Embed Size (px)
Citation preview

ORIGINAL RESEARCHpublished: 17 November 2016
doi: 10.3389/fphys.2016.00545
Frontiers in Physiology | www.frontiersin.org 1 November 2016 | Volume 7 | Article 545
Edited by:
Jean-Pierre Montani,
University of Fribourg, Switzerland
Reviewed by:
Anna Alisi,
“Bambino Gesù” Children’s Hospital
and Research Institute, Italy
Gabriella Gallo,
University of Genoa, Italy
*Correspondence:
Maria Moreno
Fernando Goglia
†These authors have contributed
equally to this work.
Specialty section:
This article was submitted to
Integrative Physiology,
a section of the journal
Frontiers in Physiology
Received: 16 September 2016
Accepted: 28 October 2016
Published: 17 November 2016
Citation:
Moreno M, Silvestri E, Coppola M,
Goldberg IJ, Huang L-S, Salzano AM,
D’Angelo F, Ehrenkranz JR and
Goglia F (2016)
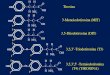
3,5,3′-Triiodo-L-Thyronine- and
3,5-Diiodo-L-Thyronine- Affected
Metabolic Pathways in Liver of LDL
Receptor Deficient Mice.
Front. Physiol. 7:545.
doi: 10.3389/fphys.2016.00545
3,5,3′-Triiodo-L-Thyronine- and3,5-Diiodo-L-Thyronine- AffectedMetabolic Pathways in Liver of LDLReceptor Deficient MiceMaria Moreno 1*†, Elena Silvestri 1 †, Maria Coppola 1, Ira J. Goldberg 2, Li-Shin Huang 2,
Anna M. Salzano 3, Fulvio D’Angelo 4, Joel R. Ehrenkranz 5 and Fernando Goglia 1*
1Department of Science and Technologies, University of Sannio, Benevento, Italy, 2Department of Medicine, Columbia
University, New York, NY, USA, 3 Proteomics and Mass Spectrometry Laboratory, Instituto per Il Sistema Produzione Animale
in Ambiente Mediterraneo, National Research Council, Napoli, Italy, 4 Biogem s.c.a r.l., Ariano Irpino (AV), Italy, 5Department
of Medicine, Intermountain Healthcare, Salt Lake City, UT, USA
3,5,3′-triiodo-L-thyronine (T3) and 3,5-diiodo-L-thyronine (T2), when administered
to a model of familial hypercholesterolemia, i.e., low density lipoprotein receptor
(LDLr)-knockout (Ldlr−/−) mice fed with a Western type diet (WTD), dramatically reduce
circulating total and very low-density lipoprotein/LDL cholesterol with decreased liver
apolipoprotein B (ApoB) production. The aim of the study was to highlight putative
molecular mechanisms to manage cholesterol levels in the absence of LDLr. A
comprehensive comparative profiling of changes in expression of soluble proteins in
livers from Ldlr−/− mice treated with either T3 or T2 was performed. From a total
proteome of 450 liver proteins, 25 identified proteins were affected by both T2 and T3,
18 only by T3 and 9 only by T2. Using in silico analyses, an overlap was observed with
11/14 pathways common to both iodothyronines, with T2 and T3 preferentially altering
sub-networks centered around hepatocyte nuclear factor 4 α (HNF4α) and peroxisome
proliferator-activated receptor α (PPARα), respectively. Both T2 and T3 administration
significantly reduced nuclear HNF4α protein content, while T2, but not T3, decreased the
expression levels of the HNFα transcriptional coactivator PGC-1α. Lower PPARα levels
were found only following T3 treatment while both T3 and T2 lowered liver X receptor
α (LXRα) nuclear content. Overall, this study, although it was not meant to investigate
the use of T2 and T3 as a therapeutic agent, provides novel insights into the regulation
of hepatic metabolic pathways involved in T3- and T2-driven cholesterol reduction in
Ldlr−/− mice.
Keywords: iodothyronines, liver, cholesterol, LDL receptor, proteomics
INTRODUCTION
Familial hypercholesterolemia (FH) is a common inherited disorder resulting from mutationsin the low-density lipoprotein receptor (LDLr) gene leading to defects in LDL cholesterol(LDL-C) clearance. Despite the use of currently available cholesterol-lowering treatments,a high proportion of FH patients do not reach treatment goals and remain at risk ofatherosclerotic cardiovascular diseases (Naoumova et al., 2004). More intensive and alternative

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
treatments are often needed for these patients as well as otherhypercholesterolemic patients who do not meet treatment goalsor who have difficulty with statin therapy. One possible approachto cholesterol reduction is via activation of hepatic thyroidreceptors (TRs).
Both 3,5,3′,5′-tetraiodo-L-thyronine (T4), and 3,5,3′-triiodo-L-thyronine (T3) (THs) reduce circulating cholesterol in animalsand humans (Klein and Danzi, 2008; Angelin and Rudling,2010). However, their therapeutic use for the treatment ofhyperlipidemia is limited due to deleterious side effects from TRactivation in extrahepatic tissues leading to altered cardiovascularfunction, muscle wasting, and bone loss (Scheiffele and Schultze,1972). In recent years, several thyromimetics (i.e., GC1 andKB2115) have been developed that selectively activate TRβ, thepredominant TR isoform in the liver that is primarily responsiblefor the effects on cholesterol and lipoprotein metabolism exertedby T3 (Johansson et al., 2005; Erion et al., 2007; Tancevskiet al., 2010; Pramfalk et al., 2011). These compounds retardatherosclerosis progression in animals and exert favorablelipid-modulating effects in humans, while lacking THs-relatedthyrotoxic side-effects (Bakker et al., 1998; Erion et al., 2007;Baxter and Webb, 2009; Pedrelli et al., 2010). However, sideeffects may limit its clinical use since the cholesterol-reducingthyrometic eprotirome (Ladenson et al., 2010) was recentlyshown to elevate circulating liver enzymes (Sjouke et al., 2014), asign of liver damage and, indeed, development of eprotirome wasterminated after cartilage damage was observed in a toxicologicalstudy in dogs.
Hepatic TR activation might lower cholesterol levels viamultiple mechanisms. Although the primary hepatic effects oncholesterol metabolism of THs was thought to be via LDLclearance through increased expression of LDLr (Ness and Lopez,1995), we and others have reported that iodothyronines [T3 and3,5-diiodo-L-thyronine (T2)] as well as GC1 markedly reducecirculating LDL-C in LDLr knockout (Ldlr−/−) mice (Goldberget al., 2012; Lin et al., 2012). These studies and those of others(Davidson et al., 1988, 1990) show that both iodothyronineslead to a reduction in hepatic secretion of apolipoprotein B(ApoB)-containing lipoproteins [very low density lipoproteins(VLDL) and LDL] while high density lipoprotein (HDL) levelsare reduced only by T3 (Goldberg et al., 2012).
This reduction is not associated with increased hepaticprotein or mRNA levels of LDL receptor related protein(LRP) 1 or the scavenger receptor-B1 (SR-B1) (Goldberget al., 2012). Cholesterol reduction was also reported tobe associated with increased expression of cholesterol-7α-hydroxylase (Cyp7a1) (Lin et al., 2012), which convertscholesterol into bile acids. These studies (Goldberg et al.,2012; Lin et al., 2012) dispelled the conventional view thatthyroid-mediated reduction of cholesterol requires LDLrand indicated that the thyroid hormone responsive element(TRE) in the LDL receptor is not required in vivo forthyroid-induced LDL reduction. Although our previousstudy (Goldberg et al., 2012) with high doses of T3 and T2 wasnot meant to investigate their use as therapeutic agents, theelicited dramatic reduction in circulating cholesterol levels inhypercholesterolemic Ldlr−/− mice opened new perspectives
in defining non-LDLr pathways that may have potential for thetreatment of hypercholesterolemia.
The goal of the following study was to gain additional insightsinto the molecular factors and pathways that contribute to theabove hypocholesterolemic actions of T2 and T3. To uncover thepathways and networks altered by these two iodothyronines, weperformed a comprehensive comparative profiling of changes inexpression of soluble proteins in livers from Ldlr−/− mice treatedwith either T3 or T2.
MATERIALS AND METHODS
Animals and Experimental ProtocolAll studies were approved by the Columbia UniversityInstitutional Animal Care and Use Committee (IACUC).Male wild-type (WT) C57BL/6 and Ldlr−/− mice of 3–4months of age, purchased from the Jackson Laboratory, wereused. WT C57BL/6 and Ldlr−/− mice (n = 5–6/group) werefed a Western-type diet (WTD) containing: 42% fat, 42.7%carbohydrate, 15.2% protein, 0.15% cholesterol; total 4.5 Kcal/g(Harlan Teklad) for 1 week. After 1 week, C57BL/6 andLdlr−/− mice were continued on the WTD and were dividedinto groups receiving vehicle (58.5% saline + 40% DMSO +
1.5% 1M NaOH) or iodothyronines—T3 (0.75mg/kg) (EMDChemicals/Calbiochem) or T2 (12.5 mg/kg) (Santa Cruz)—viadaily gavage for another week and then sacrificed. The doses ofT3 and T2 were chosen to obtain the same cholesterol reducingeffect (Goldberg et al., 2012). Mass spectrometry and NMRprofiles showed no T3 or T4 contamination in the T2. All bloodsamples obtained during the non-terminal portion of the studywere taken after a 4-h fast.
Protein Extraction and Sample Preparationfor Two-Dimensional Gel Electrophoresis(2D-E)Protein extraction and sample preparation for 2D-E wereperformed as reported in Silvestri et al. (2006). Liver tissue wassuspended in sample buffer [20 mMTris, 7 M urea, 2 M thiourea,4% CHAPS, 10 mM 1,4-dithioerythritol (DTE), 1 mM EDTA,and a mixture of protease and phosphatase inhibitors]. Thesuspensions were homogenized using a Polytron homogenizer,sonicated for 30 s, and centrifuged at 150,000 g for 45 min.The obtained supernatants contained the total liver proteinssolubilized in the isoelectrofocusing (IEF)-compatible agents.The protein content of each sample was determined by Bio-Rad’sDC method (Bio-Rad Laboratories, Hercules, CA). Total proteinextracts were prepared for each animal, and each individual wasassessed separately.
2D-ESamples of 650µg of protein were applied to immobilized pHgradient (IPG) strips (pH 4–7, 17 cm) (Bio-Rad). Samples of1mg of protein were utilized for preparative gels (IPG strips, pH4–7, 17 cm). Focusing started at 200 V, with the voltage beinggradually increased to 3500 V and kept constant for a further66,500 V/h (PROTEAN IEF System, Bio-Rad). Prior to SDS-PAGE, the IPG strips were incubated for 15 min with a solution
Frontiers in Physiology | www.frontiersin.org 2 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
of Tris-HCl buffer (pH 8.8), urea (6 M), glycerol (30%, v/v), SDS(2%, w/v), and DTT (2%, w/v). Strips were then equilibratedfor another 15 min in the same buffer containing iodoacetamide(2.5%, w/v) instead of DTT. The second-dimensional separationwas performed in 12% SDS-polyacrylamide gels. After proteinfixation, the gels were stained with colloidal Coomassie Blue,according to the manufacturer’s instructions. Molecular masseswere determined by running standard protein markers, coveringthe range 10–200 kDa. The pI values used were those given by thesupplier of the IPG strips.
Protein Visualization and Image AnalysisDigital images of the gels were recorded using a calibrateddensitometer (GS-800, Bio-Rad) and analyzed using PDQuestsoftware (Bio-Rad) (Silvestri et al., 2006). For each matchsetanalysis, maps corresponding to protein extracts from animalsof the same experimental group were organized into “ReplicateGroups” (each containing 4maps), named Ldlr−/−, Ldlr−/−+T2,and Ldlr−/−+T3. Statistical analysis was performed using aStudent’s t-test. Spots for which the p-value was <0.05 wereconsidered to display significant changes.
Protein Digestion and Mass SpectrometryAnalysisSpots from 2D-E were manually excised from gels, triturated, andwashed with water. Proteins were in-gel reduced, S-alkylated, anddigested with trypsin, as previously reported (D’Ambrosio et al.,2008). Protein digests were subjected to a desalting/concentrationstep on µZipTipC18 pipette tips (Millipore Corp., Bedford, MA,USA) and then analyzed by nano-liquid chromatography (nLC)-electrospray ionization (ESI)-linear ion trap (LIT)-tandem(MS/MS) mass spectrometry, using a LTQ XL mass spectrometer(Thermo Fischer Scientific, USA) equipped with a Proxeonnanospray source connected to an Easy-nanoLC (Proxeon,Odense, Denmark). Peptide mixtures were separated on an EasyC18 column (100 × 0.075 mm, 3 µm) (Thermo, USA) using agradient of acetonitrile containing 0.1% formic acid in aqueous0.1% formic acid; acetonitrile was ramped from 5 to 35% over 10min, from 35 to 95% over 2 min, and remained at 95% for 12min,at a flow rate of 300 nL/min. Spectra were acquired in the rangem/z 400–2000. Acquisition was controlled by a data-dependentproduct ion-scanning procedure over the 3 most abundant ions,enabling dynamic exclusion (repeat count 2; exclusion duration1 min). The mass isolation window and collision energy were setto m/z 3 and 35%, respectively.
Protein IdentificationMASCOT software package version 2.2.06 (Matrix Science, UK)(Cottrell, 2011) was used to identify spots unambiguously from aMusMusculus protein sequence database retrieved from UniProtrepository (76,058 sequences, 10/2011). Raw data from nanoLC-ESI-LIT-MS/MS were searched using a mass tolerance valueof 2 Da for precursor ion and 0.8 Da for MS/MS fragments,trypsin as proteolytic enzyme, a missed-cleavages maximumvalue of 2, and Cys carbamidomethylation and Met oxidation asfixed and variable modifications, respectively. Protein candidateswith more than 2 assigned peptide sequences, with MS/MS ion
score >30 and a peptide expectation value <0.05, were furtherevaluated by comparisonwith their calculatedmass and pI values,using the experimental values obtained from 2D-E.
In silico Biological AnalysisThe lists of differentially expressed proteins were input into theIPA platform (Ingenuity Systems, http://www.ingenuity.com) forthe identification of canonical pathways and functions differingbetween the treatments. The cutoff used was 1.5 for the foldchange and 0.05 for the p-value. Fisher’s exact test was used tocalculate a p-value indicating the probability that each biologicalfunction and/or disease assigned to that dataset might be soassigned due to chance alone. The results of Fisher’s exact testwere corrected for multiple testing using the false discovery rate(FDR). Comparison analysis was also performed to comparethe roles of the proteins among the generated lists. In order togenerate the reported networks, a list of differentially expressedproteins in the experimental conditions was overlaid onto aglobal molecular network developed from information containedin the Ingenuity Pathways Knowledge Base (IPKB). Networks ofthese focus-gene products were then algorithmically generatedon the basis of their connectivity. The IPA platform uses acurated database to construct functional regulatory networksfrom a list of individual proteins. To build networks, the programutilizes the IPKB containing large numbers of individuallymodeled relationships between proteins (obtained from theupgraded literature contained in IPA platform). The algorithmthen determines a statistical score for each network. This is doneby comparing the number of focus proteins that contribute to agiven network relative to the total number of occurrences of thoseproteins in all networks or pathways stored in the IPKB. Thena score is assigned to each network. The score is the negativelog of P, and it denotes the likelihood that the focus proteinsin the network might be found together by chance. Therefore,scores of 2 have at least 99% confidence of not being generatedby chance alone. In addition, the biological functions assignedto each network are ranked according to the significance of thatbiological function to the network.
Nuclei Preparation and WesternImmunoblot AnalysisAs already described in Cioffi et al. (2010), livers were dissectedand minced in ice-cold isolation buffer (consisting of 220 mMmannitol, 70 mM sucrose, 20 mM Tris-HCl, 1 mM EDTA, and5 mM EGTA, pH 7.4) and then were homogenized in a Potter-Elvehjem homogenizer. To isolate nuclei, the liver homogenatewas centrifuged at 500 g for 10 min at 4◦C. The obtainedpellet was subsequently resuspended and spun through a sucrosecushion (30% sucrose, 10 mM Tris-HCl, pH 7.5, 10 mM NaCl,and 3 mMMgCl2) at 1300 g for 10 min at 4◦C. The nuclear pelletwas washed with cold 10mMTris-HCl, pH 7.5, and 10mMNaCl.
For Western immunoblotting analysis, the nuclear pelletswere homogenized in lysis buffer containing 20mM Tris-HCl, pH 7.5, 150mM NaCl, 1mM EDTA, 1mM EGTA,2.5mM Na2H2P2O7, 1mM b-CH3H7O6PNa2, 1mM Na3VO4,1mM PMSF, 1mg/ml leupeptin, and 1% (w/v) Triton X-100by using an Optima TLX Ultraturrax (Beckman Coulter,
Frontiers in Physiology | www.frontiersin.org 3 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
Milan, Italy), then centrifuged at 13,400 g for 10 min at4◦C. Protein concentration was determined by using the BioRad’s DC method (Bio-Rad Laboratories, Hercules, CA). Thefollowing primary antibodies were used: anti-HNF4α (ab41,898,Abcam,mousemonoclonal); anti-PPARα (ab8934, Abcam, rabbitpolyclonal); anti-LXRα (ab41,902, Abcam, mouse monoclonal);anti-TRβ (GTX113278, GeneTex, rabbit polyclonal); anti-PGC-1α (AB3242, Merck Millipore, rabbit polyclonal); anti-α tubulin(ab4074, Abcam, rabbit polyclonal). Proteins were detected bya chemiluminescence protein-detection method based on theprotocol supplied with a commercially available kit (Millipore)and by using the appropriate secondary antibodies. Signals werequantified by means of a Bio-Rad ChemiDocTM XRS, usingdedicated software (QuantityOne, Bio-Rad Laboratories).
Serum Levels of Free T3 (FT3) and Free T4(FT4)The serum levels of FT3 and FT4 weremeasured by commerciallyavailable kits by means of immunoassay [materials and protocolssupplied by Byk-Sangtec Diagnostica (Dietzenbach, Germany)].
Statistical AnalysisData are expressed as the mean ± SD as indicated in thefigures. Comparisons between two groups were performedusing student’s t-test. Comparisons amongst three groupswere performed using one-way ANOVA and followed by theNewman-Keuls test. For all analyses, p < 0.05 was consideredstatistically significant.
RESULTS
Altered Liver Protein Expression ProfileInduced by T2 and T3Through a 2D-E-based proteomic approach, the hepaticpathways and the molecular mediators involved in the T2-and T3- induced reductions in circulating cholesterol levelsin Ldlr−/− mice were investigated. At the detection-limits set,the image software counted 450 matched proteins among thevarious maps in the liver (Figure 1A). Pair-wise comparisonswere performed to analyze the differential expression patternassociated with the T2 and T3 treatment (Ldlr−/−+T2 andLdlr−/−+T3 vs. Ldlr−/−). When the interest was limited toa differential expression of at least 2-fold and a statisticalsignificance of at least 95% (p < 0.05), 57 (about 12.4% oftotal entries), and 59 spots (about 12.8% of total entries)showed significant quantitative changes in liver following T2-and T3-treatment, respectively. Importantly, the differentialexpression produced by T2 and T3 overlapped on 33 proteinproducts (Figure 1B) corresponding to 40% of the totalamount of differentially expressed proteins (Figure 1E). Theremaining, specifically affected either by T2 (Figure 1C) orby T3 (Figure 1D), represented 29 and 31% of the totalamount of differentially expressed proteins (Figures 1F,G,respectively).
Considering that in a previous study (Goldberg et al., 2012) wedemonstrated amodulation exerted by T3 and T2 on intrahepaticcontent of apolipoproteins and that ApoE and ApoA1 are well
resolved on a standard 2D map of total soluble liver proteome,ApoE and ApoA1 were localized by means of map comparison(Fountoulakis et al., 2001) and subjected to quantitative analysis.ApoE, corresponding to spot 43 (Figure 1C), was significantlyaffected only by T2 with a differential expression of −40%vs. Ldlr−/− control levels (p < 0.1) (Figure 2). ApoA1,corresponding to spot 69 (Figure 1D), tended to be reduced inLdlr−/−+T3 mice by −40% vs. Ldlr−/− control levels, althoughwithout statistical significance (Figure 2). These two proteinstogether with other 67 spots among those automatically detectedby the matching software, were manually excised, trypsinyzed,and subjected to nanoLC-ESI-LIT-MS/MS analysis. Fifty-twoprotein spots were unambiguously identified (SupplementaryMaterial 1). Among these, 25 protein products were affected byboth T2 and T3, 9 only by T2, 18 only by T3.
Proteins detected as a mixture of components were excludedfrom further analyses and thus were not discussed further inthis report. When proteins were identified as multiple spotson the same map, putatively reflecting the occurrence of post-translational modifications, the pattern of changes was fairlysimilar among the various species.
Analysis of Potential Target Proteins of T2and T3Both T2 and T3 reduced expression of a number of hepaticproteins compared to the expression levels obtained forLdlr−/− control mice (91 and 79% of differentially expressedproteins, respectively). A significant increase produced by bothiodothyronines was observed for cellular retinol-binding protein1 also named CRBP1 (spot 1) and major urinary protein 1(MUP1) (spot 2), with the effect of T3 being more pronouncedthan that of T2 (Figure 3). Key metabolic enzymes wereamong the proteins decreased by both iodothyronines(Figure 4). Importantly, these enzymes are involved in majorhepatic processes such as amino acid metabolism [catecholO-methyltransferase (spot 4), ornithine aminotransferase,mitochondrial (spot 21) and histidine ammonia-lyase (spot 29)](Figure 4A), substrate metabolism [aldose 1-epimerase (spot 15),malate dehydrogenase (spot 17), and fructose-1,6-bisphosphatase1 (spot 19)] (Figure 4B), and cellular stress [lactoylglutathionelyase (spot 3), regucalcin (spot 13), mitochondrial aldehydedehydrogenase (spot 25)] (Figure 4C). Of note, these proteinsare regulated to the same extent by either T3 or T2, highlightinga common effect of the two iodothyronines.
T2 specifically decreased proteins such as fatty acid-bindingprotein (spot 31), succinyl-CoA ligase (spot 38), and glycerolkinase (spot 42), which are involved in lipid, oxidative, andcarbohydrate metabolism, respectively (Figure 5A). In contrast,glycerol-3-phosphate dehydrogenase (spot 68) and isocitratedehydrogenase [NAD] (spot 58), involved in oxidative andsubstrate metabolism, were significantly increased only by T3(Figure 5B). As a whole, these results suggest that, at thedoses used, T2 and T3, although modulating overlappingmetabolic events (e.g., amino acid and intra-mitochondrialenergy metabolism), also have their own specific moleculartargets.
Frontiers in Physiology | www.frontiersin.org 4 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
FIGURE 1 | Effects of T3 and T2 on the hepatic proteome in Ldlr−/− mice. (A) Representative 2D-E Coomassie Blue stained maps of total liver soluble proteins
extracted from Ldlr−/−, Ldlr−/−+T2, and Ldlr−/−+T3 mice and separated on 17 cm/pH 4–7 IPG strips in the first dimension and on 12% SDS-PAGE in the second.
Spotted and identified proteins among those with a density that differed significantly (by at least 0.5- or 2-fold; p < 0.05) between Ldlr−/− and either Ldlr−/−+T2 or
Ldlr−/−+T3 (blue spots) (B), Ldlr−/− and Ldlr−/−+T2 only (red spots) (C), and Ldlr−/− and Ldlr−/−+T3 only (green spots) (D). Overlapped circles represent the
total amount of differentially expressed proteins between the experimental groups. The blue area represents the percentage of proteins affected by both T2 and T3
(iodothyronines’ common proteomic effect in liver of Ldlr−/− mice) (E); the red area represents the percentage of proteins affected only by T2 treatment (F); the green
area represents the percentage of proteins affected only by T3 treatment (G).
IPA Analysis of Potential Target ProteinsInvolved in T2 and T3 EffectsTo further characterize the effects elicited by either T2-or T3-treatment in liver of Ldlr−/− mice, proteomic datawere analyzed by using the IPA platform that, based onknown interactions between affected proteins, defines commonfunctional and canonical pathways as well as protein networks,thereby offering additional information about the complexinteractive links between modulated proteins following thetreatments under study. The in silico analysis confirmedthat the most significant T2/T3- dependent changes alteredlipid-, amino acid-, carbohydrate-, and energy- metabolism(Supplementary Material 2). These changes are mediatedby effects on pathways such as glycolysis/gluconeogenesis,citrate cycle, pentose phosphate, glutathione, and amino acidmetabolism (Figures 6A,B). Overall, in terms of modulatedfunctions and pathways, T2 and T3 exerted a similar effect with a
few exceptions. Of particular metabolic relevance the peroxisomeproliferator-activated receptor α (PPARα)/retinoid X receptor α
(RXRα) pathway was affected only by T3 (Figure 6B).The protein network analysis for T2 produced the highest
scored node (the value being 28) corresponding to the hepatocytenuclear factor 4α (HNF4α), a nuclear receptor well knownto act as a master regulator of liver-specific gene expressionorchestrating lipid and cholesterol metabolism (Figure 7A).HNF4α is directly interconnected with some focus proteinsacquired in 2D-E analysis such as aldehyde dehydrogenase(ALDH2, ALDH1), malate dehydrogenase (MDH1), and fattyacid-binding protein (FABP), which are involved in substrateand lipid metabolism (Figure 7A). Strictly in line with thepathways analysis, as far as T3 effects are concerned, networktool revealed PPARα as the main hub of the highest scorednetwork (IPA score 30), a nuclear receptor that regulates diverseaspects of lipid metabolism, including fatty acid oxidation,
Frontiers in Physiology | www.frontiersin.org 5 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
FIGURE 2 | Effects of T3 and T2 on the intrahepatic content of ApoE
and ApoA1 in Ldlr−/− mice. Representative subsections of 2D-E images of
livers of Ldlr−/−, Ldlr−/−+T2, and Ldlr−/−+T3 mice are reported. Results
are expressed as arbitrary units (means ± SD; n = 4).
FIGURE 3 | T3- and T2-dependent differential expression in liver of
Ldlr−/− mice: increased proteins. Representative subsections of 2D-E
images of livers of Ldlr−/−, Ldlr−/−+T2, and Ldlr−/−+T3 mice. Results are
expressed as arbitrary units (means ± SD; n = 4) (p < 0.05).
and lipoprotein metabolism (Figure 7B). Within the network,PPARα directly interacts with some focus proteins acquiredin 2D-E analysis such as aldehyde dehydrogenase (ALDH2),mitochondrial glycerol-3-phosphate dehydrogenase (GPD2),maleylacetoacetate isomerase (GSTZ1), major urinary protein1 (MUP1), and peroxiredoxin-6 (PRDX6), all of which areinvolved in substrate and energymetabolism and in cellular stress(Figure 7B).
Nuclear Factors Affected By T2 and T3 inLiver of Ldlr−/− MiceConsidering that IPA network analysis highlighted HNF4α andPPARα as the highest scored nodes grouping the major numberof interactions with differentially regulated proteins by T2 andT3, respectively, Western blot analysis was performed to furtherinvestigate the putative involvement of such nuclear factors intothe hypocholesterolemic effects of both iodothyronines. Both T2and T3 administration to Ldlr−/− mice produced a significantreduction of the nuclear content of HNF4α, with the effectbeing slightly stronger in Ldlr−/−+T2 than in Ldlr−/−+T3 mice(Figures 8A,B). By contrast, only T3 administration produced apronounced and significant decrease of nuclear PPARα levels vs.those in control Ldlr−/− mice. According to what is suggested bythe in silico analysis, this result is consistent with the hypothesisthat PPARα has a specific role in the metabolic effects exerted byT3 in Ldlr−/− mice.
In view of the fact that a partnership of HNF4α withits coactivator, peroxisome proliferator-activated receptor γ
FIGURE 4 | T3- and T2-dependent differential expression in liver of
Ldlr−/− mice: decreased proteins. Representative subsections of 2D-E
images of livers of Ldlr−/−, Ldlr−/−+T2, and Ldlr−/−+T3 mice. (A) Enzymes
involved in amino acid metabolism. (B) Enzymes involved in substrate
metabolism. (C) Proteins involved in cellular stress. Results are expressed as
arbitrary units (means ± SD; n = 4) (p < 0.05).
coactivator-1α (PGC-1α) in the regulation of lipoproteinmetabolism has been characterized (Rhee et al., 2006), theeffect of T2 and T3 on the hepatic nuclear content of PGC-1αwas analyzed. As shown in Figures 8A,B, only T2 significantlyreduced expression levels of PGC-1α, hence suggesting animpaired PGC-1α/HNF4α-dependent signaling in the plasmacholesterol-lowering effects of T2.
Nuclear receptors other than HNF4α and PPARα alsoparticipate in the transcriptional regulation of key factorsinvolved in the intrahepatic cholesterol metabolism. Specifically,it has been reported that T3 exerts its hypocholesterolemiceffect mainly by binding to liver TRβ but, in the absenceof this, also acting through the liver X receptor α (LXRα)(Gullberg et al., 2000). Nuclear receptor expression levels maybe predictive for the activity of their ligands. In view of thisand considering that LXRα is a central hepatic cholesterolsensor, Western blot analysis was performed to verify whetherthe hypocholesterolemic effect of T2 and T3 in Ldlr−/− micecorrelates with LXRα and TRβ hepatic expression levels. Inparallel with the lower plasma cholesterol levels detectedin Ldlr−/−+T2 and Ldlr−/−+T3 mice (vs. Ldlr−/−), bothiodothyronines reduced LXRα nuclear content (Figures 8A,B).Of note, LXRα nuclear content tended to increase when Ldlr−/−
Frontiers in Physiology | www.frontiersin.org 6 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
FIGURE 5 | Iodothyronine specific effects on differential expression in
liver of Ldlr−/− mice. (A) Representative T2 specific affected proteins. (B)
Representative T3 specific affected proteins. Results are expressed as
arbitrary units (means ± SD; n = 4) (p < 0.05).
mice were consuming the WTD (Figures 8C,D). A significantreduction of TRβ levels (about −60% vs. Ldlr−/− control mice)was observed only in Ldlr−/−+T3 mice (Figures 8C,D).
Nuclear Factors Affected by T2 and T3 inLivers of WT MiceIn order to evaluate whether the absence/presence of LDLrmight be a major determinant of the effects of T2 and T3 onthe expression level of the aforementioned transcription factors,Western blot analyses were performed on liver tissue from WTmice fed either chow diet or WTD. The effects elicited by T2on HNF4α, PGC-1α, and LXRα levels in livers from Ldlr−/−
mice were not observed in WT mice fed chow or WTD while T3treatment, without significantly altering PPARα nuclear content,increased both HFN4α and PGC-1α hepatic levels, in WTD WTmice (Supplementary Material 3).
Finally, due to the large dose of T2 used, we sought to excludedisplacement of T3 from thyroid binding globulin as a reason forthe efficacy of the T2-treatment. To do this, we assessed serumFT3 and FT4 levels in T2 treated mice; neither was increased[the actual values being for FT3 (pg/ml): 8.55 ± 1.8; 3.2 ± 1.9∗;for FT4 (ng/dl): 1.24 ± 0.8; 0.15 ± 0.06∗, in WTD WT andWTD-WT+T2 animals, respectively; ∗P < 0.05 vs. WTD WT].Therefore, T2 did not displace T3 from the thyroid bindingglobulin. Thus, increased endogenous T3 does not account forthe effects of T2.
DISCUSSION
Although it has been hypothesized on the basis of in vitro studiesthat THs mediate lowering of LDL-C primarily by increasedhepatic expression of LDLr (Lopez et al., 2007) and that they
up regulate scavenger SR-B1 in mice (Johansson et al., 2005),very recently, we (Goldberg et al., 2012) and others (Lin et al.,2012) reported that THs can reduce serum cholesterol via a LDLr-independent mechanism. Ldlr−/− mice have modestly elevatedlevels of plasma cholesterol but when fed a WTD they developmuch higher levels of cholesterol and atherosclerosis. Whentreated with high doses of T3 or T2, WTD-fed Ldlr−/− miceshowed a dramatic decrease in LDL-C with these reductionsbeing linked to reductions in ApoB48 and ApoB100 secretion(Goldberg et al., 2012).
Utilizing 2D-E and MS, we studied a total proteome of 450liver proteins and identified 25 proteins affected by both T2 andT3, 18 only by T3 and 9 only by T2. At the used doses, both T2and T3 impact liver proteome producing a general decrease ofmodulated proteins.
When the differentially expressed proteins were mapped forthe identification of canonical pathways and biological functions(among which lipid, carbohydrate and amino acid metabolism),14 pathways/lists with the highest statistical significance wereidentified for both iodothyronines. By comparing these lists,3 non-overlapping pathways were identified for one of thetwo iodothyronines (e.g., of particular metabolic relevance,PPARα/RXRα activation for T3). This indicates the existence ofcommon proteomic targets for T2 and T3 as well as of specificones.
Of note, among the individual common proteins increasedby both iodothyronines, we identified MUP1. Although thephysiological functions of this low molecular weight secretedprotein belonging to the lipocalin family remain poorlyunderstood, recent evidence demonstrated that MUP1 could bepositively correlated with energy expenditure, metabolism, andinsulin sensitivity in mice (Hui et al., 2009). This might open newperspectives in understanding themechanisms through which T3and T2 determine the whole animal metabolic state (for recentreview see, Mullur et al., 2014; Goglia, 2015; Davis et al., 2016).
Key enzymes involved in amino acid- and substrate-metabolism and cellular stress were all individual commonproteins decreased by both iodothyronines. Specifically,the reduction of catechol O-methyltransferase, ornithineaminotransferase, and histidine ammonia-lyase indicates asuppressive effect elicited by both T2 and T3 on enzymesinvolved in amino acid degradation and ureagenesis, which isin line with the known effects of thyroid hormone on ammoniametabolism (Sochor et al., 1981; Marti et al., 1988). The reductionin the expression levels of lactoylglutathione lyase and aldehydedehydrogenase, on the other hand, indicates a lower content ofdetoxifying enzymes in the liver of Ldlr−/− mice treated witheither T2 or T3. Of note, consistent with what was previouslyreported (Silvestri et al., 2006), we observed an iodothyronine-induced decrease of the hepatic level of regucalcin, suggestinga role of calcium availability in the cellular effects of T2 and T3(Del Viscovo et al., 2012).
Only T3 increased the mitochondrial enzymes glycerol-3-phosphate dehydrogenase and isocitrate dehydrogenase.This suggests that T3, in Ldlr−/− mice, might stimulatemitochondrial respiration and substrate oxidation. Inparticular, glycerol-3-phosphate dehydrogenase is a
Frontiers in Physiology | www.frontiersin.org 7 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
FIGURE 6 | T3 and T2 affected canonical pathways in liver of Ldlr−/− mice: in silico analysis. The lists of differentially expressed proteins were input into the
IPA platform (Ingenuity® Systems, http://www.ingenuity.com) for functional enrichment analysis. Over-represented canonical pathways for Ldlr−/−+T2 (A) and
Ldlr−/−+T3 (B) vs. Ldlr−/− deregulated proteins are represented in barplots by their statistical score (negative logarithm to the base 10 of B-H corrected p-value)
with a threshold of 1.3 (yellow straight line). The ratio between deregulated and all proteins of a pathway is also reported (yellow not straight line).
well-established marker of liver thyroid status (Rauchováet al., 2011).
Analyzing the functional interaction between the identifieddifferentially expressed target proteins, the network tool revealedsub networks centered around HNF4α for T2 and around PPARα
for T3.HFN4α, an orphan member of the nuclear receptor
superfamily, is crucial for the maintenance and regulation ofhepatic lipid metabolism and it is a major in vivo regulator of theexpression of liver-specific genes involved in the production andclearance of apolipoproteins (Yin et al., 2011). Odom et al. (2004)reported that HFN4α directly binds to almost 50% of the activelytranscribed hepatic genes whereas no other transcription factorbinds to more than 1/5 of that number. In humans, Medica 16,an HFN4α antagonist, acts as a hypolipidemic agent likely due toreduced activation of HFN4α target gene expression (Hertz et al.,
2001; Sheena et al., 2005; Rufibach et al., 2006). The identificationofHFN4α as a central node in the proteomic effects of T2 suggeststhat this receptor could be involved in the hypocholesterolemiceffect elicited by T2 in Ldlr−/− mice. Indeed, accordingly with thedramatic reduction in circulating apoB-containing lipoproteins,nuclear HFN4α protein content was strongly decreased in liverfromWTD-Ldlr−/− mice following T2 treatment.
Interacting proteins are crucial in determining thetranscriptional activity of nuclear receptors. The PGC-1α/HFN4α partnership plays a crucial role in hepatic lipoproteinmetabolism (lipoprotein synthesis and export) (Rhee et al.,2006), and modulation of PGC-1α coactivation of HFN4α mayprovide a novel mechanism to manage dyslipidemia (Rhee et al.,2006). Nuclear hepatic PGC-1α content was reduced by T2,thus suggesting affected PGC-1α- HFN4α-dependent signalingin the plasma cholesterol-lowering effects of T2. This might
Frontiers in Physiology | www.frontiersin.org 8 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
FIGURE 7 | T3 and T2 affected protein networks in liver of Ldlr−/− mice. Network representation of the molecular relationships between identified, differentially
expressed proteins (Ingenuity Systems Ltd.) in liver of Ldlr−/−+T2 (A) and Ldlr−/−+T3 (B) vs. Ldlr−/− mice. Gene products are represented as nodes, and the
biological relationship between two nodes is represented as an edge (line). Indirect interactions appear as broken lines, whereas direct interactions appear as solid
lines. All edges shown are supported by at least 1 reference from the literature, from a textbook, or from canonical information stored in the Ingenuity Pathways
Knowledge Base. Human, mouse, and rat orthologues of a gene are stored as separate objects in IPKB, but are represented as a single node in the network. For
clarity, network shapes are shown.
be correlated with the decreased hepatic level of ApoE as wellas the decreased plasma levels of ApoB48 and ApoB100 anddecreased Srb1 mRNA levels (Goldberg et al., 2012). Whencompared with T2, T3 decreased HFN4α nuclear content whilenot affecting that of PGC-1α, likely indicating that the observedT2/T3-hypocholesterolemic effect depend on different molecularevents. Besides PGC-1α, another coactivator of HFN4α isFABP, which has been shown to structurally and functionallybind HNF4α to stimulate its transcriptional activity (McIntoshet al., 2013). The significantly lower level of FABP in liver fromWTD-Ldlr−/− mice only following T2 treatment is in line withthe decreased expression of HNF4α and its protein targets.Among these, of a particular interest could be the glycerol kinase(GK), an enzyme involved in supplying glucogenic precursorsfor hepatic glucose production. The GK promoter contains afunctional HNF4α binding site (Stepanian et al., 2003) and thehepatic content of this enzyme was significantly decreased onlyin T2-treated mice. By contrast, T3 treatment, while decreasingliver HNF4α levels, did not decrease those of FABP. Moreover,the lack of a significant effect of T3 on GK liver content might
suggest a differential impact of the two iodothyronines on hepaticglucose production and thus on the whole animal glycaemia.This would require further analyses above all considering thewell-known insulin antagonizing effect of T3 and the recentlydescribed insulin sensitizing action of T2 (de Lange et al., 2011;Moreno et al., 2011; Padron et al., 2014).
Reduction of PPARα levels was found only following T3treatment and it is in accordance with the lower levels of HDLand Srb1 mRNA as well as the increased hepatic triglyceridesecretion (Goldberg et al., 2012). Although PPARα activationby fibrates decreases ApoB-containing lipoproteins as well astotal serum ApoB levels in rodents and human (Peters et al.,1997; Milosavljevic et al., 2001), treatment of Ldlr−/− mice withciprofibrate markedly decreased plasma ApoB-48-carrying IDLand LDL but at the same time caused a marked accumulation ofApoB-100 carrying IDL/LDL, increased plasma cholesterol levelsand promoted aortic atherosclerosis (Fu et al., 2004). Our findingof a T3-induced decrease in PPARα levels associated with asignificant reduction in plasma cholesterol and ApoB-containinglipoproteins levels as well decreased hepatic production of
Frontiers in Physiology | www.frontiersin.org 9 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
FIGURE 8 | Effects of T3 and T2 on nuclear factors in liver of Ldlr −/− mice. (A) Representative Western blot analyses of HNF4α, PGC-1α, PPARα, and LXRα
protein levels in Ldlr−/−, Ldlr−/−+T2, and Ldlr−/−+T3 mice. Liver nuclear protein contents were normalized on α-tubulin. Protein load was 30 µg/lane. (B)
Quantitative analysis of protein expression levels. Results are expressed as arbitrary units (means ± SD; n = 3). Bars labeled with dissimilar letters are significantly
different (p < 0.05). (C) Representative Western blot analyses of LXRα and TRβ protein levels in Ldlr−/− mice fed with either a chow (Ldlr−/− C) or a western type diet
(Ldlr−/− WTD) and treated with T3 (Ldlr−/− WTD+T3). Liver nuclear protein contents were normalized on α-tubulin. Protein load was 30 µg/lane. The vertical lines
indicate cuts in the membranes to avoid to display samples not related to the used model. (D) Quantitative analysis of nuclear protein content of LXRα and TRβ in
Ldlr−/− C, Ldlr−/− WTD and Ldlr−/− WTD+T3 mice. Results are expressed as arbitrary units (means ± SD; n = 4). Bars labeled with dissimilar letters are
significantly different (p < 0.05).
ApoB-100, is in line with the above study and supports thecholesterol-lowering effect of T3 in a situation in which theIDL/LDL lipoprotein remnants cannot be efficiently cleared fromthe plasma via the LDLr pathway. In the context of our results,it is interesting to note that PPARα is a target of HNF4α(Pineda Torra et al., 2002; Martinez-Jimenez et al., 2010) andthat T3 by reducing HNF4α levels likely produces a downstreamnegative effect on PPARα which, indeed, could not be the casewith T2, that did not significantly reduce PPARα levels vs.controls. So although both T2 and T3 reduce HNF4α levels,likely they target different intrahepatic pathways all leading to thehypocholesterolemic effect in Ldlr−/− mice.
Whether specifically targeting HNF4α- and PPARα-downstream proteins will reproduce the LDL reductionfound with thyroid hormones is an area for future studies alsobased on in vitro approaches. Moreover, we should note that2D-E based proteomic analyses are intrinsically limited as theyresolve only soluble and highly expressed components of proteinmixtures, and as the detection of low and high molecular massor of basic and hydrophobic proteins is inefficient. Thus, otherfactors missing in our analysis could also be critical for thehypocholesterolemic effects of iodothyronines.
Besides the above mentioned nuclear receptors, LXRα, byregulating expression of several genes (Calkin and Tontonoz,
2010), controls cholesterol clearance via regulation of intestinalcholesterol absorption, biliary cholesterol secretion andcholesterol conversion into bile acids. Both T3 and T2 lowerednuclear LXRα protein content. One competitor receptor ofLXRα in mediating the hypocholesterolemic effect of T3, isTRβ (Gullberg et al., 2000). According to the hyperthyroidstate of the animals, hepatic nuclear content of TRβ wassignificantly decreased only in T3-treated mice. The lack of T2effects on TRβ levels, at the high dose of T2 used, consideringthe low, but not ignorable, affinity of T2 for TRβ, mightstimulate to further investigate how T2 might exert someof its transcriptomic/proteomic effects in a TR-dependentmanner (Mendoza et al., 2013; Navarrete-Ramírez et al., 2014;Orozco et al., 2014; Jonas et al., 2015), above all in view ofthe fact that, as already shown by Goldberg et al. (2012) inLdlr−/− mice, also in WT mice, the high used dose of T2significantly suppressed FT3 and FT4 levels. Moreover, as faras it concerns the differences between the actions exerted byT3 and T2, it has to be considered that only one dose of eachiodothyronine was used and T2 was administered at a muchhigher concentration than T3, so that it would be expectedthat differences in response, apart from involving differentmechanisms, might also reflect differences in the dose/responsecurve.
Frontiers in Physiology | www.frontiersin.org 10 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
T2-mediated effects on HNF4α, PGC-1α, and LXRα levelswere not observed in WT mice either when fed with Chow orWTD. On the other hand, T3 lead to increased HFN4α andPGC-1α levels inWTWTD fedmice. Thus, the absence/presenceof LDLr may be a major determinant of the effects that T2 and T3exert on the above nuclear factors.
In conclusion, although there are differences among mice,humans and rats regarding cholesterol hepatic metabolism, thisstudy, independently of the specific effects elicited by the highused doses of T2 and T3, furnishes novel information on LDLr-independent pathways and mediators that could be importanttargets for cholesterol lowering therapies.
AUTHOR CONTRIBUTIONS
MM designed the experimental approaches, supervised dataelaboration, wrote and revised the manuscript; ES designedthe experimental approaches, performed proteomic analyses,supervised data elaboration, wrote and revised the manuscript;MC performed proteomic analyses, elaborated data, wrote andrevised the manuscript; IG designed the experimental modeland approaches, supervised animal care and treatments, andrevised the manuscript; LH designed the experimental modeland approaches, supervised animal care and treatments and
revised the manuscript; AS performed MS analyses and revisedthe manuscript; FD performed in silico analyses and revised themanuscript; JE contributed to the design of the work and revisedthe manuscript; FG coordinated the experimental proceduresand revised the manuscript.
FUNDING
University of Sannio Research Grants.
ACKNOWLEDGMENTS
We thank Dr. Paul A. Dawson, Department of Pediatrics,Emory University, and Dr. Ryan E. Temel, Saha CardiovascularResearch Center, University of Kentucky, for critical readingof the manuscript. Dr. L-S Huang’s current affiliation is at theDivision of Cardiovascular Sciences, National Heart, Lung andBlood Institute, National Institutes of Health, Bethesda, MD.
SUPPLEMENTARY MATERIAL
The Supplementary Material for this article can be foundonline at: http://journal.frontiersin.org/article/10.3389/fphys.2016.00545/full#supplementary-material
REFERENCES
Angelin, B., and Rudling, M. (2010). Lipid lowering with thyroidhormone and thyromimetics. Curr. Opin. Lipidol. 21, 499–506. doi:10.1097/MOL.0b013e3283402e9c
Bakker, O., Hudig, F., Meijssen, S., and Wiersinga, W. M. (1998). Effectsof triiodothyronine and amiodarone on the promoter of the humanLDL receptor gene. Biochem. Biophys. Res. Commun. 249, 517–521. doi:10.1006/bbrc.1998.9174
Baxter, J. D., and Webb, P. (2009). Thyroid hormone mimetics: potentialapplications in atherosclerosis, obesity and type 2 diabetes. Nat. Rev. DrugDiscov. 8, 308–320. doi: 10.1038/nrd2830
Calkin, A. C., and Tontonoz, P. (2010). Liver X receptor signaling pathwaysand atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 30, 1513–1518. doi:10.1161/ATVBAHA.109.191197
Cioffi, F., Zambad, S. P., Chhipa, L., Senese, R., Busiello, R. A., Tuli, D.,et al. (2010). TRC150094, a novel functional analog of iodothyronines,reduces adiposity by increasing energy expenditure and fatty acid oxidationin rats receiving a high-fat diet. FASEB J. 24, 3451–3461. doi: 10.1096/fj.10-157115
Cottrell, J. S. (2011). Protein identification using MS/MS data. J. Proteomics 74,1842–1851. doi: 10.1016/j.jprot.2011.05.014
D’Ambrosio, C., Arena, S., Salzano, A. M., Renzone, G., Ledda, L., and Scaloni, A.(2008). A proteomic characterization of water buffalo milk fractions describingPTM of major species and the identification of minor components involved innutrient delivery and defence against pathogens. Proteomics 8, 3657–3666. doi:10.1002/pmic.200701148
Davidson, N. O., Carlos, R. C., Drewek, M. J., and Parmer, T. G. (1988).Apolipoprotein gene expression in the rat is regulated in a tissue-specificmanner by thyroid hormone. J. Lipid Res. 29, 1511–1522.
Davidson, N. O., Carlos, R. C., Sherman, H. L., and Hay, R. V. (1990). Modulationof apolipoprotein B-100 mRNA editing: effects on hepatic very low densitylipoprotein assembly and intracellular apoB distribution in the rat. J. Lipid Res.31, 899–908.
Davis, P. J., Goglia, F., and Leonard, J. L. (2016). Nongenomic actions of thyroidhormone. Nat. Rev. Endocrinol. 12, 111–121. doi: 10.1038/nrendo.2015.205
de Lange, P., Cioffi, F., Senese, R., Moreno, M., Lombardi, A., Silvestri, E.,et al. (2011). Nonthyrotoxic prevention of diet-induced insulin resistance by3,5-diiodo-L-thyronine in rats.Diabetes 60, 2730–2739. doi: 10.2337/db11-0207
Del Viscovo, A., Secondo, A., Esposito, A., Goglia, F., Moreno,M., and Canzoniero,L. M. (2012). Intracellular and plasma membrane-initiated pathways involvedin the [Ca2+]I elevations induced by iodothyronines (T3 and T2) inpituitary GH3 cells. Am. J. Physiol. Endocrinol. Metab. 302, E1419–E1430. doi:10.1152/ajpendo.00389.2011
Erion, M. D., Cable, E. E., Ito, B. R., Jiang, H., Fujitaki, J. M., Finn, P. D., et al.(2007). Targeting thyroid hormone receptor-β agonists to the liver reducescholesterol and triglycerides and improves the therapeutic index. Proc. Natl.Acad. Sci. U.S.A. 104, 15490–15495. doi: 10.1073/pnas.0702759104
Fountoulakis, M., Juranville, J. F., Berndt, P., Langen, H., and Suter, L.(2001). Two-dimensional database of mouse liver proteins. An update.Electrophoresis 22, 1747–1763. doi: 10.1002/1522-2683(200105)22:9<1747::AID-ELPS1747>3.0.CO;2-H
Fu, T., Mukhopadhyay, D., Davidson, N. O., and Borensztajn, J. (2004).The peroxisome proliferator-activated receptor α (PPARα) agonistciprofibrate inhibits apolipoprotein B mRNA editing in low densitylipoprotein receptor-deficient mice: effects on plasma lipoproteins andthe development of atherosclerotic lesions. J. Biol. Chem. 279, 28662–28669.doi: 10.1074/jbc.M403271200
Goglia, F. (2015). The effects of 3,5-diiodothyronine on energy balance. Front.Physiol. 5:528. doi: 10.3389/fphys.2014.00528
Goldberg, I. J., Huang, L. S., Huggins, L. A., Yu, S., Nagareddy, P. R., Scanlan, T.S., et al. (2012). Thyroid hormone reduces cholesterol via a non-LDL receptor-mediated pathway. Endocrinology 153, 5143–5149. doi: 10.1210/en.2012-1572
Gullberg, H., Rudling, M., Forrest, D., Angelin, B., and Vennström, B. (2000).Thyroid hormone receptor β-deficient mice show complete loss of the normalcholesterol 7α-hydroxylase (CYP7A) response to thyroid hormone but displayenhanced resistance to dietary cholesterol.Mol. Endocrinol. 14, 1739–1749. doi:10.1210/mend.14.11.0548
Hertz, R., Sheena, V., Kalderon, B., Berman, I., and Bar-Tana, J. (2001).Suppression of hepatocyte nuclear factor-4α by acyl-CoA thioesters ofhypolipidemic peroxisome proliferators. Biochem. Pharmacol. 61, 1057–1062.doi: 10.1016/S0006-2952(01)00578-0
Frontiers in Physiology | www.frontiersin.org 11 November 2016 | Volume 7 | Article 545

Moreno et al. Iodothyronines and LDL-R-Independent Hepatic Pathways
Hui, X., Zhu, W., Wang, Y., Lam, K. S., Zhang, J., Wu, D., et al. (2009).Major urinary protein-1 increases energy expenditure and improves glucoseintolerance through enhancing mitochondrial function in skeletal muscleof diabetic mice. J. Biol. Chem. 284, 14050–14057. doi: 10.1074/jbc.M109.001107
Johansson, L., Rudling, M., Scanlan, T. S., Lundåsen, T., Webb, P., Baxter, J., et al.(2005). Selective thyroid receptor modulation by GC-1 reduces serum lipidsand stimulates steps of reverse cholesterol transport in euthyroid mice. Proc.Natl. Acad. Sci. U.S.A. 102, 10297–10302. doi: 10.1073/pnas.0504379102
Jonas, W., Lietzow, J., Wohlgemuth, F., Hoefig, C. S., Wiedmer, P., Schweizer,U., et al. (2015). 3,5-Diiodo-L-thyronine (3,5-T2) exerts thyromimetic effectson hypothalamus-pituitary-thyroid axis, body composition, and energymetabolism in male diet-induced obese mice. Endocrinology 156, 389–399. doi:10.1210/en.2014-1604
Klein, I., and Danzi, S. (2008). Thyroid disease and the heart. Circulation 117:e18.doi: 10.1161/CIRCULATIONAHA.106.678326
Ladenson, P. W., Kristensen, J. D., Ridgway, E. C., Olsson, A. G., Carlsson,B., Klein, I., et al. (2010). Use of the thyroid hormone analogueeprotirome in statin-treated dyslipidemia. N.Engl. J. Med. 362, 906–916. doi:10.1056/NEJMoa0905633
Lin, J. Z., Martagón, A. J., Hsueh, W. A., Baxter, J. D., Gustafsson, J. Å.,Webb, P., et al. (2012). Thyroid hormone receptor agonists reduce serumcholesterol independent of the LDL receptor. Endocrinology 153, 6136–6144.doi: 10.1210/en.2011-2081
Lopez, D., Abisambra Socarrás, J. F., Bedi, M., and Ness, G. C. (2007). Activationof the hepatic LDL receptor promoter by thyroid hormone. Biochim. Biophys.
Acta 1771, 1216–1225. doi: 10.1016/j.bbalip.2007.05.001Marti, J., Portoles, M., Jimenez-Nacher, I., Cabo, J., and Jorda, A. (1988). Effect
of thyroid hormones on urea biosynthesis and related processes in rat liver.Endocrinology 123, 2167–2174. doi: 10.1210/endo-123-5-2167
Martinez-Jimenez, C. P., Kyrmizi, I., Cardot, P., Gonzales, F. J., and Talianidis,I. (2010). Hepatocyte nuclear factor 4α coordinates a transcription networkregulating hepatic fatty acid metabolism. Mol. Cell. Biol. 30, 565–577. doi:10.1128/MCB.00927-09
McIntosh, A. L., Petrescu, A. D., Hostetler, H. A., Kier, A. B., and Schroeder, F.(2013). Liver-type fatty acid binding protein interacts with hepatocyte nuclearfactor 4α. FEBS Lett. 587, 3787–3791. doi: 10.1016/j.febslet.2013.09.043
Mendoza, A., Navarrete-Ramírez, P., Hernández-Puga, G., Villalobos, P., Holzer,G., Renaud, J. P., et al. (2013). 3,5-T2 is an alternative ligand for the thyroidhormone receptor β1. Endocrinology 154, 2948–2958. doi: 10.1210/en.2013-1030
Milosavljevic, D., Griglio, S., Le Naour, G., and Chapman, M. J. (2001).Preferential reduction of very low density lipoprotein-1 particle number byfenofibrate in type IIB hyperlipidemia: consequences for lipid accumulationin human monocyte-derived macrophages. Atherosclerosis 155, 251–260. doi:10.1016/S0021-9150(00)00634-1
Moreno, M., Silvestri, E., De Matteis, R., de Lange, P., Lombardi, A., Glinni, D.,et al. (2011). 3,5-Diiodo-L-thyronine prevents high-fat-diet-induced insulinresistance in rat skeletal muscle through metabolic and structural adaptations.FASEB J. 25, 3312–3324. doi: 10.1096/fj.11-181982
Mullur, R., Liu, Y. Y., and Brent, G. A. (2014). Thyroid hormone regulation ofmetabolism. Physiol. Rev. 94, 355–382. doi: 10.1152/physrev.00030.2013
Naoumova, R. P., Thompson, G. R., and Soutar, A. K. (2004). Current managementof severe homozygous hypercholesterolemia. Curr. Opin. Lipidol. 15, 413–422.doi: 10.1097/01.mol.0000137222.23784.2a
Navarrete-Ramírez, P., Luna, M., Valverde,-R., C., and Orozco, A. (2014). 3,5-di-iodothyronine stimulates tilapia growth through an alternate isoform of thyroidhormone receptor β1. J. Mol. Endocrinol. 52, 1–9. doi: 10.1530/JME-13-0145
Ness, G. C., and Lopez, D. (1995). Transcriptional regulation of rat hepaticlow-density lipoprotein receptor and cholesterol 7 α hydroxylase by thyroidhormone. Arch. Biochem. Biophys. 323, 404–408. doi: 10.1006/abbi.1995.0061
Odom, D. T., Zizlsperger, N., Gordon, D. B., Bell, G. W., Rinaldi, N. J., Murray,H. L., et al. (2004). Control of pancreas and liver gene expression by HNFtranscription factors. Science 303, 1378–1381. doi: 10.1126/science.1089769
Orozco, A., Navarrete-Ramírez, P., Olvera, A., and García,-G., C. (2014).3,5-diiodothyronine (T2) is on a role. A new hormone in search of recognition.Gen. Comp. Endocrinol. 203, 174–180. doi: 10.1016/j.ygcen.2014.02.014
Padron, A. S., Neto, R. A., Pantaleão, T. U., de Souza dos Santos, M.C., Araujo, R. L., de Andrade, B. M., et al. (2014). Administration of
3,5-diiodothyronine (3,5-T2) causes central hypothyroidism and stimulatesthyroid-sensitive tissues. J. Endocrinol. 221, 415–427. doi: 10.1530/JOE-13-0502
Pedrelli, M., Pramfalk, C., and Parini, P. (2010). Thyroid hormones and thyroidhormone receptors: effects of thyromimetics on reverse cholesterol transport.World J. Gastroenterol. 16, 5958–5964. doi: 10.3748/wjg.v16.i47.5958
Peters, J. M., Hennuyer, N., Staels, B., Fruchart, J. C., Fievet, C., Gonzalez, F. J.,et al. (1997). Alterations in lipoprotein metabolism in peroxisome proliferator-activated receptor α-deficient mice. J. Biol. Chem. 272, 27307–27312. doi:10.1074/jbc.272.43.27307
Pineda Torra, I., Jamshidi, Y., Flavell, D. M., Fruchart, J. C., and Staels, B.(2002). Characterization of the human PPARα promoter: identification of afunctional nuclear receptor response element.Mol. Endocronol. 16, 1013–1028.doi: 10.1210/mend.16.5.0833
Pramfalk, C., Pedrelli, M., and Parini, P. (2011). Role of thyroid receptorβ in lipid metabolism. Biochim. Biophys. Acta 1812, 929–937. doi:10.1016/j.bbadis.2010.12.019
Rauchová, H., Mrácek, T., Novák, P., Vokurková, M., and Soukup, T. (2011).Glycerol-3-phosphate dehydrogenase expression and oxygen consumption inliver mitochondria of female and male rats with chronic alteration of thyroidstatus. Horm. Metab. Res. 43, 43–47. doi: 10.1055/s-0030-1265220
Rhee, J., Ge, H., Yang, W., Fan, M., Handschin, C., Cooper, M., et al.(2006). Partnership of PGC-1α and HNF4α in the regulation of lipoproteinmetabolism. J. Biol. Chem. 281, 14683–14690. doi: 10.1074/jbc.M512636200
Rufibach, L. E., Duncan, S. A., Battle, M., and Deeb, S. S. (2006). Transcriptionalregulation of the human hepatic lipase (LIPC) gene promoter. J. Lipid Res. 47,1463–1477. doi: 10.1194/jlr.M600082-JLR200
Scheiffele, E., and Schultze, K. W. (1972). The coronary drug project. JAMA
221:918. doi: 10.1001/jama.1972.03200210062030Sheena, V., Hertz, R., Nousbeck, J., Berman, I., Magenheim, J., and Bar-Tana, J.
(2005). Transcriptional regulation of human microsomal triglyceride transferprotein by hepatocyte nuclear factor-4α. J. Lipid Res. 46, 328–341. doi:10.1194/jlr.M400371-JLR200
Silvestri, E., Moreno, M., Schiavo, L., de Lange, P., Lombardi, A., Chambery, A.,et al. (2006). A proteomics approach to identify protein expression changes inrat liver following administration of 3,5,3’-triiodo-L-thyronine. J. Proteome Res.
5, 2317–2327. doi: 10.1021/pr060141lSjouke, B., Langslet, G., Ceska, R., Nicholls, S. J., and Nissen, S. E., Öhlander, M.,
et al. (2014). Eprotirome in patients with familial hypercholesterolaemia (theAKKA trial): a randomised, double-blind, placebo-controlled phase 3 study.Lancet Diab. Endocrinol. 2, 455–463. doi: 10.1016/S2213-8587(14)70006-3
Sochor, M., McLean, P., Brown, J., and Greenbaum, A. L. (1981). Regulation ofpathways of ornithine metabolism. Effects of thyroid hormone and diabetes onthe activity of enzymes at the “ornithine crossroads’ in rat liver. Enzyme 26,15–23.
Stepanian, S. V., Huyn, S. T., McCabe, E. R., and Dipple, K. M. (2003).Characterization of the human glycerol kinase promoter: identification of afunctional HNF-4α binding site and evidence for transcriptional activation.Mol. Genet. Metab. 80, 412–418. doi: 10.1016/j.ymgme.2003.10.003
Tancevski, I., Demetz, E., Eller, P., Duwensee, K., Hoefer, J., Heim, C., et al.(2010). The liver-selective thyromimetic T-0681 influences reverse cholesteroltransport and atherosclerosis development in mice. PLoS ONE 5:e8722. doi:10.1371/journal.pone.0008722
Yin, L., Ma, H., Ge, X., Edwards, P. A., and Zhang, Y. (2011). Hepatichepatocyte nuclear factor 4α is essential for maintaining triglyceride andcholesterol homeostasis. Arterioscler. Thromb. Vasc. Biol. 31, 328–336. doi:10.1161/ATVBAHA.110.217828
Conflict of Interest Statement: The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest.
Copyright © 2016 Moreno, Silvestri, Coppola, Goldberg, Huang, Salzano, D’Angelo,
Ehrenkranz and Goglia. This is an open-access article distributed under the terms
of the Creative Commons Attribution License (CC BY). The use, distribution or
reproduction in other forums is permitted, provided the original author(s) or licensor
are credited and that the original publication in this journal is cited, in accordance
with accepted academic practice. No use, distribution or reproduction is permitted
which does not comply with these terms.
Frontiers in Physiology | www.frontiersin.org 12 November 2016 | Volume 7 | Article 545