Embed Size (px)
Citation preview

7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 1/8
Signaling R eactions I nduced in H uma n Fibroblasts D uringA dhesion to C ementum-D erived A tta chment P rotein
MASAHIRO SAITO and A. SAMPATH NARAYANAN
ABSTRACT
Cementum-derived attachment protein (CAP) is a M r 56,000 collagenous protein which promotes the adhesion and
spreading of mesenchymal cell types. The CAP promotes the adhesion of osteoblasts and periodontal ligament cells
better than gingival fibroblasts, while epithelial cells do not adhere to CAP-coated surfaces. To understand the
mechanisms involved in CAP action, we have studied the signal transduction events induced by the CAP in human
fibroblasts during cell adhesion. Human gingival fibroblasts were serum starved for 48 h, trypsinized, and addedto non–tissue culture plastic plates previously coated with CAP. At various time points, attached cells were
examined for induction of signaling reactions. Adherence of cells to plates coated with CAP caused tyrosine
phosphorylation of proteins migrating on PAGE with molecular mass of 125–130, 85, 70, and 42–44 kDa. We
identified focal adhesion kinase p125FAK and p130Cas as components of the 125–130 kDa protein band; however,
p125FAK was the major phosphorylated component. ERK-1 and ERK-2 were detected in the 42–44 kDa protein
band, but only the ERK-2, not ERK-1, was phosphorylated. Adhesion to CAP-stimulated mitogen-activated
protein kinase (MAPK) activity and induced the expression of c-fos mRNA. Protein-tyrosine phosphorylation and
c-fos mRNA expression were not induced in unattached cells, and adhesion was not abolished by the protein
tyrosine kinase inhibitor, genestein. MAPK activity and c-fos mRNA expression were not induced in monolayer
cultures, indicating that these reactions are induced by adhesion and not necessary for cell adhesion. The kinetics
of MAPK activation were different from cells attaching on fibronectin (FN) or polylysine, and c-fos mRNA levels
increased only half as much on FN and very little on polylysine. These data demonstrated that CAP and other
adhesion molecules present in mineralized tissue matrices induce characteristic signaling events during adhesion,which may play a role in recruitment of specific cell types during wound healing and in mediating their specific
biological functions. (J Bone Miner Res 1999;14:65–72)
INTRODUCTION
C E LL ADH E S I O N I S an essential requirement for assembly
of cells into a three-dimensional ma trix, and adhesion
interactions between cells and the extracellular matrix regu-
late many cellular functions, including cell migration, d ivi-
sion, and differentiation.(1–4) Most mamma lian cells cannot
progress through the G 1 phase of the cell cycle and com-
plete cell division unless they are a dhered to a substratum;
this property of ‘‘anchorage dependence’’ is lost in cancer
cells.(5,6) Several matrix components serve as substrates for
cell adhesion; prominent among these are collagens, fibro-
nectin (FN), laminin, bone sialoprotein, and osteopontin.
Cells bind to these molecules through specific integrin re-
ceptors, and the b inding induces Na/H antipor ter, phospha-
tidyl inositol turnover, protein kinase C activity, protein-
tyrosine phosphorylation, transcription o f c-fos , NF-B
expression, and many other signaling events.(7–10) Protein
tyrosine phosphorylation is one of the ea rliest b iochemical
reactions detected, and the phosphorylated proteins include
focal adhesion kinase (p125FAK , FAK), Crk-associated ty-
rosine kinase p130Cas (Cas), paxillin, tensin, and mitogen-
activated protein kinase (MAPK).(9,11–17)
Cell adhesion is a key biological process associated with
wound healing. Healing of wounds proceeds through a se-
ries of distinct but overlapping phases in which different
matrix components and cell types participate, and the
course of healing events are regulated by interactions be-
D epartment of P athology, U niversity of Washington School of Medicine, Seattle, Washington, U.S.A.
JOURNAL OF BONE AND MINERAL RESEARCHVolume 14, Number 1, 1999
Blackwell Science, Inc.© 1999 American Society for Bone and Mineral Research
65

7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 2/8
tween many cell types, polypeptide mediators, and matrix
components.(18,19) Participation of appropriate cell types is
the crucial fa ctor that determines whether healing occurs by
repair or regeneration. Adhesion molecules as well as an-
tiadhesive proteins present in the wound matrix may par-
ticipate in this process; for example, while FN and collagens
promote the adhesion of several cell types, laminin is spe-
cific to certain cells and C1q and thrombospondin are an-
tiadhesive. That adhesion molecules may affect the courseof w ound healing and determine whether healing occurs by
regeneration or repair is illustrated by the healing of peri-
odonta l connective tissues. These tissues are destroyed by
periodontitis, a chronic inflammatory disease, and regen-
eration requires formation of new connective tissue and
bone a nd restoration of the connective tissue atta chment to
tooth surfaces. However, the most common outcome is re-
pair by an epithelial, instead of connective tissue, attach-
ment.(20,21) This is because diseased tooth surfaces cannot
promote the a ttachment or growth of fibroblasts.(22,23)
We recently reported that the cementum, which is the
calcified outer layer of teeth through which collagen fibers
of connective tissues are anchored onto tooth root surfaces,
contains a M r 56,000 collagenous protein, which promotest h e a d h e si o n a n d s pr e a d i ng o f m e se n ch y m a l c e l l
types.(24,25) The distribution of this protein, cementum-
derived attachment protein (CAP ), appears to be limited to
cementum.(26) The CAP promotes cell adhesion at least as
well as FN, and among the periodontal cells, alveolar bone
cells and fibroblast subpopulations bind and adhere to
CAP-coated surfaces with different affinities.(24,27) In con-
trast, epithelial cells do not adhere to CAP.(24) The CAP
binding to periodontal fibroblast populations appears to be
related to their ability to d ifferentiate into osteoblastic phe-
notype.(28) To understand the molecular basis for the bio-
logical functions mediated by the CA P, we ha ve studied the
signaling events transduced by this protein during cell ad-
hesion. O ur objectives were to identify biochemical signal-ing reactions associated with adhesion to CAP, to deter-
mine if the activated events contribute to adhesion, and to
determine if these events are the same a s those induced by
other adhesion molecules. Fo r t hese experiments, we used
gingival fibrobla sts in which we have previously characater-
ized signaling events induced by growth factors.(29) O ur
results show that specific signaling events are induced dur-
ing adhesion to C AP and that t he magnitude and kinetics of
their induction differ from those induced by other ad hesion
molecules.
MATERIALS AND METHODS
M aterials
Fibroblasts used for these studies were established from
human gingival explants and maintained as described pre-
viously.(24,25) The following antibodies were used: mono-
clonal antibodies against phosphotyrosine (PY-20; Trans-
duction Laboratory, Lexington, KY, U.S.A.) and FAK (2
A7, Upstate B iotechnology Inc., Lake Placid, NY, U .S.A.),
polyclonal antibodies a gainst FAK (C-903), Ca s (C -20;
Santa Cruz Biochemistry, Santa Cruz, CA, U.S.A.), and
MAPK (7884; generous gift from D r. Jean Campbell, D e-
partment of Biochemistry and Howard Hughes Institute,
U niversity o f Washington). Hea t-inactivated bo vine serum
albumin (BSA ) fraction V, sodium orthovanadat e, genistin,
and soybean trypsin inhibitor were purchased from Sigma
(St. Louis, MO, U.S.A.) and genistein from LC Laborato-
ries (Woburn, MA, U .S.A.). [-32P]ATP and [-32P]dCTP
were the products of D uPont NEN (Boston, MA, U .S.A.).
c-fos cDNA was a generous gift from D r. D avid Morris,D epartment of B iochemistry, U niversity of Washington.
Protein assay kit a nd TRIzol R NA extraction kit w ere the
products of Bio-Rad (Hercules, CA, U.S.A.) and G IBCO
B RL (G aithersburg, MD , U .S.A.), respectively. All other
chemicals and reagents were of analytical grade and ob-
tained from Sigma C hemical Co.
Purification of CAP
Bovine cementum was harvested from the teeth of ani-
mals obtained from a slaughter house and extracted in 1.0
M C H 3COOH containing 25 mM EDTA, 1 mM each of
phenylmethylsulfonyl fluoride and N-ethylmaleimide, and 1
g/ml each of pepstatin and leupeptin, followed by 4 M
guanidine H Cl, pH 7.5, containing proteinase inhibitors.
The guanidine extract was subjected to diethylaminoethyl
(D EA E)-cellulose chromatography, and proteins eluted
with 0.5 M NaC l were separated by C 18 reversed-phase high
performance liquid chromatography.(25) CAP was eluted at
34–36% CH 3CN. I t migrated as a single protein band with
M r 56,000, and amino acid sequence information for CAP
peptides have been reported elsewhere.(25) The CA P
preparation promoted fibroblast attachment at an optimal
concentration of 0.5–2.0 g/ml. We have previously show n
by Western analysis that the CAP preparations obtained in
this manner did not contain other adhesion proteins, FN,
type I, III, V, or FACI T collagens, osteopontin, and bonesialoprotein. (25) Preparations used for experiments de-
scribed here were also checked for the ab sence of contami-
nation with adhesion proteins specified above.
A ssay fo r cell adhesion and pr eparation o f
cell lysates
Non-tissue culture–treated plastic Petri dishes were
coated with a dhesive ligands as d escribed previously by in-
cubating CAP (2.5 g/ml), FN (5 g/ml), o r po ly-D -lysine
(PL; 5 g/ml) for 2 h at 37° C a nd then blo cked with 2 mg/ml
of B SA in phosphate-buffered saline. These concentrations
were chosen based on concentration curves and they pro-
moted optimal a ttachment. Fibroblasts were serum-starvedfor 24 h, trypsinized, and plated on dishes coated with ad-
hesive ligands. After incubation a t 37° C for 1 h o r specified
periods, unattached cells were removed, and attached cells
were processed as needed. For adhesion assays [35S]methio-
nine-labeled cells were used, and atta chment wa s measured
as described previously.(24–26) For other experiments, 1.5 ×
106 cells were added to 100-mm Petri plates. At the indi-
cated time, unattached cells were removed, and attached
cells were lysed in 1 ml of 1% Triton X-100, 150 mM N aC l,
SAITO AND NARAYANAN66

7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 3/8
10 mM Tris, pH 7.4 buffer, 1 mM EDTA, 1 mM EG TA,
0.5% NP-40, 0.1 mM sodium orthovanadate, 1 mM phenyl-
methylsulfonyl fluoride, 1 mM b enzamidine, 10 g/ml a pro -
tinin, and 10 g/ml leupeptin. A fter 30 minutes of incuba-
tion on ice, the cell lysates were disrupted by sonication and
cleared by centrifugation at 12,000 rpm for 15 minutes.
When unattached cells were needed, the medium contain-
ing these cells was collected, cells pelleted by centrifuga-
tion, and lysates prepared. The protein concentration o f thelysates was determined by the B radford procedure using a
Bio-Rad protein assay kit. To study the effect of kinase
inhibitors, cells were preincubated for 2 h with the inhibi-
tors prior to trypsinization. E ach experiment was per-
formed in triplicate and repeated at least three times, and
data from one representative experiment are presented.
Immunoprecipitation
Cell lysates were incubated in anti-FAK (C-903) or anti-
Cas antibodies for 3 h in ice and then with protein A beads
for 1 h. The beads were collected by centrifugation, wa shed
thrice in lysis buffer, and boiled in SD S-PAG E sample
buffer. Proteins were separated on 8% gel slabs.
Immunoblotting
Total cell lysates or immunoprecipitated proteins were
separated by 8% SD S-PAG E under reducing conditions
and electrotransferred o nto Immobilon-NC nitrocellulose
membranes (Millipore Corp., B edford, MA, U .S.A.). The
membranes were blocked with 1% BSA in T-TBS (0.1%
Tween 20, 50 mM Tris, pH 7.6, 150 mM NaCl) for 2 h and
5% nonfat milk for 1 h, respectively, for anti-FAK and
MAPK antibodies. The membranes were probed with 1:
1000 diluted primary a ntibody in respective b locking buff-
ers for 1 h and then w ith horseradish peroxidase-linkedsecondary antibody and developed for enhanced chemilu-
minescence. In some cases, bound antibodies were stripped
from the membranes in buffer containing 0.1 M glycine, pH
2.5, for 30 minutes, and reprobed with desired second an-
tibody. D ata from one representative experiment are pre-
sented.
G el-shift assay for M A PK
Fifty micrograms of total cell lysates wa s electrophoresed
onto a 12% SD S-polyacrylamide gel, electroblotted onto
nitrocellulose membrane, and incubated with 1:1000 dilu-
tion of anti-MAPK antibody. Proteins were identified by
Western analysis.
I n-gel myelin basic protein ki nase assay
Cell lysates (20 g) were separated by 10% SD S-PA G E
in gels conta ining 0.4 mg/ml of myelin ba sic protein (MB P).
The gel was washed twice in buffer A (50 mM HEPES and
5 mM 2-mercaptoethanol) containing 20% 2-propanol for 1
h to remove SDS and then denatured by treating twice in
buffer A containing 6 M guanidine-H Cl for 1 h. The gel was
renatured with two overnight changes of buffer A contain-
ing 0.04% Tween 20 at 4° C. A fter preincuba tion in buffer B
(50 mM HEPES, 100 M sodium orthovanadate, 10 mM
magnesium chloride, and 5 mM 2-mercaptoethanol) fo r 30
minutes at 30° C, phosphorylation assay of MB P w as carried
out by placing the gel in buffer B containing 50 M ATP
and 50 C i [-32P]ATP for 1 h at 30° C. R eaction wa s ter-
minated b y placing the gels in 5% trichloroacetic acid with
10 mM Na4P2O7, and the gels were washed to remove un-reacted [32P]ATP. The gels were then dried and subjected
to autoradiography. To quantitate the kinase activity, the
MAPK band was subjected to computerized densitometry
using National Institutes of Health image software analysis
(NIH, B ethesda, MD , U .S.A.).
Northern blotting
Total R NA was isolated by TR Izol reagent, a nd 15–20g
was separated onto 1% formaldehyde agarose gels. A fter
transferring to a nylon filter, it was hybridized with 32P -
labeled D NA probes as described previously.(29) The
probes were a 915 bp E co R I–Sph I fragment encoding c-fos
cD NA, or an oligonucleotide encoding 28S ribosomal R NA
(5-G CG A G AG C G CC AG C TATCCTG AG G -3).
RESULTS
A dhesion to C A P induces protein
tyrosine phosphorylation
Tyrosine phosphorylation of proteins such as FAK is one
of the early signaling events induced by adhesion(8,10);
therefore, we examined w hether C AP catalyzes tyrosine
phosphorylation of the FA K and other cellular proteins.
Human gingival fibroblasts were plated on CAP-coated
dishes for 1 h, nonadherent cells were removed, a nd lysateswere prepared. Lysates were also prepared from nonadher-
ent cells, which were o btained b y centrifugation of medium
containing unattached cells. Lysates were also prepared
from cells in monolayer to which 2.5 g/ml CA P was a dded
in solution. Western a nalysis of the lysates using a ntiphos-
photyrosine antibody revealed that cells attaching to the
CAP-coated dishes contained several protein bands cross-
reacting with the antibody. Among these, at least four
ba nds migra ting with 125–130, 85, 70, and 42–44 kD a were
enriched relative to ‘‘0’’ time cells (Fig. 1, lanes 1 and 2).
The intensity of these bands remained elevated for at least
3 h (data not shown). However, these protein bands were
faint or not d etectable in lysates of unatta ched cells (Fig. 1,
lane 4). Several other cross-reacting bands found in ‘‘0’’time cells were also absent in the unattached cells (Fig. 1,
lane 4). In cells attaching to P L, the 125–130 kD a band was
enriched relative to ‘‘0’’ time, but not to levels observed
with C AP (Fig. 1, lane 3). These bands w ere phosphorylat-
ed in monolayers even before adding CAP, and differences
could not be detected after the addition of CAP (Fig. 1,
lanes 5 and 6).
FAK is a ma jor phosphorylated protein in cells adhering
to FN and exposed to growth factors, and this molecule is
SIGNALING REACTIONS INDUCED BY CEMENTUM ATTACHMENT PROTEIN 67

7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 4/8
involved in the formation of focal contacts.(13,14,16) There-
fore, we examined if the protein band migrating with ∼125–130 kDa and t yrosine phosphorylated is FAK. I mmunopre-
cipitates of cell lysates obtained using a specific anti-FAK
antibody were subjected to immunoblotting with antiphos-
photyrosine antibody. The results showed that FAK was
present in the immunoprecipitates (Fig. 2a, lower panel). It
was not tyrosine phosphorylated in ‘‘0’’ time cells (Fig. 2a,
lane 1, upper panel), but tyrosine-phosphorylated in cells
atta ching to CA P (lane 2). The electrophoretic migration of
FAK resembles Cas, which is a substrate for phosphorylat-
ed FAK and a component of focal adhesion plaques(15);
therefore, we examined whether the 125–130 kD a protein
band contains phosphorylated Cas. The Cas was immuno-
precipitated by the anti-Cas antibody (Fig. 2b, bottom
panel); however, it was only negligibly tyrosine phosphor-ylated in cells attaching to CAP (Fig. 2b, upper panel).
Zhu and Assoian(30) have demonstrated that activation
of the MAP K is associated with cell adhesion and spread-
ing. Figure 1 (lane 2) showed that a protein ba nd migrating
with ∼42kD a w as tyrosine phosphorylated in cells adherent
to CAP. The mobility of this protein corresponded to that
of the activated MAPK ERK-2, the active form of which
requires phosphorylation of tyrosine and threonine.(31) To
examine if this protein band contains MAPK, cells were
allowed to adhere to CAP-coated dishes for various time
periods, and cell lysates w ere subjected to immunoblotting
with antiphosphotyrosine and anti-MAPK antibodies a nd
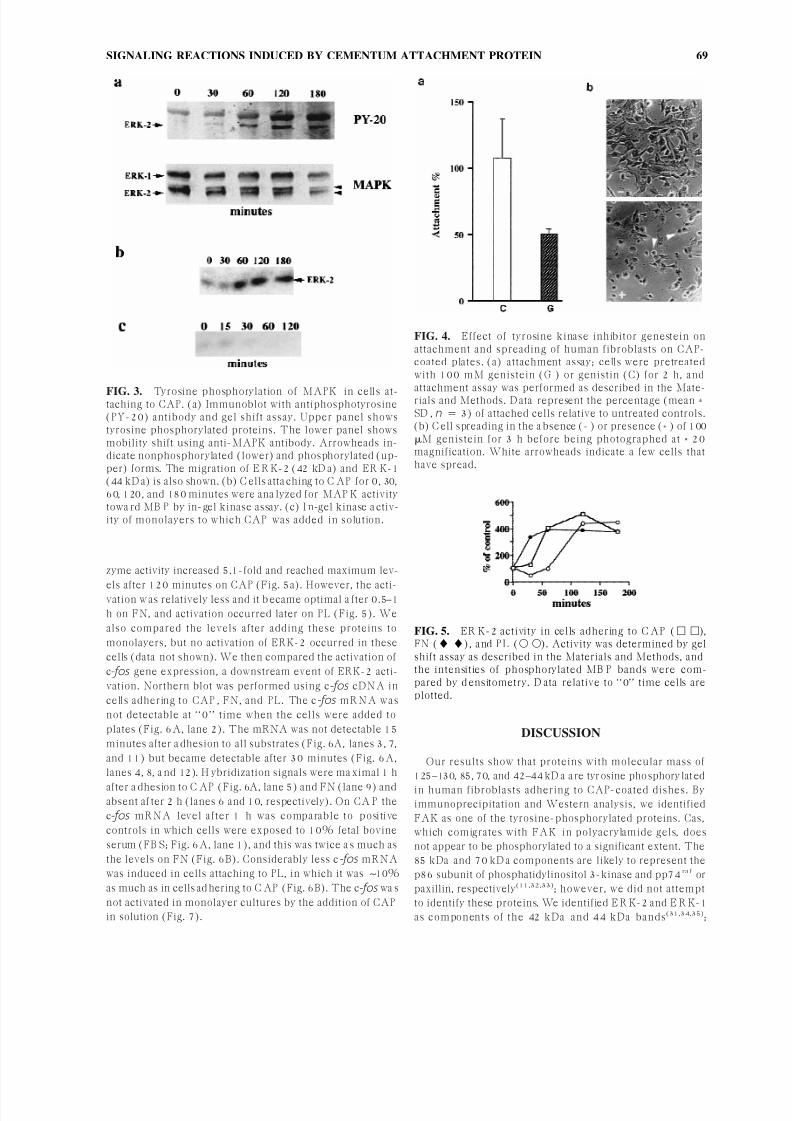
gel shift assay. The 42–44 kD a band became tyrosine pho-shorylated in 60–120 minutes after the cells a ttached to
CAP (Fig. 3a, upper panel) and remained phosphorylated
for 3 h. Western analysis using anti-MAPK antibody re-
vealed that there wa s a distinct electrophoretic mobility
shift o f this protein (Fig. 3a, lower panel). It s level reached
a maximum after 60–120 minutes in cells adhering to CA P
and decreased after 180 minutes. However, a more slowly
migrating 44 kDa band recognized by this antibody did not
manifest mobility shift. To verify if the 42 kD a protein has
MAPK activity and to determine the kinetics of its activa-
tion, we measured kinase activity by in-gel kinase assay
after fra ctionation of cell lysates on SD S-PA G E containing
its substrate, MBP . We found that the 42 kD a protein band
was a ble to phosphorylate M B P substrate 60–180 minutesafter adhesion to CAP (Fig. 3b). We also examined if this
enzyme is activated by the addition of CAP in solution;
however, although there wa s some activation 15 minutes
after C AP add ition, it was transient and no activation could
be detected at later time points (Fig. 3c).
These results showed that tyrosine phosphorylation of
FAK, ERK-2, and other proteins occur in fibroblasts ad-
hering to CAP. To determine if phosphorylation of these
proteins is necessary for cell adhesion, cells were exposed to
genestein, a competitive inhibitor of the ATP binding site
for protein ty rosine kinases, and the effect o n cell adhesion
was examined. Pretreatment of cells with genistein for 2 h
did not abrogate attachment to CAP-coated dishes com-
pletely, although the atta chment decreased to 50.0 ± 5.7%(Fig. 4a). The decrease was statistically significant (p <
0.005). Atta chment was no t inhibited by the inactive ana log
genistin (Fig. 4a), and genistein also inhibited cell spreading
(Fig. 4b).
We examined whether signaling events induced by adhe-
sion to CA P ma nifest differences from those induced when
cells adhere to other substrates. First we compared the ki-
netics of a ctivation of ER K-2 in cells attaching to FN a nd
PL. Activation was determined by gel shift assay. The en-
FIG. 1. Protein tyrosine phosphorylation in human gingi-val fibroblasts adhering to CAP-coated dishes or exposedto CAP in solution. C ells were serum starved for 48 h,trypsinized, and allowed to adhere to plates coated with 2.5g/ml of CAP (lanes 1 and 2), or 5.0 g/ml P L (lane 3).Lanes 5 and 6 represent monolayers to which medium with-out (lane 5) and with (lane 6) 2.5 g/ml CA P was a dded.After 60 minutes of incubation, cells were lysed and 50 gof protein was separated by SD S-PA G E , transferred to ni-trocellulose membrane, and subjected to Western analysisusing a ntiphosphotyrosine a ntibody. B ands w ere visualizedby enhanced chemiluminescence substrate. Cross-reactingprotein bands, which are o f great er intensity relative to ‘‘0’’time (lane 1), are indicated by the thick arrows on the right.Thin arrows on the left indicate the migration of proteinmarkers w ith mo lecular ma ss 213, 123, 85, 50, and 30 kD a,respectively, from the top. Lane 4, cells not attaching toCA P-coated plates in 60 minutes.
FIG. 2. CA P-induced tyrosine phosphorylation of FAK.Fibroblasts were allowed to ad here on CAP for 1 h, and celllysates were immunoprecipitated with (a) anti-FAK anti-body or (b) anti-Cas antibody. First, Western blotting wasdone with antiphosphotyrosine antibody (top panels). Theblots were stripped and reprobed with the respective anti-bodies used for immunoprecipitation (bottom panels).Lane 1, ‘‘0’’ time cells; lane 2, atta ched cells. The da ta pre-sented are from the same autogradiograph and the sameexposure.
SAITO AND NARAYANAN68
7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 5/8
zyme activity increased 5.1-fold and reached maximum lev-
els after 120 minutes on CAP (Fig. 5a). However, the acti-vation w as relatively less and it b ecame optimal a fter 0.5–1
h on FN, and activation occurred later on PL (Fig. 5). We
also compared the levels after adding these proteins to
monolayers, but no activation of ERK-2 occurred in these
cells (data not shown). We then compared the activation of
c-fos gene expression, a downstream event of ERK-2 acti-
vation. Northern blot was performed using c-fos cD NA in
cells adhering to CAP , FN, and PL. The c-fos mR NA w a s
not detectable at ‘‘0’’ time when the cells were added to
plates (Fig. 6A, lane 2). The mRNA was not detectable 15
minutes after a dhesion to all substrates (Fig. 6A, lanes 3, 7,
and 11) but became detectable after 30 minutes (Fig. 6A,
lanes 4, 8, a nd 12). H ybridization signals were ma ximal 1 h
after a dhesion to C AP (Fig. 6A, lane 5) and FN (lane 9) andabsent af ter 2 h (lanes 6 and 10, respectively). On CA P the
c-fos mRNA level after 1 h was comparable to positive
controls in which cells were exposed to 10% fetal bovine
serum (FB S; Fig. 6A, lane 1), and this was twice a s much as
the levels on FN (Fig. 6B). Considerably less c -fos mRNA
was induced in cells attaching to PL, in which it was ∼10%
as much as in cells ad hering to C AP (Fig. 6B). The c-fos wa s
not activated in monolayer cultures by the addition of CAP
in solution (Fig. 7).
DISCUSSION
Our results show that proteins with molecular mass of
125–130, 85, 70, and 42–44 kD a a re tyr osine pho sphory lat ed
in human fibroblasts adhering to CAP-coated dishes. Byimmunoprecipitation and Western analysis, we identified
FAK as one of the tyrosine-phosphorylated proteins. Cas,
which comigrates with FAK in polyacrylamide gels, does
not appear to be phosphorylated to a significant extent. The
85 kDa and 70 kD a components are likely to represent the
p86 subunit of phosphatidylinositol 3-kinase and pp74ra f or
paxillin, respectively(11,32,33); however, we did not attempt
to identify these proteins. We identified E R K-2 and E R K-1
as components of the 42 kDa and 44 kDa bands(31,34,35);
FIG. 3. Tyrosine phosphorylation of MAPK in cells at-taching to CAP. (a) Immunoblot with antiphosphotyrosine(PY-20) antibody and gel shift assay. Upper panel showstyrosine phosphorylated proteins. The lower panel showsmobility shift using anti-MAPK antibody. Arrowheads in-dicate nonphosphorylated (lower) and phosphorylated (up-per) forms. The migration of E R K-2 (42 kD a) and ER K-1(44 kD a) is also shown. (b) C ells atta ching to C AP for 0, 30,60, 120, and 180 minutes were ana lyzed for MAP K activitytowa rd MB P by in-gel kinase assay. (c) I n-gel kinase a ctiv-ity of monolayers to which CAP was added in solution.
FIG. 4. Effect of tyrosine kinase inhibitor genestein onattachment and spreading of human fibroblasts on CAP-coated plates. (a) attachment assay; cells were pretreatedwith 100 mM genistein (G ) or genistin (C) for 2 h, and
attachment assay was performed as described in the Mate-rials and Methods. D ata represent the percentage (mean ±SD , n 3) of attached cells relative to untreated controls.(b) C ell spreading in the a bsence (− ) or presence (+ ) of 100M genistein for 3 h before being photographed at × 20magnification. White arrowheads indicate a few cells thathave spread.
FIG. 5. ER K-2 activity in cells adhering to C AP ( ),FN (), and PL ( ). Activity was determined by gelshift assay as described in the Materials and Methods, andthe intensities of phosphorylated MB P bands were com-pared by d ensitometry. D ata relative to ‘‘0’’ time cells areplotted.
SIGNALING REACTIONS INDUCED BY CEMENTUM ATTACHMENT PROTEIN 69

7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 6/8
however, only the E R K-2 appears to be ty rosine phosphor-ylated.
The FAK is autophosphorylated in response to ligand
occupancy of integrins, clustering of integrins without li-
gands and in response to growth factors, and its activation
is associated with the formation of focal adhesions where it
is localized along with other phosphotyrosine-containing
proteins and vinculin.(9,11–17) Focal contacts are also formed
in cells adhering to CAP, and these sites are positively im-
munostained by antibodies to phosphotyrosine (data not
shown). Phosphorylation of ER K-2 shows that the MAP K
cascade, which mediates many cellular functions,(31,35) is
activated. One of the outcomes of MAPK cascade activa-
tion is induction of gene expression for AP-1 proteins, (36,37)
and, as expected, adhesion to CAP induces c-fos gene ex-pression (Fig. 6). However, peak ERK-2 activity is ob-
served after 2–3 h, while c-fos expression is optimal in 1 h,
indicating that c-fos expression is activated before ERK-2
activity reaches a maximum level, and that other factors
also contribute to c-fos expression. In contrast, Cas and
ERK-1, which are phosphorylated in other adhering cell
types,(15,30,34) are not affected in the human fibroblasts ad-
hering to CAP. We also did not observe a transient activa-
tion o f E R K-1/2 immediately in response to adhesion, al-
though it occurs rapidly and transiently, reaching a peak in
10–15 minutes in other adhering cells and in the gingival
fibroblasts in response to growth factors.(29,34)
These proteins are not phosphorylated in cells not attach-
ing to CA P, in which c-fos expression is a lso not activated.However, cell adhesion is not abrogated by treating cells
with genestein.(2) These results indicate that protein tyro-
sine phosphorylation occurs as a result o f adhesion. The
phosphorylation a ppears to require binding of CA P to its
receptor (see below) because they are not phosphorylated
in cells attaching to P L. P rotein tyrosine phosphorylation of
several proteins occurs significantly in mo nolayer cultures
even without the addition of CAP, and the level of phos-
phorylation increases when CAP is added to these cells in
solution (data not shown). This observation indicates that
phosphorylation can occur by adhesion-dependent and in-
dependent mechanisms. Our results also indicate that the
signaling events can be divided into two types based on
requirement for a dhesion a nd binding to cell surfaces: FAKand similar proteins, which are phosphorylated by binding
to substrates with or without cell adhesion, and those such
as ERK-2 activation and c-fos expression, which require
both adhesion and binding.
The kinet ics of M A P K a ct iva t ion roughly pa ra llels
spreading, and the spreading, not attachment, is inhibited
by 12-0-tetradecanolyphorbol-13-acetate (data not shown).
Cell spreading is also inhibited by genestein. These data
support the observations of Zhu and Assoian(30) that acti-
vation of the MA PK cascade may contribute to cell spread-
ing. H owever, mechanisms other than MAPK activation
appear to be necessary for cell spreading because, even
though the MAPK cascade (E RK -2) is activated, the cells
do not spread on PL for up to 6 h. Similarly, an increase inthe levels of c-fos mRNA, which is also induced b y many
agents including serum,(37,38) is dependent on cell adhesion,
and this gene, like MAPK, is not activated in unattached
cells and monolayers and very little in cells adhering to PL
(Figs. 6 and 7). These results indicate that some of the
signaling events induced during adhesion to CAP involve
specific adhesive interactions between the cells and CAP.
Western analysis shows the presence of several distinct
tyrosine-phosphorylated proteins in ‘‘0’’ time cells. Al-
FIG. 7. c-fos mRNA expression by monolayer cultures ofhuman gingival fibroblasts exposed for 1 h to 10% FBS(lane 1) a nd serum-free D ulbecco modified Eagle’s me-dium (lane 2), or to CAP for 30 minutes (lane 3) and 1 h(lane 4). Fifteen micrograms of R NA w as separated . U pperpanel, c-fos ; lower panel, 28S RNA.
FIG. 6. c-fos mRNA in adhering fibroblasts. (A) Northernanalysis with c-fos (upper panel) and 28S RNA (lowerpa nel) probes. Fift een microgra ms of t ot a l R NA w a sloaded onto gels. Lane 1, cells exposed to 10% FB S for 1 h;lane 2, ‘‘0’’ time; lanes 3–6, CAP; lanes 7–10, FN; lanes11–14, PL ; lan es 3, 7, and 11, 15 minutes; lanes 4, 8, a nd 12,30 minutes; lanes 5, 9, and 13, 1 h; lanes 6, 10, and 14, 2 h.The arrow (top panel) and arrowhead (lower panel) indi-cate c-fos and 28S mRNA, respectively. (B) c-fos mRNAlevels in cells attaching to CAP ( ), FN (o—o), and PL(. . .). *Cells exposed to FBS. Arbitrary densitometryunits normalized for 28S RNA are plotted. Approximately4 g of tota l mRNA was used for lane 3 due to low numberof cells that attached to CAP in 15 minutes in this experi-ment.
SAITO AND NARAYANAN70

7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 7/8
though we did not characterize these proteins, they a re
likely to represent proteins tyrosine phosphorylated during
normal cell metabolism. These are presumably phosphory-
lated by adhesion-independent mechanisms, by serum fa c-
tors present in culture medium, for example.
B oth C AP and FN mediate cell attachment through arg-
gly-asp sequence, and both bind to fibroblasts through 51
integrins(7,24,39) (I w a now s ki, B a rt old, a nd Na ra ya na n,
manuscript in preparation). The fibroblasts form focal con-tacts on both substrates (data not shown). Nevertheless,
there are significant differences in the type of signaling re-
actions induced by CAP and FN. For example, the kinetics
of ERK-2 activation are different; while optimal activation
occurs 30–60 minutes after adhesion to FN (Fig. 5),(30,34) it
begins and maximizes later on CA P. E R K-1 is not activated
by the CAP ; however, this appears to be a function of gin-
gival fibroblasts because it is not activated by FN as well
(data not shown). G enistein inhibits CAP-mediated fibro-
blast attachment significantly, whereas the inhibition of at-
tachment by this drug on FN is relatively less (the attach-
ment of genestein-treated fibroblasts in one experiment was
56.0 ± 11.4, 64.6 ± 28.2, and 76.4 ± 16.9% [n 3] as much
as controls on dishes coated with C AP , FN, and P L, respec-tively). Finally, adhesion on CAP induces c-fos expression
to levels comparable to those induced by FBS, while it is
stimulated to only half as much on the FN. These data
indicate tha t the C AP -mediated a dhesion is associated with
induction of signaling events in a characteristic manner.
Substrate-specific differences in signaling events have also
been reported in mouse aortic smooth muscle cells; in these
cells, adherence to extracellular matrix, not to FN, induces
FAK phosphorylation.(40)
An essential requirement for wound regeneration is the
recruitment, proliferation, and differentiation of the right
cell type(s) in preference over other cells. This is especially
important in periodontal regeneration in which epithelial
cells must be excluded and fibroblasts and cementoblastsselected from a pool of many progenitor cells.(41) Our re-
sults indicate that adhesion proteins are likely to play a key
role in the cell selection process. This could be achieved in
at least two different ways. One is the type of proteins
available in the w ounded tissue to promote adhesion, which
may be specific to certain cell types and subtypes. Alterna-
tively, cell selection may depend upon the ability of target
cells to adhere to the available adhering molecules. In ei-
ther case, cell selection is most likely to depend on the type
and magnitude of signaling events induced. The signaling
events can also be expected to determine how the target
cells respond functionally. For example, cells may grow or
differentiate depending upon the adhesion substrate, pres-
ence of a matrix, and the type of soluble mediators. (1,3,4)
Such signaling events could include the MAP K cascade, the
AP-1 component fo s , which is induced by the MAP cas-
cade,(36,37) as well as other rea ctions that participate in cel-
lular functions.
ACKNOWLEDGMENTS
We are grateful to Dr. David Morris for c-fos cDNA
probe and Dr. Jean Campbell for her generous gift of an-
tiserum against MAPK. We thank Dr. Hidenori Koyama
and members of D r. Bradford C . B erk’s Laboratory, D e-
partment of Medicine/Ca rdiology, U niversity of Washing-
ton for advice and discussions during the course of this
work. The assistance of Drs. Matthew Wright, Scott Coats,
R onald Siefert, and D aniel Williams is highly appreciated.
This research wa s supported by U .S. National I nstitutes of
Hea lth grants D E 10491 and D E 08229.
REFERENCES
1. Adams JC, Watt FM 1993 Regulation of development anddifferentiat ion by the extracellular matrix. D evelopment117:1183–1198.
2. Ekblom P , Vestweber D , Kemler R 1986 Cell–matrix interac-tions and cell adhesion during development. Annu Rev CellBiol 2:27–47.
3. G umbiner B M 1996 Cell ad hesion: The molecular b asis of tis-sue architecture and morphogenesis. Cell 84:345–357.
4. Lin CQ, Bissell MJ 1993 Multi-faceted regulation of cell dif-ferentiation by extracellular matrix. FASEB J 7:737–743.
5. Pardee AB 1989 G 1 events and regulation of cell proliferation.Science 46:603–608.
6. Stoker M, O’Neil C, B erryman S, Waxman V 1968 Anchorageand growth regulation in normal and virus transferred cells. IntJ Cancer 3:683–693.
7. Hynes R O 1992 Integrins: Versatility, modulation, a nd signal-ing in cell adhesion. C ell 69:11–25.
8. Juliano R L, H askill S 1993 Signal transduction from the extra-cellular matrix. J Cell Biol 120:577–585.
9. Miyamoto S, Teramoto H , Coso OA, G utkind JS, Burbelo PD ,Akiyama SK, Yamada KM 1995 Integrin function: Molecularhierarchies of cytoskeletal and signaling molecules. J Cell B iol131:791–805.
10. Yamada KM, Miyamoto S 1995 Integrin transmembrane sig-naling and cytoskeletal control. Curr O pin Cell B iol 7:681–689.
11. B urridge K , Turner CE , R omer L H 1992 Tyrosine phosphor-ylation of paxillin and ppFAK accompanies cell adhesion toextracellular matrix: A role in cytoskeletal a ssembly. J CellBiol 119:893–903.
12. Craig SW, Johnson R P 1996 A ssembly of focal a dhesions:Progress, paradigms, portents. Curr Opin Cell Biol 8:74–85.
13. G uan J L, Shalloway D 1992 R egulation of focal adhesion-associated protein tyrosine kinase by both cellular adhesionand oncogenic transformation. Nature 358:690–692.
14. Kornberg L, E arp H S, Parsons JT, Schaller M, Juliano R L 1992Cell adhesion or integrin clustering increases phosphorylationof a focal adhesion-associated tyrosine kinase. J Biol Chem267:23439–23442.
15. Nojima Y , Morino N, Mimura T, Ha masaki K, Furuya H , SakaiR, Sato T, Tachibana K, Morimoto C, Yazaki Y, Hirai H 1995Integrin-mediated cell adhesion promotes tyro sine phosphor-ylation of p130Ca s, a Src homology 3-containing molecule hav-ing multiple Src homology 2-binding motifs. J Biol Chem270:15398–15402.
16. Schaller MD , P arsons JT 1994 Focal a dhesion kinase a nd a s-sociated proteins. Curr Opin Cell Biol 6:705–710.
17. Wood CK , Turner CI, J ackson P , Critchley D R 1994 Chara c-terization of the paxillin-binding site a nd the C -terminal focaladhesion targeting sequence in vinculin. J Cell Sci 107:709–717.
18. G ailit J, Clark RAF 1994 Wound repair in the context of ex-tracellular matrix. Curr Opin Cell Biol 6:717–725.
19. Clark RAF 1996 The Molecular and Cellular Biology ofWound Repair. Plenum Press, New York, NY, U.S.A.
20. Minabe M 1991 A critical review o f the biologic rationale forguided tissue regeneration. J Periodontol 62:171–179.
21. Wikesjo U M, Nilveus R E , Selvig K 1992 Significance of earlyhealing events on periodontal repair: A review. J Periodontol63:158–165.
22. Olson RH , Adams D F, Layman D L 1985 Inhibitory effect of
SIGNALING REACTIONS INDUCED BY CEMENTUM ATTACHMENT PROTEIN 71

7/17/2019 5650140109_ftp
http://slidepdf.com/reader/full/5650140109ftp 8/8
periodontally diseased root extracts on the growth of humangingival fibroblasts. J Periodontol 56:592–596.
23. Polson AM 1986 The root surface and regeneration: Presenttherapeutic limitations and future biologic potentials. J ClinPeriodontol 13:995–999.
24. Olson S, Arzate H, Narayanan AS, Page RC 1991 Cell attach-ment activity of cementum proteins and mechanism of endo-toxin inhibition. J D ent Res 70:1272–1277.
25. Wu D, Ikezawa K, Parker T, Saito M, Narayanan AS 1996Chara cterization o f a collagenous cementum-derived attach-
ment protein. J Bone Miner R es 11:686–692.26. Arzate H , O lson SW, Page R C, G own AM, Narayanan AS
1992 Production of a monoclonal antibody to an attachmentprotein derived from human cementum. FASE B J 6:2990–2995.
27. Pitaru S, Narayanan SA , Olson S, Savion N, Hekmati H, Alt I,Metzger Z 1995 Specific cementum a ttachment protein en-hances selectively the attachment and migration of periodontalcells to root surfaces. J Periodontal Res 30:360–368.
28. Liu HW, Ya cobi R, Savion N, Nara yanan A S, Pitaru S 1997 Acollagenous cementum-derived attachment protein is a markerfor progenitors of the mineralized tissue-forming cell lineage ofthe periodontal ligament. J Bone Miner Res 12:1691–1699.
29. Yonemura K, Raines EW, Ahn N, Narayanan AS 1993 Mito-genic signaling mechanisms of human cementum-derivedgrowth factors. J Biol C hem 268:26120–26126.
30. Zhu X, Assoian RK 1995 I ntegrin-dependent activation of
MAP kinase: A link to shape- dependent cell proliferation.Mol Biol Cell 6:273–282.
31. Seger R, Krebs E G 1995 The MA PK signaling cascade.FASEB J 9:726–735.
32. Chen H C, G uan J L 1994 Association o f focal adhesion kinasewith its potential substrate phosphatidylinositol 3-kinase. ProcNatl Acad Sci USA 91:10148–10152.
33. Turner CE, G lenney JR Jr, Burridge K 1990 Paxillin: A new vinculin-binding protein present in focal adhesions. J C ell Biol111:1059–1068.
34. Morino N, Mimura T, Ha masaki K, Tobe K, U eki K, KikuchiK, Takehara K, Kadowaki T, Yazaki Y, Nojima Y 1995 Ma-trix–integrin interaction activates the mitogen-activated pro-tein kinase, p44erk–1 and p42erk–2. J Biol Chem 270:269–273.
35. Chen Q, Kinch MS, Lin TH, Burridge K, Juliano RL 1994Integrin-mediated cell adhesion a ctivates mitogen-activatedprotein kinases. J Biol Chem 269:26602–26605.
36. D avis R J 1993 The mitogen-activated protein kinase signaltransduction pathway. J Biol Chem 268:14553–14556.
37. K arin M 1995 The regulation of AP -1 activity by mitogen-activated protein kinases. J Biol Chem 270:16483–16486.
38. Piechaczyk N, Blandhard JM 1994 c-fos proto-oncogene regu-lation and function. Crit R ev Oncol H ematol 17:93–131.
39. Pierschbacher MD , R uoslahti E 1984 Cell atta chment activityof fibronectin can be duplicated by small synthetic fragments ofthe molecule. Nature 309:30–33.
40. Wilson L, Carrier MJ, Kellie S 1995 pp125FAK tyrosine kinaseactivity is not required for the assembly of F-actin stress fibresand focal adhesions in cultured mouse aortic smooth musclecells. J Cell Sci 108:2381–2391.
41. Pitaru S, McCulloch CAG , Narayanan AS 1994 Cellular ori-gins and differentiation control mechanisms during periodontaldevelopment and wound healing. J Periodontal Res 29:81–94.
Address reprint requests to:
A .S. Nar ayanan D epartment of Pathology
Box 357470
Uni versity of Washington School of M edicine
Seattle, W A 98195 U .S.A .
Received in original form May 11, 1998; in revised form August 4,1998; accepted August 28, 1998.
SAITO AND NARAYANAN72





























