Embed Size (px)
Citation preview

Aus dem lnstitut fur Tierpbysiologie und Tierernahrung der Universitat Gottingen
* 4C-Ausscheidung und -Retention nach Applikation von I4C-Adenin an Ratten unter besonderer Beriicksichtigung der I4CO2- Freis etzung als Parameter s to ffwechs elp hysiologis c her Vorgange
Von H. A. GREIFE und S. MOLNAR
Eingang des Ms. 15. 9. 1981
Einleitung
Anhand der Veratrnung von "CO, aus 14C-rnarkierten Substanzen kann ihre Verstoffwechse- lung kontinuierlich uber einen langeren Zeitraum verfolgt werden. Als Beispiele seien Unter- suchungen an Ratten nach Applikation von 14C-Arninosauren (BROOKES et al. 1972; BERGNER und SIMON 1976), -Fettsauren (EYO 1975; MOLNAR et al. 1977) und -Nukleinsauren-Derivaten (GREIFE und MOLNAR 1978 a,b; 1979) genannt.
Nur bei bekannten und quantitiv kalkulierbaren Stoffwechselwegen sowohl der '"C-Tracer als auch ihrer rnarkierten Metaboliten erlaubt die 14C0,-Veratrnung allein Ruckschlusse auf die jeweilige Stoffwechselsituation. Diese Bedingungen liegen fur die Purinbase Adenin vor.
Am Beispiel der Ratte sollte in dieser Arbeit die Bedeutung der Markierungsart -spezifisch 8-14C- oder uniform '"C-rnarkiert- auf die 14C0,-Freisetzung aus Adenin und die hieraus abzuleitenden Stoffwechselvorgange dernonstriert werden. Die zusatzliche Erfassung des Radiokohlenstoffs in den Exkrernenten und im Tierkorper sollte eine '"C-Bilanzierung errnog- lichen.
2 Material und Methoden
2.1 Versuchstiere und Tracerverabreichung
Als Versuchstiere dienten mlnnliche Albinoratten des institutseigenen Inzuchtstarnrnes HK 51 mit einer fittleren Korpermasse von 112 f 6 g. Bis 15 Stunden vor Versuchsbeginn erhielten die Tiere ad libitum ein Standardrattenfutter (16% Rohprotein); wahrend des Versuches stand ihnen ausschliel3lich Trankwasser zur Verfiigung.
Als Tracer wurde (8-I"C)- bzw. (U-I4)-Adenin (Radio Chemical Centre Arnersham Buchler GrnbH & Co KG, Braunschweig) in physiologischer NaC1-Losung aufgenommen (50 pCi/ml) und 0,5 ml/Tier rnit einer Tuberkulinspritze in die rechte laterale Schwanzvene injiziert oder mit Hilfe einer Oesophaguskanule direkt in den Magen der Versuchstiere verabreicht. Die Tiere erhielten vergleichbare '"C-Dosen zwischen 5,8 pCi und 6,3 KCi.
U.S. Copyright Clearance Center Code Statement: Z. Tierphysiol, Tierernahrg. u. Futtermittelkde. 47 (1982), 10-20 0 1982 Verlag Paul Parey, Hamburg und Berlin ISSN 0044-3565 / InterCode: ZlTFAA
0044-3565/82/4701-0010 $ 02.50/0

I 4 C-Ausscheidung und -Retention nach Applikation von ‘‘C-Adenin an Ratten 11
22 Versuchsdurchfuhrung und Analytik
Sofort nach Verabreichung der I4C-Tracer wurden die Ratten fur 24 Stunden in Respirations- kafige gesetzt. Als Stoffwechselkarnrnern dienten Exsikkatoren rnit Seiten- und Deckeltuben. Der Einbau von zwei in der Hohe versetzten Drahtgittern rnit unterschiedlicher Maschenweite errnoglichte die Trennung von Kot und Harn. Die kontinuierliche Messung der 14C02-Verat- rnung erfolgte rnit Hilfe eines MethandurchfluBzahlers (MeBintervall 5 Min.) bei einern Wirkungsgrad von ca. 1%. Die MeBwerte wurden uber einen Scaler-Timer analog auf einen Servogor-Schreiber und digital auf einen Kinder-Drucker ausgegeben. Eine genauere Beschrei- bung der Versuchsanlage geben die Arbeiten von EYO (1975) und GREIFE (1977). Zur quantita- tiven Bestirnrnung der ausgeatrneten I4C-Aktivitat wurde das gesarnte C 0 2 irn AnschluB an die 14C02-Verlaufsrnessung in 2 n NaOH absorbiert.
Nach Versuchsende (Versuchsdauer 24 Stunden) wurden die Tiere rnit Diathylather getotet und Kot und Harn quantitativ gesarnrnelt. Der Intestinaltrakt wurde entnornrnen, der Lange nach aufgeschnitten und der Inhalt rnit einern Hornspatel vorsichtig ausgeschabt. Urn eine Uberschatzung der “C-Retention zu verhindern, wurden Magen und Darm sauber ausge- waschen. Der Blaseninhalt wurde rnit dern Harn vereinigt, die Blase anschliegend ebenfalls gereinigt. Das Spulwasser von Darrn und Blase wurde nicht gesarnrnelt.
Der Inhalt des Magen-Darm-Traktes (MDT-Inhalt) sowie der Kot wurden nach der Gefrier- trocknung in einem Morser hornogenisiert. Der entleerte Magen-Darrn-Trakt und der verblei- bende Restkorper wurden getrennt in 6 n HCI unter Erhitzen vollstandig hydrolysiert und gefiltert. Die Filtrate wurden rnit aqua dest. auf definierte Volurnina aufgefullt, die Filter verei- nigt und bei 60°C unter Vakuurn getrocknet. Die extrahierten atherloslichen Bestandteile ergaben die Korperfett-Fraktion. Die Filterruckstande wurden nicht aufbereitet.
Die 14C-Aktivitat in den Tracerlosungen und irn Harn konnte direkt unter Verwendung eines Dioxan-Szintillatorsysterns (Quickszint 454; Fa. Zinsser) gernessen werden. Als geeignet zur weitgehend quenchfreien 14C-Bestirnrnung in der Natronlauge (Absorptionsflussigkeit des 14C02) erwies sich ein Szintillatorsystern auf Basis Pseudocurnol (1, 2,4-Trirnethylbenzol; Fa. Baker). Von den gefriergetrockneten Hornogenaten (Kot, MDT-Inhalt) wurden 150-200 rng in einern Oxyrnaten verbrannt, wobei der freiwerdende Radiokohlenstoff in phenethylarninhal- tiger Szintillatorfliissigkeit (330 rnl Phenethylarnin, 220 rnl Methanol, 400 rnl Toluol, 75 rnl aqua dest., 7,O g PPO, 0,7 g BIS-MSB) absorbiert wurde. Die Aktivitatsrnessung der durch Saurehydrolyse hornogenisierten Korperfraktionen erfolgte nach Soluene-AufschluB der Hydrolysate (GREIFE 1977) in einem Toluolszintillator (200 rng Dirnethyl-POPOP, 4 g PPO, ad 1000 ml Toluol). Die Aktivitat der atherloslichen Lipide konnte nach Abdarnpfen des Athers rnit dern gleichen Szintillatorsystern direkt ermittelt werden.
Der Einflul3 der I4C-Substanzen (8-I4C)- bzw. (U-14C)-Adenin und der Art der Verabrei- chung (i. v./oral) auf die Versuchsergebnisse wurde rnittels doppelter Varianzanalyse (Anova) untersucht. Unterschiede der Gruppenrnittelwerte wurden sowohl zwischen den I4C- Substanzen bei gleicher Applikationsart als auch innerhalb der Substanzen zwischen den Applikationsarten rnit Hilfe des t-Testes abgesichert. Zu den Mittelwerten wird in den Ergeb- nistabellen jeweils die Standardabweichung der Einzelwerte ( f -Werte) angegeben.

12 H. A. Greije und S, Moinar
3 Versuchsergebnisse
3.1 "C-Ausscheidung iiber Atemluft und Exkremente
Die Verteilung der 14C-Aktivitat auf die Ausscheidungswege und den Inhalt des Magen-Darm- Traktes wird in der Tab. 1 dargestellt. Aus uniform IJC-markiertem Adenin wurde bei gleicher
Tabellc 1
Die 14C-Aktivitatsausscheidung innerhalb von 24 Stunden nach i .v . und oraler Verabreichung von "C-Adenin an Ratten
(% der "C-Dosis)
Magen-Dan-Tra k t T i e r z a h l A temlu f t Harn KO t - I n h a l t s u m
i . v . o r a l . i . v . o r a l I4C-Adenin i . v . / o r a l i . v . o r a l i . v . o r a l i . v . o r a l
27.3 98.6 + 1.9 2 1,9 f 0.4 : 0.4 f O,6 2 0.2 + 2 .3 f 2,0
( 8-14C)-Adeni n 4 4 0,9 4,3 23 .0 91.5 0.4 0.8 2.9 2,2 + 0.2 f 0,9 -
t -Tes t i . v . / o r a l ++++ ++++ n . s . n . 5 . ++++
(U-14C)-Adenin 8 8 4,7++++2 1, 5t+++ 2 1 .3n ' ' 70 ,O++ft o , 3 n . s . 1,4n.s. 3,0n.s. 2 , 6 n . s . 29,3n.s'95,4t
t - T e s t i . v . / o r a l ++++ tttt +t n.s. ti++
+ 2.2 f 2.0 - + 0.3 0.5 t 2.1 : 2.5 0,1 f 0,8 f 0,8 f 0.7 -
___________.____._______________________~.~-~~~~-~--------------------------------------------------------------------------------~
F-Wert f u r : 145,l++++ 3,4".5. 0,on.s. 1,3"" Substanz 2300,5t+tC
Verab re i chungsar t 35 14,6++++ 4325,8++++ 12,1++ 4.6".'. 6515 ,2++++
Wechsel w i r kung 847.2"" 102,9++++ 6.1' 0 , 1 ~ , ~ . 10.4"
D i e s t a t i s t i s c h e Auswertung nach dem t -Test e r f o l g t e i n den Tabe l l en 1-3 1.) zwlschen den Verabreichungsarten ( i . v . I o r a 1 ) i n n e r -
h a l b d e r Substanzen und 2.) zwischen den SubstanZen b e i q l e i c h e r A o p l i k a t i o n s a r t
' p t 0.05 ++ p 5 0.01 +++ p 5 0,001 *'++ p 5 0,0001 ""' = n i c h t s i g n i f i k a n t
Applikationsart jeweils etwa die fiinffache Menge an 14C0, veratmet wie aus der spezifisch (8- I4C)-markierten Substanz. Unabhangig von der Markierungsart wurde nach oraler Applikation der Tracer wiederum fiinfmal so vie1 "CO, uber die Respirationsluft ausgeschieden wie nach i.v. Injektion. Absolut betrachtet fiihrte i. v. injiziertes (8-"C)-Adenin rnit 0,9% der I 'C-Dosis zur geringsten, oral verabreichten (U-"C)-Adenin rnit 21,5% zur hochsten I 'CO,-Freisetzung.
Uber den Harn wurden jeweils die Hauptaktivitatsanteile ausgeschieden, wobei die Appli- kationsart einen hoch signifikanten EinfluB ausiibte. Wahrend nach i. v. Injektion sowohl aus (8-' 'C)- als auch aus (U-'"C)-Adenin gut 20% der "C-Dosis im Harn analysiert wurden, waren es nach oraler Verabreichung der Tracer etwa 90% bzw. 70%.
Mit dem Kot wurden nach oraler Applikation der Tracer geringfiigig hohere Aktivitatsan- teile abgegeben als nach i. v. Injektion, quantitativ blieb dieser "C-Ausscheidungsweg aber von geringer Bedeutung. Auf den Inhalt des Magen-Darm-Traktes entfielen unabhangig von der Art der ''C-Markierung oder der Verabreichung zwischen 2% und 3% der "C-Dosis.
Hoch signifikante Auswirkungen auf die gesamte 14C-Ausscheidung (einschlieRlich "C im MDT-Inhalt) gingen von der Verabreichungs-, nicht aber von der Markierungsart aus. Der mit den Tracern oral applizierte Radiokohlenstoff wurde zu uber 95% ausgeschieden, dagegen unter 30% vom i.v. injizierten I4C.

“C-Aussrheidung und -Retenlion nach Applikarion 7101 ‘‘C-Adenin an Ralten 13
1200.
1000.
800- E
E 600. - - > $ Loo-
200.
3.2 Der zeitliche Verlauf der “COy-Veratmung
Wegen des geringen Wirkungsgrades des MethandurchfluBzahlers (ca. lob) wurde nur nach oraler Applikation von (U-I4C)-Adenin ein auswertbarer Verlauf der “CO,-Exhalation aufge- zeichnet (Abb. I). Bereits 5 Minuten nach der Tracerapplikation wurde “C-Aktivitat in der Respirationsluft registriert und nach etwa 20 Minuten die maximale “COY-Ausscheidung
1400 I
I 15 30 L5 60 75 90 105 120 135 150 165 Min. 180
Abb. 1. Der Verlauf der 14C0,-Freisetzung aus oral an Ratten verabreichtern (U-”C)-Adenin (MeRintervall 5 Min.)
erreicht. Die ‘TO,-Veratmung war 90 Minuten nach Versuchsbeginn auf eine relativ geringes Niveau abgesunken, iiber einen Zeitraum von insgeamt 60 Stunden war sie nachweisbar. Wie die kumulative 14COyVerlaufskurve verdeutlicht (Abb. 2), wurden innerhalb der ersten Stunde mehr als 50% der in 6 Stunden insgesamt veratmeten I4C-Aktivitat ausgeschieden, nach 3 Stunden waren es iiber 80%.
I 1 2 3 L 5 Sld. 6
Abb. 2. Der kurnulative Verlauf der “CO,-Freisetzung aus oral an Ratten verabreichtern (U-“C)-Adenin
3.3 Die I4C-Retention im Tierkorper
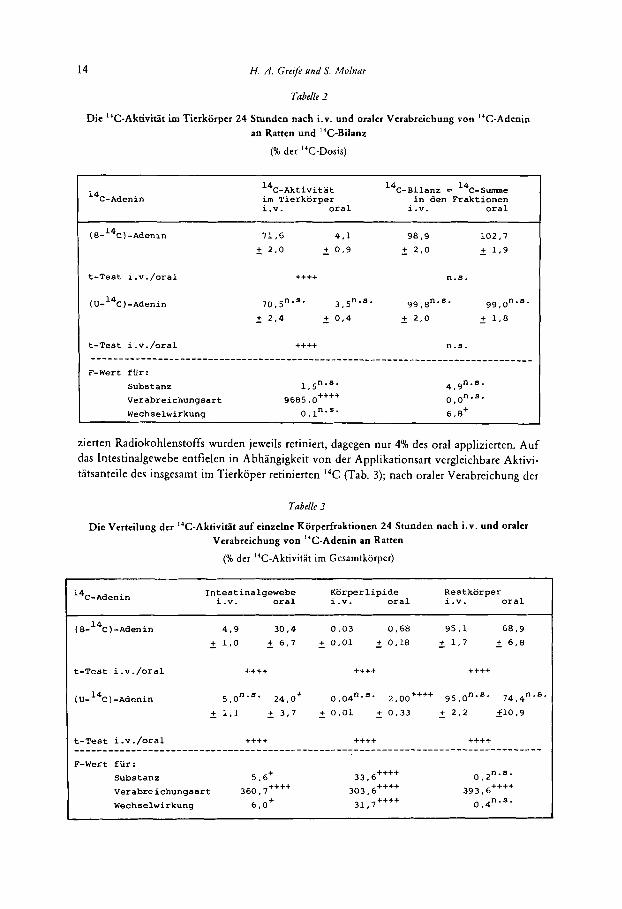
Die Hohe der im Tierkorper verbliebenen I4C-Aktivit3t wurde von der Applikationsart bestimmt, nicht aber von der Art der 14C-Markierung des Adenins (Tab. 2). Etwa 70% des inji-
14 H. A. Grefe und S. Molna?
t-Test i.v./oral ++++ ++++ ++++ .................................................................................. F-Wert fur:
Substanz 5.6+ 3 3 ,6++++ 0, 2".s-
Verabre ichungsart 360,7++++ 30 3,6++++ 39 3,6++++
Wechselwirkung 6,0+ 31,7++++ 0,4".'- I
Tabelk 2
Die "C-Aktivitat im Tierkorper 24 Stunden nach i.v. und oraler Verabreichung von I4C-Adeoin an Ratten und "C-Bilanz
(% der 14C-Dosis)
14C-Aktivitat "~-~ilanz = 1 4 ~ - ~ u m m e im Tierkorper in den Fraktionen i.v. oral i.v. oral
( 8-14C) -Adenin 71.6 4.1 98.9 102,7 - + 2.0 - + 0.9 - + 2.0 - + 1.9
t-Test i.v./oral ++++ n.s.
70,5n's' 3,5nas- 99, 8".s* 99.0"-8- - + 2.4 - + 0,4 - + 2,o - + 1.8
zierten Radiokohlenstoffs wurden jeweils retiniert, dagegen nur 4% des oral applizierten. Auf das Intestinalgewebe entfielen in Abhangigkeit von der Applikationsart vergleichbare Aktivi- tatsanteile des insgesamt irn Tierkoper retinierten I4C (Tab. 3); nach oraler Verabreichung der
Tab& 3
Die Verteilung der "C-Aktivitat auf einzelnc Korperfraktionen 24 Stunden nach i.v. und oraler Verabreichung von "C-Adenin an Ratten
(% der "C-Aktivitat im Gesamtkorper)

I 4C-Ausscheidung und -Retention nach Applikation von " C-Adenin an Ratten 15
Tracer war es die fiinf- bis sechsfache Menge von der i.v. Injektion. Nur Aktivitatsspuren waren in den Korperlipiden zu finden. Ein signifikant hoherer I4C-Einbau in diese Frakion wurde durch die orale Verabreichung der 14C-Substanzen erreicht, insbesondere von (U-I4C)- Aden i n .
Da sowohl die "C-Ausscheidung als auch die I4C-Retention analytisch erfaBt wurden, konnte zu Versuchsende eine ''C-Bilanz aufgestellt werden (Tab. 2). Ohne signifikante Grup- pendifferenzen wurden zwischen 99% und 103% des eingesetzten Radiokohlenstoffs wiederge- funden.
4 Diskussion
4.1 Ausscheidung des Radiokohlenstoffs aus I4C-Adenin
Da die i4C02-Freisetzung aus applizierten 14C-Tracern das Verhaltnis von anabolen zu kata- bolen Stoffwechselablaufen charakterisiert, wurde sie als Parameter zur Arninosaurenbedarfser- mittlung unter definierten Ernahrungsbedingungen bei wachsenden Ratten (BROOKES et al. 1972; BERGNER und SIMON 1976), Lamrnern (BROOKES et al. 1973) und Ferkeln (CHAVEZ und BAYLEY 1976 b) herangezogen. Fehlinterpretationen sind hierbei moglich, wenn venvertbare "C-Metaboliten auftreten (REEDS 1974) und wenn die Versuchsbedingungen von EinfluB auf die Pool-GroBe der verabreichten '4C-Aminosaure (CHAVEZ und BAYLEY 1976 a) sind.
Aus (8-14C)-Adenin wird als radioaktives Abbauprodukt aber ausschlieBlich 14C02 frei; die (U-14C)-markierte Substanz kann dariiber hinaus 14C-Glyoxylat liefern, wenn sie mikrobiell zersetzt wird (Abb. 3). Da nur geringe Tracermengen verabreicht wurden, ist auch kein EinfluB auf den Adenin-Pool zu envarten.
Herauszustellen sind die Auswirkungen, die von der Applikations- und der Markierungsart des Adenins auf die 14C02-Veratmung ausgehen. Beim intermediaren Purinabbau zum Allan-
0 Urikase
7 Allontoin Harnsaure Adenin
Allontoinase
0 0
II I 9 H', N ' 'NH, H,IN -'c -'NH, H,N - c - NH,
I Allontoikase Hornstoff Hornstoff Urease I + NH, + CO, + HCOOH I I, I I
O=C' 'COOH 'C=O
H N - C'-'NH 0
I H
Allan toinsaure
'11 I HC - COOH
Glyoxylot
Abb. 3. Schematische Darstellung des Adenin-Abbaus (modifiziert nach SAVAIANO et al. 1980)

16 H. A. Greqe und S. Molnar
toin wird ausschliei3lich das 6. C-Atom freigesetzt (CANELLAKIS und COHEN 1955; Abb. 3). Verschiedene Mikroorganismen konnen eine vollstandige Zersetzung der Purine zu CO, und N H , erreichen (VOGELS und VAN DER DRIFT 1976). Der als I4CO2 aus der 8-C-Position verat- mete Radiokohlenstoff zeigt somit das AusrnaB der mikrobiellen Purinzersetzung im Darm an. Einen deutlichen Hinweis auf den enteralen Abbau des Purinringes liefert die im Vergleich zur i.v. Injektion hohere 14COz-Exhalation aus oral verabreichtern (8-"C)-Adenin. Die geringe 14C02-Freisetzung aus dem injizierten Tracer kann auf eine Sekretion des Adenins in den Darm (BERLIN und HAWKINS 1968) zuriickzufuhren sein, aber auch als Folge radioaktiver Frerndsubstanzen in der Tracerlosung erklart werden. Die diinnschichtchromatographisch uberprufte radiochemische Reinheit der Isotopen betrug allerding 99%.
Die sowohl nach i. v. als auch nach oraler Applikation hohere "C0,-Veratmung aus uniform I4C-markiertem Adenin laBt zweifelsohne den SchluB zu, daB die Purinbase interrne- diar teilweise zurn Allantoin abgebaut wurde und die ''C-Aktivitatsgehalte im Harn weitge- hend auf dieses Ausscheidungsendprodukt der Purine entfallen. Abgeleitet wird diese Interpre- tation der 14C02-Freisetzung in der Tab. 4.
Der Purinring enthalt insgesamt fiinf C-Atorne. Wird er zum Allantoin abgebaut, entweicht das C-Atom in Position 6 des Ringes als I4CO2, das sind 20% des gesamten C-Gehaltes. Nehrnen wir einmal an, daB der Radiokohlenstoff im Harn ausschliefilich auf Allantoin entfallt, so IaRt sich hieraus die theoretische 14C0z-Veratmung aus (U-14C)-Adenin ableiten (Punkt 2. in der Tab. 4). Hierzu addiert werden mug die '"CO,-Freisetzung, die auf den voll- standigen mikrobiellen Purinabbau zuruckzufuhren ist (= '*CO,-Veratmung aus (8-l'C)- Adenin), urn die theoretisch rnogliche 14C0,-Exhalation aus (U-"CC)-Adenin zu errechnen (Punkt 4. in der Tab. 4).
Tabelle 4
Riickschlul3 von der "C0,-Veratmung aus (8-I4C)- und (U-I4C)-Adenin auf die renalen 14C~Ausscheidungsprodukte
I 1 4 C i m Harn bzw. 14CO--Vera tmuna I % der l 4 C - D o s i s i . v . o r a l
1. 1 4 C i m Harn aus ( U - 1 4 C ) - A d e n i n 2 1 . 3 70.0
2 . 14C02-Veratmung, WWI 1 4 C i m Harn = 1 4 C im A l l a n t o i n (=1./4) 5.3 17,5
3. 1 4 C 0 -Veratmung a u s ( 8 - 1 4 C ) -Adenin 0 . 9 4 . 3
4. 14C02-Veratmung a u s ( U-14C) -Adenin , berechnet (Summe 2 .+3. )
2
6 . 2 21.8
5. 14C02-Veratmung a u s (U-14C)-Adenin, g e m e s s e n 4.7 21.5
Der kalkulierte Wert stimmt nach oraler Applikation des (U-"C)-Tracers mit der gemes- senen "C02-Exhalation iiberein. Die ''C-Aktivitatsausscheidung irn Harn laBt sich also vollig durch die Exkretion von 14C-Allantoin erklaren. Ebenso durfte demnach die hohe renale "C- Ausscheidung aus oral verabreichtern (B-"C)-Adenin auf Allantoin entfallen.
Nach Injektion der (U-"C)-Purinbase wurde eine etwas geringere '4C0,-Freisetzung gernessen als kalkuliert werden konnte. Abgesehen von Fehlern bei der Uberschlagsrechnung,

"C-Ausscbeidung und -Retention nacb Applikafion von "C-Adenin an Rafien 17
kann diese Differenz auch durch die direkte Ausscheidung von Adenin oder der oxydierten Base 2,8-Dihydroxyadenin (BENDICH et al. 1950; CLIFFORD und STORY 1976; AKINTONWA et al. 1979) eingetreten sein. So wies die radiochrornatographische Analyse des Harns nach oraler Applikation von (8-I4C)-Adenin an Ratten irn Allantoin einen Anteil von 95% der gesarnten Harnakivitat aus, nach i.v. Injektion dagegen nur von 80% (Ho et al. 1979). Ausgeschlossen werden kann aber auch nicht eine geringe I4C-Markierung der Harnsaure und bei unvollstan- diger rnikrobieller Purinzersetzung der weiteren C-haltigen Abbauprodukte, wie von SAVAIANO et al. (1980) nach oraler Gabe von (8-I4C)-Adenin an Ratten dernonstriert wurde (siehe hierzu Abb. 3).
Die geringen I4C-Gehalte irn Kot nach oraler Verabreichung der Tracer verdeutlichen eine hohe Absorptionsrate des Adenins. So entfielen auch auf den Inhalt des Magen-Darrn-Traktes nicht hohere Aktivitatsanteile als nach i.v. Injektion der "C-Substanzen. Die Werte konnen hierbei aber durch abgeschabte Darrnepithelien uberschatzt worden sein. Auf eine schnelle Absorption weisen die hohen I4CO,-Gehalte der Aternluft kurz nach oraler Verabreichung von (U-I4C)-Adenin und die ausgepragte Peakphase der 14C0,-Verlaufskurve hin. Die am "'COp-Verlauf abzulesende rasche Metabolisierung des Tracers lagt gleichzeitig eine starke Beteiligung der Darrnflora beirn Adeninabbau ausschliegen. Zu iiberpriifen ware, o b bereits irn bekannten ,,Vorrnagensystern" der Ratte (KUNSTYR 1974) ein bedeutender Abbau der Purine einsetzt.
4.2 Retention des Radiokohlenstoffs aus I4C-Adenin
Die I4C-Retention aus rnarkiertern Adenin wurde nur durch die Applikationsart (i.v./oral) bestirnrnt. Wahrend uber 70°h des injizierten Radiokohlenstoffs irn Tierkorper verblieben, waren es nur etwa 4% nach oraler Tracerapplikation. Die rnoglichen Stoffwechselwege des Adenins werden in der Abb. 4 skizziert.
1 - Adenin-Phosphoribosyltransferase, 2- Adenosin-Kinase, 3 - Adenosin-Desaminase, 4 - Nukleosid- Phosphorylase, 5 - HypoxanthidGuanin-PRPP-Transferase, 6 - 5'-Nukleotidasen
Adenosin 1
Adenin /
I Hornsaure
1 Allantoin
Abb. 4. Mogliche Stoffwechselwege deran Ratten verabreichten Purinbase Adenin(modifiziert nach SHENOY , und CLIFFORD 1975).
Die hohe Retention von injiziertem I4C-Adenin bestatigen Arbeiten an verschiedenen Tierarten (Maus: BENNET 1953; Ratte: GREIFE und MOLNAR 1978 b; SAVAIANO et al. 1980;

18 H. A. Grefe und S. Molnar
Kaninchen: BARTLETT 1972; 1977 a; Hammel: PRICCE 1977) und dem Menschen (BARTLETT 1977 b). GREIFE und MOLNAR (1978 b) fiihren die hohe Retention des Radiokohlenstoffs aus I4C-Adenin auf eine schnelle Uberfuhrung des Adenins in AMP zuriick, ein Mechanismus, der die Zelle wirksam vor freiern Adenin zu schutzen vermag (MURRAY et aI. 1970). So war die iiber Adenin injizierte "C-Aktivitat auch vor allem in den Adenin-Nukleotiden (AMP, ADP, ATP) von Korpergeweben nachzuweisen (BENNET 1953; BARTLETT 1972; 1977 a,b),
Oral verabreichtes Adenin wird bereits im Darmlurnen mikrobiell (Desarninierung) oder in der Darmmukosa enzyrnatisch iiber AMP zurn Hypoxanthin und Xanthin abgebaut (Abb. 4) und deshalb nur zu geringen Anteilen unverandert absorbiert und in Korpernukleotide einge- baut worden sein. Hierbei ist die anteilmagig hohe Nutzung der Purinbase im Intestinalge- webe selbst herauszustellen, wie auch am Beispiel der Maus (SONODA und TATIBANA 1978) und des Meerschweinchens (MACKINNON und DELLER 1974) demonstriert wurde.
Die aus (8-I4C)-Adenin inkorporierte I4C-Aktivitat laBt direkt auf die Verwendung der Base bei der Nukleotidsynthese schliegen, weil metabolisches C02 als einziges radioaktives Abbau- produkt nur gering an Syntheseprozessen beteiligt ist (GRONEUER et al. 1974). Uber eine Fixie- rung im Oxalacetat mit Hilfe der PyruvatCarboxylase, im Malat unter Katalysierung des Mala- tenzyms oder im Succinyl-CoA durch eine Karboxylierung von PropionylCoA kann I4CO2 aber in den Citrat-Zyklus gelangt sein und damit u.a. zu einer "C-MarkieAng des Glycerinan- teils der Korperlipide gefiihrt haben. Aus oral verabreichtem (U-14C)-Adenin kann mikrobiell "C-markiertes Glyoxylat freigesetzt worden sein (siehe Abb. 3) und diese Verbindung iiber den bei Mikroorganismen bekannten Glyoxylat-Zyklus (SCHLECEL 1976) markiertes Acetyl- CoA zur Fettsynthese geliefert haben. Angesichts des augerst geringen I4C-Einbaus in die Lipidfraktion bleibt diese Annahme aber reine Spekulation.
Sowohl bei Saugern als auch beim Geflugel sind Stoffwechselschaden durch eine hohe Adeninaufnahme bekannt, wie Wachstumsdepressionen bei Adeningehalten von 0,3% in der Rattendiat (CLIFFORD und STORY 1976) und von 0,4% im Kukenfutter zeigen (BAKER und MOLI- TORIS 1974). Endogen und aus Nahrungs-Nukleinsauren wird die Purinbase Adenin wegen des Fehlens einer Adenosin-Phosphorylase in tierischen Geweben nicht freigesetzt (siehe Abb. 4). Geringe Mengen der exogen zugefiihrten freien Base konnen durch den aufgezeigten interme- diaren Schutzmechanisrnus - den raschen Aufbau zu AMP, katalysiert durch die Adenin- Phosphoribosyltransferase - abgefangen werden. Nur die genaue Kenntnis der Basenzusam- mensetzung der Nukleinsauren-Fraktion sowie des Gehaltes an freien Purinbasen ermoglicht eine Abschatzung der potentiellen Risikofaktoren bei Austausch konventioneller durch mikrobielle Proteintrager in der Tierernahrung.
Zusammenfassung
Die Purinbase Adenin wurde als (8-I'C)- bzw. als (U-I4C)-Tracer alternativ in die rechte laterale Schwanzvene (i.v.) oder in den Magen (oral) von mannlichen Albinoratten (1 12 f 6 g) verab- reicht. Nach 24 Stunden wurde die ''C-Ausscheidung uber Atemluft, Harn und Kot sowie die I4C-Retention im gesamten Tierkorper, dem Gastrointestinaltrakt (ohne Inhalt) und den Korperlipiden analysiert. Nach oraler Applikation des (U-I4C)-Tracers wurde der Verlauf der 14C02-Veratmung mit Hilfe eines MethandurchfluGzahlers kontinuierlich verfolgt.
Die rasche Veratmung von I4CO2 aus oral verabreichtem (U-'4C)-Adenin zeigte eine schnelle Absorption und Verstoffwechselung der Substanz an. Wahrend eine Freisetzung von I4CO, aus der (8-"C)-Purinbase nur durch mikrobielle Zersetzung moglich ist, wird aus der

I4CC-Ausscheidung und -Retention nach Applikation von “C-Adenin an Ratten 19
(U-14C)-Substanz 14COz aus der 6. C-Position frei, wenn sie zum Allantoin abgebaut wird. Der Vergleich der L4COz-Exhalation in Abhangigkeit von der Markierungsart 1a13t den indireken SchluS zu, da13 die renale “C-Exkretion fast ausschlieBlich auf I4C-Allantoin entfillt. Da aus oral appliziertem (8-I4C)-Adenin mehr 14C02veratmet wurde als aus i. v. injiziertem, mu13 auf einen teilweisen mikrobiellen Abbau der Purinbase im D a m geschlossen werden.
Unabhangig von der Markiemngsart wurde der mit den Tracern oral applizierte Radiokoh- lenstoff zu iiber 95% ausgeschieden, dagegen unter 30% vom i.v. injizierten 14C. Die hochste Aktivitatsabgabe erfolgte jeweils iiber den Harn, die geringste iiber den Kot.
Retiniert wurden etwa 70% der injizierten 14C-Aktivitat, aber nur 4% der oral verabreichten, wovon 20%-30% auf das Intestinalgewebe entfielen. Ein intensiver Katabolismus im D a m und der Darmschleimhaut scheint eine hohere Nutzung des oral applizierten Adenins zu verhin- dern.
Von geringer Bedeutung ist eine Fixierung von ”COz bei Syntheseprozessen; nur Aktivi- tatsspuren entfielen 2.B. auf die Korperlipide. Da l4CO2, das als einziges radioaktives Abbau- produkt aus (8-I4C)-Adenin auftritt, weitgehend veratmet wird, 1aSt die inkorporierte I4C- Aktivitat auf die Venvendung der Purinbase bei der Nukleotidsynthese schliegen.
Summary
“C-excretion and -retention after application of “C-adenine to rats with special regard to “CO,-liberation as a parameter o f metabolic process.
The purine base adenine was administered as (8-I4C)- or as (U-14C)-tracer alternatively in the right tail vein (i.v.) or in the stomach (oral) of male albino rats (1 12 f 6 g). After 24 hours the excretion of “C-activity in respiratory air, urine and faeces was determined as well as reten- tion of 14C in whole bodies, gastro-intestinal tracts (exclusive contents) and body lipids. After oral application of the (U-’’C)-tracer the course of “COz-expiration was continously followed by use of a methane counter.
A quick absorption and catabolism was indicated by the fast expiration of I4CO2 after oral application of (U-I4C)-adenine. When (U-I4C)-adenine is catabolized to allantoin I4CO2 will be liberated from the 6. C-position of the purine ring whereas only microbial degradation of (8-I C)-adenine produces 14COz. The results of the 14C0z-exhalation with respect to the kind of labelling allows one to indirectly conclude that 14C in urine is excreted almost exclusively as 14C-allantoin. Part of the purine base must have been decomposed by microbes in the gut because more 14C02 was expired after oral application of (8-I4C)-adenine as compared to i.v. injection.
Irrespective of the kind of labelling more than 95% of the orally but less than 30% of the i. v. administered radioactivity were excreted. The I4C-activity was mainly excreted in the urine whereas only small amounts appeared in the faeces.
About 70% of the I4C-dose injected were retained but only 4% of the orally applied 14C- dose were retained. After oral dosing 20%-30% of the 14C-incorporation were retained in the intestinal tissues. An intensive catabolism in the gut and gut tissue seems to prevent a more effective utilisation of orally applied adenine.
The fixation of l4CO2 for synthetic purposes is of minor importance. For instance only little “C was detected in the body lipids. I4COz, as the only radioactive catabolic product of (8-14C)-adenine, is almost completely exhaled. Therefore the 14C-incorporation indicates that the purine base was utilised for the synthesis of nucleotides.

20 H. A. Gr+ und S. Moinur
Literatur
1. AKINTONWA, A,; AUDITORE, J. V.; GREEN, L. D., 1979: Appl. Pharmacol. 47, 229-235. 2. BAKER, D. H.; MOLITORIS, B. A,, 1974: J. Nutr. 104, 553-557. 3. BARTLETT, G. R., 1972: Internationales Symposium iiber Struktur und Funktion der Erythrocyten. VI.
4. BARTLETT, G. R., 1977 a: Transfusion 17, 351-357. 5. BARTLETT, G. R., 1977 b: Transfusion 17, 367-374. 6. BENDICH, A,; BROWN, G. B.; PHILIPS, F. S.; THIERSCH, J. B., 1950: J. Biol. Chem. 183, 267-277. 7. BENNETT, E. L., 1953: Biochim. Biophys. Acta 11, 487-496. 8. BERGNER, H.; SIMON, 0.. 1976: Arch. Tierernahr. 26, 815-826. 9. BERLIN, R. D.; HAWKINS, R. A,, 1968: Am. J. Physiol. 215, 932-941.
Berlin. 355-362.
10. BROOKES, 1. M.; OWENS, F. N.; GARRIGUS, U. S., 1972: J. Nutr. 102, 27-36. 11. BROOKES, 1. M.; OWENS, F. N.; BROWN, R. E.; GARRIGUS, U. S., 1973: J. Anim. Sci. 36, 965-970. 12. CANELLARIS, E. S.; COHEN, P., 1955: J. Biol. Chem. 213, 385-395. 13. CHAVEZ, E. R.; BAYLEY, H. S., 1976 a: Br. J. Nutr. 36, 189-198. 14. CHAVEZ, E. R.; BAYLEY, H. S., 1976 b: Br. J. Nutr. 36, 369-380. 15. CLIFFORD, A. J.; STORY, D. L., 1976: J. Nutr. 106, 435-442. 16. EYO, E. S., 1975: Untersuchungen uber den Verlauf der “C-Aktivitat in der Atemluft und iiber die
Verteilung der ‘“C-Aktivitat in einigen Korperfraktionen nach oral verabreichten “CC-markierten Substanzen. Diss. sc. agr., Gottingen.
17. GREIFE, H., 1977: Untersuchungen zum Nukleinsaurestoffwechsel der Ratte unter Einsatz “C-markier- ter Purin-, Pyrimidinbasen und Nukleinsauren. Diss. sc. agr., Gottingen.
18. GREIFE, H.; MOLNAR, S., 1978 a: Z. Tierphysiol., Tiererndhrg. u. Futtermittelkde. 40, 236-247. 19. GREIFE, H.; MOLNAR, S., 1978 b: Z. Tierphysiol., Tierernahrg. u. Futtermittelkde. 40, 248-256. 20. GREIFE, H.; MOLNAR, S., 1979: Z. Tierphysiol., Tierernahrg. u. Futtermittelkde. 41, 184-197. 21. GRONEUER, K. J.; KRAMPITZ, G.; HARDEBECK, H.; FOHR, F., 1974: Z. Tierphysiol., Tierernahrg. u. Futter-
22. Ho, C. Y.; C H U , V.; SAVAIANO, D. Y.; CRANE, R. T.; WEIR, W. C.; CLIFFORD, A. J., 1979: Fed. Proc. 38,
23. KUNSTYR, I., 1974: Zbl. Vet. Med. A 21, 553-561. 24. MACKINNON, A. M.; DELLER, D. J., 1973: Biochim. Biophys. Acta 319, 1-4. 25. MOLNAR, S.; EYO, E. S.; MEULEN, U. TER, 1977: Z. Tierphysiol., Tierernahrg. u. Futtermittelkde. 39,
26. MURRAY, A. W.; ELLIOTT, D. C.; ATKINSON, M. R., 1970: Progr. Nucl. Acid Res. Mol. Biol. 10, 87-119. 27. PRIGGE, H., 1977: Beeinflussung der renalen Allantoin-Ausscheidung bei Hammeln durch Infusion ver-
28. REEDS, P. J., 1974: Br. J. Nutr. 31, 259-270. 29. SAVAIANO, D. A.; Ho, C. Y.; CHU, V.; CLIFFORD, A. J., 1980: J. Nutr. 110, 1793-1804. 30. SCHLEGEL, H. G. (Herausgeber), 1976: Allgemeine Mikrobiologie. 4. Auflage. Stuttgart: Georg Thieme
31. SHENOY, T. S.; CLIFFORD, A. J., 1975: Biochim. Biophys. Acta 521, 55-66. 32. SONODA, T.; TATIBANA, M., 1978: Biochim. Biophys. Acta 41 1 , 55-66. 33. VOGELS, G. D.; VAN DER DRIFT, C., 1976: Bacteriol. Rev. 40, 403-468.
Anscbrifi der AIIforen: I ns ti tu t fur Tierp h ysiol ogi e und Tie re rna hrung, 0 s kar-Kell ner-Weg 6,
mittelkde. 33, 289-304.
608.
96-104.
schiedener Substanzen in den Labmagen. Diss. sc. agr., Gottingen.
Verlag. S. 223.
3400 Gottingen-Weende


![シクロプロトリン...10 2011 2 3 5 3 14C [cyc-14C] 3-14C [phe-14C] mg/kg g/g 1 2 Fischer 3 [cyc-14C] 50 mg/kg 5,000 mg/kg Fischer 3 7 2 1 50 mg/kg 5,000 mg/kg 50 mg/kg / Tmax](https://img.pdfslide.net/doc/110x75/60ef4da753c03d22e86933e7/ffffff-10-2011-2-3-5-3-14c-cyc-14c-3-14c-phe-14c-mgkg.jpg)
![VE1-3.PPT [Schreibgeschützt] [Kompatibilitätsmodus] · Toxikokinetik • Resorption (Aufnahme) • Verteilung • Metabolismus (Biotransformation) • Elimination (Ausscheidung)](https://img.pdfslide.net/doc/110x75/5d4f197688c9932e758b9f6e/ve1-3ppt-schreibgeschuetzt-kompatibilitaetsmodus-toxikokinetik-resorption.jpg)




![SkeletalMuscleAMP-activatedProteinKinaseIsEssentialfor … · 2009-10-07 · 15min.Theupperaqueousphase(containing2-[14C]DG)wasusedtodetermine2-[14C]DG-Pasdescribedpreviously(29).Aportion](https://img.pdfslide.net/doc/110x75/5f3b5d9a091da77b4f678521/skeletalmuscleamp-activatedproteinkinaseisessentialfor-2009-10-07-15mintheupperaqueousphasecontaining2-14cdgwasusedtodetermine2-14cdg-pasdescribedpreviously29aportion.jpg)