Embed Size (px)
Citation preview
Nature © Macmillan Publishers Ltd 1998
8The membrane lipid phosphatidylinositol-3-phosphate (PtdIns(3)P) is constitutivelyproduced by yeast and higher eukaryotesthrough the phosphorylation of phos-phatidylinositol by phosphatidylinositol-3-OH kinase1 (PI(3)K). PtdIns(3)P isimportant for vesicular transport1, but littleis known about how it acts, and proteinsthat specifically recognize it have not yetbeen identified. Here we identify the FYVEfinger2, an evolutionarily conserved double-zinc-binding domain (see Supplementaryinformation), as a protein structure thatbinds to PtdIns(3)P with high specificity.
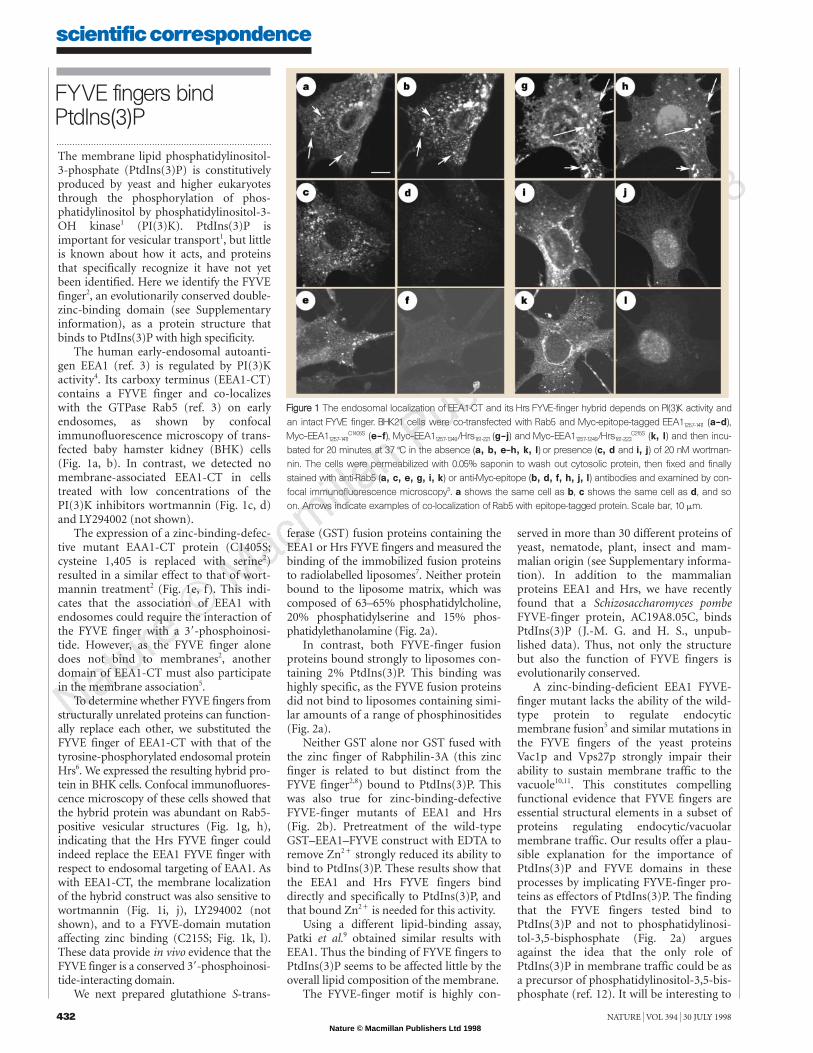
The human early-endosomal autoanti-gen EEA1 (ref. 3) is regulated by PI(3)Kactivity4. Its carboxy terminus (EEA1-CT)contains a FYVE finger and co-localizeswith the GTPase Rab5 (ref. 3) on earlyendosomes, as shown by confocalimmunofluorescence microscopy of trans-fected baby hamster kidney (BHK) cells(Fig. 1a, b). In contrast, we detected nomembrane-associated EEA1-CT in cellstreated with low concentrations of thePI(3)K inhibitors wortmannin (Fig. 1c, d)and LY294002 (not shown).
The expression of a zinc-binding-defec-tive mutant EAA1-CT protein (C1405S;cysteine 1,405 is replaced with serine2)resulted in a similar effect to that of wort-mannin treatment2 (Fig. 1e, f). This indi-cates that the association of EEA1 withendosomes could require the interaction ofthe FYVE finger with a 38-phosphoinosi-tide. However, as the FYVE finger alonedoes not bind to membranes2, anotherdomain of EEA1-CT must also participatein the membrane association5.
To determine whether FYVE fingers fromstructurally unrelated proteins can function-ally replace each other, we substituted theFYVE finger of EEA1-CT with that of thetyrosine-phosphorylated endosomal proteinHrs6. We expressed the resulting hybrid pro-tein in BHK cells. Confocal immunofluores-cence microscopy of these cells showed thatthe hybrid protein was abundant on Rab5-positive vesicular structures (Fig. 1g, h),indicating that the Hrs FYVE finger couldindeed replace the EEA1 FYVE finger withrespect to endosomal targeting of EAA1. Aswith EEA1-CT, the membrane localizationof the hybrid construct was also sensitive towortmannin (Fig. 1i, j), LY294002 (notshown), and to a FYVE-domain mutationaffecting zinc binding (C215S; Fig. 1k, l).These data provide in vivo evidence that theFYVE finger is a conserved 38-phosphoinosi-tide-interacting domain.
We next prepared glutathione S-trans-
scientific correspondence
432 NATURE | VOL 394 | 30 JULY 1998
served in more than 30 different proteins ofyeast, nematode, plant, insect and mam-malian origin (see Supplementary informa-tion). In addition to the mammalianproteins EEA1 and Hrs, we have recentlyfound that a Schizosaccharomyces pombeFYVE-finger protein, AC19A8.05C, bindsPtdIns(3)P (J.-M. G. and H. S., unpub-lished data). Thus, not only the structurebut also the function of FYVE fingers isevolutionarily conserved.
A zinc-binding-deficient EEA1 FYVE-finger mutant lacks the ability of the wild-type protein to regulate endocyticmembrane fusion5 and similar mutations inthe FYVE fingers of the yeast proteinsVac1p and Vps27p strongly impair theirability to sustain membrane traffic to thevacuole10,11. This constitutes compellingfunctional evidence that FYVE fingers areessential structural elements in a subset ofproteins regulating endocytic/vacuolarmembrane traffic. Our results offer a plau-sible explanation for the importance ofPtdIns(3)P and FYVE domains in theseprocesses by implicating FYVE-finger pro-teins as effectors of PtdIns(3)P. The findingthat the FYVE fingers tested bind toPtdIns(3)P and not to phosphatidylinosi-tol-3,5-bisphosphate (Fig. 2a) arguesagainst the idea that the only role ofPtdIns(3)P in membrane traffic could be asa precursor of phosphatidylinositol-3,5-bis-phosphate (ref. 12). It will be interesting to
FYVE fingers bindPtdIns(3)P
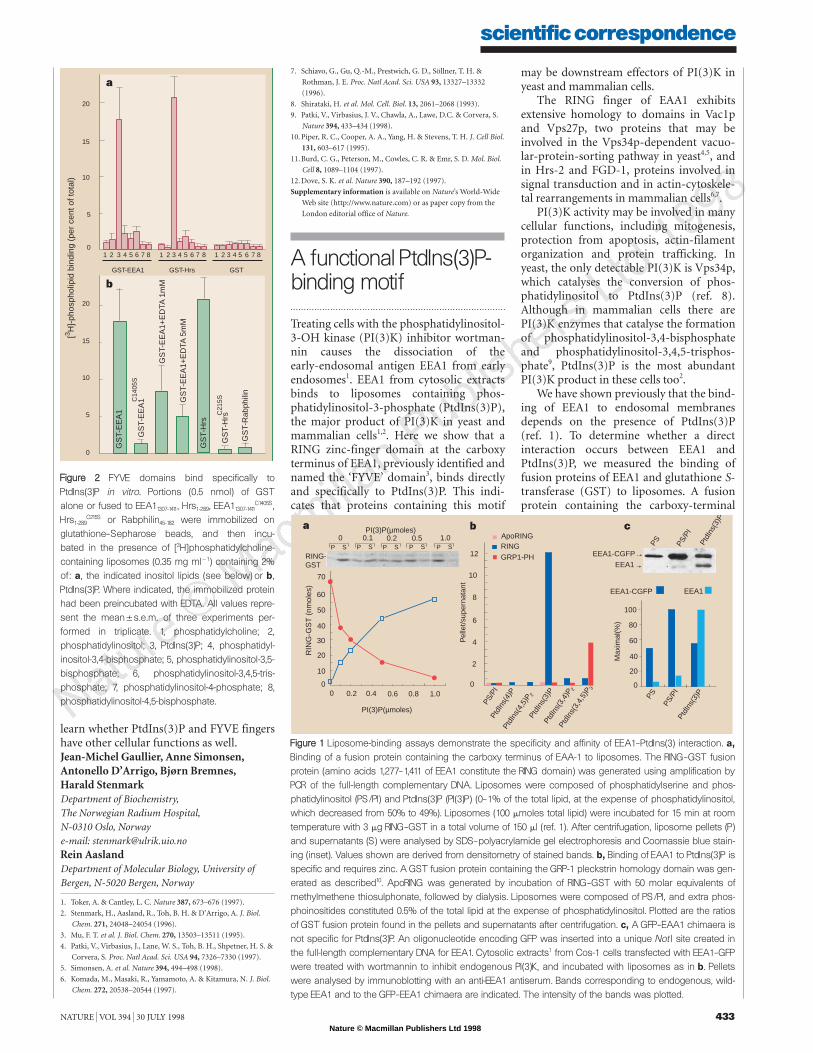
ferase (GST) fusion proteins containing theEEA1 or Hrs FYVE fingers and measured thebinding of the immobilized fusion proteinsto radiolabelled liposomes7. Neither proteinbound to the liposome matrix, which wascomposed of 63–65% phosphatidylcholine,20% phosphatidylserine and 15% phos-phatidylethanolamine (Fig. 2a).
In contrast, both FYVE-finger fusionproteins bound strongly to liposomes con-taining 2% PtdIns(3)P. This binding washighly specific, as the FYVE fusion proteinsdid not bind to liposomes containing simi-lar amounts of a range of phosphinositides(Fig. 2a).
Neither GST alone nor GST fused withthe zinc finger of Rabphilin-3A (this zincfinger is related to but distinct from theFYVE finger2,8) bound to PtdIns(3)P. Thiswas also true for zinc-binding-defectiveFYVE-finger mutants of EEA1 and Hrs(Fig. 2b). Pretreatment of the wild-typeGST–EEA1–FYVE construct with EDTA toremove Zn2& strongly reduced its ability tobind to PtdIns(3)P. These results show thatthe EEA1 and Hrs FYVE fingers binddirectly and specifically to PtdIns(3)P, andthat bound Zn2& is needed for this activity.
Using a different lipid-binding assay,Patki et al.9 obtained similar results withEEA1. Thus the binding of FYVE fingers toPtdIns(3)P seems to be affected little by theoverall lipid composition of the membrane.
The FYVE-finger motif is highly con-
FFiigguurree 11 The endosomal localization of EEA1-CT and its Hrs FYVE-finger hybrid depends on PI(3)K activity andan intact FYVE finger. BHK21 cells were co-transfected with Rab5 and Myc-epitope-tagged EEA11257-1411 (a–d),Myc–EEA11257-1411
C1405S (e–f), Myc–EEA11257-1349/Hrs161-221 (g–j) and Myc–EEA11257-1249/Hrs161-223C215S (k, l) and then incu-
bated for 20 minutes at 37 7C in the absence (a, b, e–h, k, l) or presence (c, d and i, j) of 20 nM wortman-nin. The cells were permeabilized with 0.05% saponin to wash out cytosolic protein, then fixed and finallystained with anti-Rab5 (a, c, e, g, i, k) or anti-Myc-epitope (b, d, f, h, j, l) antibodies and examined by con-focal immunofluorescence microscopy5. a shows the same cell as b, c shows the same cell as d, and soon. Arrows indicate examples of co-localization of Rab5 with epitope-tagged protein. Scale bar, 10 mm.
Nature © Macmillan Publishers Ltd 1998
8
scientific correspondence
NATURE | VOL 394 | 30 JULY 1998 433
learn whether PtdIns(3)P and FYVE fingershave other cellular functions as well.Jean-Michel Gaullier, Anne Simonsen,Antonello D’Arrigo, Bjørn Bremnes,Harald StenmarkDepartment of Biochemistry,The Norwegian Radium Hospital,N-0310 Oslo, Norwaye-mail: [email protected] AaslandDepartment of Molecular Biology, University ofBergen, N-5020 Bergen, Norway
1. Toker, A. & Cantley, L. C. Nature 387, 673–676 (1997).
2. Stenmark, H., Aasland, R., Toh, B. H. & D’Arrigo, A. J. Biol.
Chem. 271, 24048–24054 (1996).
3. Mu, F. T. et al. J. Biol. Chem. 270, 13503–13511 (1995).
4. Patki, V., Virbasius, J., Lane, W. S., Toh, B. H., Shpetner, H. S. &
Corvera, S. Proc. Natl Acad. Sci. USA 94, 7326–7330 (1997).
5. Simonsen, A. et al. Nature 394, 494–498 (1998).
6. Komada, M., Masaki, R., Yamamoto, A. & Kitamura, N. J. Biol.
Chem. 272, 20538–20544 (1997).
7. Schiavo, G., Gu, Q.-M., Prestwich, G. D., Söllner, T. H. &
Rothman, J. E. Proc. Natl Acad. Sci. USA 93, 13327–13332
(1996).
8. Shirataki, H. et al. Mol. Cell. Biol. 13, 2061–2068 (1993).
9. Patki, V., Virbasius, J. V., Chawla, A., Lawe, D.C. & Corvera, S.
Nature 394, 433–434 (1998).
10.Piper, R. C., Cooper, A. A., Yang, H. & Stevens, T. H. J. Cell Biol.
131, 603–617 (1995).
11.Burd, C. G., Peterson, M., Cowles, C. R. & Emr, S. D. Mol. Biol.
Cell 8, 1089–1104 (1997).
12.Dove, S. K. et al. Nature 390, 187–192 (1997).
Supplementary information is available on Nature’s World-Wide
Web site (http://www.nature.com) or as paper copy from the
London editorial office of Nature.
[3 H]-
phos
phol
ipid
bin
ding
(pe
r ce
nt o
f tot
al)
20
15
10
5
01 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8 1 2 3 4 5 7 86
GST-EEA1 GST-Hrs GST
20
15
10
5
0
a
b
GS
T-E
EA
1
GS
T-E
EA
1+E
DTA
1m
M
GS
T-E
EA
1
GS
T-H
rs
GS
T-H
rs
GS
T-R
abph
ilin
GS
T-E
EA
1+E
DTA
5m
M
C14
05S
C21
5S
FFiigguurree 22 FYVE domains bind specifically toPtdIns(3)P in vitro. Portions (0.5 nmol) of GST alone or fused to EEA11307-1411, Hrs1-289, EEA11307-1411
C1405S,Hrs1-289
C215S or Rabphilin45-182 were immobilized onglutathione–Sepharose beads, and then incu-bated in the presence of [3H]phosphatidylcholine-containing liposomes (0.35 mg ml11) containing 2%of: a, the indicated inositol lipids (see below) or b,PtdIns(3)P. Where indicated, the immobilized proteinhad been preincubated with EDTA. All values repre-sent the mean5s.e.m. of three experiments per-formed in triplicate. 1, phosphatidylcholine; 2,phosphatidylinositol; 3, PtdIns(3)P; 4, phosphatidyl-inositol-3,4-bisphosphate; 5, phosphatidylinositol-3,5-bisphosphate; 6, phosphatidylinositol-3,4,5-tris-phosphate; 7, phosphatidylinositol-4-phosphate; 8,phosphatidylinositol-4,5-bisphosphate.
Treating cells with the phosphatidylinositol-3-OH kinase (PI(3)K) inhibitor wortman-nin causes the dissociation of theearly-endosomal antigen EEA1 from earlyendosomes1. EEA1 from cytosolic extractsbinds to liposomes containing phos-phatidylinositol-3-phosphate (PtdIns(3)P),the major product of PI(3)K in yeast andmammalian cells1,2. Here we show that aRING zinc-finger domain at the carboxyterminus of EEA1, previously identified andnamed the ‘FYVE’ domain3, binds directlyand specifically to PtdIns(3)P. This indi-cates that proteins containing this motif
A functional PtdIns(3)P-binding motif
EEA1-CGFP
EEA1
EEA1-CGFP EEA1
Max
imal
(%)
100
80
60
40
20
0
PS
PtdI
ns(3
)P
PtdI
ns(3
,4,5
)P3
PtdI
ns(3
,4)P
2
PS/P
I
PS/P
IPt
dIns
(4,5
)P2
PtdI
ns(3
)P
PtdI
ns(3
)P
PtdI
ns(4
)P
PS PS/P
I
ApoRINGRING
GRP1-PH
Pel
let/s
uper
nata
nt
12
10
8
6
4
2
00 0.2 0.4 0.6 0.8 1.0
70
60
50
40
30
10
0
20
PI(3)P(µmoles)
PI(3)P(µmoles)
RING-GST
0 0.1 0.2 0.5 1.0
a b c
P S P S P S P S P S
RIN
G-G
ST
(nm
oles
)
FFiigguurree 11 Liposome-binding assays demonstrate the specificity and affinity of EEA1–PtdIns(3) interaction. a,Binding of a fusion protein containing the carboxy terminus of EAA-1 to liposomes. The RING–GST fusionprotein (amino acids 1,277–1,411 of EEA1 constitute the RING domain) was generated using amplification byPCR of the full-length complementary DNA. Liposomes were composed of phosphatidylserine and phos-phatidylinositol (PS/PI) and PtdIns(3)P (PI(3)P) (0–1% of the total lipid, at the expense of phosphatidylinositol,which decreased from 50% to 49%). Liposomes (100 mmoles total lipid) were incubated for 15 min at roomtemperature with 3 mg RING–GST in a total volume of 150 ml (ref. 1). After centrifugation, liposome pellets (P)and supernatants (S) were analysed by SDS–polyacrylamide gel electrophoresis and Coomassie blue stain-ing (inset). Values shown are derived from densitometry of stained bands. b, Binding of EAA1 to PtdIns(3)P isspecific and requires zinc. A GST fusion protein containing the GRP-1 pleckstrin homology domain was gen-erated as described10. ApoRING was generated by incubation of RING–GST with 50 molar equivalents ofmethylmethene thiosulphonate, followed by dialysis. Liposomes were composed of PS/PI, and extra phos-phoinositides constituted 0.5% of the total lipid at the expense of phosphatidylinositol. Plotted are the ratiosof GST fusion protein found in the pellets and supernatants after centrifugation. c, A GFP–EAA1 chimaera isnot specific for PtdIns(3)P. An oligonucleotide encoding GFP was inserted into a unique Not I site created inthe full-length complementary DNA for EEA1. Cytosolic extracts1 from Cos-1 cells transfected with EEA1–GFPwere treated with wortmannin to inhibit endogenous PI(3)K, and incubated with liposomes as in b. Pelletswere analysed by immunoblotting with an anti-EEA1 antiserum. Bands corresponding to endogenous, wild-type EEA1 and to the GFP–EEA1 chimaera are indicated. The intensity of the bands was plotted.
may be downstream effectors of PI(3)K inyeast and mammalian cells.
The RING finger of EAA1 exhibitsextensive homology to domains in Vac1pand Vps27p, two proteins that may beinvolved in the Vps34p-dependent vacuo-lar-protein-sorting pathway in yeast4,5, andin Hrs-2 and FGD-1, proteins involved insignal transduction and in actin-cytoskele-tal rearrangements in mammalian cells6,7.
PI(3)K activity may be involved in manycellular functions, including mitogenesis,protection from apoptosis, actin-filamentorganization and protein trafficking. Inyeast, the only detectable PI(3)K is Vps34p,which catalyses the conversion of phos-phatidylinositol to PtdIns(3)P (ref. 8).Although in mammalian cells there arePI(3)K enzymes that catalyse the formationof phosphatidylinositol-3,4-bisphosphateand phosphatidylinositol-3,4,5-trisphos-phate9, PtdIns(3)P is the most abundantPI(3)K product in these cells too2.
We have shown previously that the bind-ing of EEA1 to endosomal membranesdepends on the presence of PtdIns(3)P(ref. 1). To determine whether a directinteraction occurs between EEA1 andPtdIns(3)P, we measured the binding offusion proteins of EEA1 and glutathione S-transferase (GST) to liposomes. A fusionprotein containing the carboxy-terminal





























