Embed Size (px)
Citation preview

NATURE BIOTECHNOLOGY VOL 17 DECEMBER 1999 http://biotech.nature.com 1223
RESEARCH
The use of cysteine proteinase inhibitorsto engineer resistance against potyviruses
in transgenic tobacco plantsRafael Gutierrez-Campos 1, Juan Antonio Torres-Acosta1, Luis Jorge Saucedo-Arias 2,
and Miguel Angel Gomez-Lim2*
1Departamento de Química, Centro Básico, Universidad Autónoma de Aguascalientes, Ave. Universidad 940, C.P. 20 100, Aguascalientes, Ags. México.2Departamento de Ingeniería Genética, Centro de Investigación y Estudios Avanzados, Unidad Irapuato, Apartado Postal 629, Irapuato, Mexico.
*Corresponding author ([email protected]).
Received 19 March 1999; accepted 23 September 1999
As the processing mechanism of all known potyviruses involves the activity of cysteine proteinases, weasked whether constitutive expression of a rice cysteine proteinase inhibitor gene could induce resistanceagainst two important potyviruses, tobacco etch virus (TEV) and potato virus Y (PVY), in transgenic tobaccoplants. Tobacco lines expressing the foreign gene at varying levels were examined for resistance against TEVand PVY infection. There was a clear, direct correlation between the level of oryzacystatin message, inhibi-tion of papain (a cysteine proteinase), and resistance to TEV and PVY in all lines tested. The inhibitor was inef-fective against tobacco mosaic virus (TMV) infection because processing of this virus does not involve cys-teine proteinases. These results show that plant cystatins can be used against different potyviruses andpotentially also against other viruses, whose replication involves cysteine proteinase activity.
Keywords: genetic engineering, transgenic plants, potyvirus, tobacco etch virus, tobacco, proteinase inhibitors, virus resistance
Potyviruses comprise a large and agriculturally important group ofaphid-transmissible plant viruses1. Conventional breeding programsto develop resistance against potyviruses are effective but also pro-tracted and expensive. An alternative approach is to use the tech-niques of plant genetic transformation. Resistance has been achievedby transformation with natural viral resistance genes, antiviral pro-teins, or sense and antisense RNAs corresponding to segments ofviral genomes2. Pathogen-derived resistance (PDR), which involvestransformation with genes derived from the pathogen itself, has pre-viously been used to induce resistance to potyvirus infection3–5.However, the effectiveness of this strategy is limited by the specifici-ty for the viral strain from which the transgene was isolated, thelarge number of different potyviruses, and the genetic variationwithin a particular potyvirus2.
A novel approach to control viral diseases may involve the use ofproteinase inhibitors such as cysteine proteinase inhibitors or cys-tatins. Cystatins inhibit sulfhydryl proteinase activities and havemainly been studied in animal cells, in which three major familieshave been categorized based on sequence similarities: stefins, cys-tatins, and kininogens6. Although cystatins have been found in sev-eral plant tissues, such as seeds7, tubers8, and ripening fruits9, thephysiological function of these proteins is not well understood. Atleast two functions have been proposed: regulation of proteinturnover and host plant defense against insect predation10. In thisrespect, it has been demonstrated that plant cystatins inhibit diges-tive cysteine proteinases from insect guts and therefore possessinsecticidal activity11,12. Transgenic plants expressing cystatins fromcorn and rice (oryzacystatin I)13,14 showed an enhanced resistanceagainst insect gut proteinases and plant colonization by the parasitenematode Globodera pallida15. Oryzacystatin I also seems to protectagainst herpes simplex virus type 1, both in vitro and in vivo16.
The potyvirus genome encodes a polyprotein that is processed byproteolysis into individual gene products17. Three proteinases have
been shown to be involved in the processing mechanism, and at leastone of them (helper-component proteinase, HC-Pro) seems to be ofthe cysteine type18,19. An essential step for the replication ofpotyviruses is the activity of proteases identified as cysteine pro-teinases20, which process the 346 kDa polyprotein. We hypothesizedthat as cystatins are very powerful and specific inhibitors of cysteineproteinases21, they might provide a novel and specific method tocontrol virus infection. In the current work, we transformed tobaccoplants with the oryzacystatin gene and then challenged the trans-genic plants with tobacco etch virus (TEV) and potato virus Y(PVY), two important members of the potyvirus group.
Results and discussionConstruction of the chimeric gene and plant transformation. Weconstructed a chimeric gene with the coding sequence of the Oryzasativa cystatin (oryzacystatin I) gene22 fused to the 35S promoterfrom the cauliflower mosaic virus and nos 3´-end sequences (35S-oryzacystatin). Tobacco (Nicotiana tabacum L. var. Xanthi) leaf disksgrown axenically were cocultivated with Agrobacterium tumefaciensstrain pGV2260 containing the 35S-oryzacystatin gene23.Transformed tissue was selected for kanamycin resistance and regen-erated into plants23. To reduce the possibility of selecting chimericplants, leaf disks were taken from kanamycin-resistant plants, andwhole plants were regenerated from them in the presence of a higher(double) concentration of kanamycin. As a preliminary verificationthat the plants contained exogenous DNA, we extracted total DNAand performed PCR using primers deduced from the neomycinphosphotransferase II (NPTII) sequence24. Those lines from which a700 bp fragment was amplified and whose identity as NPTII was ver-ified by sequencing, were selected (data not shown).
Detection and expression of the oryzacystatin gene in trans-genic plants. The presence of the oryzacystatin gene in the selectedPCR-positive primary transformants (R0) was verified in 11 inde-
© 1999 Nature America Inc. • http://biotech.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://b
iote
ch.n
atu
re.c
om
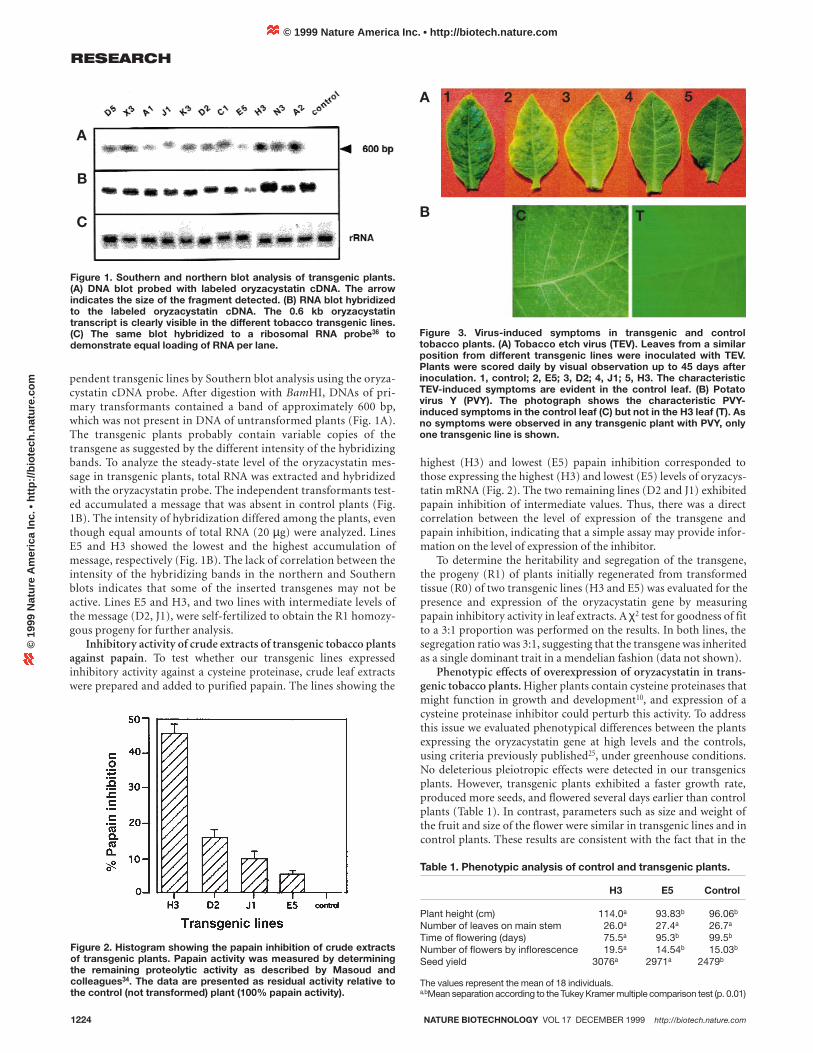
pendent transgenic lines by Southern blot analysis using the oryza-cystatin cDNA probe. After digestion with BamHI, DNAs of pri-mary transformants contained a band of approximately 600 bp,which was not present in DNA of untransformed plants (Fig. 1A).The transgenic plants probably contain variable copies of thetransgene as suggested by the different intensity of the hybridizingbands. To analyze the steady-state level of the oryzacystatin mes-sage in transgenic plants, total RNA was extracted and hybridizedwith the oryzacystatin probe. The independent transformants test-ed accumulated a message that was absent in control plants (Fig.1B). The intensity of hybridization differed among the plants, eventhough equal amounts of total RNA (20 µg) were analyzed. LinesE5 and H3 showed the lowest and the highest accumulation ofmessage, respectively (Fig. 1B). The lack of correlation between theintensity of the hybridizing bands in the northern and Southernblots indicates that some of the inserted transgenes may not beactive. Lines E5 and H3, and two lines with intermediate levels ofthe message (D2, J1), were self-fertilized to obtain the R1 homozy-gous progeny for further analysis.
Inhibitory activity of crude extracts of transgenic tobacco plantsagainst papain. To test whether our transgenic lines expressedinhibitory activity against a cysteine proteinase, crude leaf extractswere prepared and added to purified papain. The lines showing the
highest (H3) and lowest (E5) papain inhibition corresponded tothose expressing the highest (H3) and lowest (E5) levels of oryzacys-tatin mRNA (Fig. 2). The two remaining lines (D2 and J1) exhibitedpapain inhibition of intermediate values. Thus, there was a directcorrelation between the level of expression of the transgene andpapain inhibition, indicating that a simple assay may provide infor-mation on the level of expression of the inhibitor.
To determine the heritability and segregation of the transgene,the progeny (R1) of plants initially regenerated from transformedtissue (R0) of two transgenic lines (H3 and E5) was evaluated for thepresence and expression of the oryzacystatin gene by measuringpapain inhibitory activity in leaf extracts. A χ2 test for goodness of fitto a 3:1 proportion was performed on the results. In both lines, thesegregation ratio was 3:1, suggesting that the transgene was inheritedas a single dominant trait in a mendelian fashion (data not shown).
Phenotypic effects of overexpression of oryzacystatin in trans-genic tobacco plants. Higher plants contain cysteine proteinases thatmight function in growth and development10, and expression of acysteine proteinase inhibitor could perturb this activity. To addressthis issue we evaluated phenotypical differences between the plantsexpressing the oryzacystatin gene at high levels and the controls,using criteria previously published25, under greenhouse conditions.No deleterious pleiotropic effects were detected in our transgenicsplants. However, transgenic plants exhibited a faster growth rate,produced more seeds, and flowered several days earlier than controlplants (Table 1). In contrast, parameters such as size and weight ofthe fruit and size of the flower were similar in transgenic lines and incontrol plants. These results are consistent with the fact that in the
1224 NATURE BIOTECHNOLOGY VOL 17 DECEMBER 1999 http://biotech.nature.com
RESEARCH
Table 1. Phenotypic analysis of control and transgenic plants.
H3 E5 Control
Plant height (cm) 114.0a 93.83b 96.06b
Number of leaves on main stem 26.0a 27.4a 26.7a
Time of flowering (days) 75.5a 95.3b 99.5b
Number of flowers by inflorescence 19.5a 14.54b 15.03b
Seed yield 3076a 2971a 2479b
The values represent the mean of 18 individuals.a,bMean separation according to the Tukey Kramer multiple comparison test (p. 0.01)
Figure 1. Southern and northern blot analysis of transgenic plants.(A) DNA blot probed with labeled oryzacystatin cDNA. The arrowindicates the size of the fragment detected. (B) RNA blot hybridizedto the labeled oryzacystatin cDNA. The 0.6 kb oryzacystatintranscript is clearly visible in the different tobacco transgenic lines.(C) The same blot hybridized to a ribosomal RNA probe36 todemonstrate equal loading of RNA per lane.
A
B
C
Figure 2. Histogram showing the papain inhibition of crude extractsof transgenic plants. Papain activity was measured by determiningthe remaining proteolytic activity as described by Masoud andcolleagues34. The data are presented as residual activity relative tothe control (not transformed) plant (100% papain activity).
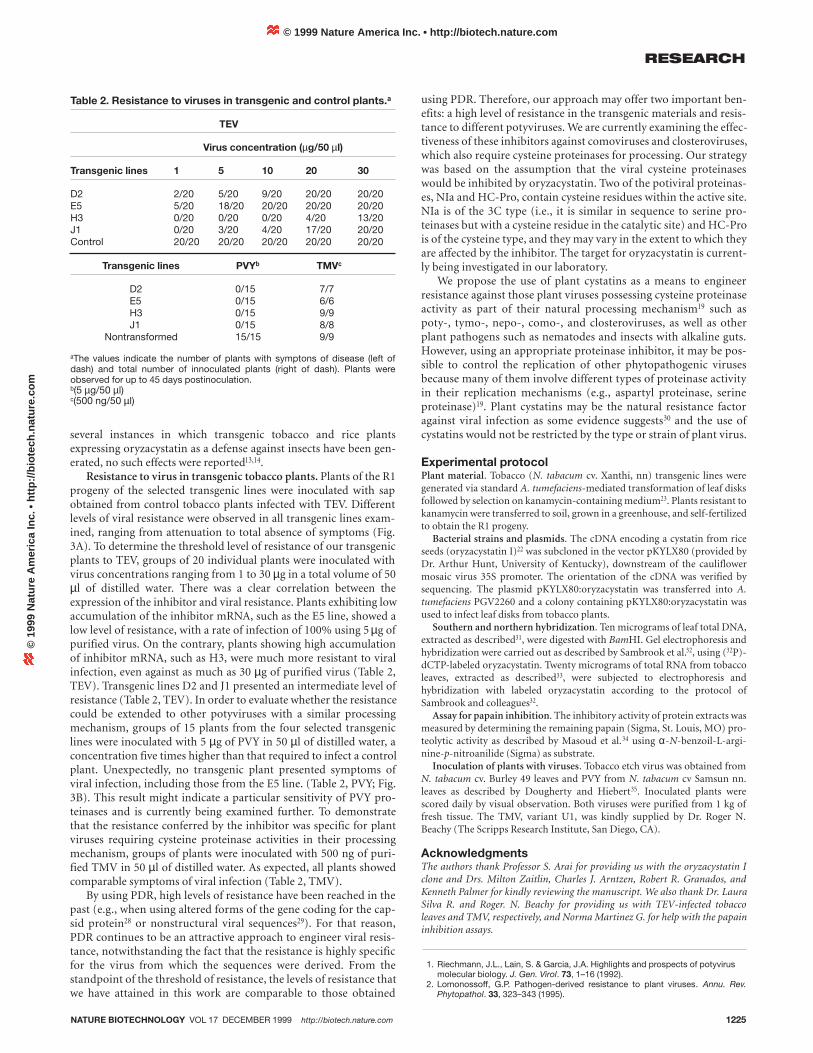
Figure 3. Virus-induced symptoms in transgenic and controltobacco plants. (A) Tobacco etch virus (TEV). Leaves from a similarposition from different transgenic lines were inoculated with TEV.Plants were scored daily by visual observation up to 45 days afterinoculation. 1, control; 2, E5; 3, D2; 4, J1; 5, H3. The characteristicTEV-induced symptoms are evident in the control leaf. (B) Potatovirus Y (PVY). The photograph shows the characteristic PVY-induced symptoms in the control leaf (C) but not in the H3 leaf (T). Asno symptoms were observed in any transgenic plant with PVY, onlyone transgenic line is shown.
1 2 3 4 5
C TB
A
© 1999 Nature America Inc. • http://biotech.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://b
iote
ch.n
atu
re.c
om
NATURE BIOTECHNOLOGY VOL 17 DECEMBER 1999 http://biotech.nature.com 1225
several instances in which transgenic tobacco and rice plantsexpressing oryzacystatin as a defense against insects have been gen-erated, no such effects were reported13,14.
Resistance to virus in transgenic tobacco plants. Plants of the R1progeny of the selected transgenic lines were inoculated with sapobtained from control tobacco plants infected with TEV. Differentlevels of viral resistance were observed in all transgenic lines exam-ined, ranging from attenuation to total absence of symptoms (Fig.3A). To determine the threshold level of resistance of our transgenicplants to TEV, groups of 20 individual plants were inoculated withvirus concentrations ranging from 1 to 30 µg in a total volume of 50µl of distilled water. There was a clear correlation between theexpression of the inhibitor and viral resistance. Plants exhibiting lowaccumulation of the inhibitor mRNA, such as the E5 line, showed alow level of resistance, with a rate of infection of 100% using 5 µg ofpurified virus. On the contrary, plants showing high accumulationof inhibitor mRNA, such as H3, were much more resistant to viralinfection, even against as much as 30 µg of purified virus (Table 2,TEV). Transgenic lines D2 and J1 presented an intermediate level ofresistance (Table 2, TEV). In order to evaluate whether the resistancecould be extended to other potyviruses with a similar processingmechanism, groups of 15 plants from the four selected transgeniclines were inoculated with 5 µg of PVY in 50 µl of distilled water, aconcentration five times higher than that required to infect a controlplant. Unexpectedly, no transgenic plant presented symptoms ofviral infection, including those from the E5 line. (Table 2, PVY; Fig.3B). This result might indicate a particular sensitivity of PVY pro-teinases and is currently being examined further. To demonstratethat the resistance conferred by the inhibitor was specific for plantviruses requiring cysteine proteinase activities in their processingmechanism, groups of plants were inoculated with 500 ng of puri-fied TMV in 50 µl of distilled water. As expected, all plants showedcomparable symptoms of viral infection (Table 2, TMV).
By using PDR, high levels of resistance have been reached in thepast (e.g., when using altered forms of the gene coding for the cap-sid protein28 or nonstructural viral sequences29). For that reason,PDR continues to be an attractive approach to engineer viral resis-tance, notwithstanding the fact that the resistance is highly specificfor the virus from which the sequences were derived. From thestandpoint of the threshold of resistance, the levels of resistance thatwe have attained in this work are comparable to those obtained
using PDR. Therefore, our approach may offer two important ben-efits: a high level of resistance in the transgenic materials and resis-tance to different potyviruses. We are currently examining the effec-tiveness of these inhibitors against comoviruses and closteroviruses,which also require cysteine proteinases for processing. Our strategywas based on the assumption that the viral cysteine proteinaseswould be inhibited by oryzacystatin. Two of the potiviral proteinas-es, NIa and HC-Pro, contain cysteine residues within the active site.NIa is of the 3C type (i.e., it is similar in sequence to serine pro-teinases but with a cysteine residue in the catalytic site) and HC-Prois of the cysteine type, and they may vary in the extent to which theyare affected by the inhibitor. The target for oryzacystatin is current-ly being investigated in our laboratory.
We propose the use of plant cystatins as a means to engineerresistance against those plant viruses possessing cysteine proteinaseactivity as part of their natural processing mechanism19 such aspoty-, tymo-, nepo-, como-, and closteroviruses, as well as otherplant pathogens such as nematodes and insects with alkaline guts.However, using an appropriate proteinase inhibitor, it may be pos-sible to control the replication of other phytopathogenic virusesbecause many of them involve different types of proteinase activityin their replication mechanisms (e.g., aspartyl proteinase, serineproteinase)19. Plant cystatins may be the natural resistance factoragainst viral infection as some evidence suggests30 and the use ofcystatins would not be restricted by the type or strain of plant virus.
Experimental protocolPlant material. Tobacco (N. tabacum cv. Xanthi, nn) transgenic lines weregenerated via standard A. tumefaciens-mediated transformation of leaf disksfollowed by selection on kanamycin-containing medium23. Plants resistant tokanamycin were transferred to soil, grown in a greenhouse, and self-fertilizedto obtain the R1 progeny.
Bacterial strains and plasmids. The cDNA encoding a cystatin from riceseeds (oryzacystatin I)22 was subcloned in the vector pKYLX80 (provided byDr. Arthur Hunt, University of Kentucky), downstream of the cauliflowermosaic virus 35S promoter. The orientation of the cDNA was verified bysequencing. The plasmid pKYLX80:oryzacystatin was transferred into A.tumefaciens PGV2260 and a colony containing pKYLX80:oryzacystatin wasused to infect leaf disks from tobacco plants.
Southern and northern hybridization. Ten micrograms of leaf total DNA,extracted as described31, were digested with BamHI. Gel electrophoresis andhybridization were carried out as described by Sambrook et al.32, using (32P)-dCTP-labeled oryzacystatin. Twenty micrograms of total RNA from tobaccoleaves, extracted as described33, were subjected to electrophoresis andhybridization with labeled oryzacystatin according to the protocol ofSambrook and colleagues32.
Assay for papain inhibition. The inhibitory activity of protein extracts wasmeasured by determining the remaining papain (Sigma, St. Louis, MO) pro-teolytic activity as described by Masoud et al.34 using α-N-benzoil-L-argi-nine-p-nitroanilide (Sigma) as substrate.
Inoculation of plants with viruses. Tobacco etch virus was obtained fromN. tabacum cv. Burley 49 leaves and PVY from N. tabacum cv Samsun nn.leaves as described by Dougherty and Hiebert35. Inoculated plants werescored daily by visual observation. Both viruses were purified from 1 kg offresh tissue. The TMV, variant U1, was kindly supplied by Dr. Roger N.Beachy (The Scripps Research Institute, San Diego, CA).
AcknowledgmentsThe authors thank Professor S. Arai for providing us with the oryzacystatin Iclone and Drs. Milton Zaitlin, Charles J. Arntzen, Robert R. Granados, andKenneth Palmer for kindly reviewing the manuscript. We also thank Dr. LauraSilva R. and Roger. N. Beachy for providing us with TEV-infected tobaccoleaves and TMV, respectively, and Norma Martinez G. for help with the papaininhibition assays.
1. Riechmann, J.L., Lain, S. & Garcia, J.A. Highlights and prospects of potyvirusmolecular biology. J. Gen. Virol. 73, 1–16 (1992).
2. Lomonossoff, G.P. Pathogen-derived resistance to plant viruses . Annu. Rev.Phytopathol. 33, 323–343 (1995).
RESEARCH
Table 2. Resistance to viruses in transgenic and control plants.a
TEV
Virus concentration (µg/50 µl)
Transgenic lines 1 5 10 20 30
D2 2/20 5/20 9/20 20/20 20/20E5 5/20 18/20 20/20 20/20 20/20H3 0/20 0/20 0/20 4/20 13/20J1 0/20 3/20 4/20 17/20 20/20Control 20/20 20/20 20/20 20/20 20/20
Transgenic lines PVYb TMVc
D2 0/15 7/7E5 0/15 6/6H3 0/15 9/9J1 0/15 8/8
Nontransformed 15/15 9/9
aThe values indicate the number of plants with symptons of disease (left ofdash) and total number of innoculated plants (right of dash). Plants wereobserved for up to 45 days postinoculation.b(5 µg/50 µl)c(500 ng/50 µl)
© 1999 Nature America Inc. • http://biotech.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://b
iote
ch.n
atu
re.c
om

3. Ling, K., Namba,S. & Gonsalvez, D. Protection against detrimental effects ofpotyvirus infection in transgenic tobacco plants expressing the papaya ringspotvirus coat protein gene. Bio/Technology 9, 752–758 (1991).
4. Regner, F., da Camara Machada, A., Steinkellner, H. & Mattanovih, D. Coat pro-tein resistance to plum pox potyvirus in Nicotiana clevelandii and Nicotiana ben-thamiana. Plant Cell Reports 11, 30–33 (1992).
5. Audy, P., Palutaikis, P., Slack, S.A. & Zaitlin, M. Replicase-mediated resistance topotato virus Y in transgenic tobacco plants. Mol. Plant-Microbe Interact. 7,15–22 (1993).
6. Barrett, A.J. The cystatins: a new class of peptidase inhibitors. Trends Biochem.Sci. 12, 193–196 (1987).
7. Fernandes, K.S.V. et al. The resistance of cowpea seeds to bruchid beetles is notrelated to levels of cysteine proteinase inhibitors. Plant Mol. Biol. 23, 215–219(1993).
8. Waldron, C., Wegrich, L.M., Merlo, P.A.O. & Walsh, T.A. Characterization of agenomic sequence coding for potato multicystatin, an eight-domain cysteineproteinase inhibitor. Plant Mol. Biol. 23, 801–812 (1993).
9. Dopico, B., Lowe, A.L., Wilson, I.D., Merodio, C. & Grierson, D. Cloning and char-acterization of avocado fruit mRNAs and their expression during ripening andlow-temperature storage. Plant Mol. Biol. 21, 437–449 (1993).
10. Turk, V. & Bode, W. The cystatins: protein inhibitors of cysteine proteinases.FEBS Lett. 285, 213–219 (1991).
11. Heins, M.E., Osuala, C.I. & Nielsen, S.S. Isolation and partial characterization of asoybean cystatin cysteine proteinase inhibitor of coleopteran digestive proteolyt-ic activity. J. Agric. Food Chem. 39, 1515–1520 (1991).
12. Orr, G.L., Strickland, J.A. & Walsh, T.A. Inhibition of Diabrotica larval growth by amulticystatin from potato tubers. J. Insect Physiol. 40, 893–900 (1994).
13. Hosoyama, H., Irie, K., Abe, K. & Arai, S. Oryzacystatin exogenously introducedinto protoplasts and regeneration of transgenic rice. Biosci. Biotechnol.Biochem. 58, 1500–1506 (1992).
14. Irie, K. et al. Transgenic rice established to express corn cystatin exhibits stronginhibitory activity against insect gut proteinases. Plant Mol. Biol. 30, 149–157 (1996).
15. Urwin, P.E., Atkinson, H.J., Waller, D.A. & McPherson, M.J. Engineered oryzacys-tatin-I expressed in transgenic hairy roots confers resistance to Globodera palli-da. Plant J. 8, 121–131 (1995).
16. Aoki, H. et al. Antiviral effect of oryzacystatin, a proteinase inhibitor in rice,against herpes simplex virus type 1 in vitro and in vivo. Antimicrob. AgentsChemother. 39, 846–849 (1995).
17. Dougherty, W.G. & Carrington, J.C. Expression and function of potyviral geneproducts. Annu. Rev. Phytopathol. 26, 123–143 (1988).
18. Kasschau, K.D. & Carrington, J.C. Requirement for HC-Pro processing duringgenome amplification of tobacco etch potyvirus. Virology 209, 268–273 (1995).
19. Spall, V.E., Shanks, M. & Lomonossoff, G.P. Polyprotein processing as a strategy
for gene expression in RNA viruses. Semin. Virol. 8, 15–23 (1997).20. Dougherty, W.G. & Semler, B.L. Expression of virus-encoded proteinases: func-
tional and structural similarities with cellular enzymes. Microbiol. Rev. 57,781–822 (1993).
21. Machleidt, W., Assfalg-Machleidt, I. & Auerswald, A. in Innovations in proteasesand their inhibitors (ed. Avilés, F.X.) 349–367 (Walter de Gruyter, New York; 1993).
22. Abe, K., Emori, Y., Kondo, H., Suzuki, K. & Arai, S. Molecular cloning of a cysteineproteinase inhibitor of rice (oryzacystatin). Homology with animal cystatins andtransient expression in the ripening process of rice seeds. J. Biol. Chem. 262,16793–16797 (1987).
23. Horsh, R.B. et al. A simple and general method for transferring genes into plants.Science 227, 1229–1231 (1985).
24. Hamill, J.D., Rounsley, S., Spencer, A., Todd, G. & Rhodes, M.J.C. The use of thepolymerase chain reaction in plant transformation studies. Plant Cell Rep. 10,221–224 (1991).
25. Gan, S. & Amasino, R.M. Inhibition of leaf senescence by autoregulated produc-tion of cytokinin. Science 270, 1986–1988 (1995).
28. Lindbo, J.A. & Dougerthy, W.G. Pathogen-derived resistance to a potyvirus:immune and resistant phenotypes in transgenic tobacco expressing alteredforms of a potyvirus coat protein nucleotide sequence. Mol. Plant-MicrobeInteract. 2, 144–153 (1992).
29. Maiti, I.B., Murphy, J.F., Shaw, J.G., & Hunt, A.G. Plants that express a potyvirusproteinase gene are resistant to virus infection. Proc. Natl. Acad. Sci. USA 90,6110–6114 (1993).
30. Shih, D.S., Bu, M., Price, M.A. & Shih, C.T. Inhibition of cleavage of a plant viralpolyprotein by an inhibitor activity present in wheat germ and cowpea embryos.J. Virol. 61, 912–915 (1987).
31. Dellaporta, S.L., Wood, J., & Hicks, J.B. A plant DNA minipreparation: version II.Plant Mol. Biol. 1, 19–21 (1983).
32. Sambrook, J., Fritsch, E.F. & Maniatis T. Molecular cloning: a laboratory manual.2nd edn (Cold Spring Harbor Press, New York; 1989).
33. López-Gómez, R. & Gómez-Lim, M.A. A method for extracting intact RNA fromfruits rich in polysaccharides using ripe mango mesocarp. Hortscience 27,440–442 (1992).
34. Masoud, S.A., Johnson, L.B., White, F.F. & Reeck, G.R. Expression of a cysteineproteinase inhibitor (oryzacystatin-I) in transgenic tobacco plants. Plant Mol.Biol. 21, 655–663 (1993).
35. Dougherty, W.G. & Hiebert, E. Translation of potyvirus RNA in a rabbit reticulo-cyte lysate: reaction conditions and identification of capsid protein as one of theproducts of in vitro translation of tobacco etch and pepper mottle virus viralRNAs. Virology 101, 466–474 (1980).
36. Gerlach, W.L. & Bedbrook, J.R. Cloning and characterization of ribosomal RNAgenes from wheat and barley. Nucleic Acids Res. 7, 1869–1885 (1979).
1226 NATURE BIOTECHNOLOGY VOL 17 DECEMBER 1999 http://biotech.nature.com
RESEARCH
© 1999 Nature America Inc. • http://biotech.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://b
iote
ch.n
atu
re.c
om





























