Embed Size (px)
Citation preview

1
The Role of Vegetative Reproduction in the Recruitment and
Survival of Populus fremontii and Salix spp. in a
Californian Riparian System
Mandy Tu
Section of Evolution and Ecology, One Shields Avenue,
University of California, Davis, CA 95616
E-mail: [email protected]; Phone 530-752-1092; FAX 530-752-1449
Current Address:
The Nature Conservancy’s Invasive Species Initiative
821 SE 14th Avenue Portland, Oregon 97214
[email protected], 530-802-8100

2
ABSTRACT
The importance of vegetative reproduction of woody species in the family
Salicaceae has been assumed for a long time. Most studies of reproduction in
this family however, have concentrated on seed dispersal and seedling survival,
and scientific studies on the success of vegetative propagules are largely lacking.
The initial recruitment and establishment of riparian woody vegetation on a
newly-formed sandbar along the lower Cosumnes River, located in California’s
Central Valley, was followed from 1996 to 1998. The establishment and survival
of woody species populations were monitored using permanent belt-transects.
Cottonwoods (Populus fremontii ssp. fremontii) and willows (Salix exigua, S.
gooddingii, S. laevigata, S. lasiolepis) were the only woody colonizers. Seed
viability and dispersal were not limiting factors in their regeneration.
Nevertheless, the successful recruitment of P. fremontii from broken-off branches
is six times more important than recruitment from seeds at this site. Salix spp.
reproduced exclusively vegetatively from deposited branches.
Keywords riparian succession, Populus fremontii, Salix ssp., seedling
establishment, vegetative propagation

3
INTRODUCTION
Vegetation succession following regular disturbance events such as large
floods has received much attention within the last 30 years (Bliss and Cantlon
1957, Leopold et al. 1964, Nanson and Beach 1977, Malanson 1993, Naiman and
Décamps 1997). In riparian systems, fluvial geomorphic processes have been
documented to continually create, change, and destroy environments suitable for
the establishment of riparian plant species (McLeod and McPherson 1973, Fonda
1974, Sands 1977, Frye and Quinn 1979, Warner and Hendrix 1984, Asplund and
Gooch 1988, Auble and Scott 1998). On sandbars, as well as on gravel bars and
point bars, the processes of sedimentation and erosion may continually alter the
colonization substrate (McBride and Strahan 1984, Taylor et al. 1999). The degree
of water drawdown may also influence the survival of established plants (Fenner
et al. 1984, 1985, Stromberg and Patten 1991, Rood et al. 1994, 1998). Most studies
of vegetation succession along rivers have looked at natural rivers or stream
systems (Frye and Quinn 1979, Fyles and Bell 1986, Dunham 1989). In controlled
river systems, several studies have concluded that certain rates of water flow are
necessary for the successful establishment of native riparian species (Fenner et al.
1985, Stromberg and Patten 1991, 1996, Segelquist et al. 1993, Braatne et al. 1996,
Begg et al. 1998, Mahoney and Rood 1998, Rood et al. 1994, 1998).
In California’s Central Valley, riparian forests once extended up to three
miles wide along each river in pre-European settlement times (Thompson 1961).
Since the early 1900’s however, California rivers and streams have been

4
especially highly impacted by anthropoegenic forces such as agriculture,
damming, and urban development (Mount 1995). Hunter et al. (1999) deduced
that only 3.3% of once-existing riparian areas remain in the Central Valley, and
are mostly much degraded. Since the Cosumnes River remains as the only
undammed river in the Central Valley and still retains yearly flooding regimes, it
provides a unique opportunity to study the early (pioneering) stages of plant
succession following disturbance. Previous studies have examined plant
establishment on sandbars in the southwestern United States (Shafroth et al.
1994, 1998, Taylor et al. 1999) and in other parts of California (McBride & Strahan
1984, Shanfield 1984, Strahan 1984, McCarten 1989, Stromberg et al. 1991), but
none have studied these events along an uncontrolled river in California’s
Central Valley. By understanding some aspects of the reproductive ecology of
woody pioneering species, recommendations for the restoration of these highly
degraded environments, by simulating natural processes, can be implemented.
The goal of this paper is to describe the early recruitment of woody tree
species on a sandbar at the Cosumnes River Preserve, where the most pristine
patches of riparian forests remain in California’s Central Valley (Reiner 1996,
Wicinas 1998). This paper addresses the early colonization stage of plant
establishment and survival on a newly deposited substrate. I hypothesized that
the primary means of recruitment of woody trees would be from seeds, as the
seed production by the studied species is prolific and most of the published
literature from the western and southwestern United States have focused on the

5
successful establishment of woody seedlings (McBride & Strahan 1984, Strahan
1984, Shafroth et al. 1994, 1998, Stromberg et al. 1991). At the same time, records
on vegetative reproduction are mostly anecdotal. Only very few studies have
determined the relative importance of sexual versus asexual propagules of
Salicaceae in native stands (Braatne et al. 1996). The present study quantifies the
magnitude of these two modes of Salicaceae reproduction.
METHODS
Study Site
The Cosumnes River Preserve is a Nature Conservancy preserve located
at 38˚ 15'N 121˚ 23'W, approximately 20 miles south of the city of Sacramento in
California’s Central Valley. The Cosumnes River is the only river in California's
Central Valley that has been left undammed (Mount 1995, Reiner 1996) and still
experiences seasonal flooding. This flooding may last between one to six months
in duration (Reiner, pers. comm.). The vegetation in this area is structured by
both stand age as well as by elevation in relation to the river floodplain (Tu,
unpubl. data, Conard et al. 1977). Dominant trees in this area are P. fremontii in
the lower wetter and younger areas, Fraxinus latifolia, Acer negundo and Salix
ssp. in the mid-layers, to mature Quercus lobata trees in the more upland and
older sites.
In the fall of 1995 a levee was intentionally broken by The Nature
Conservancy just south of a 15-year-old cottonwood forest (hereafter referred to

6
as the Accidental Forest). In the ensuing winter floods, a new sandbar
established perpendicular to where the levee was breached, on an old abandoned
farm-field.
Study Species
Populus fremontii S. Watson ssp. fremontii – This common tree of
streamsides and alluvial bottomlands is widespread from California throughout
the West to the Rocky Mountains (Hickman 1993). P. fremontii was prominent in
the pristine riparian forests of the Sacramento Valley (Thompson 1961), and is
still abundant in wet areas throughout California’s Central Valley (Griffin and
Critchfield 1972). This species has seedlings that are easily recognized by their
two almost cordate-shaped cotyledons. P. fremontii is known to recruit easily
from both seeds (Fenner et al. 1984) and from branches (Pope et al. 1990). For the
duration of the study, it could easily be determined if new individuals that
arrived on the sandbar were derived from either branches (by their initial large
size) or from seeds.
Salix exigua Nutt., Salix gooddingii C. Ball, Salix laevigata Bebb, Salix
lasiolepis Benth. – These dioecious trees and shrubs of moist habitats are also
widespread throughout California and the West in lowland areas (Hickman
1993). For the purposes of this study, all species of this genus were combined for
ease of analysis. Salix species have been documented to recruit via both seeds
(Niiyama 1990, Shafroth et al. 1994) and by clonal growth (Douglas 1987).

7
Seedling Recruitment and Survival
On the newly-created north-south oriented sandbar, five east-west
running belt-transects which run perpendicular to the water line (parallel to the
river) were established during the early summer of 1996. The five belt-transects
varied in length from 8 to 12 meters, and were 0.5 meters wide. These transects
were monitored for seed germination and survival of all woody tree species four
times from 1996-1998. A total area of 23 square meters was sampled for
seedlings.
Vegetative Branch Recruitment and Survival
In addition to these transects, within the boundaries of the entire transect
area (1,000 m2), vegetative growth from pieces of broken-off branches of both P.
fremontii and Salix spp. were tagged, mapped and monitored for survival from
1996 to 1998. These sprouting branches were measured four times between 1996
to 1998. Stem diameter was measured at the soil surface. Height was measured
to the apical meristem. Survival of seedlings and branches was calculated from
both the first and second cohort years (1996 and 1997).
RESULTS
Seedling Recruitment and Survival
The germination of woody species growing on this sandbar began in late
May of 1996. Seeds of P. fremontii have a short viability time of 2-3 weeks (Tu,
unpub. data; Fenner et al. 1984) in which there is almost complete viability and

8
the tendency to germinate immediately after dispersal (McBride and Strahan
1984). Seeds of P. fremontii were dispersed over the entire surface of the
sandbar, but were more concentrated in the moist zones either at the edge of the
outflow or in small depressions which were scattered across the sandbar surface.
In all three years of this study, no seedlings of any Salix spp. were found.
In the first field-season (1996) after the levee breach, I mapped and
followed the fates of 7,898 P. fremontii seedlings that had germinated within the
five belt-transects. By the end of the first field-season, all but 37 of the seedlings
had died. There was no significant correlation of surviving seedlings with
relative elevation on the sandbar (Tu, unpub. data). In the following year only 7
seedlings of the original seedling cohort remained, and by the summer of 1998,
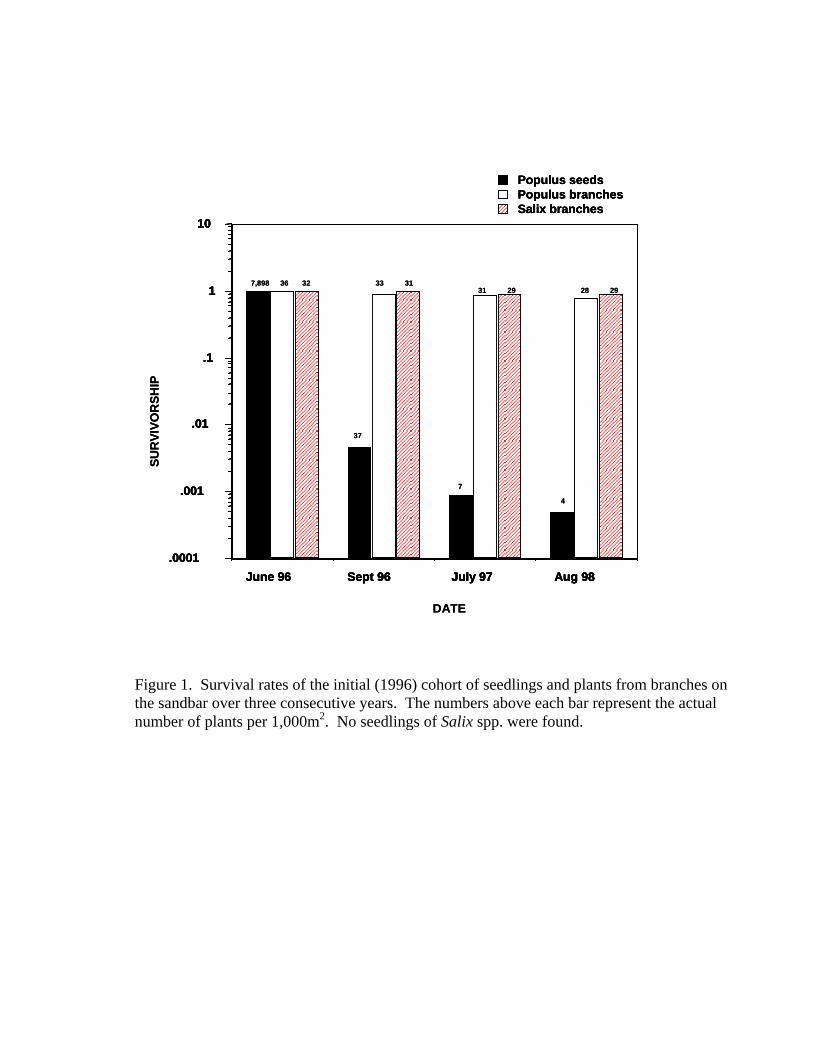
only 4 seedlings had survived. Figure 1 shows the survival of the first cohort of
P. fremontii seedlings, the actual numbers of germinated seedlings from the first
initial year of establishment, and the total numbers of survivors over the first
three years.
Branch Recruitment and Survival
In contrast with the low numbers of survivors of P. fremontii seedlings
and complete absence of Salix spp. seedlings, there were many surviving plants
from re-sprouting branches of both P. fremontii and Salix spp. which had been
washed down from the river within the boundaries of the studied area (Figure 1).
These branches lay either on top of, or embedded up to 0.3 meters deep (pers.
obs.) on the deposited sandbar. The proportions of survivors from the initial

9
year of colonization from seeds is extremely low (Table 1). For P. fremontii, only
0.004 seed/m2, or one surviving seedling per every 250 m2, survived. In
comparison, the number of surviving branches of P. fremontii was much higher,
at 0.025 branches/m2, or one surviving branch per 40 m2. No seedlings of Salix
were found during the three years of this study, but there were relatively high
numbers of Salix from broken-off branches (0.032/m2, or one surviving
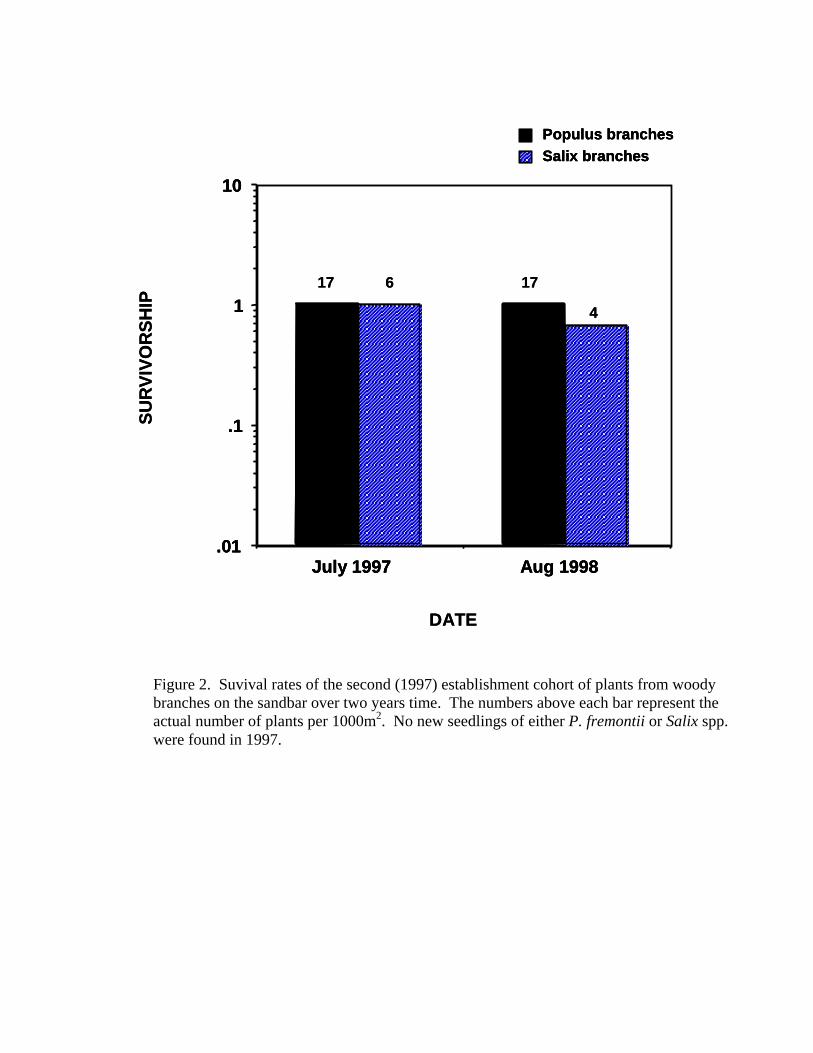
branch/31.25 m2) surviving over three years. The second establishment cohort
year (1997) similarly concluded that vegetative branches had high rates of
survivorship (Figure 2).
Growth Rates of Seeds versus Branches
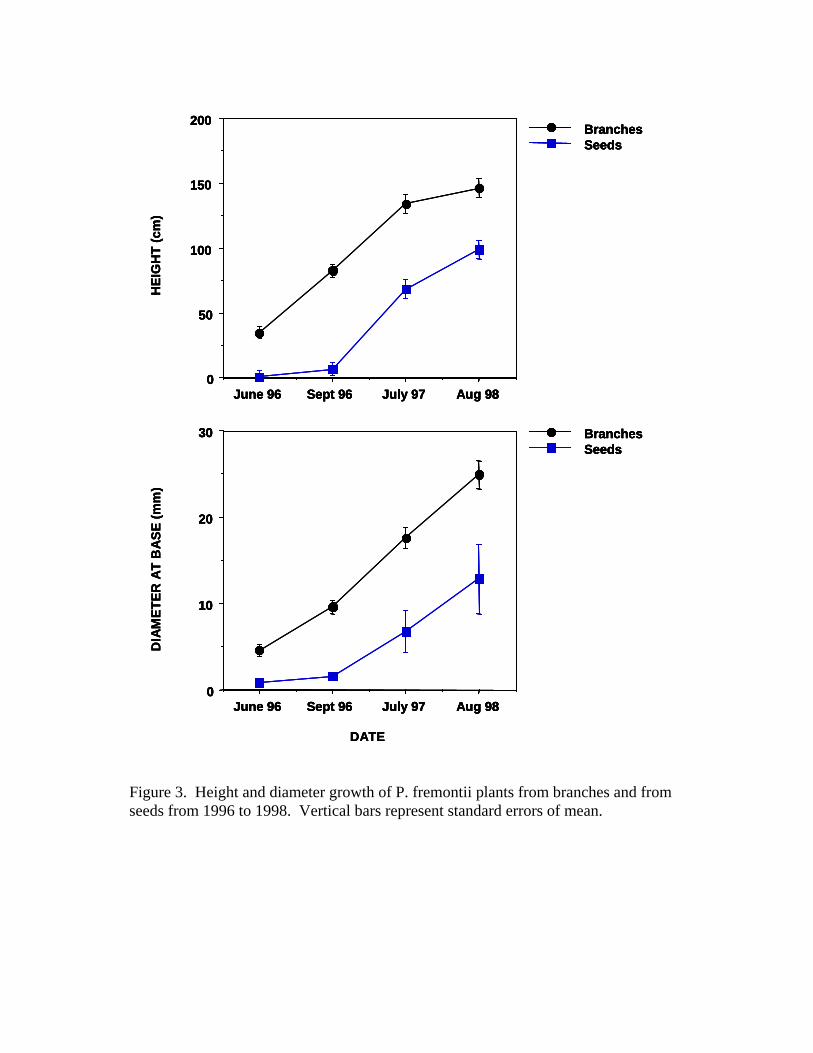
Throughout the duration of this research, it could easily be determined if
the origin of P. fremontii plants were from seeds or from branches. Both the
height and the diameter of plants derived from branches were significantly
larger than those derived from seeds (Figure 3). In the initial year of plant
establishment, all seedlings were observed to have been derived directly from
seeds. It was similarly easy to determine which plants came from branches,
because most plants from branches were quite large (over 0.3 meters tall, unpub.
data) at the initial colonization period. By the third year of observation, all new
plants on the sandbar are assumed to have been derived from branches, as all
new inputs were sizable (over 0.3 meters tall, unpub. data). No new seeds or
seedlings were observed in any of the following years of the study.

10
DISCUSSION
The natural regeneration of native woody riparian species on this sandbar
was not wholly unexpected. In fact, this levee breach was instigated in hopes of
“naturally creating” a young forest dominated by P. fremontii. Historical photos
show that the Accidental Forest, the nearby 15-year-old stand of trees dominated
by P. fremontii, initially established following an accidental levee breach during
the high floods of 1983. What was unexpected however, was the rejection of our
hypothesis that the primary means of recruitment of woody tree seedlings would
be from seeds. On this sandbar, millions of seeds of P. fremontii had washed-up
along the edges and into small depressions on the sandbar. The exact locations
of approximately 8,000 P. fremontii seedlings were recorded and mapped. After
the long, hot summer of 1996, the majority of seedlings had died, apparently
because of desiccation. This was evident as the remnants of many young
seedlings remained as dried little pieces of plant material. By the end of that first
year, only 37 seedlings remained. After three years, only 4 of the initial 8,000 P.
fremontii seedlings had survived. McBride and Strahan (1984), Mahoney and
Rood (1998) and Rood (1998) have all noted that the survival of P. fremontii is
highly dependent upon sufficient soil moisture during the early seed
germination and seedling establishment stages. McLeod and McPherson (1973)
have also documented this dependency for Salix. If the roots of P. fremontii
seedlings are not sufficiently long enough at the time of water drawdown, or if
there is not enough sufficient soil moisture at the time of seed dispersal and

11
germination, all seedlings will die. At this site, grazing by herbivores did not
appear to have an affect on the recruitment and survival of woody species (pers.
obs.). The leading cause of mortality appears to have been lack of moisture as
most dead seedlings remained intact and could be accounted for in the following
years.
In contrast to the low survival of seedlings, there were much greater rates
of survival for plants resprouting from branches of P. fremontii and Salix spp.
This is a phenomenon that has been eluded to by many botanists and restoration
ecologists in riparian systems (Henderson 1991, Shafroth et al. 1994, McBride and
Strahan 1984). Frequently in the the riparian literature (Malanson 1993), mention
is made of vegetative reproduction playing some part in vegetation succession
(Douglas 1989, Shafroth et al. 1994). Rood et al. (1994) even excavated several
hundreds of small cottonwoods to determine their method of origin. They found
that clonal reproduction was able to account for approximately half of the
regeneration efforts in several cottonwood forests in Alberta. Fay et al. (1999)
concluded similarly that Populus euphratica in Spain was exclusively clonal.
Even with this knowledge however, sexual reproduction is still commonly cited
as the primary mode of natural regeneration in Salicaceae (Bradley et al. 1986,
Niiyama 1990, Stromberg 1996), and studies with hard evidence of vegetative
reproduction (rates of survival, rates of growth) remains lacking. Studies of
aspens (Populus trichocarpa, P. grandidentata) however, are the exception in that

12
it has been shown conclusively that vegetative reproduction is common (Barnes
1966, Braatne et al. 1996).
In this study, it was easy to determine whether propagules were of sexual
or of vegetative origin (Figure 3). We emphasize that the primary mode of
woody regeneration, at least in this Mediterranean climate, is from vegetative
propagation, not from seeds. In comparing the total numbers of survivors of
both branches and seeds on this sandbar, to those of the neighboring Accidental
Forest and to a nearby 35-year-old mixed-riparian forest, there are a few
noticeable differences. In regards to Salix, there were slightly higher numbers of
Salix individuals in the Accidental Forest (Table 1). However, this higher
number may be a result of additional root or stem suckers. There were no
individuals of Salix found in any of the 35-year-old mixed-riparian forested sites.
Nor were there any seedlings of Salix found on the sandbar. Hence it appears
that Salix reproduces almost solely by vegetative means.
P. fremontii also appears to reproduce primarily by vegetative means.
Even though the number of survivors after three years from branches outnumber
those which survive by seed almost 6 to 1, recruitment by seed has been shown
to be a valuable source of propagules in most other studies in the riparian
literature (McBride and Strahan 1984, Stromberg and Patten 1991, Taylor et al.
1999) and should not be discounted completely. However, this study shows that
broken-off branches were the primary source of propagules for recruitment. In
comparing the total numbers of branches and seeds from this sandbar to the

13
Accidental Forest, there are almost three times as many young trees of P.
fremontii per square meter in the Accidental Forest as compared to the 3-year-
old sandbar. We speculate that this may be partially explained by the much
higher flood peak flows of 1983 (801.8 m3 in January through June) versus the
floods of 1996 (352.9 m3 in January through June; data from CA Dept. of Water
Resources). The higher water flows may have washed in more branches, and
may have retained a higher water table for a longer time, which would have
benefited the survival of P. fremontii seedlings. Another aspect of P. fremontii
regeneration to be considered is their natural suckering ability or the ability to
put-out multiple stems (Rood et al. 1994, Fay et al. 1999). It is not known how
many genetic individuals comprise the Accidental Forest. However, it is known
that the branches from the 3-year-old sandbar were each washed-down
individually. How related those branches are to one another is not known.
In the Accidental Forest, there are no indications of the P. fremontii trees
dying of old age, windthrow, or other natural disturbances. There is however,
evidence of disturbance and herbivory by beaver (unpub. data). In the 35-year-
old forest, the large trees of P. fremontii are starting to die, presumably from old
age and/or windthrow. Thus explaining the low total numbers of P. fremontii
individuals in the 35-year-old forest.
Additionally, new inputs from branches may occur following the initial
creation of the sandbar habitat. In the second year of monitoring, 17 new
branches of P. fremontii and 6 new branches of Salix appeared on the sandbar.

14
Furthermore, there was very low mortality of these additional branches. P.
fremontii had complete survival of the second cohort of branches and Salix had
60% survival of the branches (4 of 6 survived). No new seedlings of either genus
were found after the initial year of establishment. This may be due to increased
competition for light and nutrients from other species.
Being able to reproduce vegetatively may be a selective advantage for
both Populus and Salix spp. Both genera produce copious amounts of small,
wind-dispersed, short-lived seeds. If these seeds do not land in a safe-site (sensu
Harper) and have adequate moisture at the seedling stage, there is a very narrow
window of opportunity for successful establishment. By reproducing
vegetatively from branches, there is an increased chance of success because of
increased amounts of stored nutrients, increased competitive ability for light (by
being taller), and perhaps increased water availability (by being buried deep in
the soil).
The recruitment of woody tree species in riparian systems is not well
understood. By rejecting our initial hypothesis that the primary means of
recruitment of woody trees would be from seeds, shows us that there are many
complexities involved in tree recruitment. Much emphasis in the literature has
focused on woody seedling establishment, and how conditions for establishment
may be altered (Stromberg and Patten 1991, Mahoney and Rood 1998). By
acknowledging that natural regeneration of some species may occur primarily by
vegetative propagules, opens the door for many restoration possibilities.

15

16
ACKNOWLEDGEMENTS
Many thanks to Marcel Rejmánek for his unending insightfulness and
guidance during this research. Many additional thanks to E. Grotkopp, S.
Brewer, and R. Reiner for comments, and the staff at the Cosumnes River
Preserve for logistical assistance. Cheerful field assistance was provided by R.
Nelson, R. Mølholm and L. Lopez. This work was supported by funds from The
Nature Conservancy, The Nature Conservancy’s Rodney Johnson/Katherine
Ordway Student Grant Fellowship Fund, The California Native Plant Society,
and The Davis Herbaria Society.

17
REFERENCES Asplund, K. K. and M. T. Gooch. 1988. Geomorphology and the
distributional ecology of Fremont Cottonwood (Populus fremontii) in a
desert riparian canyon. Desert Plants 9: 17-27.
Auble, G. T. and M. L. Scott. 1998. Fluvial disturbance patches and
cottonwood recruitment along the upper Missouri River, Montana.
Wetlands 18: 546-556.
Barnes, B. V. 1966. The clonal growth habit of american aspens. Ecology 47:
439-447.
Begg, C. S., W. Archibold, and L. Delanoy. 1998. Preliminary investigation
into the effects of water-level control on seedling recruitment in riparian
cottonwoods, Populus deltoides, on the South Saskatchewan River.
Canadian Field Naturalist 112: 684-693.
Bliss, L. C. and J. E. Cantlon. 1957. Succession on river alluvium in northern
Alaska. American Midland Naturalist 58: 452-69.
Bradley, C. E. and D. G. Smith. 1986. Plains cottonwood recruitment and
survival on a prairie meandering river floodplain, Milk River, southern
Alberta and northern Montana. Canadian Journal of Botany 64: 1433-1442.
Braatne, J. H., S. B. Rood, and P. E. Heilman. 1996. Life history, ecology, and
conservation of riparian cottonwoods in North America. In R. F.
Stettler, H. D. Bradshaw, Jr., P. E. Heilman, and T. M. Hinckley (eds.)

18
Biology of Populus and its implications for management and
conservation. Part I, Chapter 3. NRC Research Press, National
Research Council of Canada, Ottawa, ON, Canada.
Conard, S. G., R. L. MacDonald, and R. F. Holland. 1977. Riparian vegetation
and flora of the Sacramento Valley. P. 47-55. In A. Sands (ed.) Riparian
Forests in California. University of California, Davis, CA, USA.
Douglas, D. A. 1989. Clonal growth of Salix setchelliana on glacial river
gravel bars in Alaska. Journal of Ecology 77: 112-26.
Dunham, K. M. 1989. Vegetation-environment relations of a Middle
Zambezi floodplain. Vegetatio 82: 13-24.
Fay, M. F., M. D. Lledó, M. M. Kornblum, and M.B. Crespo. 1999. From the
waters of Babylon? Populus euphratica in Spain is clonal and probably
introduced. Biodiversity and Conservation 8: 769-778.
Fenner, P., W. W. Brady, and D. R. Patton. 1984. Observations on seeds and
seedlings of Fremont Cottonwood. Desert Plants 6: 55-58.
Fenner, P., W. W. Brady, and D. R. Patton. 1985. Effects of regulated water
flows on regeneration of Fremont Cottonwood. Journal of Range
Management 38: 135-138.
Fonda, R. W. 1974. Forest succession in relation to river terrace development
in Olympic National Park, WA. Ecology 55: 927-942.
Friedman, J. M., M. L. Scott, and W. M. Lewis. 1995. Restoration of riparian

19
forest using irrigation, artificial disturbance, and natural seedfall.
Environmental Management 19: 547-557.
Frye, R. J. and J. A. Quinn. 1979. Forest development in relation to
topography and soils in a flood-plain of the Raritan River, New Jersey.
Bulletin of the Torrey Botanical Club 106: 334-45.
Fyles, J. W. and M. A. M. Bell. 1986. Vegetation colonizing river gravel bars
in the Rocky Mountains of southeastern British Columbia. Northwest
Science 60: 8-14.
Griffin, J. R. and W. B. Critchfield. 1972. The distribution of forest trees in
California. U.S.D.A. Forest Service Research Paper PSW-82.
Henderson, L. 1991. Alien invasive Salix spp. (willows) in the grassland
biome of South Africa. South African Forestry Journal 157: 91-95.
Hickman, J. C. (ed.) 1993. The Jepson Manual. University of California,
Berkeley, CA, USA.
Hunter, J. C., K. B. Willett, M. C. McCoy, J. F. Quinn, and K. E. Keller. 1999.
Prospects for preservation and restoration of riparian forests in the
Sacramento Valley, California, USA. Environmental Management 24:
65-75.
Leopold, L. B., M. G. Wolman, and J. P. Miller. 1964. Fluvial Processes in
Geomorphology. W.H. Freeman, San Francisco, CA, USA.
Mahoney, J. M. and S. B. Rood. 1998. Streamflow, requirements for
cottonwood seedling recruitment - An interactive model. Wetlands 18:

20
634-645.
Malanson, G. P. 1993. Riparian Landscapes. Cambridge University Press,
Cambridge, MA, USA.
McBride, J. R. and J. Strahan. 1984. Establishment and survival of woody
riparian species on gravel bars of an intermittent stream. American
Midland Naturalist 112: 235-245.
McCarten, N. F. 1989. Plant community development, site quality analysis
and river dynamics in the design of riparian preserves on the middle
Sacramento River, California. In D. L. Abell (ed.) Proceedings of the
California Riparian Systems Conference: Protection, management, and
restoration for the 1990s; 1988 Sept 22-24; Davis, CA. General Technical
Report PSW-110. Berkeley, CA, USA.
McLeod, K. W. and J. K. McPherson. 1973. Factors limiting the distribution of
Salix nigra. Bulletin of the Torrey Botanical Club 100: 102-110.
Mount, J. F. 1995. California rivers and streams, the conflict between fluvial
process and land use. University of California Press, Berkeley, CA,
USA.
Naiman, R. J. and H. Décamps. 1997. The ecology of interfaces: Riparian
zones. Annual Review of Ecology and Systematics 28: 621-658.
Nanson, G. C. and H. F. Beach. 1977. Forest succession and sedimentation on
a meandering-river floodplain, northeast British Columbia, Canadian
Journal of Biogeography 4: 229-251.

21
Niiyama, K. 1990. The role of seed dispersal and seedling traits in
colonization and coexistence of Salix species in a seasonally flooded
habitat. Ecological Research 5: 317-331.
Pope, D. P., J. H. Brock, and R. A. Backhaus. 1990. Vegetative propagation of
key southwestern woody riparian species. Desert Plants 10: 91-95.
Reiner, R. 1996. The Cosumnes River Preserve: 1987-95 fertile ground for
new conservation ideas. Fremontia 24: 16-19.
Rood, S. B., C. Hillman, T. Sanche, and J. M. Mahoney. 1994. Clonal
reproduction of riparian cottonwoods in southern Alberta. Canadian
Journal of Botany 72: 1766-1774.
Rood, S. B., A. R. Kalischuk, and J. M. Mahoney. 1998. Initial cottonwood
seedling recruitment following the flood of the century of the Oldman
River, Alberta. Wetlands 18: 557-570.
Sands, A. (ed.) 1977. Riparian forests in California: their ecology and
conservation. University of California Institute of Ecology, Publication
No. 15, Davis, CA, USA.
Segelquist, C. A., M. L. Scott, and G. T. Auble. 1993. Establishment of Populus
deltoides under simulated alluvial groundwater declines. American
Midland Naturalist 130: 274-285.
Seigel, R. S. and J. H. Brock. 1990. Germination requirements of key
southwestern woody riparian species. Desert Plants 10: 3-8.
Shanfield, A. N. 1984. Alder, cottonwood, and sycamore distribution and

22
regeneration along the Nacimiento River, California. In R. E. Warner
and K. M. Hendrix (eds.) California riparian systems: ecology,
conservation, and productive management. University of California
Press, Berkeley, CA, USA.
Shafroth, P. B., G. T. Auble, J. C. Stromberg, and D. T. Patten. 1998.
Establishment of woody riparian vegetation in relation to annual
patterns of streamflow, Bill Williams River, Arizona. Wetlands 18:
577-590.
Shafroth, P. B., M. L. Scott, J. M. Friedman, and R. D. Laven. 1994.
Establishment, sex structure and breeding system of an exotic riparian
willow, Salix x rubens. American Midland Naturalist 132: 159-172.
Strahan, J. 1984. Regeneration of riparian forests of the Central Valley. In R.
E. Warner and K. M. Hendrix (eds.) California riparian systems:
ecology, conservation, and productive management. University of
California Press, Berkeley, CA, USA.
Stromberg, J. C. and D. T. Patten. 1996. Instream flow and cottonwood growth
in the eastern Sierra Nevada of California, USA. Regulated Rivers 12:
1-12.
Stromberg, J. C. and D. T. Patten. 1991. Instream flow requirements for
cottonwoods at Bishop Creek, Inyo County, CA. Rivers 2: 1-11.
Stromberg, J. C., D. T. Patten, and B. D. Richter. 1991. Flood flows and
dynamics of Sonoran riparian forests. Rivers 2: 221-235.

23
Taylor, J. P., D. B. Wester, and L. M. Smith. 1999. Soil disturbance, flood
management, and riparian woody plant establishment in the Rio
Grande floodplain. Wetlands 19: 372-382.
Thompson, K. 1961. Riparian forests of the Sacramento Valley, California.
Annals of the Association of American Geographers 51: 294-315.
Warner, R. E. and K. M. Hendrix (eds.) 1984. California riparian systems:
ecology, conservation, and productive management. University of
California Press, Berkeley, CA, USA.
Wicinas, D. 1998. A mellifluous roar. Nature Conservancy 48: 20-26.

24
Table 1. The total number of surviving individuals per square meter from the
initial recruitment cohort, as compared to two other different-aged stands.
Sampled stands P. fremontii Salix spp.* 3-year-old sandbar
Seeds 0.004 0.000
Branches 0.025 0.032 15-year-old Accidental Forest 0.090 0.048 (2,700 m2) 35-year-old forest 0.009 0.000 (1,800 m2) *Includes Salix exigua, S. gooddingii, S. lasiolepis and S. laevigata.

25
Figure 1. Survival rates of the initial (1996) cohort of seedlings and plants from
branches on the sandbar over three years time. The numbers above each bar
represent the actual number of plants per 1,000 m2. No seedlings of Salix spp.
were found.
Figure 2. Survival rates of the second (1997) establishment cohort of plants from
woody branches on the sandbar over two years time. The numbers above each
bar represent the actual number of plants per 1,000 m2. No new seedlings of
either P. fremontii or Salix spp. were found in 1997.
Figure 3. Height and diameter growth of P. fremontii plants from branches and
from seeds from 1996 to 1998. Vertical bars represent standard errors of mean.
June 96 Sept 96 July 97 Aug 98.0001
.001
.01
.1
1
10
Populus seedsPopulus branchesSalix branches
DATE
SUR
VIVO
RSH
IP
7,898 36 32
37
33 31
7
31 29
4
28 29
June 96 Sept 96 July 97 Aug 98.0001
.001
.01
.1
1
10
June 96 Sept 96 July 97 Aug 98June 96 Sept 96 July 97 Aug 98.0001
.001
.01
.1
1
10
.0001
.001
.01
.1
1
10
Populus seedsPopulus branchesSalix branches
Populus seedsPopulus branchesSalix branches
DATE
SUR
VIVO
RSH
IP
7,898 36 32
37
33 31
7
31 29
4
28 29
Figure 1. Survival rates of the initial (1996) cohort of seedlings and plants from branches on the sandbar over three consecutive years. The numbers above each bar represent the actual number of plants per 1,000m2. No seedlings of Salix spp. were found.
.
July 1997 Aug 1998.01
.1
1
10
Populus branchesSalix branches
DATE
SUR
VIVO
RSH
IP
17 6
4
17
.
July 1997 Aug 1998July 1997 Aug 1998.01
.1
1
10
.01
.1
1
10
Populus branchesSalix branchesPopulus branchesSalix branches
DATE
SUR
VIVO
RSH
IP
17 6
4
17
Figure 2. Suvival rates of the second (1997) establishment cohort of plants from woody branches on the sandbar over two years time. The numbers above each bar represent the actual number of plants per 1000m2. No new seedlings of either P. fremontii or Salix spp. were found in 1997.
June 96 Sept 96 July 97 Aug 980
10
20
30 BranchesSeeds
DATE
DIA
MET
ER A
T B
ASE
(mm
)
June 96 Sept 96 July 97 Aug 980
50
100
150
200 BranchesSeeds
HEI
GH
T (c
m)
June 96 Sept 96 July 97 Aug 980
10
20
30 BranchesSeeds
DATE
DIA
MET
ER A
T B
ASE
(mm
)
June 96 Sept 96 July 97 Aug 98June 96 Sept 96 July 97 Aug 980
10
20
30
0
10
20
30 BranchesSeedsBranchesSeeds
DATE
DIA
MET
ER A
T B
ASE
(mm
)
June 96 Sept 96 July 97 Aug 980
50
100
150
200 BranchesSeeds
HEI
GH
T (c
m)
June 96 Sept 96 July 97 Aug 98June 96 Sept 96 July 97 Aug 980
50
100
150
200
0
50
100
150
200 BranchesSeedsBranchesSeeds
HEI
GH
T (c
m)
Figure 3. Height and diameter growth of P. fremontii plants from branches and from seeds from 1996 to 1998. Vertical bars represent standard errors of mean.





























