Embed Size (px)
Citation preview

Murine genetic models of obesity, including Lepob/Lepob andmutations associated with ectopic expression of agouti (such asAy and Avy), have defects in energy homeostasis resulting from alack of coordination between energy intake and energy expen-diture1. The mechanism for the Lepob/Lepob animals is absenceof leptin, the hormone that communicates information to thebrain regarding the extent of long-term energy stores held in theform of adipose tissue. It has also been known for many yearsthat Ay and Avy mice are hyperphagic and hypometabolic2,3.These findings have been reproduced for the comparableMC4R–/– model4–6. The Ay and MC4R–/– mice, with time, devel-op resistance to centrally administered leptin7,8. However, as theprincipal cause for the development of obesity, this explanationis incomplete. For example, young, lean MC4R–/– mice stillrespond to the anorexigenic effects of leptin7, and MC4R–/– miceexhibit only a fraction of the neuroendocrine and energy home-ostasis phenotypes seen in Lepob/Lepob or Leprdb/Leprdb mice.Furthermore, the double Lepob/LepobAy/a mutant has a moreextreme obesity phenotype than either single mutant, indicat-ing leptin-independent roles for the melanocortin system9. What,then, are the normal roles for the melanocortinergic circuitryin the daily maintenance of energy homeostasis, and how dolesions in this circuitry lead to obesity?
Proopiomelanocortin (POMC) neurons in the arcuatenucleus of the hypothalamus (ARC) are a site of dense leptinreceptor expression10, and are important for leptin’s actionson both feeding behavior11 and brown fat thermogenesis12. Inaddition to expressing leptin receptors, POMC neuronsrespond to a variety of other factors, including dopamine,gonadal steroids, glucose and insulin13–15; this implies a broad-
Melanocortin-4 receptor is requiredfor acute homeostatic responses toincreased dietary fat
Andrew A. Butler1, Daniel L. Marks2, Wei Fan1, Cynthia M. Kuhn3, Maria Bartolome3 and RogerD. Cone1
1 Vollum Institute, Oregon Health Sciences University, 3181 SW Sam Jackson Park Road, Portland, Oregon 97201, USA2 Department of Pediatric Endocrinology, Oregon Health Sciences University CDRCP, Portland, Oregon 97201, USA3 Department of Pharmacology and Cancer Biology, Duke University Medical Center, Durham, North Carolina 27710, USA
Correspondence should be addressed to R.D.C. ([email protected])
In response to moderately increased dietary fat content, melanocortin-4 receptor-null mutant(MC4R–/–) mice exhibit hyperphagia and accelerated weight gain compared to wild-type mice. Anincreased feed efficiency (weight gain/kcal consumed) argues that mechanisms in addition to hyper-phagia are instrumental in causing weight gain. We report two specific defects in coordinatingenergy expenditure with food intake in MC4R–/– mice. Wild-type mice respond to an increase in thefat content of the diet by rapidly increasing diet-induced thermogenesis and by increasing physicalactivity, neither of which are observed in MC4R–/– mice. Leptin-deficient and MC3R–/– mice regulatemetabolic rate similarly to wild-type mice in this protocol. Melanocortinergic pathways involvingMC4-R-regulated neurons, which rapidly respond to signals not requiring changes in leptin, thusseem to be important in regulating metabolic and behavioral responses to dietary fat.
er role for these neurons in communicating peripheral infor-mation relevant to energy homeostasis to centers controllingfeeding and metabolism. For example, hypophagia associatedwith gastric infusion of a liquid diet is blocked by the intrac-erebroventricular administration of the melanocortin antago-nist SHU9119 (ref. 16). Together, these data suggest aninvolvement of the melanocortin system in the acute coordi-nation of feeding with energy expenditure over time periodsduring which leptin levels appear stable.
Previous studies have not used the MC4R–/– mouse toexamine the role of the MC4R in acutely integrating energyintake with each component of energy expenditure (basalmetabolism, diet-induced thermogenesis, activity); these stud-ies instead focused on the long-term consequences of manip-ulation of caloric intake or exposure to high-fat diets4,5,7. Toidentify specific roles of the MC4-R in coordinating energyintake and expenditure, we examined the feeding and specificenergy expenditure responses of MC4R–/– mice to a variety ofperturbations from homeostasis. These studies identify a spe-cific role for the MC4-R in acute ingestive, DIT and physicalactivity responses to increased caloric intake.
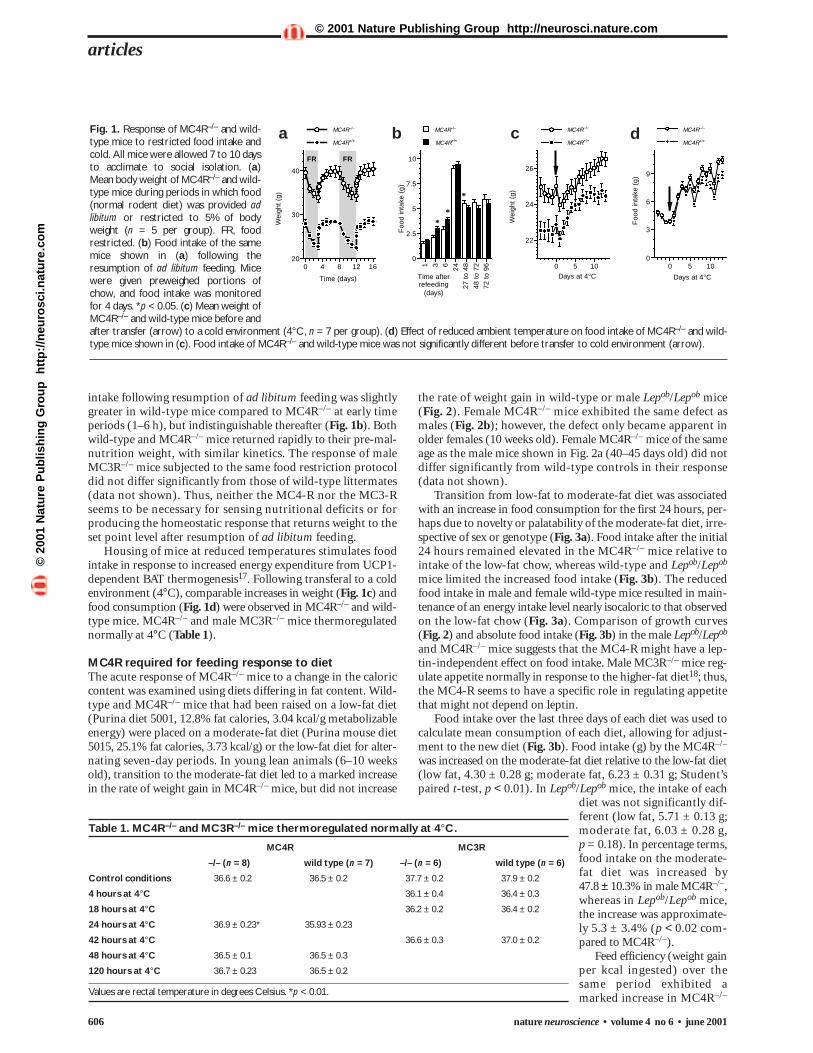
RESULTSNormal adaptation to cold and to food restrictionTo examine whether the MC4-R is necessary for sensing andresponding to nutritional deficits, we analyzed the effects oflimiting food intake to 5% of body weight in male MC4R–/–
mice and wild-type controls (Fig. 1). In terms of absoluteweight loss, the response of MC4R–/– and wild-type mice to 72 h or 96 h nutritional deprivation was similar (Fig. 1a). Food
articles
nature neuroscience • volume 4 no 6 • june 2001 605
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://n
euro
sci.n
atu
re.c
om
© 2001 Nature Publishing Group http://neurosci.nature.com
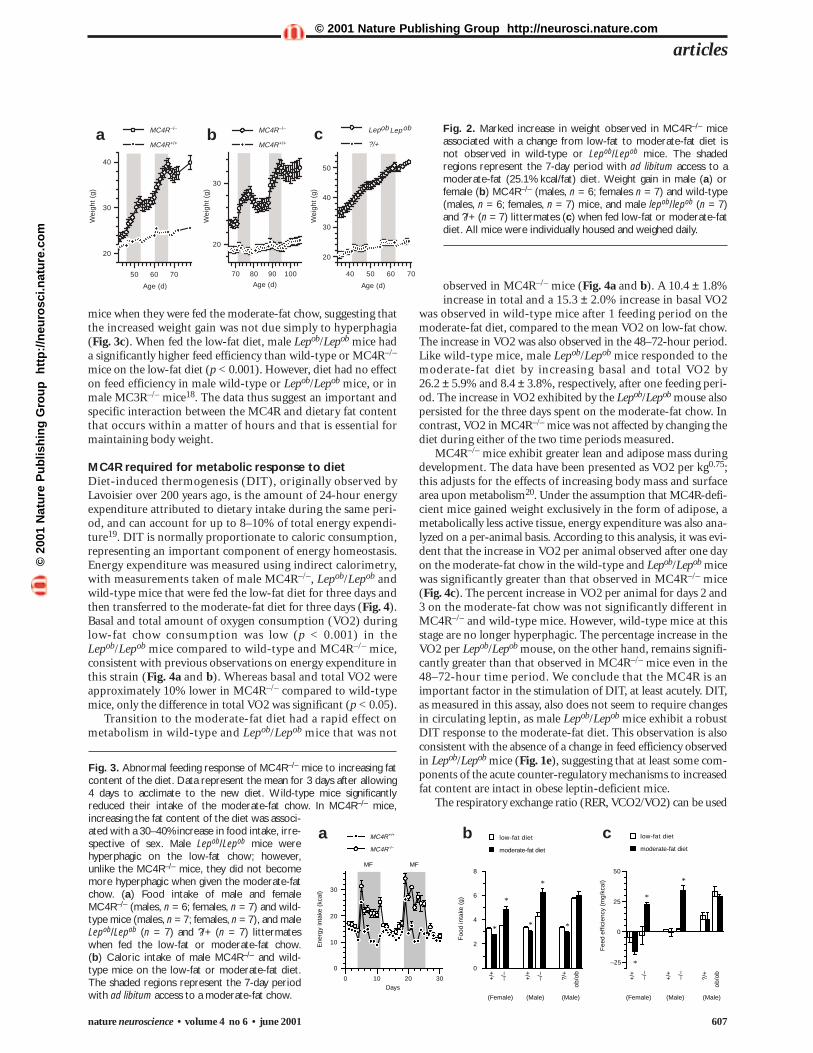
the rate of weight gain in wild-type or male Lepob/Lepob mice(Fig. 2). Female MC4R–/– mice exhibited the same defect asmales (Fig. 2b); however, the defect only became apparent inolder females (10 weeks old). Female MC4R–/– mice of the sameage as the male mice shown in Fig. 2a (40–45 days old) did notdiffer significantly from wild-type controls in their response(data not shown).
Transition from low-fat to moderate-fat diet was associatedwith an increase in food consumption for the first 24 hours, per-haps due to novelty or palatability of the moderate-fat diet, irre-spective of sex or genotype (Fig. 3a). Food intake after the initial24 hours remained elevated in the MC4R–/– mice relative tointake of the low-fat chow, whereas wild-type and Lepob/Lepob
mice limited the increased food intake (Fig. 3b). The reducedfood intake in male and female wild-type mice resulted in main-tenance of an energy intake level nearly isocaloric to that observedon the low-fat chow (Fig. 3a). Comparison of growth curves (Fig. 2) and absolute food intake (Fig. 3b) in the male Lepob/Lepob
and MC4R–/– mice suggests that the MC4-R might have a lep-tin-independent effect on food intake. Male MC3R–/– mice reg-ulate appetite normally in response to the higher-fat diet18; thus,the MC4-R seems to have a specific role in regulating appetitethat might not depend on leptin.
Food intake over the last three days of each diet was used tocalculate mean consumption of each diet, allowing for adjust-ment to the new diet (Fig. 3b). Food intake (g) by the MC4R–/–
was increased on the moderate-fat diet relative to the low-fat diet(low fat, 4.30 ± 0.28 g; moderate fat, 6.23 ± 0.31 g; Student’spaired t-test, p < 0.01). In Lepob/Lepob mice, the intake of each
diet was not significantly dif-ferent (low fat, 5.71 ± 0.13 g;moderate fat, 6.03 ± 0.28 g, p = 0.18). In percentage terms,food intake on the moderate-fat diet was increased by 47.8 ± 10.3% in male MC4R–/–,whereas in Lepob/Lepob mice,the increase was approximate-ly 5.3 ± 3.4% (p < 0.02 com-pared to MC4R–/–).
Feed efficiency (weight gainper kcal ingested) over thesame period exhibited amarked increase in MC4R–/–
intake following resumption of ad libitum feeding was slightlygreater in wild-type mice compared to MC4R–/– at early timeperiods (1–6 h), but indistinguishable thereafter (Fig. 1b). Bothwild-type and MC4R–/– mice returned rapidly to their pre-mal-nutrition weight, with similar kinetics. The response of maleMC3R–/– mice subjected to the same food restriction protocoldid not differ significantly from those of wild-type littermates(data not shown). Thus, neither the MC4-R nor the MC3-Rseems to be necessary for sensing nutritional deficits or forproducing the homeostatic response that returns weight to theset point level after resumption of ad libitum feeding.
Housing of mice at reduced temperatures stimulates foodintake in response to increased energy expenditure from UCP1-dependent BAT thermogenesis17. Following transferal to a coldenvironment (4°C), comparable increases in weight (Fig. 1c) andfood consumption (Fig. 1d) were observed in MC4R–/– and wild-type mice. MC4R–/– and male MC3R–/– mice thermoregulatednormally at 4°C (Table 1).
MC4R required for feeding response to dietThe acute response of MC4R–/– mice to a change in the caloriccontent was examined using diets differing in fat content. Wild-type and MC4R–/– mice that had been raised on a low-fat diet(Purina diet 5001, 12.8% fat calories, 3.04 kcal/g metabolizableenergy) were placed on a moderate-fat diet (Purina mouse diet5015, 25.1% fat calories, 3.73 kcal/g) or the low-fat diet for alter-nating seven-day periods. In young lean animals (6–10 weeksold), transition to the moderate-fat diet led to a marked increasein the rate of weight gain in MC4R–/– mice, but did not increase
Fig. 1. Response of MC4R–/– and wild-type mice to restricted food intake andcold. All mice were allowed 7 to 10 daysto acclimate to social isolation. (a)Mean body weight of MC4R–/– and wild-type mice during periods in which food(normal rodent diet) was provided adlibitum or restricted to 5% of bodyweight (n = 5 per group). FR, foodrestricted. (b) Food intake of the samemice shown in (a) following theresumption of ad libitum feeding. Micewere given preweighed portions ofchow, and food intake was monitoredfor 4 days. *p < 0.05. (c) Mean weight ofMC4R–/– and wild-type mice before andafter transfer (arrow) to a cold environment (4°C, n = 7 per group). (d) Effect of reduced ambient temperature on food intake of MC4R–/– and wild-type mice shown in (c). Food intake of MC4R–/– and wild-type mice was not significantly different before transfer to cold environment (arrow).
articles
606 nature neuroscience • volume 4 no 6 • june 2001
0
2.5
5
7.5
10
Foo
d in
take
(g)
1 3 6 24
27 t
o 48
48 t
o 72
72 t
o 96
Time afterrefeeding
(days)
MC4R–/–
20
30
40
Wei
ght
(g)
0 4 8 12 16
**
FR
*
FR
Time (days)
22
24
26
Wei
ght
(g)
0 5 10Days at 4°C
0
3
6
9
Foo
d in
take
(g)
0 5 10
Days at 4°C
MC4R+/+
MC4R–/–
MC4R+/+
MC4R–/–
MC4R+/+
MC4R–/–
MC4R+/+ a b c d
Table 1. MC4R–/– and MC3R–/– mice thermoregulated normally at 4°C.
MC4R MC3R
–/– (n = 8) wild type (n = 7) –/– (n = 6) wild type (n = 6)
Control conditions 36.6 ± 0.2 36.5 ± 0.2 37.7 ± 0.2 37.9 ± 0.2
4 hours at 4°C 36.1 ± 0.4 36.4 ± 0.3
18 hours at 4°C 36.2 ± 0.2 36.4 ± 0.2
24 hours at 4°C 36.9 ± 0.23* 35.93 ± 0.23
42 hours at 4°C 36.6 ± 0.3 37.0 ± 0.2
48 hours at 4°C 36.5 ± 0.1 36.5 ± 0.3
120 hours at 4°C 36.7 ± 0.23 36.5 ± 0.2
Values are rectal temperature in degrees Celsius. *p < 0.01.
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://n
euro
sci.n
atu
re.c
om
© 2001 Nature Publishing Group http://neurosci.nature.com
observed in MC4R–/– mice (Fig. 4a and b). A 10.4 ± 1.8%increase in total and a 15.3 ± 2.0% increase in basal VO2
was observed in wild-type mice after 1 feeding period on themoderate-fat diet, compared to the mean VO2 on low-fat chow.The increase in VO2 was also observed in the 48–72-hour period.Like wild-type mice, male Lepob/Lepob mice responded to themoderate-fat diet by increasing basal and total VO2 by 26.2 ± 5.9% and 8.4 ± 3.8%, respectively, after one feeding peri-od. The increase in VO2 exhibited by the Lepob/Lepob mouse alsopersisted for the three days spent on the moderate-fat chow. Incontrast, VO2 in MC4R–/– mice was not affected by changing thediet during either of the two time periods measured.
MC4R–/– mice exhibit greater lean and adipose mass duringdevelopment. The data have been presented as VO2 per kg0.75;this adjusts for the effects of increasing body mass and surfacearea upon metabolism20. Under the assumption that MC4R-defi-cient mice gained weight exclusively in the form of adipose, ametabolically less active tissue, energy expenditure was also ana-lyzed on a per-animal basis. According to this analysis, it was evi-dent that the increase in VO2 per animal observed after one dayon the moderate-fat chow in the wild-type and Lepob/Lepob micewas significantly greater than that observed in MC4R–/– mice(Fig. 4c). The percent increase in VO2 per animal for days 2 and3 on the moderate-fat chow was not significantly different inMC4R–/– and wild-type mice. However, wild-type mice at thisstage are no longer hyperphagic. The percentage increase in theVO2 per Lepob/Lepob mouse, on the other hand, remains signifi-cantly greater than that observed in MC4R–/– mice even in the48–72-hour time period. We conclude that the MC4R is animportant factor in the stimulation of DIT, at least acutely. DIT,as measured in this assay, also does not seem to require changesin circulating leptin, as male Lepob/Lepob mice exhibit a robustDIT response to the moderate-fat diet. This observation is alsoconsistent with the absence of a change in feed efficiency observedin Lepob/Lepob mice (Fig. 1e), suggesting that at least some com-ponents of the acute counter-regulatory mechanisms to increasedfat content are intact in obese leptin-deficient mice.
The respiratory exchange ratio (RER, VCO2/VO2) can be used
Fig. 2. Marked increase in weight observed in MC4R–/– miceassociated with a change from low-fat to moderate-fat diet isnot observed in wild-type or Lepob/Lepob mice. The shadedregions represent the 7-day period with ad libitum access to amoderate-fat (25.1% kcal/fat) diet. Weight gain in male (a) orfemale (b) MC4R–/– (males, n = 6; females n = 7) and wild-type(males, n = 6; females, n = 7) mice, and male lepob/lepob (n = 7)and ?/+ (n = 7) littermates (c) when fed low-fat or moderate-fatdiet. All mice were individually housed and weighed daily.
mice when they were fed the moderate-fat chow, suggesting thatthe increased weight gain was not due simply to hyperphagia(Fig. 3c). When fed the low-fat diet, male Lepob/Lepob mice hada significantly higher feed efficiency than wild-type or MC4R–/–
mice on the low-fat diet (p < 0.001). However, diet had no effecton feed efficiency in male wild-type or Lepob/Lepob mice, or inmale MC3R–/– mice18. The data thus suggest an important andspecific interaction between the MC4R and dietary fat contentthat occurs within a matter of hours and that is essential formaintaining body weight.
MC4R required for metabolic response to dietDiet-induced thermogenesis (DIT), originally observed byLavoisier over 200 years ago, is the amount of 24-hour energyexpenditure attributed to dietary intake during the same peri-od, and can account for up to 8–10% of total energy expendi-ture19. DIT is normally proportionate to caloric consumption,representing an important component of energy homeostasis.Energy expenditure was measured using indirect calorimetry,with measurements taken of male MC4R–/–, Lepob/Lepob andwild-type mice that were fed the low-fat diet for three days andthen transferred to the moderate-fat diet for three days (Fig. 4).Basal and total amount of oxygen consumption (VO2) duringlow-fat chow consumption was low (p < 0.001) in theLepob/Lepob mice compared to wild-type and MC4R–/– mice,consistent with previous observations on energy expenditure inthis strain (Fig. 4a and b). Whereas basal and total VO2 wereapproximately 10% lower in MC4R–/– compared to wild-typemice, only the difference in total VO2 was significant (p < 0.05).
Transition to the moderate-fat diet had a rapid effect onmetabolism in wild-type and Lepob/Lepob mice that was not
Fig. 3. Abnormal feeding response of MC4R–/– mice to increasing fatcontent of the diet. Data represent the mean for 3 days after allowing4 days to acclimate to the new diet. Wild-type mice significantlyreduced their intake of the moderate-fat chow. In MC4R–/– mice,increasing the fat content of the diet was associ-ated with a 30–40% increase in food intake, irre-spective of sex. Male Lepob/Lepob mice werehyperphagic on the low-fat chow; however,unlike the MC4R–/– mice, they did not becomemore hyperphagic when given the moderate-fatchow. (a) Food intake of male and femaleMC4R–/– (males, n = 6; females, n = 7) and wild-type mice (males, n = 7; females, n = 7), and maleLepob/Lepob (n = 7) and ?/+ (n = 7) littermateswhen fed the low-fat or moderate-fat chow. (b) Caloric intake of male MC4R–/– and wild-type mice on the low-fat or moderate-fat diet.The shaded regions represent the 7-day periodwith ad libitum access to a moderate-fat chow.
nature neuroscience • volume 4 no 6 • june 2001 607
articles
20
30
40
50
Wei
ght
(g)
40 50 60 70
Age (d)
?/+
Lepob ob
Wei
ght
(g)
20
30
70 80 90 100
Age (d)
20
30
40
Wei
ght
(g)
50 60 70
Age (d)
MC4R–/– MC4R–/–
MC4R+/+ MC4R+/+
Lepa b c
0
10
20
30
Ene
rgy
inta
ke (
kcal
)
0 10 20 30Days
MC4R
MC4R
0
2
4
6
8
Foo
d in
take
(g)
+/+
–/– +/+
–/– ?/+
ob/o
b
moderate-fat diet
low-fat diet
(Female) (Male) (Male)
*
*
*
*
*
–25
0
25
50
Fee
d ef
ficie
ncy
(mg/
kcal
)
+/+
–/–
+/+
–/–
?/+
ob/o
b
moderate-fat diet
low-fat diet
(Female) (Male) (Male)
*
*
*
MFMF
+/+
–/–
a b c
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://n
euro
sci.n
atu
re.c
om
© 2001 Nature Publishing Group http://neurosci.nature.com
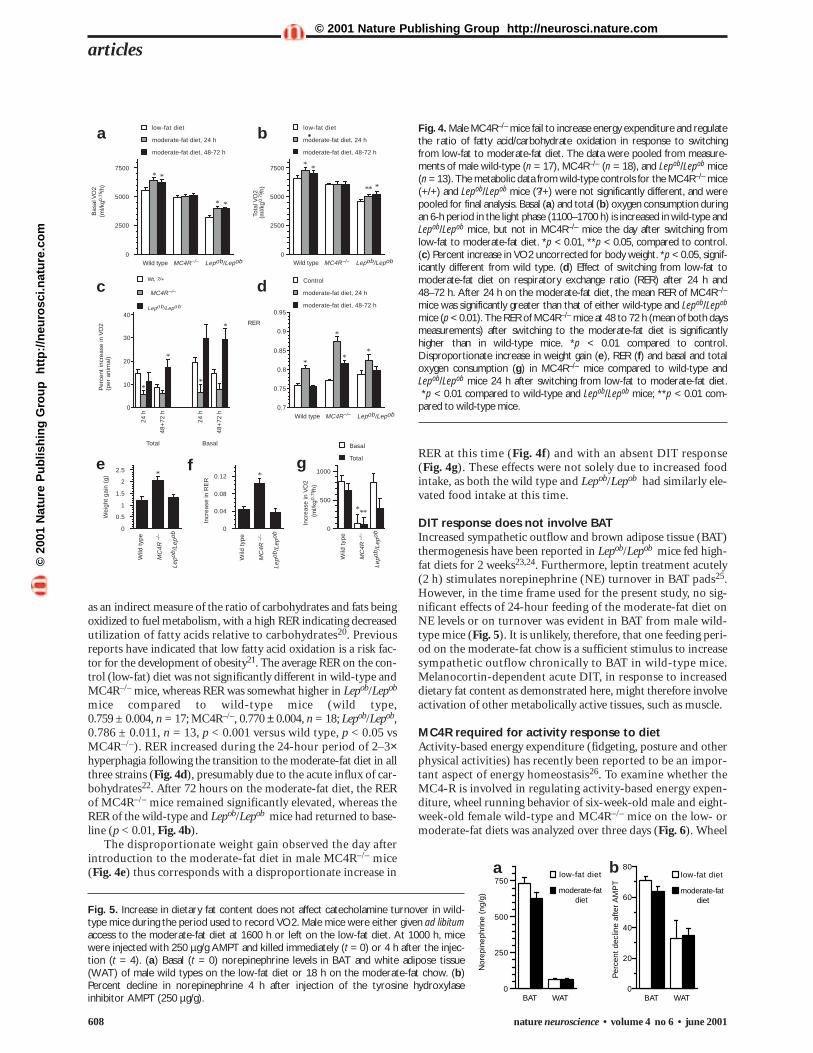
as an indirect measure of the ratio of carbohydrates and fats beingoxidized to fuel metabolism, with a high RER indicating decreasedutilization of fatty acids relative to carbohydrates20. Previousreports have indicated that low fatty acid oxidation is a risk fac-tor for the development of obesity21. The average RER on the con-trol (low-fat) diet was not significantly different in wild-type andMC4R–/– mice, whereas RER was somewhat higher in Lepob/Lepob
mice compared to wild-type mice (wild type, 0.759 ± 0.004, n = 17; MC4R–/–, 0.770 ± 0.004, n = 18; Lepob/Lepob,0.786 ± 0.011, n = 13, p < 0.001 versus wild type, p < 0.05 vsMC4R–/–). RER increased during the 24-hour period of 2–3×hyperphagia following the transition to the moderate-fat diet in allthree strains (Fig. 4d), presumably due to the acute influx of car-bohydrates22. After 72 hours on the moderate-fat diet, the RERof MC4R–/– mice remained significantly elevated, whereas theRER of the wild-type and Lepob/Lepob mice had returned to base-line (p < 0.01, Fig. 4b).
The disproportionate weight gain observed the day afterintroduction to the moderate-fat diet in male MC4R–/– mice(Fig. 4e) thus corresponds with a disproportionate increase in
RER at this time (Fig. 4f) and with an absent DIT response(Fig. 4g). These effects were not solely due to increased foodintake, as both the wild type and Lepob/Lepob had similarly ele-vated food intake at this time.
DIT response does not involve BATIncreased sympathetic outflow and brown adipose tissue (BAT)thermogenesis have been reported in Lepob/Lepob mice fed high-fat diets for 2 weeks23,24. Furthermore, leptin treatment acutely (2 h) stimulates norepinephrine (NE) turnover in BAT pads25.However, in the time frame used for the present study, no sig-nificant effects of 24-hour feeding of the moderate-fat diet onNE levels or on turnover was evident in BAT from male wild-type mice (Fig. 5). It is unlikely, therefore, that one feeding peri-od on the moderate-fat chow is a sufficient stimulus to increasesympathetic outflow chronically to BAT in wild-type mice.Melanocortin-dependent acute DIT, in response to increaseddietary fat content as demonstrated here, might therefore involveactivation of other metabolically active tissues, such as muscle.
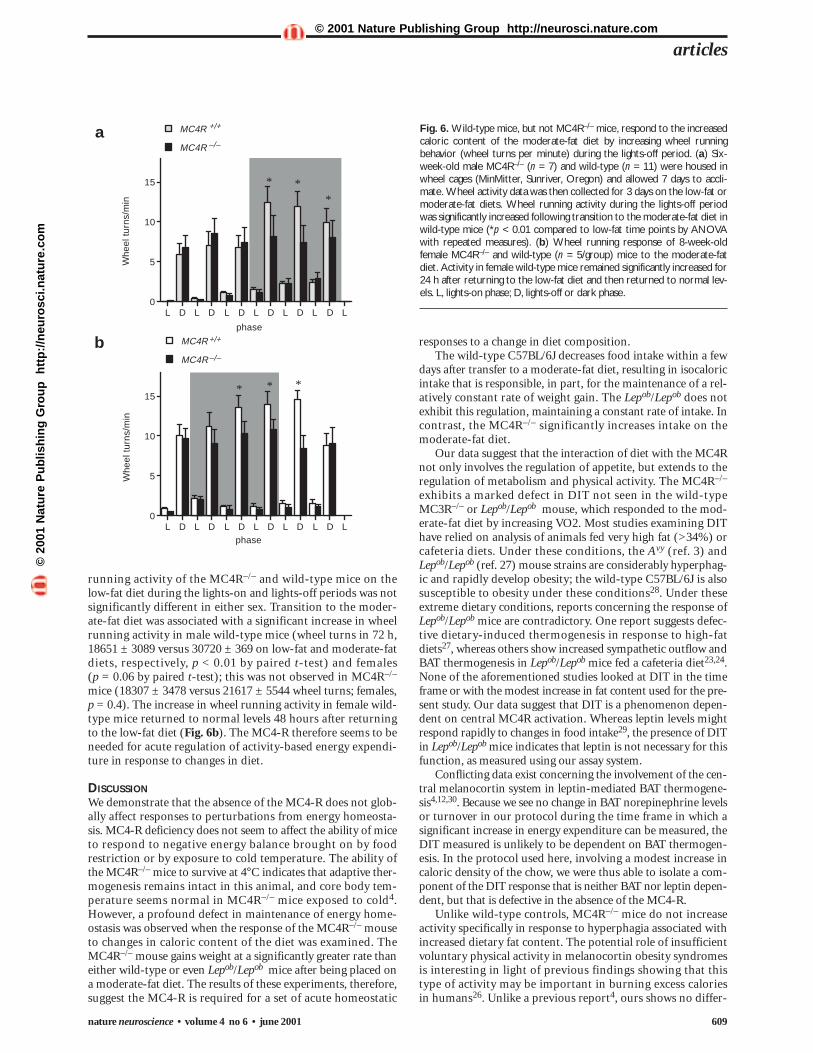
MC4R required for activity response to dietActivity-based energy expenditure (fidgeting, posture and otherphysical activities) has recently been reported to be an impor-tant aspect of energy homeostasis26. To examine whether theMC4-R is involved in regulating activity-based energy expen-diture, wheel running behavior of six-week-old male and eight-week-old female wild-type and MC4R–/– mice on the low- ormoderate-fat diets was analyzed over three days (Fig. 6). Wheel
articles
608 nature neuroscience • volume 4 no 6 • june 2001
0
2500
5000
7500
Bas
al V
O2
moderate-fat diet, 48-72 h
moderate-fat diet, 24 h
low-fat diet
0
0.5
1
1.5
2
2.5
Wei
ght
gain
(g)
0
0.04
0.08
0.12
Incr
ease
in R
ER
0
500
1000
Incr
ease
in V
O2
Total
Basal
*
***
*
0
2500
5000
7500
Tota
l VO
2
moderate-fat diet, 48-72 h
moderate-fat diet, 24 h
low-fat diet
* *
* *
* *
***
0.7
0.75
0.8
0.85
0.9
0.95
RER
moderate-fat diet, 48-72 h
moderate-fat diet, 24 h
Control
*
*
**
0
10
20
30
40
Per
cent
incr
ease
in V
O2
(per
ani
mal
)
24 h
48+
72 h
24 h
48+
72 h
Lepob/Lepob
MC4R
Wt, ?/+
Total Basal
*
*
*
*
Lepo
b /Le
pob
MC4R Lepob/LepobWild type
(ml/k
g
/h)
0.75
(ml/k
g
/h)
0.75
(ml/k
g
/h)
0.75
–/– MC4R Lepob/LepobWild type –/–
–/–
MC4R Lepob/LepobWild type –/–
Wild
typ
e
MC
4R–/
–
Lepo
b /Le
pob
Wild
typ
e
MC
4R–/
–
Lepo
b /Le
pob
Wild
typ
e
MC
4R–/
–
Fig. 4. Male MC4R–/– mice fail to increase energy expenditure and regulatethe ratio of fatty acid/carbohydrate oxidation in response to switchingfrom low-fat to moderate-fat diet. The data were pooled from measure-ments of male wild-type (n = 17), MC4R–/– (n = 18), and Lepob/Lepob mice (n = 13). The metabolic data from wild-type controls for the MC4R–/– mice(+/+) and Lepob/Lepob mice (?/+) were not significantly different, and werepooled for final analysis. Basal (a) and total (b) oxygen consumption duringan 6-h period in the light phase (1100–1700 h) is increased in wild-type andLepob/Lepob mice, but not in MC4R–/– mice the day after switching fromlow-fat to moderate-fat diet. *p < 0.01, **p < 0.05, compared to control.(c) Percent increase in VO2 uncorrected for body weight. *p < 0.05, signif-icantly different from wild type. (d) Effect of switching from low-fat tomoderate-fat diet on respiratory exchange ratio (RER) after 24 h and48–72 h. After 24 h on the moderate-fat diet, the mean RER of MC4R–/–
mice was significantly greater than that of either wild-type and Lepob/Lepob
mice (p < 0.01). The RER of MC4R–/– mice at 48 to 72 h (mean of both daysmeasurements) after switching to the moderate-fat diet is significantlyhigher than in wild-type mice. *p < 0.01 compared to control.Disproportionate increase in weight gain (e), RER (f) and basal and totaloxygen consumption (g) in MC4R–/– mice compared to wild-type andLepob/Lepob mice 24 h after switching from low-fat to moderate-fat diet.*p < 0.01 compared to wild-type and Lepob/Lepob mice; **p < 0.01 com-
pared to wild-type mice.
a b
c d
e f g
0
20
40
60
80
Per
cent
dec
line
afte
r A
MP
T
BAT WAT0
250
500
750
Nor
epin
ephr
ine
(ng/
g)
BAT WAT
moderate-fat diet
low-fat diet low-fat diet
moderate-fat diet
Fig. 5. Increase in dietary fat content does not affect catecholamine turnover in wild-type mice during the period used to record VO2. Male mice were either given ad libitumaccess to the moderate-fat diet at 1600 h or left on the low-fat diet. At 1000 h, micewere injected with 250 µg/g AMPT and killed immediately (t = 0) or 4 h after the injec-tion (t = 4). (a) Basal (t = 0) norepinephrine levels in BAT and white adipose tissue(WAT) of male wild types on the low-fat diet or 18 h on the moderate-fat chow. (b)Percent decline in norepinephrine 4 h after injection of the tyrosine hydroxylaseinhibitor AMPT (250 µg/g).
a b
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://n
euro
sci.n
atu
re.c
om
© 2001 Nature Publishing Group http://neurosci.nature.com
nature neuroscience • volume 4 no 6 • june 2001 609
running activity of the MC4R–/– and wild-type mice on thelow-fat diet during the lights-on and lights-off periods was notsignificantly different in either sex. Transition to the moder-ate-fat diet was associated with a significant increase in wheelrunning activity in male wild-type mice (wheel turns in 72 h,18651 ± 3089 versus 30720 ± 369 on low-fat and moderate-fatdiets, respectively, p < 0.01 by paired t-test) and females (p = 0.06 by paired t-test); this was not observed in MC4R–/–
mice (18307 ± 3478 versus 21617 ± 5544 wheel turns; females,p = 0.4). The increase in wheel running activity in female wild-type mice returned to normal levels 48 hours after returningto the low-fat diet (Fig. 6b). The MC4-R therefore seems to beneeded for acute regulation of activity-based energy expendi-ture in response to changes in diet.
DISCUSSIONWe demonstrate that the absence of the MC4-R does not glob-ally affect responses to perturbations from energy homeosta-sis. MC4-R deficiency does not seem to affect the ability of miceto respond to negative energy balance brought on by foodrestriction or by exposure to cold temperature. The ability ofthe MC4R–/– mice to survive at 4°C indicates that adaptive ther-mogenesis remains intact in this animal, and core body tem-perature seems normal in MC4R–/– mice exposed to cold4.However, a profound defect in maintenance of energy home-ostasis was observed when the response of the MC4R–/– mouseto changes in caloric content of the diet was examined. TheMC4R–/– mouse gains weight at a significantly greater rate thaneither wild-type or even Lepob/Lepob mice after being placed ona moderate-fat diet. The results of these experiments, therefore,suggest the MC4-R is required for a set of acute homeostatic
responses to a change in diet composition.The wild-type C57BL/6J decreases food intake within a few
days after transfer to a moderate-fat diet, resulting in isocaloricintake that is responsible, in part, for the maintenance of a rel-atively constant rate of weight gain. The Lepob/Lepob does notexhibit this regulation, maintaining a constant rate of intake. Incontrast, the MC4R–/– significantly increases intake on themoderate-fat diet.
Our data suggest that the interaction of diet with the MC4Rnot only involves the regulation of appetite, but extends to theregulation of metabolism and physical activity. The MC4R–/–
exhibits a marked defect in DIT not seen in the wild-typeMC3R–/– or Lepob/Lepob mouse, which responded to the mod-erate-fat diet by increasing VO2. Most studies examining DIThave relied on analysis of animals fed very high fat (>34%) orcafeteria diets. Under these conditions, the Avy (ref. 3) andLepob/Lepob (ref. 27) mouse strains are considerably hyperphag-ic and rapidly develop obesity; the wild-type C57BL/6J is alsosusceptible to obesity under these conditions28. Under theseextreme dietary conditions, reports concerning the response ofLepob/Lepob mice are contradictory. One report suggests defec-tive dietary-induced thermogenesis in response to high-fatdiets27, whereas others show increased sympathetic outflow andBAT thermogenesis in Lepob/Lepob mice fed a cafeteria diet23,24.None of the aforementioned studies looked at DIT in the timeframe or with the modest increase in fat content used for the pre-sent study. Our data suggest that DIT is a phenomenon depen-dent on central MC4R activation. Whereas leptin levels mightrespond rapidly to changes in food intake29, the presence of DITin Lepob/Lepob mice indicates that leptin is not necessary for thisfunction, as measured using our assay system.
Conflicting data exist concerning the involvement of the cen-tral melanocortin system in leptin-mediated BAT thermogene-sis4,12,30. Because we see no change in BAT norepinephrine levelsor turnover in our protocol during the time frame in which asignificant increase in energy expenditure can be measured, theDIT measured is unlikely to be dependent on BAT thermogen-esis. In the protocol used here, involving a modest increase incaloric density of the chow, we were thus able to isolate a com-ponent of the DIT response that is neither BAT nor leptin depen-dent, but that is defective in the absence of the MC4-R.
Unlike wild-type controls, MC4R–/– mice do not increaseactivity specifically in response to hyperphagia associated withincreased dietary fat content. The potential role of insufficientvoluntary physical activity in melanocortin obesity syndromesis interesting in light of previous findings showing that thistype of activity may be important in burning excess caloriesin humans26. Unlike a previous report4, ours shows no differ-
articles
0
5
10
15
Whe
el t
urns
/min
L D L D L D L D L D L D L
MC4R
MC4R
0
5
10
15
Whe
el t
urns
/min
L D L D L D L D L D L D L
MC4R
MC4R
phase
phase
***
*
**
+/+
+/+
–/–
–/–
Fig. 6. Wild-type mice, but not MC4R–/– mice, respond to the increasedcaloric content of the moderate-fat diet by increasing wheel runningbehavior (wheel turns per minute) during the lights-off period. (a) Six-week-old male MC4R–/– (n = 7) and wild-type (n = 11) were housed inwheel cages (MinMitter, Sunriver, Oregon) and allowed 7 days to accli-mate. Wheel activity data was then collected for 3 days on the low-fat ormoderate-fat diets. Wheel running activity during the lights-off periodwas significantly increased following transition to the moderate-fat diet inwild-type mice (*p < 0.01 compared to low-fat time points by ANOVAwith repeated measures). (b) Wheel running response of 8-week-oldfemale MC4R–/– and wild-type (n = 5/group) mice to the moderate-fatdiet. Activity in female wild-type mice remained significantly increased for24 h after returning to the low-fat diet and then returned to normal lev-els. L, lights-on phase; D, lights-off or dark phase.
a
b
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://n
euro
sci.n
atu
re.c
om
© 2001 Nature Publishing Group http://neurosci.nature.com

ence in the activity of young MC4R–/– mice relative to wild-type controls when fed the standard low-fat diet. This dis-crepancy probably reflects differences in technique. In thepresent study, the mice were allowed at least seven days to accli-mate to the new environment, whereas previously, activity wasexamined in a novel environment. Given that the MC4R–/–
mouse has an abnormal response to novelty and mild stres-sors31, the reduction in spontaneous locomotor behavior mightrepresent a stress-related response, rather than be a true depic-tion of basal home-cage activity.
Mice fed very high-fat diets (42–43% kcal from fat) for aslong as two weeks exhibit no increase in basal leptin levels com-pared to mice fed a high-carbohydrate diet with a lower fatcontent (approximately 25% kcal from fat)32. Analysis of areaunder the curve from frequent sampling does show that a mod-erate (35%) increase in plasma leptin occurs within 24 hoursafter a switch to a high-fat diet32. It is not certain whether anincrease in leptin would be observed with the moderateincrease in fat in our protocol. In any event, our data showingan intact DIT response in the Lepob/Lepob mouse suggest thatat least this response is not leptin dependent. Leptin infusionover an eight-day period has also been reported to have noeffect on wheel running activity in rats33.
How might increased fat intake lead to an increase in MC4-R activity if leptin is not initially involved? It is possiblethat an intermediate of fatty acid metabolism, such as malonyl-CoA34, may directly communicate information to MC4-R-depen-dent hypothalamic circuits via arcuate POMC and NPY/AGRPneurons. The data presented here argue that melanocortin obe-sity syndromes may be caused by acute defects in coordinatingintake with expenditure. This may result from defective sensingof nutrient levels by structures like the arcuate nucleus, withdefective leptin signaling in the same circuits occurring alongwith, or perhaps even secondarily to the development of obesity.
METHODSMice. MC4R–/– and wild-type controls are from the originalC57BL/6Jx129 colony5 within the Vollum Institute. Studies compar-ing feeding in MC4R–/– and wild-type males used mice bred three gen-erations into the C57BL/6J strain. Feeding and metabolic studies infemales used mice bred five generations into the C57BL/6J strain. MaleLepob/Lepob and +/? littermates, in the C57BL/6J background, werepurchased from Jackson Laboratories (Bar Harbor, Maine). Mice weregroup housed in a 12-h light/dark cycle. For studies measuring foodintake, mice were housed individually and food intake was estimatedby measuring the weight of food remaining in the wire feeders in theroof of the cage. Mice were weaned at 21 days and allowed ad libitumaccess to Laboratory Rodent Diet (Purina 5001, PMI Nutrition Inter-national, Brentwood, Missouri). To minimize error due to spillage,large intact pellets were used and the litter was inspected for signifi-cant (>0.1 g) particles of food at the time of weighing. Low-fat Purinadiet 5001 (23.0% protein, 4.5% fat, 5.3% crude fiber, 49% carbohy-drate, total digestible nutrient 76%, 3.04 kcal/g metabolizable energy)and moderate-fat Purina diet 5015 (17.0% protein, 11.0% fat, 3%crude fiber, 53.5% carbohydrate, total digestible nutrient 88%, 3.73kcal/g metabolizable energy) were used.
Indirect calorimetry. VO2 and VCO2 were simultaneously determinedby indirect calorimetry (Oxymax, Columbus Instruments, Columbus,Ohio). Mice were housed in separate chambers at 24 ± 1°C. Mice werefirst acclimatized to the chambers for two days. Measurements wererecorded for 4–5 h during the middle of the light cycle (1100–1600 h).Samples were recorded every 3 min with the room air reference takenevery 30 min and the air flow to chambers 500 ml/min. Basal oxygenconsumption was determined for individual curves as the average of thelowest plateau regions corresponding to resting periods. Total oxygen
consumption was the result of all samples recorded corresponding toperiods of movement as well as inactivity. RER was calculated as themolar ratio of VO2:VCO2.
Catecholamine turnover. Catecholamine turnover in brown and whiteadipose tissue was measured as described previously25. Briefly, micewere given an intraperitoneal injection of 250 µg/g α-methyl-p-tyro-sine methyl ester (AMPT, Sigma-Aldrich, St. Louis, Missouri) dissolvedin PBS. Mice were killed either immediately (time 0) or 4 h after injec-tion, and interscapular brown and epididymal white fat was collectedfor catecholamine analysis.
Motor activity. Animals were housed individually in metabolic cagesequipped with a running wheel (Mini-Mitter, Sunriver, Oregon). Wheelrevolutions were quantified by recording the magnetic switch closuresof a magnet placed on the revolving wheel.
Statistical methods. Data sets were analyzed for statistical significanceusing PRISM (GraphPad, San Diego, California) for ANOVA or EXCEL(Microsoft, Redmond, Washington) for Student’s t-test. Repeated mea-sures ANOVA was used to analyze data sets where samples had been col-lected over several days (VO2, wheel turns).
ACKNOWLEDGEMENTSThis work was supported by NIH grant PPGDK55819 to R.D.C. We thank
K. Khong, E. Douthit and K.E. Miles for their help.
RECEIVED 11 APRIL; ACCEPTED 25 APRIL 2001
1. Leibel, R. L., Chung, W. K. & Chua, S. C .J. The molecular genetics of rodentsingle gene obesities. J. Biol. Chem. 272, 31937–31940 (1997).
2. Yen, T. T., McKee, M. M. & Stamm, N. B. Thermogenesis and weight control.Int. J. Obes. 8 (Suppl. 1), 65–78 (1984).
3. Frigeri, L. G., Wolff, G. L. & Teguh, C. Differential responses of yellow Avy/Aand agouti A/a (BALB/c X VY) F1 hybrid mice to the same diets: glucosetolerance, weight gain, and adipocyte cellularity. Int. J. Obes. 12, 305–320(1988).
4. Marie, L. S., Miura, G. I., Marsh, D. J., Yagaloff, K. & Palmiter, R. D. Ametabolic defect promotes obesity in mice lacking melanocortin-4 receptors.Proc. Natl. Acad. Sci. USA 97, 12339–12344 (2000).
5. Huszar, D. et al. Targeted disruption of the melanocortin-4 receptor results inobesity in mice. Cell 88, 131–141 (1997).
6. Cowley, M. A. et al. Integration of NPY, AGRP, and melanocortin signals inthe hypothalamic paraventricular nucleus: evidence of a cellular basis for theadipostat. Neuron 24, 155–163 (1999).
7. Marsh, D. J. et al. Response of melanocortin-4 receptor-deficient mice toanorectic and orexigenic peptides. Nat. Genet. 21, 119–122 (1999).
8. Halaas, J. L. et al. Physiological response to long-term peripheral and centralleptin infusion in lean and obese mice. Proc. Natl. Acad. Sci. USA 94,8878–8883 (1997).
9. Boston, B. A., Blaydon, K. M., Varnerin, J. & Cone, R. D. Independent andadditive effects of central POMC and leptin pathways on murine obesity.Science 278, 1641–1644 (1997).
10. Cheung, C. C., Clifton, D. K. & Steiner, R. A. Proopiomelanocortin neuronsare direct targets for leptin in the hypothalamus. Endocrinology 138,4489–4492 (1997).
11. Seeley, R. J. et al. Melanocortin receptors in leptin effects. Nature 390, 349(1997).
12. Satoh, N. et al. Satiety effect and sympathetic activation of leptin aremediated by hypothalamic melanocortin system. Neurosci. Lett. 249, 107–110(1998).
13. Smith, A. I. & Funder, J. W. Proopiomelanocortin processing in the pituitary,central nervous system, and peripheral tissues. Endocr. Rev. 9, 159–179(1988).
14. Low, M. J., Simerly, R. D. & Cone, R. D. Receptors for the melanocortinpeptides in the central nervous system. Curr. Opin. Endocrinol. Diabet. 1,79–88 (1994).
15. Baskin, D. G. et al. Insulin and leptin: dual adiposity signals to the brain forthe regulation of food intake and body weight. Brain. Res. 848, 114–123(1999).
16. Hagan, M. M. et al. Role of the CNS melanocortin system in the response tooverfeeding. J. Neurosci. 19, 2362–2367 (1999).
17. Enerback, S. et al. Mice lacking mitochondrial uncoupling protein are cold-sensitive but not obese. Nature 387, 90–94 (1997).
18. Butler, A. A. et al. A unique metabolic syndrome causes obesity in themelanocortin-3 receptor-deficient mouse. Endocrinology 141, 3518–3521(2000).
articles
610 nature neuroscience • volume 4 no 6 • june 2001
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://n
euro
sci.n
atu
re.c
om
© 2001 Nature Publishing Group http://neurosci.nature.com

nature neuroscience • volume 4 no 6 • june 2001 611
19. Schutz, Y., Bessard, T. & Jecquier, E. Thermogenesis measured over a wholeday in obese and non-obese women. Am. J. Clin. Nutr. 40, 542–552 (1984).
20. Ferrannini, E. The theoretical basis of indirect calorimetry: a review.Metabolism 37, 287–301 (1988).
21. Zurlo, F. et al. Low ratio of fat to carbohydrate oxidation as predictor ofweight gain: study of 24-h RQ. Am. J. Phys. 259, E650–657 (1990).
22. Nakaya, Y. et al. Respiratory quotient in patients with non-insulin-dependentdiabetes mellitus treated with insulin and oral hypoglycemic agents. Ann.Nutr. Metab. 42, 333–340 (1998).
23. Mercer, S. W. & Trayhurn, P. Effect of high fat diets on energy balance andthermogenesis in brown adipose tissue of lean and genetically obese ob/obmice. J. Nutr. 117, 2147–2153 (1987).
24. Himms-Hagen, J., Hogan, S. & Zaror-Behrens, G. Increased brown adiposetissue thermogenesis in obese (ob/ob) mice fed a palatable diet. Am. J. Physiol.250, E274–E281 (1986).
25. Collins, S. et al. Role of leptin in fat regulation. Nature 380, 677 (1996).26. Levine, J. A., Eberhardt, N. L. & Jensen, M. D. Role of nonexercise activity
thermogenesis in resistance to fat gain in humans. Science 283, 212–214(1999).
27. Trayhurn, P., Jones, P. M., McGuckin, M. M. & Goodbody, A. E. Effect of
overfeeding on energy balance and brown fat thermogenesis in obese (ob/ob)mice. Nature 295, 323–325 (1982).
28. Surwit, R. S., Kuhn, C. M., Cochrane, C., McCubbin, J. A. & Feinglos, M. N.Diet-induced type II diabetes in C57Bl/6J mice. Diabetes 37, 1163–1167 (1988).
29. Saladin, R. et al. Transient increase in obese gene expression after food intakeor insulin administration. Nature 377, 527–532 (1995).
30. Haynes, W. G., Morgan, D. A., Djalali, A., Sivitz, W. I. & Mark, A. L.Interactions between the melanocortin system and leptin in control ofsympathetic nerve traffic. Hypertension 33, 542–547 (1999).
31. De Souza, J., Butler, A. A. & Cone, R. D. Disproportionate inhibition offeeding in A(y) mice by certain stressors: a cautionary note.Neuroendocrinology 72, 126–132 (2000).
32. Ziotopoulou, M., Mantzoros, C. S., Hileman, S. M. & Flier, J. S. Differentialexpression of hypothalamic neuropeptides in the early phase of diet-inducedobesity in mice. Am. J. Physiol. Endocrinol. Metab. 279, E838–E845 (2000).
33. Eckel, L. A. et al. Chronic administration of OB protein decreases food intakeby selectively reducing meal size in female rats. Am. J. Physiol. 275, R186–193(1998).
34. Loftus, T. M. et al. Reduced food intake and body weight in mice treated withfatty acid synthase inhibitors. Science 288, 2379–2381 (2000).
articles©
2001
Nat
ure
Pu
blis
hin
g G
rou
p
htt
p:/
/neu
rosc
i.nat
ure
.co
m© 2001 Nature Publishing Group http://neurosci.nature.com





























