Embed Size (px)
Citation preview

How information is transmitted between generations
Read chapter 6

How information is transmitted between generations
Darwin unfortunately never understood what genes were or how information passed between generations.
Even though Darwin didn’t know it, Gregor Mendel was figuring out the laws of inheritance while Darwin was developing his theory of evolution.

Mendel’s work
Mendel carried out classic research on inheritance by crossing pure-breeding strains of peas.
The results of his crosses revealed two basic laws of inheritance.
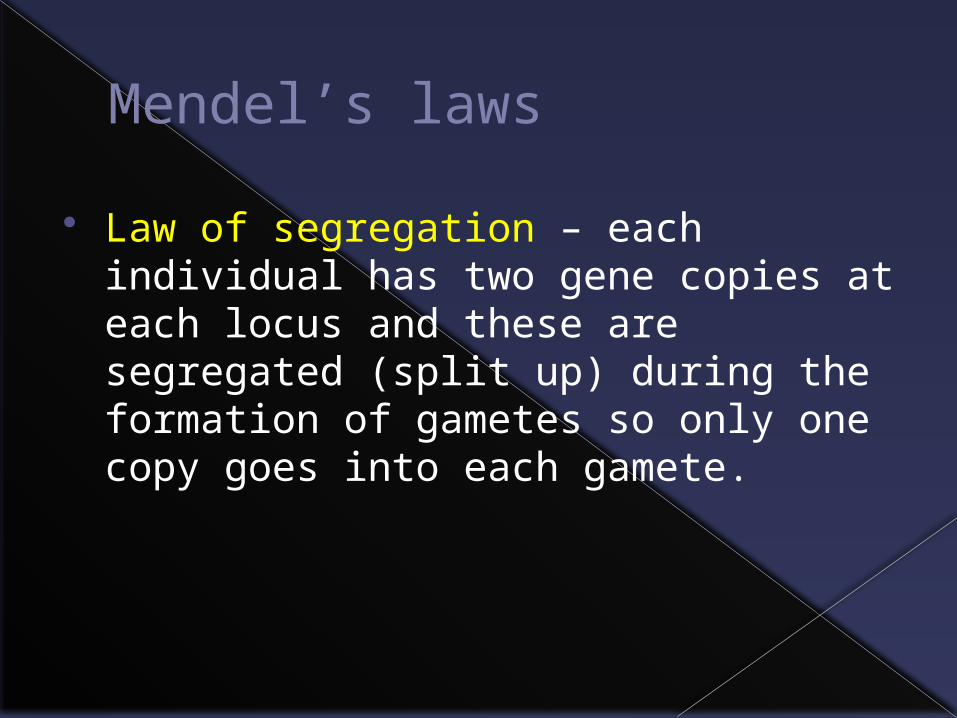
Mendel’s laws
Law of segregation – each individual has two gene copies at each locus and these are segregated (split up) during the formation of gametes so only one copy goes into each gamete.


Mendel’s laws
Mendel also showed by carrying out crosses in which he tracked multiple traits simultaneously that different traits were inherited independently.
Mendel’s Law of independent assortment – alleles at different loci are inherited independently i.e. what allele is passed down at one locus has no influence on what allele is passed down at another.

Blending vs particulate inheritance
What Mendel’s work showed was that copies of discrete particles (what we today call genes) are passed from one generation to the next.
Inheritance is thus particulate and is not a process of blending (e.g. as occurs when paints are mixed)


Blending inheritance
An early criticism of Darwin’s work was that any new favorable mutation would be so diluted by the effects of blending inheritance that it could never become common.
How then could evolution occur?


Particulate inheritance.
With particulate inheritance that problem disappears. Genes do not blend together and so a favorable variant can increase over time as a result of selection.

Where do new alleles come from?
DNA made up of sequence of nucleotides. Each nucleotide includes a sugar, phosphate and one of four possible nitrogenous bases (adenine and guanine [both purines], and thymine and cytosine [both pyrimidines]).


Where do new alleles come from?
The opposite strands of the DNA molecule are complementary because the strands are held together by bonds between the opposing bases and adenine bonds only with thymine and cytosine only with guanine.
Thus, knowing the sequence on one strand enables one to construct the sequence on the other strand.
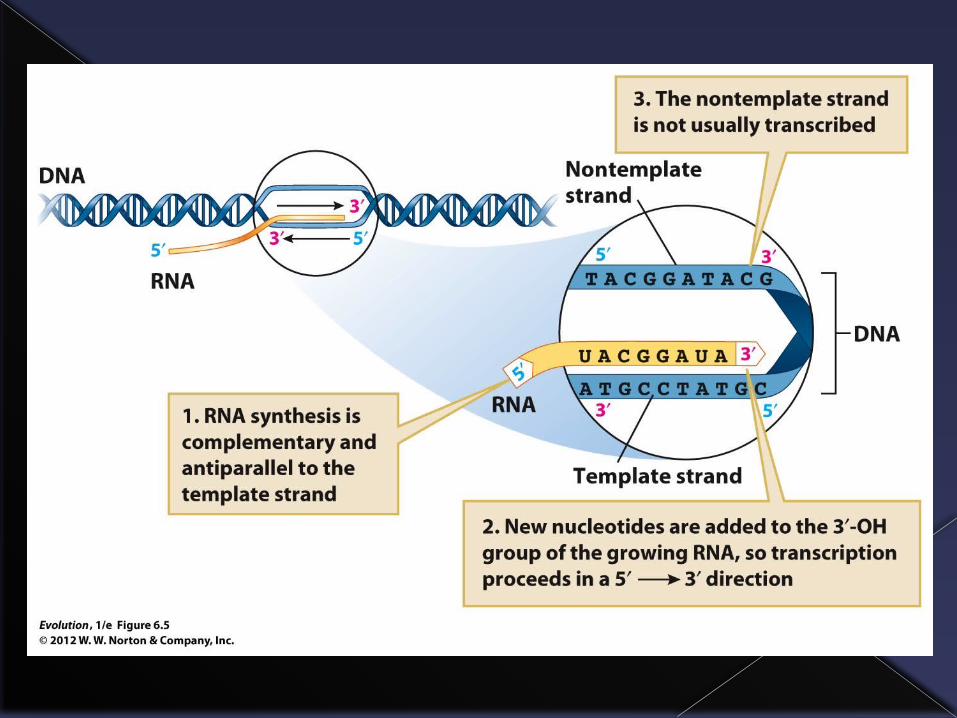

Where do new alleles come from?
Sequence of bases in DNA codes for protein structure as each three base sequence codes for one amino acid in the protein chain.
[To refresh yourself on basic DNA structure and protein synthesis see any Introductory Biology textbook]


Transcription and translation
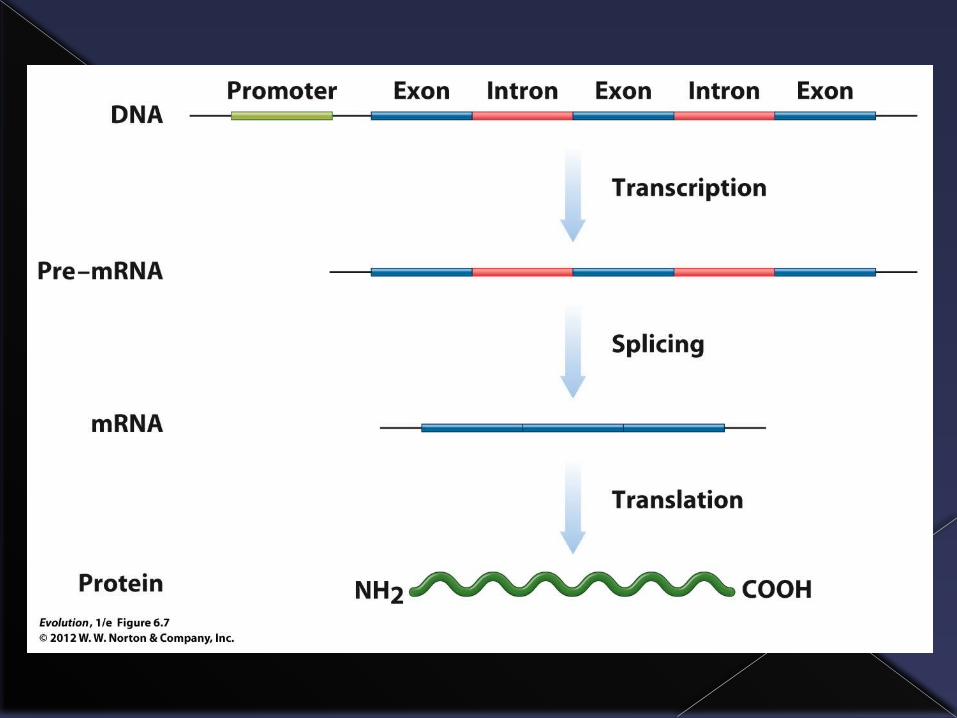
In the processes of transcription and translation a section of DNA (a gene) is first transcribed into to an RNA copy, which then has all the all the non-coding portions (the introns) removed, and is then translated into a protein.
One gene codes for one protein.

Alleles and genotypes
Different versions of a gene are called alleles.
The combination of genes an individual has at a locus (a genes physical location on a chromosome) is referred to as its genotype.
[genotype may also refer to all the alleles an individual has at all its loci]

Heterozygotes and homozygote
An individual who has two different alleles at its loci is a heterozygote.
An individual with two copies of the same allele is referred to as a homozygote.

Dominant and recessive alleles
If the heterozygote is phenotypically identical to one of the homozygotes we say the allele in that homozygote is dominant and the other allele is recessive.
If the heterozygote is phenotypically intermediate between the two homozygotes we say the alleles are codominant.


Punnett Squares
A grid called a punnett square can be used to figure out what genotypes will be produced as a result of a cross between individuals.
Each side of the punnett square lists the possible gametes an individual can produce and the squares in the grid show the genotypes different combinations of gametes will produce.


Genetic variability and recombination
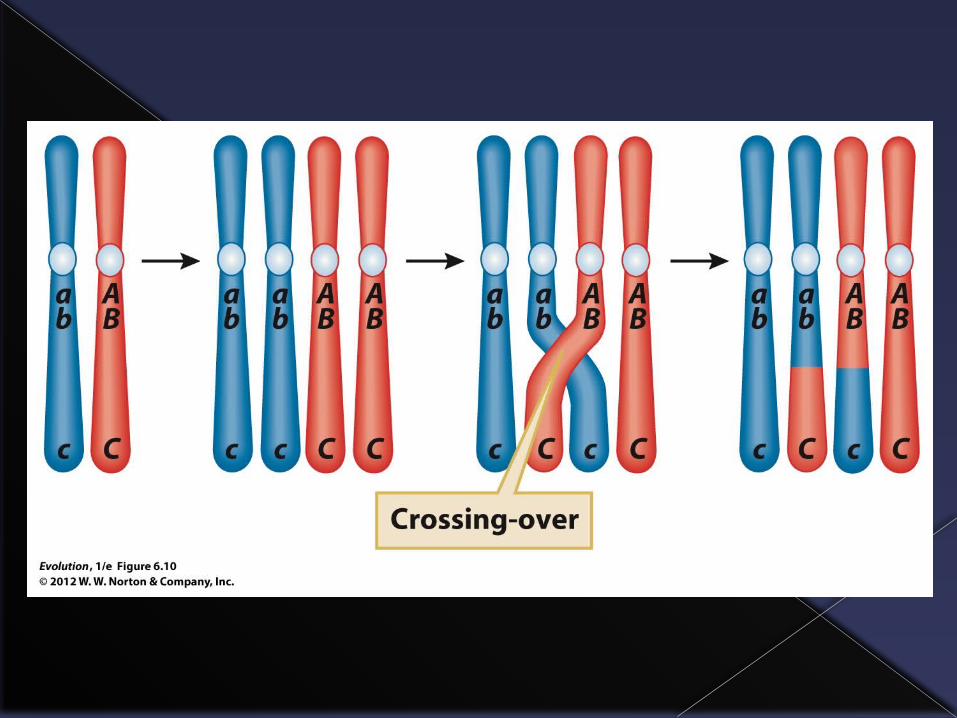
A major source of genetic variability comes from the production of new combinations of genes as a result of sexual reproduction.
The process of crossing-over in which pieces of chromosomes are exchanged between chromosomes produces new and unique chromosomes.

Genetic variability and mutation
Recombination remixes existing variation, but where does variation ultimately come from?
Mutations – changes in the DNA sequence of an organism are the ultimate source of all variation

Where do new alleles come from?
When DNA is synthesized an enzyme called DNA polymerase reads one strand of DNA molecule and constructs a complementary strand.
If DNA polymerase makes a mistake and it is not repaired, a mutation has occurred.

Types of mutations
A mistake that changes one base on a DNA molecule is called a point mutation.
Two forms:› Transition: one pyrimidine (T or C) substituted
for the other pyrimidine or one purine substituted for the other purine (A or G).
› Transversion: purine substituted for pyrimidine or vice versa
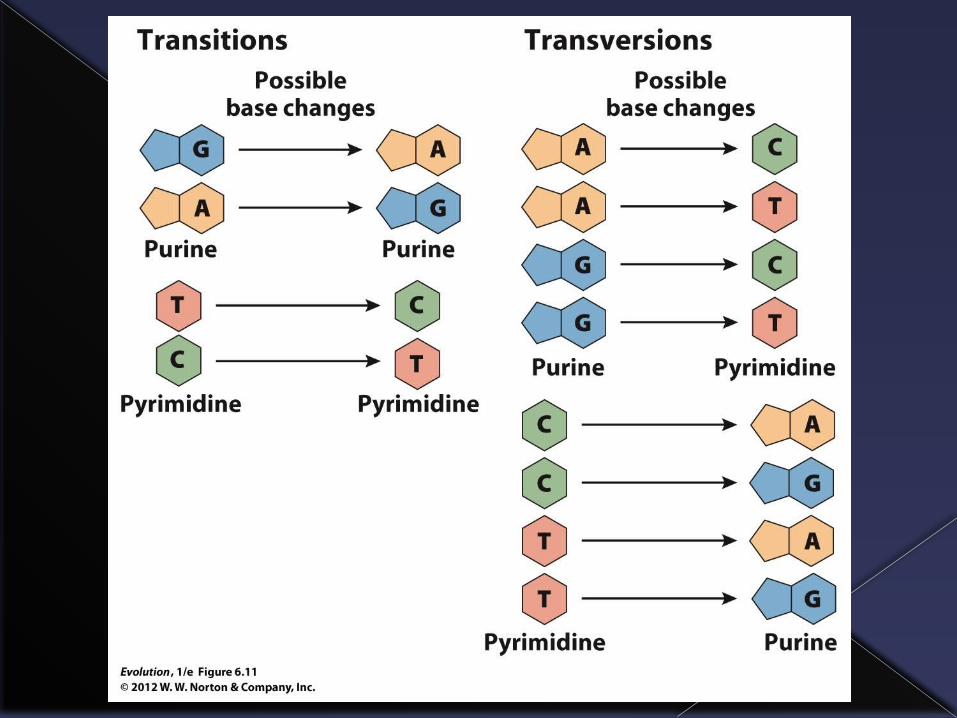

Types of mutations
Not all mutations cause a change in amino acid coded for. These are called silent mutations.
Mutations that do cause a change in amino acid are called replacement mutations.


Types of mutations
Another type of mutation occurs when bases are inserted or deleted from the DNA molecule.
This causes a change in how the whole DNA strand is read (a frame shift mutation) and produces a non-functional protein.
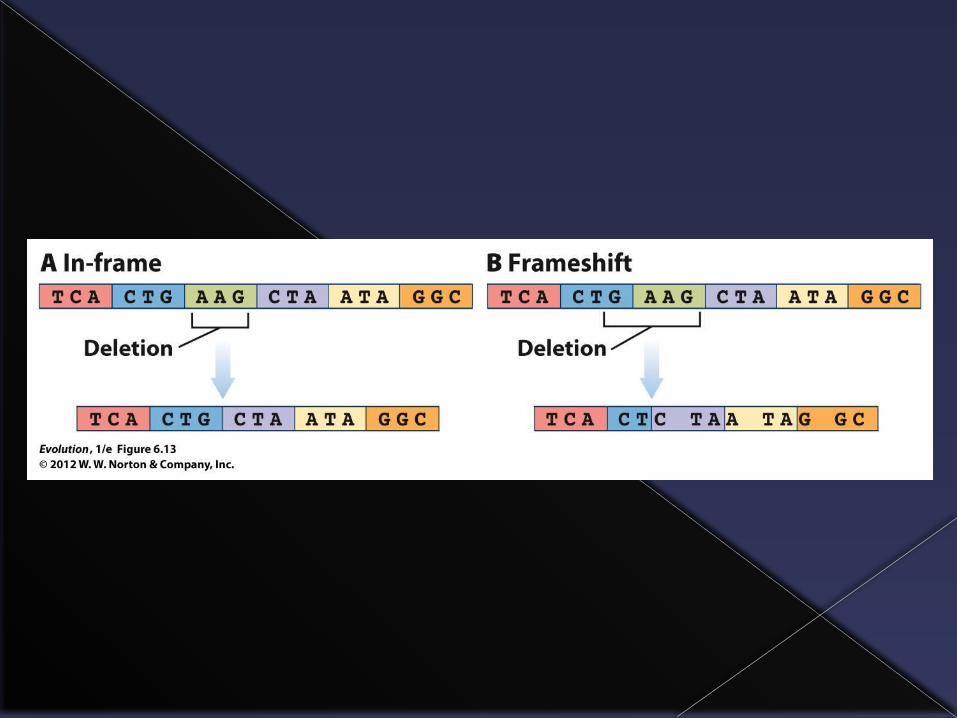

Another source of variation: Gene duplication
Duplication results from unequal crossing over when chromosomes align incorrectly during meiosis.
Result is a chromosome with an extra section of DNA that contains duplicated genes.


Gene duplication
Extra sections of DNA are duplicates and can accumulate mutations without being selected against because the other copies of the gene produce normal proteins.
Gene may completely change over time so gene duplication creates new possibilities for gene function.

Globin genes
Hemoglobin (the oxygen-carrying molecule in red corpuscles) consists of an iron-binding heme group and four surrounding protein chains (two coded for by genes in the Alpha cluster and two in the Beta cluster).

Example of gene duplication: Globin genes
Ancestral globin gene duplicated and diverged into alpha and beta ancestral genes about 450-500 mya.
Later transposed to different chromosomes and followed by further subsequent duplications and mutations.

From Campbell and Reese Biology 7th ed.

Globin genes
Lengths and positions of exons and introns in the globin genes are very similar. Very unlikely such similarities could be due to chance.

4.9
Exons (blue), introns (white), number in box is number of nucleotides.

Globin genes
Different genes in alpha and beta families are expressed at different times in development.
For example, in very young human fetus, zeta (from alpha cluster) and epsilon (from beta cluster) chains are present initially then replaced. Similarly G-gamma and A-gamma chains present in older fetuses are replaced by beta chains after birth.

4.8
Gestation (weeks) Post-birth(weeks)
Fetal hemoglobin has a higher affinity for oxygen than adult hemoglobin.Enhances oxygen transfer from mother to offspring.

D o mutations arise when needed?
It is important to realize that mutations arise randomly and are not biased towards making the organism better adapted.
In fact, most mutations are deleterious.

Luria and Delbruck’s experiment
The random nature of mutations was demonstrated by an elegant experiment by Luria and Delbruck in 1943.
At the time it was unclear if mutations occurred at random or if the environment somehow induced mutations when there was a need for them.

Luria and Delbruck’s experiment
Luria and Delbruck worked on the bacterium Escherischia coli and a bacteriophage virus that killed E. coli.
When E. coli on petri dishes was exposed to the virus most E. coli were infected and killed but a few resistant bacteria survived and produced colonies.

Luria and Delbruck’s experiment
L+D wanted to distinguish between two competing hypotheses.
1. Random mutation: before being exposed to phage some bacteria would have randomly developed mutations. These would survive and produce colonies.

Luria and Delbruck’s experiment
2. Acquired hereditary resistance. At time of exposure to phage all bacteria would be vulnerable. However, exposure to the phage would induce resistance in some small percentage of bacteria that would produce resistant colonies.

L+D experiment

Luria and Delbruck’s experiment
L+D grew phage sensitive bacteria to high densities in a broth and spread them on a plate covered with phage.
They then counted the number of colonies that grew after 24-48 hours.
Using just this count data they could distinguish between the two hypotheses.

Luria and Delbruck’s experiment
How were they able to use the count data to distinguish between the two hypotheses?
Logic depends on the timing of when mutations would have happened.

Luria and Delbruck’s experiment
If the presence of phage induced mutations in the bacteria then mutations would not occur until after the bacteria were exposed to the phage.
Because there were lots of bacteria in each culture and a nontrivial number should develop resistance by the law of large numbers we would expect a similar number of colonies to develop on each plate.

Luria and Delbruck’s experiment
In contrast under the random mutation model beneficial mutations could have arisen at any time before phage exposure.

Luria and Delbruck’s experiment
Thus we would expect in some cases for the mutation to have arisen well before exposure to phage and for the bacteria to have produced lots of descendents with the mutation.
In other cases the mutation may have arisen more recently and so few descendents with the mutation would be present.

Luria and Delbruck’s experiment
Therefore, under the random mutation scenario we would expect there to be a lot of variation from plate to plate in the number of resistant colonies seen.


Luria and Delbruck’s experiment
L+D repeated their experiment many times and counted the resistant colonies they found.
When they analyzed their data they found that the variance was much larger than the mean which supported the random mutation hypothesis and allowed them to reject the induced mutation hypothesis.

Ratio of beneficial to deleterious mutations
Very little data is available of the proportions of mutations that are beneficial versus deleterious.
However a recent study on a virus in which specific point mutations were artificially induced have shown that most are deleterious, about 40% neutral and only 2-3% beneficial.


Ratio of beneficial to deleterious mutations
Even though most mutations are neutral or deletrious the few beneficial mutations when combined with natural selection are what lead to evolution taking place.





























