Embed Size (px)
Citation preview

1
A deletion in NRT2.1 attenuates Pseudomonas syringae-induced hormonal perturbation, resulting
in primed plant defenses
Víctor Flors Herrero
Área de Fisiología Vegetal, Departamento de Ciencias Agrarias y del Medio Natural, ESTCE
Universitat Jaume I, Campus de Riu Sec. 12071, Castellón, Spain
Tel: +34 (964) 729417
FAX: +34 (964) 728066
E-mail: [email protected]
Plant Physiology Preview. Published on December 12, 2011, as DOI:10.1104/pp.111.184424
Copyright 2011 by the American Society of Plant Biologists
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

2
A deletion in NRT2.1 attenuates Pseudomonas syringae-induced hormonal perturbation, resulting
in primed plant defenses
Camañes Gemma1, Pastor Victoria1, Cerezo Miguel1, García-Andrade Javier2, Vicedo Begonya1,
García-Agustín Pilar1, Flors Victor1
1Área de Fisiología Vegetal. Departamento de Ciencias Agrarias y del Medio Natural, ESTCE
Universitat Jaume I, Campus de Riu Sec. 12071, Castellón, Spain 2Instituto de Biología Molecular y Celular de Plantas, Universidad Politécnica de Valencia-
Consejo Superior de Investigaciones Científicas, Ciudad Politécnica de la Innovación 46022,
Valencia, Spain
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

3
Funding source:
Generalitat Valenciana GV/2007/099, Plan Promoción Bancaja-UJI P1.1A2007-07 and P1.1B2007-
42
Corresponding author and e-mail address:
Víctor Flors; [email protected] (last position in order of appearance)
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

4
Abstract
For an efficient defense response against pathogens, plants must coordinate rapid genetic
reprogramming to produce an incompatible interaction. NRT2 gene family members are sentinels of
nitrate availability. In this study, we present an additional role for NRT2.1 linked to plant resistance
against pathogens. This gene antagonizes the priming of plant defenses against the bacterial pathogen
Pseudomonas syringae pv tomato DC3000 (Pst). The nrt2 mutant (which is deficient in two genes,
NRT2.1 and NRT2.2) displays reduced susceptibility to this bacterium. We demonstrate that modifying
environmental conditions that stimulate the de-repression of the NRT2.1 gene influences resistance to
Pst independently of the total level of endogenous nitrogen. Additionally, hormonal homeostasis
seemed to be affected in nrt2, which displays priming of salicylic acid (SA) signaling and concomitant
irregular functioning of the jasmonic acid (JA) and abscisic acid (ABA) pathways upon infection.
Effector-triggered susceptibility and hormonal perturbation by the bacterium seem to be altered in nrt2,
probably due to reduced sensitivity to the bacterial phytotoxin coronatine. The main genetic and
metabolic targets of coronatine in Arabidopsis remain largely unstimulated in nrt2 mutants. In addition,
a Pseudomonas syringae strain defective in coronatine synthesis (Pst COR-) showed the same virulence
towards nrt2 as the coronatine-producing strain did. Taken together, the reduced susceptibility of nrt2
mutants seems to be a combination of priming of SA-dependent defenses and reduced sensitivity to the
bacterial effector coronatine. These results suggest additional functions for NRT2.1 that may influence
plant disease resistance by downregulating biotic stress defense mechanisms and favoring abiotic stress
responses.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

5
Introduction
In higher plants, nitrate uptake through the roots is mainly achieved by two gene families, NRT1 and
NRT2. In Arabidopsis, there are 53 NRT1 genes and 7 NRT2 genes (Tsay et al., 2007). NRT1 codes for
components of the nitrate low-affinity transport system (LATS), which regulates nitrate uptake mainly
when it is present at high concentrations in the root environment. NRT1 can combine its LATS function
with a high-affinity transport system (HATS) function (Ho et al., 2009), and this dual role is regulated
by phosphorylation. Functional characterization has shown that NRT2 genes belong to the family of
nitrate high affinity transport systems (HATS) (Gansel et al., 2001). The NRT2.1 gene is activated when
nitrate concentrations are below 1 mM, and it mediates systemic signals to the shoot in split root
systems depending on the nitrate concentration (Cerezo et al., 2001; Gansel et al., 2001). The net
contribution to total nitrate uptake of NRT2 is low compared with NRT1 (Little et al., 2005; Tsay et al.,
2007). Interestingly, mutations in the NRT2.1 gene have no influence on total nitrate or nitrogen content
in normal fertilization conditions (Orsel et al., 2004). In addition, individual NRT2.1 mutations in
Arabidopsis seem to be compensated for by NRT2.2; therefore, it is necessary to have both genes
blocked to induce a significant reduction of nitrate uptake mediated by HATS (Li et al., 2007).
Therefore, additional roles for the NRT2 gene family have been proposed. Recently, the term
transceptor has been applied to membrane proteins that fulfill dual nutrient transport/signaling
functions . Although it is now clear that NRT1.1 can be considered a transceptor there is also evidence
suggesting a signaling role for NRT2.1 (Gojon et al., 2011). NRT2 genes are related to environmental
sensing and signal transduction that integrates the metabolism nitrogen with carbon metabolism (Little
et al., 2005). NRT1.1 and NRT2.1 can both perceive small amounts of nitrate and transmit signals to the
plant in order to integrate growth with nutrient availability (Krouk et al., 2010). Moreover, the 150-bp
promoter region of NRT2.1 is sufficient to mediate induction by nitrate and repression by nitrogen
metabolites (Castaings et al., 2010).
NAR2.1/NRT3.1 has been described as a trans-membrane protein that is necessary for NRT2.1
function, but it has no transporter activity by itself (Okamoto et al., 2006; Tsay et al., 2007). This
finding demonstrates complex regulation of the NRT2.1 gene, the role of which as a nitrate transporter
has been questioned, and which has been proposed to be an environmental signal sensor that controls
the development of the root system and coordinates it with nutritional cues (Little et al., 2005; Tsay et
al., 2007).
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

6
Nitrate uptake mediated by HATS has also been demonstrated to be regulated by hormonal and
metabolic signals in wheat (Cai et al., 2007). ABA and Gln enhance NRT2.1 and NAR2.1 gene
expression when nitrate is not present in the nutrient solution, but they have no effect when nitrate
fertilization is appropriate. This phenomenon suggests the existence of feedback between hormonal
signaling and NRT2.1-mediated environmental sensing (Cai et al., 2007).
Nitrate transporter mutants have been shown to display unusual phenotypes in response to abiotic stress
(Yao et al., 2008), but there is little or no evidence that links nitrate HATS activity to resistance to
biotic stresses. One of the few links between nitrogen transporters and biotic stress resistance is found
in the gene NAR2/NRT3.1, also known as WR3 (wound responsive3), which is induced in response to
wounding through a JA-independent pathway (www.arabidopsis.org).
Several findings have indicated that the nutritional status of a plant affects its defensive capacity
against the bacterium Pseudomonas syringae (Modolo et al., 2006). When the main nitrogen source is
ammonium, Arabidopsis is more sensitive to Pseudomonas syringae infection. Feeding plants with
ammonium as the sole nitrogen source prevents nitric oxide (NO) formation and inhibits the
hypersensitive reaction (HR), which correlates with altered susceptibility. By contrast, low levels of
nitrate favor NO formation through enhancement of the NO-synthase activity of nitrate reductase (NR;
Modolo et al., 2005). Overfertilization leading to elevated internal nitrogen levels also contributes to
rice plant susceptibility (Long et al., 2000). This result could be explained by inhibition of the NO-
synthase activity of NR and stimulation of its reductase activity (Modolo et al., 2005). Although the
reasons for these alterations of basal plant resistance are not yet clear, net nitrate rates influence the
dual activity of NR and alter the generation of reactive oxygen species (ROS) and NO. This result may
have a direct effect on plant defense responses against pathogens. Lozano-Juste and León (2010) have
shown that nia1nia2noa1 triple mutants (genes involved in nitrate reductase enzyme and NO synthesis
respectively) display enhanced ABA sensitivity and drought tolerance, due to the uncoupling of NO
from ABA-triggered responses linked to defense responses. In addition to these indications, alterations
in amino-acid transporters, which are tightly regulated by nitrogen status, have direct effects on disease
resistance. For example, the lysine transporter mutant lht1 displays constitutive activation of SA-
dependent signaling and resistance to Pseudomonas syringae (Liu et al., 2010)
Although the constitutive expression of defense genes results in resistant plants, this property usually
has a negative impact on normal plant development (van Hulten et al., 2006). Therefore, among plant
defense strategies, priming has emerged as a notably effective one (Ton et al., 2005; Conrath et al.,
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

7
2006; Conrath, 2011). Plants, upon appropriate stimulation, can activate faster and stronger defense
signals by potentiating their basal immune system. Interestingly, van Hulten and coworkers (2006)
have demonstrated that primed responsiveness to SA in the edr1 mutant correlates with elevated levels
of basal resistance against Pst with no major loss of plant growth or seed production. Although the
mechanisms underlying priming are not fully known, the accumulation of dormant mitogen-activated
protein kinases and chromatin remodeling are two processes likely to be involved in this phenomenon
(Conrath, 2011). Although a link has not yet been established, the upregulation of SA-dependent
responses may be under the control of such processes.
In this study, we demonstrate that alterations in the NRT2 gene family modify basal susceptibility to the
bacterial pathogen Pst. The nrt2 mutant is less susceptible to Pst, and this reduced susceptibility
correlates with both SA pathway priming and reduced sensitivity to pathogenic effectors. These results
suggest an additional role for membrane-located nitrate transporters in environmental stress perception
and signal transduction.
Results
Disruption of NRT2.1 and NRT2.2 results in reduced susceptibility to Pst
Little et al., (2005) demonstrated that NRT2.1 acts as a nitrate sensor and signal transducer to
coordinate the development of the root system with nutritional and abiotic cues. Given the similarities
and close link between biotic and abiotic sensing (Jakab et al., 2005; Ton et al., 2005), the levels of
basal resistance against the hemi-biotrophic pathogen Pst were tested in the mutant nrt2. To this end,
normally fertilized Ws and nrt2 plants were inoculated by dipping at two different stages of growth.
Both adult plants and seedlings of nrt2 showed reduced susceptibility to the bacteria, as evidenced by
reduced symptom severity and reduced bacterial proliferation (Fig 1A, B and C).
Because total levels of nitrogen in a plant have a strong influence on pathogen colonization and
proliferation ( Hoffland et al., 1999; Ali et al., 2009), we determined whether alterations in NRT2 gene
expression or short periods of nitrate deprivation influence total N content, because it can induce the
expression of several NRT2 family members (de-repression: Lejay et al., 1999). Therefore, we
transferred hydroponically growing plants to nitrate-free medium before inoculation. Forty-eight hours
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

8
of deprivation was not sufficient to modify total N levels in plants that had been grown with 1 mM of
NH4NO3 for 6 weeks (Fig S1A). We also confirmed that NRT2.1 expression was enhanced not only in
roots but also in leaves in both ecotypes tested (Ws and Col-0; Fig S1B, C). Interestingly, after the
induction of NRT2 gene expression by nitrate depletion, Ws plants became more sensitive compared to
normally fertilized Ws plants, whereas the level of resistance of nrt2 plants did not change (Fig1E).
This finding might be attributable to the plants’ lack of responsiveness to root environmental sensing.
Therefore, the de-repressed (active transcription) versus repressed (non transcribed) state of NRT2.1
affects the basal resistance level of Arabidopsis against Pst.
It is noteworthy that nrt2 is a mutant that lacks the function of both the NRT2.1 and NRT2.2 genes (Li
et al., 2007). To clarify whether the reduced susceptibility of nrt2 is due to one or both genes, we tested
basal resistance in individual mutants, such as lin1 (blocked in NRT2.1; Little et al., 2005), the
knockout mutant Salk_043543 and the double knockout mutant Salk_035429, all in the Col-0
background. The single NRT2.1 mutant lin1 and the double knockout (Salk_035429) also showed
reduced susceptibility, but the single mutant Salk_043543 did not, suggesting that mutation of both
genes may not be necessary to obtain significantly reduced bacterial growth (Fig 1D). The fact that
Salk_035429 and lin1 are altered in their basal resistance reinforces the hypothesis that these genes
play additional roles in stress sensing and/or signal transduction rather than being only involved in
nitrate transport at low concentrations. In fact, NRT2.1 expression after nitrate depletion was lower on
Col-0 than in Ws, and this finding correlates well with the naturally greater resistance of Col-0. This
natural genetic variation has been recently reported to be linked to two QTLs, one of which regulates
SA-induced PR1 expression (Ahmad et al., 2011).
nrt2 shows primed SA-dependent responses upon Pst infection
Using chemical agents, such as β-aminobutyric acid (BABA), benzothiadiazole (BTH) or azelaic acid,
to prime SA-dependent defenses results in enhanced resistance against Pst (Conrath et al., 2006; Jung
et al., 2009). The mutant nrt2 was found to display slightly increased levels of PR1 and PR5 compared
to those of Ws in the absence of infection (Fig 2A). Interestingly, when the infection is present nrt2
showed stronger and faster induction of these SA marker genes (Fig 2A), mainly at early time-points.
Accordingly, SA levels were higher in nrt2 at 48 h after infection (Fig 2B).
Salk_035429 and lin1 in the Col-0 background showed enhanced PR1 and SA responses concomitant
with their reduced susceptibility to the bacterium (Fig S2). To assess the relevance of SA priming in
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

9
nrt2, we crossed lin1 and Salk_035429 with sid2.1 (isochorismate synthase deficient, Wildermuth et
al., 2001). We tested two independent lines of the double and triple mutants lin1-sid2.1 and
Salk_035429 -sid2.1, and all of them displayed wild-type levels of resistance, which demonstrates that
enhancement of the SA pathway is necessary component of the reduced susceptibility of nrt2 (Fig 2C).
The potential hypersensitivity of nrt2 to SA was tested by treating the plants with β-aminobutyric acid.
BABA was effective at protecting both wild-type and nrt2 to the same levels (Fig S3). Therefore, the
mutant retains its sensitivity to the SA priming induced by BABA.
nrt2 has reduced sensitivity to Pseudomonas syringae effectors
Among the many impacts of bacterial effectors, manipulation of plant hormones is one of the main
targets. The role of ABA in Pst-Arabidopsis interactions is complex and not fully understood (Ton et a.,
2009). It is known that Pst can take control of stomatal movements during pre-invasive and post-
invasive stages by hijacking ABA signaling (Melotto et al., 2008; Ton et al., 2009; Torres-Zabala et al.,
2009). We checked for a contribution of ABA to nrt2-reduced susceptibility by studying the expression
of several ABA marker genes and ABA levels. We monitored three different ABA-responsive genes,
specifically, ABI1, RD22 and RAB18. The expression of ABI1 was upregulated by infection in nrt2, but
it remained unchanged in Ws. By contrast, RD22 was downregulated by the bacterium. Surprisingly,
both ABA marker genes showed lower expression in nrt2 than in Ws in the absence of challenge.
RAB18 showed the most dramatic change, as it was upregulated by the bacterium in Ws but displayed
lower expression in the mutant after infection (Fig 3A). This situation may lead to deficient ABA
signaling manipulation by the bacterium (Ton et al., 2009). Accordingly, the levels of ABA remained
unchanged during infection in the nrt2 mutant, whereas ABA levels increased in wild-type plants (Fig
3B).
To study the abnormal ABA signaling in nrt2, its ability to sense this hormone was verified by treating
Ws and nrt2 plants with ABA 48 h before the infection (Fig 3C). ABA treatment increased the disease
rate in both wild-type and mutant plants; however, bacterial growth only showed significative
differences in ABA-treated nrt2 plants, probably due to the high inoculum used for all the experiments.
The observation of the altered behavior of ABA marker genes in nrt2 in the absence of infection
prompted us to test the response of nrt2 plants to drought stress; however, the mutant showed wild-type
levels of water loss and tolerance (Fig S4). Therefore, ABA, which is implicated in abiotic stress
signaling, seems to function normally in nrt2 plants. Subsequently, we studied whether coronatine
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

10
sensitivity was altered in this mutant because it has been reported that coronatine alters ABA
functioning during infection (Torres-Zabala et al., 2007; Melotto et al.,2008). To this end, several
bioassays were performed. In the first experiment, the resistance of nrt2 mutants against Pst COR- was
tested in both the Ws and Col-0 backgrounds. Interestingly, the absence of coronatine eliminated the
reduced susceptibility in all individual and double mutants compared with their corresponding wild-
type plants (Fig 4, Fig S5). We subsequently complemented the Pst COR- by treating both genotypes
with coronatine (Fig 4B; Brooks et al., 2004). The result confirmed that the initial reduced
susceptibility phenotype of nrt2 was reestablished; therefore, it is clear that the presence of coronatine
makes a contribution to the reduced susceptibility of this mutant.
This indication that nrt2 displays altered effector sensitivity was further verified by treating Ws and
nrt2 plants grown in MS medium with the effector coronatine (Fig 5). Treatment with coronatine
significantly induced H2O2 (a marker of coronatine's effect) in Ws, whereas nrt2 plants remained
unaltered after treatment (Fig 5A, B). This finding suggests that H2O2 may be a target of coronatine in a
pathway requiring NRT2.1. An additional experiment confirmed the insensitivity of nrt2 plants to
coronatine. Ws and nrt2 plants were sprayed with coronatine. After the treatment, detached leaves of
Ws remained turgid, while nrt2 leaves presented clear wilting at 4 and 6 hours post-treatment (hpt), as
did water-treated leaves (Fig 5C). This result might be due to an induction of stomatal closure at 4 and
6 hpt by coronatine in Ws, while in nrt2 plants the stoma would remain unaltered. We also infected
Arabidopsis by direct infiltration to circumvent possible stomatal aperture defects in the mutant.
Surprisingly, nrt2 remained resistant, suggesting that coronatine sensitivity may contribute to its
enhanced resistance but is not essential for it (Fig S6). Finally, we performed the classical test of
sensitivity of germination to coronatine. In this case, nrt2 showed wild-type sensitivity upon coronatine
treatment, which was visible as purple leaves and a delay in germination (Fig 5D).
Another major target of coronatine is the COI1 receptor and subsequent JA pathway activation
(Fonseca et al., 2009). Accordingly, the JA pathway showed slower activation in nrt2 compared with
Ws. The JA marker genes VSP2 and MYC2, which are not reduced in the mutant in the absence of
infection, showed a lower level of induction at 24 hpi in nrt2 upon infection (Fig S7A). The fact that
these genes are not altered without pathogen confirms that the negative influence of SA priming in the
mutant is only present when the plant is challenged. In addition, infected mutants displayed reduced JA
levels compared to infected wild-type plants (Fig S7B).
Therefore, the downregulation of ABA and JA signaling after Pst infection in nrt2 could be a
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

11
consequence of a partial loss of bacterial effector sensitivity rather than a defect in those pathways.
Gene expression analysis confirms the reduced sensitivity of nrt2 to coronatine.
We tested whether nrt2 plants present abnormal responses to direct coronatine treatment. We observed
that coronatine treatment upregulated such genes as PDF1.2, CORI1 and CORI3, however such
induction was always lower in nrt2 compared with wild-type (Fig S7 C). In contrast to Pst infection
(Fig 3A), RAB18 was downregulated by direct treatment with coronatine in Ws, whereas in nrt2 it
remained unaltered (Fig S7 C). This result reinforces the hypothesis that nrt2 is less sensitive to
coronatine-induced alterations in the plant.
To characterize the molecular response of nrt2 to Pst further, whole-genome transcriptional profiles of
Ws and nrt2 plants mock-treated or infected with Pst were obtained using Arabidopsis ATH1 arrays
from Affymetrix. An examination of the differentially expressed genes in Ws and nrt2 upon infection
compared with mock treatment revealed a group of 574 genes that were differentially expressed in the
mutant but were unchanged in Ws, whereas 550 genes were up- or downregulated in Ws as a
consequence of infection but remained unaltered in nrt2 (Fig S8, Table S2).
Following the line of evidence suggesting that nrt2 has reduced sensitivity to bacterial effectors, we
selected genes from the array that are induced by coronatine. This set of genes was described by Tsai et
al. (2010) after a comparative full genome analysis of plants infected by a coronatineless Pseudomonas
syringae strain or by a strain with functional coronatine. In addition, a second comparative analysis
performed by Tsai and coworkers discriminated from among all of the coronatine-inducible genes a
group of 31 genes that were downregulated by the priming agent BABA. We looked at the behavior of
this set of genes in our study and, interestingly, 18 out of 31 genes repressed by BABA were also less
induced in infected nrt2 plants compared with wild-type-infected plants (Fig S8B, Table S3). This
finding supports the idea that in the absence of NRT2, the plant displays behavior similar to that of
BABA-primed plants.
Discussion
In the present work, we investigated additional roles for the NRT2.1 gene. Mutations affecting NRT2.1
reduce the susceptibility of Arabidopsis to the virulent pathogen Pst. nrt2 mutants displays primed SA
signaling upon infection (Fig 1 and 2). In addition, these plants present defects in sensitivity to the
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

12
bacterial effector coronatine and downregulation of the JA pathway, probably as a consequence of SA
priming, reduced coronatine sensitivity or both (Fig S7). This result suggests that one or several
components of the NRT2 gene family may function as environmental sensors, coordinating not only
root and nutritional cues but also abiotic and biotic responses by enhancing abiotic responses and
downregulating plant defenses against biotic challenges. The term transceptor has been applied to such
genes as NRT1.1 that act as transporters but also coordinate other features of plant development and N
metabolism (Gojon et al., 2011). In fact, mutants defective for NRT1 family members are impaired in
NRT2.1 regulation, which highlights the tight regulatory link between nitrate perception by NRT1.1 and
the function of other NRT family members. NRT2.1 and NRT2.2 have been proposed to play roles in the
integration of sugar and amino-acid metabolism. Other studies point to NRT2.1 as a putative abiotic
stress sensor that coordinates the development of the root system with the root environment and
nutrition (Little et al., 2005; Tsay et al. 2007). It is worth mentioning that downstream nitrate uptake
mutants, such as nia1nia2noa1, display hypersensitivity to ABA and faster stomatal closure upon
drought stress, being subsequently more drought-tolerant (Lozano-Juste and León, 2010).
In our research, we were interested in studying links between nitrate perception and biotic stress
responses. Several reports have described changes in plant susceptibility to Pst that depend on the
nitrogen, nitrate or ammonium content of the plant (Long et al., 2000; Modolo et al., 2005, 2006). To
avoid this possible interference, we performed all experiments with normally fertilized plants (unless
otherwise mentioned). Arabidopsis fertilized with ammonium nitrate at 1 mM has its LATS activated
and NRT2.1, NRT2.2 and NIA repressed (Krouk et al., 2010). In this condition, we show that mutants
lacking either NRT2.1 or NRT2.2 have reduced susceptibility (Fig 1). This phenotype is due to the lack
of the nitrate HATS because activation (de-repression) versus downregulation (repression) of these
genes has clear consequences on wild-type Ws susceptibility to the bacterium.
One mechanism potentially able to explain the reduced susceptibility of nrt2 is found in the SA
signaling pathway. nrt2 mutants show priming of SA signaling at early stages of infection, as
confirmed by SA marker gene induction and hormone accumulation. SA priming is one of the main
mechanisms of induced resistance against Pst (Zimmerli et al., 2000; Conrah et al., 2006). There are
several priming inducers (such as BTH, BABA and azelaic acid) that clearly reduce bacterial disease
symptoms and plant tissue colonization by stimulating the SA pathway (Conrath et al., 2006; Jung et
al., 2009). Interestingly, nrt2 mutants experience constitutive priming because basal levels of SA in the
absence of infection are the same as in Ws, and this finding has also been reported for other mutants,
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

13
such as edr1 (enhanced disease resistance1) (van Hulten et al., 2006). Mutants with direct activation of
SA signaling, such as cpr1, are highly resistant to biotrophic pathogens, but this condition slows
growth and reduces seed production (van Hulten et al., 2006). The nrt2 mutant displays no phenotype
in the absence of the pathogen, and it shows enhanced defense responses upon infection similarly to
edr1. Therefore, NRT2.1, bacterial perception and SA seem to be linked. Genevestigator stimulus
analysis showed that both programmed cell death (PCD) (a possible mechanism of HR) and flg22
induce NRT2.1 expression strongly (data not shown). Recently, Ward et al. (2010) have highlighted by
metabolomic analysis that nitrogen containing compounds and amino acids are strongly altered upon
Pst infection. Increasing evidence points to Gln reallocation in the cytoplasm as a checkpoint that
clearly interacts with SA signaling. Wu et al. (2009) reported that Gln strongly represses BABA
priming against Pst by downregulating PR1. In addition, lysine and histidine transport (LHT1)
mutants show strongly affected disease resistance (Liu et al., 2010). The LHT1 gene antagonizes SA-
dependent signaling, and it is upregulated by pathogen attack. lht1 mutants show upregulated PR1,
NRT1.1 and AMT1.1, among others. In addition, the elevated resistance of lht1 seems not to be directly
linked to global N status as it is with NRT2.1 (Liu et al., 2010). Thus, it seems clear that some of the
genes regulating nitrogen and amino-acid metabolism or transport have strong regulatory functions that
play roles in plant-pathogen interactions.
Several studies suggest that hormones transfer nitrate signals, which would explain the context of
nitrate's influence on plants. INF1, an elicitin from Phytophthora infestans, induces NRT2.1 expression
in tobacco, and it stimulates many SA-dependent responses and HR. Thus, SA signaling and NRT2.1
seem to be coordinately upregulated by this pathogenic elicitor (Kawamura et al., 2009). It is likely that
such oomicetes that combine necrotrophyc and biotrophyc lifestyles target certain plant genes related to
nutrition in order to improve the plant's nutrient sources to their advantage. It is noteworthy that a tenth
of the genome is under the control of nitrate (Gutierrez et al., 2003), and interestingly, a nitrate-
regulated biomodule of genes is also controlled by plant hormones (Krouk et al., 2010).
Our experiments disproved the hypothesis that irregular functioning of the ABA signaling pathway
upon infection is responsible for the reduced susceptibility of nrt2 mutants (Fig 3). The results indicate
that ABA signaling functions normally in these mutants because they show wild-type responses to
drought stress. In addition, external treatment with ABA increases the susceptibility of nrt2 mutants to
Pst, as was previously reported for wild-type plants (Mohr and Cahill, 2003).
ETS (effector triggered susceptibility, Jones and Dangl, 2006) is one of the main roles of the bacterial
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

14
effector coronatine. Several studies have demonstrated that coronatine targets genes to stimulate the JA
signaling pathway in order to repress SA signaling and take advantage of plant colonization. In
addition, coronatine also hijacks ABA functions, such as stomatal closure in the pre-invasive stages by
opening stomata and during post-penetration stages by closing stomata (Torres Zabala et al., 2007; Ton
et al., 2009). In all cases, coronatine plays an active role in bacterial infection, but in our experiments,
we found that nrt2 mutants were partially unable to sense coronatine. This bacterial effector failed to
induce H2O2 in nrt2 mutants, and it was also unable to reduce wilting in detached mutant leaves,
probably due to a failure of stomatal closure (Fig 4 and 5). In addition, nrt2 shows a wild-type degree
of basal resistance to Pst COR-, indicating that the presence of the effector is necessary to observe the
reduced susceptibility of nrt2 mutants. Tsai et al. (2010) obtained similar results using the chemical
priming agent BABA. These researchers determined that a functional coronatine response was needed
to express BABA priming and induce resistance against Pst. These observations, together with the fact
that many genes induced by coronatine are downregulated in nrt2, explain why the reduced
susceptibility of nrt2 disappears when the bacterium lacks coronatine. To conclude, nrt2 mutant is less
sensitive to coronatine-triggered susceptibility, and this property reduces the ability of the bacterium to
colonize the mutant. Another interesting result of these experiments is that hydrogen peroxide is a
putative target of coronatine, and this interaction is NRT2.1–dependent, as nrt2 is totally impaired in
H2O2 production upon coronatine treatment.
Tsai et al. (2010) reported that priming by BABA against Pst acts through the specific inhibition of
coronatine-triggered responses, and this inhibition resembles nrt2 phenotypes that match partial
coronatine insensitivity with SA-dependent priming and reduced susceptibility to the bacterium.
Among all of the potential contributions to the reduced susceptibility of nrt2 mutants, SA priming
seems to be the major player. In this study, we demonstrated the absence of the resistance phenotype in
the triple and double mutants Salk_035429 -sid2.1 and lin1-sid2.1 (fig 2). However, the reduced
sensitivity to coronatine may also contribute to the resistance; therefore, the phenotype could be a
combination of both. The fact that most experiments performed with NRT2 mutants in the Col-0 and
Ws backgrounds have obtained almost identical results reinforces our main conclusions. However, the
reduced SA-signaling priming and the lower resistance of lin1 and Salk_035429 mutants is justified
because Col-0 displays natural SA-priming and thus enhanced resistance to Pst compared with Ws
(Ahmad et al., 2011). Regarding the SA-JA crosstalk, it is likely that the SA priming is also a
consequence of the absence of repression by JA signaling, which would be less stimulated, due to the
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

15
reduced coronatine sensitivity. This interpretation is further confirmed by the delay in MYC2 induction
in nrt2 compared with wild-type because MYC2 mediates the suppression of SA-dependent defenses by
coronatine (Fig 6). This crosstalk has been recently revised by Pieterse et al. (2009).
Indirectly, nrt2 mutants show attenuated Pst-induced hormonal perturbations that result in the priming
of plant defenses and reduced susceptibility.
In conclusion, although the full implications of the nitrate transporter NRT2 in biotic and abiotic
stresses remain to be clarified, the implications may be linked though the enhancement of abiotic stress
sensing and coordinated biotic stress repression. At the same time, this transporter could be a direct or
indirect target used by bacterial pathogens to take advantage of plant colonization. This research
suggests that NRT2.1 is a novel transceptor of the nitrate transport family that regulates plant disease-
resistance signaling.
Material and Methods
Plant material and growth conditions
Wild-type Arabidopsis thaliana Wassilewskija (Ws) and the Ws mutant nrt2.1-2.2 (referred to as nrt2
in the text) (Filleur et al., 2001) were generously provided by Alain Gojon (INRA Montpelier France),
the knockout lines nrt2.1-2.2 (referred to as Salk_035429 in the text) and nrt2.2 (referred to as
Salk_043543 in the text) in the Col-0 background were obtained from the SALK institute (Alonso et
al., 2003) and the EMS lin1 line was generously supplied by Jocelyn Malamy (University of Chicago,
EEUU).
All plant genotypes were germinated in soil, and 2 weeks after germination, the seedlings were
individually transferred to 33-ml pots containing commercial potting soil (TKS1, Floragard GmbH;
http://www.floragard.de) (adult plants), or approximately 50 seedlings were germinated in 33-ml pots
containing commercial potting soil (seedling plants). The plants were cultivated at 20 ºC day/18 ºC
night with 8.5 h of light (105 μE m-2 s-1) per 24 h and 60% relative humidity.
Growth conditions for starvation experiments
Wild-type Arabidopsis thaliana (Ws) and the mutant nrt2 were grown hydroponically, as described in
Lejay et al. (1999). The seeds were germinated directly on top of modified Eppendorf tubes filled with
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

16
pre-wetted sand. Plants were grown until the age of 6 weeks on a 1 mM NH4NO3 nutrient solution
(repressed plants), which prevented any growth difference between the two genotypes (Lejay et al.,
1999). Before inoculation experiments, the plants were transferred for 48 h to nitrogen-free solution
(de-repressed plants).
Bacterial strains and bacterial growth assays
The bacterial strains were Pseudomonas syringae pv tomato DC3000 (Pst) and Pseudomonas syringae
pv tomato DC3000 cma (Pst COR-; Brooks et al., 2004). Bacteria were grown overnight at room
temperature in King's B solid medium with appropriate antibiotics and diluted to the desired
concentration with 10 mM MgSO4 for plant inoculation. These bacteria were used to infect 5-week-old
(or otherwise mentioned) Arabidopsis plants by dipping in a suspension of 2.5 x 107 colony-forming
units (c.f.u)/ml using 0.02% Silwet L-77 (Tornero and Dangl, 2001). After incubation at 28 ºC for 3
days, the number of rifampicin-resistant colony-forming units per gram of infected leaf tissue was
determined, and bacterial proliferation over the 3-day time interval was calculated. Three days after the
challenge inoculation, the percentage of leaves with symptoms was determined per plant (n= 20–25).
Leaves showing necrotic or water-soaked lesions surrounded by chlorosis were scored as diseased.
Double and triple mutant generation and selection of homozygous lines
The sid2.1 mutant was crossed with Salk_035429 and lin1 (all in the Col-0 background), and the
resulting F1 generation was self-crossed. Homozygous F2 seedlings were selected by PCR, and F3
seeds were used for the experiments. DNA was isolated from leaves of individual plants (Edwards et
al., 1991). To identify plants that were homozygous for the sid2.1 mutation, the following primers were
used: sid2.1-F 5’-gctctgcagcttcaatgc-3’ and sid2.1-R 5’-cgaagaaatgaagagcttgg. A fragment of 243 nt
was obtained and subsequently digested by the enzyme Tru1 (MBI Fermentas, Vilnius, Lithuania) and
separated on a 2 % agarose gel to evaluate the fragment sizes of 154 nt and 89 nt in the mutant. Next,
homozygous sid2.1 plants were selected to find the other mutations in homozygosity. In this case, the
primers used for Salk_035429 and lin1 were, respectively, LP 5'-gcaagcgactatcatcactcc-3', RP 5'-
gttctccatgagcttcgtgag-3' and LB 5'-atttgccgatttcggaac-3', following the SALK institute instructions for
Salk_035429 and lin-F 5'-atcaaatctcaaacttgcaaagaac-3' and lin-R 5'-ggggaccaaaagatcacacacggatc-3'. To
identify plants homozygous for lin1, the amplification product from the wild-type allele is cleaved with
BamH1 (MBI Fermentas, Vilnius, Lithuania), and the product from the lin1 mutant allele is uncut.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

17
These fragments were separated in a 2 % agarose gel to evaluate the fragment sizes.
Quantitative real-time RT-PCR analysis of transcripts
Gene expression analysis by quantitative real-time RT-PCR was performed using RNA samples
extracted from leaf tissue using the Total Quick RNA cells and tissues kit (Talent;
http://www.spin.it/talent). Arabidopsis leaf tissue samples for RNA isolation were collected at 24 and
48 h after inoculation. Leaf tissue from five plants each of the mutant and wild-type plants was
collected. For quantitative real-time RT-PCR experiments, 1.5 μg of total RNA was digested using 1
unit of RQ1 RNase-Free DNase (Promega; http://www.promega.com) in 1 μl of DNase buffer and up to
10 μl of Milli-Q water and was incubated for 30 min at 37 ºC. After the incubation, 1 μl of RQ1 DNase
stop buffer was added, and the solution was incubated again at 65 ºC for 10 min to inactivate the
DNase. Highly pure RNA was used for the RT reaction. The RT reaction was performed by adding 2 μl
of RT buffer, 2 μl of 5 mM dNTP, 2 μl of 10 μM oligo(dT)15 primer (Promega), 1 μl of 10 U μl–1
Rnasin RNase inhibitor (Promega) and 1 μl of Omniscript reverse transcriptase (Qiagen,
http://www.qiagen.com/). The reaction mixture was incubated at 37 ºC for 60 min. Less than 10 % of
the volume of the RT reaction was used for the quantitative PCR. Forward and reverse primers (0.3
μM) were added to 25 μl of QuantiTectTM SYBR Green PCR reaction buffer (Qiagen), as were 2 μl of
cDNA and Milli-Q sterile water up to 50 μl total reaction volume. Quantitative PCR was carried out
using the Smart Cycler II sequence detector (Cepheid; http://www.cepheid.com). PCR cycling
conditions consisted of an initial HotStarTaq (Qiagen; http://www.qiagen.com) polymerase activation
step at 95 ºC for 15 min followed by 45 cycles of 95 ºC for 15 sec, 60ºC for 30 sec and 72 ºC for 30
sec. Melting-curve analysis was performed at the end of the PCR reaction to confirm the products'
purity. Differences in cycle numbers during the linear amplification phase between samples containing
cDNA from treated and untreated plants were used to determine differential gene expression. The β-
tubulin and At1g13320 genes of Arabidopsis were used as housekeeping genes. Normalized values of
gene expression are presented on a log2 scale. A list of the primers used in the qRT-PCR is shown in
Table S1.
Determination of ABA, SA and JA levels
Fresh material was frozen in liquid nitrogen and lyophilized. Before extraction, a mixture of internal
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

18
standards containing 100 ng [2H6]- ABA, 100 ng [2H4]-SA and 100 ng of dihydrojasmonic acid
(Pinfield-Wells et al., 2005) was added. Dry tissue (0.05 g) was immediately homogenized in 2.5 ml of
ultrapure water. After centrifugation (5000 g, 40 min), the supernatant was recovered and adjusted to
pH 2.8 with 6 % acetic acid and subsequently partitioned twice against an equal volume of diethyl
ether. The aqueous phase was discarded, and the organic fraction was evaporated in a Speed Vacuum
Concentrator (Eppendorf; www.eppendorf.com) at room temperature, and the solid residue was
resuspended in 1 ml of a water/methanol (90:10) solution and filtered through a 0.22-μm cellulose
acetate filter. A 20-μl aliquot of this solution was directly injected into the HPLC system. Analyses
were carried out using a Waters Alliance 2690 HPLC system (Milford, MA, USA) with a nucleosil
ODS reversed-phase column (100 x 2 mm i.d.; 5 ml; Scharlab, Barcelona, Spain; http://
www.scharlab.es). The chromatographic system was interfaced to a Quatro LC (quadrupole-hexapole-
quadrupole) mass spectrometer (Micromass; http://www.micromass.co.uk). MASSLYNX NT software
version 4.1 (Micromass) was used to process the quantitative data from calibration standards and the
plant samples.
Microarray analysis
Leaves of wild-type Arabidopsis thaliana Wassilewskija and the Ws mutant nrt2 infected with Pst and
not infected were taken for total RNA preparation using TRIZOL (Invitrogen) reagent. Total RNA was
purified with the RNeasy Mini kit (Qiagen). Double-stranded cDNA was synthesized with the one-
cycle cDNA Synthesis Kit (Affymetrix) and was purified with the GeneChip Sample Cleanup Module
(Affymetrix). The purified cDNA was used to prepare biotin-labeled cRNA using a GeneChip IVT
Labeling Kit according to the manufacturer's instructions. The biotin-labeled cRNA was fragmented at
94 ºC for 35 min, which yielded the probes used for hybridization. The probes were hybridized with the
Affymetrix ATH1 array, and washing and scanning were carried out according to the assay procedure.
The hybridization image was analyzed with Affymetrix Microarray Suite 5.0 software, and the data
were normalized. Three biological replicates from three different experiments were used for
transcriptomic comparisons. All basic operations, array hybridization, image analysis and statistical
analysis were performed by Progenika Co® (Bilbao, Spain). Bioinformatic analysis was performed by
using the PartekGenomics Suite dChip (www.dchip.org) and the software Affy and affyPLM from the
consortium BioConductor (www.bioconductor.org).
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

19
Chemical treatments
Two-week-old seedlings were individually transferred to 33-ml pots. At the age of five weeks, plants
were soil-drenched with water (control) or a solution of BABA or ABA at a final concentration of 250
μM or 80 μM, respectively. Two days after the chemical treatment, the plants were inoculated, as
described above.
Growth conditions and coronatine treatment in in vitro assays
Approximately 15 sterilized Col-0 seeds were sown per well in sterile 12-wells plates containing filter-
sterilized MS medium amended with 1% sucrose. Seedlings were cultivated under standard growth
conditions (15 h day cycle; 20ºC /17ºC) with a light intensity of 105 μE m-2 s-1. After 7 days, the
growth medium was replaced with fresh MS medium. One day later, the plants were challenged with
coronatine (Sigma C8115) at a final concentration of 0.5 ng/μL in the growth medium. At day 9, the
plants were stained with 3,3'-diaminoenzidine (DAB; Sigma, D-8001) to quantify hydrogen peroxide
induction (Thordal-Christensen et al., 1997). Samples were stained for 8 h and were subsequently de-
stained with 95% ethanol. Microscopic analysis was performed using a microscope (Nikon Eclipse
11000) with a VIS filter. Hydrogen peroxide was quantified as the relative number of brown pixels in
digital photographs using GIMP 2.6 software.
Supplementary Material
Supplementary Figure 1. Total nitrogen levels and NRT2 transcript levels in leaves and roots of Ws and
Col-0 plants.
Supplementary Figure 2. Effect of Pst infection on PR1 gene expression levels and SA accumulation in
five-week-old Col-0 wild-type plants, Salk_035429 and lin1 mutants.
Supplementary Figure 3. Disease rate and bacterial proliferation in Ws wild-type plants and nrt2
mutants infected with Pst upon BABA treatment.
Supplementary Figure 4. Water loss in Ws wild-type and nrt2 plants.
Supplementary Figure 5. Bacterial proliferation in Col-0 wild-type, lin1 and Salk_035429 plants
infected with Pst and the coronatineless strain Pst COR-
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

20
Supplementary Figure 6. Bacterial proliferation in Ws wild-type and nrt2 plants infected by infiltration
with Pst.
Supplementary Figure 7. Effect of Pst infection on VSP2 and MYC2 gene expression levels and JA
accumulation in Ws wild-type and nrt2 plants.
Supplementary Figure 8. Analysis of differentially expressed genes in Ws and nrt2 upon infection.
Study of the reduced sensitivity of nrt2 to coronatine by array analysis and direct treatment with the
effector.
Supplementary Table 1. List of primers employed in qRT-PCR analysis.
Supplementary Table 2. List of genes and fold induction analysis present in the Venn diagram.
Supplementary Table3. List of genes that are induced by coronatine and down-regulated in nrt2-
Pst/nrt2-mock vs. Ws-Pst/Ws-mock. qRT-PCR confirmation of several genes present in the array.
ACKNOWLEDGMENTS
We thank the SCIC of the Universitat Jaume I for its technical support. We also thank Brigitte Mauch-
Mani and Juan Antonio López-Ráez for proofreading and critically reviewing the manuscript. We also
thank the funding provided by Generalitat Valenciana GV/2007/099, Plan Promoción Bancaja-UJI
P1.1A2007-07 and P1.1B2007-42.
LITERATURE CITED
Ahmad S, Van Hulten M, Martin J, Pieterse C, Van Wees S, Ton J (2011) Genetic dissection of basal defense responsiveness in accessions of Arabidopsis thaliana. Plant Cell Env. 34:191-1206
Ali FS, Zayed G, Saad OA, Abdul-Mohsen (2009) Optimisation of nitrogen fertiliser level for
maximum colonisation of mycorrhizae on roots of coriander plants. African Crop Science Conference
Proceedings 9:117 - 122
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J,
Barajas P, Cheuk R, Gadrinab C, Heller C, Jeske A, Koesema E, Meyers CC, Parker H,
Prednis L, Ansari Y, ChoyN, Deen H, Geralt M, Hazari N, Hom E, Karnes M, Mulholland
C, Ndubaku R, Schmidt I, Guzman P, Aguilar-Henonin L, Schmid M, Weigel D, Carter
DE, Marchand T, Risseeuw E, Brogden D, Zeko A, Crosby WL, Berry CC, Ecker JR
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

21
(2003) Genome-Wide Insertional Mutagenesis of Arabidopsis thaliana. Science 301: 653-657
Brooks DM, Guzman GH, Kloek AP, Alarcón-Chaidez F, Sreedharan A, Rangaswamy V,
Peñaloza-Vázquez A, Bender CL, Kunkel BN (2004) Identification and characterization of a
well-defined series of coronatine biosynthetic mutants of Pseudomonas syringae pathovar
tomato DC3000. Mol Plant-Microbe Interact. 17:162-174
Cai C, Zhao XQ, Zhu YG, Li B, Tong YP, Li ZS (2007) Regulation of the high-affinity nitrate
trasnport system in wheat roots by exogenous abscisic acid and glutamine. Journal of
Integrative. Plant Biol 49: 1719-1725
Castaings L, Marchive C, Meyer C, Krapp A (2010) Nitrogen signalling in Arabidopsis: how to
obtain insights into a complex signalling network. J Exp Bot 62: 1391-1397
Cerezo M, Tillard P, Filleur S, Munos S, Daniel-Vedele F, Gojon A (2001) Major alterations of the
regulation of root NO3- uptake are associated with the mutation of Nrt2.1 and Nrt2.2 genes in
arabidopsis. Plant Physiol 127: 262-271.
Conrath U (2011) Molecular aspects of defence priming. Trends Plant Sci 16: 524-531
Conrath U, Beckers GJM, Flors V, Garcia-Agustin P, Jakab G, Mauch F, Newman MA, Pieterse
CMJ, Poinssot B, Pozo MJ, Pugin A, Schaffrath U, Ton J, Wendehenne D, Zimmerli L,
Mauch-Mani B and Prime-A-Plant Grp (2006) Priming: Getting ready for battle. Mol Plant-
Microbe Interact 19: 1062-1071.
Edwards K, Johnstone C, Thomson C (1991) A simple and rapid method for the preparation of plant
genomic DNA for PCR analysis. Nucleic Acids research. 19: 1349.
Filleur S, Dorbe MF, Cerezo M, Orsel M, Granier F, Gojon A, Daniel-Vedele F (2001) An
Arabidopsis T-DNA mutant affected in Nrt2 genes is impaired in nitrate uptake. FEBS Lett 489:
220–224.
Fonseca S, Chico JM, Solano R (2009) The jasmonate pathway: the ligand, the receptor and the core
signalling module. Curr Op Plant Biol 12: 539-547.
Gansel X, Munos S, Tillard P, Gojon A (2001) Differential regulation of the NO3- and NH4
+
transporter genes AtNrt2.1 and AtAmt1.1 in Arabidopsis: relation with long-distance and local
controls by N status of the plant. Plant J 26: 143-155
Gojon A, Krouk G, Perrine-Walker F, Laugier E (2011) Nitrate transceptor(s) in plants. J Exp Bot
62:2299-2308
Gutierrez RA, Gifford ML, Poulthey C, Wang R, Shasha DE, Coruzzi GM, Carwford NM (2003)
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

22
Insights into the genomic nitrate response using genetics and the Sungear Sofware system. J
Exp Bot 58: 2359-2357
Ho CH, Lin SH, Hu HC, Tsay YF (2009) CHL1 functions as a nitrate sensor in plants. Cell 138:1184-
1194
Hoffland E, van Beusichem ML, Jeger MJ (1999) Nitrogen availability and susceptibility of tomato
leaves to Botrytis cinerea. Plant and Soil 210: 263–272
Jakab G, Ton J, Flors V, Zimmerli L, Métraux JP, Mauch-Mani B (2005) Enhancing Arabidopsis
Salt and Drought Stress Tolerance by Chemical Priming for Its Abscisic Acid Responses. Plant
Physiol 139:267-274
Jones JDG, Dangl JL (2006) The Plant Immune System. Nature 444: 323-329
Jung HW, Tschaplinski TJ, Wang L, Glazebrook J, Greenberg JT (2009) Priming in Systemic Plant
Immunity. Science 324: 89-91.
Kawamura Y, Hase S, Takenaka S, Kanayama Y, Yoshikawa H, Takahashi H (2009) INF1 elicitin
activates jasmonic acid- and ethylene-mediated signaling pathways and induces resistance to
bacterial wilt disease in Tomato. J Phytopathol 157:287-297
Krouk G, Crawford NM, Coruzzi GM, Tsay YF (2010) Nitrate signaling: adaptation to fluctuating
environments. Curr Opinion Plant Biol 13: 266-273
Lejay L, Tillard P, Lepetit M, Olive FD, Filleur S, Daniel-Vedele F, Gojon A (1999) Molecular and
functional regulation of two NO3- uptake systems by N- and C- status of Arabidopsis plants.
Plant J 18: 509-519
Li W, Wang Y, Okamoto M, Crawford NM, Siddiqi MY, Glass AD (2007) Dissection of the
AtNRT2.1:AtNRT2.2 inducible high-affinity nitrate transporter gene cluster. Plant Physiol 143:
425–433
Little DY, Rao H, Oliva S, Daniel-Vedele F, Krapp A, Malamy JE (2005) The putative high-affinity
nitrate transporter NRT2.1 represses lateral root initiation in response to nutritional cues. Proc
Natl Acad Sci USA 102: 13693–13698
Liu G, Ji Y, Bhuiyan NH, Pilot G, Selvaraj G, Zou J, Wei Y (2010) Amino acid homeostasis
modulates salicylic acid-associated redox status and defense responses in Arabidopsis. Plant
Cell doi:10,1105/tpc.110.079392
Long DH, Lee FN, TeBeest DO (2000) Effect of Nitrogen Fertilization on Disease Progress of Rice
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

23
Blast on Susceptible and Resistant Cultivars. Plant Disease 84: 403-409
Lozano-Juste J, León J (2010) Enhanced abscisic acidmediated responses in nia1nia2noa1-2 triple
mutant impaired in NIA/NR- and AtNOA1-mediated nitric oxide biosíntesis in Arabidopsis.
Plant Physiol. 152:891–903
Melotto M, Underwood W, He SY (2008) Role of stomata in plant innate immunity and foliar
bacterial diseases. Annu Rev Phytopathol 46: 101-122
Modolo LV, Augusto O, Almeida IMG, Magalhaes JR, Salgad I (2005) Nitrite as the major source of
nitric oxide production by Arabidopsis thaliana in response to Pseudomonas syringae. FEBS
Lett 579: 3814-3820
Modolo LV, Augusnt O, Almeida IMG, Pinto-Maglio CAF, Oliveira HC, Seligman K, Salgado I
(2006) Decreased arginine and nitrite levels in nitrate reductase-deficient Arabidopsis thaliana
plants impair nitric oxide synthesis and the hypersensitive response to Pseudomonas syringae.
Plant Sci 171: 34-40
Mohr PG, Cahill DM (2003) Abscisic acid influences the susceptibility of Arabidopsis thaliana to
Pseudomonas syringae pv. tomato and Peronospora parasitica. Func. Plant Biol. 30: 461-469
Okamoto M, Kumar A, Li W, Wang Y, Siddiqi MY, Crawford NM, Glass AD (2006) High-affinity
nitrate transport in roots of Arabidopsis depends on expression of the NAR2-like gene
AtNRT3.1. Plant Physiol 140: 1036-1046
Orsel M, Eulenburg K, Krapp AA, Daniel-Vedele F (2004) Disruption of the nitrate transporter
genes AtNRT2.1 and AtNRT2.2 restricts growth at low external nitrate concentration. Planta
219: 714–721
Pieterse CMJ, Leon-Reyes A, Van der Ent S, Van Wees SCM (2009) Networking by small-molecule
hormones in plant immunity. Nat Chem Biol 5: 308-316
Pinfield-Wells H, Rylott EL, Gilday AD, Graham S, Job K, Larson TR, Graham IA (2005)
Sucrose rescues seedling establishment but not germination of Arabidopsis mutants disrupted in
peroxisomal fatty acid catabolism. Plant J 43: 861-872.
Thordal-Christensen H, Zhang Z, Wei YD, Collinge DB (1997) Subcellular localization of H2O2 in
plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery
mildew interaction. Plant J 11: 1187-1194
Ton J, Flors V, Mauch-Mani B (2009) The multifaceted role of ABA in disease resistance. Trends
Plant Sci 14: 310-317
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

24
Ton J, Jakab G, Toquin V, Flors V, Iavicoli A, Maeder MN, Metraux JP, Mauch-Mani B (2005)
Dissecting the beta-aminobutyric acid-induced priming phenomenon in Arabidopsis. Plant Cell
17: 987-999
Tornero P, JL Dangl JL (2001) A high throughput method for quantifying growth of phytopathogenic
bacteria in Arabidopsis thaliana. Plant J 28: 475-481
Torres-Zabala M, Bennett MH, Truman1WH, Grant MR. (2009) Antagonism between salicylic and
abscisic acid reflects early host–pathogen conflict and moulds plant defence responses. Plant J
59: 375-386
Torres-Zabala, M, Truman W, Bennett MH, Lafforgue G, Mansfield JW, Egea PR, Bogre L,
Grant M (2007) Pseudomonas syringae pv. tomato hijacks the Arabidopsis abscisic acid
signaling pathway to cause disease. EMBO J 26: 1434-1443
Tsai CH, Singh P, Chen CW, Thomas J, Weber J, Mauch-Mani B, Zimmerli L (2011) Priming for
enhanced defence responses by specific inhibition of the Arabidopsis response to coronatine.
Plant J. 65: 469-479
Tsay YF, Chiu CC, Tsai CB, Ho CH, Hsu PK (2007) Nitrate transporters and peptide transporters.
FEBS Lett 581: 2290–2300
van Hulten M, Pelser M, van Loon LC, Pieterse CMJ, Ton J (2006) Costs and benefits of priming
for defense in Arabidopsis. Proc. Nat. Acad. Sci. USA 4: 5602-5607
Yao J, Shi WM, Xu WF (2008) Effects of salt stress on expression of nitrate transporter and
assimilation-related genes in tomato roots. Rus J Plant Physiol 55: 232-240
Ward JL, Forcat S, Beckmann M, Bennett M, Miller SJ, Baker JM, Hawkins ND, Vermeer CP,
Lu C, Lin W, Truman WM, Beale MH, Draper J, Mansfiel JW, Grant M (2010) The
metabolic transition during disease following infection of Arabidopsis thaliana by Pseudomonas
syringae pv. Tomato. Plant J 63:443-457
Wildermuth MC, Dewdnye J, Wu G, Ausube FM (2001) Isochorismate synthase is required to
synthesize salicylic acid for plant defence. Nature 414:562-566
Wu C.C, Sinch P, Chen M.C, Zimmerli L (2009) L-Glutamine inhibits beta-aminobutyric acid-
induced stress resistance and priming in Arabidopsis. J Exp Bot 61:995-1002
Zimmerli L, Jakab C, Metraux JP, Mauch-Mani B (2000) Potentiation of pathogen-specific defense
mechanisms in Arabidopsis by β-aminobutyric acid. Proc Nat Acad Sci USA 97:12920-12925.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

25
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

26
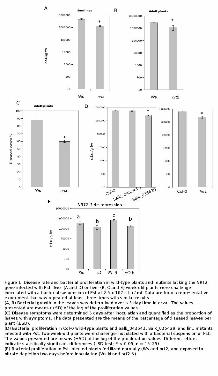
Figure 1. Disease rate and bacterial proliferation in wild-type plants and mutants lacking the NRT2
gene infected with Pst. Two- (A and D) or five- (B, C and E) week-old plants were challenge-
inoculated with a bacterial suspension of Pst at 2.5 x 107 c.f.u./ ml. Data are from a representative
experiment that was repeated at least three times with similar results.
(A, B) Bacterial growth in the leaves was determined over a 3-day time interval. The values presented
are means (±SD) of the log of the proliferation values.
(C) Disease symptoms were determined 3 days after inoculation and quantified as the proportion of
leaves with symptoms. The data presented are the means of the percentage of diseased leaves per plant
(±SD).
(D) Bacterial proliferation in Col-0 wild-type plants and Salk_043543, Salk_035429, and lin1 mutants
infected with Pst. Two-week-old plants were challenge-inoculated with a bacterial suspension of Pst.
The values presented are means (±SD) of the log of the proliferation values. Asterisk indicates
statistically significant differences (LSD test; P < 0.05, n = 15–25).
(E) Bacterial proliferation in Pst-infected plants fertilized normally (Ws and nrt2) and exposed to
nitrate depletion two days before inoculation (Ws-N and nrt2-N).
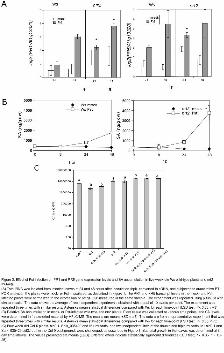
Figure 2. Effect of Pst infection on PR1 and PR5 gene expression levels and SA accumulation in five-
week-old Ws wild-type plants and nrt2 mutants.
(A) Total RNA was isolated from infected leaves at 24 and 48 hours after inoculation (hpi), converted
to cDNA, and subjected to quantitative RT-PCR analysis. The plants were mock- or Pst–inoculated, as
described in Figure 1. The PR1 and PR5 transcript levels in the mock- and Pst-infected plants were
normalized to the expression of At1g13320 measured in the same sample. The experiment was repeated
using β-tubulin with similar results. The data shows the average of two independent experiments
obtained with a pool of 10 plants per point. The experiment was repeated three times with similar
results. Asterisks mean statistical differences compared with Ws for each time-point (LSD test; P<
0.05, n=3)
(B) Relative SA accumulation in mock- or Pst-infected wild-type and nrt2 plants. Plant tissue was
collected at various time points, and SA levels were determined in freeze-dried material by HPLC-MS.
The results are means ±SD (n=5). The data are from a representative experiment that was repeated
three times with similar results. Asterisks mean statistical differences compared with Ws for each time-
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

27
point (LSD test; P< 0.05, n=5)
(C) Four-week-old Col-0 plants, sid2.1, Salk_035429 and lin1 mutants, and two independent lines of
the double and triple mutants lin1-sid2.1 and Salk_035429-sid2.1, all in the Col-0 background, were
challenged, as described in Figure 1. Bacterial growth in the leaves was determined at 3-day time
interval. The values presented are means (±SD). Different letters indicate statistically significant
differences (LSD test; P < 0.05, n = 15–25).
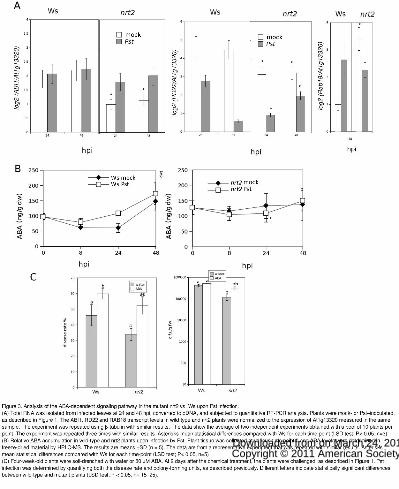
Figure 3. Analysis of the ABA-dependent signaling pathway in the mutant nrt2 vs. Ws upon Pst
infection.
(A) Total RNA was isolated from infected leaves at 24 and 48 hpi, converted to cDNA, and subjected
to quantitative RT-PCR analysis. Plants were mock- or Pst–inoculated, as described in Figure 1. The
ABI1, RD22 and RAB18 transcript levels in wild-type and nrt2 plants were normalized to the
expression of At1g13320 measured in the same sample. The experiment was repeated using β-tubulin
with similar results. The data show the average of two independent experiments obtained with a pool of
10 plants per point. The experiment was repeated three times with similar results. Asterisks mean
statistical differences compared with Ws for each time-point (LSD test; P< 0.05, n=3)
(B) Relative ABA accumulation in wild-type and nrt2 plants upon infection by Pst. Plant tissue was
collected at various time points, and ABA levels were determined in freeze-dried material by HPLC-
MS. The results are means ±SD (n = 5). The data are from a representative experiment that was
repeated with similar results. Asterisks mean statistical differences compared with Ws for each time-
point (LSD test; P< 0.05, n=5)
(C) Four-week-old plants were soil-drenched with water or 80 μM ABA. At 2 days after the chemical
treatment, the plants were challenged, as described in Figure 1. Pst infection was determined by
quantifying both the disease rate and colony-forming units, as described previously. Different letters
indicate statistically significant differences between wild-type and mutant plants (LSD test; P < 0.05,
n= 15–25).
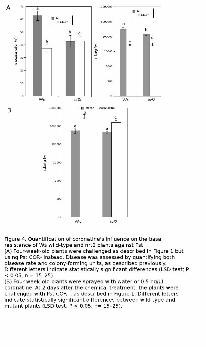
Figure 4. Quantification of coronatine's influence on the basal resistance of Ws wild-type and nrt2
plants against Pst.
(A) Four-week-old plants were challenged as described in Figure 1 but using Pst COR- instead. Disease
was assessed by quantifying both disease rate and colony-forming units, as described previously.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

28
Different letters indicate statistically significant differences (LSD test; P < 0.05, n = 15–25).
(B) Four-week-old plants were sprayed with water or 0.5 ng/μl coronatine. At 2 days after the chemical
treatment, the plants were challenged with Pst COR- , as described in Figure 1. Different letters
indicate statistically significant differences between wild-type and mutant plants (LSD test; P < 0.05,
n= 15–25).
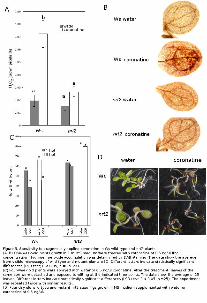
Figure 5. Sensitivity to exogenously applied coronatine in Ws wild-type and nrt2 plants.
(A, B) One-week-old plants grown in 1 ml MS medium were treated with coronatine at 0.5 ng/μl final
concentration. Hydrogen peroxide accumulation was determined by DAB staining. The data show the
average from visible microscopy of wild-type and mutant plants ±SD. Different letters indicate
statistically significant differences (LSD test; P < 0.05, n = 15–25).
(C) Four-week old plants were sprayed with water or 0.5 ng/μl coronatine. After the treatment, leaves
of the same size were detached and exposed to wilting at the indicated time points. The data show the
average of 25 leaves. Different letters indicate statistically significant differences (LSD test; P < 0.05, n
=25). The experiment was repeated twice with similar results.
(D) Four-day-old wild-type and mutant nrt2 seedlings grown in MS medium supplemented with water
or coronatine at 0.5 ng/μl.
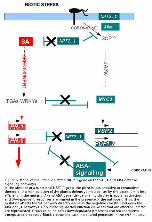
Figure 6. Model of the influence of the NRT2.1 gene on the SA, JA and ABA defense-signaling
pathways.
In the absence of a functional NRT2.1 gene, the plant is less sensitive to coronatine; therefore, the
manipulation of the plant's defensive metabolism by the bacterium is less efficient. In the mutant, JA-
and ABA-dependent signaling display irregular activation, and SA-dependent responses are primed in
the presence of the pathogen. Thus, the mutation affects the SA pathway directly and also the negative
crosstalk between the ABA and JA pathways. Only those signal transduction elements that are affected
in nrt2 are represented. Green color indicates downregulated processes and red represents upregulated
processes. Black crosses indicate the altered processes in the nrt2 mutant.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

29
Supplementary Figure 1.
(A) Total nitrogen levels in 6 week-old plants fertilized normally (Ws and nrt2) and plants subjected to
48 hours of nitrogen deprivation to induce NRT2.1 expression (Ws-N and nrt2-N).
(B, C) The NRT2.1 transcript levels in leaves and roots of Ws and Col-0 plants were normalized to the
expression of At1g13320 measured in the same sample. The experiment was repeated by using β-
tubulin with similar results. The data show the average of two independent experiments obtained with a
pool of 10 plants per point. The experiment was repeated three times with similar results. Asterisks
indicate statistically significant differences (LSD test; P < 0.05, n = 15–25).
Supplementary Figure 2. Effect of Pst infection on PR1 gene expression levels and SA accumulation in
five-week-old Col-0 wild-type plants, Salk_035429 and lin1 mutants.
(A) Total RNA was isolated from infected leaves at 24 and 48 hpi, converted to cDNA, and subjected
to quantitative RT-PCR analysis. Plants were mock- or Pst–inoculated, as described in Figure 1. The
PR1 transcript levels in mock- and Pst-infected plants were normalized to the expression of At1g13320
measured in the same sample. The experiment was repeated using β-tubulin with similar results. The
data show the average of two independent experiments obtained with a pool of 10 plants per point. The
experiment was repeated three times with similar results. Asterisks mean statistical differences
compared with wild-type (LSD test; P< 0.05, n=3)
(B) Relative SA accumulation in mock- or Pst-infected Col-0 wild-type, Salk_035429 and lin1
mutants. Plant tissue was collected at various time points, and SA levels were determined in freeze-
dried material by HPLC-MS. The results are means ±SD (n=5). The data are from a representative
experiment that was repeated three times with similar results. Asterisks mean statistical differences
compared with wild-type for each time-point (LSD test; P< 0.05, n=5)
Supplementary Figure 3. Disease rate and bacterial proliferation in Ws wild-type plants and nrt2
mutants infected with Pst upon BABA treatment and water loss in Ws wild-type and nrt2 plants.
Four week-old plants were soil-drenched with water or 250 μM BABA. After two days of chemical
treatment, the plants were inoculated with a bacterial suspension of Pst at 2.5 x 107 c.f.u./ ml. Pst
infection was determined by quantifying both the disease rate and colony-forming units. Different
letters indicate statistically significant differences (LSD test; P < 0.05, n = 15–25).
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

30
Supplementary Figure 4. Five-week-old plants were subjected to dehydration. The plants were flooded
and left to drain at day 0. Water loss was calculated by measuring the weight of the plants (n=25). The
values are means of water loss (g/plant; ±SD) after different numbers of days of dehydration. The
results shown are derived from a representative experiment that was repeated three times, yielding
comparable results.
Supplementary Figure 5. Bacterial proliferation in Col-0 wild-type plants, lin1 and Salk_035429
mutants infected with Pst or Pst COR-.
Four-week-old plants were challenge-inoculated with a bacterial suspension of Pst at 2.5 x 107 c.f.u./
ml. Disease was determined by quantifying both disease rate and colony-forming units as described
previously. Different letters indicate statistically significant differences (LSD test; P < 0.05, n = 15–25).
Supplementary Figure 6. Bacterial proliferation in Ws wild-type and nrt2 plants infected with Pst.
(A) Five-week-old plants were challenge-inoculated with a bacterial suspension of Pst at 105 c.f.u./ ml
by infiltration with a syringe without a needle. Bacterial growth in the leaves was determined over a 3-
day time interval. The values presented are means (±SD) of the log of the proliferation values.
Different letters indicate statistically significant differences (LSD test; P<0.05, n=15-25).
(B) The picture shows a representative sample of Pst-infected Ws and nrt2 plants three days after
inoculation.
Supplementary Figure 7. Effect of Pst infection on VSP2 and MYC2 gene expression levels and JA
accumulation in Ws wild-type and nrt2 plants.
(A) Total RNA was obtained, as described previously. The VSP2 and MYC2 transcript levels in wild-
type and nrt2 plants were normalized to the expression of At1g13320 measured in the same sample.
The experiment was repeated using β-tubulin with similar results. The data show the average of two
independent experiments obtained with a pool of 10 plants per point. The experiment was repeated
three times with similar results.
(B) Relative JA accumulation in wild-type and nrt2 plants upon infection by Pst. The plants were
inoculated, as described in Figure 1. Plant tissue was collected at various time points, and JA levels
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

31
were determined in freeze-dried material by HPLC-MS. The results are means ±SD (n = 5). The data
are from a representative experiment that was repeated with similar results.
(C) Transcript levels of several representative marker genes in Ws and nrt2 after coronatine treatment.
Plants were treated with either water or coronatine at 0.5 μM, and four hours after the treatment, leaf
material was harvested for analysis. Total RNA was obtained, and CORI1, CORI3, PDF1.2 and RAB18
transcript levels in wild-type and nrt2 plants were analyzed, as described previously. The data show the
average of two independent experiments obtained with a pool of 10 plants per point. The experiment
was repeated two times with similar results. Asterisks mean statistical differences compared with Wt
(LSD test; P< 0.05, n=3).
Supplementary Figure 8. Analysis of genes differentially expressed in wild-type and nrt2 plants after
inoculation with Pst or coronatine treatment.
(A) Venn diagram representing common up- or downregulated genes in wild-type and nrt2 plants upon
infection.
(B) Number of genes that are responsive to coronatine present in the Venn diagram for Ws-Pst
infected/Ws-mock and nrt2-Pst/nrt2-mock.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon March 24, 2018 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.





![A Deletion in NRT2.1 Attenuates Pseudomonas syringae ... · A Deletion inNRT2.1 Attenuates Pseudomonas syringae-Induced Hormonal Perturbation, Resulting in Primed Plant Defenses1[C][W]](https://img.pdfslide.net/doc/110x75/5e012c764c6b0c39e752c5c1/a-deletion-in-nrt21-attenuates-pseudomonas-syringae-a-deletion-innrt21-attenuates.jpg)























