Embed Size (px)
Citation preview
letters
nature structural biology • volume 6 number 8 • august 1999 755
The tetramerizationdomain of the Mntrepressor consists of tworight-handed coiled coilsIrene M.A. Nooren1, Robert Kaptein1, Robert T. Sauer2
and Rolf Boelens1
1Department of NMR Spectroscopy, Bijvoet Center for BiomolecularResearch, Utrecht University, Padualaan 8, 3584 CH, Utrecht, TheNetherlands. 2Department of Biology, Massachusetts Institute ofTechnology, Cambridge, Massachusetts 02139, USA.
The tetrameric Mnt repressor is involved in the geneticswitch between the lysogenic and lytic growth ofSalmonella bacteriophage P22. The solution structure of itsC-terminal tetramerization domain, which holds togetherthe two dimeric DNA-binding domains, has been deter-mined by NMR spectroscopy. This structure reveals anassembly of four a-helical subunits, consisting of a dimerof two antiparallel coiled coils with a unique right-handedtwist. The superhelical winding is considerably strongerand the interhelical separation closer than those found inthe well-known left-handed coiled coils in fibrous proteinsand leucine zippers. An unusual asymmetry arises betweenthe two monomers that comprise one right-handed coiledcoil. A difference in the packing to the adjacent monomerof the other coiled coil occurs with an offset of two helicalturns. The two asymmetric monomers within each coiledcoil interconvert on a time scale of seconds. Both withrespect to symmetry and handedness of helical packing, theC2 symmetric four-helix bundle of Mnt differs from otheroligomerization domains that assemble DNA-bindingmodules, such as that in the tumor suppressor p53 and theE. coli lac repressor.
Supercoiling of a-helices, as first recognized by Crick1,2, iswidely found in both fibrous and globular proteins3,4. Tworight-handed amphipathic a-helices that wind around oneanother in a left-handed fashion comprise the so-called coiled-coil motif. While undistorted a-helices have ~3.6 residues perturn, the left-handed twist in these coiled coils reduces theperiodicity to 3.5 with respect to the supercoil axis (heptadrepeat). Winding of the supercoil in a right-handed fashionleads to a superhelical periodicity that exceeds that of undis-torted helices. Recently, a de novo designed peptide has beenshown to form a stable four-stranded right-handed coiledcoil5. However, right-handed coiled coils have not beenobserved as stable oligomeric folds of peptide chains in natu-rally occurring proteins. Between non-identical a-helices,right-handed coiled coils have been found in a few globularproteins, but mostly not as isolated folds (SCOP database6).Right-handed coiled coils have been proposed for several,mainly membrane proteins based on modeling of amphipathichelices (as referenced by Harbury et al.5). Non-intertwinedright-handed supercoils have been identified in the structureof the dimeric transmembrane domain of glycophorin A inaqueous detergent micelles7.
Here, we present the solution structure of the tetramerizationdomain of the Mnt repressor of Salmonella bacteriophage P22and show that it forms a unique assembly of two right-handed,
two-stranded coiled coils. The Mnt repressor maintains the lyso-genic growth of the P22 prophage8. Its dimeric N-terminaldomain contacts operator DNA and the C-terminal domain(residues 52–82; Mnt-C) is responsible for tetramerization andstrong DNA binding9. Treatment with chymotrypsin separatesthe two domains by cleavage of the linker region9. Biochemicaland biophysical experiments have demonstrated that the chy-motryptic C-terminal fragment forms a stable assembly of thefour subunits with a high a-helical content9.
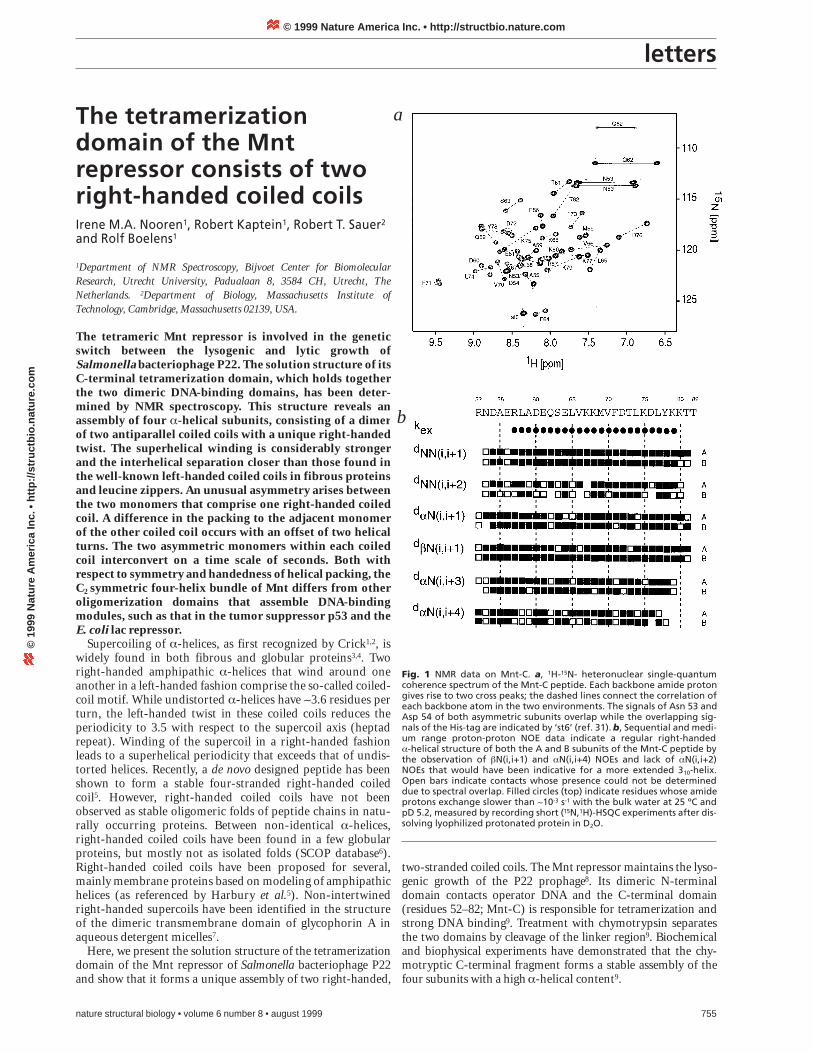
Fig. 1 NMR data on Mnt-C. a, 1H-15N- heteronuclear single-quantumcoherence spectrum of the Mnt-C peptide. Each backbone amide protongives rise to two cross peaks; the dashed lines connect the correlation ofeach backbone atom in the two environments. The signals of Asn 53 andAsp 54 of both asymmetric subunits overlap while the overlapping sig-nals of the His-tag are indicated by ‘st6’ (ref. 31). b, Sequential and medi-um range proton-proton NOE data indicate a regular right-handeda-helical structure of both the A and B subunits of the Mnt-C peptide bythe observation of bN(i,i+1) and aN(i,i+4) NOEs and lack of aN(i,i+2)NOEs that would have been indicative for a more extended 310-helix.Open bars indicate contacts whose presence could not be determineddue to spectral overlap. Filled circles (top) indicate residues whose amideprotons exchange slower than ~10-3 s-1 with the bulk water at 25 ºC andpD 5.2, measured by recording short (15N,1H)-HSQC experiments after dis-solving lyophilized protonated protein in D2O.
a
b
© 1999 Nature America Inc. • http://structbio.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://s
tru
ctb
io.n
atu
re.c
om
letters
Mnt-C structureAnalysis of the NMR spectra of Mnt-C revealed an asymmetricassembly of the four subunits. Two sets of NMR resonances Aand B are visible in the NMR spectra (Fig. 1a), correspondingto different chemical environments of two A monomers andtwo B monomers, respectively. Uniform 15N and 13C labeling ofthe Mnt-C peptide was used to obtain unambiguous resonanceassignments of both the A and B polypeptide chains.
The intra- and inter-subunit NOEs of the symmetricallyassembled monomers were discriminated using 13C/15N-fil-tered-NOE experiments. The intra-monomeric NOE connec-tivities demonstrated a continuous regular right-handeda-helical structure in both the A and B chains (Fig. 1b). Theinter-subunit NOEs between the N- and C-terminal region ofthe A strands showed that the two A helices are arranged in an
756 nature structural biology • volume 6 number 8 • august 1999
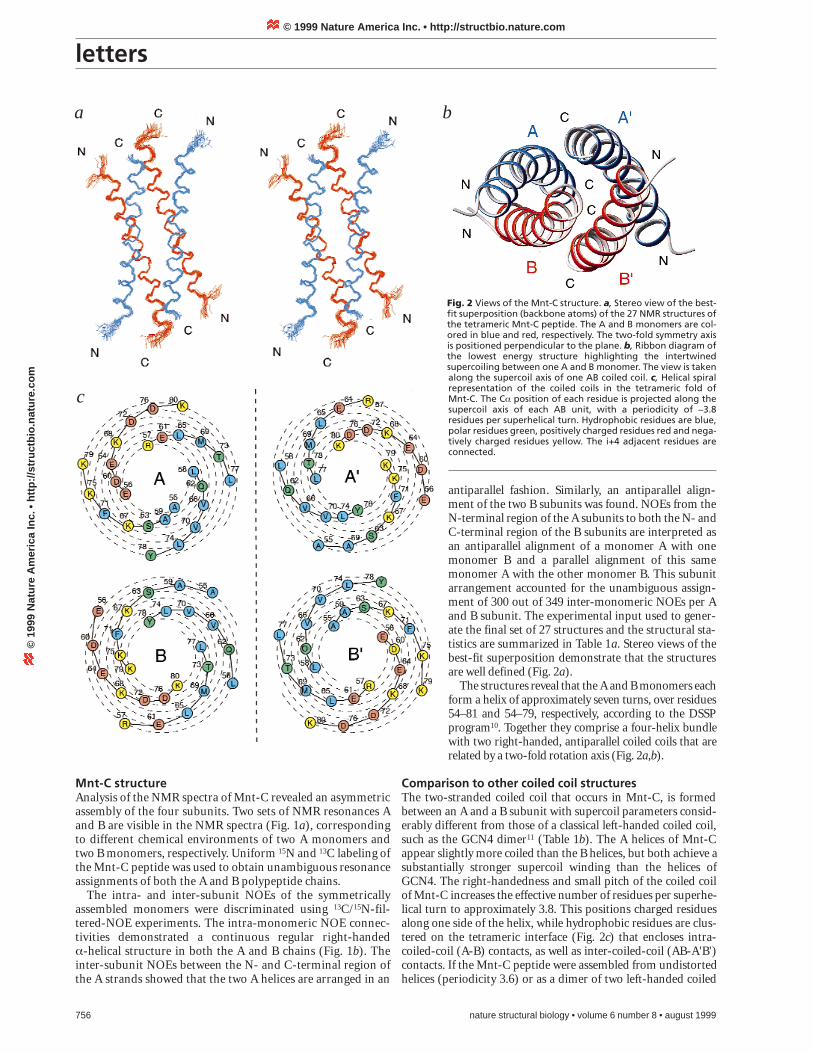
antiparallel fashion. Similarly, an antiparallel align-ment of the two B subunits was found. NOEs from theN-terminal region of the A subunits to both the N- andC-terminal region of the B subunits are interpreted asan antiparallel alignment of a monomer A with onemonomer B and a parallel alignment of this samemonomer A with the other monomer B. This subunitarrangement accounted for the unambiguous assign-ment of 300 out of 349 inter-monomeric NOEs per Aand B subunit. The experimental input used to gener-ate the final set of 27 structures and the structural sta-tistics are summarized in Table 1a. Stereo views of thebest-fit superposition demonstrate that the structuresare well defined (Fig. 2a).
The structures reveal that the A and B monomers eachform a helix of approximately seven turns, over residues54–81 and 54–79, respectively, according to the DSSPprogram10. Together they comprise a four-helix bundlewith two right-handed, antiparallel coiled coils that arerelated by a two-fold rotation axis (Fig. 2a,b).
Comparison to other coiled coil structuresThe two-stranded coiled coil that occurs in Mnt-C, is formedbetween an A and a B subunit with supercoil parameters consid-erably different from those of a classical left-handed coiled coil,such as the GCN4 dimer11 (Table 1b). The A helices of Mnt-Cappear slightly more coiled than the B helices, but both achieve asubstantially stronger supercoil winding than the helices ofGCN4. The right-handedness and small pitch of the coiled coilof Mnt-C increases the effective number of residues per superhe-lical turn to approximately 3.8. This positions charged residuesalong one side of the helix, while hydrophobic residues are clus-tered on the tetrameric interface (Fig. 2c) that encloses intra-coiled-coil (A-B) contacts, as well as inter-coiled-coil (AB-A'B')contacts. If the Mnt-C peptide were assembled from undistortedhelices (periodicity 3.6) or as a dimer of two left-handed coiled
Fig. 2 Views of the Mnt-C structure. a, Stereo view of the best-fit superposition (backbone atoms) of the 27 NMR structures ofthe tetrameric Mnt-C peptide. The A and B monomers are col-ored in blue and red, respectively. The two-fold symmetry axisis positioned perpendicular to the plane. b, Ribbon diagram ofthe lowest energy structure highlighting the intertwinedsupercoiling between one A and B monomer. The view is takenalong the supercoil axis of one AB coiled coil. c, Helical spiralrepresentation of the coiled coils in the tetrameric fold of Mnt-C. The Ca position of each residue is projected along thesupercoil axis of each AB unit, with a periodicity of ~3.8residues per superhelical turn. Hydrophobic residues are blue,polar residues green, positively charged residues red and nega-tively charged residues yellow. The i+4 adjacent residues areconnected.
a
c
b
© 1999 Nature America Inc. • http://structbio.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://s
tru
ctb
io.n
atu
re.c
om

letters
nature structural biology • volume 6 number 8 • august 1999 757
coils (periodicity 3.5), a hydrophobic and uncharged interfaceformed along the whole length of the tetrameric interface wouldbe less clear, and unambiguous inter-subunit NOEs between theantiparallel A and B helices would not be satisfied. The superhe-lical periodicity of Mnt-C also contrasts with that found for thedesigned right-handed coiled peptides that have a repeat of 11residues per three helical turns5.
The pairwise helical packing in the coiled coil of Mnt-C isantiparallel and is more complex than the regular parallel pack-ing that occurs in a classical left-handed coiled coils such asGCN4 (Fig. 3a). Side chains of different size along the length ofthe supercoil interface of Mnt-C give rise to an irregular separa-tion in the interface. The Mnt-C coiled coil displays an interheli-cal separation closer than that in a leucine zipper (Table 1b). Thecoiled-coil interface of Mnt-C has valine and serine residues inthe center, and small residues, such as Ala 55 and Ala 59, at theN-terminus that pack tightly against elongated side chains, suchas Leu 74 and Tyr 78, at the C-terminus (Fig. 2c). This type ofhelical packing has been observed in Ala-coils that involveantiparallel left-handed coiled coils with Ala or another smallresidue in every seventh position of the sequence allowing closeinterhelical distances of 7.5–8.4 Å, as found in a set of unrelatedglobular or membrane proteins12. In Mnt-C, typically, thetyrosines in the C-terminal region of both the A and B subunitspack against Ala 55 and Ala 59 of the B and A subunits, respec-tively. The aromatic ring of Tyr 78 is positioned on the outer faceof the coiled coil and takes part in multiple intra- and inter-coiled-coil hydrophobic and hydrogen bonding interactions.This explains the critical role of Tyr 78 in tetramerization13.Whereas the deletion mutant Mnt(1–78) still forms tetramers,Mnt(1–77) is dimeric in solution. In the dimeric Mnt(1–76)mutant, the C-terminal part is poorly structured, but shows atendency to form an a-helix14.
Symmetry of subunit packingFor parallel two-, three- and four-stranded coiled coils as foundfor native GCN4 and de novo designed peptides mimickingGCN415 the C2, C3 and C4 symmetry axes, respectively, are colin-ear with the supercoil axes. In each of the two-stranded antipar-allel coiled coils of Mnt-C, an ‘approximate’ local two-foldsymmetry axis is present in the middle near Val 66, perpendicu-lar to the plane in Fig. 3a. The Mnt-C tetramer appears as adimer of two two-stranded coiled coils, involving an unusualasymmetry between the subunits that comprise one coiled coil.The A/B asymmetry of the Mnt-C homotetramer arises from adifference between the A-A' and B-B' interfaces in the arrange-ment of the two coiled coils AB and A'B' (Fig. 3b). While thesame side chains are involved in both the A-A' and B-B' interface,the A or B side chain contacts other residues of the symmetryrelated partner (A' or B'). The difference involves a relative shiftof two helical turns. The interhelical angles and separationsbetween the long axes of the two equivalent anti-parallel Ahelices are similar to those measured between the B helices (165°and 10.5 Å, respectively) and both the symmetric A helices andthe symmetric B helices are related by the same two-fold symme-try axis. The C2 axis of the tetramer is, however, positioned nearresidue 69 for the A helices and near residue 65 for the B helices.
It is possible to model a fully D2 symmetric homotetramer byshifting the A-B coiled coils one helical turn with respect to eachother. In this case, the ‘approximate’ C2 axes of the two two-stranded coiled coils coincide. Apparently however, a fully sym-metric assembly of the Mnt-C peptide compatible with thesupercoiling does not provide the lowest free energy. The distrib-
Fig. 3 Interhelical packing in the Mnt-C tetramer. a, Comparison of thecoiled-coil interface in the Mnt-C and GCN4 peptides. The ‘approximate’two-fold symmetry axis of the Mnt-C coiled coil is positioned perpendicu-lar to the plane in the top view. The views at the bottom are taken alongthe supercoil axis, which for GCN4 forms the symmetry axis. b, Comparison of the A-A’ and B-B’ interface in the Mnt-C tetramer.Different colors of side chains denote non-adjacent i+4 residues. The C2
symmetry axis that relates A to A’ and B to B’ is positioned perpendicularto the plane. c, Schematic representation of the packing of the two‘approximate’ symmetric coiled coils seen in (a), colored black and redrespectively. The two right-handed coiled coils can undergo a relativesliding that exchanges the interhelical A-A’ and B-B’ packing seen in (b)and other contacts across the tetrameric interface as depicted betweenthe A monomer (ball) and B monomer of the other coiled coil (bar).
a
b
c
© 1999 Nature America Inc. • http://structbio.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://s
tru
ctb
io.n
atu
re.c
om

letters
ution of the inter-subunit NOEs (Table 1a) and interhelical dis-tances (8.0 Å and 10.5 Å) illustrate that the contacting surfacewithin one coiled coil is significantly larger than between thetwo coiled coils. This is consistent with chemical cross-linkingexperiments9 that suggested a ‘dimer of dimers’ type ofoligomerization for Mnt-C. In contrast to the truncatedMnt(1–76) mutant14, slow amide proton solvent exchange ratesare observed for both helices in Mnt-C (Fig. 1b). ROESY exper-iments also revealed an interconversion of the A and B forms ona time scale of seconds. In view of the slow hydrogen-deuteriumexchange, it is not likely that the A-B exchange process involvescomplete unfolded states. Hence, a plausible model to accountfor the exchange of the A and B forms is a relative sliding orrepacking of the two compact right-handed coiled coils (Fig.3c). While the helices remain intact, the asymmetric A-A' and B-B' packing schemes (Fig. 3b) interchange. Thus, the exchangetakes place between two identical structures of the homo-tetramer (AB-A'B' « BA-B'A').
Comparison to other four-helix bundlesTwo other homotetrameric four-helix bundles that assembleDNA-binding domains have been reported, the tetramerizationdomain of the human tumor-suppressor protein p5316,17 andthat of the lac repressor18. Both show D2 (222) symmetry with allfour helices in an identical environment. The asymmetric sub-unit assembly of the Mnt-C homotetramer restricts its overallsymmetry to C2. The arrangement of helices in Mnt-C also dif-fers from that in the oligomerization domains of p53 and lacrepressor, both in helical curvature and handedness of interheli-cal packing (Fig. 4). Similar to Mnt-C, the helices in the four-helix bundle of p53 assemble as a dimer of dimers but arearranged symmetrically and at a different angle. Four-helixbundles occur as homotetramers, homo- or heterodimers, andas single polypeptide chains, in many functional and structuralsettings4,19. The topology of the four-helix bundle of Mnt-C isstrikingly similar to that of the four-helix bundle of the yeasttranscription factor IIA (TFIIA), the structure of which hasbeen determined in the ternary complex with the TATA-boxbinding protein and DNA20 (Fig. 4). The four-helix bundle ofTFIIA consists of two hairpins of non-identical subunits that
758 nature structural biology • volume 6 number 8 • august 1999
show a right-handed intertwined supercoiling similar to that ofMnt-C.
Left-handed coiled-coil structures as well as four-helix bun-dles form widespread model systems used in de novo peptide orprotein design to understand interhelical packing and stability inprotein structure21. The right-handed coiled coil and asymmet-ric interhelical packing of the tetramerization domain of the Mntrepressor constitute a new and rather stable peptide fold thatmay be used as a starting point for new directions in peptidedesign.
MethodsThe preparation of the isotope enriched Mnt-C peptide from a His-tagged constuct of the Mnt repressor, as well as a detailed descrip-tion of the NMR analysis and structure determination will bereported elsewhere22. Lyophilized Mnt-C was dissolved in 50 mM K-pi pH 5.2, 200 mM NaCl, 0.01 % Na-azide, 5 % D2O to a final con-centration of 3 mM. NMR experiments were carried out at 25 °C oneither a Bruker AMXT-600 or Varian Unity Plus 750, both equippedwith a triple-resonance, z-gradient probe. Sequential and side-chainassignments were achieved using 1H, 15N and 13C heteronuclear cor-relation experiments. 2D 13C/15N-filtered NOESY and 3D 13C-filteredNOESY-HSQC experiments23 discriminate intra- and intersubunitNOEs between the symmetric monomers. The heterotetramer wasobtained by a equimolar mixture of unlabeled (12C/14N) and labeled(13C/15N) Mnt-C. Chemical exchange was measured using 2D ROESYexperiments24 (tm = 50–100 ms), recorded at 10–45 °C.
Distance restraints were collected from a 2D NOESY (tm = 60 ms)and a 3D NOESY-(15N,1H)-HSQC and NOESY-(13C,1H)-HSQC (tm = 75 ms), recorded at 25° C. NOEs were subdivided in three distanceclasses: strong (1.8–2.8 Å), medium (1.8–3.5 Å) and weak (1.8–5.5 Å),using secondary structure elements for calibration. NOEs corre-sponding to residues close to the symmetry axis were treated asambiguous intra- or intersubunit NOEs. Symmetry of the A and Bmonomers respectively was maintained using the X-PLOR non-crys-tallographic symmetry term25 and pseudo-NOE distance restraints26.From randomized atomic coordinates, 30 structures were calculatedusing X-PLOR V3.85125,27. The selected 27 structures were analyzedusing PROCHECK28, whereas XHELIX29 was used for obtaining(super)helical parameters. Molecular figures were generated usingMOLMOL30.
Coordinates. The coordinates of the ensemble of structures havebeen deposited in the Protein Data Bank (accession code 1QEY).
Fig. 4 Comparison of the C2 symmetric Mnt-Cfour-helix bundle to these of the D2 symmetrictetramerization domains of lac repressor andp53, and to the four-helix bundle of yeast TFIIAwith a right-handed superhelix. Note also thedifferences in handedness and crossing angle ofinterhelical packing.
© 1999 Nature America Inc. • http://structbio.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://s
tru
ctb
io.n
atu
re.c
om

letters
AcknowledgmentsThis research was supported by the Council of Earth and Life Sciences of theNetherlands Organization for Scientific Research (ALW/NWO). We thankA.George and C. Waldburger for their help in protein expression andpurification. D. Walther is gratefully acknowledged for his help in the analysisof helical parameters.
Correspondence should be addressed to R.B. email: [email protected]
Received 10 February, 1999; accepted 13 April, 1999.
1. Crick, F.H.C. Acta. Crystallogr. 6, 685–689 (1953).2. Crick, F.H.C. Acta. Crystallogr. 6, 689–697 (1953).3. Lupas, A. Trends Biochem. Sci. 21, 375–382 (1996).4. Kohn, W.D., Mant, C.T. & Hodges, R.S. J. Biol. Chem. 272, 2583–2586 (1997).5. Harbury, P.B., Plecs, J.J., Tidor, B., Alber, T. & Kim, P.S. Science 282, 1462–1467
(1998).6. Murzin, A.G., Brenner, S.E., Hubbard, T. & Chothia, C. J. Mol. Biol. 247,
536–540 (1995).7. MacKenzie, K.R., Prestegard, J.H. & Engelman, D.M. Science 276, 131–133
(1997).8. Susskind, M.M. & Youderian, P. In Lambda II (eds Hendrix, R.W., Roberts, J.W.,
Stahl, F.W. & Weisberg, R.) 347–366 (Cold Spring Harbor Press, Cold SpringHarbor, New York; 1983).
9. Waldburger, C.D. & Sauer, R.T. Biochemistry 34, 13109–13116 (1995).
nature structural biology • volume 6 number 8 • august 1999 759
10. Kabsch, W. & Sander, C. Biopolymers 22, 2577–2637 (1983).11. O’Shea, E.K., Klemm, J.D., Kim, P.S. & Alber, T. Science 254, 539–544 (1991).12. Gernert, K.M., Surles, M.C., Labean, T.H., Richardson, J.S. & Richardson, D.C.
Protein Sci. 4, 2252–2260 (1995).13. Knight, K.L. & Sauer, R.T. Biochemistry 27, 2088–2094 (1988).14. Burgering, M.J. et al. Biochemistry 33, 15036–15045 (1994).15. Harbury, P.B., Zhang, T., Kim P.S. & Alber, T. Science 262, 1401–1407 (1993).16. Lee, W. et al. Nature Struct. Biol. 1, 877–890 (1994).17. Clore, G.M. et al. Science 267, 1515–1516 (1995).18. Friedman, A.M., Fischmann, T.O. & Steitz, T.A. Science 268, 1721–1727 (1995).19. Harris, N.L. Presnell, S.R. & Cohen, F.E. J. Mol. Biol. 1356–1368 (1994).20. Tan, S., Hunziker, Y., Sargent, D.F. & Richmond, T.J. Nature 381, 127–134
(1996).21. Betz, S.F., Bryson, J.W. & DeGrado, W.F. Curr. Opin. Struct. Biol. 5, 457–463 (1995).22. Nooren, I.M., George, A.V.E., Kaptein, R., Sauer, R.T. & Boelens, R. J. Biomol.
NMR, in the press (1999)23. Slijper, M., Kaptein, R. & Boelens, R. J. Magn. Reson. B 111, 199–203 (1996).24. Bax, A. & Davis, D.G. J. Magn. Reson. 63, 207–213 (1985).25. Brünger, A.T. X-PLOR. A system for X-ray Crystallography and NMR (Yale
University Press, New Haven, Connecticut; 1992).26. Nilges, M. Proteins Struct. Funct. Genet. 17, 295–309 (1993).27. Nilges, M., Clore, G.M. & Gronenborn, A.M. FEBS Lett. 239, 129–136 (1988).28. Laskowski, R.A., MacArthur, M.W., Moss, D.S. & Thornton, J.M. J. Appl.
Crystallogr. 26, 283–291 (1993).29. Walther, D., Eisenhaber, F. & Argos, P. J. Mol. Biol. 255, 536–553 (1996).30. Koradi, R., Billeter, M. & Wüthrich, K. J. Mol. Graphics 14, 52–55 (1996).31. Milla, I.M., Brown, B.M. & Sauer, R.T., Protein Sci. 2, 2198–2205 (1998).32. Chothia, C., Levitt, M. & Richardson, D. J. Mol. Biol. 145, 215–250 (1981).
Table 1 Structural statistics for the Mnt-C tetramer and a comparison of a-helical and superhelical parameters for two-stranded coiled coils
a, NMR statistics<SA>1
R.m.s. deviations from experimental data2
Distance restraints (Å) (1,439) 0.008 ± 0.001 Dihedral restraints (°)(60) 0.006 ± 0.012
R.m.s. deviations from idealized geometryBonds (Å) 0.003 ± 0.0001 Angles (º) 0.360 ± 0.003Impropers (º) 0.129 ± 0.007
Ramachandran analysis4
Most favored region 97.3 ± 0.1%Additional allowed regions 2.7 ± 0.1%Generally allowed or disallowed regions 0.0 ± 0.0%
Coordinate precision3
Backbone residues (Å) 0.25 ± 0.05 All heavy atoms (Å) 0.79 ± 0.07 Only core heavy atoms (Å) (Ala, Leu, Met, Tyr, Val) 0.44 ± 0.08
b, Coiled coil characteristicsClassical parallel left-handed, GCN45 Anti-parallel right-handed Mnt-C6
a-Helix parameters7
a-Helix radius (Å) 2.30 2.30 ± 0.01/2.30 ± 0.01Residues per a-helical turn 3.63 3.66 ± 0.01/3.65 ± 0.01Rise per residue (Å) 1.51 1.50 ± 0.23/1.52 ± 0.27
Superhelix parameters8
Interhelical distance9 (Å) 9.4 8.0 ± 0.1Residues per supercoil turn 100 67 ± 2/65 ± 2Supercoil pitch (Å) 141 79 ± 3/94 ± 6Crossing angle10 (º) 25 147 ± 2
1<SA> denotes the final ensemble of 27 simulated annealing structures, selected based on low total energy criteria.2The number of experimental restraints is given per A and B monomer. The distance restraints comprise 545 intra-A, 501 intra-B, 28 inter A, 16 interB, 187 intra-coiled-coil AB, 69 AB inter-coiled-coil AB, 49 ambiguous intra-inter subunit NOE restraints and 44 A and 44 B hydr ogen bond distancerestraints (representing 22 hydrogen bonds for each chain) as indicated by slow amide proton solvent exchange. For monomer A and B, respectively,27 and 26 f dihedral angle restraints and 3 and 4 dihedral restraints on c1 were used.3R.m.s. deviations from the mean structure for the well-defined region (residues 57–79 of monomers A and 57–78 of monomers B).4Residues 54–81 of monomers A and 54–79 of monomers B.5GCN4-p111, pdb entry 2zta.6Parameters are given as averages in the ensemble of structures. For the asymmetric Mnt-C helices, parameters are given for its individual helices(A/B).7The DSSP program10 was used to identify the residues that comprise the a-helices.8Superhelical parameters were obtained by fitting the Ca backbone to a supercoil parametrization suggested by Crick1,2. Supercoiling is measuredwith respect to the supercoil axis of the AB coiled coil.9The interhelical distance represents the closest approach distance.10The crossing angle is defined according to Chothia et al.32.
© 1999 Nature America Inc. • http://structbio.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://s
tru
ctb
io.n
atu
re.c
om





























