Embed Size (px)
Citation preview

9The NucleusNuclear Organization
Nuclear Envelope and Molecular TraffickingNucleolus and rRNA Processing

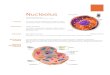
9 The Structure and Function of the Nuclear Envelope
• The nuclear envelope has a complex structure consisting of two nuclear membranes, an underlying nuclear lamina, and nuclear pore complexes.
Nuclear membranes are a system of two concentric membranes (inner and outer) that surround the nucleus.
The nuclear envelope separates the contents of the nucleus from the cytoplasm and provides the structural framework of the nucleus.

9.1 The nuclear envelope
The selective traffic of proteins and RNAs through the nuclear pore complexes establish the internal composition of the nucleus and also play a critical role in regulating eukaryotic gene expression.

9 Structure of the Nuclear Envelope• The critical function
of the nuclear membranes is to act as a barrier that separates the contents of the nucleus from the cytoplasm.
• The nuclear lamina underlies the inner nuclear membrane and is a fibrous meshwork that provides structural support to the nucleus.
• Lamins are 60- to 80-kilodalton fibrous proteins that make up the nuclear lamina.

9 Structure of the Nuclear Envelope
• Lamins are composed of intermediate filament proteins which combine with each other to form higher-order structures.
• Attachment of Lamins to the nuclear envelope is mediated by prenylation and binding to specific inner nuclear membrane proteins such as emerin and the lamin B receptor.
• Nuclear Lamina diseases: Hutchinson-Gilford progeria

9 Nuclear Lamina Defects
• X-linked Emery-Dreifuss muscular Dystrophy Results form mutations in the nuclear
envelope protein, emerin.
• Non-Sex Linked Emery Dreifuss Muscular Dystrophy
Results from mutations in the LMNA gene which is a single gene that encodes lamin A and C.
• Hutchinson-Guilford Progeria Disease Results from a 150 bp deletion within the lamin
A gene.

9 The Nuclear Pore Complex
• A nuclear pore complex consists of a structure with eightfold symmetry organized around a large central channel.

9 The Nuclear Pore Complex
• Nuclear pore complexes are the only channels through which small polar molecules, ions, and macromolecules can travel between the nucleus and the cytoplasm.
• Depending on their size, molecules can travel through the nuclear pore complex by one of two different mechanisms.
Passive transport
Active transport –energy dependent

9 Selective Transport of Proteins to and from the Nucleus
• Nuclear export signals are specific amino acid sequences that target proteins for export from the nucleus.
• Protein import through the nuclear pore complex begins when a specific importin binds to the nuclear localization signal of a cargo protein in the cytoplasm.

9 Selective Transport of Proteins to and from the Nucleus
• Nuclear localization signals are specific amino acid sequences that are recognized by transport receptors and direct the transport of proteins through the nuclear pore complex.
• Nuclear localization signals have been identified in many proteins.

9 Selective Transport of Proteins to and from the Nucleus
• Nuclear transport receptors are proteins that recognize nuclear localization signals and mediate transport across the nuclear envelope.
• Karyopherin is a nuclear transport receptor.
• Importins transport macromolecules to the nucleus from the cytoplasm.
• Exportins transport macromolecules from the nucleus to the cytoplasm.

9.10 Distribution of Ran/GTP across the nuclear envelope
Ran is one of several types of small GTP-binding proteins that regulate movement of macromolecules through the nuclear pore.

9.12 Nuclear export

9 Regulation of Nuclear Protein Import
• The transport of proteins to the nucleus is yet another level at which the activities of nuclear proteins can be controlled.
• In one mechanism of regulation, transcription factors associate with cytoplasmic proteins that mask their nuclear localization signals.

9 Transport of RNAs
• Most RNAs are exported from the nucleus to the cytoplasm.
• RNAs are transported across the nuclear envelope as ribonucleoprotein complexes, or RNPs.

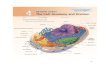
13The Plasma Membrane

9 13.1 Bilayer structure of the plasma membrane• Studies of the red blood
cell plasma membrane provided the first evidence that biological membranes consist of lipid bilayers.
• The fundamental structure of the plasma membrane is the phospholipid bilayer, which forms a stable barrier between two aqueous compartments.
• Proteins embedded within the phospholipid bilayer carry out the specific functions of the plasma membrane.

9 The Phospholipid Bilayer• Phosphatidylcholine -- glycerol phospholipid with a head group formed
from choline.
• Phosphatidylethanolamine -- glycerol phospholipid with a head group formed from ethanolamine.
• Phosphatidylserine -- glycerol phospholipid with a head group formed from serine.
• Sphingomyelin -- phospholipid consisting of two hydrocarbon chains bound to a polar head group containing serine.

9 The Phospholipid Bilayer
• The fluid mosaic model of membrane structure is now generally accepted as the basic paradigm for the organization of all biological membranes.
• Phosphatidylinositol is a phospholipid localized to the inner half of the plasma membrane, which plays an important role in cell signaling.
• Glycolipids are lipids consisting of two hydrocarbon chains linked to a polar head group containing carbohydrates.
• Cholesterol, a lipid consisting of four hydrocarbon rings, is a major membrane constituent of animal cells.

9 Membrane Proteins
• Peripheral membrane proteins are proteins that dissociate from the membrane following treatments with polar reagents that do not disrupt the phospholipid bilayer.
• Integral membrane proteins can be released only by treatments that disrupt the phospholipid bilayer.
• Transmembrane proteins span the lipid bilayer with portions exposed on both sides of the membrane.

9 13.4 Solubilization of integral membrane proteins by detergents

9 13.6 Integral membrane proteins of red blood cells
• The most abundant peripheral membrane protein of red blood cells is spectrin, which is the major cytoskeletal protein of erythrocytes.
• There are two major integral membrane proteins in red blood cells—glycophorin and band 3.

9 Membrane Proteins
• Some proteins are anchored in the plasma membrane by covalently attached lipids or glycolipids.
• Glycosylphosphatidylinositol, or GPI, are glycolipids containing phosphatidylinositol that anchor proteins to the external face of the plasma membrane.
• Other proteins are anchored to the inner leaflet of the plasma membrane by covalently attached lipids.

9 13.10 Mobility of membrane proteins

9 Mobility of Membrane Proteins
• Lipid composition can perturb the free diffusion of membrane proteins.
• The basolateral domain is the surface region of a polarized epithelial cell that is in contact with adjacent cells or the extracellular matrix.
• The apical domain is the exposed free surface of a polarized epithelial cell.

9 13.12 Structure of lipid rafts

9 13.13 The glycocalyx• The glycocalyx is formed by oligosaccharides of
glycolipids and transmembrane glycoproteins.
• Selectins are cell adhesion molecules that recognize oligosaccharides exposed on the cell surface.

9 13.15 Permeability of phospholipid bilayers• The internal composition
of the cell is maintained because the plasma membrane is selectively permeable to small molecules.
• Only small, relatively hydrophobic molecules are able to diffuse across a phospholipid bilayer at significant rates by using passive diffusion.
• Passive diffusion is the simplest mechanism by which molecules can cross the plasma membrane.
• Specific transport proteins mediate the selective passage of small molecules across the membrane, allowing the cell to control the composition of its cytoplasm.

9 Phagocytosis
• Phagolysosomes, which are phagosomes fused to lysosomes, contain lysosomal acid hydrolases that digest the ingested material.
• The ingestion of large particles by phagocytosis plays distinct roles in different kinds of cells.

9 Receptor-Mediated Endocytosis
• Receptor-mediated endocytosis, a form of pinocytosis, provides a mechanism for the selective uptake of specific macromolecules.
• Clathrin-coated pits are specialized regions of the plasma membrane where specific cell surface receptors are found.
• Dynamin, a membrane-associated GTP-binding protein, assists in the budding off of pits from the plasma membrane.

9 Receptor-Mediated Endocytosis
• Clathrin assembles into a basketlike structure that distorts the membrane, forming invaginated pits.
• Clathrin coated pits occupy about 1-2% of the surface area of plasma membranes.

9 Receptor-Mediated Endocytosis
• Caveolae are small invaginations of the plasma membrane.
• Caveolin is a protein that interacts with lipid rafts and forms caveolae.
• Macropinocytosis is a process where large vesicles can mediate the uptake of fluids.

9 Protein Trafficking in Endocytosis
• Endosomes are vesicles with tubular extensions, located at the periphery of the cell, that fuse with clathrin-coated vesicles which have shed their coats.
• An important feature of early endosomes is that they maintain an acidic internal pH as the result of the action of a membrane H+ pump.

9 Protein Trafficking in Endocytosis
• Recycling to the plasma membrane is the major fate of membrane proteins taken up by receptor-mediated endocytosis.
• Ligands and membrane proteins destined for degradation in lysosomes are transported from early endosomes to late endosomes, which are located near the nucleus.
• Receptor down-regulation is a phenomenon where receptor-ligand complexes are removed from the plasma membrane, thereby terminating the response of the cell to growth factor stimulation.