
55
4.2 Bacillus thuringiensis бактериялары қалыптасқан Жетісу Алатауы
топырақтарының морфо-генетикалық сипаттары және
құрамдарымен қасиеттері
62

3
4.3 Жетісу және Іле Алатауларының орманның күңгірт-сұр
топырақтарының микроорганизмдерінің сандық және сапалық
кӛрсеткіштері мен температура ӛзгерісіне байланыстылығы
66
4.4 Жетісу және Іле Алатауларының жабайы жемісті орман
топырақтарымен бунақденелілерден бӛлініп алынған,
энтомопатогенді Bacillus thuringiensis бактериялары
71
4.5 Бӛлініп алынған дақылдар уыттылығының әр-түрлі
бунақденелілерге әсері
90
4.5.1 Bacillus thuringiensis бактерия штаммдарының жапырақ
ширатқыш кӛбелегінің жұлдызқұрттарына қарсы биологиялық
белсенділіктертері
90
4.5.2 Bacillus thuringiensis бактерия штаммдарының алма күйе
кӛбелегінің жұлдызқұрттарына биологиялық белсенділіктертері
92
4.5.3 Бунақденелілерден бӛлініп алынған Bacillus thuringiensis
энтомопатогенді бактерия штаммдарының биологиялық
белсенділіктертері
94
4.5.4. Bacillus thuringiensis бактерия штаммдары негізіндегі дақылдық
сұйықтықтарды дайындау және олардың биологиялық
белсенділіктері
98
5 АЛМА КҤЙЕ КӚБЕЛЕГІНЕ ЭНТОМОПОТОГЕНДІ
БАКТЕРИЯЛАР НЕГІЗІНДЕ ЖАСАЛҒАН ЖАРТЫЛАЙ
СҦЙЫҚ ПРЕПАРАТТЫҚ ҤЛГІЛЕРДІҢ
ЭКОНОМИКАЛЫҚ ТИІМДІЛІГІ
99
ҚОРЫТЫНДЫ 101
ПРАКТИКАЛЫҚ ҦСЫНЫСТАР 103
ПАЙДАЛАНЫЛҒАН ӘДЕБИЕТТЕР ТІЗІМІ 104

4
НОРМАТИВТІК СІЛТЕМЕЛЕР
Диссертациялық жұмыста келесі нормативтік құжаттарға сілтемелер
қолданылған:
ҚРСТ 2.11-68 құрлымдаушы құжаттардың біріңғай жүйесі. ережелерді бақылау.
ҚРСТ 2.105–95 құрлымдаушы құжаттардың біріңғай жүйесі. мәтінді құжаттарға
жалпы талаптар.
ҚРСТ 6.38–90 құжаттамаларды біріңғайлау жүйесі. құжаттамаларды
ұйымдастырушы-бӛлу жүйесі. құжаттарды ресімдеу талаптары.
ҚРСТ 7.32–2001 ғылыми-зерттеу жұмыстар туралы есеп. құрлымдаушы және
ресімдеу ережелері.
ҚРСТ 7.1–2003 библиографиялық жазба. библиографиялық сипаттау. Жалпы
талаптар мен құрастыру ережелері.
ҚРСТ 27593–88 топырақтар. терминдер және анықтаулар.
ҚРСТ 28168–89 топырақтар. үлгілерді алу.
ҚРСТ 26213–91топырақтағы гумус мӛлшерін И.В. Тюрин әдісімен анықтау.
ҚРСТ 2610 –84 топырақтағы жалпы азотты-И.Г.Кьельдал әдісімен анықтау.
ҚРСТ 26264–84 топырақтағы жалпы фосфор мен калийді анықтау.
ҚРСТ 26205-84 топырақтағы жеңіл гидролизденетін азотты-Тюрин-Кононова
әдісімен, жылжымалы фосфорды-Мачингин әдісі, жылжымалы калийді-
жалынды фотометрде анықтау.
ҚРСТ 26423-85, 26428-85, 26425-85, 26426-85, 26424-85, 25428-85 топырақтың
су сүзіндісінің иондық құрамын анықтау.
ҚРСТ 26425-85 топырақтың pH кӛрсеткішін анықтау.
ҚРСТ 17.4.4.01-84 топырақтағы катиондар сиымдылығын-Бобко-Аскинази-
Алешин әдісімен анықтау.

5
АНЫҚТАМАЛАР
Бұл диссертациялық жұмыста келесі анықтамаларға сәйкес сілтемелер
кӛрсетілген:
Топырақ – тау жынысы, организмдер, климат, жер бедері және уақыттың
кешенді әрекеттесуінен түзілген күрделі кӛп функционалды және кӛп құрамды,
кӛп фазалы құнарлық қабілеті бар ашық жүйелі құрылым.
Топырақ кескіні - бір-бірін ауыстырып отыратын генетикалық қабаттар
жиынтығы.
Топырақ ҥлгілері – топырақ қабатынан белгілі бір мӛлшерде алынған
топырақ.
Гумус – құрамында жоғарғы ӛсімдіктерге қажетті қоректік заттары бар
топырақтың негізгі ерекше органикалық бӛлігі.
Су сҥзіндісі – топырақты дистилденген сумен шайқағаннан кейін сүзіліп
алынған ерітінді.
Топырақ генезисі – топырақтардың және оларға тән ерекшеліктердің
(құрылым, құрам, қасиет, құбылымдар) жаратылуы, түзілуі және дамуы.
Бактериялар (гр.bakterion-таяқша) — тек микроскопта ғана кӛрінетін аса
ұсақ бір жасушалы ағза.
Микроорганизм –тек қана микроскоппен кӛруге болатын ӛте ұсақ
организмдер.
Биопрепараттар – адамдар, жануарлар және ӛсімдіктердің түрлі
ауруларын анықтауда, оларды емдеу және алдын ала сақтандыру мақсатында
қолданылатын ерекше биологиялық қосылыстар.
Токсин – кейбір микроорганизмдер, ӛсімдіктер мен жануарлар түзетін улы
заттар. Химиялық табиғаты бойынша полипептидтер мен ақуыздарға жатады.
Патоген – жұқпалы аурулар қоздырғыш.
Вируленттілік - патогенділік кӛрсеткіші.
Серотип – бактериялардың бір түрдің тұр тармағының бір-бірінен
антигендік құрылымдары бойынша айырылатын түрлері.

6
БЕЛГІЛЕУЛЕР МЕН ҚЫСҚАРТУЛАР
ҚРСТ – Қазақстан республикасының стандарты
КС – кӛлемдік салмақ
ТСК – топырақтың сіңіру кешені
ЕТС – ең тӛменгі сусиымдылық
Гқ – гумин қышқылы
Фқ – фульво қышқылы
C:N – кӛміртегі мен азоттың қатынасы
HCO–
3 – бикарбонат
CO32-
– карбонат
Cl- – хлор
SO42-
– сульфат
Са2+
– калций
Mg2+
– магний
Na+
– натрий
K+
– калий
C – кӛміртегі
N – азот
P – фосфор 0C – цельци градус (ауа температурасын ӛлшейтін шама)
% – пайыз
мг–экв–миллиграмм эквивалент
см – сантиметр
B. – Bacillus
Bt – Bacillus thuringiensis
ЕПА – ет пептонды агар
КА – картопты агар
«А» – Балық гидролизаты қоректік ортасы энтомопатогенді
микроорганизмдерді ӛсіруде қолайлы орта
МҰТС – мемлекеттік ұлттық табиғи саябақ
ҚазӚҚ және КҒЗИ – Қазақ ӛсімдік қорғау және карантин ғылыми зерттеу
институты

7
КІРІСПЕ
Тақырыптың ӛзектілігі. Топырақ табиғатта микроорганизмдердің
тіршілік етуінің шынайы ортасы. Олар топырақтың түзілу және ӛзіндік тазару
процесіне белсенді қатысып, табиғатта заттар алмасудағы орны орасан зор.
Топырақтағы микроорганизмдердің белсенділігі ӛте алуан түрлі. Олардың
қатысуымен ӛсімдік және жануар текті органикалық заттардың гумификациясы
мен минералдануы жүреді, органикалық және минералды қосылыстардың
тотығу және қалпына келуі, атмосфералық азоттың фиксациясы, нитрификация
және денитрификсация үрдісі, минералдардың ыдырауы және түзілуі жүреді.
Топырақ микроорганизмдерінің мүмкіндігін зерттеу ӛзекті мәселе болып
табылады. Топырақтан бӛлініп алынған Bacillus thuringiensis энтомопатогенді
бактерияларының қабыршаққанатты зиянкестерге тиімділігін анықтау ӛте
маңызды болып табылады.
ҚР Экологиялық қауіпсіздігі концепциясында «биоалуантүрлілікке нақты
қауіп тӛнуінің негізгі мәселесі ормандарды жою, топырақ эрозиясы, ӛсімдік пен
жануарлар түрлерін шектен тыс пайдалану болып табылады» деп кӛрсетілген.
Дегенмен, биоалуантүрліліктің сақталуы жӛнінде сӛз қозғағанда,
бунақденелілер мен микроорганизмдер де орман биоценозының қажетті
бӛлшегі (компоненті) және белгілі бір жағдайларда олардың кӛп шығын
келтіруі мүмкін екендігін ұмытпаған жӛн. Мысал ретінде 2000 жылдардың
басында Жоңғар Алатауында алманың күйе кӛбелегінің жаппай кӛбеюін айтуға
болады. Олар алма ағаштарына үлкен зиян келтіріп, нәтижесінде бүгінгі таңда
ағаштардың 20-25 % ӛлген немесе ӛте әлсіреген жағдайда. Зиянкестермен
күресу жұмыстары (әуеден бүрку арқылы) Қазақ ӛсімдік қорғау және карантин
ҒЗИ-ның ғылыми әдістемелік жетекшілігімен 2003-2005 жж жүргізілді. Бұл
жұмысқа мемлекеттен 168 млн теңге бюджет қаражаты жұмсалды. Институт
қызметкерлерінің бақылаулары бойынша 2010-2011 жж. алма күйе кӛбелегі Іле
Алатауында жаппай кӛбейіп, алма ағаштарын үлкен апатқа ұшыратқан.
Іле және Жетісу Алатаулары мәдени алманың дүниежүзілік шыққан
орталығы екендігін, ал біздің жергілікті жабайы алмамыз ғаламшардағы барлық
мәдени алма сорттарының арғы атасы екендігін әлемдік қауымдастықтар
әлдеқашан мойындаған. Біздің парызымыз, осы керемет табиғи феномендерді
антропогенді күйзелістерден және зиянды организмдерден қорғау. Осыған
байланысты, Іле және Жетісу Алатаулары топырағының биоалуантүрлілігін
зерттеу аса ӛзекті мәселе болып табылады.
Агроэкожүйенің фитосанитарлық жағдайын қалыптастыруда пайдалы
және зиянды фауна популяциялары мӛлшерінің динамикасын мақсатты бағытта
реттеудің оларды жобалаудағы маңызы зор. Зиянкестердің сан мӛлшерін
шектейтін, талшықты заттарды қолданумен байланысы жоқ шаралардың
әртүрлі топтары бар. Зиянды бунақденелердің сан мӛлшерін биологиялық
бақылау олардың ӛлуі мен мӛлшерінің азаюы және пайдалы түрлердің
тіршілігіне қолайлы жағдай тудыру мақсатында тірі организмдерді немесе
олардың тіршілік ӛнімдерін қолдануды қарастырады. Химиялық пестицидтерді

8
қолданудың кӛпжылдық тәжірибе нәтижелерінде олардың кӛпшілігінің кең
кӛлемде токсикологиялық кері әсері бар екендігі анықталды. Пестицидтер
қоршаған ортада жинақталуы және айналуы, табиғи биоценозды бұзуы,
жануарлардың пайдалы түрлері мен адам денсаулығына зиян келтіруі мүмкін.
ЮНЕСКО мәліметтері бойынша пестицидтер мен жер биосферасының
жалпы кӛлемде ластануы мұнай ӛнімдері, белсенді заттар, фосфаттар,
минералды тыңайтқыштар, ауыр металлдар, азот, күкірт, кӛмірқышқылы және
басқа заттар қосындылары секілді заттардан кейін 8-9 орынды алып отыр. Ал
пестицидтердің дүниежүзілік қолданылуы қазіргі уақытта шамамен жылына 3
млн. тонна.
Осыларға байланысты қазіргі кезеңде зиянды бунақденелердің сан
мӛлшерін биологиялық (микробиологиялық) әдістермен реттеу ӛсімдік қорғау
стратегиясының құнды негізі болып отыр. Ӛсімдік қорғаудың бұл тәсілдерінің
экономикалық және экологиялық ерекшеліктері, қоршаған ортаның ластану
деңгейінің жоғарылығын ескере отырып, ауыл шаруашылығы ӛндірісінде
кеңінен және белсенді түрде енгізу қажет екендігі айқындалып отыр.
Ғылыми зерттеудің қажеттілігі
1) Жетісу және Іле Алатауларының жабайы жемісті ормандарындағы
әлемдегі барлық мәдени алма сорттарының генетикалық негізі болып
саналынатын Сиверс жабайы алмасының генетикалық ресурстарын сақтау.
Себебі, бұл біздің ұлттық мақтанышымыз және байлығымыз, әрі ол бүкіл
адамзаттың мақтанышы болып табылады.
2) Соңғы он жылдықтарда оларда зиянды бунақденелілердің сан
мӛлшерінің күрт артуы, бұрынғы химиялық күресу шараларының орнына
биологиялық күрес шараларын жүргізудің қажеттілігі.
Зерттеу мақсаты мен міндеттері
Зерттеу мақсаты – Жетісу және Іле Алатауларының жабайы жемісті орман
топырақтарынан және бактериоздық белгілерімен ӛлген бунақденлілерден
энтомопатогенді Bacillus thuringiensis бактерияларын бӛліп алу және жабайы
жемісті орман алқаптарының топырақтарының экологиялық жағдайын бағалау
және де осы топырақтардан бӛлініп алынған Bacillus thuringiensis
энтомопатогенді бактерияларының қабыршаққанатты зиянкестерге тимділігін
анықтау.
Зерттеу міндеттері:
1. Жетісу және Іле Алатауларының жабайы жемісті орман
топырақтарының морфо-генетикалық сипаттарын, химиялық құрамдарын,
физико-химиялық қасиеттерін зерттеу.
2. Жабайы жемісті орман топырақтарының микрофлорасын зерттеу.
3. Жабайы жемісті орман топырақтарынан және топырақтағы
бактериоздық белгілерімен ӛлген бунақденелілерден Bacillus thuringiensis
энтомопатогенді бактерияларды бӛліп алу, олардың түр құрамын анықтау және
морфологиялық және биохимиялық ерекшеліктерін зерттеу.
4. Жетісу және Іле Алатауларының қабыршаққанатты зиянкестеріне қарсы
дайындалған жергілікті штаммдардың биологиялық белсенділігіне баға беру.

9
Ғылыми жаңалығы – Жетісу және Іле Алатауларының жабайы жемісті
орман белдеуінде қалыптасқан орманның күңгірт-cұр топырақтарының, оны
түзуші факторлардың бір-бірімен сәйкесті әрекеттесу ерекшеліктерін анықтау
және олардан Bacillus thuringiensis энтомопатогенді бактерияларын бӛліп алу
жұмыстары бұрын бізге дейін жүргізілмеген.
Кӛптеген жылдар бойы зиянды бунақденелілермен күресу үшін Bacillus
thuringiensis бактериясының негізінде жасалған биопрепараттар қолданылып
келеді. Бұл препараттармен ӛсімдіктерді ӛңдегенде бунақденелілер олардың
зиянды әсеріне ұшырамайды. Бактериялардың әсер етуші негізі бунақденелерге
уытты болып келетін ақуызды эндотоксиннің споралары мен кристаллдарын,
экзотоксин және кейбір ферменттер.сондықтан Bacillus thuringiensis – ӛсімдік
қорғаудың қауіпсіз құралы деуге болады. Ол қоршаған ортаға түскенде олар
табиғи реттеуіш механизмін сақтайды және гомеостаз бен экожүйенің тұрақты
дамуының нақты факторлары ретінде жүреді.
Бұл топтың бактериялары топырақтан, бунақденелілерден және қоршаған
ортаның басқа нысандарынан бӛлінеді. Қазіргі уақытта Bacillus thuringiensis
(Bt) бактериясының 80 астам түрлері белгілі, олардың ішінде бунақденелілерге
әртүрлі әсер ететін штаммдар кездеседі .
Қазіргі таңда биопрепараттар нарығының 90-95 % Bt бактериясының
споралы-кристаллды кешенінен тұрады. Дүниежүзінде Bt бактериясы негізінде
20 жуық ӛнеркәсіптік препарат үлгілері жасалған. Тек Ресейдің ӛзінде ғана
оннан астам осындай препараттар шығарылған.
Ӛкінішке орай, біздің елімізде Ақ кӛбелек отандық биопрепаратынан басқа
энтомопатогенді микроорганизмдердің жергілікті штаммдары негізінде
жасалған биопрепараттар жоқ. Бұл биопрепарат Қазакстан Республикасы
териториясында қолдануға рұқсат етілген пестицидтер тізіміне енгізлгеніне
қарамастан, осы күнге дейін мемлекеттік тұрғыдан қаражаттың бӛлінбеуіне
байланысты ауыл шаруашылығында кең кӛлемде қолданылмай келеді.
Жҧмыстың жариялануы және сыннан ӛтуі. Зерттеу қорытындылары
«Выявление активных видов энтомопотогенных микроорганизмов,
осуществляющих биологический контроль над вредителями в Джунгарском
Алатау» (Санкт-Петербург, 2015 ж.), « Оценка вирулентности некоторых
штаммов энтомопатогенной бактерии Bacillus thuringiensis в отношении
гусениц яблонной моли после хранения » (Алматы, 2012 ж.), «Скрининг
новых изолятов энтомопатогенных бактерий Bacillus thuringiensis » (Алматы,
2014 ж.), «Использование различных методов хранения энтомопатогенных
микроорганизмов», (Алматы, 2014ж.) «Использование новых штаммов
энтомапотагеннх бактерий группы Bacillus thuringiensis ssp. для регуляции и
численности яблонной моли» (Алмалыбак, 2014) атты халықаралық ғылыми-
тәжірибелік конференцияларындада баяндалған.
Зерттеудің негізгі нәтижелері кӛрініс тапқан 10 ғылыми еңбектеб
жарық кӛрген. ҚР БҒМ саласындағы бақылау комитетімен ұсынылған ғылыми
басылымдарда – 4. Scopus компаниясының деректер базасына кіретін

10
басылымда – 1. халықаралық ғылыми конференцияларда – 5.Оның ішінде
халықаралық ғылыми - тәжірибелік конференцияларда – 1.
Жҧмыстың қҧрылымы және кӛлемі. Диссертациялық жұмыс
компьютерлік текстпен 112 бетте баяндалған, кіріспеден, 5 бӛлімнен, 5 негізгі
ғылыми тараулардан, қорытындыдан, ӛндіріске ұсыныс пен қосымшалардан
тұрады. Пайдаланған әдебиеттер тізімі 160. Диссертацияға 25 кесте мен 37
сурет кіреді.

11
НЕГІЗГІ БӚЛІМ
1 Әдебиетке шолу
1.1 Энтомопатогенді Bacillus thuringiensis бактериялары бӛлініп
алынған топырақ аймақтарына шолу
Топырақ – барлық материалдық игіліктердің кӛзі. Ол азық-түлік, малға
жем, ким үшін талшық, құрылыс материалдарын береді. Топырақтың ең
маңызды байлық екенін айта отырып, К.Маркс, еңбек – байлықтың әкесі болса,
топырақ – анасы деген. Топырақтың физикалық касиеттері су, ауа ӛткізгіштігі,
су сиымдылығы, құрамы мен құрылым ерекшелігіне байланысты. Топырақтын
негізгі бӛлігін бастапқы топырақ түзуші аналық жыныстан бастау алатын
минералды құрамы анықтаса (85-99 %), біршама бӛлігін топырақ түзілу
процесінде, негізінен осында ӛскен ӛсімдіктер қалдықтары (шіріген тамыр,
түскен жапырақ, т.б.) ыдыраған кезде синтезделіп, түзілетін органикалық.
(қара-шірінді) заттар кұрайды (1-15 %). Жануарлар дүниесі (негізінен
омырткасыздар) тіршілігінің әрекетінен ондағы органикалық заттардың
ыдырауы жылдамдайды. Адамның шаруашылық әрекеті (орманды кесу, шӛп
егу, жер жырту, мелиорация, органикалық және минералды тыңайтқыштар
қолдану) топырақ түзуші кейбір факторларға (мысалы, ӛсімдікке) әсер етіп,
топырақ түзілу процесінің бағытымен қарқындылығын тез ӛзгертеді [1].
Іле Алатауының солтүстік беткейінің орталық бӛлігі үшін,
әртүрлі бағдардағы беткейлердің айтарлықтай айырмашылықтары болған
кездегі, ландшафт белдеулерінің кең спектірлі топырақ-ӛсімдік
жамылғысының тік белдеулігі тән [2].
Топырақ эрозиясы әлбетте антропогендік әрекет әсер еткенде
жоғарылайды және әдеттегі табиғи жағдайларға қарағанда топырақ
жамылғысы ондаған, ал кей жағдайларда жүздеген есе тезірек жүруі мүмкін.
Іле Алатауының топырақ-ӛсімдік жамылғысын зерттеумен түрлі
жылдары Северцева 1873; Аболина 1930; Соседов 1940,1959; Глазовская
1946,1949; Чупахина 1968; Науменко 1996 және т.б. атақты ғалымдар
айналысқан.
Іле Алатауының солтүстік беткейінде топырақ-ӛсімдік жамылғысының
таралуы биіктік пен беткей экспозицияның ӛзгешеліктеріне сәйкес жүреді.
Оның ерекше қасиеттері жер бедерімен динамикалық-геологиялық
үрдістерімен, топырақ түзуші жыныстардың қасиеттерімен құрамымен,
топырақтың жасымен, адамзаттың әрекетімен және т.с.с жағдайларымен
анықталады. Беткейлердің экспозициясы топырақ-ӛсімдік жамылғысының тік
белдеулігінде айтарлықтай ауытқуларды тудырады. Оның оңтүстік
беткейлерде жер бедерін тудыратын үрдістер қарқынды жүреді, ол барлық
биіктік белдеулерінде кездесетін сәл дамыған кристалды таулы-далалық
топырақтың дамуына алып келеді [3].
Іле Алатауында әдетте келесідей тіктік ландшафты белдеулерге бӛлінеді
[4]: Гляциальды-нивальды белдеу (3400 метрден жоғары). Бұл белдеу
шектерінде топырақ жамылғысы тек кейбір жерлерде ғана кездеседі және

12
тастақ әрі сәл дамыған болып келеді. Белдеу таулы ӛзендердің қоректенуі
және қатты жауын-шашын аккумуляциясы ауданы ретінде маңызды рӛл
атқарады.
Альпілік белдеу (3400-3300 метрден 3100-3000 метрге дейін). Топырақ
жамылғысы алпілік таулы-шалғынды, қаңқалы, жұқа қалыңдықты – 20-дан 50
см-ге дейін. Олар мұздықтардың әрекетінен пайда болған кең тараған бедер
түрлері -торгтік аңғарлар, олармен қоса солтүстік, солтүстік–шығыс,
солтүстік–батыс экспозиция беткейлерін басып жатқан қарлар. Қар
жамылғысының асты – қиыршық тас пен қойтастардан тұрады.
Субальпілік белдеу (3100-3000 м-ден 2800-2700 м дейін). Бұл
белдеудің солтүстік экспозиция беткейлеріде қалыңдығы 40-50 см болатын
таулы-шалғындық субальпілік топырақ қалыптасқан. Топырақ асты –
ірікристалды материалдар. Беткейдің оңтүстік экспозициясында таулы-
далалық топырақ кең таралған. Оның таулы-шалғындық альпілік топыраққа
қарағанда қалыңдығы жұқа және қатты тастылық.
Шыршалы ормандардың және биік таулы субальпілік шалғындар белдеуі
(2800-2700 метрден 1800 метрге дейін). Бұл белдеудің солтүстік экспозиция
беткейлері Тянь-Шань шыршаларының қылқанды ормандары және
ормансыз учаскелерін биік шӛпті шалғындармен жабады. Шыршалы орман
белдеуінің жоғарғы бӛлігінде бұталар (түркістандық және сібірлік арша,)
кездеседі. Шыршалы орманның орталық және тӛменгі бӛліктерінің шӛп
жамылғысы алаңдарда қалақай, папоротник және тағы басқа түрлері
кездеседі. Оңтүстік, оңтүстік-шығыс және оңтүстік-батыс экспозиция
беткейлері шалғынды – далалық және бұталы ӛсімдіктермен жамылған [5, 16-
19 б].
Шыршалы ормандар әдетте таулы–орманның күңгірт түсті
күлденбеген топырағында, кӛк терек ормандары таулы–орманның күңгірт сұр
топырақтарында ӛседі. Барлық таулы–орманның күңгірт сұр
топырақтары күлденген топырақтар қатарына жатады. Шалғындардың
астында – топырақтар таулы орманның – шалғынды, қалың гумусты
горизонттылығымен (60-тан 90 см-ге дейін) сипатталады. Бұл топырақтар
кӛптеген жағдайларда сілтісізденген. Бұл белдеуде кристалды
жыныстардың жер бетіне шығулары кӛптеп таралған.
Түрлі шӛпті – астық тұқымдас шалғынды дала және таудың жалпақ
жапырақты орман ландшафттар белдеуі (1800 метрден 1200 метрге дейін).
Бұл биіктік белдеуінде ағаштар мен бұталардың 40 жуық түрі бар.
Ауданның басым бӛлігінде бақшалар және бақтар орналасқан. Солтүстік
экспозиция беткейлерінде кӛктеректі орманның астында таулы
орманды топырағы, ал бұталы - шалғынды даладан тӛмен орманның сұр
топырағымен сілтізденген қара топырақ таралған. Белдеудің тӛменгі
бӛлігінде айтарлықтай гумусты (16-17 %) таудың қара топырағы дамыған.
Бұталы далалық белдеу (1200 метрден 850 метрге дейін). Белдеудің
жоғарғы бӛлігінде әр түрлі топырақта түрлі шӛпті-бұталы, боз шӛпті
ӛсімдіктер астында, әдетте таудың қара топырағы, ал қара-қоңыр

13
топырақтарда ксерофитті шӛптер ӛседі. Белдеудің тӛменгі астықты-жусанды
дала бӛлігінде қара топыраққа ұқсас келетін топырақ таралған.
Белдеулердің анық алмасуы Іле Алатауының орталық бӛлігіне ғана тән,
шығыс және батыс бӛліктерінде ландшафттардың ормансызданып далалануы
жоғарылайды.
Іле Алатауында топырақтанушылар топырақтың 20 типін бӛлген.
А.Ф.Литовченко оларды гидрологиялық әсерлер бойынша 4 топқа біріктіріп
(таулы-шалғындық, таулы-орманды, орманды-далалы топырақ және таулы қара
топырақ) олар әрбір тобына сәйкесінше сипаттамалар берген [6].
И.С.Соседов Іле Алатауының топырағын 2 топқа біріктіруге болады деп
есептеген: орта және биік таулы белдеулерде кӛп таралған ірі кристалды
грунттарда қалыңдығы жұқа тасты топырақтар және аласа таумен-тауалды
белдеуде майда грунтта қалыптасқан топырақ. Жалпы алғанда, Іле Алатауының
топырағы балшықты механикалық құрамымен, жақсы құрылымымен, жоғары
ылғал сыйымдылығымен, елеулі тоңуымен сипатталады. Жоғары кеуектілікке
шамамен 75-85 % таудың-шалғынды альпілік, тау-орманының қара түрлі-
түсті, тау-орманның күңгірт-сұр топырағы және қалың қара топырақ
қабаты тән, ал кеуектілік тӛменгі (50% жуық) – түйіртпексіз таулы-далалық
топырағында кездеседі. Топырақтың кӛлемдік салмағы тау-орман
топырағының 0-10 см қабатында 0,52 г/см3-тен тӛменгі қабатына қарай 1,6
г/см3 ӛзгереді [6,85-96 б.б]. Бұл топырақтардан ең маңызды
сипаттамаларының бірі олардың ең аз далалық ылғал сыйымдылығы болып
табылады. Түрлі типті топырақтардың ылғал сыйымдылығы жайлы толық
мәліметтері А.Д. Карбышеваның жұмысында [7] келтірілген. Іле Алатауы
аумағының басым бӛлігінің топырақтары мен грунттары жоғары
инфильтрациялық және су аккумуляциялау қасиеттерімен ерекшелінеді.
Осының салдарынан беттік су ағыны әдетте шымданған беткейлерде
болмайды және фильтрациясы 0,15- 0,20 мм/мин аспайтын құмбалшықты,
балшықты беткейлерде, беткейлердің жартасты бӛліктерінде, жолдар мен
соқпақтарда, жылжымалық телімдерде және т.с.с. жерлерде байқалады [6,382
б].
Тау орманның күрең –сұр топырақтары Жетісу Алатауының таулы–
орманды-дала аймақтың орта белдеуіне қалыптасқан. Абсолютты биктігі
шамамен 2000-2500 м. Олар әдетте жаппай белдеу түзбейді, керісінше орманды
дала аймағының басқа топырақтарының арасында жекеленген алқаптар түрінде
орналасқан.
Тек Константиновка, Тополевка және Лепсі ауылдарында жаппай таралуы
байқалады. Онда орманды-дала ӛсмдіктерінің арасында жапырақты орман
басты орын алады (алма-қайыңды, кӛктерек).
Жер бедері орташа тік бӛлімдермен (15-200) әртүрлі бағыттардағы
беткейлерімен, ең бастысы солтүстік және солтүстік батыс экспозициясында
айқын байқалады. Сонымен қатар кейбір қыраттардың плато тәріздес шыңдары
мысалы, Қайқан, Белтерен, Суық жайлау, Шұбарағаш, Қотырқайың және т.б.
түрінде кӛрініс тапқан.

14
Топырақ түзуші жыныстар лесс тәріздес шӛгінді. Шығу тегі эолды-
делювияльды. Құрамы құмбалшықтармен балшықтар. Сипатталған
топырақтарда ағаш ӛсімдіктері басым: кӛктеректен тұратын жапырақты
ормандар, теректер, қайыңдар, шетендер, жабайы алмалар, мойыл, ырғай,
бӛріқарақат және басқалар.
Тау-орманының күңгірт–сұр топырақтарының морфологиялық белгілері
бойынша кейбір қара топырақ типіне, оның күлденген және сілтісізденген
типшелеріне жақын. Олардың гумусты қабаттары қара-сұр және күнгірт–сұр
реңге ие болады. Кӛп жағдайда топырақ беті орман тӛсенішімен жабылған.
Күлденген қабат мүлдем жоқ, бірақ кремний сеппесі әрқашанда екінші гумус
қабатында (A2B) байқалады.
Сипатталған орманның күңгірт-сұр топырағы А қабатында жоғары
гумустылығымен (10,57 %), бірақ оның астындағы қабатшада В1 күріт
тӛмендегенділігімен (2,70 %), гумустық қабатының (A+B1), жұқалығымен
(30см) ерекшеленеді [8].
Жетісу Алатауы Сібір мен Орта Азия таулары аралығындағы ӛзіндік кӛпір
тәрізді. Солтүстігінде сібірлік шалғынды-далалы зона, оңтүстігінде далаға тән
ландшафттар тараған. Таудың биіктік белдеулері тау алды мен тау іші
жазықтарында 500—600 м-ден 1200—1400 м-ге дейінгі аралықты қамтитын
шӛлейт зонасынан басталады. Мұнда суармалы егістік алқаптар мен
жайылымдар орналасқан. Кӛкӛніс, жеміс ағаштары, мал азықтық шӛптер мен
астық дақылдары ӛсіріледі. Дала зонасы 1200-1400 м-ден 1800-2000 м-ге дейін
биіктерде орналасқан. Мұнда шыршалы ормандар мен самырсын ӛседі.
Олардың арасындағы шалғындар мал жаюға жарамды. Таудың биік белдеуі
солтүстікте 2200— 2400 м-ден, ал оңтүстікте 2400-2500 м-ден биікте жатыр.
Мұнда субальпі шалғындары, қар мен мұздықтар кездеседі. Биік таудың кейбір
бӛліктерін малшылар жайылым ретінде пайдаланады. Жануарлар дүниесінде
алтайлық және тянь-шаньдық түрлер кӛп [9].
Жетісу Алатауы - Қазақстандағы ірі, әрі құрылымы жағынан күрделі тау
жүйелерінің бірі. Жетісу тау жүйесінің солтүстік және солтүстік-батыс
жалғасы. Ол солтүстік-шығысында Алакӛл казаншұңқыры мен оңтүстік-
батысында Іле ӛзені аңғары аралығында созыла орналасқан. Ұзындығы 450 км,
ені 100-250 км. Жетісу Алатауын батыста Кӛксу, шығыста Боротола ӛзендері
бӛліп жатыр. Осы екі бӛлік Солтүстік және Оңтүстік Жетісу Алатауы болып,
екі үлкен тау жотасын құрайды [10]. Оңтүстік Жетісу тау жүйесінің Сарышоқы
мен Борохоро жоталары түгелдей дерлік Қытай жерінде жатыр. Қазақстан
шекарасына таянған жерде Орашар Барлық тауымен (3300 м) жалғаса отырып,
Жайыр Майлы атты аласа жотаға ауысады. Осы жоталардың арасында кезінде
тектоникалық козғалыстардан пайда болған үлкен аңғар бар. Ол кей жерлерде
батпақты, қамысты, кей жерде құрғақ дала боп кетеді. Аңғар бойында үйінді
тастар кӛп. Жергілікті халық оны қорым дейді. Осы аңғар аркылы Ебінұр
кӛлінің қазаншұңкыры (Қытайда) Алакӛл қазаншұңқырымен шектеседі. Осы
алқап Жетісу (Жоңғар) қақпасы деп аталады. Ерте замандардан бастап осы
қақпа арқылы Шығыс Еуропа мен Азияны, Қазақстанды жалғастырған Жібек

15
жолы ӛткен, онымен Шыңғысхан жаулаушылары, Жоңғар шапқыншылары
жүрген [11]. Жетісу Алатауының Қазақстандық бӛлігі, осы кақпаның
батысынан басталады да, батысқа қарай Боротола ӛзеніне дейін Қытай -
Қазақстан шекарасы бойымен созылады. Осы жерде Жетісу Алатауы
асимметриялық құрылымдағы алып таулы жотаға айналады. Жоңғар-
Алатауының табиғи паркі ӛсімдік және жануарлар әліміне ӛте бай болып,
ғылыми бағытта қызығушылық тудыратын кӛптеген эномикалық және реликті
түрлерімен ерекшеленеді. Жетісу Алатауының флорасы ӛсімдіктердің 2168
түрімен берілген, оның 76 эндемикалық түрі тек қана осы жотада ғана
кездеседі. Жоңғар Алатауының флорасында 21 сирек кездесетін түрлері де бар.
Зерттеу жұмысының ӛзектілігіне: Жоңғар Алатау тауларында ӛсетін
қылқанжапырақты ӛсімдіктер түрлерінің ӛсіп, дамуының морфологиялық
ерекшеліктерін мен таралуын және шайыр жолдарының анатомиялық
ерекшеліктерін зерттеу жатады. Эксперименталдық зерттеу нәтижесінде
қылқанжапырақты ӛсімдіктердің 3 түрлерінің анатомиялық құрылысының
ерекшеліктері анықталды, олардың таралуы зерттелінді. Қылқан жапырақтылар
- сүректі ағаш және бұта түрінде ӛсетін ашықтұқымды ӛсімдіктер класы.
Қазақстанда 2 тұқымдасы бар: қарағай тұқымдасы— қарағай, балқарағай,
шырша, майқарағай және кипарис тұқымдасы— арша, кипарис; мәңгі жасыл
(қыста қылқаны түсіп қалатын балқарағайдан басқасы) ӛсімдіктер. Тұқымы
арқылы, кейде вегетативтік жолмен ӛніп-ӛседі. Жапырағы қылқанды, жіңішке,
түкті не қабыршақты болып біткен, сағақсыз, ӛркендері қысқа, шоқтанып
орналасқан. Мамыр-шілдеде гүлдеп, шілде-тамызда жеміс береді. Қылқан
жапырақтардың сүрегінен құрылыс материалдары жасалады, қағаз жасайтын
үгінді, канифоль, скипидар. Шырша – қарағай тұқымдасына жататын биіктігі
30-50 м, баяу ӛсетін қылқанжапырақты ағаш. Қазіргі кездегі флорада бұл класс
ашық тұқымдылардың ішіндегі түрлерінің саны жағынан ең кӛбі (600 дай түрі
бар). Негізінен солтүстік ендікте кең таралған, бұл жерлерде бір немесе бірнеше
түрлерден тұратын туыстары: қарағай, шырша, май қарағай, балқарағай үлкен
қылқанжапырақты орман түзеді. Оңтүстік ендікте қылқан жапырақтылар
қоңыржай климатты облыстарда орман түзеді (Огненная Земля, Патагония,
Жаңа Зеландия, Тасмания). Тропикалық облыстарда олар тек таулы жерлерде
ӛседі [12].
1.2 Топырақ жәндіктері мен микроорганизмдері
Топырақта тіршілік ететін тірі ағзалардың зат және энергия алмасуы тек
қана ӛзара емес, сонымен қатар топырақтың басқа да құрама бӛліктерімен
байланысты болады.
Микроағзаларды зерттеудің негізін қалаған И.В. Вернадский топырақтың
пайдалы генофондын кез-келген ластану түрлерінен сақтаудың мәселелерін
шешудің ӛте маңызды екендігін атап ӛткен. Ол топырақ биотасының тіршілігі
және сонымен қатар топырақ ағзалардың химиялық құрамының арасындағы
байланыстарды кӛрсеткен.
Барлық топырақ биотасы келесі топтарға жіктеледі:

16
1. Микрофлора-бактериялар, актиномицеттер, саңырауқұлақтар,
балдырлар.
2. Микрофауна (0,002-0,2 мм) - инфузориялар, тамыраяқтылар.
3. Мезофауна (0,2-2,0 мм)- нематодтар, кенелер.
4. Макрофауна (2-20 мм) - энхитреидтер, моллюскалар, қоңыздар және
олардың личинкалары.
5. Мегафауна (20-200 мм) - жауын құрттар, омыртқалылар.
6. Топырақта уақытша мекен ететіндер - тышқан тәрізділер.
ТМД топырақтарының мезофаунасы М.С. Гиляровтың еңбектерінде
кеңінен қарастырылган. Ол топырақта ӛмір сүретін жануарлардың, осы ортамен
байланыстылығына қарап үш негізгі топтарға бӛлген: геобионттар – ӛмір сүру
циклі толығымен топырақта ӛтеді; геофилдер – ӛмір цикілінің біраз бӛлігі
топырақта ӛтетін жануарлар; геоксендер – топырақта кездейсоқ кездесетін
қолайсыз жағдайда қашып тығылатын немесе қорғанатын уақытша топырақ
тұрғындары. Геофилдер әдетте жоғары гумусты топырақтарда басым [13,14].
Іле Алатауының солтүстік беткейімен тау алды жағында топырақтардың
мезофаунасы З.А Тукенованың зерттеу еңбегінде егжей - тегжейлі
қарастырылған.
Автордың мәліметі бойынша Іле Алатауының топырақтарының
мезофауналық потенциалы тау етегі топырақтағы кәдімгі сұр топырақтардан
таудың қара және орманның күңгірт-сұр топырақтары бағытында артқан. Егер
қара топырақтарда негізгі доминант омыртқасыздардан жауын құрттары мен
құмырсқалар болса, күңгірт- қара қоңыр топырақтарда доминант болып тек
жауын құрттары саналады. Ал құмырсқалар субдоминантты деңгейінде
кездеседі. Мұнда субдоминанттар басым, мысалы, шегірткелер. Ал сирек
кездесетіндерге жапырақ жегіштер жатады.
Орманның күнгірт-сұр және қара топырақтарында сапрофагтар мен
(ӛсімдіктің ыдыраған қалдықтарымен қоректенетін организмдер) зоофагтар
(басқа жануарлармен қоректенетін жануарлар) басым. Олар мезофаунаның 85
% құрайды, мӛлшерлерінің ара қатынасы 1:1 [15].
Топырақ жәндіктері мен микрорганизмдері жүйесі табиғаттағы зат
айналымында, топырақ құрамының құрылу процесінде, оның құнарлылығының
түзілуінде белсенді рӛл атқарады. Олар барлық табиғи органикалық
қосылыстарды ыдырата алады. Яғни, олар топырақта ӛскен ӛсімдіктердің
барлық қалдықтарын ыдыртып, ӛсімдіктерге сіңетін әр түрлі минералды
заттармен қатар, қарашірінді түзеді. Мұндай организмдерге, мысал ретіндс,
бұршақ тұқымдас ӛсімдіктердің тамырларында ӛмір сүріп, ауадағы азотты осы
ӛсімдіктердің тамырларына жинайтын түйнек бактерияларын жатқызуға
болады. Бүкіл әлемдегі ауыспалы егіс жүйесінде бұршақ тұқымдас
ӛсімдіктердің ерекше орын алатыны да сондықтан. Топырақ құнарын тиімді
пайдалану үшін, оның құрамындағы организмдерге жағдай жасап, ауыспалы
егіс жүйесі мен агротехниканы дұрыс колдану қажет. Топырақтың 10 — 15 см-
ге дейінгі беткі құнарлы қабатында га-на 10 т бактериялар, сол шамада

17
саңырауқұлақтар, 4 т жауын құрты,140 кг балдырлар,17 кг-дай әртүрлі
жәндіктер кездеседі [16].
Н. Сейткали Іле және Жетысу Алатауының жабайы жемісті орман
топырақтарын зерттей отырып, мынадай мәліметтерге қол жетізген. Топырақ
микроорганизімдер үшін табиғи субстрат болып табылады. Ол жерден
актиномицеттердің әртүрлерін бӛліп алуға болады. Бірақ актиномицеттердің
биомассасының басым кӛп бӛлігі споралар болып табылады. Қоректік
орталарға егу жүргізу әдістемесі бойынша популияциялардың колониясын
санау кезінде биомассаның тек 1-4 % мицелийлер құрады. Ол органикалық
заттары мол микрозоналарда табылды. Зерттеу нәтижелері бойынша
крахмалды-казеинды ортада топырақтағы актинобактериялардың саны 0,11 ±
0,001 млн. кл/г болатындығы анықталды. Олар гетеротрофтар, яғни дайын
органикалық қосылыстармен қоректенеді, соның нәтижесінде олар табиғаттағы
зат алмасу үрдісіне қатынасады. Зерттеу мәліметтері топырақтағы
саңырауқұлақтардың 9,8 ± 1,6 млн. кл/г болғандығын кӛрсетті. Олар әртүрлі
субстарттардың бетінде ӛсетін хлорофилсіз микроорганизмдер. Ашытқы
организмдерін санау сабуро қоректік ортасында жүргізілді. Олардың
топырақтағы саны 0,011± 0,001 млн. кл/г болды. Бұлай болғанда топырақтағы
ашытқылардың құрамы, бактериялар мен саңырауқұлақтарға қарағанда
айтарлықтай тӛмен [17].
Топырақ табиғатта микроорганизмдердің тіршілік етуінің шынайы ортасы
және бастапқы резервуары болып табылады (азот, кӛміртек, күкірт, темір және
т.б.). Олар топырақтың түзілу және ӛзіндік тазару үрдісіне белсенді қатысып,
табиғатта заттар алмасудағы орны орасан зор. Топырақ таулы ортадан желдің,
судың тірі организмдердің және олардың органикалық қосындыларының, яғни
ӛсімдіктермен жануарлардың ӛнімінен пайда болуынан түзіледі. Топырақ
микроорганизмдері сулы және коллоидты қабықтарда тіршілік етеді және
топырақ бӛліктерін бұзады. Оларда саңырауқұлақтардың гифтерімен
қозғалғыш бактерияларының жылжуы және кӛшіп жүруі олардың арасында
микроскопиялық жіңішке сулы қабаттардың болуына әкеледі.
Топырақ микрофлорасының сапалық құрамы ӛте әртүрлі: Олар әртүрлі
бактериялар, актиномицеттер, спирохет, архебактериялар, микоплазм,
саңырауқұлақтар, вирустардан тұрады. Әртүрлі топтағы микроорганизмдердің
құрамы мен ара қатынасы топырақ түріне, оны ӛңдеу әдісіне, құрамындағы
органикалық заттарға, ылғалдылыққа, климаттық жағдайларға және тағы басқа
себептерге байланысты болып келеді. Құмды топырақтарда аэробты
микроорганизмдер тіршілік етеді, ал балшықты, ылғалды (оттегінің кіруі қиын)
ол жерде негізінен анаэробты микроорганизмдер тіршілік етеді. Топырақ
микроорганизмдері 25-45 0С аралығында кӛбейе алады, ал термофильдіктер –
одан да жоғары тампературада қалыпты ӛмір сүре алады.
Микроорганизмдер күрделі биоценозда болады, олар ӛздерінің араларында
ӛсімдіктермен бірге антагонистикалық және симбиотикалық ара қатынаста
сипатталынады. Ӛсімдіктердің тамыр аймағында бактерия ӛте кӛп: олар
қарқынды кӛбею және жоғарғы белсенділікті резосфера деп аталатын зонасын

18
түзеді. Топырақтың ризосфералық зонасының микрофлорасы бай, әртүрлі және
әртүрдегі ӛсімдік үшін ӛзіндік ерекшелігі болады. Микроорганизмдер
ӛсімдіктердің тамырлық бӛліну ара қатынасында дұрыс хемотаксисті болады
және олар органикалық қосылыстардың минералдану үрдісіне қатысады,
сонымен қатар ӛсімдіктерді тез қабылдайтын минералды заттармен, яғни
дәрумендермен, ауксиндермен қамтамасыз етеді. Олар ӛсімдіктердің
заталмасуына белсенді қатысады. Топырақтың ӛзіндік тіршілігінде ауа және
тамақ үшін үлкен күрес болып жатады. Кӛбіне бір топтағы бактериялардың
тіршілігі басқа топтағы бактериялардың антибиотикалық бактериоциондық
әсерінен басылып қалады.
Бір жылда топырақтағы микроорганизмдердің мӛлшері бірнеше
миллиардқа дейін жетеді. Олардың ең кӛбі тазартылған топырақта - 4,8-5,2
млрд. Аз мӛлшерде орман топырағында, құмда (1,2-0,9 млрд).
Микроорганизмдердің тірі үлесі 1 гектар топырақта орташа 1000 кг. құрайды.
Микроорганизмдердің жыл мезгілінің ӛзгерісіне байланысты: кӛктемде саны
ӛседі, жазда максимумға жетеді, жаздың ортасында азаяды, (күн сәулесінің
белсенділігінің әсеріне байланысты) күзде тағы да кӛбейеді, ал қыста азаяды.
Микроорганизмдердің топырақта таралуы әртүрлі. Топырақтың
құнарлануына қарамастан жоғарғы 1-2 см қабатында микробтар аз. Бұл күн
сәулесінің ультракүлгін сәулелеріне және құрғату үрдістеріне байланысты.
Микрофлора 10-20 см тереңдікте жақсы дамыған. Бұл қабатта негізінен
биохимиялық үрдістер жүріп жатады, яғни органикалық заттарға ауысу, бір-
бірлерін ауыстыратын микроорганизмдердің тіршілік етуінің әртүрлілігі. Одан
тереңдеген сайын топырақ микрофлорасы азайып, 4-5 м тереңдікте
микроорганизмдер ӛте сирек кездеседі. Артезиандық құдықтардан алынатын су
стерилді болып келгендіктен, құрамында бактериялар қоретенетін
амминоорганикалық қосылыстар болмайды.
Топырақ микрофлорасының құрамындағы бактерияларды физиологиялық
топтарға бӛлінген. Олар әртүрлі органикалық заттардың біртіндеп ыдырау
үрдісіне қатысады.
1. Бактериялар – аммонификаторлар, олар ӛсімдік қалдықтарының шіруіне
әкеледі. Шіру үрдісіне мына бактериялар қатысады: B.subtilis, B.mesentericus,
Serratia marcescens, бактерия тұқымы: Proteus; саңырауқұлақ тобы: Aspegillus,
Mucor, Penicillium; анаэробты: C.bsporogenes, C.putrificum; уробактериялар:
Urobacillus pasteuri, Sarcina ureae.
2. Нитрифицирлейтін бактериялар: Nitrosomonas және Nitrobacter.
Nitrosomanas аммиакты азот қышқылына дейін тотықтандырып, нитрит түзеді.
Ал Nitrobacter азотты азот қышқылына дейін қышқылдандырып, нитрат түзеді.
Нитрификцирлейтін бактерияларды 1899 ж. топырақ микробиологияның негізін
қалаушы С.Н. Виноградский ашты. Оның айтуы бойынша нитрификация үрдісі
2-басқышта жүреді және нитрификцирлейтін бактериялар автотрофты және
спецификалық қозғалыс әсерін иемденген. Нитрификация үрдісіндегі
микробтардың маңызын Виноградскии былай сипаттаған: «Микробтар зат
алмасудың дұрыс ӛзгеруіндегі негізгі агенттер болып тұрады; олар әртүрлі

19
реактивтерді тасымалдайтын тірі тасымалдаушылар. Оларсыз әртүрлі керек
үрдістер, яғни зат алмасу үрдісі болмас еді».
3. Еркін тіршілік ететін азот фикцирлейтін бактериялар ауадан
атмосфералық азотты алады және тіршілік ету үрдісінде молекулярлы азоттан
ақуызды басқа да азотты қосылыстарды түзеді. Оларды ӛсімдіктер қолданады.
4. Жасымықты ыдырататын бактериялар ашыту түрлерін тудырады. Олар
микробтардың кӛмірсутекті органикалық қосылыстарды ыдыратқан кезде
байқалады (сүт қышқылы, спирт, май қышқылы, прапион қышқылы, сірке
қыщқылы).
5. Күкірт, темір, фосфор және тағы басқа элементтер айналымына
қатысатын бактериялар – күкірт бактериялары, темір бактериялары және т.б.
Олар әртүрлі қышқылдану және қалпына келу үрдістеріне қатысады.
Топырақ кабаты әр түрлі тірі организмдер тіршілік ортасы ретінде
пайдаланатын табиғи орта. Онда топырақ микроорганизмдерінен — бактерия,
саңырауқұлақ, сәулелі саңырауқұлак, т.б. және кӛптеген омыртқасыз
жануарлардан қарапайым жәндіктер, кұрттар, былқылдақденелілер, ересек
бунакденелілер мен олардың дернәсілдері, жер қазғыш омырткалылар, т.б.
кездеседі. Олардың санының ӛзгеруі жыл маусымына да байланысты, кӛктем
мен күзде кӛбірек болса, қыс пен жазда азаяды. Қарапайым жәндіктер ӛздігінен
тіршілік ететін, бір жасушалы ӛте ұсақ организмдерден тұрады. Топырақ
арасындағы тұнба суларда бірнеше мыңға жуық түрлері мекеңдейді. Олар 4
класқа бӛлінеді: талшықтылар, саркодиналар ,споралылар және инфузориялар.
Қарапайым жәндіктер кәсіптік бағалы жануарлардың негізгі қорегі, ал басқа
түрлері паразит түрінде адамдар мен жануарларға ауру жұқтырады. Мыс,
талшықтылар, споралылар құстар мен адамдарды безгек ауруына
шалдықтырады. Жұмыр кұрттардың бір түрлері топырақтың арасында (50 -
1200 млн/га-ға дейін) шоғырланып, топырақтың қалыптасу үрдісіне қатысады,
ал екіншілері ӛсімдіктерді зақымдап, адамдар мен омыртқалы жануарларды
ауруға шалдықтырады.
Сақиналы кұрттардың да кейбір түрлері топырақта мекендейді. Мысалы,
жауын кұрты топырақты ӛз ішегінен ӛткізіп, оны қопсытып, араластырып, ауа
мен ылғал ӛткізгіштігін жақсарта отырып, топырақ құрамының қальптасу
үрдісіне қатысады. Топырақты буынакаяқтылар арасында шаянтәрізділер,
ӛрмекшітәрізділер, бунакденелілер кластарынан ӛрмекшілер, кенелер, бұ-
заубастар, шаяндар, сымқұрттар, т.б., хордалылардан жыландар, кесірткелер,
кірпілер, суырлар, сарышұнақтар, қосаяқтар, тышқандар мекендейді. Кірпілер
мен жертесерлер зиянкестер мен кеміргіштерді жойып пайда келтірсе, кейбір
түрлері адамдар мен жануарлар арасында ауру қоздырғыштарын таратады
(сарышұнақтар, тышқандар, т.б;).Топырақ жәндіктері мен микрорганизмдері
жүйесі табиғаттағы зат айналымында, топырақ құрамының құрылу үрдісінде,
оның құнарлылығының түзілуінде белсенді рӛл атқарады. Олардың барлық
табиғи органикалық қосылыстарды табиғи немесе бірнеше органикалық
қосылыстарды ыдырата алады. Яғни, олар топырақта ӛскен ӛсімдіктердің
барлық қалдықтарын ыдыртып, ӛсімдіктерге сіңетін әр түрлі минералды

20
заттармен қатар, қарашірінді түзеді. Мұндай организмдерге, мысал ретінде,
бұршақ тұқымдас ӛсімдіктердің тамырларында ӛмір сүріп, ауадағы азотгы осы
ӛсімдіктердің тамырларына жинайтын түйнек бактерияларын жатқызуға
болады. Бүкіл әлемдегі ауыспалы егіс жүйесінде бұршақ тұқымдас
ӛсімдіктердің ерекше орын алатыны да сондықтан. Топырақ құнарын тиімді
пайдалану үшін, оның құрамындағы организмдерге жағдай жасап, ауыспалы
егіс жүйесі мен агротехниканы дұрыс колдану қажет. Топырақтың 10 — 15 см-
ге дейінгі беткі құнарлы қабатында гектарына 10 т бактериялар, сол шамада
саңырауқұлақтар, 4 т жауын құрты, 140 кг балдыр,17 кг-дай әртүрлі жәндіктер
кездеседі [18].
1.3 Кристалл тҥзуші бактерияларды зерттеудің тарихы
Тұрақтанған шаруашылықтың, соның ішінде бал арасы және жібек құртын
ӛсірудің туындауы арқасында адамдардың жәндіктер патологиясына деген
қызығушылығы пайда болды. Бал арасы аурулары жӛнінде Аристотелден
байқауға болады, ол біздің эрамызға дейінгі ІV ғасырда ӛмір сүрген.
Бунақденедідердың ауруының алғашқы тіркеуі XIX ғасырдың бас жағына
жатады. Осы күндері кейбір зерттеушілермен анықталғаны, саңырауқұлақтар
және насекомдар инфекциясының арасындағы байланыстары байқалды. [19,
20].
Бунақденелілердегі инфекциялық аурулардың алғашқы сипаттамасының
ресми приоритеті, әдетте Аугостино Бассиге жатқызылады (1835-1839), ондағы
кӛретініміз, жібек құртының Bomdux mori ақ мускардин қоздырушысы
саңырауқұлақ болып табылады, олар оны Botrtis bassiana деп атаған. А.Бассидің
жұмысынан кейін бунақденедідердың ауруларын анықтауға арналған кӛптеген
зерттеулері бар [21].
Споратүзуші энтомопатогенді бактерияларды Луи Пастер ӛзінің тұт жібек
құртының ауруларын зерттеу кезінде байқаған болатын. Бірақ, споралы
кристалл түзуші микроорганизмдер жӛніндегі алғашқы кӛз жеткізерлік
жұмыстар Жапонияда жарияланған болатын. 1901-1902 жылдары жапон
оқымыстысы Ишивата тұт жібек құрттарынан спора түзуші бактерияларды
бӛліп алды, оны Bacllus sotto деп анықтады. 1908 жылдан бастап Ивабуши
Жапонияда «sotto» - тұт жібек құрты құртының параличін қоздырғышы
жӛнінде жазды. Еуропада Bacllus thuringiensis жӛніндегі мәліметтер «Ephestia
kuehniella Zell» қамбалық отшаларды әкелгеннен кейін пайда болды. Сонымен,
Тюрингия провинциясында (Германия) жоғарыда аталған насекомның ауру
құртынан неміс ғалымы Берлинер 1911-1915 жылдары спора түзетін
бактерияларды анықтады және оны Bacillus thuringiensis деп атады [22].
Одан кейінірек соған ұқсас штамм анықталды және оны Маттес жіті
зерттеді. Ол бактерияны қамба отшасына қарсы пайдаланғысы келді, бірақ
онысы жүзеге аспады.
Ӛткен жүз жылдықтың отызыншы жылдары Еуропада жүгері сабағының
кӛбелегіне күресі жӛнінде (Puraista nubilalis Hbn.) халықаралық компанияда
кеңінен зерттеулер жүргізілді. Метальников, Хегула, Шорин және Гуш

21
тәжірибелерінде бірқатар бактериялардың түрлерін пайдаланды және сонымен
қатар Bt, ол жаңа атпен анықталған B. ephestiae, B. gelechiae, B. galleriae, B.
Cazaubon [23]. Барлық изоляттардың бірдей ұқсастығы олардың жәндіктерге
тигізген ықпалының тиімділігі бойынша кӛрініп тұр. 1928 жылы Гуш Bt.
зертханада және далада сынады және құрттар санының айтарлықтай
тӛмендеуіне қол жеткізді [24].
Айта кету керек, осындай тиімділікке, қашан бактериялдық препараттар
ӛндірісінде стандартталуы және оның бӛлек партиялары ӛзінің сапасы бойынша
ажыратылмаған кезде қол жеткізілді, ӛйткені жәндіктерге бактериялардың
ықпалының қағидасы белгілі емес еді. Осыған қарамастан, осы препарат
«Спореин» Париждегі Либек зертханасында жақсы сапада болды, оны, 1950
жылы осы препараттың қайтадан тиімді қолданылғанын Жако тәжірибесі анық
жақсы кӛрсетеді [23].
Еуропада 50-ші жылдары Bt. тек қана венгер зерттеушілері және оқушы
Гуша – Клемент жұмыс жасады, олар осы бактерияны америкалық ақ кӛбелекке
қарсы тиімді пайдаланды [25].
АҚШ-та 1951 жылы Штейнхауз ӛзінің Матес Bt. штаммы жӛнінде
бақылауларын жариялады, ол ӛзінің бунақденелілерге деген тиімділігін 20 жыл
бойы жасанды қоректік ортада ӛсіргеніне қарамай сақтап қалған. Осы мәлімет,
бұл бактерияға деген зерттеушілердің үлкен қызығушылығын тудырды.
Әртүрлі елдерде энтомопатогенді кристалл түзуші бациллдердің алуан түрінің
анықталуымен және жаңа сипаттамасымен айналыса бастады. Сонымен K.
Toumanoff, C. Vago тұт жібек құртынан Е.В. Талалаев Bacillus cereus subs.
Alesti. анықтады, Н.А. Красильников және А.Б. Гукасян Томск облысының
орманында қабыршақ қанатты насекомдардан жаңа түрін - Bacillus tuviensis
Kras et Guk анықтады [26-29].
50-ші жылдары ВИЗР-де үлкен бал арасы отша құрттарының (Galleria
mellonella) эпизотиясы кезінде – ауруын тудырушы Bt ssp. Galleria қоздырғыш
анықталды [30-32].
Бациллалардың Bt тобының біркелкіленуінің бірінші сызба нұсқасын А.Н.
Heimpel және T. Angus жасады. Авторлар АМК білім беру тәрізді қасиеттердің
негізінде, фосфолипаза және пигменттің үш түрін сипаттады: Bacillus
thuringiensis, Bacillus finitimus, Bacillus entomocidus. Ӛз кезегінде, әрбір түрге
олар бірқатар алуантүрліліктерді енгізді [33].
Энтомопатогендік кристалл түзуші бациллалардың жіктелуі бойынша
негізгі зерттеулер француз ғалымы de Barjak және Bonneffoi A. жүргізді. Олар
Bt. маңызды 35-тен астам белгілерін зерттеді, соның ішінде біркелкілену
мақсатында алғаш рет серодиагностиканы пайдаланды, ол бұрамалы
агглютинацияға негізделген [34]. Bt бактерияларының әртүрлі зерттеушілермен
алынған 24 штаммын зерттеп және жинақтай отырып, оларды алты
сероварларға бӛлді. Ары қарай олармен басқа жаңа сероварлар сипатталды. de
Barjak және A. Bonneffoi кристалл түзуші бактериялар штаммының екеуін
сипаттады - Bt. subsp. aisawae (серовар 7) және subsp. morrisoni (серовар 8).
Олар серологиялық зерттеулер кӛмегімен, тӛртінші сероварға жататын

22
штаммдарды бекітті, олар антигендік құрылымымен айрықшаланады. 10
сероварға жататын штаммдар - Bt. subsp. darmstadiensis 1968 жылы Krieg, de
Barjak сипаттады. Грузияда ӛлген тұт жібек құрттарынан Bt. subsp. caucasicus
бӛлініп алынды, ол сонымен қатар антигендік құрылым бойынша 10 серовар
ретінде сипатталған. Африкянның штаммының subsp. darmstadiansis кейбір
биохимиялық қасиеттері жағынан айырмашылығы бар: оң лецитиназдық
белсенділік, пигмент түзілуі және термотұрақты экзотоксиннің болмауымен.
Кейінірек, жіті зерттеу кезінде олардың антигендік формулалары анықталды:
10 а - Bt. subsp. caucasicus, 10 a 10 b - Bt. subsp. darmstadiansis [35,36,37,38,39].
Bt. subsp. toumonoff (11 серовар) Krieg, Bt. subsp. thompsoni (12 cеровар) de
Barjak және Thomson [40,41].
De Barjak және Lemelle үшінші серовар ішіндегі жаңа нұсқаның антигенді
формуламен За3b - Bt. subsp. kurstaki, тіршілікті байқады, сонда de Barjak және
Bonneffoi - subsp. caucasicus бесінші серовар ішінде, subsp.galleriaе -
биохимиялық және серологиялық қасиеттерімен ерекшеленеді [42,43].
1970-1977 жылдары Ресейде жаңа штаммдар сипатталды. Томск облысы
және Алтай аймағының жәндіктер популяциясынан Bt. subsp. galleriae бӛлініп
алынды және сипатталды. Г.М. Иванов және А.Б.Гукасян капуста кӛбелегінің
құрттарынан жаңа сероварлар бӛліп алды: Bt. subsp. aragusuniensis және subsp.
brassicae. Сібірде В.П. Ходырев эпифитті микрофлорадан жаңа штамм
анықтады - subsp.foliates, ол биохимиялық қасиеті жағынан subsp.sotto-ға ӛте
ұқсас, тек оның сарыуыздық ортада қызыл-сары пигмент бӛлетін қасиеті бар.
Серологиялық қасиеттерді зерттеу кезінде сарысуға қарсы 14 сероварларды
пайдаланып, осы штаммның антигенді құрылымына зерттеулер жасалды
[44,45,46].
70-ші жылдары Орта Азияда республикаларында энтомопатогенді
микроорганизмдер іздестіруге анағұрлым мән беретін болды. Сонымен, Е.Н.
Троицкая, Е.Н. Михайлов, Г.А. Плужников фитофагтардан кӛп мӛлшерде
кристалл түзуші бациллалар алды, олардың арасында жаңа түрі болды және
оны Bt. subsp. Asiae-mediae деп атады [47].
Жапон оқымыстысы алынған штаммның антигендік құрылымын зерттей
отырып, жаңа серовар анықтады, оны олар Bt. subsp. Kyushiensis (11а серовар:
11с) деп атады. Осы штамм toumanoff-тан кейбір биологиялық қасиеттерімен
ерекшеленді [48].
70-80-ші жылдары L.Goldberg және L. Margalit алғаш рет Израилде маса
личинкасынан Bt. subsp. Israelensis (Н14) анықтады, олар маса және шіркей
личинкасы үшін жоғары инсектицидті болып келеді. De Luece, Simonson және
Larson зерттеулерінің арқасында тағы да екі жаңа түрлер пайда болды: Bt.
subsp.dacota және subsp. Indiana. Bt. subsp. Ostriniae (серовар 8а: 8С) және
wuhanensis (жарақсыз белгілі бір серотипті) жапондық ғалымдармен жаңа түр
ретінде ұсынылды. Жапондық ғалымдар үш жаңа сероварлар анықтап және оны
сипаттады [49].
Bt subsр. tohokuensis (серовар 17), subsр. kamomotoensis (серовар 18) және
subsр. tochigiensis (серовар 19). Осыны ғалымдар 1989 жылы типтік жолақтар

23
үшін 5Н антигендік субфакторлардың қатысуын анықтады (3a, 3b. 3c, 3d және
3e) Н3 сероварында. Олармен екі жаңа түрастыларының аттары ұсынылды Bt.
subsр. sumiyoshiensis (Н серовар 3а:3d) және Bt subsр. furuoraensis (Н серовар
3а:3d:3e) [50].
1983 жылы Bt subsр. tenebrionis бактериясы анықталды, ол қаттықанатты
жәндіктердің патогендеріне (отр. Coleoptera) – С патотипке жатқызылған.
Қаттықанаттыларға тән δ-эндотоксиндердің Lepidoptera-лардан айырмашылығы
бар – ӛзіндік қасиеті бар ақуыздар, қабілеттілігі кристалдарда тек қана
протоксиндер түрінде болмайды, және де сонымен қатар молекулярлық
массасы 55-67 кДа, белсенді токсиндер түрінде болады. Олар бактериялық
ӛнімдердің кристалл түзілу кезеңінде ақуыз протеиназаларымен
модификациялану нәтижесінде пайда болады [51].
1.4 Энтомопатогенді бактериялар негізінде алынған биопрепараттар
1956 жылы Bt сұйық қоректік ортадағы мәдени ӛсіру жӛніндегі алғашқы
мәліметтері пайда болды, 1958 жылы Г. Ванкова кейбір жағдайлардағы терең
заводтық Bt ферментациясы жӛніндегі бір ғана еңбекті жариялады. Сол
уақыттан бері сатылымда, құрамында споралар және Bt кристаллдары бар
ӛндірістік препараттар пайда болды [52,53].
XX жүзжылдықтың 70-80 жылдары әлемде келесі ӛндірістік препараттар
ӛндірілді: энтобактерин, дендробациллин, инсектин, токсобактерин,
битоксибацеллин, лепидоцид, БИП және гомелин – бұрынғы Кеңес Үкіметінде;
биотрол, бактан, агритол, пароспорин, турицид, дипел – АҚШ-та; спореин,
бактоспеин и плантибак –Францияда; бактукал – Югославияда; батурин –
Чехословакияда.
Иркутскінің мемлекеттік университетінде микробиологиялық препарат
жасап шығарылды – Bt. subsp. dendrolimus Tal. кристалл түзуші бактериялар
негізінде түзілетін дендробациллин, cеротип 4, шығарылудың алғашқы
жылдарында, ол сібір жібекқұртымен күрес үшін орман шаруашылығына
арналған болатын. Сонымен қатар дендробациллин ауылшаруашылығында да
жеміс және мақта дақылындағы зиянкестермен күресте үлкен сұраныспен
қолданыста болды.
Тек қана Bt. subsp.israilensis (Н-14) штаммдар негізінде келесі препараттар
алынды – текнар (Швейцария), вектобак (АҚШ, бактимас (Франция), москитур
(Чехословакия), скитал (Англия), булмосцид (Болгария).
1963 жылы КСРО-да Bt ssp. Galleria негізіндегі энтобактериннің алғашқы
ӛндірістік шығарылымы ӛндіріле бастады. Препарат жеміс-жидек
дақылдарындағы, капустадағы, бақ және шаруашылық егістіктерінде және
ормандарда жапырақкеміруші насекомдардың комплексіне қарсы қолданылды.
Энтобактеринмен кезек алмастырушы, оның культурадағы жоғары
фагосезімталдығына байланысты ӛндірісін тоқтатты, 80-ші жылдары
лепидоцид пайда болды. Лепидоцид Bt ssp. Kurstaki негізінде пайда болды,
штамм Z-52, диірмен отшасының (Ephestia kuеhniella) құртынан Э.Р. Зурабова
анықтаған. Айта кету керек, 1970– ші жылдан кейін, кӛптеген

24
энтомопатогендік бактериалдық препараттар әлемде осы Bt түрастының
негізінде жасап шығарды, ол алғаш рет 1962 жылы бӛлінді [54]. Bt ssp. kurstaki
бір спораға эндотоксиннің кристаллдарын екіден бастап беске дейін ӛндіреді.
Сондықтан лепидоцид инсектицидтік белсенділігі бойынша дендробациллинді
және энтобактеринді жиі басып озды, ол дегеніміз біздің бұрынғы елімізде осы
препараттың танымалдылығының себебі болып табылады [55].
Патовар А- ға сонымен қатар Bt ssp. Thuringiensis жатады. Осы түрасты β-
экзотоксиннің анағұрлым қарқындылығымен әйгілі. Алғашқы осы негіздегі Bt
препараты – битоксибацилин (БТБ) – бұрынғы КСРО-да жасалып шығарылған;
кейінірек турингин пайда болды, ол спора және кристалсыз тек β-
экзотоксиннен тұрады [56].
Шіркейдің және масаның санын азайтатын бактокулицид Ресейде
ӛндіріледі [57]. Бұл препарат Bt ssp.israliensis бациллалары негізінде жасалған.
Түрасты спора және эндотоксиннің белгісіз формадағы кристаллын, β-
экзотоксинді жасап шықпайды. Эндотоксин масаның личинкаларында ішек
жасушасының деструктивтік ӛзгерістерін тудырады. Масаның Aedes және Culex
туыстарының личинкалары бактокулицидке анағұрлым сезімтал болып келеді .
Соңғы уақыттарда бактокулицидтің әсер ету спектрлері кеңейіп келеді, оны
күріштің (Cricotopus silvestris Fabr) және шампиньонның (Lycoriella fumosum L)
масашаларына қарсы пайдалануға мүмкіндіктер кӛрсетілді [58,59]. 90-шы
жылдары Ғылыми-зерттеу институтының ауылшаруашылық
микробиологиясында (Санкт-Петербург қ.) жаңа тиімділігі жоғары –
қаттықанатты жәндіктерге қарсы селективті әсер ету қабілеті бар Bt негізіндегі
бацикал препараты жасалынып шығарылды [60].
Ертеректе сол жерде Р Bt subsp. tenebrionis (Н-8а, 8b) штаммы негізінде
децимид препараты жасап шығарылды. Децимид 1962- 1996 жылдары ауыл
шаруашылығы үшін пайдалануға рұқсат берілген препараттардың тізіміне
енгізілген [61].
А.Я. Лескова, ӛңделетін ӛсімдіктерде және оңтүстік пен солтүстік
аудандардың топырағында сақталатын биопрепараттар туралы кеңінен
зерттеулер жүргізіп мынадай қорытындыға келеді, онда инсоляция және
жаңбыр ӛсімдіктердегі спора және токсиндердің сақталу кезеңін қысқартады
[62]. А.Г. Кольчевский күн және жаңбыр инсоляциясының капуста және картоп
жапырақтарындағы битоксибациллин (БТБ) - препаратының сақталу ықпалын
зерттеді. Оның байқағаны, ауа райы ашық болғанда (жарық 98 мың люкс)
тіршілік қабілетті спораның мӛлшері, БТБ-мен ӛңделген ӛсімдіктерде, 5 сағат
ішінде 81,1 %-ға, ауа райы бұлтты болғанда (жарық 23 мың люкс) – 63,7 %-ға
тӛмендеген [63].
Жібек тұты ӛсімдігінің ӛңделген жапырақтарында және топырақта Bt
тобының бактериялар сақталу мерзімі, сонымен қатар жібек тұты ағашы үшін
улылық сақталу мерзімі анықталды. Алғаш рет жібек тұты жапырағының
дендробациллиннің дақыл-ӛнімдерінің дамуын ингибирлеу қабілеті анықталды
[64].

25
1.5 Bacillus thuringiensis бактерияларымен жасалынып шығарылатын
токсиндер
1962 жылы А. Хемпел Bt токсиндердің келесі жіктелуін берді:
1) α- экзотоксин – ӛсіп келе жатқан бактерияның ферменті, соның ішінде
фосфолипаза С, анығы, Clostridium welchii α-токсинмен аналогия бойынша;
2) β- экзотоксин – аздап суда еритін термотұрақты нуклеотидті табиғаттың
токсині, ол бактериалды клеткамен қоршаған ортаға шығарылады;
3) γ- экзотоксин – біркелкіленбеген фосфолипаза;
4) δ- эндотоксин – параспоральды ақуыздық кристаллдар [65, 66].
Осы уақытта тек δ- β- токсиндер жақсы зерттелген. α- және γ- токсиндер
жӛнінде мәлімет аз [67]. А. Криг α- токсинді термолабильді ақуыз ретінде
сипаттаған, насекомдар және тышқанға қарсы белсенді, α- экзотоксинді
сонымен қатар «тышқан факторы» деп атайды. Ол штаммы B.cereus тәрізді, Bt
штаммында да ӛндіріледі [68].
Ең негізгі δ-токсиннен бӛлек термотұрақты β-экзотоксин насекомдардың
патогенезінде ерекше рӛл атқарады. Ол алғаш рет 1959 жылы thuringiensis
түрастының Bt дақылды сұйықтығында байқалған болатын [69]. Авторлар
бекіткендей, токсин суда ериді, сонымен бірге жоғары және тӛмен
температураларға тӛзімді. Экзотоксин түзу Н1, Н4, Н8-Н10 сероварларындағы
типтік штаммдарда байқалды. Бұл кристалл түріндегі токсин, ақуыз болып
есептелмейді, ӛйткені 15 мин бойы 120 °C автоклавтау кезінде ӛзінің
белсенділігін сақтап қалады [70].
A.Burgerion зерттеулері кӛрсеткендей экзотоксин қабылданатын
бунақденелілердің личинкасына қарсы анағұрлым тиімді. Ол насекомдардың
ӛзіндік түлеуін тежейді. Мысалы, капустаның ақ кӛбелегінің тұмсығында
атрофия жүреді, соның нәтижесінде имагоның тамақтану мүмкіндігі толығымен
жойылады. Колорад қоңызында астыңғы ернінің щупиктері жойылады, ал
тілінің ұшы созылып кетеді, аталған деформациялар тұқым қуалайды.
Личинкалардың токсиндерге анағұрлым сезімталдығы, аз мӛлшерде –
жұмыртқадағы ұрықтар, қуыршақтар және ересек бунақденелілер [71].
β- экзотоксин кристаллдық эндотоксинге қарағанда ӛте кең кӛлемде ықпал
етуге қабілетті болады. Ол тек қабыршақ қанаттылырға ғана улы болып қоймай
(соның ішінде совок), сонымен қатар түзу қанаттылырға да, қос қанаттылар
және кейбір қоңыздарға да улы болады. Белоктық токсинге қарама-қарсылығы,
насекомдарда термотұрақты экзотоксиннің салыстырмалы баяу реакциясын
тудырады, ықпалы ӛте баяу дамығандықтан, ол ешуақытта бірден ӛліп
қалмайды.
ТМД елдерінде термотұрақты Bt. Экзотоксинді зерттеу бойынша үлкен
тәжірибе жинақталған [72-76].
Бактерия ықпалының ӛзіндік қасиеті кешенді, әртүрлі метаболиттермен
байланысқан, олардың бастысы ақуыздық пароспоральдық түзілу болып
табылады, ол кристалл деп аталады. Кристалдар, дельта-эндотоксиндердің
ақуызынан тұрады (немесе Cry ақуыздар), формасы, ӛлшемі және
бунақденелілерге ӛзіндік қасиеті жағынан ерекшеленеді. Энтомоцидтік

26
ақуыздардың кӛпшілігінің молекулярлық массасы 130-145 кДа болып келеді. Bt
бактерияларының инсектицидтілігінің дәстүрлі сипаты пароспоралдық қосылу
бунақденелілердің биотесттеріне және морфологиясына негізделеді. Ӛзіндік
қасиеті жоғары токсиндер белгілі, олар Lepidoptera, Coleoptera және Diptera
отрядтарының белгілі бір ӛкілдерін ӛлтіруге қабілетті [77].
Ангус Ханнейдің сӛзі бойынша Bt sotto түзілетін параспоральдық денеге
талдау жүргізгенде (кристалдар), олардың құрамында токсин бар болғанын, сол
арқылы бактерия бунақденелілерді залалдайтындығын байқағаннан кейін ғана
түсіндірілген, Bt вегетативті жасушада параспоральды кристалдық қосылудың
болуын 1915 жылы Берлинер анықтаған. Бірақ, оның табиғаты мен ықпалының
механизмін байыппен зерттеу XX ғасырдың екінші жартысында басталды. Осы
кезде негіз салушы білімдер бипирамидальды кристаллдарды зерттеу кезінде
алынған болатын, қабыршақ қанаттылар отрядының бунақденелілері үшін
патогенді [78,79].
Осы кезде ӛндіретін Bt δ- эндотоксиндердің кӛптеген мӛлшері белгілі.
Дельта- эндотоксиндердің кӛптеген гендері плазмидтерде кездеседі [80].
Инсектицидті кристалдық ақуыздың генінің бірінші рет клондалуынан кейін Bt
токсиндердің әртүрлі гендерінің бӛлінуі бойынша кӛптеген мӛлшерде
мәліметтер жинақталған, ол дегеніміз оларды жүйелендіру қажеттігін кӛрсетеді
[81]. Кристалды инсектицидтік ақуыздарды кодтайтын гендері және токсиндері
Cry ретінде белгіленді. Бастапқыда Cry-ақуыздар олардың белсенділік спектрі
және молекулярлық массасы негізінде жіктеледі (м.м.) [26]. Алғашқыда осы
құрлымда, Cry I қабыршақ қанаттыларға қарсы белсенді болады және м.м. 135
кDа болады, ал Cry II – қабыршақ қанаттылар және қос қанаттылар керісінше
және м.м. 65 кDа болады. м.м. 65 кDа болғанда, бірақ белсендісі қатты
қанаттыларға қарсы ақуызды Cry III деп белгіледі [55].
δ- эндотоксин спора түзу кезінде бактериалды жасушаның ішінде пайда
болады. Параспоральды денелер, немесе кристалл шындығында протоксин
тәрізді болады. Қабылдайтын қожайындарға улы болу үшін, ол тек қана
жасушасынан ажырап қана қоймай, сонымен бірге бунақденелілердің ішегіне
протеолитикалық ферменттер ықпалымен белсенділік танытуы керек.
Кристалл түзуде Bt-тің тұрақты белгісі болады және оның патогенділігінде
маңызды рӛл атқарады. Осы түрдің кейбір изоляттары ӛсіру кезінде де жасанды
қоректік ортада 50 жыл бойына ӛзінің улы кристаллдарды ӛндіру қасиетін
сақтап келген [82]. Кристаллдар ӛзінің химиялық құрамы бойынша ақуыздық
қосылыс болады, оның құрылымына 18 аминқышқылы енеді. Кристаллдары
жоғары температураның әсеріне тӛзімділігі аз, суда ерімейді.
Кӛптеген зерттеушілердің жұмыстарында әсер ету механизмі және
кристаллдың улылығы анықталды. Кристаллдың әсері қабыршақ
қанаттылардың құрттарында жақсы зерттелген. Залалдандыру үрдісі кезінде
улы кристаллдар ішектің перистальтикасын тоқтатады және эпителиді
бүлдіреді, соның әсерінен бактериялар ішектен дене қуысына жеңіл еніп кетеді,
олар сол жерде тез кӛбейіп және септицемия тудырады, соңы ӛлімге әкеледі
[83,84,85].

27
Bt әртүрлі штаммдарында кристаллдық ақуыздың улылығы
бунақденелілердің әртүрлі түрлері үшін ауытқиды.
Осылай, кристаллдар B. thuringiensis subsp. thuringiensis рН 11,8 [86],
кезінде, B. thuringiensis subsp. sotto кезінде рН 10,0 [95]. B. thuringiensis subsp.
alesti – кезінде рН 11,0-12,2 [87], B. thuringiensis subsp.dendrolimus – кезінде рН
9,5 [88, 89], B. thuringiensis subsp. kurstaki – кезінде рН 10-11 протеаз әсерінен
ішек сӛлінде ериді [90].
Басқа зерттеушілер айтқандай, орта ішектің протеиназдары қабыршақ
қанаттылырдың кей түрлеріндегі қабылдау механизмінде айтарлықтай рӛл
атқармайды. Broorhart G.L, Kramer K.L. оңтүстік қамба шырақшасы құртының
Вt. бактерияларына тӛзімділік механизмін зерттей отырып, орта ішектің
протеиназ ерекшеліктерімен байланысты, тӛзімді және қабылдайтын
құрттардағы протеолитикалық белсенділігі үшін рН мәні бойынша ақуыз
жинақтау бойынша айтарлықтай айырмашылықтар байқалмады [91].
A. Heimpel, T.A. Angus қабыршақ қанаттылырдың қабылдайтын түрлерінің
арасында (ішектің сілтілі рН) екі типте бӛледі, кристаллдарды жұтқаннан кейін
байқалатын белгілері арқылы ерекшеленеді. Алғашқы типіне қоректің кенеттен
тоқтау кезінде болатын және дененің жалпы жансыздануы жатады. Осы
белгілер гемолимфаның сілтілігінің жоғарылауынан, ішектің рН тӛмендеуімен
және ішек эпителиінің бұзылуымен болады. Кейбір осы типке жататын түрлері:
тұт жібек құрты (Bombux mori), емен жібек құрты (Antheraea pernyi), сонымен
қатар Protoparae spp [92].
А.Я. Лескова зерттеулері бойынша Subsp. galleria (Н-5) штаммдарының
насекомдардың сегіз отрядының 88 түрінің сегізінде әсерін сезіне отырып,
оларды қабылдау деңгейіне қарай тӛрт топқа бӛлді. Бірінші топқа қабыршақ
қанаттылар жатады, жоғары қабылдағыштығына қарай – оларға нағыз күйелер,
шұбараша күйесі, жапырақ ораушылар, нимфалидтер мен ақ кӛбелектер. Екінші
топқа қабыршақ қанаттылар ӛкілдері жатады, құрттардың ӛлімі 40-70 %
кӛрсеткіштен аспады. Олар шырақшалар тұқымына жатады, үшінші топқа
жататындары қабылдамайтын және тӛзімді болып келеді – сыналған
насекомдардан тӛртінші топта түзу қанаттылар, қатты қанаттылар, екі
қанаттылар ӛкілдері жатады [93].
Ішек қуысының бактерияларының патогенді қасиеті болуы мүмкін және
жаппай аурудың себебі болуы мүмкін.
В.П. Поспелов зерттеулері бойынша жаппай эпизоотии зерттей отырып,
қорытындылағаны, жиі себепкері бунақденелілерде тіршілік ететін
бактериялар. Олар әлсіреген физиологиялық жағдайдан қоршаған ортаның
әсерінен патогенді болады.
Ары қарай В.П. Поспеловтың идеясы бактерия-симбионаттар жӛніндегі
потанциалдық патогендер ретінде зерттеушілер жанында дамыды.
Кӛрсетілгендей, әртүрлі факторлар (температура, ылғал, интоксикация, екінші
реттік инфекциа) «патогенде» ішекте тіршілік ететін бактериялар ӛтуін
ынталандырады [94,95].

28
Әсіресе, жиі бактериоздан зертханада немесе инсектарийдегі ӛсірілетін
бунақденелілер зардап шегеді, оларға табиғаттағыдай жағдай жасау мүмкін
емес.
Н.П. Исакова капуста кӛбелегінің ішек флорасының рӛлін, Вt subsp.
Galleriae зерттеді. Алынған мәліметтер ішек флорасының инфекция қабылдау
деңгейінің сандық байланысын кӛрсетті. Ішектің бактериямен кӛп қаныққаны
кезінде Вt патологиялық үрдіс болған [96,97].
Т.В. Завезенова, Р.В. Барайщук, Л.В. Кирюшкин сынды ғалымдар
бацилланың тоғыз серотипіне жататын эндотоксиндерінің отыз штаммының
морфологиялық сипатын жіті зерттеді. Жүргізген тәжірибелерде құрттарды
таза кристаллдармен залалдау бойынша Galleria mellonella, әртүрлі
морфологиялық топтарға жатқызды, кристаллдар морфологиясы арасындағы
тікелей байланыс және оның улылығымен бекітілмеген [98].
В.А. Бурлак токсин белсенділігі және бунақденелілер реакциясы кристалл
формасына қарағанда ӛлшеміне байланысты болып келеді деп тұжырымдаған.
Ол G.mellonella құрттарда Вt subsp. kurstaki штаммына сынақ жүргізді,
морфологиялық колония типін түзетін және параспоралық енудің 2
морфологиялық әртүрлі топтары. Ӛлімнің аздығын (15-20 %) штаммдар
туындатады, бар кристаллдар бипирамидальды формада 72°шыңдық ұшымен
(колонияның тегіс типі), (55 % дейін кӛбі) - штаммдар, кристаллдары шың
бұрышымен октаэдра формасында 80±1°. ҚХР әртүрлі аудандарының
топырағынан Вt изоляттар алынды, олар 17 серотипке жатады.
Бунақденелілердің 9 түріне d-эндотоксиндерге талдау және сынақ жүргізілді:
қабыршақ қанаттылар, қоңыздар, қос қанаттылар. Мозайкалық-бипирамидалық
кристаллдар ақуызымен 130-140 кДа және 65-68 кДа жоғары улы екендігі
белгілі болды. Куб, полиэдр, шар тәріздес кристаллдар қоңыз личинкасына
қарсы тиімді болды. Полиморфты кристаллдар жоғары улы Prodenia liture
[99,100].
H. Herman, H. Whitely Ақуыз кристаллдарын генетиканың заманауи
әдістерін қолданып зерттеді. Bt Олар айтқандай 14 cry гендері белгілі, ол ақуыз
токсинінің синтезін кодтайды. Осы гендер және осыған сай ақуыздар 4 класқа
бірікті: белок, қабыршақ қанаттылырға улы, I классқа, қос қанаттылар – II және
IV, қатты қанаттылар – III [101].
М. Hafer, H.S. Salama Bt ssp. kurstaki тиімділікті жоғарылатуға
биохимиялық әсер ету жолымен, жәндік ішегіндегі жағдайды оңтайландыруға
бағытталған, патогеннің улы бӛліктерін босату үшін қажетті. Ол үшін калидің
карбонатын енгізді, мақта совкасына қарсы эндотоксин потенциалын
күшейтуші. Зерттеу кӛрсеткендей, параспоральды кристаллдар орта ішектің
асқорыту сӛлінің әсеріне ұшырап және ферментативті белсенділікпен
қарқынды коррелирлейді [102].
Осы кезде әдебиетте Bt эндотоксиндер құрылымы жӛніндегі ақпараттар
кеңінен жинақталды, зерттеу үшін заманауи биохимияның, молекулярлық
биологияның, гендік инженерияның, сонымен қатар электронды микроскопия
әдістері қолданылады.

29
В. Я. Ягудин, А.С. Крылов сынды зерттеушілер ақуыздың әртүрлі Н-
серотиптердің Bt параспоральды кристаллдарын электрофорез әдісімен
зерттеді. Кристаллдарда бактериялар 1, 3, 4, 5, 6 және 10 серотиптер байқалды
1-2 белоктан м.м. 130-145 кДа және бір қосымша фракциямен м.м. 55-120 кДа.
Зерттелген ақуыздардың құрамында кем дегенде 3антиген типі кездеседі, бӛлек
немесе әртүрлі сарысуға қарсы тестілеу арқылы анықталады. Т.А. Смирнова
спора және кристаллдардың бірқатар Bt штаммдарына электрондық-
микроскопиялық талдау жүргізді. Соның нәтижесінде, Bt кристаллдардың
ультрақұрылымдық ұйымының гетерогенділігіне тән, ол түрастымен
байланысты; эндотоксиннің формасы және кристаллдардың нәзік құрылымы
патотиппен коррелирленеді, оны энтомоцидтік әсердің белгілі бір спектрімен
штамм скринингі үшін қолдануға болады [103].
Патотипті эндотоксиннің кристаллдар құрылымының корреляциясы
жӛнінде басқа ғалымдар да ұсыныс жасайды. Осылай, кристалдар түзетін,
қабыршақ қанатты бунақденелілердің (Lepidoptera отряд) қабыршақ
қанаттылар отрядының ӛкілдері үшін улы бактериялар, А патотипке, ал
қосқанаттылар үшін улысы (Diptera отряд) - Д патотипке жатады. Т.Г. Юдина
бунақденелілерге қатысты Bt полипептидтерінің және инсектицидті
белсенділіктерінің арасындағы антимикробты корреляцияны анықтады. Автор
мынадай қорытындыға келеді, полипептидтердің антимикробты белсенділігі
бацилл енгізуде осы бактериялар үшін жәбірленуші- бунақденелінің денесінде
соңғы микрофлорадан басым болып және бірқатар жағдайларда бациллярлық
споралардың ӛсіп кетуін оңтайлы жағдаймен қамтамасыз етеді [104, 105,106].
1.6 Қазақстандағы зиянды жәндіктерге қарсы микробиологиялық әдіс
бойынша Bt қолдана отырып, ғылыми зерттеулер жҥргізу
Қазақстанда алғаш рет кристалл түзуші бактериялар 1962 жылы ҚазСРО
ҒА микробиология және вирусология институтында М.В. Яловицин және
ҚазӚҚКҒЗИ мамандары ашқан.
Авторлар зиянды бунақденелілер үшін табиғи эпизоотия кезінде ӛлген
алма күйесі, жусан жапырағын жегішінің бацилласы және т.б.
бунақденелілерден уыттыларын бӛліп алды. Білімнің сол деңгейі үшін
толықтай жеткілікті зерттелді және олардың қасиеті, түрлері анықталды, ал
жусан жапырағын жегішінің штаммы негізінде аздаған мӛлшерде ВТС
бактериалды препараты алынды. Жүргізілген жұмыстарды тек қана
Қазақстандағы микроорганизмдердің осы тобының (серотиптер таралуының
ӛзіндік қасиеті, биологиялық қасиетінің ерекшеліктері, әсер ету спектрі,
анағұрлым уытты немесе токсигенді штаммдар таңдауы және т.б.) бастамасы
ретінде қарастыруға болады. Ӛкінішке қарай, кӛрсетілген авторлармен алынған
штаммдар, ары қарай жоғалып кетті. Бірақ, қазір осы облыста біздің
мамандарға жергілікті жоғары уытты штаммдармен жұмыс кезінде кӛп күш
жұмсалуда [107,108].

30
Қазақстан Республикасының МАН микробиология және вирусология
Институтында ӛсімдіктердің ауруы және зиянкестерімен күресте тиімді
биопрепараттарды жасаудың негізі жасалды [109].
Алматы облысының Іле Ауданының Ынтымақ ауылында ҚазҒЗИ және ӚК
жеміс-жидек және дақылдары жұмысшыларымен 2004-2005 жылы ағаш
ӛскіндерін барлау кезінде емен орман алаңында америка ақ кӛбелегінің ұясы
табылды, ондағы құрттар 100 %-ға ӛлген. Осы зиянкестердің ӛлу себебін
анықтау үшін ҚазҒЗИжәнеӚК биотехнология лабораториясында және ИСиЭЖ
СО РАН бунақденелілер патологиясы зертханасында (Новосибирск қ.) жіті
түрде микробиологиялық зерттеулер жүргізілді. Ауру құрттардан Bt subsp.
kurstaki кристалл түзуші бактерияларының штаммдары алынды және
біркелкілендірілді. Алынған штаммның 2127-3k биологиялық тиімділігінің
зиянкестер түріне тәуелділігі Z-52 –лепидоцидтің продуценті эталонынан 1,5-3
есе жоғары болып шықты.
2006 жылы алынған бактериялар негізінде алғашқы қазақстандық
биопрепарат «Ақ кӛбелек жасалынып шығарылды». 2007-2008 жылы препарат
тіркеуден ӛтті және ауылшаруашылық және жеміс-жидек дақылдарының
зиянды қабыршақ қанатты бунақденелілеріне қарсы сынақтар жүргізілді. Сынақ
мәліметтері бойынша препараттың биологиялық тиімділігі 90-100 %-ға жетті ол
зиянкес насекомдардың түріне байланысты. Осы уақытта (2009 жылдан бастап)
препарат, кейбір қабыршақ қанаттыларға қарсы ӛндірісте қолданысқа жіберілді
және Қазақстан Республикасының территориясында 10 жылдық мерзімге
пайдалануға рұқсат етілген Пистицидтер анықтамасына енгізілді (улы
химикаттар) [110].

31
2 ЗЕРТТЕУ МАТЕРИАЛДАРЫ МЕН ТӘСІЛДЕРІ
2.1 Зерттеу нысандары
Жетісу және Іле Алатауының жабайы жемісті орман топырақтарынан
бӛлініп алынған Bacillus thuringiensis энтомопотогенді барктериялары.
Топырақ — жер қыртысының құнарлы беткі қабаты. Сол себептен
топырақта алуан түрлі микроорганизмдер жиналады. Микроорганизмдердің
топырақтағы құрамымен мӛлшері жыл маусымында ӛзгеріске түсіп отырады.
Мәселен, қыста топырақтағы бактерия саны азаяды, кӛктемде қатты артып,
жаздың басында барынша кӛбейеді де, содан соң едәуір кеміп, күздікүні қайта
кӛбейеді. Әсіресе топырақтың 1 г микроорганизмдердің кӛптеген түрлері
кездеседі. Микроорганизмдер жердің беткі бетінде аз, себебі күн кӛзі тура
түсуінен кеуіп, немесе ӛліп қалады. Жерден 5-20 метр тереңдікте мыңдаған,
милиондаған микроорганизмдер кездеседі. Топырақтың беткі қабаттарында
аэробты микроорганизмдер кездессе, ал терең қабаттарында анаэробты
микроорганизмдер кездеседі. Зерттеу жұмысында зерттеу нысаны ретінде
жабайы жемісті орман топырағынан бӛлініп алынған энтомопатогенді бактерия
штаммдары қолданылды [111].
Бунақденелілер микрофлорасы. Бунақденелілердің микрофлорасы екіге
бӛлінеді: бұл клеткаішілік және клеткааралық. Бунақденелінің жасушаішілік
микрофлорасы жасуша ядросын жаулап алады немесе цитоплазмасында
сақталып қалады. Бұл микроорганизмдер жәндіктің симбионттарын кӛрсетеді.
Жәндіктердің құрсақ клеткасында ӛмір суретін симбионттар мицетом деп
аталады. Мицетомдар бірнеше жәндіктерде кездеседі олар: кӛпаяқтылар,
қоңыздар, шилан. жасушаішілік микроорганизмдер дақылдары жасанды
қоректік орталарда ӛспейді. жасушааралық микроорганизмдер жасушаішілік
микроорганизмдерге қарағанда алуан түрлі болып келеді. Олар жәндіктер
ішегінің үстінгі жабындарында болады. Ауру жұқтырған жәндіктерді кӛбінесе
олардың жаппай ӛлімге ұшыраған орындардан, ал кейде популяциясын мұқият
қарап шығу арқылы жинайды. Табиғатта ӛлген жәндіктерді ағаштардың
түбінен, жапырақ астынан кездестіруге болады. Жәндіктердің ӛлекселері
жапырақтың беткі қабатына бекініп тұруы мүмкін. Ӛлген жәндіктерді
топырақтың беткі қабатынан және ӛсімдіктердің қалдықтарының арасынан да
кездестіруге болады. Бунақденелілердің мүшелері мен құрылысы ӛте нәзік
болып келеді. Субстраттан ажыратқанда тез бұзылып кететіндіктен, оны
кӛбінесе субстратпен бірге жинаймыз. Ӛлі жәндіктерді беті мақта тығынмен
жабылған құрғақ пробиркаларда сақтайды. Пробрика сыртына жинақталған
орны туралы мәлімет жазылады. Кейде кристалл түзуші бактерияларды
тасымалдайтын тірі жәндіктер де болуы мүмкін екендігін білуіміз керек. Сол
себептен талдау жасау үшін 50-200-ге дейін жәндіктің үлгісін жинап,
зертханаға жібереді. Зертханада жәндіктердің барлығын зерттеп, ауру және
ӛлімге ұшырағандарын бӛліп алады. Жәндіктердің жарты бӛлігін эфирмен
ұйықтатып микрофлорасын тексереді [112].
Bacillis thuringiensis штаммдарының уыттылығын және инсектицидті
әсері бақылау үшін тест-объекті ретінде жапырақ ширатқыш кӛбелегінің

32
жұлдызқұрттары (Archips crataegana) және алма күйе кӛбелегі (Yponomeuta
malinellus Zell.) қолданылады.
Жапырақ ширатқыш кӛбелектің жұлдызқұрттары (Archips crataegana)
16-аяқты, сирек түктері бар, басы қою қоңыр немесе қара, денесі сұр, қара
түсті. Ұзындығы 1 см дейін жетеді, түрлі ӛсімдіктердің жапырақтарымен
қоректенеді. Жапырақ ішінде ӛрмек құрып ішінде оралып тұрады.
Маусымның аяғында қуыршаққа айналып, екі аптадан кейін кӛбелекке
айналады (сурет 1) [113].
Жапырақ ширатқыш кӛбелегінің
жұлдызқұрттары(Archips
crataegana)
Алма күйе кӛбелегінің
жұлдызқұрттары (Yponomeuta
malinellus Zell) қолданылады
Сурет 1- Жапырақ ширатқыш кӛбелегінің жұлдызқұрттары (Archips
crataegana) және алма күйе кӛбелегі (Yponomeuta malinellus Zell)
2.2 Далалық тәжірибе жҥргізу әдістемесі
2.2.1 Зерттеулер жүргізілген аймақтар топырақтарының морфо-гентикалық
сипаттары, құрамдары мен қасиеттерін анықтау
Далалық жағдайда кескін қазу;
Далалық жағдайда топырақты зерттеу топырақ кесінділерінде жүргізіледі.
Топырақ кесінділері негізгі (терең), жартылай кесінділер (жартылай
шұңқырлар) және прикопкалар болып бӛлінеді. Негізгі кескін топырақтың
толық таксономиялық атауын анықтау үшін қолданылады. Онда барлық
топырақ қабаттары айырылып зерттелінеді. Оның тереңдігі әдетте 150-300см
аралығында болады. Негізгі кесінділерді бедердің ең қалыпты деген
мүсіндерінде жартылай кесінділермен құмбалшықтар топырақ түрлерінің
таралу шекараларын анықтау мақсатында пайдаланылады. Олардың тереңдігі
әдетте сәйкесінше 75-100 см және 25-75 см.
Топырақ кесіндісі ұзындығы 120-150 см, ені 60-80 см. Кескінді қазып
болған соң, оған сипаттама бермес бұрын күн сәулесі түсетін қабырғасын
пышақтың ұшымен жоғарыдан тӛмен аршып оның табиғи қалыптағы күйіне
келтіру қажет. Кескінді түсіне, жаңа жарандыларға, құрылымына және басқа да
морфологиялық белгілеріне қарай генетикалық қабаттарға бӛліп, олардың

33
жатқан тереңдігі мен қалыңдығы ӛлшенеді. «Топырақтың қайнауын» тексеру
үшін 10 %-дық НСІ ерітіндісін топырақ үстіне тамызады. Одан соң әр қабатты
морфологиялық тұрғыдан сипаттауға кіріседі. Оның түсін, ылғалдылығын,
гранулометриялық құрамын, тамыр жүйесінің таралу сипатын, құрылымын,
тұтқырлығын, ондағы жаңа заттар мен кірме заттарды, бір қабаттан екіншісіне
ауысу сипатын анықтайды. Кескінге морфологиялық сипаттама бергеннен соң
қабаттардан топырақ үлгілері алынып, топырақтың типі, типшесі, тегі, түрі
анықталып, оның толық атауы күнделікке жазылды.[114,115,116].
Алынған топырақ үлгілерін зертханаға әкеліп құрғатады да ұнтақтап, тор
кӛзінің диаметрі 1 мм електен ӛткізеді. Әрбір үлгіні картоннан жасалған жеке
қораптарға салады. Дайындалған үлгілерден топырақтың агрохимиялық
кӛрсеткіштері анықталады (сурет 2) [117].
Сурет 2- Зерттеу жүргізілетін аймақтың топырақ кескіні
Топырақтың морфологиялық белгілерін зерттеу кезінде оның құрамын,
топырақтың қарашірікті (А), ӛтпелі (В) және аналық жыныс (С) қабаттары
баяндалып жазылды [118].
Топырақтағы құбылыстарды бағалау тӛмендегідей зерттеу әдістерінің
нәтижелерінен алынған мәліметтерге сүйене отырып, жүргізілді: топырақтан
үлгілерді алу ҚРСТ 28168–89, топырақтағы гумус мӛлшерін И.В. Тюрин
әдісімен анықтау ҚРСТ 26213–91, топырақтағы жалпы азотты-И.Г. Кьельдал
әдісімен анықтау ҚРСТ 26107–84, топырақтағы жалпы фосфор мен калийді

34
анықтау ҚРСТ 26264–84, топырақтағы жеңіл гидролизденетін азотты-Тюрин-
Кононова әдісімен, жылжымалы фосфорды-Мачингин әдісімен, жылжымалы
калийді-жалынды фотометрде анықтау ҚРСТ 26205-84, 85 топырақтың су
сүзіндісінің иондық құрамын анықтау ҚРСТ 26423-85, 26428-85, 26425-85,
26426-85, 26424-85, 25428, топырақтың pH кӛрсеткішін анықтау ҚРСТ 26425-
85, топырақтағы катиондар сиымдылығын-Бобко-Аскинази-Алешин әдісімен
анықтау ҚРСТ 17.4.4.01-84.
Топырақтағы ылғалдылықты анықтау
- 1 г абсолютты құрғақ топырақты зерттеу үшін, зерттеуге алынған
топырақтан 10 г орташа үлгі таңдау;
- бос бюксті ӛлшеу;
- дымқыл топырақ салынған бюксті ӛлшеу;
- кептіргіш пеште 105°С тұрақты салмаққа жеткенше қыздырып топырақ
салынған бюксті ӛлшеу;
-абсолютты құрғақ топырақты ӛлшеу;
- формула бойыншыа 1 г дымқыл топырақтағы абсолютты құрғақ топырақ
мӛлшерін анықтау:
W%=100 × a:b
а – буланған судың массасы
в – құрғақ топырақ массасы
2.3 Зертханалық тәжірибе жҥргізу әдістемесі
2.3.1 Зерттеу жүргізілген аймақтардың топырақтарының жалпы
микрофлорасын зерттеу
Топырақ үлгілерін мұқият тазаланған және спиртпен сүртілген пышақпен
немесе күрекпен алады.
Алдымен 2-3–сантиметр беткі қабатын алып тастау керек. Сонан кейін
зертелініп отырған аумақтан монолит алынады. Монолиттің ұзындығы үлгі
алынып отырған қабаттың қалыңдығымен сәйкес болуы керек. 200 м2 ге дейінгі
аумақтан әрқайсысының салмағы 0,5 кг болатын 10 үлгі алынады.
Сынамаларды залалсыздандырылған мӛшекте мұқият араластырыды,
араластырлған сынамадан орташа салмағы шамамен 0,5 кг етіп алып оны
залалсыздандырылған матадан жасалған дымқыл пергаментті қалтаға салады.
Қалтаның аузын мықтап байлап, оны полиэтиленді қалтаға салады.
Полиэтиленді қалтаға үлгінің қай ауданнан алынғандығы және агротехникалық
фонның жергілікті бедері жазылған және топырақтың қасиеті мен қандай
ӛсімдіктер ӛсетіндігі жазылған этикетканы да салады.
Егістікті емес жерлерден топырақ алған кезде, алдын-ала 30х30 см2
аумақтағы ӛсімдік жамылғысы мен түскен жапырақтарды тазартады. Бұнымен
қоса топырақтың шымды қабатын 2 см ге дейін аршу керек. Сосын зертеліп
отырған учаскенің әртүрлі жерлерінен сынамалар алынып, орташа үлгі
дайындалады. Егер үлгілер ормандағы немесе шағын аумақтағы учаскеден
алынса, үлгілердің салмағы 50-100 г болуы мүмкін. Сондай ақ топырақ үлгілері
кравт қағазынан жасалған пакетке немесе мақталы тығынмен жабылатын үлкен

35
пробиркаларға салады. Талдауға дейін топырақ үлгілері тоңазытқышта
сақталуы керек.
Микробиологиялық талдауға ҥлгілерді дайындау Жақсылап араластырылған топырақты спиртпен сүртілген және от
жалынына ұсталған құрғақ әйнектің үстіне тӛгеді. Топырақты шпателмен
мұқият араластырады және тегістеп жаяды. Пинцетті пайдаланып тамырлардан
және басқада бӛгде қоспалардан арылтады. Жұмысқа кіріспестен бұрын
шпатель мен пинцентті жалынға қыздырады және ауада аздап салқындатады.
Әйнектегі топырақтың әр жерінен аз кӛлемдегі топырақты алып, оны алдын ала
дайындалған стерилді фарфор чашкада, үлгінің орташа салмағын 1 г етіп
техникалық таразымен ӛлшейді.
Топырақ агрегаттарын бұзылысқа ұшырату үшін және топырақ
бӛліктерінен бактерия клеткаларын десорбцияға ұшырату үшін үлгіге арнаулы
ӛңдеу жұмыстарын жүргізеді. Алдын ала сыйымдылығы 250 мл болатын екі
колба дайындайды, біреуінде 100 мл стерилденген құбыр суы болады, екіншсін
бос қалдырады. Бірінші колбадан 0,4-0,8 мл су алып, онымен фарфор
чашкадағы ӛлшеп алынған топырақты паста тәрізді күйге келгенге дейін
суландырады, сосын қоспны стерилді резеңке пестикпен немесе резеңке қолғап
киіп, қолмен 5 мин ұнтақтайды. Бірінші колбадағы суды толығымен пайдалана
отырып, ұнтақталған топырақ массасын (от жалынының маңайында) бос
колбаға кӛшіреді. Колбаны топырақ суспензиясымен бірге шайқағыш апаратта
5 мин шайқайды. Осыдан кейін суспензияны ірі бӛлшектер (түйіршіктер) шӛгу
үшін 30 сек қояды да, іле-шала оны препарат дайындауға немесе ӛсіруге
пайдаланды. Мұнда бастапқы алынған суспензиядағы зерттеліп отырған
материал 100 есе сұйылтылады.
Микроорганизмдерді зерттеу мен есепке алу үшін Петри табақшаларына
беттік егу жүргізіледі [119].
Әдістің мәні зерттеліп отырған белгілі микроорганизм суспензиясын
Петри табақшасына қатты қоректік ортаға егіп, ӛскен колонияларды санау
болып табылады. Кохтың қағидасы бойынша мұндағы әр колония бір
клетканың кӛбею нәтижесі ретінде саналады. Зерттелетін суспензияның белгілі
кӛлемін қатты қоректік ортаға егу арқылы, ӛсіп шыққан колониялардың саны
негізінде бастапқы құрамында қанша микроорганизм клеткалары болғандығын
айтуға мүмкіншілік береді. Кӛбінесе Кох әдісі бойынша жүргізілген
микроорганизмдерді сандық анықтау нәтижелерін клетка саны арқылы емес,
колония түзетін бірлік (КТБ) ретінде кӛрсетеді [120].
Қарапайым бояу, жағынды дайындау техникасы
Материалдар және құрылғылар. Майсыздандырылған заттық шынылар,
фильтрленген қи қоймалжың немесе ұнның пішендік тұндырмасы, шыны
таяқша, спирттік жандырғыштар, сіріңке, бактериологиялық бояғыштар
(Лефнер кӛгерткіші немесе фуксин ерітіндісі), стакандағы су, пинцет, фильтр
қағазының жолақтары, бояу үшін пипетка.
Жұмыс тәртібі: Заттық шыныны жайлап қыздырады. Шыны таяқшамен
жоғарыда келтіріген сұйықтықтардың бірін алады және шыны бетін жұқа

36
қабатпен жағады. Жағындыны кептіреді. Сосын оның құрғақ ыстықпен
белгілеуді жүргізеді; заттық шыныны пинцетпен алып, спиртті жандырғыш
үстінен ырғақты шеңберлі қозғалысын 3-5 рет жүргізеді. Бұл кезде
микроағзалар ӛледі де, шыны бетіне берік жабысады. Шыныны суытқаннан
кейін пипеткамен бояғыштан тамшы тамызып, 1-2 минутқа қалдырады, сосын
стакандағы суға пинцетпен ұстап тұрып шыныны шайып алады. Судың артық
мӛлшерін фильтр қағазымен сорып алады және ауада немесе спиртті
жандырғыштың жалынына абайлап кептіреді. Бұдан кейін препарат
иммерсонды жүйе кӛмегімен зерттеуге дайын [121].
Қоректік орталарды дайындау
Ет-пептонды сорпасын (бульон) дайындау. Ет-пептонды сорпаның сұйық
фазасында кӛптеген гетеротрофты микроорганизмдердің дамуы мүмкін.
Сорпаны дайындау үшін, сүйектерден, сіңір және майдан толық тазартылған
сиыр етінен дайындалады. Бір литр суға ет турағыштан ӛткізілген 500 гр ет
салады да, бір тәулікке суыққа тұнуға қалдырады. Тұндырманы дәкеден ӛткізіп,
етті сығып, салқын су моншасына салады. Моншадағы суды қайнатуға
жеткізеді және қайнаған судағы тұндырманы 1/2 сағат ұстайды.
Сонан соң тұндырманы мақтадан тоқылған матадан жасалған шүберек
арқылы фильтрлейді. Сорпаға 5 г ас тұзын және 10 г пептон қосады. Сосын
сорпасын 1 атмосфералық қысым кезінде автоклавта 20 минут ұстап, ерігіш
ақуыздарды тұндырады. Ақуыздардың толық тұнуы және сорпаның толық
мӛлдірлендіру үшін, оларды автоклавқа салу алдында, сорпаға су мӛлшерімен
тең бір тауық жұмыртқасының ақуызын салады. Ақуыздар тұнғаннан кейін
сорпалы қағаз фильтрінде (қатпарлы) сүзеді және индикатор қағазымен рН-ын
тексереді. Әдетте сорпа әлсіз сілтілік ортаға ие және бактерияларды ӛсіруге
жарамды. Сондықтан оны, 10 % натрий бикарбонаты ерітіндісін қосып, әлсіз
сілтілік реакцияға дейін нейтралдайды.
Содан кейін сорпаны колбаға құйып, қағаз қалпағы бар дәкелік тығынмен
жабады және атмосфералық қысымда 20 минут залалсыздандырады. Бірқатар
жағдайларда, толық тұндырғаннан кейін де сорпа мӛлдір болмайды. Онда
коллоидты бӛлшектердің тұнуын, нейтрализацияланғаннан кейін жоғарыда
келтірілген әдіспен сорпа дайындау - ұзақ процесс және еңбек кӛлемдігін қажет
етеді, мұны сорпа кубиктерін қолдану арқылы жеңілдетуге болады. Бұл
жағдайда 1л суда қайнату кезінде 2 сорпа кубигін еріту жеткілікті.
Содан соң сорпаны дәкелік фильтрде сүзеді және кӛрсетілген әдіспен
залалсыздандырады. Сорпаға ас тұзын салмайды, бірақ ортаның рН-ын
тексереді. Тәжірибе нәтижелері, концентрлі сорпа дайындаудың қажет
еместігін кӛрсетті, себебі дақылдарды бұл сорпада ӛсіру нашар жүреді.
Ет-пептонды агар (ЕПА) дайындау. ЕПА әмбебап қатты ортаға ие, ол
шамамен 50-100 % температура кезінде ғана ериді және кез-келген
температуралық жағдайда іс жүзінде жұмыс істеуге мүмкіндік береді. ЕПА
дайындаудың бастапқы ортасы ет-пептонды сорпа болып табылады. Дайын
сорпаға 2 % агар қосып, ашық винттелген және винттелмеген қақпақпен агар
толық ерігенше автоклавта қыздырады. Ыстық орталы 10 %-тік сода

37
ерітіндісімен әлсіз сілтілік реакцияға дейін нейтралдайды. Коллоидты
бӛлшектерді тұндыруды, ет-пептонды сорпа дайындау кезінде келтірілген
әдіспен жүргізеді және автоклавта ыстық күйінде қатпарлы фильтр арқылы
(агар ыстық күйінде шамамен 40-50 0C температурада қатады) фильтрлейді.
Фильтрленген ЕПА атмосфералық қысымда 20 минут бойы
залалсыздандырады.
Ескерту. Фильтрленетін ортаны жоғары температурада ұстап тұрып, ЕПА
фильтрлейді [122].
Қоректік орталарды құю. Әдетте, қоректік орталарда пробиркаға немесе
Петри шынысына керегінше құяды. Агарды ашық вентильді автоклавта немесе
су моншасында балқытқан ыңғайлы. Желатин су моншасында 25-30-50 0С
кезінде қыздырғанда оңай балқиды. Ортаны балқытқаннан соң залалсыздықты
сақтай отырып, қажетті кӛлемдегі ыдысқа құяды. Бұл үшін құю кезінде колба
мойнын спиртті жандырғыш жалынынан жүргізіп дәкелі тығынды, құюдан
кейін күйдіреді. Жұмыс кезінде тығынды аты жоқ саусақ пен шынашақ
арасында ұстап тұрады. Пробирканы ортасымен (орта сипатына байланысты)
құйғаннан кейін тӛменде келтірілген әдістердің бірімен залалсыздандырады.
Тіке агар немесе желатин алу үшін, пробирканы штативке бекітеді, ал
айқасқан орта бетін алу үшін, оларды тығынға тимейтіндей етіп тіреуге
еңкейтіп орналастырады. Сұйық ортаны құюда да осы ереже сақталады.
Қоректік орталарға микроорганизмдерді егу әдістері
Егу 3 кезеңнен тұрады: сұйылту дайындау, Петри табақшасына тығыз
қоректік орталарға отырғызу, ӛскен колонияларды санау.
Сұйылту дайындау. Жекеленген колониялар алу үшін микроорганизмдер
бар дақылды немесе материалды сұйылтады. Сұйылтуды стерилді құбыр
суында, сұйылтудың тұрақты коэффицентін пайдалана отырып дайындайды,
әдетте бұл коэффицент 10 санына тең. Сұйылту дайындау үшін стерилді құбыр
суын стерилді құрғақ пробиркаларға 9 мл-ден құяды. Одан кейін стерилді
пипетканың кӛмегімен алынған бастапқы суспензияның 1 мл-ін 9 мл стерилді
суы бар пробиркаға құяды, бұл бірінші сұйылту, 1:10. Бірінші сұйылтудан
алынған суспензияны жаңа стерилді пипеткамен араластырады. Осы
пипеткамен алынған суспензияның 1 мл алып, екінші пробиркаға кӛшіреді, бұл
екінші сұйылту, 1:100. Осылайша қалған сұйылтуларды дайындайды.
Петри табақшасына егу. Суспензияны беттік және тереңдік әдістермен
егуге болады. Беттік әдіспен егу алдында стерилді Петри табақшаларға
ерітілген қоректік ортаны 20-30 мл-ден құяды. Егуді белгілі сұйылтудан
жүргізеді. Стерилді пипеткамен сәйкес сұйылтудың белгілі кӛлемін 0,1 мл
енгізеді. Суспензияны қоректік орта бетіне Дригальский шпателінің кӛмегімен
жаяды. Табақшаларды термостатқа 28-30 0С температурасында бірнеше күнге
қалдырады.
Зерттелетін суспензияның белгілі кӛлемін қатты қоректік ортаға егу
арқылы ӛсіп шыққан колониялардың саны негізінде бастапқы құрамында қанша
микроорганизм клеткалары болғандығын айтуға мүмкіншілік береді. 100 мл

38
стерилді құбыр суына 10 г топырақ нұсқасын салып, тӛмендегідей сызба-нұсқа
бойынша сұйылту жүргізеді (сурет 3):
Сурет 3- Микроорганизмдердің суспензиясынан сұйылту дайындаудың және
егудің сызба-нұсқасы
Өскен колонияларды санау. Ӛсу жылдамдығына байланысты Петри
табақшасында ӛскен колониялардың санын дақылдаудың 1-5 тәулігінен кейін
колониялардың орташа санын анықтайды да формула бойынша 1 мл-дегі
клеткалар санын Мак-Креди кестесімен (формула 1) есептейді [123].
v
аМ
n10 (1)
мұндағы, М - 1мл-дегі микроорганизм клеткаларының саны; а - Петри
табақшасындағы микроорганизмдер колониясының орташа саны; 10 - сұйылту
коэффициенті; n - егу жүргізілген сұйылтудың реттік саны; v - егуге алынған
суспензияның кӛлемі.
Топырақта әртүрлі топ ӛкілдері тіршілік етеді. Топырақ микрофлорасын
зерттеу үшін Кох әдісі бойынша сұйылту жүргізіледі.
Жасанды қоректік ортаға себу әдісімен топырақтағы
микроорганизмдер санын анықтау
Топырақ суспензиясын дайындау. Бұл үшін ішінде зарарсыздандырылған 3
мл суы бар бірнеше пробиркаларды, ішінде 90 мл зарарсыздандырылған суы
бар колбаны және сыйымдылығы 200 мл болатын зарасыздандырылған, құрғақ
екінші колбаны дайындайды.
Зерттелетін топырақты зарарсыздандырылған әйнек бетіне салады.
Әйнекті алдын ала спиртпен сүртіп, спиртовка жалынына қарып алады.
Топырақты шпатилмен жақсылап араластырады, механикалық қоспалардан
тазартады. Зарарсыздандыру ережесін сақтай отырып, әйнек бетіне 15 гр

39
топырақ салады. Одан соң оны зарарсыздандырылған кордон тостағаншаға 10 г
топырақты ӛлшеп салады. Бұған алғашқы колбадан зарарсыздандырылған
судан 1-2 мл су кұйып, 5 минут езеді. Тостағаншада езілген топырақты, екінші
құрғақ колбаға салады. Сонда 1:10-ға тең бірінші сұйылтуды алады. Колбадағы
топырақ суспензиясын 5 минут бойына шайқап, 30 минуттай тұндырады да,
зарарсыздандырылған суы бар 1-ші пробиркаға құяды. Сонда 1:100 сұйылту
алынады. Дәл осылайша бірнеше сұйылтуды – 1:100, 1:1000, 1:10000, 1:
1000000 жүргізеді. Сұйылту дәрежесі топырақтағы микроағзалар санына,
топырақтың типіне, генетикалық горизонтқа, жыл маусымына, топырақ
үлгісінің ылғалына байланысты болады. Әрбір жаңа сұйылтуды атқару үшін
жаңа зарарсыздандырылған пипетканы қолданады. Бактерияларды бӛліп алу
және санын есептеу үшін әр түрлі сұйылтылған пробиркадағы топырақ
суспензиясын ЕПА, КАА (крахмалды амиакты агар) агарлы топырақ Әшби
орталығына себеді.
Саңырауқұлақтар мен ашытқыларды бояу және санау үшін СА
(суслоагарды) немесе түрлі ӛзгертілген Чапек ортасын пайдаланады.
Су машинасында балқытылған қоректік ортаны, зарасыздандыру ережесін
сақтай отырып, Петри қабықшысына қалыңдығы 0,5-0,8 см етіп құяды. Әрбір
сұйылтуға осындай Петри табақшасының 3-5 даярлайды. Орта қатқаннан соң
оның жабынынан судың тамшыларын жоғалту үшін Петри табақшасын 60-700
С температурасы бар термостатта кептіреді. Жабын әйнекке топырақ нӛмерін,
себу мерзімін, сұйылту дәрежесін және тәжірбиенің қайталау санын жазып
қояды.
Топырақ микроорганизмдерін бӛліп алу үшін, топырақ суспензиясын
қоректік ортаның бетіне себеді. Алдымен пипеткамен бір мл сұйылтылған
суспензия алынады. Әр сұйылтуды қоректік ортаға енгізген алдында
зарарсыздандырылған жаңа пипетка алынуы тиіс. Залалсыздандырылған
пипеткамен тиісті сұйылтудан 1 мл топырақ суспензиясын алып, Петри
табақшасындағы қатты қоректік ортаның дәлдеп ортасынан құяды.
Шпательмен осы тамшыны агар бетіне жақсылап жояды. Топырақ
суспензиясын себу үшін әдетте 0,1- 0,2 мл микропипеткаларды қолданады.
Себілген Петри табақшасын қақпағын тӛмен қаратып, 28-37 0С термостатқа
қояды. Петри табақшасындағы бактериялар колониясын 3-5, саңырауқұлақтар
мен ашытқыларды 5-7, актиномицеттер 7-15 тәуліктен кейін санайды. Егерде
Петри табақшасынан 50-200 колония бактерия мен актиномицеттер, 30-50
колония саңырауқұлақтар табылса, нақты нәтиже деп есептейді.
Петри табақшасындағы колонияларын санау.
Бұл үшін Петри табақшасының табанын тұщы немесе сиямен бір неше
сектор немесе сегментке бӛледі. Егер де қоректік орта мӛлдір болса,
колонияларды табақша астынан түзіп тұратын жарық сәулесімен санайды.
Саналған колонияларды сиямен нүкте салу арқылы белгілеп қояды. Ӛте майда
колонияларды үлкейтетін шыны – лупамен санайды. Тиісті сұйылтудан
себілген Петри табақшасындағы микроағзалар колониясын есептеп,

40
табақшадағы колониялар санының орта кӛрсеткішін анықтайды (формула 2),
оны 1г құрғақ топыраққа шағады:
А = (б× в × г) / Д (2)
Бұнда а – 1г құрғақ топырақтағы микроағзалар жасушаларының саны. б –
Петри табақшасындағы микроағзалар колониясының орта саны. в – тиісті
сұйылту. г – пипеткадағы 1 мл сұйықтағы тамшы саны, д - анализге алынған
топырақ салмағы. Осымен қатар бұл анализ кезінде топырақ ылғалдығын
анықтайды [124].
2.3.2 Энтомопатогенді микроорганизмдерді топырақтан және
бунақденелерден бӛліп алу
Топырақ-кристалтүзуші бактериялар үшін табиғи орта, бұл ортада олар
кӛбейеді, қолайсыз жағдай туындағанда ӛздерін сақтап қалады, сонымен бірге
жәндіктер осы бактерияларды тасмалдаушы болып табылады, дегенмен, бұл
бактерияларға қолайлы жағдай туындаса, иесінде ауру тудырып иесінің ӛлуіне
алып келеді. Bacillus thuringiensis басқа табиғи субстраттардан бӛліп алу
қарастырылмаған.Топырақтың сынамасын топырақ бурымен алады. Пышақпен
топырақты іріктеп алады немесе спиртпен сүртіп тазаланған күрекпен алу
қажет. Зерттелетін аумақты 16 шаршы метрден артық болмаған мӛлшердегі
телімдерге бӛледі. Bac.thuringiensis табылған аумақтан 15 см-ге дейінгі
тереңдіктен топырақ сынамасын алады. Алдымен топырақтың 2-3 сантиметр
жоғарғы қабатын алады, кейін 2 метрге дейінгі тереңдіктен әр 25 сантиметрден
сынамалар алады. Топарақ сынамаларынан кристалл түзуші бактерияларды
бӛліп алу үшін арнайы А қоректік ортасы пайдаланылады. құрамы: пептон 0,7
%, балық гидролизаты 0,4 %, NaCl 0,65 %, агар-агар 1,5 %.
Топырақтан дайындалған суспензияны 70 C0
температураға жеткізіп
қыздырымыз. Петри табақшасындағы арнайы А қоректік ортасына 0,25 мг
суспензия құйып, шпательмен жаймалап беттік егу жүргізіледі. Шпательдегі
жұғындыдан екінші және үшінші петри табақшаларына қайталап егу
жүргізіледі. Термостатта 5-6 тәулік 28 0С ӛсіру жүргізіледі.
Ӛскен колонияларды морфологиялық сипаттау үшін қолданбалы заттық
шыныға жұғынды препарат жасап, микроскопта (х 900-1000), иммерсия
майының астында бақылау жүргізіледі. Мұнда мақсат Bt колониясынан спора
және кристалдарды анықтау.
Бунақденелерден бактерияларды таза ортаға бӛліп алуда бӛгде микрофлора
түсіп кетпес үшін жәндіктің денесін залалсыздандырамыз. Объектінің беткі
қабатын 96 % спиртпен немесе сулема ерітіндісімен (1:1000) стерильді сумен
және 2 %-дық карболол суымен шайып, эфирге немесе судың асқын тотығына
батырады. Бұл ерітінділермен ӛңдеу уақыты 1-5 минут. Ірі жұлдызқұрттарды
спиртке батырып, спирт шамының алауынан ӛткізеді. Кепкен материалмен
жұмыс жасайтын болсақ, материалды стерильді сумен сулап алады.
Жұғындыны х 400-1000 есе үлкейтіп қарайды. Бактериоз болған жағдайда
жұғындыда бір-бірінен пішіні жағынан әртүрлі кӛп мӛлшерде бактерия болады.
Қажет болған жағдайда жұғындыны бояп қарайды. Стерильденген жәндіктерді

41
стерильді суы бар пробиркаларға салып, стерильді шыны таяқшамен біртекті
гомогенді суспензия алғанша ұнтақтайды. Оқшауланған колонияларды алу
үшін кезекті сұйылтулар дайындалады. Сұйылтуды стерильденген құбыр
суымен немесе физиологиялық ерітінді (0,65 %-тік Nacl ерітіндісі) кӛмегімен
жасайды. Сұйылтуға арналған суды дайындау үшін стерильденген құрғақ
пробиркаларға құбыр суынан 9 мл-ден ӛлшеп құяды. Содан соң зерттелетін
суспензиядан стерильді пипеткамен 1 мл алып, 9 мл суы бар пробиркаға құяды.
Бұл – бірінші сұйылту, яғни 10-1
. Алынған сұйылтуды жаңа стерильді
пипеткамен мұқият араластырады. Осы пипеткамен сол суспензиядан 1 мл
алып екінші пробиркаға құяды. Бұл екінші сұйылту, яғни 10-2
. Сұйылту саны
зерттелетін микроорганизм популяциясының тығыздығына байланысты
болады. Тығыздығы жоғары болған сайын, сұйылту саны да кӛп болады. Әр
сұйылтудан 0,05 мл алып қоректік ортасы бар Петри табақшаларының бетіне
сеуіп стерилденген шпательмен жаймалап егіп шығамызда Петри
табақшаларын тӛнкеріп, термостатқа 28-30 0С-қа 4-5 тәулікке қоямызда 2
тәуліктен бастап колониялардың ӛсуіп қарап, оларға сипаттама береміз. Мұнда
колонияның әр тәулік сайын ӛскен диаметрі есептелініп отырады. 3 тәулікте
колонияларға бекітілген препарат жасап, бірнеше бояумен бояп, олардың
вегетатаивті жасушаларын қарауға болады [125]
2.3.3 Bacillus thuringiensis энтомопотогенді бактерияларының дақылдық-
морфологиялық, биохимиялық қасиеттерін анықтау
Bacillus thuringiensis бактериясының морфологиялық белгілерін анықтау.
Bt колониялары ақ немесе ақшыл сұр түсті. Жоғарғы беті түйіршікті,
бұдыр немесе тегіс болуы мүмкін. Колония жиектері (шеттері) әдетте тісті
немесе қалақты, анда-санда толқынды немесе тегіс болып келеді. Қырынан
қарағандағы пішіні тегіс. Bt бактериясы түрлерінің жалпы ерекшеліктері бар.
Ол дара немесе алқалы, ұзындығы 6-8 мкм қылдарының арқасында
жылжымалы болып келетін 3-6х0,8-1,3 мкм мӛлшерлі дұрыс грамды
таяқшалары. Таяқшалар бір тәулік ӛсірілгеннен кейін споралар түзіледі.
Таяқшаның бір ұшынан эндоспора, ал қарама қарсы бӛлігінен ақуызды кристал
түзіледі. Спорангийден кристалдардың толық бӛртуі дақылдарды 28 °С-та
инкубациялаудың 5-6 тәулігінде іске асады. Споруляция барысында
кристалдың қалыптасуы Bt бактериясын B.cereus тобының қалған барлық
ӛкілдерінен ажырататын белгі болып табылады.
Микроорганизмдердің дақылдық ерекшеліктерін қатты қоректік орталарда
ӛсу белгілерімен анықтады. Әрбір микроораганизмге дақылдық қасиеттер тән
болғандықтан, маңызды диагностикалық белгі болып табылады. Қатты
қоректік ортаның бетінде микроорганизмдерді егу түріне қарай колония,
штрих, жаймаланып ӛсуі мүмкін. Колония деп бір клеткадан ӛсіп шыққан, бір
түрге жататын клеткаларды айтады.
Қоректік орталар микроорганизмдердің таза дақылын бӛліп алуда,
дақылдық қасиеттерін зерттеуде маңызы зор. Бактериологиялық жұмыстардың,
зерттеу нәтижелерінің сапалы анықталуына негіз болып табылады [126].

42
Бактериялардың дақылдық-мофологиялық қасиеттері «А» қоректік
ортасында зерттелді. «А» қоректік ортасының құрамы келесідей:
1.Пептон- 1 %;
2. Балық гидролизаты – 0,4 %;
3. NaCl – 0,5 %;
4. Агар – 1,5-2,0 %;
5. H2O – 100 мл.
Беттік колониялардың түзілуі қатты субстратта кӛпшілік
микроорганизмдердің ӛсу ерекшеліктері. Мұндай колониялар әртүрлілігімен
ерекшеленеді. Оларды сипаттауға келесі белгілерді есепке алады:
– пішіні – дӛңгеленген, амеба тәрізді, бұрыс пішінді, ризоидты және т.б.;
– ӛлшемі – диаметрі 1-8 мм, нүктелік колониялар –1 мм-ден кем;
– беті – тегіс, кедір-бұдыр, қыртысталған, әжімді;
– оптикалық ерекшелігі жылтыр, мӛлдір, жартылай мӛлдір, мӛлдір емес;
– түсі – түссіз, ақ, сары, алтын түстес, қызыл, қоңыр, қара және т.б;
– кӛлденең кӛрінісі – тығыз, тӛмпек, кратер тәрізді, агарға ене ӛскен;
– колониялардың шеті – тегіс, толқын тәрізді, ризоидты;
– консистенциясы – майлы, қамыр тәрізді, жұмсақ немесе қатты.
Bacillus thuringiensis дақылдарының морфологиясын анықтау үшін бояу
әдістері. Циль бойынша карболды фуксинмен бояу.
Бояуды дайындау. Ұнтақтағышта 10 мл ректификат спиртімен үгітілген 1г
негізгі фуксинге 5 г фенол қосады (карбол қышқылы). Қоспаны жақсылап
ұнтақтап 100 мл дистилденген су қосып, ұдайы араластырып отырады.
Ерітіндіні қағаз фильтр арқылы сүзіп алады. Бір тәуліктен соң пайдалануға
жарамды. Жұмыс алдында 10 рет дистилденген сумен араластырады. Бұл
ерітінді қараңғы шыныда немесе жабық пробиркада ұзақ уақыт сақталады.
Бояу техникасы: заттық шыныдағы кепкен жағындының бетіне карболды
фуксин ерітіндісін тамызып, 1-2 мин ұстайды. Содан кейін ерітіндіні тӛгіп,
жағындыны құбыр суымен шаяды. Нәтижесінде кристаллдар мен жасушалар
қызыл түске боялады, ал споралар боялмайды.
Эозинмен бояу:
1) Карбол қышқылы– 5 г;
2) Эозин– 2 г;
3) Дистилденген су – 100 мл.
Бояу дайындалған соң бір тәуліктен кейін қолдануға жарамды. Бекітілген
жағындыға заттық шыныны жабатындай етіп пипеткамен бояу жағады. 2-3 мин
аралығында бояйды. Содан соң бояуды қараңғы шынысы бар ыдысқа тӛгеді.
Заттық шыныдағы жағындыны дистилденген сумен шайып, кептіреді.
Кристаллдары таңқурай түске боялады, ал споралар боялмайды, тек жиектері
қызыл болады.
Пешков әдісі бойынша бояу. Заттық шыны бетіне дистилденген су
тамызып бетіне тұзақпен бактерия дақылын жағады. Ауада кептіріп, спирт шам
жалынынан ӛткізіп бекітеді. Сосын бетіне Леффлер бойынша метилен кӛгін
тамызып, 3 мин бетінде буы шыққанша спирт шам жалынында ұстайды. Содан

43
соң сумен шайып, үстіне сафронин қызылын тамызып 1 мин ұстап, сумен
шайып, иммерсиялық май тамызып қарайды [127].
Дақылдар себілген табақшалар термостатта 30 °C та 5-6 тәулік бойы
инкубацияланды, кейіннен ӛсіп шыққан микрофлораларға микроскопиялық
талдау жүргізілді. Ол үшін энтомопатогенді бактерия штаммдарының
колониясынан заттық әйнекке жағынды алынды.
Жағынды дайындау. Майсыздандырылған таза заттық әйнекке құбыр
суынан тамшы тамыздық. Бактериологиялық ілмектің кӛмегіме
микроорганизмдер дақылы бар түтік шыныдан (пробиркадан) микробты
массаның аз мӛлшері алынып, тамшыға енгізілді. Ілмекпен тамшыны әйнек
бетіне мұқият жағамыз, шамамен 4 см2. Қою суспензияны сумен араластырып,
сосын жағындыны бӛлме температурасында кептіреміз: әдетте бӛлме
температурасындағы ауада немесе препаратты оттық жалына жоғары ұстап,
әлсіз қыздыру арқылы. Кептірілген препаратты оттық жалынының үстінде
ұстап бекітеміз (фиксаждаймыз): препаратты сырт жағынан 3-4 рет ақырын
жалынның үстінен ӛткізіп аламыз.
Жағындыны бояу барысында оған бірнеше тамшы бояғыш зат тамызамыз.
Бояғыш затқа байланысты боялу ұзақтығы әрқалай (1-5 минут) болады.
Бояғаннан кейін препаратты сумен жаямыз, сорғыш қағазбен құрғатамыз және
ауада кептіреміз.
Эозин қосылған бояғыш құрамы: карбол қышқылы – 5 г; эозин – 2 г;
дистилденген (тазартылған) су – 100 мл. Бояғыш дайындалғаннан кейінгі бір
тәуліктен соң қолдануға жарамды. Бекітілген жағындыға тамызғыштың
кӛмегімен (пипеткамен) заттық әйнектің бетін жабатындай етіп бояғышты
тамызамыз да 2-3 минутқа қалдырамыз. Сосын бояғышты қайтадан қарақоңыр
әйнекті ыдысқа кері тӛгеміз. Заттық әйнектегі жағындыны дистилденген сумен
шайып, кептіреміз. Кристаллдарды таңқурай түстес етіп бояймыз, споралар
боялмайды, қызыл жиекті болып қалады.
Микроскоп арқылы майлы иммерсиямен (х 900-1000) боялған және
кептірілген жағындыларда споралар, параспоралы денелер (эндотоксин
кристаллдары) және бейтаныс микрофлоралар анықталады. Изоляттарды
микроскопиялық талдау үшін Axispor 40 (Carl Zeiss) сәулелі микроскопы
қолданылды.
Микробтардың дақылдық белгілері олардың қатты қоректік орталарда
ӛсуін сипаттау арқылы анықталды. Дақылдық ерекшелік микробтың әр түріне
тән, сондықтан маңызды диагностикалық белгі болып табылады.
Қоректік орталар зерттеліп отырған материалдан микробтардың таза
дақылдарын бӛліп алу және олардың ерекшеліктерін анықтау үшін қажет,
сонымен қатар, бактериологиялық жұмыстардың негізі болып табылады [128].
Бактериялардың дақылды–морфологиялық ерекшеліктері ЕПА (ет
пептонды агар) қатты қоректік ортасында (етпептонды сорпа (ЕС) 100 мл, агар-
агар 1,5-2,0 %) зерттелді.
Колониялардың ерекшеліктерін анықтау үшін бактериялар Петри
табақшасына құйылған ЕПА-ға себілді. Колониялар келесі белгелері бойынша

44
сипатталды: кӛлемі, сыртқы пішіні, мӛлдірлігі, шетінің сұлбасы бедері, беті,
түсі, құрылымы және консистенциясы.
Споракристалды қоспалардың бунақденелілерге ӛзіндік әсерін бағалау
үшін штаммдар қоректік ортада 6 тәулік бойы 30 °C температурада, спорлар
мен кристалдар толық түзілгенге дейін ӛсірілді.
Bacillus thuringiensis бактериясының биохимиялық қасиетін анықтау
Бактерияның биохимиялық белсенділігі микробты клеткалар ӛндіріп,
қоршаған ортаға шығаратын ферменттердің сипаты мен саны бойынша
анықталды. Микробтарды бақылау үшін кӛмірсулар мен ақуыздарды ыдырауын
белсендіретін сахаролитті және протеолитті ферменттерді анықтау аса маңызды
орын алады.
Сахаролитті ферменттерді табу үшін бактерияның зерттеліп отырған
культурасы (себіндісі) Гисс қоректік ортасына (1 % пептон, 0,5 % NaCl, 0,5 %
кӛміртегі, 1 мл Андраде индикаторы) себілді. Ортада сахарозаға, салицинге,
маннозаға қарым қатынасы сыналды. Инокуляция және инкубация
температурасы 28 °C. Оң реакция ортаның қызаруымен талданды.
Ацетилметилкарбинолды анықтау (АМК) Фогес-Проскуаер тәсілі
бойынша жүргізілді: 28 °C-та глюкозопептонды суда инкубацияланған 3 мл
тӛрт тәуліктік культураға 3,0 мл 20 % -ды ащы натрий қосылды. Шыны
түтікшелер қарқынды сілкіленді және екі сағат бойы тік қалпында қалдырылды.
Оң реакция орта бетінің қызғылт түске боялуымен белгіленді.
Лецитовителлинді реакция (ЛВР) Бехер әдісімен жүргізілді: 10 %-ды
жұмыртқа сарысының эмульсиясы физиологиялық ерітіндіде дайындалды.
Қатты қоректік ортада реакция түзілуі үшін 10 мл суспензияны 50 °C- дейін
салқындатылған 90 мл «А» қоректік ортасына қосып, Петри табашасына
құйдық. Себу жұмыстары ине арқылы жүзеге асырылды, реакция 28 сағаттан
соң есептелді, инкубация температурасы 28 °C. Оң реакция колония
айналасындағы агардың ағарған аймақтарымен сипатталды.
Каталаза И.И. Ашмарин бойынша келесі тәсілмен анықталды: заттық
әйнекке қатты ортадан алынған себінді бӛлшегін орналастырып, оған 1-2
тамшы сутек тотығын қосамыз. Оң реакция оттегі кӛпіршіктерінің бӛлінуімен
белгіленді.
Уреазаны анықтау Кристенсен ортасында жүргізілді (0,1 % пептон; 0,5 %
NaCl; 0,2 % калий фосфаты; 0,6 мл 0,2 %-ды фенолрот сулы ерітіндісі (рН 6,8).
Орта негізі автоклавта залалсыздандырылды. 50 °C дейін суытылған ортаға
асептика ережелерін сақтай отырып, 2 % мӛлшерде 20 %-ды несепнәр ерітіндісі
қосылды. Кристенсен ортасы несепнәрмен инокуляцияланып, 28 °C
инкубацияланды. Оң реакция ортаның қызаруымен сипатталды.
Крахмал гидролизі «А» қоректік ортасына 0,2 % ерігіш крахмал қосу
арқылы анықталды. Орта түйреу әдісімен инокуляцияланды. Инкубациядан
кейін агардың үстіне Люголь ерітіндісін тӛгеміз. Оң реакция ӛсу аймағының
айналасында түссіз учаскенің пайда болуымен сипатталды.
Протеолиттік ерекшелік казеинді агарға нүктелі себу барысында
гидролиз аймақтары бойынша анықталды. Оны даярлау үшін «А» ортасына 1/3

45
кӛлемде зарарсыздандырылған майсыздандырылған сүт қосады. Оң реакция
колонияның ӛсу аймағының айналасында түссіз учаскенің пайда болуымен
белгіленді.
2.3.4 Bacillus thuringiensis бактериясының штаммдарының биологиялық
белсенділігін анықтау
Bacillus thuringiensis штаммдарының бунақденелілерге уыттылығын, яғни
биологиялық белсенділігін тексеру бактерия дақылдары қолданылды. Ол үшін
бактерия дақылдарынан сұйылтулар жасалды. Пробиркада қиғаш агарда ӛсіп
тұрған бактерия дақылдарына 4 мл-ден дистилденген су құйылып, жақсылап
араластырылды. Сұйылтулардың титрін анықтау үшін Том Горяев
камерасында споралар саналды. Спора титрі жоғары болды. Бунақденелілерді
залалдауға қажетті спора титрі 1x107болғандықтан, пробиркаларға 8 мл-ден
қайта су қосылды. Дайын болған суспензияларға жапырақтарды малып, 2-3
жасар жұлдызқұрттарға беріп, ауа кіретін тесіктері бар стақандарға салынды.
Әр стақанға 10 жұлдызқұрттан салынды. Әрқайсысы үш қайталаудан жасалды.
Жұлдызқұрттарды күнделікті бақылап, 5-7 күн 25 оС температурада ұстады
(сурет 4). Содан кейін ӛлгендері пайыздық кӛрсеткішпен тіркеледі [129].
Сурет 4- Жұлдызқұрттарды залалдау схемасы
Бунақденелілерді залалдау ауыз қуысы арқылы жүргізілді. Ол үшін
азықтық ӛсімдіктер 1х10 8 мӛлшердегі споракристалды суспензиямен ӛңделді.
Тәжірибенің әр нұсқасы 4-5 қайталаумен қойылды. Бір қайталауға – сынақ
бунақденесінің 10 данасы алынды. Олардың ӛлуі күн сайын 5 тәулік бойы

46
тіркелді. Биологиялық тиімділікті есептеу үшін Аббот формуласы (формула 3)
қолданылды:
100
А
БАЭ (3)
мұнда: Э – сынақ нысанының сан мӛлшерінің азаюы пайызбен кӛрсетілген
тиімділік;
А – есеп жүргізілген күні бақылаудағы тірі дарақтар саны;
Б – есеп жүргізілген күні тәжірибедегі тірі дарақтар саны.

47
3 ЗЕРТТЕУ ЖҤРГІЗІЛЕТІН АЙМАҚТАРДЫҢ ТАБИҒИ-КЛИМАТ
ЖАҒДАЙЛАРЫ
3.1 Климат
Климат топырақтың түзілуіне ӛзінің барлық метеорологиялық
кӛрсеткіштерімен әсер етеді. Оларға адамның ӛндірістік әрекетінің әсеріде бар.
Климаттық жағдайлар кез-келген територияның физико-географиялық
үрдістерінің жүруіне шешуші әсер етеді. Климаттық жағдайлар, алуан түрлі
топырақ үрдістерінің бағыты мен қарқындылығына тікелей әсер етуші негізгі
фактор болып табылады.
Іле Алатауы бойынша
Қазақ гидрометеорология ғылыми зерттеу институты ғалымдарының пікірі
бойынша Алматының климаты жерорта теңізінің климат жағдайына ұқсастау,
адам мен жануарлардың және ӛсімдіктердің тіршілік етуіне қолайлы болып
келеді. Жауын-шашын мен температура мӛлшерлері қыс маусымында күзге
қарағанда салыстырмалы түрде құрғақ.
Қарастырылып отырған ауданның климаты континенталды, жергілікті
биіктіктің тӛмендеуінен ауа температурасы жән ауа ылғалдылығы
тапшылығының тӛмендеуі, күн радиациясы мен атмосфера мӛлдірлігінің
ӛсуі байқалады [130]. Аймақтың климаттық сипаттамалары толық және жеке
элементтері бойынша О.М. Челпанов, О.А. Дроздов, Н.Ф. Гельмгольц, А.С.
Утешев және т.б. ғалымдардың еңбектерінде қарастырылған. Іле
Алатауының климаттық жағдайлары [131,132] еңбектерде толығырақ
жалпылай қорытылған.
Іле Алатауының құрлықтың терең ішінде орналасуына байланысты
солтүстік, солтүстік–батыс және батыстан полярлық және арктикалық ауа
массаларының енуіне қолайлы. Аумақта, жалпы, атмосфералық циркуляциямен
қатар, жергілікті таулық-аңғарлық циркуляция (әсіресе мамыр-қыркүйек
айларында) орын алады [133].
Қатты ағындылардың қалыптасуына елеулі әсер етуші негізгі климаттық
сипаттамалар–атмосфералық жауын-шашын, қар жамылғысы, ауа
температурасы мен ылғалыдылық, инсоляция, жел жылдамығы тӛменде
1 және 2 кестелерде берілген. Климаттық сипаттамалардың кестедегі сандық
мәндері [134, 135] жұмыстарынан келтірілген.
Іле Алатауының ауа температурасы ӛзгеруінің негізгі заңдылығы болып
оның мәндерінің жергілікті жер биіктігінің ӛзгеруіне байланысты азаюы,
сонымен бірге қысқы уақытта 1400-1700 метр биіктікке дейін температура
инверсиясының байқалуы табылады. Сонымен қатар ауа температурасының
экспозициялық айырмашылығы ӛте айқын байқалады: оңтүстік экспозиция
беткейлеріне ауа температурасының анағұрлым жоғарғы мәндері ие, одан
тӛменірек ауа температурасы батыс экспозиция беткейлерінде және ең
тӛменгі ауа температурасы солтүтік экспозиция беткейлеріне тән болып
табылады [136].
Ауа температурасын ӛте айқын ӛзгергіштік пен айтарлықтай
үлкен жылдық амплитудалардың байқалуымен сипаттауға болады. Орташа

48
жылдық ауа температурасы Іле Алатауында жазықтықта және аласа
таулы-тауалды аймақтарда 8-9,3 0С ал 3017 метрде (Мыңжылқы
метеобекеті) жоғары таулы аймақта ―минус‖ 2 0С, 3750 м деңгейінде
«минус» (6-6,5) °С [135] (кесте 1) дейін жетеді. Орташа жылдық температура
нормалар мәліметтерін салыстырғанда, ғаламдық жылыну қарастырылып
отырған ауданның ауа температурасына әсер еткендігін кӛруге болады.
Түрлі бекеттердегі орташа жылдық ауа температурасы 0°С тан 0,5 °С дейін
кӛтерілген, орташа кӛтерілу 0,23 °С құрады.
Кесте 1- Орташа айлық және орташа жылдық ауа температурасы (°С) [104]
Станция
Биіктігі, м
Айлар
Жыл
I
II
III
IV
V
VI
VII
VIII
IX
X
XI
XII
Алматы,ОГ
М С
847
-6,4
-4,7
2,1
10,8
16,3
20,7
23,4
22,5
17,2
9,5
1,2
-4,2
9,0
Усть-
Горельни
к
1913
-6,2
-5,5
-2,1
3,7
8,5
12,5
15,1
14,2
9,7
4,3
-1,2
-4,1
4,2
Мыңжылқы
3017
-9,6
-6,7
-7,1
-2,1
1,8
5,4
7,8
7,5
3,7
-5,4 -6,6
-9,4
-1,8
Ү. Алматы
2516
-8,8
-8,2
-4,5
1,2
5,2
9,3
11,5
10,8
7,1
1,8
-3,6
-6,6
1,2
Есік
1098
-4,6
-4,2
1,5
9,7
14,8
19,4
22,3
20,8
15,9
8,5
1,2
-2,7
8,5
Каменское
1317
-3,1
-3,2
1,3
8,7
13,6
18,5
21,2
20,1
15,2
8,610
3,1
-0,2
8,5 Шелек
606
-7,2
-4,7
3,4
12,2
17,5
22,2
24,5
23,2
17,5
1,5
-4,5
9,7
Ұзынағаш
814
-8,8
-7,2
0,6
10,2
15,4
20,2
22,9
21,4
15,5
7,8
0,2
-5,4
7,8
Атмосфералық жауын-шашын. Іле Алатауының солтүстік беткейі басқа
да тау массивтерінің шеттік аймақ беткейлері сияқты едәуір ылғалдылықпен
және жергілікті жер биіктігінің ӛсуімен жауын-шашын шамасының едәуір
жоғарылауымен сипатталады (кесте 2). Оны ғалымдар әдетте атмосфералық
фронттардың тауға жақын жерде шиелінісуімен, жылдың жылы мерзімінде
конвекциялық бұлттанудың туындауымен, сонымен қатар арктикалық,
полярлық және тропиктік ауа массаларының әсеріне ұшырауымен түсіндіреді
[137]. Жауын-шашын мӛлшері жота ұзындығы бойынша әртүрлі. Іле
Алатауында жергілікті жердің биіктігінен жауын-шашынның жылдық
суммасының ӛзгеру сипаты жайлы әртүрлі кӛзқарастар бар [138, 139, 140,141].
Қазіргі уақытта жауын-шашын мӛлшері биіктіктің 3500-3600м-ге дейін
үздіксіз ӛседі, одан жоғарылағанда тӛмендейді [137]. Жотаның орталық
бӛлігінде орналасқан Кіші Алматы ӛзені алабында жауын-шашын 847 м абс.
биіктікте 600 мм-ден (Алматы, ГМО), ал 3600 м абс биіктік 1260 мм-ге
дейін ӛзгереді, одан ары жауын-шашын мӛлшері азаяды. Жауын-шашын
мӛлшері жотаның орталық бӛлігінен шығыс пен батысқа қарай азаяды,
ылғалдың кӛп бӛлігі жотаның батыс және орталық бӛлігінде
қалатындықтан, шығыс бӛлігі батысқа қарағанда аз ылғалданған.
Іле Алатауына жауын-шашынның кӛктемгі мӛлшері күздікінен жоғары.
Жауын-шашынның екі максимумы және бір минимумы қыста байқалады.
«Қазгидромет» мәліметтері бойынша Іле Алатауындағы жауын-шашынның
айлық, маусымдық және жылдық мӛлшерінің 1891 – 2000 жж. аралығында

49
есептелген мәндері (кестеде 3) келтірілген [142].
Кесте 2 - Орташа айлық, маусымдық және жылдық жауын-шашын мӛлшері,мм
Метеобекет
Айлар
Жыл
XI-III
IV-X
I
II
III
IV
V
VI
VII
VII
IX
X
XI
XII
Шелек
17
16
20
28
33
25
19
11
12
23
26
22
252
101
151
Есік
34
37
71
11
14
10
15
61
38
28
32
60
61
44
673
246
427
Алматы (Кам.плато)
37
45
86
88
57
42
43
73
65
44
873
278
595
Алматы, ОГМС
28
35
67
98
11
59
38
27
27
54
53
35
624
221
403
Ұзынагаш
Усть-Горельник
24
28
28
32
42
67
69
11
71
18
47
13
35
98
23
63
20
42
35
55
35
50
29
32
458
902
158
209
300
693
Ү.Алматы
22
27
55
98
15
12
11
72
43
46
42
33
819
179
640
Мыңжылқы
21
23
49
95
15
15
12
93
47
47
40
27
883
160
723
Жетісу Алатауы бойынша
Жетісу Алатауы Іле Алатауы сияқты дүниежүзілік мұхиттан шалғай
орналасқан. Оның климатына Арктиканың суық ауасы, Тұран ойпатының
ыстық ауасы және Сібір антициклоны әсер етеді. Тау етегіндегі шӛлейттер мен
шӛлдерге континентті климат тән. Биік таулардың беткейлерінде климат
сипаты қоңыржай: қыс суық, жаз қоңырсалқын. Қаңтардың орташа
температурасы –-3,1°-3,6 °С аралығында, шілдеде +22,8°+23,3 °С (кесте-2).
Жетісу қақпасы арқылы күшті Ебі желі соғады. Ол Жетісу Алатауындағы
Барлықтау мен Майлытау жоталарынан келетін циклон мен Жетісу қақпасының
оңтүстік-шығысы арқылы ӛтетін антициклонның соқтығысуынан пайда болады.
Ебі желі жылы, оның жылдамдығы 60-80 м/с дейін жетеді. Кӛбінесе 1-2, кейде
тіпті 3-7 тәулік бойы соғады. Осы қақпа арқылы Сайқан тауларынан соғатын
суық сайқан желі де ӛтеді.
Жауын-шашын мӛлшері жыл мезгілдері бойыншада ӛзгеріп отырады(кесте
3). Орташа жылдық атмосфералық жауын-шашын мӛлшері бұл аймақта онша
кӛп емес 35-217 мм. Жаз айларында ауаның салыстырмалы ылғалдары ӛте
тӛмен, бар болғаны 25-29 %. Тұрақты қар жамылғысы желтоқсан айының
бірінші жартысында қалыптасады. жауын-шашынның түсу мӛлшерінің
максимумы кӛктемге тән (сурет 5). Жауын-шашынның орташа жылдық
мӛлшері 588- 600 мм. Қыста қар қалың жауады, ал тау шындары мұздықтармен
кӛмкерілген.
Кесте 3 - Орташа жылдық жауын-шашынның түсу мӛлшері және оның жыл
мезгілі бойынша бӛлінісі Метеостанция
Биіктігі
Қыс Кӛктем Жаз күз жылдық
М
м
4
%
м
м
7
%
м
м
%
%
м
м
%
%
мм
Қапал 1239 3
45
1
14
2
217
3
36
2
203
3
31
1
106
1
20
600
Қос-ағаш 1570 1
102
1
17
1
174
3
30
1
139
2
27
1
153
2
26
588

50
Сурет 5 - Орташа жылдық жауын-шашынның жыл мезгілі бойынша түсу
мӛлшері
Зерттелініп отырған аймақта ауаның жылдық орташа температурасы.
Оның мӛлшері оңтүстіктен солтүстікке қарай арта түседі. Теріс температуралар
негізінен желтоқсан, қаңтар және ақпан айларына тән. Ең суық ай-қаңтар,
ақпан. қаңтар айында -3,1-3,6 0С. Орташа тәуліктік температурасының +10
0С
жоғары болатын күндердің саны 142-181. Күздің бірінші үсіктері қазан айының
бірінші жартысында, ал кӛктемнің соңғысы-сәуірдің аяғында болуы мүмкін.
Орташажылдық ауа температурасы бұл аймақта 6,6-7,6 0С аралығында. Ең
ыстық ай шілде температурасы 23,1-23,3 0С (сурет 6).
Ауа температурасы сәуірде күрт кӛтеріліп, қазанда күрт түседі. Сәуірдің
ортасы мен аяғында температура 10 0Сжоғары болып ол 65-170 күнге созылады.
Осы кезеңнің тиімді температуралар жиынтығы 2660-2790 0С. Соңғы үсіктер
сәуірдің аяғында немесе мамырдың басында болуы мүмкін, ал біріншілері
қыркүйектің үшінші декадасында, немесе қазанның басында (кесте-4).
Кесте 4- Саркан қаласының Айлық орташа температурасы, ºС (2013-2015жж)
Айлар Ауаңың жылдық орташа температурасы, ºС
2013ж 2014ж 2015ж
Қаңтар -3,6 -3,1 -3,2
Ақпан -3,2 -3,8 -3,1
Наурыз 1,4 1,2 1,3
Сәуір 12,9 8,6 11,5
Мамыр 15,4 17 16,8
Маусым 20,2 24 22,3
Шілде 23,1 23,3 22,8
Тамыз 22,6 22,8 22,2
Қыркүйек 16,7 14,5 14,1
Қазан 7,9 7,6 -
Қараша 2,8 2,4 -
Желтоқсан -1,6 -1,8 -

51
Сурет 6 - Саркан қаласының айлық орташа температурасы, ºС (2013-
2015жж)
3.2 Жер бедері және топыраққҧраушы жыныстар
Жер бедерін түзуші ӛзара қарама-қарсы бағытта әрекеттесетін
эндогенді және экзогенді үрдістердің әсерінен болады. Сыртқы күштер жер
қабығын гравитациялық тепе-теңдікке алып келсе, ішкі күштер
ауырлық күшіне қарамастан, оны жүйелі түрде бұзып отырады.
Бұндай күрделі ӛзара әрекеттесудің нәтижесінде жер беті мүсіндері үнемі
ӛзгеріп, түрлі кедір-бұдырлықтар туындайды. Басқаша айтқанда, бұл
күштердің әрекеттесуі жер бедерінің қалыптасуы мен даму үдерістерін
анықтайды. Жер бедерінің кез келген қайта қалыптасуы қашанда оның
алғашқы материалы ретінде қарастырылатын тау жыныстарының бұзылуы
мен орын ауысуынан басталады. Сәйкесінше, аймақтың геологиялық
құрылымын, тау жыныстарының ерекшеліктері мен қалыптасу
жағдайларын, сонымен қатар олардың қозғалу заңдылықтары жайлы білім –
жер беті қабатының қалыптасу заңдылықтарын түсіну үшін шешуші мәнге ие
[143].
Іле Алатауы — Іле Алатауына аумақтың 75 % алып жатқан гранит,
порфир, порфирит және тағы басқа интрузивтік тау жыныстарының кең
таралуы тән. Аймақтың таулы бӛлігінде шығу тегі магмалық болып
табылатын палеозойлық кристалды жыныстар кӛптеп таралса, тауалды
бӛлігінде қоңыр-қызғылт саздар және суӛткізгіштігінің тӛмендігімен
сипатталатын құмбалшықты қуатты шӛгінділері кең таралған.
Аңғарлардың елеулі бӛліктері жұмыр тасты-малтатасты тӛрттік шӛгінді
жамылғыларынан тұрады [144].

52
Солтүстік Хан тәңірі таудың орографиялық құрылымында ұзын, массивті
және биік тау жоталары – Іле Алатауы, Күнгей-Алатау және т.б. айқын
байқалады Іле Алатауы кӛптеген жоталар сияқты ендік бағытта созылып
жатыр. Батысында биік емес қыратты (1500 м дейін) кең үстірттерді (Қордай,
Арғайты) бейнелейтін аласа таулы қыраттан басталатын жота Жетіжол
жотасы мен Кіндіктас тауларынан Шоқпар және Қопа ойысымен бӛлінген
[131].
Іле Алатауының табиғи шекаралары: шығысында – Шарын ӛзені,
оңтүстікте – Шілік және Шоңкемін ӛзендері, ал солтүстік бӛлігі тауалды
жазық жолағымен шектескен [145].
Ең жоғарғы абсолюттік биіктіктері орталық бӛлігінде орын алған
жотаның, жоғарғы нүктесі – Талғар шыңы (4973 м.абс).
Талғар шыңынан шығысқа қарай Іле Алатауы Аманжол ӛткеліне дейін
ендік бағытта созылады. Бұл жерде оның биіктігі біршама тӛмендейді
(4000-3500 м-ге дейін). Аманжолдан шығысқа қарай Шілік ӛзеніне дейін
ең биік нүктелері 3000-3200 м-ге дейін жететін Сарытау сілеміне ұштасқан.
Шілік және Шарын ӛзендерінің аралығында жотаның ең тӛмен сілемдері –
биіктіктері 2300 м-ге жететін Сӛгеті мен Торыайғыр таулары орналасқан.
Іле Алатауының жалпы ұзындығы 250 шақырымға жуық,
оныңи шамамен 150 шақырымын орташа биіктігі шамамен 4000 м болатын
тік болып келетін қырқалар алып жатыр.
Іле Алатуы жер бедерін биік таулы-мұздықты, ортатаулы және
аласатаулы-тауалы деп бӛлуге болады [146,147]. Тауларда гидрологиялық
элементтердің биіктік белдеулігін айқындайтын ең басты фактор жер бедері
болып табылады. Биіктік белдеулерінің ауысуымен шайылу жағдайлары
да ӛзгеріске ұшырайды.
Іле Алатауында биіктік белдеулік айқын байқалады.
Биік таулы белдеу (2800-3000 метрден жоғары). Бұл тау аумағының
шамамен 40 % алып жатыр, негізінде жотаның ӛсі бойында орналасқан.
Белдеудің жер бедері альпілік, терең тілімденумен ерекшеленеді. Солтүстік
беткейлерде мұз бен қар, ал оңтүстікте – жартас пен сусымалар жатыр
[146,11-18 б]. Биік таулы-мұздықты белдеу бедерінің сыртқы түрі кӛп
амплитудалы тектоникалық қозғалыстардың, беткейлік денудацияның, ежелгі
және қазіргі мұздану әсерінен түзілген. Жоғарғы бӛліктегі ӛзен аңғарлары
трог пішініне ие (Мыңжылқы шатқалы, Үлкен Алматы ӛзені аңғары, Үлкен
Алматы кӛлінен жоғары және т.б.). Бұл кең трогтық аңғарлар әдетте
ақырғы мореналармен аяқталады.
Биік таулы белдем үшін беткейлердің тӛменгі бӛліктерінде
және ӛзендердің араларында борпылдақ-сынықты материалдардың кӛп
жиналуы тән. Беткейлердің жоғарғы бӛліктерінде ежелгі жыныстар ашық
жартас түрінде кӛрініп жатыр. Үлкен құламалар үгілу ӛнімдерін тау етегіне
орын ауыстыруына қолайлы жағдай туғызады (бұл биіктік белдемінің
жалпы ауданының 25 % -еңістік бұрышы 15 тен 20 дейін, ал 36,9 % -
еңістік бұрышы 35° жоғары) [148, 75-77 б; 149].

53
Орта таулық белдем (1400-1500 метрден 2800-3000 метрге дейін, Іле
Алатауы аумағы таулы ауданының шамамен 25 %). Бұл белдемнің 2000
метрден жоғары бӛлігінде бедердің 25-35 дан 400-қа дейінгі құламалы
беткейлерімен шатқалдары және V-тәрізді пішінді аңғарлар желісімен айқын
тілімдену тән. 2000 метрден аласалай келе беткейлердің құламалылығы 200-
300–қа дейін тӛмендейді [148, 77-80б]. Беткейлерде шыңға дейін дерлік
топырақ-ӛсімдік жамылғысы бар. Бірақ кӛптеген жартас, шақшатастар
секілді жалаңаш телімдер бар. Орта таулы белдемде ӛзеннің су және
тасынды ағындылары қалыптасуына еріген қар, нӛсер және жер асты сулары
ӛз әсерін тигізеді.
Аласа таулы–тау алды белдем (800-900 метрден 1400-1500 метрге
дейін, Іле Алтауы аумағының шамамен 35 %). Бұл белдеу жотаның орталық
сілемесінен солтүстік бағытта созылып жатқан тау қырқалармен сипатталады.
Шатқалдар түрінд келетін ӛзен аңғарларының тереңдіктері 300-500 м-ге
жетеді. Басты ағынсулардың ысыранды конустары айқын байқалады. Бұл
белдеуге үштік және қойтас-ірі малтатасты тӛрттік шӛгінделерінің сығысып
қатпарланған қабаттарынан тұратын жайпақ массивті қырқалар –
«бӛктерлер» тән. Жалпы алғанда, аласа таулы белдемдерде биіктіктердің
амплитудалары шамалы, бірақ айтарлықтай кӛлденең тілімделген. Аумақтың
5,8 пайызында телімдену 600-700 метрге дейін жетеді [149,15-18б]. Аласа
таулы-тау алды белдемі қатты ағындының қалыптасуындағы ролі биік
таулы-мұздықты және орта таулы белдеулермен салыстырғанда
айтарлықтай тӛмен. Беткейлер еңістігінің тӛмендеуі, шамалы ғана тік
тілімдену, жауын-шашынның аз мӛлшері, шымдану үдерісінің кең орын
алуы және тағы басқа табиғи салдарынан болады.
Л.Ю.Чигринец 2002-жылы зерттеліп отырған аумақтың жер бедері туралы
ауқымды ақпарат беретін және ӛзгерістерді бағалау үшін пайдалануға болатын
Іле Алатауы ауданының үш ӛлшемді орографиялық үлгісі ГАЖ-
технологиясын қолданып, құрастырылған блок диаграммадан кӛруге болады.
Жоғарыда аталып ӛткендей геологиялық жағдайлар қатты ағындының
қалыптасуындағы ең негізгі факторлардың бірі болып табылады. Ландшафтық-
климаттық жағдайлармен қатар таужыныстарының біртектілігі немесе
біртексіздігі, түйіршіліктің ӛлшемі, су ӛткізгіштігі, майысу қасиеттері,
жарықшақтығы, химиялық тұрақтылығы, ерігіштігі және т.б. физикалық және
химиялық қасиеттеріне тәуелді болып табылатын жыныстардың тұрақтылығы
маңызды мәнге ие. Г.С. Ананьевтің [150] пікірі бойынша басқа
жағдайлар қалыпты кезінде молекулалық тіркелу күштері бӛлшектердің тік
ӛлшемдеріне кері пропорционал, яғни ірі түйіршікті жыныстар
ұсақтүйіршіктілерге қарағанда бұзылуға кӛбірек қабілетті.
Іле Алатауы геологиясын зерттеулерге кӛптеген жұмыстар арналған.Ең
танымалдары Н.Г. Кассин [ 151], Д.Н. Казанли [152], Г.Ц. Медоев
[153], Н.Н. Костенко [154], А.А. Абдуллин [155], Е.Д. Шлыгин [156], У.М.
Ахметсафин және т.б. еңбектері.
Жотаның баурайында тӛрттік шӛгінділермен түрлі қалыңдықта

54
ысыранды-конустары қалыптасқан. Жергілікті жер биіктіктерінің
ӛсуімен тӛрттік шӛгінділердің таралуы мен тереңдігі азаяды да, ірілігі
ұлғаяды. Оңтүстік бағдарлы беткейледе солтүстік бағдарлылармен
салыстырғанда борпылдақ шӛгінділер қалың болып келеді [157].
Жетісу Алатауы- Қазақстанның шығыс бӛлігіндегі тау жүйесінің
солтүстік және солтүстік-батыс жалғасы болып табылады. Ол
солтүстігінде Алакӛл қазаншұңқыры мен оңтүстігінде Іле ӛзені арнасы
аралығында ендік бағытта созылып жатыр. Ұзындығы 450 км, ені 100-200 км.
Жоңғар Алатауын батыста Кӛксу ӛзені, шығыстаБуратал ӛзені кесіп ӛтеді. Бұл
екі бӛлік Солтүстік және Оңтүстік Жоңғар Алатауы деп аталатын екі үлкен тау
жотасын құрайды. Жер бедері орташа тік бӛлімдермен (15-200) әртүрлі
рубтардың бұзылған беткейлерімен, ең бастысы солтүстік және солтүстік батыс
экспозициясында кӛрсетілген. Сонымен қатар кейбір қыраттардың плато
тәріздес шыңдарымен, мысалы, Қайқан, Бельтерен, Суық жайлау, Шұбарағаш,
Қотырқайың және т.б.
Қазақстан шекарасының маңында Қытай аумағында жатқан Үрқашар
Барлықтау тауына (3300 м, Жайыр және Майлыдеп аталатын аласа тауларға
ауысады. Бұл тау жоталары аралығында кең аңғарлы ойыс орналасқан. Тау
аңғарларында кішігірім тас үйінділері (қорымдар) кӛп. Ойыс Ебінұр кӛлін
Алакӛл қазаншұңқырымен жалғастырып жатыр, бұл табиғи асуды Жоңғар
қақпасы деп атайды. Жоңғар Алатауының Қазақстандық бӛлігі Жетісу
қақпасынан батысқа қарай басталады. Бұл бӛлігінде негізінен Жетісу Алатауы
ассиметриялық құрылымы бар алып тау жотасына айналады. Жетісу Алатауы
қатар жатқан екі басты жотадан (Солтүстік және Оңтүстік) тұрады, оларды
Кӛксу ӛзені бӛліп жатыр. Солтүстік жотаның ең биік нүктесі — Бесбақан (4622
м). Солтүстік тау жотасы солтүстік-шығысқа қарай баспалдақ тәрізді аласара
отырып, біртіндеп кең жазыққа ұласады. Оның жоғарғы сатысында Тастау,
Ақанжайлау, Қотыртас, Мыңшұқыр (2800-3000 м) орналасқан. Олардың
аралығында кіші, Суықтау, Қарашоқы, Желдіқарағай (2000 м) тау жоталары.
Үшінші саты — Шыбынды, Қарасарық, Қырықкӛл тау алды аңғарлары (1500-
1600 м), олардан солтүстік-шығысқа қарай Текелі және Сайқан таулары (1100
м) орналасқан. Оңтүстік жотаның ең биік нүктесі — Мұзтау (4370 м). Оңтүстік
тау жотасының жер бедері де сатылы, бірақ солтүстік жотамен салыстырғанда
күшті тілімделген. Оның батысында және оңтүстік-батысында Итшоқы,
Қотырқайың, Алтынемел, Суаттау тау алды жоталары орналасқан. Олар Іле
ӛзені аңғарына жақындап келеді. Солтүстік беткейдің жер бедері тегіс, кӛлбеу
болады, тау тӛбелері тегіс. Тау қыраттарын бӛліп жатқан аңғарларының
беткейлері тік жарқабақты, тілімделген. Оңтүстік беткейлері жарлауыт болып
келеді. Тау шыңдары солтүстікпен салыстырғанда тегіс емес, кӛтеріңкі болады.
Топырақ түзуші жыныстар лесс тәріздес құмбалшық. Шығу тегі
пролювияльды-делювияльды құмбалшықтармен балшықтар. Тау жоталары
кембрийге дейінгі граниттер мен кристалдық гранитті тақтатастардан құралған.
Жетісу Алатауының солтүстік және солтүстік-батыс бӛліктерінде силур мен
девонның құмтастары, сазды тақтатастарынан тұратын негізгі жыныстардан

55
басқа тӛменгі және орта палеозойдың шӛгінді жыныстары да кездеседі. Тау
жүйесінің оңтүстік және оңтүстік-батыс бӛліктерінде жоғарғы палеозойдың
кӛптеген жыныстары таралған. Олар тас кӛмір және пермь дәуірлерінің
құмтасы, тақтатасы, әктасы мен конгломераттарынан тұрады. Тау аралық
қазаншұңқырлар мен аңғарларда, тау етектерінде палеоген мен неогеннің
шӛгінді жыныстары кездеседі. Жетісу Алатауы каледон және герцин тау түзілу
кезінде қатпарлы тауларға айналды, ал мезозой мен тӛменгі кайнозойда сыртқы
күштер әсерінен бұзылып, біртіндеп тегістелді. Қарқынды жүрген альпі
қатпарлығы нәтижесінде кейіннен қатпарлы-жақпарлы биік тауға айналды. Жер
бедерінің қалыптасуына ежелгі және қазіргі мұз басулар да ықпал етті. Мұз
басу іздері әртүрлі жерлерде байқалады.
3.3 Ӛсімдіктері
Іле Алатауының солтүстік беткей ӛсімдіктері айтарлықтай әйгілі
оқымысты ғалымдар П.П. Семенов, А.В. Шинтников, М.Г. Попов, П.П.
Поляков және т.б. еңбектерінде айтарлықтай сипатталынған. Олар бойынша
бұл аймақта ӛсімдіктердің таралуы тіктік белдеулерге сүйенген, яғни
ӛсімдіктер бірін бірі тіктік белдеулерде жүйелі түрде заңдылықпен ауыстырып
отырады.
Іле Алатауының жабайы жемісті орман белдеуіндегі орманның күнгірт-сұр
топырағының ӛсімдіктері: кӛктерек, аққайың, долана, шырша, жабайы алма,
жабайы алмұрт, қарақат, шӛптесін ӛсімдіктер құрайды. Біржылдық және кейде
кӛпжылдық ӛсімдіктер дамыған. Олардың биіктіктері 30-50 см маңында.
Жетісу Алатауының табиғи паркі ӛсімдік және жануарлар әліміне ӛте бай.
Жетісу Алатауының флорасы ӛсімдіктердің 2168 түрімен берілген, оның 76
эндемикалық түрі тек қана осы жотада ғана кездеседі. Жетісу Алатауының
флорасында 21 сирек кездесетін түрлері де бар.
Жетісу Алатауы жабайы жемісті орман белдеуіндегі орманның күнгірт-сұр
топырағының ӛсімдіктері: жабайы алма, кӛктерек, аққайың және шетеннен
тұратын жалпақ жапырақты орман. Мұнда шыршалы ормандар мен самырсын
ӛседі. Орманшаларда долана, ырғай және жидекті бұталар- қарақат, таңқурай,
қарабӛлдірген кездеседі.

56
4 ЗЕРТТЕУ НӘТИЖЕЛЕРІ
Микроорганизмдердің топырақта таралуы топырақтың гранулометриялық
және химиялық құрамдарына, ӛңдеу дәрежесіне, ӛсімдік жамылғысына тәуелді.
Тағыда маңызды кӛрсеткіштер - ол ылғалдылық, рН, температуралық жағдай
және жыл мезгілі. Санды және сапалы кӛрсеткіші бойынша органикалық затқа
бай топырақ басымдылық кӛрсетеді, оның ішінде біздің зерттеуге алған жабайы
жемісті орман топырағы десек болады.
Зерттеу мақсатына байланысты Іле және Жетісу Алатауларының жабайы
жемісті орман ағаштарының астында қалыптасқан топырақтардың
микрофлораларының сандық және сапалық кӛрсеткішін анықтау жүргізілді.
Ұзақ жылдар бойы топырақтану ғылымында топырақты физикалық және
химиялық зат ретінде, яғни тек ӛсімдіктер ӛсетін орта деп танылып, онда
тіршілік ететін микроорганизмдер мүлде ескерілмеді. Кӛрнекті орыс
ғалымдары В. В. Докучаев, П. А. Қостычев және В. Р. Вильямстың еңбектерінің
нәтижесінде топырақтың аса күрделі орта екендігін, онда миллиондаған
микроорганизмдер және басқа да жәндіктер тіршілік ететіндігін дәлелденді.
4.1 Bacillus thuringiensis бактериялары қалыптасқан Іле Алатауы
топырақтарының морфо-гентикалық сипаты және қҧрамдары мен
қасиеттері
Кескін (26.10.2014) Іле Алатауының солтүстік беткейінде Медеу және
Бутаковка шатқалдары аралығындағы аралас орман белдеуінде қазылған.
Беткей экспозиция солтүстік-шығыс ылдилығы 30o. Абсолюттік биктігі 1300-
1600 м. Ӛсімдіктері- долана, қайың, жабайы алма, Тянь-Шан шыршасы, кӛп
жылдық шӛптесін ӛсімдіктер.
Тӛменде (сурет 7) орманның күңгіріт-сұр топырағының кескіні
бейнеленген.
Сурет 7 - Іле Алатауы жабайы жемісті орман белдеуіндегі орманның
күңгірт-сұр топырағының кескін бейнесі

57
Зерттеуге алынған топырақ кескіні бір-бірінен айқын дифференцияланған
тӛрт қабаттан тұрады. Олар беткі гумусті-аккумулятивті (А), ӛтпелі В1 және B2
қабатшалары және соңғының астында аналық тау жынысына дейінгі тереңдікте
ВС қабаты қалыптасқан. Олардың жалпы қалыңдығы 105см. Бұл топырақ
кескіннің жауын-шашын мӛлшерімен ӛсімдік қалдықтарының жеткілікті
болуына қарамастан қалыңдығының онша терең еместігін кӛрсетеді. Негізгі
себеп, беткей ылдилығының жоғары болуында. Дегенмен бұндай жағдайдың
ӛзінде гумус қабаты жақсы жетілген. Оның қалыңдығы (A+B1) 30 см құрайды.
A 0-15см
15
B1 15-30см
15
B2 30-80 см
50
BC 80-105см
25
C1 105-120см
15
Күңгірт сұр, дымқыл, сәл нығыздалған, шашыраңқы,
ұсақ кесекті-дәнді, тамырлармен күшті кескінделген,
агрегаттардың біразы моншақ тәрізді тамырлармен
байланысқан, ауыр құмбалшықты, жауын
құрттарының қуыстық іздері және копролиттері
кӛптеп кездеседі, ауыр құмбалшықты, 10 %- тік HCl
дан қайнамайды, келесі қабатқа ӛту түсіне қарай
айқын.
Күрең-сұр, ӛте дымқыл, кей жерлерінде су топырақ
агрегаттарының аралығында және олардың бетінде
жылтырайды, ұсақ тамырларымен кесінделген,
жекешеленген ірі тамырларда кездеседі, ауыр құм
балшықты, 10 %- тік HCl дан қайнамайды,
саңырауқұлақ жіпше торлары кездеседі, келесі қабатқа
ӛту түсіне қарай біртіндеп.
Сұр-күрең, ӛте дымқыл, орташа нығыздалған, ұсақ
кесекті, ауыр құмбалшықты, ұсақ ӛсімдік
тамырларымен сирек кесінделген, жауын құрттарының
іздері (қуыстары) және копролиттері кездеседі,
агрегаттардың беттері жылтыр қабыршақтармен
қапталған, 10 %-тік HCl дан қайнамайды, келесі
қабатқа ӛту құрлымы мен түсіне қарай біртіндеп.
Түсі әртүрлі, сұр-күрең сарғыштау дақты, сәл дымқыл,
күшті нығыздалған, тамырлармен сәл кескінделген,
түтікті, диаметрі 1 см дӛңгелек қуыстар жауын құрты
копролиттерімен толтырылған, жауын құрттарының
іздері кездеседі, ауыр құмбалшықты, 10 %- тік HCl дан
қайнамайды, келесі қабатқа ӛту түсіне қарай біртіндеп.
Түсі әртүрлі, сұр-күрең және сарғыштау дақты, сәл
дымқыл, күшті нығыздалған, түтікті, диаметрі 1см
дӛңгелек қуыстар жауын құртының копролиттерімен
толтырылған, жауын құрттар іздері кездеседі, ауыр
құмбалшықты, саңырауқұлақтар түрінде, келесі
қабатқа ӛту түсіне қарай біртіндеп.

58
Түйіртпектілігі суға тӛзімді ұсақ кесекті-дәнді. Олар бір-бірімен ӛсімдік
тамырларымен құрсауланған. В2 қабатшасы жоғарғылардан күрең ӛңінің
басымдылығы және қалыңдылығымен (50 см) ерекшеленеді. Аналық тау
жынысы үстіндегі қалыптасқан ӛтпелі ВС қабатында топырақ түзілу
үдерстерінің қарқындылығы бәсеңдеген. Оны ӛсімдік тамырларының
сиректенгендігі кӛрсетеді. Бұл қабат мезофаунаның әсіресе жауын
құрттарының қысқы тұрағына айналған. Олар топырақ түзілуге әлі іліне
қоймаған аналық тау жынысы лесс С қабатында да кездеседі.
Топырақтың түзілуі кескіндегі су құбылымының шайылымды жағдайында
жүрген. Оны кескіннің барлық қабаттарының карбонаттан мүлдем
шайылғандығын тұз қышқылынан қайнамайтындығы кӛрсетеді.
Жоғарыда сипатталған орманның күңгірт-сұр топырағы ӛзінің жоғары
гумустылығымен, әсіресе А қабатында (10,57 %), бірақ оның астындағы В1
қабатшада күріт тӛмендейтінділігімен (2,70 % дейін), жалпы гумустық
қабаттың (A+B1), жұқалығымен (30 см) ерекшеленеді (кесте 5 ).
Топырақтағы гумустық кескіннің бұлай болып ӛзгеруін ӛсімдік
жамылғысы құрамымен түсіндіруге болады. Шӛптесін ӛсімдік тамырларының
негізгі бӛлігі топырақтың беткі қабатында орналасса, орман ағаштары мен
бұталардның тамырлары тереңірек қабаттарда жайғасқан. Осыларға сәйкес
ӛсімдік қалдықтарының мӛлшері мен сапасының кескін бойында әртүрлі
болуы, олардың гумификациялануының бағыттары мен қарқындылығының
бірдей болмауы нәтижесінде топырақ кескінінің беткі қабатында ӛте жоғары
гумусты, ал оның астында сәл гумустенген қабаттардың қалыптасуына әкелген.
Зерттелініп отырған топырақтағы жалпы азоттың мӛлшері А қабатында ӛте
жоғары (0,569 %), ал оның астында орташа (0,18 %). Оның кескін бойы ӛзгеруі
гумустың ӛзгеруімен тығыз байланысқан. Себебі гумин- және
фульвоқышқылдары негізінен кӛміртегінен тұрады. Оның элементтік
құрамында кӛміртегі мен азоттың қатынасы орта есеппен сәйкесінше 56:4 және
42:3 тең [158], яғни топырақта гумус мӛлшері артқан сайын азоттың да мӛлшері
арта түседі.
Топырақта жалпы фосфордың мӛлшері кӛтеріңкі, оның мәні топырақтың
беткі қабатында 0,25 % құрайды. Топырақ түзілу үрдісі бейтарапты ортада
жүреді (pH-7,0). Бұл кӛрсеткіш мезо және микрофаунаның ӛсіп-ӛнуіне оңтайлы.
Топырақтың алмасуға қабілетті катиондарының жиынтығы айтарлықтай
жоғары (20-23 мг-экв100 г топыраққа шаққанда). Бұл жағдай топырақтың
жоғары гумустілігі және ауыр құмбалшықтылығымен түсіндіріледі. Себебі
топырақтың органикалық бӛлігінің иондарды сіңіру қабілеттілігі минералдық
бӛлігінен 10-30 есе артық екендігінде, онымен қоса топырақта тозаң және ұсақ
шаң фракцияларының басымдылығы топырақтың беттік меншікті аумағын,
яғни олардың қоршаған ортамен (газ және сұйық) әрекеттесуін күрт ұлғаитады.
Бұл топырақ бӛлшектерінің сіңіру аумағын арттырып қана қоймай, ондағы
алюмосликаттар еншісінде арттырады, ал оларда әрдайым гумус мӛлшері
кӛтеріңкі.

59
Кесте 5 - Іле Алатауының жабайы жемісті орман белдеуіндегі орманның күңгірт-сұр топырағының химиялық құрамы
Үлгі
тер
еңд
ігі,
см
%
рН
Сің
іру с
ый
ым
ды
лы
ғы,
мг/
экв 1
00г
топ
ырақ
та
Сусү
зін
діс
інің
100г топырақта сіңірілген негіздер, мг-экв/%. Жылжымалы формасы мг/100г Г
ум
ус
Жал
пы
азо
т
Жал
пы
фосф
ор
Са2+
Mg2+ Na
+ K
+
Жең
іл
гид
роли
зден
етін
азот
Р2О5 К2О
0-15 10,57 0,569 0,25 7,2 23,35 19,60/84 3,20/13,7 0,54/2,3 0,01/0,04 9,80 4,16 74,40
15-25 2,98 0,192 0,21 7,0 19,32 16,40/85 2,40/12,4 0,46/2,4 0,06/0,31 3,70 1,80 23,04
40-50 2,42 0,162 0,18 7,0
1,45 19,20

60
Сіңірілген катиондар негізінен калциймен берілген. Оның еншісі 85 %
құрайды. Магнийдің мӛлшері аз (13 %), ал натрий тіптен аз (2,3%). Соңғы
кӛрсеткіш топырақтың кебірленбегендігін кӛрсетеді. Сіңірілген калийдің
мӛлшері 0-15 см және 15-25 см тереңдікте болар-болмас сәйкесінше 0,01және
0,06 мг-экв немесе 0,04 және 0,31 % құрайды. Бұл ион топырақтағы калийдің
потенциялдық қоры болып есептелінеді.
Топырақтағы азоттың ӛсімдікке тиімді түрлерінің жиынтығы жеңіл
гидролизденетін азот. Оның мӛлшері біз зерттеп отырған топырақтың беткі
қабатында жоғары- 100 г топырақта 9,8 мг. Ал оның астындағы қабатында
күрт тӛмендеген (3,1-мг 100 г топырақта). Жылжымалы фосфордың мӛлшері
беткі қабатта кӛтеріңкі, ал оның астындағы қабатында орташа. Бұл топырақ
алмаспалы калиймен қамтамасыздануы беткі қабаты ӛте жоғары қамтамасыз
етілген, ал астыңғы қабат орташа.
Сӛйтіп зерттелініп отырған Іле Алатауының солтүстік беткейінде жабайы
алма ағаштары белдеуіндегі орманның күңгірт-сұр топырағының түзілуі
шӛптесін және орман ағаштары астында бейтарапты ортада, қолайлы
гидротермиялық жағдайда жүрген. Онда гумус ӛте жоғары, азот, фосфор және
калий мӛлшерлері жоғары және орташа деңгейлерде. Бұл жағдайлар сіңірілген
катиондар сыйымдылығынын кӛтеріңкілігін қалыптастырған. Онда калций
ионы мейілінше басым. Топырақтың беткі қабатының жылжымалы қоректік
заттармен қамтамасыз етілуі азотпен жоғары, фосформен-кӛтеріңкі,
каллиймен-ӛте жоғары деңгейлерде, ал оның астындағыда азотпен-жоғары,
қалғандарында орташа деңгейлерде қамтамасыз етілген.
Келесі кестеде (кесте 6) топырақ кескінінің гранулометриялық құрамы
кӛрсетілген. Онда зерттелініп отырған топырағымыздың механикалық
элементтері-фракциялар құрамында ірі шаңның басым екендігі байқалтады
(43-46 %). Оның мӛлшері топырақ массасының жартысына жақын. Топырақ
құрамында аздаған (3-4 %) ұсақ құм кездеседі. Ал ірі және орташа құм жоқтың
қасы. Топырақ құрамында кӛптілігі бойынша екінші болып тозаң, сонан соң
ұсақ шаң тұр. Бұл екі фракция, әрине коллоидтармен бірге топырақтың
катиондар сіңіру қабілеттілігі деңгейін анықтайды. Физикалық балшық
(<0,01мм) мӛлшері топырақтың ауыр құмбалшықтылығын кӛрсетеді. Топырақ
құрамында тозаң және ірі шаң фракцияларының басым екендігі айқын
байқалады. Бұлар зерттелініп отырған топырақтың гранулометриялық құрамы
тозаңды-ірі шаңды ауыр құмбалшықты екендігін кӛрсетеді.
Кесте 6 - Іле Алатауының жабайы жемісті орман белдеуіндегі орманның
күнгірт-сұр топырағының гранулометриялық құрамы Үлгі
тереңдіг
і, см
Абсолютті құрғақ топырақтағы фракциялар (мм) мӛлшері,% <0,001мм
фракциялар
жиынтығы Құм Шаң Тозаң
1-0,25 0,25-0,05 0,05-0,01 0,01-
0,005
0,005-
0,001 <0,001
0-15 0,20 3,44 43,21 12,05 19,83 21,27 53,15
15-25 0,15 3,03 46,16 14,00 15,58 21,08 50,66
40-50 0,14 4,29 46,86 10,08 17,27 21,36 48,71

61
6 – кестенің жалғасы 60-70 0,34 7,37 41,61 12,7 15,09 22,89 50,68
90-105 0,04 9,31 41,53 11,84 14,52 22,76 49,12
105-120 0,06 4,41 47,97 11,61 14,46 21,49 47,56
Келесі (кесте 7) орманның күңгірт сұр топырағының су сүзіндісінің
құрамы кескін қабаттарының тұзданбағандығын, тұз мӛлшерінің ӛте аз (<0,05
%) екендігін кӛрсетеді. Су сүзіндісінде қалыпты карбонаттардың (CO2-
3) және
сульфат ионының мүлдем жоқ екендігін, бикарбонат және хлор иондарының
концентрацияларының улы мӛлшерлерінен (сәйкесінше 100 г топырақта 0,8
және 0,14 мг-экв) тӛмен екендігін кӛрсетеді. Топырақ ертіндісінде натрий
және калий иондарының мӛлшері ӛте тӛмен. Топырақ түзілу және үгілу
процестерінің барысында түзілген бұл иондар топырақ кескінінен шайылып
кетіп отырған. Топырақ кескінінде калциймен магнийдің басқаларға қарағанда
айтарлықтай мӛлшерде болуы олардың биогендік карбонаттар құрамында
бекініп қалуымен (суда ӛте нашар еритіндігімен) түсіндіріледі. Кестенің соңғы
екі бағаналарын бір-бірімен салыстырсақ, жалпы олардың мӛлшерлерінің аз
екендігін, ондағы тығыз қалдықтың тұздар мӛлшерінен 1,5 еседей артық
екендігін оңай байқауға болады. Бұл айырмашылық топырақта тұздардан да
басқа еріген заттардың бар екендігімен түсіндіріледі. Олар негізінен
топырақтың суда еритін органикалық заттарымен (гумус қышқылдары және
олардың тұздары гуматтар және фульваттар) берілген.
Кесте 7 - Іле Алатауы жабайы жемісті орман белдеуіндегі орманның күңгірт-
сұр топырағының (ауалы-құрғақ) су сүзіндісінің құрамы, мг-экв/%
Топырақ ылғалдылығы топырақ түзілу үрдістеріне тікелей және жанама
әсер етеді. Жоғарыда бұл топырақ белдеуін сипаттағанда түсетін жауын-
шашын мӛлшерінің булануынан басым екендігі айқындалған. Міне, соған
байланысты, біз зерттеп отырған топырақ жыл мезгілдерінің барлық
кезеңдерінде ылғалды. Ал оның күз, қыс және кӛктем айларында оның
Үлгі
тер
еңд
ігі,
см
Сілтілік
Cl-
SO42-
Са2+
Mg2+
Na+ K
+
Тұзд
ар ж
иы
нты
ғы,
%
Ты
ғыз
қал
ды
қ,%
Жалпы
НСО3-
СО32-
тен
0-15
0,52 0,00 0,10 Жоқ 0,30 0,15 0,05 0,12 0,049 0,072 0,032 0,000 0,004 Жоқ 0,006 0,002 0,001 0,005
15-25
0,26 0,00 0,10 Жоқ 0,20 0,10 0,04 0,03 0,027 0,048 0,016 0,000 0,004 Жоқ 0,004 0,001 0,001 0,001
40-50
0,24 0,00 0,10 Жоқ 0,15 0,10 0,07 0,03 0,025 0,036
0,015 0,000 0,004 Жоқ 0,003 0,001 0,002 0,001

62
мӛлшерінің артық болуы топырақ кескінінде «шайылу» су құбылымының
қалыптасуына әкеледі.
Орманның күңгірт-сұр топырағы кескінінің кӛктем және күз
кезеңдеріндегі далалық ылғалдылығы мәліметтерінен топырақ, әсіресе оның
беткі 0-25 см қабатының ылғалдылығының мейілінше жоғары екендігін кӛруге
болады (сурет 8) Кескіннің ылғалдылығы жоғарыдан тӛмен қарай біртіндеп
кеміген. Егер беткі A қабатында оның мӛлшері кӛктемде 31,12 % болса, күзде
бұл кӛрсеткіш 33,43 % -ға дейін кӛтерілген. топырақтың BC қабатында (105-
120 см) кӛктемде 14 %, күз мезгілінде 13,43 % дейін тӛмендеген. Біріншісі
топырақтың далалық су сиымдылығына шамалас болса, екіншісі солу
ылғалдылығына (8-12 %) жақындаған. Соңғы ылғалдылық топырақ кескінінің
жауын-шашынмен ылғалдау тереңдігін кӛрсетеді. Топырақта ӛсімдік
тамырларымен мезо-және микрофауналарының шоғырлануы 0-50 см
қабатында болғандықтан жоғарыда байқалған дымқылдылық мӛлшері
олардың ӛсіп дамуына оңтайлы екендігін кӛрсетеді.
Сурет 8 - Іле Алатауы жабайы жемісті орман белдеуіндегі орманның
күңгірт сұр топырақтарының далалық ылғалдылығы
4.2 Bacillus thuringiensis бактериялары қалыптасқан Жетісу Алатауы
топырақтарының морфо-генетикалық сипаттары және қҧрамдарымен
қасиеттері
Орманның күңгірт-сұр топырақтары Жетісу Алатауының орманды
белдеуінің орта шенінде 2000-2500 м абсолютты биктіктерде қалыптасқан.
Олар әлбетте үздікті белдеу түрінде орманды дала аймағының басқа
топырақтары арасында жекеленген алқаптар түрінде орналасқан. Олардың
үздіксіз таралуын тек Константиновка, Тополевка, және Лепсі ауылдары
маңында байқауға болады. Онда орманды-дала ӛсмдіктерінің арасында
жапырақты орман басты орын алған (алма, қайың, кӛктерек).
Жер бедері негізгі беткейлердің тіктігі орташа (15-200) әртүрлі бағытты.
Беткейлердің экспозициясы.негізінен солтүстік және солтүстік батыс.

63
Сонымен қатар кейбір қыраттар плато тәріздес болып келеді. Мысалы,
Кайқан, Бельтерен, Суық жайлау, Шұбарағаш, Қотырқайың және т.б.
Топырақ түзуші жыныстар лесс тәріздес құмбалшық. Шығу тегі
делювияльды. Сипатталынып отырған топырақтарда ағашты ӛсімдіктері
басым: олар кӛктеректен тұратын жапырақты ормандар, теректер, қайыңдар,
шетендер, жабайы алмалар, мойыл, ырғай, бӛріқарақат және басқалары.
Бұл ӛңірдің орманның күңгірт–сұр топырақтарының морфологиялық
белгілері кейбірі қара топырақтың күлденген және сілтісізденген типшелеріне
жақын. Олардың гумустенген қабаттары қара-сұр және күнгірт–сұр реңге ие.
Кӛп жағдайда топырақтың беті орман тӛсенішімен жабылған. Күлденген
қабаты мүлдем жоқ, бірақ топырақ агрегаттарының үстінде кремнийлі сеппе
әрқашанда екінші гумус қабатында (A2B) байқалады. Бұл олардың Іле
Алатауларының аттас топырақтарынан морфологиялық ерекшелігіне
жатқызуға болады.
Орманның күңгірт–сұр топырақтарын сипаттау үшін тӛмендегі кескіннің
бет пішінің морфогенетикалық белгілерін келтіреміз.
Кескін Константиновка ауылынан оңтүстік–шығысқа қарай 5 км жерде
орналасқан. Кескіннен тура осындай қашықтықта солтүстік-батыста Красная
ӛзені Тентек ӛзенінне қосылады. Жер бедер беткейі тік (20-250). Қара шоқы
тауының солтүстік беткейі Красная ӛзеннің аңғарымен күшті кескінделген.
Абсолюттік биіктігі 2000 м.
Ӛсімдіктері: жабайы алма, кӛктерек, аққайың және шетеннен тұратын
жапырақты қалың орман. Бұталардан долана, ырғай, ал жидекті бұталардан-
қарақат, таңқурай, қарабүлдірген кездеседі.
Кескіннің тереңдігі 150см. Топырақ 10 % HCl ертіндісінен бетінен бастап
түбіне дейін қайнамайды. Кескіннің 15 см-нен бастап 90 см-ге дейінгі
тереңдіктерде кремний сеппелері айқын байқалады.
А0 0-5 см Ағаштардың жапырақтарынан және жартылай ыдыраған майда
бұтақтардан тұратын орман тӛсеніші;
A1 5-15 см Қара-cұр, құмбалшықты, ылғалды, бос, ірі дәнді, келесі
қабатқа ӛтуі айқын;
A2B 15-40 см Күреңді күңгірт-сұр, құмбалшықты, ірі дәнді, ылғалды, бос,
жауын құрттарымен қазғыланып ӛңделген, 30 см-ден бастап топырақ
агрегаттарының қырларында аздаған кремнийлі сеппесі байқалады, келесі
қабатқа ӛтуі біртіндеп;
B1 40-70 см Күңгірт-сұр, құмбалшықты, жаңғақты, ылғалды, нығыздалған,
жауын құрттарымен қазылған, агрегаттардың қырларында кремнийдің мол
сеппесі байқалады;
B2 70-90 см Күңгірт дақты сұрлау-күрең, ылғалды, құмбалшықты, ірі
жаңғақты-кесекті, нығыздалған, агрегаттардың қырларында аздаған кремнийлі
сеппесі, келесі қабатқа ӛтуі айқын;
BC 90-150 см Қызғылттау-күрең, балшықты, ылғалды, нығыздалған.
Топырақ кескіннің морфологиялық сипатынан, оның айқын байқалатын
алты генетикалық (A0, A1, A2, B1, B2 және BC) қабаттарынан тұратындығы,

64
топырақ бетін айтарлықтай қалың (5 см) орман тӛсенішімен жабылғандығын,
оның астында жұқа (10 см) жақсы жетілген гумусты-аккумулятивті қабаттың
қалыптасқандығын, ал келесі A2B және B1 қабаттарда қарқынды сілтісіздену
үрдісінің нәтижесінде топырақ агрегаттарының қырларында кремний
сеппелері кездеседі. Айтылған қабаттар 70 см тереңдікке дейін жауын
құртымен қазбаланып жақсы ӛңделген. Соңғы жағдай топырақтың құнарлық
деңгейінің жоғарылығының белгісі. Оның қаншалықты екендігін осы кескін
қабаттарынан алынған топырақ үлгілерінің құрамына жүгінейік (кесте 8 )
Кестеден орманның күңгірт-сұр топырағының беткі A1 қабатының
жоғары деңгейде гумустелгендігін (8,30 %), оның мӛлшері кескін бойы тӛмен
қарай біртіндеп азайып 15-40 см тереңдікте 3,88 % жететіндігін байқауға
болады. Осыған сәйкес топырақ жалпы азотпен фосфорға айтарлықтай
байытылған (0,52 және 0,25 % сәйкесінше). Су сүзіндісінің мәліметтері
топырақ түзілу үрдісінің қышқылды ортада жүретіндігін кӛрсетеді. Сініру
сиымдылығы кӛтеріңкі жағдайда (20-30 мг-экв 100 г топырақта) сіңірілген
натрийдің еншісі тӛмен (2,5 %). Бұл топырақта кебірлену белгісінің жоқтығын
кӛрсетеді. Топырақ ӛсімдіктерге тиімді қоректік азот, фосфор және калий
элементтерімен мейілінше байытылған.
Кесте 8 - Жетісу Алатауы орманының күңгірт-сұр топырағының химиялық
құрамымен физико-химиялық қасиеттері
Топырақтың гранулометриялық құрамының, оның кескін бойы бірдей
орташамен ауыр құмбалшықты шекаралықтағы шӛгінді тау жынысы лӛсте
қалыптасқандығын кӛрсетеді. Лестің ӛсімдік тамырларымен топырақты
мекендейтін жан-жануарлардың бәріне ӛте қолайлы екенін ескерсек, біз
сипаттап отырған топырағымыздағы орман ағаштары оларға ең бір қолайлы
топырақтүзуші жынысында қалыптасқандығын кӛреміз (кесте 9).
Үлгі
тер
еңд
ігі,
см
Гум
ус,
%
Жал
пы
азо
т, %
Жал
пы
фосф
ор ,
%
Су с
үзі
нд
ісін
ің р
Н
Сің
іру с
ый
ым
ды
лы
ғы,
мг/
экв 1
00 г
топ
ырақ
та 100 г топырақта сіңірілген
негіздер, мг-экв/%
Жылжымалы
түрлері, мг/100г
Са2+
Mg2+
Na+ K
+
Жең
іл г
ид
роли
з
ден
етін
азо
т Р2О5 К2О
0-5 15,5 0,8 0,28 6,8 30,2 29,4/8
0
7,35/
12,2
0,65
/2,8
0,08
/0,18
10,2
5
5,12 82,20
5-15 8,30 0,518 0,24 6,4 21,6
5
18,45
/80
5,25
/12,8
0,48
/2,4
0,01
/0,04
8,60 4,06 79,20
15-
40
3,88 0,185 0,20 6,4 18,7
2
15,60
/81
3,82
/12,2
0,42
/2,1
0,05
/0,29
2,70 1,50 21,01
40-
70
2,22 0,152 0,15 6,4 15,2 3,42
/12,2
1,42 17,20

65
Кесте 9 - Жетісу Алатауының жабайы жемісті орман белдеуіндегі орманның
күңгірт-сұр топырағының грануламетриялық құрамы
Зерттеліп отырған орманның күңгірт-сұр топырағының құрамында суда
ерімейтін тұздар мӛлшерінің ӛте аз екендігін мәліметінен айқын кӛруге
болады (кесте 10 ). Онда тұздар жиынтығы (0,05 %) дан (0,098 %) аралығында
ауытқыған. Бұл ӛте тӛмен кӛрсеткіш топырақта түзілген тұздардың әрдайым
шайылып кетіп отыратындығынан топырақтың тұздануына жол
бермейтіндігін кӛрсетеді.
Кесте 10 - Жетісу Алатауының орманды күңгірт-сұр топырағының (ауалы-
құрғақ) су сүзіндісінің құрамы, мг-экв/%
Сипатталынып отырған кескіндердің гидротермиялық құбылым
жағдайларының мәліметтерінен олардағы генетикалық горизонттарының
арасындағы ылғалдылықтың айырмашылықтарын айқын кӛруге болады. Оның
мұндай айырмашылықтары вегетациялық кезеңінің басында, әсіресе, кескіннің
Үлгі
тер
еңд
ігі,
см
Абсолютті құрғақ топырақтағы фракциялар мӛлшері %
Диаметрі
<0,001 мм
фракциялар
жиынтығы
Құм Шаң Тозаң
1-0,25
мм
0,25-
0,05мм
0,05-
0,01мм
0,01-
0,005
Мм
0,005-
0,001
мм
<0,001
Мм
0-5 Жоқ 12,6 43,21 10,05 12,03 8,2 30,15
5-15 10,03 38,70 14,20 15,05 12,08 45,60
15-40 9,50 42,85 10,08 11,28 15,36 45,71
40-70 7,37 41,63 12,4 15,09 15,30 45,80
70-90 9,31 41,50 11,42 14,25 15,30 45,82
90-150 9,20 45,90 11,65 14,72 15,20 49,50
Үлгі
тер
еңд
ігі,
см
Сілтілік
Cl-
SO42-
Са2+
Mg2+
Na+ K
+
Тұзд
ар ж
иы
нты
ғы,
%
Ты
ғыз
қал
ды
қ,
%
Жалпы
НСО3-
СО32-
тен
0-5
0,60 Жоқ 0,12 Жоқ 0,40 0,17 0,12 0,18 0,098
0.0
80 0,072 0,004 0,006 0,004 0,004 0,008
5-15
0,52 0,10 0,30 0,15 0,07 0,12 0,05
0,0
74 0,032 0,004 0,006 0,002 0,001 0,005
15-40
0,28 0,10 0,22 0,14 0,04 0,05 0,025
0,0
47 0,014 0,004 0,004 0,001 0,001 0,001
40-70
0,26 0,09
0,12 0,12 0,07 0,04 0,027
0,0
33 0,015 0,004
0,003 0,002 0,002 0,001

66
жоғарғы горизонттарында айқын кӛрінеді. Сурет- 9 да берілген орманның
күнгіртң-сұр топырағы кескінінің кӛктем және күз кезеңіндегі далалық
дымқылдылығы мәліметтерінен топырақ әсіресе оның беткі 0-25см қабатының
ылғалдылығының мейілінше жоғары екендігін кӛруге болады. Кескіннің
дымқылдылығы жоғарыдан тӛмен қарай біртіндеп кеміген. беткі A1 қабатында
ылғалдылық мӛлшері кӛктемде 32,20 % болса, күзде 30,15 %,болды. ең
тӛменгі BC қабатында (90-150см) кӛктемде 10,54 %, күз мезгілінде 12,40 %
дейін тӛмендеген. Мұнда топырақтың дымқылдылығы жоғарғы горизонт
қабаттарда біршама жоғары болды да тереңдеген сайын біртіндеп азайған.
Аталған кескін құрылысындағы ӛзгерістер зертханалық зерттеулермен
алынған (сурет 9).
Сурет 9 - Жетісу Алатауы жабайы жемісті орман белдеуіндегі орманның
күңгірт-сұр топырақтарының далалық ылғалдылығы
4.3 Жетісу және Іле Алатауларының орманның кҥңгірт-сҧр
топырақтарының микроорганизмдерінің сандық және сапалық
кӛрсеткіштері мен температура ӛзгерісіне байланыстылығы
Микробиологиялық талдауға алынған топырақ үлгілерінен дайындалған
суспензиялар тӛменде кӛрсетілген қоректік орталарға егілді.
Қатты ортаға егу 104-10
8 дәрежесіндегі сұйылтымдарынан, екі
қайталымда жүргізілді. Егілген Петри табақшалары 27-30 0С термостатқа
орналастырылып, ӛсіп шыққан колония сипаттамасы 4-5 тәуліктен бастап
жүргізілді (сурет 10).

67
Іле Алатауы жабайы жемісті орманның күңгірт-сұр топырақтарын зерттеу
барысында алынған мәліметтер ЕПА қоректік ортасында топырақтағы
аммонификаторлардың саны 15,6±0,9 млн. кл/г шамасында болды (кесте 11).
ЕПА КАА
Сабуро ЕПА
Сурет 10 - Іле және Жетісу Алатауы жабайы жемісті орман
топырақтарының жалпы микрофлорасы
Кесте 11 - Іле Алатауы жабайы жемісті орман белдеуіндегі күңгірт-сұр
топырақтарының микроорганизмдерінің физиологиялық топтары және
олардың сандық мӛлшері
Микроорганизмдердің физиологиялық
топтары
Топырақтағы
микроорганизмдердің саны,
млн. КОЕ/г
Аммонификаторлары 15,6±0,9
Спора түзуші микроорганизмдер 0,061±0,004
Олиготрофты микроорганизмдер 0,15±0,01
Минералды азотты
пайдаланатын микроорганизмдер 9,2±0,8
Олигонитрофильді
микроорганизмдер 0,84±0,07
Актинобактериялар 0,11 ± 0,001
Микромицеттер 9,8 ± 1,6
Ашытқы организмдер 0,011± 0,001
Олар топырақта азоттың органикалық формасын пайдаланады.
Аммонификаторлардың санының біздің топырақтарда біршама жоғары болуы
осы топырақта гумус пен азот мӛлшерінің едәуір мӛлшерде болуына
байланысты (10 % және 0,4 %). Аммонификаторлар топырақ түзлу процесіне
маңызды рӛл атқарады. КАА қоректік ортасында топырақтағы минералды

68
азотты пайдаланатын бактериялардың саны 9,2±0,8 млн. кл/г болды.
Минералдық азоты пайдаланатын микроорганизмдердің рӛлі-табиғатта
минералды азоттың атомдары әртүрлі тотығу дәрежесінде N5+ (N205, азот
ангидридті) тен N3- (NH3, аммиак) ке дейін кездеседі. Азоттың минералдық
қосылыстары бактериялар әсерінен аммиакқа оңай айналады. Бұл топқа
жататын микроорганизмдер қатарына азот фиксациялаушы бактериялар
жатады, (мысалы, Rhizobiut, Azotobacter, Clostridium, Klebsiella және т.б).
Спора түзуші микроорганизмдердің топырақтағы саны 0,061±0,004 млн.
кл/г. Бактериялардың кейбір түрлері сыртқы ортаның қолайсыз жағдайына аса
тӛзімді келетін спора түзеді. Спора - бактерия жасушаларында орналасқан
дӛңгелек, кейде сопақша келетін жарқырауық түйір. Спора кӛбінесе таяқша
тәрізді бактерияларда - бацилдерде пайда болады. Егер әдеттегі бактерия
жасушалары +50-60°С-та түгелдей ӛліп қалатын болса, олар спора күйінде
+100 °С ыстықты да кӛтереді және ӛте құрғақшылыққа тӛзе бере-ді.
Бактериялар спора күйінде қолайлы жағдай туғанша, ұзақ уақыт, кейде
бірнеше жылдарға дейін, тіршілік қабілетін жоймайды. Біздің зерттеу нысаны
ретінде алынған энтомопатогенді Bacillus thuringiensis бактериялары да осы
топқа жатады. Олиготрофты микроорганизмдердің топырақтағы саны
0,15±0,01 млн. кл/г болды. Бұлар қоректік ортаның тӛменгі
концентрациясында тіршілік етеді. Сондай-ақ Эшби ортасында олиготрофты
микроорганизмдердің топырақтағы саны кӛрнекті түрде жоғары яғни олардың
саны 0,84±0,07 млн. кл/г (кесте 11).
Топырақ барлық ӛсімдіктермен микроорганизмдерге табиғи субстрат
болып табылады. Ол жерден актиномицеттердің әртүрлерін бӛліп алуға
болады. Бірақ актиномицеттердің биомассасының басым кӛп бӛлігі споралар
болып табылады. Қоректік орталарға егу жүргізу әдістемесі бойынша
популияциялардың колониясын санау кезінде биомассаның тек 1-4 %
мицелийлер құрады. Ол органикалық заттары мол микрозоналарда табылды.
Зерттеу нәтижелері бойынша крахмалды-казеинды ортада топырақтағы
актинобактериялардың саны 0,11 ± 0,001 млн. кл/г болатындығы анықталды.
Олар гетеротрофтар, яғни дайын органикалық қосылыстармен
қоректенетіндер, соның нәтижесінде олар табиғаттағы зат алмасу үрдісіне
қатынасады. Зерттеу мәліметтері топырақтағы саңырауқұлақтардың мӛлшері
9,8 ± 1,6 млн. кл/г болғандығын кӛрсетті. Олар әртүрлі субстарттардың бетінде
ӛсетін хлорофилсіз микроорганизмдер. Ашытқы организмдерін санау сабуро
қоректік ортасында жүргізілді. Олардың топырақтағы саны 0,011± 0,001 млн.
кл/г болды. Бұлай болғанда топырақтағы ашытқылардың құрамы, бактериялар
мен саңырауқұлақтарға қарағанда айтарлықтай тӛмен.
Жетісу Алатауы жабайы жемісті орман топырақтарын зерттеу барысында
алынған мәліметтер ЕПА қоректік ортасында топырақтағы
аммонификаторлардың саны 14,2±0,5 млн. кл/г. Бұл Іле Алатауы жабайы
жемісті орман топырақтарының бактерияларына қарағанда салыстырмалы
түрде аздау болған. КАА қоректік ортасында топырақтағы минералды азотты
пайдаланатын бактериялардың саны 8,6±0,6 млн. кл/г болды. Спора түзуші

69
микроорганизмдердің топырақтағы саны 0,050±0,004 млн. кл/г. Олиготрофты
микроорганизмдердің топырақтағы саны 0,30±0,02 млн. кл/г болды. (кесте –
12).
Кесте 12 - Жетісу Алатауы жабайы жемісті орман белдеуі топырақтарының
микроорганизмдерінің физологиялық топтары және олардың сандық мӛлшері
Микроорганизмдердің
физиологиялық топтары
Топырақтағы микроорганизмдердің
саны, млн. КОЕ/г топырақ
Аммонификаторлары 14,2±0,5
Спора түзуші микроорганизмдер 0,050±0,004
Олиготрофты микроорганизмдер 0,30±0,02
Минералды азотты пайдаланатын
микроорганизмдер 8,6±0,6
Актинобактериялар 0,09 ± 0,002
Микромицеттер 8,5 ± 1,1
Ашытқы организмдер 0,020± 0,001
Зерттеу нәтижелері бойынша крахмалды-казеинды ортада топырақтағы
актинобактериялардың саны 0,09 ± 0,002 млн. кл/г болатындығы анықталды.
зерттеу мәліметтері топырақтағы саңырауқұлақтардың 8,5 ± 1,1 млн. кл/г
болғандығын кӛрсетті. Ашытқы организмдерін санау сабуро қоректік
ортасында жүргізілді. Олардың топырақтағы саны 0,020± 0,001 млн. кл/г
болды. Бұлай болғанда топырақтағы ашытқылардың құрамы, бактериялар мен
саңырауқұлақтарға қарағанда айтарлықтай тӛмен.
Іле Алатауы жабайы жемісті орман белдеуіндегі орманның күнгірт-сұр
топырағындағы бактериялардың жалпы сандық мӛлшерінің жыл мерзіміне
байлансты ӛзгерісі айқын байқалады (кесте 13).
Кесте 13 - Іле Алатауы жабайы жемісті орман белдеуіндегі орманның күңгірт-
сұр топырақтары бактерияларының жалпы мӛлшерінің динамикасы
Бұл топырақтың 5-10 см қабатындағы бактериялардың сандық мӛлшері
кӛктемнің мамыр айында 18,2×106 болса, жаз мезгілінің шілде айында
14,6×105 дейін тӛмендеген, ал күздің қазан айында олардың мӛлшері сәл ӛсіп
15,6×106 жеткен. 10-20 см қабатындағы бактериялардың сандық мӛлшері
кӛктем мезгілінде яғыни мамыр айында бактериялардың сандық мӛлшері
9,7×106
болса, жаз мезгілінің шілде айында азайып 8,2×105
жеткен, күздің
қазан айында олардың мӛлшері қайтадан ӛсіп 8,8×106 жеткен. Ал 10- 20
Үлгі терең
дігі,
см
Күні 1 г топырақ
тағы
бактерия
лардың саны
Күні 1 г топырақ
тағы
бактерия
лардың саны
Күні 1 г топырақ
тағы
бактерия
лардыңсаны
5-10
10-20
20-30
V.2014
18,2×106
9,7×106
5,7×106
VII.2014
14,6×105
8,2×105
3,2×104
X.2014
15,6×106
8,8×106
4,2×106

70
қабатындағы бактериялардың сандық мӛлшері кӛктем мезгілінде яғни мамыр
айында бактериялардың сандық мӛлшері 5,7×106 болса, ал жаз мезгілінде
шілде айында 3,2×104 болды, күз мезгілінде қазан айында олардың мӛлшері
4,2×106 болды. Сӛйтіп, Іле Алатауы орманның күнгірт-сұр топырақтарындағы
бактерияларының жалпы мӛлшері, осы аймақ топырақтарына тән екендігі
және олардың мӛлшерінің кескін бойы күрт тӛмендейтіндігімен, жыл
мезгілдері бойынша кӛктемнен жазға және күзге қарай азайғандығы
байқалады.
Жетісу Алатауы жабайы жемісті орман белдеуіндегі орманның күңгірт-
сұр топырақтарының (кесте 14) 5-10 см қабатындағы бактериялардың сандық
мӛлшері кӛктем мезгілінде, яғни мамыр айында 16,5×106 болса, ал жаз
мезгілінде шілде айында 12,3×105 дейін азайған, күздің қазан айында олардың
біраз мӛлшері ӛсіп, 14,1×106 жеткен. Сонымен қатар, 10-20 см қабатындағы
бактериялардың сандық мӛлшері кӛктем мезгілінде, яғыни мамыр айында
бактериялардың сандық мӛлшері 8,5×106 болса, ал жаз мезгілінде шілде
айында 6,8×105 болды, күз мезгілінде қазан айында олардың мӛлшері 7,5×10
6
болды. Ал 20-30 қабатындағы бактериялардың сандық мӛлшері кӛктем
мезгілінде яғни мамыр айларында бактериялардың сандық мӛлшері 5,2×106
болса, ал жаз мезгілінде шілде айында 2,4×104 болды, күз мезгілінде қазан
айында олардың мӛлшері 4,7×106 болды.
Кесте 14 Жетісу Алатауы жабайы жемісті орман белдеуіндегі орманның
күңгірт сұр топырақтары бактерияларының жалпы мӛлшерінің динамикасы
Топырақтағы бактериялардың кескін бойы бұлай болып ӛзгеруін ӛсімдік
жамылғысы құрамымен, ауаның температурасымен, ылғалдылық және
аэрациямен түсіндіруге болады. Микроорганизмдердің қарқынды кӛбейюі
топырақ жағдайларына тығыз байланысты. Микроорганизмдердің ӛскен
ортасы қолайлы болғанда, олар тез кӛбейеді. Топырақтың беткі қабаты ӛсімдік
жамылғысымен толық қамтамасыз етілген болып, онда қоректік заттар
жеткілікті болады. Сонымен қатар кӛктем, күз мезгілінде мұнда жылумен
ылғалдылықта жеткілікті болады. Микроорганизмдер клеткаларының 85
проценттейі судан тұрады. Сондықтан олар-дың тіршілік әрекеті ылғалды
жерде ғана жүреді. Сонымен бірге, осы уақыттарда температурада
оптимальды жағдайда болады. Осыларға сәйкес, кӛктем және күз мезгілінде
бактериялардың сандық мӛлшері жаз мезгіліне қарағанда салыстырмалы түрде
жоғары болып табылады.
Үлгі
тереңдігі,
см
Күні 1 г топырақ
тағы
бактерия
лардың саны
Күні 1 г топырақ
тағы
бактерия
лардың саны
Күні 1 г топырақ
тағы
бактерия
лардың саны
5-10
10-20
20-30
V.2014
16,5×106
8,5×106
5,2×106
VII.2014
12,3×105
6,8×105
2,4×104
X.201
4
14,1×106
7,5×106
4,7×106

71
Бактериялар мӛлшерінің мұндай ӛзгеріске ұшырауына топырақтың
температуралық және дымқылдылық жағдайына тікелей байланысты.
Топырақтағы микроорганизмдердің саны жыл маусымына байланысты
ӛзгеріп отырады. Қыста азайып кетсе, жазда қайтадан кӛбейеді. Кӛптеген
ғалымдар қыс кезінде топырақта тіршілік тежеледі, тіпті тоқтап қалады деген
пікір айтып жүр. Ал Н.А.Красильниковтың байқауынша, қалың қар астындағы
топырақ тоңға айналмай, ондағы микробиологиялык процестер үздіксіз жүріп
жатады. Кӛктемде топырақта бактериялардың дамып ӛсуіне оптималды
гидротермиялық жағдайдың болуы, олардың жалпы сандық мӛлшерінің жыл
мерзімдерінде ең жоғарғы кӛрсеткішіне тең. Ал жаз айында топырақта
дымқылдылықтың азайуы және температураның шамадан тыс жоғарылауына
байланысты бактериялардың дамуына теріс әсерін тигізген. Сол себепті
олардың саны кӛктемге қарағанда едәуір тӛмендеген. Күзде температураның
тӛмендеуімен, күз айларындаға жауын-шашынның топырақта ылғалдылықтың
кӛтерілуіне байлансты бактериялардың жалпы саны аздап кӛбейген.
4.4 Жетісу және Іле Алатауларының жабайы жемісті орман
топырақтары мен бунақденелілерден бӛлініп алынған, энтомопатогенді
Bacillus thuringiensis бактериялары Соңғы онжылдықта Қазақ ӛсімдік қорғау және карантин ҒЗИ-ның
биотехнология зертханасында, энтомопатогенді бактериялар негізінде
биологиялық препараттардың продуценттері зерттелуде.
Табиғи жағдайда микроорганизмдер тек аралас популяция түрінде
кездеседі. Олардың ішінен керекті бактерияларды таза дақылдар түрінде бӛліп
алады. Таза дақылдарды әр түрлі әдіспен тікелей табиғи субстраттан (судан,
топырақтан, ӛсімдіктерден, жәндіктерденесінен) немесе элективті және
жинағыш қоректік ортадан бӛліп алынады [159].
Зерттеу жұмысында Жетісу және Іле Алатауының жабайы жемісті
орманның күнгірт - сұр топырағынан Кох әдісімен энтомопатогенді
бактерияларының 48 штамының таза дақылы бӛлініп алынды. Оларды бӛліп
алу үшін оның әртүрлі биіктіктерінде 640 m, 1000 m, 1500 m орналасқан
топырақтарының әрқайыссынан 5 нүктеден жалпы 15 үлгі алынды (cурет 11).
Сурет 11 - Зерттеуге алынған топырақ үлгілері

72
Топырақтан дайындалған суспензияны 70 C0
температураға жеткізіп
қыздырылады. Петри табақшасындағы арнайы А қоректік ортасына 0,25 мг
суспензия құйып, шпательмен жаймалап, беттік егу жүргізіледі. Шпательдегі
жұғындыдан екінші және үшінші Петри табақшаларына қайталап егу
жүргізіледі. Оларды термостатта 5-6 тәулік бойы 28 0С ӛсіру жүргізіледі.
Егу алдында стерильді Петри табақшаларына 10-15 мл-ден ерітілген
қоректік ортаны құйып, ішіндегі қоректік орта қатқанша кӛлденең күйде
қалдырылып, оның жоғарғы бетіндегі конденсацияланған суды жою үшін
табақшалардың қақпағын тӛмен қаратып, бір күнге залалсыздандырылған
боксқа қойылды. Дайын болған қореттік ортаға стерильді пипеткамен 0,1 мл-
ден 0,05 мл аралығында топырақ суспензиясынан алып егіп,
заласыздандырылған шпательмен жаймаланды. Бұл әдісті бірнеше рет
қайталап таза дақыл бӛліп алмғанша жүргізіледі. Ӛсіп шыққан колонияларды
санау, Bt.-ды санауды 3-4 күннен кейін микроскоптық анализ арқылы B.cereus
және Bt екенін анықтағаннан соң жүргізіледі. Ең жақсы сұйылту болып «А»
ортада 30-50 колониядан 100-150 колонияға дейін ӛсіп шыққан, сұйылту
болып табылады (сурет 12 а). Ерекше колонияны таңдап алып, тұзақпен жаңа
қоректік ортаға штрихтап егеміз (сурет 12 б). Петри табақшасында штрих пен
ӛскен колонияларды қиғаш «А» қоректік ортасына егіп алып сақтауға
жібереміз (сурет 12 в).
а Б В
Сурет 12 - Топырақтан бӛліп алынған таза культуралар
Таза культураларды терең қабатта ӛсіру әдісінде қоректік орта
араластырылып отырғызылады, ал бактерия клеткалары тӛменгі қабатта ӛседі.
Бұл әдісте арнайы құрылғы шайқағыш қолданылады. Таза культураларды «А»
сұйық ортасы пептон – 1 %, балық гидролизаты – 4 %, NaCl – 5 %, сұйық
қоректік ортасында шайқағышта ӛсіреміз, алды мен алынған таза культураға
сұйылту жасап, оның титрі Том Гораев камерасында саналып есептелінді.
Сұйылтуды стандарттық мӛлшермен 4 мл дистилденген суға жасалынды.
Қоректік ортаның рН-7. 500 мл колбаларға 250 мл ден қоректік ортаны құйып
сұйылтылған бактериалды таза дақылдан 0,5 мл ден қоректік ортаға еңгізіп,

73
шайқағышқа 5 тәулікке 28 0С температураға қойылды. 3 тәуліктен соң
колбаларды алып бекітілген препарат Пешков әдісімен дайындалып боялады.
Оларды микроскоппен зерттеп, тазалығы тексеріледі. Сонымен бірге
вегетативті клеткалардың спора түзу кезеңін байқауға болады. Колбалар
қайтадан, шайқағышқа 3 тәулікке қойылады, сонан соң сұйық қоректік ортасы
бар колбаларда штаммдардың ӛскенін, қоректік орталар түсінің сарғыш-қоңыр
түске ӛзгеруінен байқауға болады (cурет 13а). Вегетативті клеткалары
тізбектелген, таяқшалар түрінде байқалады (cурет 13б). Микроскоптау арқылы
олардың кристаллдары мен споралары зерттелді. Зерттелген микроскоптық
аймақта ортада бӛгде микроорганизмдердің болмауы, дақылдың тазалығын
білдіреді. Микроорганизмдердің жақсы дамуына қоректік орталар құрамының
дұрыс құрастырылғаны, оларға оптималды жағдай жасайды (сурет 14,15).
Микроорганизмдер егілген қоректік ортаға температурасы үнемі бақылап
отырылуы қажет, Bt дақылдарына қолайлы температура 28-30 0С болады.
А Б
Сурет 13 - Бӛліп алынған таза ӛсінділерді колбаларда терең қабатта ӛсіру
Сурет 14 - А қоректік ортасында Bacillus thuringiensis энтомопатогенді
бактерияларын петри табакшаларының ортасына егілген түрі

74
Сурет 15 - А қоректік ортасында Bacillus thuringiensis энтомопатогенді
бактерияларын пробиркаларға егу
Жетісу және Іле Алатауы МҰТС жүргізілген маршруттық бақылаулар
барысында Bacillus thuringiensis энтомопатогенді бактериясының бар жоғын
анықтау мақсатында Сиверс алмасының түбінен топырақ үлгілері алынды.
Сонымен, барлығы 30 үлгі алынып, олардан 48 бактерия изоляттарының таза
ӛсіндісі бӛлініп алынды. Изоляттар микроморфологиялық зерттеулер
бойынша Bacillus thuringiensis түріне жатқызылды.
Жетісу және Іле Алатауларының жабайы жемісті орман
топырақтарындағы энтомопатогенді Bacillus thuringiensis және спора түзуші
бактериялардың 1 г топырақтағы мӛлшері салыстырмалы зерттелді (сурет 16).
Зерттеу нәтижелері бойынша Іле Алатауында спора түзуші бактерия мен
Bacillus thuringiensis бактерияларының сандық мӛлшері Жетісу Алатауына
қарағанда салыстырмалы жоғары екендігін кӛрсетті. Мұнда Жетісу Алатауы
топырақтарында pH- 6,8, Іле Алатауында pH-7,2, яғни ортаның мұндай
кӛрсеткіші спора түзуші бактериялармен Bacillus thuringiensis
бактерияларының кӛбеюі үшін ӛте қолайлы жағдай болып табылады.
Сурет 16 - Іле Алатауында спора түзуші бактерия мен Bacillus
thuringiensis бактерияларының сандық мӛлшерлері

75
Жетісу және Іле Алатауының бӛктеріндегі жүргізілген зерттеулер
барысында бунақденелілердің бактериоз белгілері бойынша 21 үлгі алынды.
олардың ішінде басым бӛлігі алма күйе кӛбелегі және жапырақ ширатқыш
кӛбелегінің ӛлі денелері. Осы 21 үлгіден Bacillus thuringiensis
бактерияларының таза дақылдарының 7 изоляты бӛлініп алынды (сурет 17).
Сурет 17 - Бактериоздық белгімен ӛлген жұлдызқұрт
Топырақ пен бактериоздық белгімен ауырып ӛлген жәндіктерден бӛліп
алынған ӛсінділердің морфо-культуралдық қасиеттерін қарастыру үшін
сонымен бірге олардың түрлі қоректік орталарда ӛсуі мен кристалл түзу
қабілетін бақылау үшін құрамы әр түрлі тығыз қоректік орталарға нүктелеп
егіп, ӛсірілді: «А» ортасы, қоректік агар-ПА, қорексіз агар-ГА, ЕПА (сурет-
18).
А Б
В Г
а – Қоректік агарда(ПА); б – «А» ортада; в – қорексіз агарда(ГА); г –ЕПА
Сурет 18 - B. Thuringiensis бактерия штаммдарының әртүрлі тығыз
қоректік орталардағы макроморфологиясы

76
Bacillus thuringiensis бактерияларының қоректік ортадағы культуралдық
қасиетін зерттегенде қоректік ортада ӛскенде айырмашылықтар болмады.
Колонияларының бұрыс пішінді, сары, ақшыл сұр түсті, шеті толқынды,
конфигурациялары дӛңгеленгендігі анықталды. Рельефі – тегіс,
консистенциясы – паста тәрізді.
Bacillus thuringiensis бактерияларының сурет 18- де кӛрсетілгендей әр
түрлі тығыз қоректік орталарға ӛсіргенде «А» қоректік ортасында ең жақсы
кӛрсеткіш кӛрсетті. Мұнда«А» қоректік ортасында ӛскен колониялар қоректік
агарлы ортадағы колониялардан екі есесіндей артық (cурет 19).
Сурет 19 - Жетісу Алатауының жабайы жемісті орман топырақтарынан
2013-жылы бӛлініп алынған Bacillus thuringiensis бактерия штаммдарының
әртүрлі тығыз қоректік орталарда 60 сағат ӛсу динамикасы
Сурет 20 - Жетісу Алатауының жабайы жемісті орман топырақтарынан
2014-жылы бӛлініп алынған Bacillus thuringiensis бактерия штаммдарының
әртүрлі тығыз қоректік орталарда 60 сағат ӛсу динамикасы

77
Сурет 21 - Іле Алатауының жабайы жемісті орман топырақтарынан
2013-жылы бӛлініп алынған Bacillus thuringiensis бактерия штаммдарының
әртүрлі тығыз қоректік орталарда 60 сағат ӛсу динамикасы
Сурет 22- Іле Алатауының жабайы жемісті орман топырақтарынан 2014-
жылы бӛлініп алынған Bacillus thuringiensis бактерия штаммдарының әртүрлі
тығыз қоректік орталарда 60 сағат ӛсу динамикасы
Микроорганизмдердің дақылдық ерекшеліктері қатты қоректік орталарда
ӛсу белгілерімен анықталды. Әрбір микроорганизмге дақылдық қасиеттер тән
болғандықтан олар маңызды диагностикалық белгі болып табылады. Қатты
қоректік ортаның бетінде микроорганизмдерді егу түріне қарай колония,
штрих, жаймаланып ӛсуі мүмкін.
Қоректік ортаның бетіне ӛсіп шыққан колониялар пішіні, ӛлшемі, беті,
түсі, колониялардың шеті, консистенциясы бойынша суреттелді. Дақылдар
эозин ерітіндісімен боялды. Микроскоптау нәтижелері вегетативті клеткалары
жағындыда бірітіндеп немесе тізбектеліп орналасқан ірі таяқша, споралары
сопақша болатынын кӛрсетті. Вегетативті жасушалардың ӛлшемдері 0,8-1,5 х
3,5-5 мкм аралығында (сурет 23).

78
Сурет 23 - Bacillus thuringiensis штамының вегетативті клеткаларының
кӛрінісі (х100)
Морфологиясы Пешков әдісі бойынша бояумен анықталды. Заттық
шыны бетіне дистилденген су тамызып, бетіне тұзақпен бактерия дақылын
жағып, ауада кептіріп, спирт шам жалынынан ӛткізіп бекітілді. Сосын бетіне
Леффлер бойынша метилен кӛгін тамызып, 3 мин бетінде буы шыққанша
спирт шам жалынында ұсталды. Содан соң сумен шайып, үстіне сафронин
қызылын тамызып 1 мин ұстап, сумен шайып, иммерсиялық май тамызып
қарадық. «А» қоректік ортасында бактерия дақылдарының колониясы
дӛңгелек, дұрыс емес пішінді (S – пішінді), сұрғылт-сарғыш түсті, тегіс
пішінді, беті тегіс немесе бұдыр болды. Бӛлініп алынған изоляттардың
ӛлшемдері 0,7 – 1,7 см аралығында болды. Консистенциясы паста тәрізді.
Спора ӛлшемі 0,8 х 1,3 мкм. Дақылдар екі типті кристаллдар түзеді: ірі
бипирамидалы және ұсақ шаршы тәрізді. 28-30ºC термостатта 5-6 тәулік ӛскен
соң споралар мен кристаллдар толығымен түзіледі (сурет 24).
Сурет 24 - Bacillus thuringiensis штамының кристаллының кӛрінісі
(х100)

79
Тӛмендегі кестелерде минерал май және резеңкелі тығын астында
сақталған штамдардың дақылдық-морфологиялық қасиеттері зерттелді.
Дақылдық қасиеттері колонияларына қарап сипатталды. Морфологиясы
Пешков әдісі бойынша бояп зерттелінді (сурет 25,26).
Дақылдық қасиеттері Морфологиялық қасиеттері
Bacillus thuringiensis BSJA3 – 13
Пішіні дӛңгеленген, беті тегіс,
түсі сары, шеті ризоидты.
Споралары сопақ, кристаллдары ірі
шашырап орналасқан.
Bacillus thuringiensis BSJA5 – 13
Бұрыс пішінді, беті тегіс, құрғақ, түсі
ақшыл сары, шеті ризоидты.
Споралары мен кристаллдары
бірдей кӛлемді.
Bacillus thuringiensis BSJA6 – 13
Бұрыс пішінді, беті тегіс, түсі
сары,шеті ризоидты.
Кристаллдары мен споралары
шашырап орналасқан. Кристалдары
ұсақ.
Сурет 25 - Жетісу Алатауының жабайы жемісті орман топырақтарынан бӛліп
алынған Bacillus thuringiensis энтомопатогенді штаммдарының дақылдық-
морфологиялық белгілері

80
Дақылдық қасиеттері Морфологиялық қасиеттері
Bacillus thuringiensis BSIA1 – 13
Бұрыс пішінді, беті әжімді,
құрғақ, түсі ақшыл сары, шеті
ризоидты.
Споралары сопақ, кристаллдары
ірі шашырап орналасқан.
Bacillus thuringiensis BSIA6 – 14
Бұрыс пішінді, беті тегіс,
құрғақ, түсі ақшыл сары, шеті
толқынды.
Споралары мен кристаллдары
бірдей кӛлемді.
Bacillus thuringiensis BSIA7 – 14
Бұрыс пішінді, беті тегіс, түсі
сары, шеті толқынды.
Кристаллдары мен споралары
шашырап орналасқан. Кристалдары
ұсақ.
Сурет 26 - Іле Алатауының жабайы жемісті орман топырақтарынан бӛліп
алынған Bacillus thuringiensis энтомопатогенді штаммдарының дақылдық-
морфологиялық қасиеттері

81
Дақылдардың негізгі бӛлігі қоректік ортада ӛскенде айырмашылықтар
болмады. Колониялары бұрыс пішінді, сары, ақшыл сұр түсті, шеті
толқынды, конфигурациялары дӛңгеленген. Рельефі – тегіс, консистенциясы
– паста тәрізді.
Қоректік орта бетіне ӛскен калонияларды микроскоп арқылы зерттеу
барысында олардың токсиндері мен спораларын грамм әдісімен бояу арқылы
кӛруге болады.
Дақылдардың морофологиялық қасиеттерінде де айтарлықтай
айырмашылықтар болмады, спора, кристалл түзу қабілеттері болды. Пешков
әдісімен бояғанда споралары сопақша кӛк түске боялды, ал кристалдары
қызыл түске боялса, олар ұсақ және ірі ромб тәрізді болды.
Топырақтан бӛлініп алынған Bacillus thuringiensis бактерия
штамдарының биохимиялық қасиеттеріне талдау жасайтын болсақ, Bacillus
thuringiensis энтомопогендік бактерияларының улы әрекеті бірнеше
факторлар негізінде қарастырылды. Bacillus thuringiensis энтомопатогендік
споралық бактериялардың дақылдарына ерекше параспоралық
қосылыстардың түзілуі бірнеше зерттеушілермен оның энтомопатогендік
әрекетінің маңызы жасалмастан бұрын айтылған болатын. Ең бірінші Хенней
бұл түзілулерді оны түзетін бактериялардың энтомоцидтік әрекетімен
байланысына негіздеді. Зерттелініп жатқан энтомопатгенді бактериялардың
таза дақылдарының кӛмірсуларға қажеттілігін анықтау түрлі қанттарды
қоректік орталарға қолданумен жүргізілді. Кӛмірсуға бай орталарда
энтомопатогенді бактерия ӛсу барсында дақылдарының қышқыл түзу, газ
түзуімен байқалды. Қышқыл түзілуін қышқылдық белсенділігінің ӛзгеруімен
(рН) ал газ түзілуін беткі қабатта пайда болған кӛпіршіктермен байқалаы.
Лецитиназа тығыз қоректік ортада анықталды. Рекция жүргізу үшін 90 мл
МПА ны, 500 С салқындатылғаннан кейін, оған жұмыртқаның сарысын қосып
оны Петри табақшаларына құйямыз. Фильтрленген қағаздардың стерильды
дисксі (d=15) мм Петри табақшасындағы қоректік ортаның бетіне қоямыз.
Зерттелетін культуралардың бірдей бӛліністегі дискінің ортасына ілмекпен
алып егіп, 24 сағатқа инкубирлеуге термостатқа (28 0С) қоямыз. Қалыпты
әрекеттесу кезінде дикстің аумағында агардың колониясы түзіледі.
Негізгі биохимиялық дифференциалдық қасиеттерін Bt, Н. Баржактың,
және Бонфуаның тәжірибелері негізінде жасалынды. Тәжірибеде ӛсінділерінің
ацетилметилкарбинолға(АМК), реакциясы, кӛміртегін қажет ету ерекшелігі,
уреаза түзуі, пигмент түзу және крахмалға гидролиз процесстерін жүргізу
арқылы зерттелінді.
Ацетилметилкарбинол(АМК) анықтау Кларк қоректік ортасында
жүргізілді. Кларк қоректік ортасының құрамы: пептон – 0,5 %; глюкоза – 1,0
%; К2HPO4 – 0,1 % немесе NaCL 0,65 %. 3 мл ӛсінділерін Кларк қоректік
ортасына сеуіп, 280С 3 тәулік бойы 5 % альфа нафтол ерітіндісінен 0,6мл
қосамыз. Ацетилметилкарбинол қызыл түске боялуы 2-5 минуттан кейін
немесе 1 сағатта белгілі болады. Ацетилметилкарбинолды синтездеуінің жүруі
ортаның қызыл түске бояйды.

82
Бактерия ӛсінділерінің қышқыл түзуі манноза мен салицинада
кӛмірсулары негізінде анықталды. Негізгі ерітінді құбыр суында дайындалды.
Оның құрамы: пептон, 0,1 % К2HPO4. 1,0 %, кӛміртегі мен спиртті 1 %
кӛлемінде қосамыз. Реакция pH=7-7.4 жүргізілді. pH ӛзгерісін анықтау үшін
бромкреозолпур индикаторының 2 мл 1,6 % спирттік ертінді түрінде 1 мл
ортаға қосамыз. Ӛсіндіні ӛңдеу үшін 3 - 4 мл кӛлемінде пробиркаға себеміз
және оны термостатқа қоямыз. Екі-үш тәуліктен кейін қышқыл pH 6,8
түзілгенде орта қызыл түске ӛзгерді және pH 5,2 болғанда орта сары түске
ӛзгерді. Бұл рН 6,8 және рН 5,2 кӛрсеткіштеде ортада қышқыл түзілуі
байқалды. Мұны түстердің ӛзгеруімен байқаймыз (сурет27 ). Ал сахарозада
қышқыл түзуі байқалмады.
Сурет 27 - Кӛмірсуларды пайдалану әрекеттері (салицина, манноза)
Уреазаның қалыптасуын келесі құрамды қоректік ортада анықтадық:
пептон 0,1 %; NaCl, KHPO 0,1 % ; фенолрот 0,6 % (0,2 % - дық су
ерітіндісінде); агар- 2,0 %. pH = 6,8. Орта автоклавта стерильденеді. Бӛлек 20
%-дық карбамид ерітіндісі дайындалып, бір сағаттан кейін 10 мл карбамидке
50 0С салқындатылған 90 мл қоректік ортаны қосып, қосындыны пробиркаға
құйып, ӛсінділерін штрихпен қоректік ортаға ектік. Реакция жүргенін ортада
қызыл түстің пайда болуымен байқаймыз (сурет 28).
Сурет 28 - Ӛсінділердің уреазаға әсері

83
Крахмалда гидролиз үрдісінің жүруі үшін ӛсінділерін ЕПА 2 %-дық
қоректік ортасына нүктелеп ектік. Ӛсінділерді ӛсіру барысында 3-4 күнге
дейін қоректік ортаға люголь ерітіндісінен бірнеше тамшы тамызып отырдық.
Сондықтан, орта йодтың әсерінен кӛк түске боялады. Гидролиз үрдісі жүре
бастаса, нүктелеп егілген ӛсінділер айналасында жарықтану пайда болып,
колониялардың ӛсуі байқалады (сурет 29).
Сурет 29 - B. thuringiensis ӛсінділерінің крахмал ортасында гидролизденуі
Жарықтану айналасы мен ӛскен колонияның айналасының диаметрін 5
тәулікке дейін әр тәулікте ӛлшеп жазып отырылды (сурет 30).
Протеолитикалық белсенділікті модифицирленген Аисон әдісі негізінде
казеин агар ортасында анықталынды. Сонан соң казеин агар ортасын
дайындау үшін «А» ортасына 1/3 кӛлемде стерильді майсыз сүтті қосамыз
Казеин агарлы ортасына бӛліп алған ӛсінділерді нүктелеп егіп, колонияларды
сол ортада ӛсіру барысында колония айналасының жарықтануы арқылы
фермент түзу барысы анықталынды. Фермент белсенділігін 1 мм-мен ӛлшенді.
Сурет 30 - Энтомопатогенді дақылдардың протеолитикалық қасиеті

84
Барлық ӛсінді сүт пептонды ортада 4 тәулік бойы ӛсірілді. 3-ші тәулікте
крахмалға гидролиз үрдісі жүрді, 4 тәулікте каталазаға оң әсер берді. ЕПА
қоректік ортасында ӛсіргенде 3 тәулікте қоректік орта бетінде жұқа
қабықшалар пайда бола бастады (cурет 31).
Сурет 31 - ЕПА қоректік ортасында дақылдардың жіңішке қабық түзуі
Энтомопатогенді бактериялар ӛздері ӛсетін ортада кӛмірсуларды қажет
етеді. Кӛмірсулар дақылдардың спора мен кристалл түзулуіне жұмсалатын
энергия кӛзі болып табылады (кесте 15,16).
Бӛліп алынған энтомопатогенді таза ӛсінділер ацетилметилкарболды
синтездейді, лецитовителленге оң әсер етеді. Эталонды дақылдарында уреаза
ферменті байқалмады. Салицинада басым кӛбі қышқыл түзеді, BSJA3– 13,
BSJA10– 13, BSJA1 – 14, BSIA15– 13, BSIA1 – 14, қышқыл түзбейді. Маннозада
кӛп сандысы қышқыл түзеді, BSJA11– 13, BSJA5 – 14, BSIA13– 13, BSIA15– 13,
BSIA18– 13, қышқыл түзбейді. Дақылдар сахарозада қышқыл түзбейді.
Эскулинді гидролиздейді. Саруызды ортада дақылдар пигмент түзбейді.
Протеолитикалық және аминқышқылдық реакциясы оң. Дақылдардың
кристалл ақуызының құрылысы ӛзара ерекшеленеді. Оларды клондау
барысында β-эндотоксинді ӛзінде сақтап қалады. Электрофорез әдісінде спора
кристалдық дақыл белогының молекулярлы массасы екі түрлі 60-40 кДа. Ромб
тәрізді.

85
Кесте 15 - Жетісу Алатауы жабайы жемісті орман топырақтарынан бӛлінген бактериялардың биохимиялық қасиеттері
Штаммдар Ацетил-
метилкар
бинол
(АМК)
Лецитови
телинді
реакция
(ЛВР)
Протеолиз Амилаза Уреаза Каталаза Қышқыл түзуі
Сахароза Манноза Салицин
BSJA1 – 13 + + + + – + – + +
BSJA2 – 13 + + + + – + – + +
BSJA3– 13 + + + + – + – + –
BSJA4 – 13 + + + + – + – + +
BSJA5 – 13 + + + + – + – + +
BSJA6 – 13 + + + + – + – + +
BSJA7 – 13 + + + + – + – + +
BSJA8 – 13 + + + + – + – + +
BSJA9 – 13 + + + + – + – + +
BSJA10– 13 + + + + – + – + –
BSJA11– 13 + + + + + + – – + BSJA12– 13 + + + + + + – + + BSJA1 – 14 + + + + – + – + – BSJA2 – 14 + + + + – + – + + BSJA3 – 14 + + + + + + – + + BSJA4 – 14 + + + + – + – + + BSJA5 – 14 + + + + + + – – + BSJA6 – 14 + + + + + + – + + BSJA7 – 14 + + + + – + + + BSJA8 – 14 + + + + + + – + +
Ескерту: ++ - биохимиялық реакциясы қарқынды;
+- биохимиялық реакциясы оң
– - биохимиялық реакциясы теріс

86
Кесте 16 - Іле Алатауы жабайы жемісті орман топырақтарынан бӛлінген бактериялардың биохимиялық қасиеттері
Штаммдар Ацетил-
метилкар
бинол
(АМК)
Лецитови
телинді
реакция
(ЛВР)
Протеолиз Амилаза Уреаза Каталаза Қышқыл түзуі
Сахароза Манноза Салицин
1 2 3 4 5 6 7 8 9 10 BSIA1 – 13 + + + + – + – + ++ BSIA2 – 13 + + + + – + – + ++ BSIA3 – 13 + + + + – + – + ++ BSIA4 – 13 + + + + – + – + ++ BSIA5 – 13 + + + + – + – + ++ BSIA6 – 13 + + + + – + – + ++ BSIA7 – 13 + + + + – + – + ++ BSIA8 – 13 + + + + – + – + ++ BSIA9 – 13 + + + + – + – + ++ BSIA10 – 13 + + + + – + – + ++
BSIA11– 13 + + + + + + – + +
BSIA12– 13 + + + + – + – + +
BSIA13– 13 + + + + + + – – +
BSIA14– 13 + + + + – + – + +
BSIA15– 13 + + + + + + – – –
BSIA16– 13 + + + + – + – + +
BSIA17– 13 + + + + + + – + +
BSIA18– 13 + + + + + + – – +
BSIA19– 13 + + + + – + – + +
BSIA20– 13 + + + + – + – + +
BSIA1 – 14 + + + + + + – + –

87
16 – кестенің жалғасы
1 2 3 4 5 6 7 8 9 10
BSIA2 – 14 + + + + + + – + +
BSIA3 – 14 + + + + – + – + +
BSIA4 – 14 + + + + + + – + +
BSIA5 – 14 + + + + + + – + +
BSIA6 – 14 + + + + – + – + +
BSIA7 – 14 + + + + – + – + +
BSIA8 – 14 + + + + + + – + +
Ескерту: ++ - биохимиялық реакциясы қарқынды;
+- биохимиялық реакциясы оң
– - биохимиялық реакциясы теріс

88
4.5 Бӛлініп алынған дақылдар уыттылығының әр-тҥрлі
бунақденелілерге әсері
Жетісу және Іле Алатауы жабайы жемісті орманның күңгірт-сұр
топырақтарынан және бактериоздық белгісі бар жәндіктерден бӛлініп алынған
энтомопотогенді Bacillus thuringiensis бактерия штаммдарының
бунақденелілерге уыттылығын, яғни биологиялық белсенділігін тексеру үшін
бактерия дақылдарынан сұйылтулар жасалды. Пробиркада қиғаш агарда ӛсіп
тұрған бактерия дақылдарына 4 мл-ден дистилденген су құйылып, жақсылап
араластырылды. Сұйылтулардың титрін анықтау үшін Том Горяев камерасында
споралар саналды. Спора титрі жоғары болды. бунақденелілерді залалдауға
қажетті спора титрі 1x107
болғандықтан, пробиркаларға 8 мл-ден қайта су
қосылды. Дайын болған суспензияларға жапырақтарды малып, 2-3 жасар
жұлдызқұрттарға беріп, ауа кіретін тесіктері бар стақандарға салынды. Әр
стақанға 10 жұлдызқұрттан салынды. Әрқайсысы үш қайталаудан жасалды.
Жұлдызқұрттарды күнделікті бақылап, 5-7 күн 25 оС температурада ұстады.
Содан кейін ӛлгендері пайыздық кӛрсеткішпен анықталынды [160].
4.5.1 Bacillus thuringiensis бактерия штаммдарының жапырақ ширатқыш
кӛбелегінің жұлдызқұрттарына қарсы биологиялық белсенділіктері
Жапырақ ширатқыш жұлдыз құрттары алма және орман ағаштарының
зиянкесі. Ол ағаштардың жапырақтарын жеп, сол ортада ӛмір сүреді. Тӛменде
(кесте 20) Жетісу Алатауының жабайы жемісті орманның күнгірт-сұр
топырақтарынан бӛлініп алынған Bacillus thuringiensis энтомопатогенді
бактерия штаммдарының насекомдарға уыттылығы зертханалық жағдайда
сыналды.Тәжірибеде спора титры 1х107 қолданылды. Мұнда жұлдыз
құрттардың 5-тәуліктегі ӛлу мӛлшері салыстырмалы түрде жоғары болған;
BSJA1 – 14, BSJA4– 14, штаммдарының ӛлу мӛлшері 90 % кӛрсетті. BSJA1 – 13,
BSJA2 – 13, BSJA8 – 13, BSJA10 – 13, BSJA2 – 14, BSJA3 – 14, штаммдары 92 ,5
% кӛрсетті. BSJA3 – 13, BSJA9 – 13, BSJA8 – 14, 95 % кӛрсетті. BSJA5 – 13,
BSJA5 – 14, штаммдары 97,5 % жоғары кӛрсеткіш кӛрсетті. BSJA6 – 13, 100 %
кӛрсетті. Бақылау варианты 2127-3к, 100 % кӛрсетті (кесте 17).
Кесте 17 - Жетісу Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛлініп алынған Bacillus thuringiensis энтомопатогенді
бактерия штаммдарының жапырақ ширатқыш кӛбелектің жұлдызқұрттарына
биологиялық белсенділігі Тәжірибе
нұсқасы
Спора
титры
Жұлдыз құрттардың ӛлу мӛлшері, %
1-тәулік 2-тәулік 3-тәулік 4-тәулік 5-тәулік
1 2 3 4 5 6 7
BSJA1 – 13 1х107 22,5±1,25 75±0,75 82,5±0,38 90±0,5 92,5±0,75
BSJA2 – 13 1х107 27,5±0,88 62,5±0,38 82,5±0,38 85±0,5 92,5±0,75
BSJA3 – 13 1х107 42,5±0,75 72,5±0,75 82,5±0,38 95±0,5 95±0,5
BSJA4 – 13 1х107 12,5±0,38 67,5±1,25 77,5±1,25 82,5±1,75 82,5±1,75

89
20 –кестенің жалғасы
1 2 3 4 5 6 7
BSJA5 – 13 1х107 22,5±0,88 60±1 80±0,5 87,5±0,38 97,5±0,38
BSJA6 – 13 1х107 40±1,5 60±1 80±0,5 90±0,5 100
BSJA7 – 13 1х107 20±1 80±0,5 80±0,5 80±0,5 80±0,5
BSJA8 – 13 1х107 30±1,5 75±1,25 90±1 92,5±0,75 92,5±0,75
BSJA9 – 13 1х107 20±1 70±1 82,5±0,38 92,5±0,38 95±0,5
BSJA10 – 13 1х107 17,5±0,38 80±1 85±1 87,5±0,75 92,5±0,75
BSJA11 – 13 1х107 7,5±1,13 75±1 82,5±1,25 82,5±1,25 87,5±1,38
BSJA12 – 13 1х107 5±0,75 80±1 80±0,5 80±0,5 80±0,5
BSJA1 – 14 1х107 32,5±0,38 75±1 60±3 90±0,5 90±0,5
BSJA2 – 14 1х107 32,5±8,8 62,5±0,88 92,5±0,75 92,5±0,75 92,5±0,75
BSJA3 – 14 1х107 22,5±0,88 80±0,5 90±1 90±1 92,5±0,75
BSJA4 – 14 1х107 17,5±1,25 62,5±1,25 87,5±1,38 90±1 90±1
BSJA5 – 14 1х107 22,5±1,25 80±1 95±0,75 97,5±0,38 97,5±0,38
BSJA6 – 14 1х107 15±0,75 55±1 75±1,25 82,5±1,13 82,5±1,13
BSJA7 – 14 1х107 15±1,5 45±0,75 77,5±1,38 82,5±1,13 85±1
BSJA8 – 14 1х107 27,5±1,25 70±1 85±0,5 95±0,5 95±0,5
2127-3к 1х107 57,5±4,7 87,5±2,5 90,0±4,1 95,0±2,8 100
Іле Алатауының жабайы жемісті орманның күңгірт-сұр топырақтарынан
бӛлініп алынған Bacillus thuringiensis энтомопатогенді бактерия
штаммдарының бунақденелілерге уыттылығы зертхана жағдайында сыналды.
Мұнда (кесте 18) жұлдыз құрттардың 5-тәуліктегі ӛлу мӛлшері; BSIA5 – 13,
BSIA6 – 14, 90 % кӛрсетті. BSIA1 – 13, BSIA6 – 13, BSIA7 – 13, BSIA7 – 14, 95 %
жоғары кӛрсеткіш кӛрсетті. Бақылау варианты 2127-3к, 100 % кӛрсетті.
Кесте 18 - Іле Алатауының жабайы жемісті орманның күнгірт-сұр
топырақтарынан бӛліп алған Bacillus thuringiensis энтомопатогенді бактерия
штаммдарының жапырақ ширатқыш кӛбелектің жұлдызқұрттарына
биологиялық белсенділігі Тәжірибе
нұсқасы
Спора
титры
Жұлдыз құрттардың ӛлу мӛлшері, %
1-тәулік 2-тәулік 3-тәулік 4-тәулік 5-тәулік
1 2 3 4 5 6 7
BSIA1 – 13 1х107 60±1 77,5±0,75 90±0,5 90±0,5 92,5±0,75
BSIA2 – 13 1х107 40±1,5 65±1,75 80±1 80±1 82,5±1,13
BSIA3 – 13 1х107 45±1 75±1,5 80±1 80±1 87,5±1,38
BSIA4 – 13 1х107 37,5±12,5 67,5±0,88 72,5±1 80±1 80±1
BSIA5 – 13 1х107 55±1 70±0,5 80±0,5 90±0,5 90±0,5
BSIA6 – 13 1х107 62,5±0,38 82,5±0,38 85±0,5 90±0,5 92,5±0,75
BSIA7 – 13 1х107 62,5±0,75 80±1,5 85±1 87,5±0,88 92,5±0,75
BSIA8 – 13 1х107 47,5±1,25 75±1 80±1 82,5±1,25 82,5±1,25
BSIA9 – 13 1х107 45±1 67,5±0,88 77,5±0,88 82,5±1,25 82,5±1,25
BSIA10 – 13 1х107 35±0,5 60±0,5 72,5±0,38 77,5±0,75 80±1
BSIA11 – 13 1х107 2,5±0,38 27,5±1,25 55±0,75 60±1 62,5±1,25
BSIA12 – 13 1х107 2,5±0,38 25±0,5 55±0,5 67,5±1,38 72,5±0,75
BSIA13 – 13 1х107 7,5±0,75 37,5±1,25 60±0,5 60±0,5 67,5±0,88
BSIA14 – 13 1х107 10±0,5 30±0,5 52,5±0,75 62,5±0,75 67,5±0,88

90
18 –кестенің жалғасы
1 2 3 4 5 6 7
BSIA15 – 13 1х107 2,5±0,38 25±0,5 42,5±0,75 55±1 62,5±0,75
BSIA16 – 13 1х107 10±1 25±1 45±0,75 52,5±0,75 52,5±0,75
BSIA17 – 13 1х107 7,5±0,75 25±1 52,5±0,75 55±0,5 67,5±0,88
BSIA18 – 13 1х107 10±0,5 30±0,5 52,5±0,38 62,5±0,75 62,5±0,75
BSIA19 – 13 1х107 12,5±0,75 32,5±0,75 55±0,5 62,5±0,75 62,5±0,75
BSIA20– 13 1х107 12,5±0,75 32,5±0,75 62,5±0,75 67,5±0,88 67,5±0,88
BSIA1 – 14 1х107 7,5±0,38 20±1 52,5±0,75 72,5±0,75 72,5±0,75
BSIA2 – 14 1х107 10±0,5 32,5±0,75 57,5±1,25 80±0,5 80±0,5
BSIA3 – 14 1х107 12,5±0,38 40±1 57,5±0,88 77,5±0,88 77,5±0,88
BSIA4 – 14 1х107 12,5±0,88 35±0,5 60±1 80±0,5 80±0,5
BSIA5 – 14 1х107 5±0,75 30±1 80±0,5 82,5±0,38 82,5±0,38
BSIA6 – 14 1х107 15±0,75 42,5±1,25 80±1 90±0,5 90±0,5
BSIA7 – 14 1х107 12,5±0,38 55±1,25 85±1 95±0,5 95±0,5
BSIA8 – 14 1х107 12,5±1,25 50±0,5 62,5±1,25 82,5±0,38 82,5±0,38
2127 -3к 1х108 20,0±4,1 52,5±11,1 75,0±6,4 95,0±1,3 100
4.5.2 Bacillus thuringiensis бактерия штаммдарының алма күйе кӛбелегінің
жұлдыз құрттарына биологиялық белсенділіктертері
Жетісу Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛлініп алынған Bacillus thuringiensis бактерия штаммдарының
алма күйе кӛбелегінің жұлдыз құрттарына биологиялық белсенділігі
зертханалық жағдайда анықталды. Мұнда (кесте 19) жұлдыз құрттардың 5-
тәуліктегі ӛлу мӛлшері 80 % дан жоғары болған. BSJA10 – 13, BSJA4 – 14, BSJA5
– 14, 90 % . BSJA5 – 13, BSJA9 – 13, BSJA1 – 14, BSJA2 – 14, BSJA3 – 14, 92,5 %,
кӛрсетті. BSJA8 – 14, 95 %. BSJA3 – 13, 97,5 % жоғары белсенділік кӛрсетті.
Бақылау варианты 2127-3к, 95 % кӛрсетті.
Кесте 19 - Жетісу Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛлініп алынған Bacillus thuringiensis энтомопатогенді
бактерия штаммдарының алма күйе кӛбелегінің жұлдыз құрттарына
биологиялық белсенділігі. Тәжірибе
нұсқасы
Спора
титры
Жұлдыз құрттардың ӛлу мӛлшері, %
1-тәулік 2-тәулік 3-тәулік 4-тәулік 5-тәулік
1 2 3 4 5 6 7
BSJA1 – 13 1х107 17,5±0,88 65±1 77,5±0,75 85±0,5 85±0,75
BSJA2 – 13 1х107 25±0,75 55±1 70±1 80±0,5 87,5±0,75
BSJA3 – 13 1х107 35±0,5 55±0,5 77,5±0,38 92,5±0,38 97,5±0,38
BSJA4 – 13 1х107 15±0,5 40±0,25 60±0,5 75±0,75 77,5±0,75
BSJA5 – 13 1х107 25±1 50±0,5 65±0,5 82,5±0,75 92,5±0,75
BSJA6 – 13 1х107 32,5±1,25 60±1 82,5±0,38 90±0,5 100
BSJA7 – 13 1х107 20±0,5 50±0,5 70±0,5 82,5±0,38 87,5±0,38
BSJA8 – 13 1х107 27,5±0,75 55±1,25 77,5±0,38 87,5±0,75 87,5±0,75
BSJA9 – 13 1х107 25±0,5 57,5±0,75 80±0,38 87,5,5±0,38 92,5±0,38
BSJA10 – 13 1х107 22,5±0,38 55±0,5 70±0,5 82,5±0,38 90±0,5
BSJA11 – 13 1х107 12,5±0,75 40±1 60±0,5 75±0,5 82,5±0,38

91
19 – кестенің жалғасы
1 2 3 4 5 6 7
BSJA12 – 13 1х107 12,5±0,75 47,5±0,75 70±0,5 80±0,5 82,5±0,38
BSJA1 – 14 1х107 20±0,5 52,5±0,38 72,5±0,38 87,5±0,38 92,5±0,75
BSJA2 – 14 1х107 22,5±0,75 50±1 77,5±0,75 82,5±0,38 92,5±0,75
BSJA3 – 14 1х107 20±0,5 52,5±0,75 77,5±0,75 87,5±0,38 92,5±0,38
BSJA4 – 14 1х107 17,5±1,25 50±1 67,5±1,25 82,5±0,38 90±0,5
BSJA5 – 14 1х107 22,5±0,88 52,5±1,38 70±1 85±1 90±0,5
BSJA6 – 14 1х107 15±0,5 45±0,5 70±1 77,5±0,88 85±1
BSJA7 – 14 1х107 15±0,5 45±0,75 65±0,75 80±1 85±0,75
BSJA8 – 14 1х107 25±1 55±1 77,5±0,75 87,5±0,38 95±0,5
2127-3k 1х108 30,0±5,7 40,0±8,2 80,0±8,2 95,0±2,1 95,0±2,1
Іле Алатауының жабайы жемісті орман топырақтарынан бӛлініп алынған
Bacillus thuringiensis бактерия штаммдарының алма күйе кӛбелегінің жұлдыз
құрттарына биологиялық белсенділігі зертханалық жағдайда анықталды. Мұнда
(кесте 20) залалдаудың 5-тәулігінде, BSIA2 – 14, BSIA4 – 14, BSIA5 – 14, 80 %.
BSIA2 – 13, BSIA4 – 13, BSIA10 – 13, BSIA8 – 14, 82,5 %. BSIA8 – 13, BSIA9 – 13,
85 % кӛрсетті. BSIA3 – 13, BSIA7 – 13, BSIA6 – 14, BSIA7 – 14, насекомдардың
ӛлу мӛлшері87,5 % кӛрсетті. BSIA1 – 13, BSIA5 – 13, BSIA6 – 13, 92,5 % жоғары
кӛрсеткішке жетті. Бақылау варианты 2127-3к, 90 % кӛрсетті.
Кесте 20 - Іле Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛліп алған Bacillus thuringiensis энтомопатогенді бактерия
штаммдарының алма күйе кӛбелегінің жұлдыз құрттарына биологиялық
белсенділігі Тәжірибе
нұсқасы
Спора
титры
Жұлдыз құрттардың ӛлу мӛлшері, %
1-тәулік 2-тәулік 3-тәулік 4-тәулік 5-тәулік
1 2 3 4 5 6 7
BSIA1 – 13 1х107 42,5±0,75 70±0,5 80 90±0,5 92,5±0,75
BSIA2 – 13 1х107 27,5±0,38 52,5±1,25 72,5±0,75 77,5±0,38 82,5±0,75
BSIA3 – 13 1х107 40±0,5 65±0,5 77,5±0,38 85±0,75 87,5±0,88
BSIA4 – 13 1х107 35±1 57,5±0,75 77,5±0,75 82,5±0,75 82,5±0,75
BSIA5 – 13 1х107 50±0,5 65±0,5 85±0,5 90±0,5 92,5±0,38
BSIA6 – 13 1х107 55±0,5 82,5±0,38 87,5±0,38 92,5±0,75 92,5±0,75
BSIA7 – 13 1х107 52,5±0,75 77,5±0,38 85±0,75 87,5±0,88 87,5±0,88
BSIA8 – 13 1х107 35±0,5 60 77,5±0,38 82,5±0,75 85±1
BSIA9 – 13 1х107 42,5±1,25 62,5±1,25 72,5±1,25 80±1 85±1
BSIA10 – 13 1х107 37,5±0,75 52,5±0,75 72,5±0,38 80±1 82,5±0,75
BSIA11 – 13 1х107 5±0,5 30±0,5 52,5±0,75 60±1 62,5±1,25
BSIA12 – 13 1х107 7,5±0,38 25±0,5 45±0,5 62,5±1,13 70±0,5
BSIA13 – 13 1х107 10±0,5 32,5±0,88 50±0,5 60±0,5 60±0,5
BSIA14 – 13 1х107 7,5±0,38 25±0,5 50±0,5 60±0,5 62,5±0,75
BSIA15 – 13 1х107 5±0,5 25±0,5 45±0,5 57,5±0,38 62,5±0,75
BSIA16 – 13 1х107 7,5±0,75 25±1 42,5±0,75 55±1 60±1
BSIA17 – 13 1х107 7,5±0,75 27,5±0,75 52,5±0,75 62,5±0,75 70±0,5
BSIA18 – 13 1х107 10±0,5 30±0,5 50 55±0,5 60±0,5
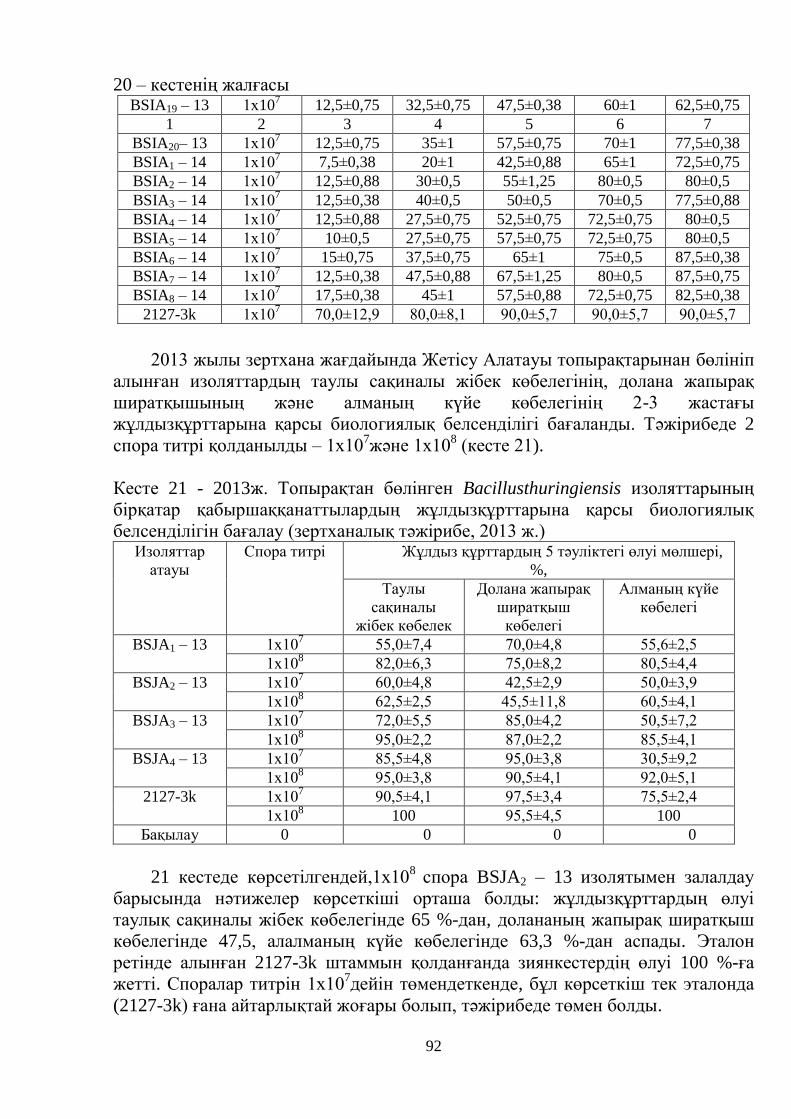
92
20 – кестенің жалғасы
BSIA19 – 13 1х107 12,5±0,75 32,5±0,75 47,5±0,38 60±1 62,5±0,75
1 2 3 4 5 6 7
BSIA20– 13 1х107 12,5±0,75 35±1 57,5±0,75 70±1 77,5±0,38
BSIA1 – 14 1х107 7,5±0,38 20±1 42,5±0,88 65±1 72,5±0,75
BSIA2 – 14 1х107 12,5±0,88 30±0,5 55±1,25 80±0,5 80±0,5
BSIA3 – 14 1х107 12,5±0,38 40±0,5 50±0,5 70±0,5 77,5±0,88
BSIA4 – 14 1х107 12,5±0,88 27,5±0,75 52,5±0,75 72,5±0,75 80±0,5
BSIA5 – 14 1х107 10±0,5 27,5±0,75 57,5±0,75 72,5±0,75 80±0,5
BSIA6 – 14 1х107 15±0,75 37,5±0,75 65±1 75±0,5 87,5±0,38
BSIA7 – 14 1х107 12,5±0,38 47,5±0,88 67,5±1,25 80±0,5 87,5±0,75
BSIA8 – 14 1х107 17,5±0,38 45±1 57,5±0,88 72,5±0,75 82,5±0,38
2127-3k 1х107 70,0±12,9 80,0±8,1 90,0±5,7 90,0±5,7 90,0±5,7
2013 жылы зертхана жағдайында Жетісу Алатауы топырақтарынан бӛлініп
алынған изоляттардың таулы сақиналы жібек кӛбелегінің, долана жапырақ
ширатқышының және алманың күйе кӛбелегінің 2-3 жастағы
жұлдызқұрттарына қарсы биологиялық белсенділігі бағаланды. Тәжірибеде 2
спора титрі қолданылды – 1х107және 1х10
8 (кесте 21).
Кесте 21 - 2013ж. Топырақтан бӛлінген Bacillusthuringiensis изоляттарының
бірқатар қабыршаққанаттылардың жұлдызқұрттарына қарсы биологиялық
белсенділігін бағалау (зертханалық тәжірибе, 2013 ж.) Изоляттар
атауы
Спора титрі Жұлдыз құрттардың 5 тәуліктегі ӛлуі мӛлшері,
%,
Таулы
сақиналы
жібек кӛбелек
Долана жапырақ
ширатқыш
кӛбелегі
Алманың күйе
кӛбелегі
BSJA1 – 13 1х107 55,0±7,4 70,0±4,8 55,6±2,5
1х108 82,0±6,3 75,0±8,2 80,5±4,4
BSJA2 – 13 1х107 60,0±4,8 42,5±2,9 50,0±3,9
1х108 62,5±2,5 45,5±11,8 60,5±4,1
BSJA3 – 13 1х107 72,0±5,5 85,0±4,2 50,5±7,2
1х108 95,0±2,2 87,0±2,2 85,5±4,1
BSJA4 – 13 1х107 85,5±4,8 95,0±3,8 30,5±9,2
1х108 95,0±3,8 90,5±4,1 92,0±5,1
2127-3k
1х107 90,5±4,1 97,5±3,4 75,5±2,4
1х108 100 95,5±4,5 100
Бақылау 0 0 0 0
21 кестеде кӛрсетілгендей,1х108
спора BSJA2 – 13 изолятымен залалдау
барысында нәтижелер кӛрсеткіші орташа болды: жұлдызқұрттардың ӛлуі
таулық сақиналы жібек кӛбелегінде 65 %-дан, долананың жапырақ ширатқыш
кӛбелегінде 47,5, алалманың күйе кӛбелегінде 63,3 %-дан аспады. Эталон
ретінде алынған 2127-3k штаммын қолданғанда зиянкестердің ӛлуі 100 %-ға
жетті. Споралар титрін 1х107дейін тӛмендеткенде, бұл кӛрсеткіш тек эталонда
(2127-3k) ғана айтарлықтай жоғары болып, тәжірибеде тӛмен болды.

93
4.5.3 Бунақденелілерден бӛлініп алынған Bacillus thuringiensis
энтомопатогенді бактерия штаммдарының биологиялық белсенділіктері
2013 жылы күнтізбе жоспарына сәйкес бактериялардың қазақстандық
штаммдары негізінде жасалған бірқатар микробиологиялық жартылай
препараттық үлгілерінің алманың күйе кӛбелегіне қарсы биологиялық
белсенділігін анықтау үшін сынақтар жүргізілді. Алманың күйе кӛбелегінің сан
мӛлшері аз болғандықтан, оған қарсы тек бір ғана препарат, яғни, Қазақстан
Республикасының территориясында қолдануға рұқсат етілген Ақ кӛбелек
сыналды. Ӛңдеуден кейінгі үшінші тәулікте тиімділік 100 % құрады.
Сонымен қатар, Іле Алатауының тау бӛктеріндегі қараусыз қалған алма
бақтарында алманың күйе кӛбелегі мен долананың жапырақ ширатқыш
кӛбелегінің жұлдызқұрттарына қарсы жартылай препараттық үлгілер сыналды.
Танаптық жағдайда алманың күйе кӛбелегінің жұлдызқұрттарына қарсы
жүргізілген сынақтар жоғары нәтижелер кӛрсетті (сурет 32;33). k-Ym07/КОХ,
k-Ym07/К және ЗПТ-07 штаммдары негізінде жасалған культуралды
сұйықтықтармен залалдау барысында зиянкестің ӛлуі сәйкесінше 97,5; 95 және
92,5 % құрады. Ал ОЗШ-07 штаммы негізінде дайындалған жартылай
препараттық үлгі эталон деңгейінде максималды тиімділік кӛрсетіп, бұл
нұсқаларда жұлдызқұрттардың ӛлуі 100 % құрады. Бұл тәжірибе 1 мл
сұйықтықтың спора титрі 1х108болды.
Сурет 32 - Жартылай препараттық үлгілердің алма күйе кӛбелегінің
жұлдызқұрттарына қарсы биологиялық белсенділігін бағалау, титр
1х108спора/мл
Сурет 33 - Алма күйе кӛбелегі жұлдызқұрттарының тәжірибе нұсқасында
ӛлген дарақтары

94
Келесі ұқсас тәжірибеде долана жапырақ ширатқыш кӛбелегінің
жұлдызқұрттарына қарсы екі спора титрі қолданылды – 1х108және
1х107спора/мл.
1х108спора титрімен залалдағанда алдыңғы тәжірибедегідей жоғары
нәтижелер алынды. Зиянкестердің ӛлуі 87,5 тен 97,5 %-ға дейін ауытқыды. Бір
ерекшелігі, бұл тәжірибеде k-Ym07/АК штаммы негізінде жасалған сұйық
жартылай препараттық үлгі алдыңғы тәжірибеге қарағанда әлдеқайда жоғары
нәтиже беріп, зиянкестердің ӛлуі 92,5 % құрады (сурет 34-35).
Сурет 34 - Жартылай препараттық үлгілердің долананың жапырақ
ширатқыш кӛбелегінің жұлдызқұрттарына қарсы биологиялық белсенділігін
бағалау, титр 1х108спора/мл
Споралар титрін 1х107тӛмендету барысында долана жапрықширатқышы
жұлдызқұрттарының ӛлуі 57,5 тен 80 %-ға дейін ауытқып, k-Ym07/АК және k-
Ym07/К штаммдары негізіндегі үлгілерде зиянкестің ӛлуі 57,5 және 67,5
%болды, ал эталон ретінде алынған 2127-3k штаммда бұл кӛрсеткіш 80 % жетті.
Ӛзге препараттық үлгілермен залалдағанда жұлдызқұрттардың ӛлуі 75 және
77,5 % құрады.
Сурет 35 - Долана жапырақ ширатқышы жұлдызқұрттарының тәжірибе
нұсқасында ӛлген дарақтары
Сонымен қатар, танаптық жағдайда Жамбыл облысының Қордай
ауданынан жиналған америкалық ақ кӛбелектің жұлдызқұрттарында тәжірибе
қойылды. Америкалық ақ кӛбелек біздің елде карантинді нысан болып

95
табылады және республика деңгейінде таралса, біздің бақтар үшін үлкен қауіп
тӛндіреді. Бүгінгі таңда зиянкес Алматы облысының Іле, Еңбекшіқазақ,
Қарасай және Талғар аудандарында, сонымен қатар Жамбыл облысының
Қордай ауданында кездеседі. Бұл зиякес 250 астам ағаш және бұта тектестерді,
соның ішінде алманы да зақымдайды. Осыған байланысты, біздің бұл нысанды
елемеуімізге болмады. Энтомопатогенді бактерия штаммдары негізінде
дайындалған культуралды сұйық препараттық үлгілердің уыттылығын анықтау
барысында алынған нәтижелер жоғары болды. Тәжірибеде 1х107және
1х108спора/мл титрлері қолданылды (сурет 36).
Сурет 36 - Америкалық ақ кӛбелектің жұлдызқұрттарына қарсы
қойылған танаптық зерттеулер
36 - суретте кӛрсетілгендей, споралар титрі 1х108спора/мл құрайтын
культуралды сұйықтықпен залалдағанда жұлдызқұрттардың ӛлуі жоғары
болып, 80 нен 100 %-ға дейін ауытқыды. k-Ym07/КОХ, ОЗШ-07, k-Ym07/К,
ЗПТ-07 және 2127-3k (эталон) штаммдары негізінде жасалған КС қолданғанда
келесідей нәтижелер алынды: 97,5; 100; 95; 100 және 100 % сәйкесінше. k-
Ym07/АК штаммы негізіндегі КС әсері әлсіздеу болып, небәрі 80 % тиімділік
кӛрсетті. Споралар титрін 1х107спора/мл тӛмендеткенде сұйық препараттық
үлгілердің де тиімділіктері тӛмендеді: 75; 92,5; 85; 80 және 90 %, сәйкесінше; ал
КЖ k-Ym07/АК штаммы негізіндегі КС нәтиже одан да тӛмен болды – 60 %.
Жүргізілген зерттеулер нәтижесінде Bacillus thuringiensis штаммдары
негізінде жасалған культуралды сұйықтықтардың қабыршаққанатты
бунақденелердің жұлдызқұрттарына қарсы биологиялық белсенділігі жоғары
екендігі анықталды.
4.5.4. Bacillus thuringiensis бактерия штаммдары негізіндегі дақылдық
сұйықтықтарды дайындау және олардың биологиялық белсенділіктері
Энтомопатогенді микроорганизмдер штаммдарының биологиялық
белсенділігін анықтау мақсатында алма күйе кӛбелегі мен долана жапырақ
ширатқышының жұлдызқұрттарында жүргізілген зертханалық сынақтар
нәтижесі бойынша жартылай препараттық үлгілер жасау үшін 5
энтомопатогенді бактерия штаммдары іріктеліп алынды.

96
Құрамы әртүрлі орталарда ӛсінділердің түсімділігі бойынша сұйық
қоректік орталарды таңдау жұмыстары жүргізілді: ашытқыполисахаридті орта
(АПС), етпептонды сорпа (ЕПС) және Лурия-Бертрани (ЛБ) ортасы.
Қоректік орталарда алынған барлық ӛсінді сұйықтықтары шамамен біркелкі
сипатта болды, дегенмен ең жақсы нәтижелер, яғни, споралар мен кристаллдардың
толықтай түзілуі ЛБ ортасында жүзеге асты, сонымен қатар, бұл ортада споралар
титрі ӛзгелерімен салыстырғанда 2 есе жоғары болды. Мұнда споралардың түзілуі 3-4
тәулікте байқалса, АПС ортасында 3 тәулікте тек тірі клеткалар байқалды. АПС та
споралардың толық түзілуі тек 6-7, ал ЕПС – 5-6 тәуліктерде жүзеге асты.
Ӛсінділердің микроморфологиясы Axiostarplus микроскопында жүргізілді.
Бактерия клеткалары дараланып, жұптасып немесе тізбектеліп орналасқан,
ұштары домалақ келген таяқшалар. Сұйық қоректік орталарда 2-3 тәулік
ӛскеннен кейін, олар ромб пішінді ірі кристалл-токсиндер түзе бастады.
Жүргізілген жұмыстар нәтижесінде зертхана жағдайында кӛбейту үшін ЛБ
ортасы таңдалып алынды. Сонымен, бұл қоректік ортада k-Ym07/КОХ, k-
Ym07/АК, k-Ym07/К, ЗПТ-07, ОЗШ-07 және 2127-3k(эталон) штаммдары
негізінде 1000 мл-ден ӛсінділер сұйықтығы дайындалды. Споралар титрі 1х108.
Дайындалған культуралды сұйықтықтардың (КС) (6 Bacillus thuringiensis
бактерия штаммдары негізінде) тіршілікке қабілетті споралар титрі Пастера-
Кох әдісімен анықталды.Себу үшін х1000 сұйылту қолданылды(кесте 22).
Кесте 22 - Bacillus thuringiensis штаммдары негізінде жасалған дақылды
сұйықтар спораларының тіршілікке қабілеттілігін анықтау Штамм атауы Бастапқы дақылды
сұйықтықтағы
споралардың титрі
Тіршілікке қабілетті
споралар титрі
Тіршілікке қабілетті
споралардың пайызы
(%)
k-Ym07/КОХ 3,2х105 3,0х10
5 93,7
ОЗШ-07 4,8 х105 4,7 х10
5 97,9
k-Ym07/К 3,6 х105 3,4 х10
5 94,4
ЗПТ-07 3,7 х105 3,7 х10
5 100
k-Ym07/АК 2 х105 1,6 х10
5 80,0
2127-3k
(эталон)
4,5 х105 4,4 х10
5 97,8
22 кестеде кӛрсетілгендей, сұйық қоректік ортада дайындалған
ӛсінділердің жалпы титрі ӛте жоғары, сонымен қатар олардың тіршіліке
қабілеттілігі де жоғары болды.
Дақылды сұйықтар спораларының тіршілікке қабілеттілігі анықталды.
2013 жылы жүргізілген тәжірибелер нәтижелеріне сүйене отырып, 2014 жылы 2
штамм іріктелді – LAu-12 және BT1-12. Осы штаммдар негізінде 5 литр
культуралды сұйықтық дайындалды. Эталон ретінде отандық Ақ кӛбелек
биопрепаратының штамм-ӛнімі – 2127-3k алынды.
Дайындалған дақылдарды сұйық жартылай препараттық үлгілердің
тіршілікке қабілетті спораларының титрі Пастера-Коха әдісімен анықталды.
Себу үшін х1000 сұйылту қолданылды. Нәтижелер кесте 23–те келтірілді.

97
Кесте 23 - Bacillus thuringiensis штаммдары негізінде жасалған дақылды сұйық
жартылай препараттық үлгілер спораларының тіршілікке қабілетілігін анықтау Штамм атауы Жартылай препараттық үлгінің тіршілікке қабілетті споралар
титрі
Дақылды сұйық үлгінің
спора титрі (х1000
бастапқыдан)
Тіршілікке
қабілетті споралар
титрі
Тіршілікке қабілетті
споралар үлесі (%)
LAu-12 2,2х105 2,1х10
5 97,8
BT1-12 3,4 х105 3,3 х10
5 97,9
2127-3k (эталон) 3,6 х105 3,5 х10
5 97,8
23-кестеде кӛрсетілгендей, сұйық қоректік орталарда дайындалған
үлгілердің споралар титрімен олардың тіршілікке қабілеттілігі жоғары болып,
барлық жағдайларда 95 % асты.
Дайындалған дақылдық сүйықтықтың биологиялық белсенділігі зертхана
және танаптық жағдайларда алманың күйе кӛбелегінің жұлдызқұрттарына
қарсы сыналды.
Дайындалған дақылдық сүйықтықтың биологиялық белсенділігін бастапқы
анықтау жұмыстары зертхана жағдайында алма күйе кӛбелегінің 2-3 жастағы
жұлдызқұрттарына қарсы жүргізілді.Тәжірибеде 1х108, 1х10
7 және 1х10
6
спора/мл титрлары қолданылды. Нәтижелер (кесте 24) - де келтірілді.
Кесте 24 - Зертхана жағдайында Bacillus thuringiensis штаммдары негізінде
дайындалған препараттық сұйық үлгілердің алма күйе кӛбелегінің
жұлдызқұрттарына қарсы биологиялық белсенділігі Споралар
титрі
Штамм Жұлдыз құрттардың тәулік бойы ӛлу мӛлшері, %
1 2 3 4 5
1х108 LAu-12 42,5±17,1 60,0±5,7 60,0±5,7 72,5±7,5 72,5±7,5
BT1-12 95,0±2,8 95,0±2,8 95,0±2,8 95,0±2,8 95,0±2,8
2127-3k
(эталон)
37,5±12,5 70,0±17,7 85,0±8,6 92,5±7,5 100
НСР.05 31,3 31,3 24,6 18,2 14,1
1х107 LAu-12 20,0±4,1 37,5±6,2 50,0±4,1 55,0±6,4 55,0±6,4
BT1-12 40,0±12,2 47,5±8,5 47,5±8,5 57,5±6,2 57,5±6,2
2127-3k
(эталон)
32,5±6,2 42,5±6,2 65,0±6,4 72,5±2,5 72,5±2,5
НСР.05 22,1 19,0 17,6 14,4 14,4
1х
106
LAu-12 5,0±2,8 5,0±2,8 10,0±7,1 15,0±5,0 15,0±5,0
BT1-12 5,0±5,0 10,0±6,4 25,0±6,4 32,5±6,2 32,5±6,2
2127-3k
(эталон)
10,0±5,7 12,5±7,5 27,0±4,7 30,0±5,7 40,0±4,1
Бақылау 0 0 0 0 0
НСР.05 0,0 0,0 16,4 15,2 13,8
24-кестеде кӛрсетілгендей, дақылдық сүйықтықтың залалдауынан кейінгі
бесінші тәулікте зиянкестің ӛлуі BT1-12 штаммы негізіндегі үлгіде 95 %
жетті,ал LAu-12 штаммы негізінде жасалған дақылдық сүйықтық 72,5 %

98
тиімділік кӛрсетті. Эталон ретінде алынған 2127-3k штаммы негізінде жасалға
КС тиімділігі 100 % жетті.Споралар титрін 1х107спора/мл дейін тӛмендеткенде
зиянкестердің ӛлуі залалдаудан кейінгі бесінші тәулікте бұл кӛрсеткіштер
тӛмен және орташа болып, BT1-12 мен LAu-12 штаммдары негізіндегі үлгілерде
сәйкесінше 57,5 және 55 %, ал 2127-3k – 72,5 % құрады. Бақылау нұсқасында
жұлдызқұрттардың ӛлуі тіркелмеді.
Зертхана жағдайында жүргізілген сынақтар нәтижесінде танаптық
ӛңдеулер үшін 1х108 спор/мл спора титрі алынды.
2014 жылы МҰТС-ның Черновск инспекторлық учаскесінде (Алматы обл.,
Алакӛл ауд., Лепсі округы) жүргізілген бағыттық бақылаулар барысында
Сиверс алмасы резерваттарының зиянкестермен зақымдалуы анықталды.
Нәтижесінде, 2013 жылдағыдай ӛте аз мӛлшерде алманың күйе кӛбелегі мен
жапырақ ширатқыш кӛбелектері тіркелді. Осыған байланысты, жеміс
ағаштарын алма күйе кӛбелегіне қарсы дақылдық сұйықтармен ӛңдеу Іле
Алатау МҰТС-да қараусыз қалған алма бақтарында жүргізілді (Чапай шатқалы,
теңіз деңгейінен 1200 м биіктікте). Жұмыс сұйықтығының титрі 1х108 спор/мл.
Жұлдызқұрттардың ӛлуі бүркуден кейінгі бесінші тәулікте есептелді.
Нәтижелер 36 суретте келтірілді.
Сурет 37 - Танаптық жағдайдағы дақылдық сұйықтың алма күйе
кӛбелегінің жұлдызқұрттарына қарсы биологиялық белсенділігі
37 суретте келтірілгендей, BT1-12 штаммы негізінде жасалған дақылдық
сұйықтармен залалдау барысында жұлдызқұрттардың ӛлуі 90 %, ал LAu-12
штаммы негізіндегі КС бұл кӛрсеткіш 67 % құрады. Эталонның биологиялық
белсенділігі 100 % болды.Ал бақылау нұсқасында (ӛңделмеген) зиянкестердің
ӛлуі тіркелмеді.
Жүргізілген тәжірибелер нәтижесі бойынша, биопрепарат жасау үшін
перспективті штамм ретінде BT1-12 Ресей Федерациясының Санкт-Петербург
қаласындағы ӛсімдіктер мен олардың зиянкестеріне қарсы уытты
микроорганизмдердің Мемлекеттік коллекциясында тіркеуден (депонирование)
ӛтті.

99
5 АЛМА КҤЙЕ КӚБЕЛЕГІНЕ ЭНТОМОПОТОГЕНДІ
БАКТЕРИЯЛАР НЕГІЗІНДЕ ЖАСАЛҒАН ЖАРТЫЛАЙ СҦЙЫҚ
ПРЕПАРАТТЫҚ ҤЛГІЛЕРДІҢ ЭКОНОМИКАЛЫҚ ТИІМДІЛІГІ
Bacillus thuringiensis энтомопатогенді бактерия штаммдары негізінде
дайындалған дақылды сұйық үлгілерінің алманың күйе кӛбелегіне тиімділігі
2015-жылы Алматы облысында шаруа қожалығының алма бағында
«Старкримсом» сортында жүргізілді.
Шаруашылықта алма ағашының зиянкесі алма күйе кӛбелегінің жаппай
кӛбейюі салдарынан 2014 және 2015 жж жеміс ӛнімділігі 50 %-ға дейін
тӛмендеген. Бұл жағдайлардың алдын алу үшін біз энтомопатогенді бактерия
штаммдары негізінде дайындалған дақылды жартылай сұйық препараттық
үлгілерді 6 л мӛлшерін сумен сұйылтып, жұлдыз құрттардың жаппай шыға
бастаған кезінде бүрку арқылы берілді. Нәтижесінде, егер алма күйе
кӛбелегінің шабуылына ұшырамаған аумақтан бір гектарға шаққанда 26400 кг
алма жемісі жиналса, алма күйе кӛбелегінің шабуылына ұшыраған аумқтағы
алма ағаштарынан 13200 кг жеміс жиналған. Ал жартылай препаратпен ӛңдеген
аумақта ӛнім мӛлшері 25608 кг құрады. Егер бір гектарға немесе біздің
жағдайда 2200 алма ағашына ӛңдеуге кететін препараттың құны 70000 теңге,
алманың 1кг кӛтерме базардағы бағасы 120 теңгеде шаруашылықта алманың
зиянкестерінің әсері болмаса оның осы сортын ӛсірудің тиімділігі 151,4 %
құрайды, ал алма күйе кӛбелегінің шабуылыны ұшыраған жағдайда
шаруашылықтың тиімділігі 25,7 %-ға дейін құлдырайды.
Осындай жағдайда алма ағаштарын деркезінде энтомопатогенді бактерия
штаммдары негізінде дайындалған дақылдарды жартылай сұйық препараттық
үлгілермен ӛңдеу алынатын жеміс мӛлшерін, ӛңдемегенге қарағанда едәур
кӛтерді (13200 кг-нан 25608 кг-ға дейін). Бұл әрекеттің тиімділігі 74,6 %
құрайды. Яғни жартылай сұйық препаратпен ӛңдеудің экономикалық тиімділігі
жоғары.

100
Кесте 25 – Алма күйе кӛбелегіне энтомопотогенді бактериялар негізінде жасалған жартылай сүйық препараттық үлгінің
«Старкримсом» сортына қолданудың экономикалық тиімділігі (бір гектарға шаққанда)
Үлгілері Ӛнімді
ӛндіруге
кеткен жалпы
шығын, теңге
Алынған
ӛнім
мӛлшері,
кг
Ӛнімнің
/кг
бағасы,
теңге
Алынған
ӛнімнің
бағасы,
теңге
Түскен
таза
кіріс,
теңге
Тиімділік
дәрежесі,
%
Алма күйе кӛбелегінің шабуылына
ұшырамаған алма ағаштары
1260000 26400 120 3168000 1908000 151,4
Алма күйе кӛбелегінің шабуылына
ұшыраған алма ағаштары
1260000 13200 120 1584000 324000 25,7
Алма күйе кӛбелегінің шабуылына
ұшыраған алма ағаштарын препаратпен
ӛңдегеннен кейін
1760000 25608 120 3072960 1312960 74,6
Бір гектардағы жеміс ағаштарының саны 2200 түп, олардың орыналасуы 1,5×3м
Бір алма ағашын баптауға кеткен шығын-573теңге
Бір алма ағашын жартылай сұйық препараттық үлгімен ӛңдеуге кеткен шығын -32теңге

101
ҚОРЫТЫНДЫ
1. Орманның күңгірт-сұр топырақтары Іле Алатауы жабайы жемісті орман
белдеуінде шӛптесін және орман ағаштары астында, бейтарапты, қолайлы
гидротермиялық, «шайылымда» су құбылымы, карбонатсыз, тұщы топырақ
ертіндісі жағдайында қалыптасқан. Топырақ кескіні айқын дифференцияланған
жұқа (0-15 см), жоғары гумусты (10,57 %), гумусты-аккумулятивтік А қабаты,
жұқа (15-30 см), тӛменгі гумусты (2,70 %) В1, түсі ашықтау копролитты В2 және
BC қабаттарынан тұрады. Топырақта азот, фосфор және калий мӛлшерлері
жоғары және орташа деңгейлерде. Жетісу Алатауы жабайы жемісті орманның
күңгірт-сұр топырақтары терең кескінді (150 см дейін), карбонатсыз, шайылу су
құбылымымен сәл қышқыл орта жағдайында айқын дифференциялданған бес
генетикалық қабаттардан (A0, A1, A2B, B1, B2 және BC) тұрады; оның беткі A1,
A2B қабаттары ӛте жоғарға гумусты (сәйкесінше 15,5 және 8,30 %). Қоректік
элементтерге байытылған; гранулометриялық құрамы – жеңілдетілген ауыр
құмбалшықты, кескін суда еритін тұздардан арылған. Бұндай жағдай Bacillus
thuringiensis энтомопатогенді бактериялары ӛмір сүруіне қолайлы жағдай
туғызады.
2. Іле және Жетісу Алатауларының жабайы жемісті орман белдеуінде
топырақ түзуші факторларға талдау жасай отырып, онда қалыптасқан
орманның күңгірт-сұр топырақтарының құрам-қасиеттерінің топырақ
мезофауналарымен микроорганизмдерге қолайлы екендігін, біріншілерінде
жауын құрттары доминант болса, екіншілерде аммонификатрлар, минералды
азотты пайдаланатын микроорганизмдер мен микромициттердің басымдылығы
анықталды.
3. Орманның күңгірт-сұр топырақтарында бактериялардың жалпы сандық
мӛлшері кескін бойы жоғарыдан 18,2 x 106 – ден (5-30 см қалыңдықта) 5,7х10
6 –
дейін (50-70 cм қалыңдықта) тӛмендеген және жыл маусымдары бойынша
кӛктемнен (11,2x106) жазға қарай азайып (8,7x10
6) күзге қарай сәл
кӛтерілетіндігі (9,5x106) анықталды.
4. Орманның күңгірт-сұр топырақтарынан бӛлініп алынған
энтомопатагенді Bacillus thuringiensis бактерияларының саны 1 г топырақта
0,010 - 0,014 КОЕ немесе жалпы бактериялар санның 15-20 % құрайды.
Топырақтардағы бунақденелілердің бактериоздық белгілері бар алма күйе
кӛбелегі және жапырақ ширатқыш жұлдыз құрттарының ӛлі денелерінен
бӛлініп алынған Bacillus thuringiensis бактерияларының морфологиялық
сипаттары бұрыс пішінді, сары, ақшыл-сұр түсті, шеті толқынды дӛңгеленген,
бедері тегіс паста тәрізді екендігі, биохимиялық зерттеулер Ацетил-метилкар
бинол, Лецитови телинді реакция, Протеолиз, Амилаза, Каталазаға реакциясы
оң, сахороза, манноза, салцинге қоректік ортада қышқыл тузуі байқалады.
5. Bacillus thuringiensis штаммдарының жапырақ ширатқыш кӛбелегінің
жұлдыз құрттарына ең жоғарғы биологиялық белсенділік BSIA6 – 13, BSIA5 –
13, BSIA3 – 13де байқалды; олардан жұлдыз құрттардың ӛлу мӛлшері

102
сәйкесінше 100, 98 және 96 % құрды; ал алма күйе кӛбелегіне әсері BSIA3 – 13
және BSIA8 – 14штамдарында байқалып сәйкесінше 98 және 96 % құрды.
6. Алманың күйе және жапырақ ширатқыш кӛбелегінің жұлдыз
құрттарына жартылай препараттық үлгілерінің (1мл сұйықтықтың спора титрі
1x108) танаптық жағдайдағы әсері k-Ym07/КОХ, k-Ym07/К және ЗПТ-07
штамдары негізінде жасалған дақылдық сұйықтықтармен залалдау зянкестердің
ӛлуін сәйкесінше 98, 95 және 93 % құрды.
7. Іле Алатауының МҰТС-да (Чапай шатқалы) ВT1-12 штамы негізінде
дайындалған сұйықтқтардың (спора титрінің 1x108/мл) алманың күйе кӛбелегі
мен жапырақ ширатқыш кӛбелегі жұлдыз құрттарын бүрку олардың ӛлуі 90 %,
ал LAu-12 штамы сәйкесінше 67 % құрады.

103
ПРАКТИКАЛЫҚ ҦСЫНЫСТАР
Қазақстанның Іле Алатау МҰТС-да қараусыз қалған алма бақтарында,
Алматы обылысында алма ӛсірумен айналысатын фермерлік шаруашылықтар
мен шаруа қожалықтарына зерттеу нәтижелерін тұжырымдай келіп келесідей
ұсыныстар жасаймыз:
1. Жүргізілген тәжірибелер нәтижесі бойынша, биопрепарат жасау үшін
перспективті штамм ретінде BT1-12 Ресей Федерациясының Санкт-Петербург
қаласындағы ӛсімдіктер мен олардың зиянкестеріне қарсы уытты
микроорганизмдердің Мемлекеттік коллекциясында тіркеуден (депонирование)
ӛтті.
2. Топырақтан бӛлініп алынған энтомопотогенді бактериялардың жоғары
белсенділік кӛрсеткен 10 штаммы іріктеліп алынып жалғасты зерттеу
мақсатында ҚазӚҚ және КҒЗИ Биотехнология зертханасының коллекциясына
ұсынылды.
3. Ең жоғары белсенділігімен ерекшеленген LAu-12, BT1-12, BSJA3 – 13 және
BSJA6 – 13 болашағы бар бактериялар болашақта биопрепарат шығаруға
ұсынылады
4. Энтомопотогенді бактериялар негізінде жасалған биопрепараттар
экологиялық жағынан қоршаған ортаға зиянсыз және экономикалық тиімділігі
жоғары және ол ӛндірістің рентабельділігін қамтамасыз етеді.

104
ПАЙДАЛАНЫЛҒАН ӘДЕБИЕТТЕР ТІЗІМІ
1 Информационно-аналитический отчет по контрольной и
правоприменительной деятельности Иртышского департамента экологии за
2009 г. – Павлодар, 2009. – 120 с.
2 Чупахин В.М. Физическая география Казахстана. – Алма-Ата: Мектеп,
1968. – С. 28-31.
3 Насыров Р.М. Науменко А.А., Соколов А.А. География почв
Заилийского Алатау. – Алма-Ата: Қазақ университеті, 1991. – 76 с.
4 Чупахин В.М. Природные районирование Казахстана. – Алма-Ата.
Наука, 1993. – 263 с.
5 Литовченко А.Ф. Экспериментальное изучение элементов водного
баланса горных водосборов. - Киев: Вища школа, 1986. – 189 с.
6 Литовченко А.Ф. Инфильтрационные свойства почвогрунтов северного
склона Заилийского Алатау. // Вопросы гидрологии горных районов. – Алма-
Ата: КазГУ, 1971. – 391 с.
7 Карбышева А.Д. Агрогидрологические свойства почв Казахской ССР. –
Алма-Ата: Изд.АН КазССР, 1971. – 112 с.
8 Кубенкулов К., Сейткали Н., Сағитов.А.О. Энтомопотогенді Bacillus
thuringiensis бактериялары қалыптасқан Іле Алатауы жабайы жемісті орманның
топырақ жамылғысына сипаттама // Журнал «Ізденстер, нәтижелер» КазНАУ. –
№2 (066). – 2015. – C. 201-208.
9 Почвы Казахской ССР. Вып 4. Алма-Атинская обалсть. – Алма-Ата,
1962. -422c.
10 Қазақстанның физикалық географиясы; Алматы: Атамұра, 2008. 37-40
бет.
11 Нұрбеков Ж. Жетісу –Семиречие; Алматы: «Эффект» баспасы, 2009. –
б 184-190.
12 Әметов Ә.Ә. Ботаника. – Алматы,2000. – б 320-324.
13 Гиляров М.С , Стриганова Б.Р. Рол почвенных беспозвоночных в
разложении растительных остатков и круговороте веществ // Итого науки и
техники. Почвенная зоология. – 1978. – № 5.– C. 8-69.
14 Гиляров М.С.,Криволуцкий Д.А. Жизн в почве. / –Ростов-на-Дону: Изд-
во РГУ, 1995. –240 c.
15 Тукенова З.А Влияние почвенно-экологических условий на мезофауну
почв Заилиского Алатау: aвтореф. к.б.н, – Алматы, 2002. – 30c).
16 Новиков Г.В., Дудурев А.Н. Санитарная охрана окружающей среды
современного города. – М.: Химия, 2001; 239с.
17 Сейткали Н., Кубенкулов К., Сағитов А.О. Іле және Жетысу Алатауы
жабайы жемісті орман топырақтарының биологиялық кӛрсеткіштері //
«Известия НАН РК» Серия аграрных наук. –Алматы, 2015. –№4(28). –С 64-69.
18 Новиков Г.В., Дудурев А.Н. Санитарная охрана окружающей среды
современного города. – М.: Химия, 2001; –45

105
19 Бахвалов С. А. Взаимоотношения шелкопряда-монашенки Ocneria
monacha L. (Lepidoptera, Lymantriade) и поражающего его вируса
ядерногополиэдроза: // aвтореф. канд. биол. наук. – Новосибирск, 1987. – 22 с.
20 Гулий В.В. Вирусные инфекции пилильщиков–массовых вредителей
лесов в среднем Приобье //aвтореф. канд. биол. наук. –Новосибирск, 1967.–19 с.
21 Полтев В.И., Гриценко Г. Н., Егорова А.И., Кальвиш Т.К. и др.
Микрофлора насекомых. – Новосибирск: Наука СО, 1969. – 272 с.
22 Berliner E.1915. Uber die Schlaffsucht der Mehlmottanraupe (Ephestia
kuhniella Zell) und ihren Erreger Bacillus thuringiensis n-sp. // Z.Angew. Entomol.,2.
– 1915.– P.29-56.
23 Я. Вейзер // Микробиологические методы борьбы с вредными
насекомыми. – Москва:. Колос, 1972. – C. 193-217.
24 Metalnikov S., Metalnikov S.S., C. R. Soc. Biol., – 1993. – 113. – Р.169-
172.
25 Klement Z., Jb. Zentranst. landw. Versuchswesen, Budapest, 1951. – Vol. 3.
– Р.118-127.
26 Shteinhaus E.A. Possible use of Bacillus thuringiensis Berliner as an aid in
the biological control of the alfalfa caterpillar // Hilgardia. – 1951. – Vol.20(18), – Р.
359-381.
27 K.Toumanoff, C.Vago. C.Etude histopatologue des vers a soil atteints de
Bacillus cereus alesti.// Ann. Inst. Pasteur. – 1953. – Vol.84. – P. 376-385.
28 Талалаев.Е.В. О воспроизведении эпизоотии септицемии у гусениц
сибирского шелкопряда // Энтомолог. обозрение.-1957.-36.2. – С.845-859.
29 Красильников Н.А. и Гукасян А.Б. Bacillus tuviensis–новый возбудитель
заболеваний сибирского шелкопряда // Микробиология. – 1964.-30 c.
30 Зурабова Э.Р. // Исследов. по биол. методу борьбы с вредителями
сельского и лесного хозяйства. – Новосибирск, 1964. – С.53-56.
31 Исакова Н.П. // Микроб. Методы борьбы с вредными насекомыми. М.:
Изд. АН ССР, 1964. – С.49-56.
32 Федоринчик Н.С. // Биолог. Средства защиты растений. М.: Колос,
1974. – С. 265-280.
33 Heimpel A.H и Angus T. The taxonomy of insect pathologenes related to
Bacillus cereus F2 of F2. // Canad. J.Microbiol, – P.531-541.
34 D'Barjak H., Bonneffoi A. Classification des souches de Bacillus
thuringiensis // C.R. Acad. Sci. – 1967. – Vol.204. – P. 1811.
35 D'Barjak H., Bonneffoi A. Essai de classification biochimique de 64 Bacillus
des groupes 2 et 3 represent // Species differentes . // Annuls. Inst. Past. – 1972. – P.
122.
36 Krieg A, Barjak H, Bonneffoi A. A new serotype of Bacillus thuringiensis
isolated Germany: Bacillus thuringiensis var. darmstadiensis. // I. Inver., Pathol.,
1968 – Vol. 10 – P. 428-430.
37 Африкян Э.К. Энтомопатогенные свойства спорообразующих бактерий
и их практических значение // Микроорганизмы и их роль в урожайности
растений. – М., 1961. – С. 55

106
38 Африкян Э.К. Энтомопатогенные свойства спорообразующих бактерий
и их практических значение // Изв. биол. ин-та. АН Арм. ССР. – 1968. – №16. –
С. 23-38.
39 Зурабова Э.Р., Рыжкова А.С. Дубинина Т.Д. Изучение разновидностей
серовара 10 Bacillus thuringiensis // Биол. журнал Армении . – 1980. №33(4). – С.
374-378.
40 Krieg A. Bac.thuringiensis Berliner Mitt. Biol. Bundesanstalt Land. // Forst
wirtsch. – Berlin-Dahlem, 1961. – Vol.1, №3. – Р.3-79.
41 De Barjak H, Thompson I. A new serotype of Bac.thuringiensis var.
thimpsoni (serotype 11). // J. Invertebr. Pathology. – 1970. – Vol.15(1). – P. 14.
42 De Barjak H., Lemille F. Fresence of flagellar antigenic Subfactors in
serotype 3 of Bacillus thuringiensis. // J. Invertebr. Phatholog. – 1970. – V.15. – P.
139-140.
43 De Barjak H., Bennefoi A. Essai de classification biohemique de 64 Bacillus
des groups 2 et 3 respresentent // Species differentes // Anuls. Inst. Past. – 1972. – С.
122.
44 Бурцева Л.И. Биологические особенности бактерий Bacillus
thuringiensis: автор. б.ғ,к дис. – Л., 1970. – 22 с.
45 Иванов Г.М., Гукасян А.Б. кристаллоносные бактерии из капустной
совки // Биология гетеротрофных микроорганизмов. –Красноярск, 1971. –С.
264-269.
46 Ходырев В.П. Новая разновидность Bacillus thuringiensis // Микробиол.
иследования в Западной Сибири. –Наука, 1976. –С.116-119.
47 Троицкая Е.Н., Михайлов Е.Н., Плужников Г.А. Новая разновидность
Bacillus thuringiensis // Узбек. Биол. журнал. –1973. - №.3. – С. 47.
48 OhbaM., Aizawia K. new subspecies of Bacillus thuringiensis possessing
11a:11c flagellar antigenic structure: Bacillus thuringiensis subs. Kyushuensis // J.
Invertebr. Pathol. – 1989. – Vol.54(2). – P.208-212.
49 Ohba M., Aizaiwia K. Properties of Bacillus thuringiensis subsр.
wuhanensis // Proc.Assoc.Plant. Prot., Kyushu. – 1979. – Vol.25. – P.125-128.
50 Ohba M., Aizaiwia K Hew flagellar (H) antigenic subfactors in Bacillus
thuringiensis H seratipe 3 with description of two new subspecies,
Bt.subsp.sumigshsiensis (H serotype 3а:3d) and Bt.subsp. funuoraonsis (H serotype
3а:3d:3e) // J.Inverteb. Pahtol. – 1989. – Vol.54(2). – P. 208-212.
51 Carrol J., Ellar D.L. Proteolytie processing of a coleopteran – specifie-
endotoxin produced by Bacillus thuringiensis var. tenebrionsis // Biochem. J. – 1989.
– Vol.261, №1. – P.99-105.
52 Lemoingne M., Bonnefoi A., Beguin S., Crion P., Martouret D., Schenk A.,
Vago C. // Entomofago – 1956. – Vol. 1. – P. 19-34.
53 Vancova J. Rultivirung von Bacillus thuringiensis in
versuchshetriehsmastab. // Trans. Internat. Cont. Insect. Pathol and Biol. Control –
Praha, 1958. –p 26
54 Beegle C.C. // Prog. 18.Int. Congr. Entomol. – Vancouver, 1988. - P.249.

107
55 Патогены насекомых: структурные и функциональные аспекты, под
ред. Глупова В.В. – М.: «Круглый год», 2001. – 736 c.
56 Кандыбин Н.В. // Бактериальные средства борьбы с грызунами и
вредными насекомыми. – М.: Агропромиздат, – 1989. – 175 с.
57 Доолоткельдиева Т. Энтомопатогенные кристаллофорные бактерии
Кыргызстана и их значение // Издательский центр МОК «Технология». – 2001.
–160 с.
58 Рогатин А.В., Байжанов М. // Изв. АН Каз ССР. – 1984. – С. 22-25.
59 Ущеков А.Г. // Защита растений. – 1994. №3. – С. 57-58.
60 Кандыбин Н.В., Смирнов А.В. Малотоннажное производство
биопрепаратов: проблема становления // Защита и карантин растений. -1997. –
Vol 8. С.16-19
61 Рогатых Н.П., Саламаха О.В. Демицид – новый биопрепарат против
личинок колорадского жука // Защита растений. – М, -1993. – C 12-67.
62 Лескова А.Я. Энтомопатогенные бактерии и их роль в защите растений.
– Новосибирск, 1987. – C 10-20.
63 Кольчевский А.Г. Факторы инактивации битоксибациллина в
биоценозах овощных культур и кортофеля. Пути повышения их сохранности на
растениях: aвтореф. канд. –Л., 1983. – С. 24.
64 Чичигина И.П. Вредность бактериальных инсектицидных препаратов,
приготовленных на основе Bacillus thuringiensis для тутового шелкопряда и
пути ее снижения: aвтореф. канд. –Самарканд, 1989. – С. 24.
65 Heimpel A.M. // J. Invert. Pathol. – 1967. – Vol.9. – P. 364-491.
66 Беттхер К. // Микробиология. 1961. Т. 30. – С. 673-678.
67 Lasey L.A., Undeen A.H. // Annu. Rev. Entomol. – 1986. – Vol(48). –
P.631-660.
68 Kaska M. The toxicity of extracellular proteases of the bacterium Serratia
marcescens for larvae of greater wax moth,Galleria mellonella // Invert. Pathol. –
1976. – Vol. 27. – P. 271.
69 Lysenko O. The mechanism of pathogenicity of Pseudomonas aeruginosa
(Schr.) // Insect Phatology. – 1963. – V. 5. – P. 78-97.
70 Лескова А.Я., Рыбина Л.М. // Энтомопат. Бактерии и их роль в защите
растений. –Новосибирск, 1987. – С.31-42.
71 Burgerjon A., Martouret D. Определение круга хореев Bacillus
thuringiensis и его значение. // Микроорганизмы в борьбе с вредными
насекомыми и клещами. –М.: Колос, 1976. – С. 243.
72 Тонконоженко А.П. Патогенность некоторых бактериальных
препаратов для Musco domestica // Ветеринария. -1967. –Vol 2.
73 Тонконоженко А.П. Изучение патогенности микроорганизмов группы
Bacillus thuringiensis – Bacillus cereus для некоторых синонтропных насекомых:
aвтореферат канд. –М., 1968. -24 с.
74 Строева И.А. Хрянина Р.А. Итоги испытаний // биопрепаратов //
Защита растений . -1989. – С. 18-20.

108
75 Саубенова О.Г., Черкашин А.Н., Байжанов М. Действие препарата
Bacillus thuringiensis (серотип 14) на личинках кровососущих комаров в
зависимости от некоторых абиотических факторов // Животный мир Казахстана
и проблемы его охраны: cб. науч. тр. –Алма-ата, 1982. – С. 156-157.
76 Узденов У.В., Ромашева Л.Ф., Федорова С.Ж. Действие на клопов
C.lectularius и клещей D.gallinae термостабильного экзотоксина и препарата
Берлинер из местных штаммов группы Bacillus thuringiensis //
Кристаллообразующие микроорганизмы и перспективы их использования в
птицеводстве: cб. науч. тр. – Фрунзе: Илим, 1981. – С. 45-53.
77 Hofte H., Whiteley H.R. // Microb. Reviews. – 1989.-Vol. 53, №.2.-P.242-
255.
78 Angus T.A. A bacterial toxin paralysing silkworm larvae // Nature. – 1954. –
Vol.173. –P. 545-546.
79 Hannay C.L. Crystalline inclusions in aerobic spore forming // Nature. –
1953. –Vol.172. – P. 1004.
80 Ramoska W.A., Hopkins T.L. Effects of mosquito larval feeding behavior
on Bacillus sphaericus efficacy // J. Invert. Pathol. – 1981. - Vol. 37. – P. 269-272.
81 Rao A.S., Mfhajan S.K. Instability of mosquito larvicidal activity of Bacillus
sphaericus // Biotehnologi. – 1990. – Vol.12. – P.7-10.
82 Hannay C.L. The parosporal body of Bacillus laterosporus Laubach. // J.
Biophys and Biochem. Citol. – 1957. – Vol.3(6). – P.1001-10.
83 Полетов В.И. Использование энтомопатогенных бактерий в
микробиологической борьбе с сибирским шелкопрядом // Сибирский
шелкопряд. – Новосибирск, 1960. – С.103-114.
84 Смирнова Т.А. Структурно-функциональная характеристика
энтомопатогенных бактерий Bacillus thuringiensis и их фагов: aвтореф. докт. –
М., 1991. - 50 с.
85 Tanada I. Susceptibiliti of the imported cabbage worm to Bacillus
thuringiensis Berliner. // Prog. Hawaiian Entom. Soc. – 1953. –№ 15. – P.159-166.
86 Hannay C.L., Fits-Games P.C. The protein crystals of B. thuringiensis
Berliner // J.Cen.Microbiol. – 1965. – №1. – P. 694-710.
87 Fits-James P.S., Yung E.U. Comparison of species and varieties of the genus
Bacillus structure and nuclicacid content of spores. // J. Bacteriol. – 1958. – Vol.78. –
P.743-754.
88 Батурин В.В., Батурина Л.И. Особенности инфекционного процесса у
чешуекрылых и прямокрылых насекомых при заражении их кристаллофорными
бактериями группы B. thuringiensis // Микроорганизмы в защите растений от
вредных насекомых: cб. научных трудов – Иркутск, 1978,-С. 97-107.
89 Сытин А.Г. Селекция споровой энтомопатогенной бактерии методом
хроматографии на бумаге // Исследования по биол. методу борьбы с
вредителями сельского и лесного хозяйства: cб.научн.трудов – Новосибирск,
1964.

109
90 Tojo A., Aizawa K. Dissolution and degradation of B. thuringiensis –
Endotoksin by Gut Juice Protcase of the silkworm. Bombux mori // Applied and
Environmental Microbiologi. – 1983. – Vol. 45, No. 2. – P. 576-580.
91 Brookhart G.L., Kramer K.L. Resistance to Bacillus thuringiensis by the
Indian meal mooth, plodia interpunetella comparison of midgut proteinases from
suscertible and resistant larvae. // J. Invertlbr. Pathol. – 1990. – Vol. 55(2). – P. 235-
244.
92 Heimpel A. Angus T.A. The site of action crystalliferous bacteria in
Lepidoptera larval. // J. Insect Pathol. – 1959. – Vol.1. – P. 152-170.
93 Лескова А. Я. Патогенные свойства Bacillus thuringiensis и условия
эффективного использования их для борьбы с вредными насекомыми.
aвтореф.д-ра биол. наук Л., 1975.- 45с.
94 Поспелов В.П. Фляшерия (септицемия)у гусениц озимой совки Agrotia
segetum Schiff // Изв. отд. приклад. энтомологии. . –1926.
95 Поспелов В.П. Микроорганизмы и симбионаты и их отношение к
болезням насекомых // Защита растений от вредителей. -1929, 6, 1.
96 Исакова Н.П. Патогенез болезни насекомых, вызываемый Bacillus
thuringiensis var. galleriae // Микробиологические методы борьбы с вредными
насекомыми. . – М.: Изд. АН СССР, 1963.
97 Исакова Н.П. Бактерии кишечного тракта насекомых и роль в их
патогенезе заболеваний // Биологические средсвтва защиты растений: cб. научн.
тр. –М.: Колос, 1974. –С. 308-320.
98 Завезенова Т.В., Барайщук Г.В., Кирюшкин А.В. Энтомоцидные
кристаллы бацилл группы thuringiensis // Микроорганизмы в защите растений
от вредных насекомых: cб. науч. тр. –Иркутск, 1978. –С. 30-35.
99 Бурлак В. А. Действие Bacillus thuringiensis subs. kurstaki на гусениц
Galleriae mellonella L. и различие морфологии их включений. // Всесоюзная
конференция «Микроорганизмы – стимуляторы и ингибиторы роста растений и
животных»: тез. докл. Ч.1. – Ташкент, 1989, -С.34.
100 Li Kong-sen, Dai Shun-ying, Li Xiagang, ihao Ying-Chun. Biologi and
toxicity of Bacillus thuringiensis from soils in China. // 19 Int. Congr. Entomol.
Beijing, June 28-jule 4. 1992. Proc: Abstr. Beijing, 1992. –P. 396.
101 Herman H., Whiteley H. Insecticidal crystal protein of Bacillus
thuringiensis. // Microbiol. Rev. – 1989. – Vol. 53,№. 2. –P 242-255.
102 Hafer M., Salama H.S. Persistence and mode of action of Bacillus
thuringiensis against Heliothis armigere (lepidopnera: Noctuidae) // Proc. 18th
Int.Congr. Entomol., Vancouver. July 3rd
-9th, 1988. Abstr and author Index –
(Vancouver, 1988). –P. 260.
103 Ягудин В.Д., Крылов А.С. Изучение антигенной структуры
эндотоксинов Bacillus thuringiensis // Прикладная биохимия и микробиология. –
1990. –С. 852-92.
104 Егоров Н.С., Юдина Т.Г. Биологические активные соидинения
определяющие токсичность микробных инсектицидов на основе Bacillus
thuringiensis. –М.: ВНИИСЭНТИ, –1989. – С. 50.

110
105 Юдина Т.Г., Милько Е.С., Егоров Н.С. Чувствительность диссоцантов
Micrococcus luteus к действию δ-эндотоксинов Bacillus thuringiensis //
Микробиология. –1996. –65.2. –С. 365-369.
106 Юдина Т.Г. Экологическая роль белков включений бактерий рода
Bacillus и рода Xenorhabdus // Биол. и интегрир. защита леса: тезисы докл.
Междунор. Симпозиума (1-7 сентября 1998). –Пушкино-Москов. обл. 1998. – С.
118-119.
107 Яловицин М.В. Энтомопатогенные микроорганизмы и применение их
в борьбе с вредителями: aвтореферат диссертации. Алма-ата, 1969.
108 Масиевская Л.М. Сводный отчет Каз НИИЗР, 1967.
109 Тулемисова К.А. Микробиологические основы создания препаратов
для защиты растений: aвотреф. канд. –М.: МГУ, 1989. – С. 50.
110 Копжасаров Б.Б. Биологические особенности американской белой
бабочки (Hyphantria cunea Drury) на юге-востоке Казахстана и разработка
защитных мероприятий против неѐ: aвотреф. канд. –Алматы, 2007. – С 27.
111 Смирнов О.В., Гришечкина С.Д. Полифункциональная активность
Bacillus thuringiensis berliner // Сельскохозяйственная биология. - 2011. - №3. -
С. 123-126.
112 Добрица А.П., Корецкая Н.Г., Гайта В.И., Коломбет Л.В., Дербышев
В.В., Жиглецова С.К. Разработка биопестицидов против колорадского жука //
Ж. Рос.хим. общества им. Менделеева. – 2001. - Т. XLV, №5-6. . – С 12.
113 Кузнецов В.И. Сем. Tortricidae (Olethreutidae, Cochylidae) —
Листовертки. / Насекомые и клещи— вредители сельскохозяйственных
культур.. Чешуекрылые. СПб.: Наука, 1994. Т. 3, ч. 1. – С. 51—234.
114 Асанов Ж.А. Ауыл шаруашылық негіздері (топырақ тану, егін
шаруашылығы, агрохимия) курсы боинша методикалық нұсқау. – Алматы,1987.
115 Мырзаханов Н. Ауыл шаруашылық негіздеріне арналған тәжірибе
құралы; oқулық / –Алматы: Республикалық баспа кабинеті, 1994.
116 Жамалбеков Е., Білдебаева Р. Топырақтану және топырақ
географиясы мен экологиясы. – Алматы,2000.
117 Елешев. Р., Смағұлов. Т., Балғабаев. Ә., Р.Рамазанова. Агрохимиялық
зерттеулер әдістемесі. – Алматы, 2008. –213 б.
118 «Топырақтану» пәні бойынша лабораториялық жұмыстарды жүргізу
әдістемесі: оқу-әдістемелік құралы. – Орал.2004.
119 Звягинцева. Д.Г. Методы почвенной микробиологии и биохимии., –
М; МГУ, 1991.
120 Шигаева М.Х., Қанаев А.Т. Микробиология және вирусология. Қазақ
Университеті, 2007.
121 Ильин В.Б., Степанова М.Д., Гармаш Г.А. Некоторые аспекты
загрязнения среды:тяжелые металлы в системе почва-растениеИзв. СО АН
СССР. Сер биол.-1980-Вып. 3. 89-99 с.
122 Гринь А.В.,Ли С.К.,Зырин Н.Г., Обухов А.И.,Платонов
Г.В.Поступление тяжелых металлов в растения в зависимости от их содержания

111
в почвах// Миграция загрязняющих веществ в почвах и сопредельных средах:
тр 2-го Всесоюз. Совещ. – Л, 1980. –C 198-202.
123 Аристовская Т.В. Микробиология подзолистых почв. – М.; Л., 1965.
124 Волков С.Н. Землеустройство в условиях земельной реформы:
экономика, экология, право. – М.:Былина, 1998.
125 Власов Г. М. Четвертичное оледенение Камчатки / Г. М. Власов, Ю.Ф.
Чемеков // Вопросы географии Дальнего Востока / под ред. Е. М. Лавренко.-Вл.
–М. –1990.
126 Плохинский Н.А. Биометрия. – М.: Изд-во МГУ, 1970. – 367с.
127 Практикум по микробиологии: учеб.пос. для студ. высшуч. заведений
/под ред. Нетрусова А.И. – М: Изд. Центр «Академия». – 2005. – 608 с.
128 Лабинская А.С. // Практикум по микробиологическим методам
исследования. – М. 1978.
129 Временные методические указания по критериям эффективности
энтомофагов и энтомопатогенов. – М. 1988. – 27 с.
130 Литовченко А.Ф. Экспериментальное изучение элементов
водного баланса горных водосборов. – Киев: Вища школа, 1986. – 189 с.
131 Ресурсы поверхностных вод СССР. Бассейн оз.Балхаш. –Л.:
Гидрометеоиздат, 1970. – Т.1, вып.2. – 643 с.
132 Соседов И.С. Методика территориальных воднобалансовых
обобщений в горах. – Алма-Ата: Наука, 1976. -150 с.,
133 Ресурсы поверхностных вод СССР. Бассейн оз.Балхаш. –Л.:
Гидрометеоиздат, 1970. – Т.13, вып.2. – 643 с.
134 Научно прикладной справочник по климату СССР. Серия
3. Многолетние данные, Казахская ССР. книга 1. –Л.: Гидрометеоиздат,
1989. – Вып. 18, ч.1-6. – 514 с.
135 Справочник по климату Казахстана. Многолетние данные.
Температура воздуха.– Алматы, 2004. - Вып.14, раздел 1– 563с.
136 Литовченко А.Ф., Мазур Л.П. Методика расчета суммарного
испарения с горных склонов // Тр. КазНИГМИ. - 1977. - Вып. 58. – С. 46-50
137 Мазур Л.П. Исследование и расчет атмосферных осадков на
горных водосборах Заилийского Алатау // Вопросыгидрологического и
гидравлического режима рек Казахстана. – Алматы: КазГУ, 1993. – С. 12-18
138 Коровин В.Н. Вертикальная зональность запасов твердых осадков
в горных районах Казахстана // Вопросы гидрологии горных районов. –
Алма-Ата, 1971. – С. 46-54.
139 Справочник поклимату Казахстана. Многолетние данные.
Атмосферные осадки. Раздел 2.– Алматы: 2004. - Вып.14. – 72с.
140 Мазур Л.П. К методике расчета снегозапасов на горных водосборах
// Развитие и преобразование географической среды Казахстана. – Алма-
Ата: Изд.КазГУ, 1982. – С. 106-115.
141 Судаков П.А., Вилесев Е.Н. Режим снежного покрова и
его распределение на Малаалматинских ледниках// Гляциологические
исследования в период МГГ. – Алма-Ата: Наука, 1962. – Вып.2. – С. 99-114.

112
142 Справочник по климату Казахстана. Многолетние данные.
Атмосферные осадки..– Алматы, 2004. - Вып.14. раздел 2. – С 13-15.
143 Жандаев М.Ж. Речные долины. – Алма-Ата, Казахстан, 1984. -184 с.
144 Геология СССР. Южный Казахстан. –М.: Недра, 1971. –Т.10, ч.1.
– 534 с.
145 Жандаев М.Ж. Геоморфология Заилийского Алатау и
проблемы формирования речных долин. – Алма-Ата: Наука, 1972. – 162 с
146 Соседов И.С. Исследование баланса снеговой влаги на горных
склонах. – Алма-Ата: Наука, 1967. -199 с.
147 Чупахин В.М. Физическая география Казахстана. – Алма-Ата:
Мектеп, 1968. – 260с.
148 Джанпейсова Р. Эрозия и дефиляция почв Казахстана. – Алма-
Ата: Наука Казахской ССР, 1977. – 282 с.
149 Литовченко А.Ф., Мочалов В.П. Полевые экспериментальные
исследования потерь дождевых вод на инфильтрацию в горах северного склона
Заилийского Алатау. – Алма-Ата, 1964. – 195 с.
150 Ананьев Г.С. Денудационная устойчивость горных парод в
разных климатических условиях // Сб. Геоморфология. – М.: Наука, 1985. - №2.
– С. 12-16.
151 Кассин Н.Г. Материалы по палеогеографии Казахстана. – Алма-
Ата: Изд. АН КазССР, 1947. – 257 с.
152 Казанли Д.Н. Новейшие движения Заилийского Алатау // Вестник
АН КазССР. – Алма-Ата, 1948. – №1(34). – С.23-27.
153 Медоев Г.Ц. Геологические и геоморфологические строение
гор Сюгаты (Заилийский Алатау). – Алма-Ата: Изд. АН КазССР, 1951. – С. 3-
105.
154 Костенко Н.Н. Основы стратиграфии антропогена Казахстана. –
Алма-Ата: Изд. АН КазССР, 1963. – 75 с.
155 Абдуллин А.А. Геология Казахстана. – Алма-Ата: Наука. 1981. –312с.
156 Шлыгин Е.Н. Геологическая история и геологическое
строение Казахстана // Очерк по физической географии Казахстана. - Алма-
Ата: Наука, 1952. – С. 59-127.
157 Соседов И.С. Методика территориальных воднобалансовых
обобщений в горах. – Алма-Ата: Наука, 1976. -150 с.
158 Орлов Д.С Химия почв. – М. : МГУ,1985. – C 268-278.
159 Толысбаев Б.Т., Иванова Г.М., Музапбаров Б., Тулемисова
Ж.К.Практикум по микробиологии и ветеринарной санитарии. – Алматы:
РИМЦ Минсельхоз РК, 1996.
160 Временные методические указания по критериям эффективности
энтомофагов и энтомопатогенов. – М. 1988. – 27 с.

113
ҚОСЫМША А

114
ҚОСЫМША А жалғасы

115
ҚОСЫМША Ә
Кесте A1- Іле Алатауы жабайы жемісті орман белдеуіндегі орманның күңгірт
сұр топырағының далалық ылғалдылығы (10.2014)
Үлгінің
тереңдігі,
см
Топырақ ылғалдылығы %
Қайталымдар Орташа
мән
Стандарттық
ауытқуы 1 2 3
0-15 34,60 33,30 32,40 33,43 33,43±0,011
15-25 24,20 23,80 24 24,00 24,00±0,002
40-50 21,60 21,50 21,7 21,60 21,60±0,001
60-70 19,30 19,40 19,20 19,30 19,30±0,001
90-105 16,90 17 16,80 16,90 16,90±0,001
105-120 13,50 13,40 13,40 13,43 13,43±0,001
Кесте A2- Жетісу Алатауы жабайы жемісті орман белдеуіндегі орманның
күңгірт сұр топырақтарының далалық ылғалдылығы
Үлгінің тереңдігі, см Топырақ ылғалдылығы %
кӛктем күз
0-15см 30,15 32,20
15-40см 23,50 26,15
40-70см 20,80 22,40
70-90см 19,70 20,25
90-150см 10,54 12,40

116
ҚОСЫМША Б
Іле Алатауының биіктік белдеулігіне топырақтық және агротопырақтық
сипаттама
(А.Соколов, Д.Стороженко)
Биктік
белдеулік
Аумағы
Млн,га
Негізгі
топырақтары
Қара
шірінділік
қабат
АВ,см
А-дағы
гумус,%
Қара шірінділік
Құрамы
C:N
С.г.к:
Сф,қ,
Та етіегіндегі
шолді дала
16,0 Құба топырақ
ашық
Құба топырақ
кәдімгі
Құба топырақ
күңгірт
Ашық қара-қоңыр
карбонатты
18-20
25-30
60-70
30-40
1-1,5
1,5-2,5
2,5-3,5
2-3
6-8
7-8
8-9
7-9
0,5-0,7
0,7-0,8
0,8-0,9
0,9-1,0
Аласа таулық
дала
9,7 Таулық құба
Таулық құба
күңгірт қара-қоңыр
Қара таулық
кәдімгі
50-70
40-50
50-70
2-3
3-5
6-8
8-9
8-9
8,5-9
0,7-0,9
1,1
1,2
Орташатаулық
шалғынды
орман
4,7 Сілтілі Т
Күрең Т
Сұр ормандық Т
Күңгірт Т
80-100
60-100
70-80
35-50
10-13
4-12
15-17
8-12
8-10
9-11
11-12
11
1,2-1,4
0,8-1,0
0,9
0,1-0,9
Биік таулық
шалғын және
шалғынды дала
3,0 Шалғынды
субальпілік шалғын
Т.
Биік таулық
шалғынды далалық
Т.
35-55
55-65
30-50
11-12
12-18
13-15
9-14
11-12
8-13
0,8-0,9
0,8-1,0

117
ҚОСЫМША В
2013-2014-жылдар Жетісу Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛліп алған Bacillus thuringiensis энтомопатогенді бактерия
колонияларының тәулік бойы ӛсу кӛрсеткіші
Бактерия штаммдары Колониялардың тәулік бойы ӛсуі (cm)
1 2 3 4
BSJA1 – 13 0,6-0,6 1,1-1,1 2,0-2,0 2,3-2,3
BSJA2 – 13 0,6-0,7 1,1-1,2 1,8-2,0 2,1-2,2
BSJA3– 13 0,6-0,6 1,1-1,1 1,8-2,0 2,2-2,3
BSJA4 – 13 0,8-0,8 1,3-1,4 2,3-2,4 3,2-3,3
BSJA5 – 13 0,6-0,7 1,3-1,4 2,3-2,4 2,6-2,7
BSJA6 – 13 0,7-0,8 1,1-1,2 1,8-1,9 2,3-2,4
BSJA7 – 13 0,6-0,6 1,1-1,1 2,3-2,4 3,0-3,1
BSJA8 – 13 0,6-0,7 1,0-1,2 1,9-2,0 2,3-2,5
BSJA9 – 13 0,6-0,7 0,9-1,1 1,9-2,0 2,2-2,3
BSJA10– 13 0,7-0,8 1,1-1,2 2,0-2,1 2,6-2,7
BSJA11– 13 0,6-0,8 1,1-1,3 2,0-2,2 2,2-2,5
BSJA12– 13 0,6-0,7 1,1-1,2 2,0-2,1 2,3-2,4
BSJA1 – 14 0,6-0,7 0,9-1,1 1,9-2,1 2,3-2,5
BSJA2 – 14 0,5-0,6 0,8-0,9 1,6-1,7 2,1-2,2
BSJA3 – 14 0,7-0,8 1,1-1,1 2,3-2,4 2,5-2,1
BSJA4 – 14 0,6-0,8 1,1-1,2 2,0-2,1 2,5-3,1
BSJA5 – 14 0,6-0,7 1,1-1,1 2,0-2,1 2,3-2,4
BSJA6 – 14 0,7-0,8 1,1-1,2 2,0-2,2 2,2-2,5
BSJA7 – 14 0,5-0,6 0,8-0,9 1,7-1,8 2,1-2,2
BSJA8 – 14 0,6-0,7 0,9-1,1 1,9-2,0 2,4-2,5

118
ҚОСЫМША Г
2013-2014-жылдар Іле Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛліп алған Bacillus thuringiensis энтомопатогенді бактерия
колонияларының тәулік бойы ӛсу кӛрсеткіші Бактерия
штаммдары
Колониялардың тәулік бойы ӛсуі (cm)
1 2 3 4
BSIA1 – 13 0,6-0,7 1,1-1,2 2,0-2,1 2,3-2,4
BSIA2 – 13 0,7-0,8 1,1-1,2 1,9-2,1 2,2-2,3
BSIA3– 13 0,6-0,6 1,1-1,1 1,9-2,0 2,3-2,4
BSIA4 – 13 0,7-0,8 1,2-1,3 2,2-2,3 2,7-2,9
BSIA5 – 13 0,6-0,7 1,2-1,3 2,2-2,3 2,6-2,7
BSIA6 – 13 0,7-0,8 1,1-1,2 1,8-1,9 2,4-2,4
BSIA7 – 13 0,6-0,6 1,1-1,2 2,3-2,4 3,0-3,1
BSIA8 – 13 0,6-0,7 1,0-1,2 2,0-2,1 2,3-2,5
BSIA9 – 13 0,6-0,7 1,0-1,1 2,0-2,1 2,2-2,3
BSIA10– 13 0,7-0,8 1,1-1,2 2,1-2,1 2,6-2,7
BSIA11– 13 0,6-0,8 1,1-1,2 2,0-2,2 2,2-2,5
BSIA12– 13 0,6-0,7 1,1-1,2 2,0-2,1 2,3-2,4
BSIA13– 13 0,6-0,6 1,1-1,2 2,0-2,1 2,3-2,5
BSIA14– 13 0,5-0,6 1,0-1,1 1,6-1,7 2,2-2,3
BSIA15– 13 0,6-0,7 1,1-1,2 2,0-2,1 2,4-2,4
BSIA16– 13 0,6-0,8 1,0-1,1 2,1-2,1 2,6-2,7
BSIA17– 13 0,6-0,6 1,1-1,2 2,0-2,1 2,3-2,5
BSIA18– 13 0,6-0,7 1,1-1,2 2,0-2,2 2,2-2,5
BSIA19– 13 0,6-0,7 1,0-1,1 2,0-2,1 2,8-2,9
BSIA20– 13 0,6-0,6 1,1-1,2 2,1-2,1 2,9-2,9
BSIA1 – 14 0,6-0,7 0,9-1,0 1,9-2,0 2,8-2,9
BSIA2 – 14 0,5-0,6 0,8-0,9 1,6-1,8 2,6-2,8
BSIA3 – 14 0,7-0,8 1,1-1,1 2,3-2,4 3,4-3,5
BSIA4 – 14 0,6-0,8 1,2-1,3 2,0-2,1 3,0-3,1
BSIA5 – 14 0,6-0,7 1,1-1,2 2,0-2,1 2,9-3,0
BSIA6 – 14 0,7-0,8 1,1-1,2 2,0-2,1 2,7-2,9
BSIA7 – 14 0,5-0,6 0,8-0,9 1,6-1,7 2,6-2,7
BSIA8 – 14 0,6-0,7 1,0-1,1 1,9-2,0 2,9-3,0

119
ҚОСЫМША Ғ
2013-2014-жылдар Жетісу Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛліп алған Bacillus thuringiensis бактерия штаммдарының
әртүрлі тығыз қоректік орталарда 60 сағат ӛсіргеннен кейінгі ӛсу динамикасы Бактерия штаммдары ГА ПА ЕПА «А»
BSJA1 – 13 0,6 1,1 2,0 2,3
BSJA2 – 13 0,3 1,2 2,0 2,2
BSJA3– 13 0,2 1,1 2,0 2,3
BSJA4 – 13 0,8 1,4 2,4 3,3
BSJA5 – 13 0,5 1,4 2,4 2,7
BSJA6 – 13 0,4 1,2 1,9 2,4
BSJA7 – 13 0,3 1,1 1,4 3,1
BSJA8 – 13 0,2 1,2 2,1 2,5
BSJA9 – 13 0,3 1,1 2,0 2,3
BSJA10– 13 0,5 1,2 2,1 2,7
BSJA11– 13 0,3 1,3 2,2 2,5
BSJA12– 13 0,5 1,2 2,1 2,4
BSJA1 – 14 0,2 1,1 2,1 2,5
BSJA2 – 14 0,3 1,5 1,7 2,2
BSJA3 – 14 0,5 1,4 2,0 3,1
BSJA4 – 14 0,2 1,2 1,5 2,6
BSJA5 – 14 0,5 1,1 2,1 2,4
BSJA6 – 14 0,4 1,2 1,2 2,5
BSJA7 – 14 0,2 1,4 1,8 2,2
BSJA8 – 14 0,3 1,1 2,0 2,5

120
ҚОСЫМША Д
2013-2014-жылдар Іле Алатауының жабайы жемісті орманның күңгірт-сұр
топырақтарынан бӛліп алған Bacillus thuringiensis энтомопатогенді бактерия
колонияларының тәулік бойы ӛсу кӛрсеткіші Бактерия штаммдары ГА ПА ЕПА «А»
BSIA1 – 13 0,5 1,2 2,0-2,1 2,4
BSIA2 – 13 0,3 1,2 1,9 2,3
BSIA3– 13 0,2 1,1 1,9 2,4
BSIA4 – 13 0,4 1,3 2,2 2,9
BSIA5 – 13 0,3 1,2 2,2 2,7
BSIA6 – 13 0,2 1,1 1,8 2,4
BSIA7 – 13 0,5 1,2 2,3 3,1
BSIA8 – 13 0,3 1,0 2,1 2,5
BSIA9 – 13 0,4 1,0 2,1 2,3
BSIA10– 13 0,4 1,1 2,1 2,7
BSIA11– 13 0,3 1,2 2,2 2,5
BSIA12– 13 0,3 1,2 2,1 2,4
BSIA13– 13 0,3 1,1 2,1 2,5
BSIA14– 13 0,2 1,0 1,6 2,3
BSIA15– 13 0,4 1,1 2,0 2,4
BSIA16– 13 0,3 1,0 2,1 2,7
BSIA17– 13 0,4 1,2 2,0 2,5
BSIA18– 13 0,3 1,1 2,0 2,5
BSIA19– 13 0,4 1,0 2,0 2,9
BSIA20– 13 0,3 1,1 2,1 2,9
BSIA1 – 14 0,6 1,0 1,3 2,9
BSIA2 – 14 0,5 1,2 1,5 2,8
BSIA3 – 14 0,7 1,4 2,3 3,5
BSIA4 – 14 0,6 1,2 2,0 3,1
BSIA5 – 14 0,6 1,2 2,1 3,0
BSIA6 – 14 0,7 1,1 2,0 2,9
BSIA7 – 14 0,5 1,2 1,6 2,7
BSIA8 – 14 0,6 1,1 1,9 3,0

121
ҚОСЫМША Е
Жетісу Алатауы орманның күңгірт-сұр топырақтарының топырақ кескіні

122
ҚОСЫМША Ж
Зертханалық жағдайда топырақ микроорганизмдерін бӛліп алу





























