Embed Size (px)
Citation preview

Development 111, 771-778 (1991)Printed in Great Britain © The Company of Biologists Limited 1991
771
A member of the RXR nuclear receptor family is expressed in neural-crest-
derived cells of the developing chick peripheral nervous system
ANNIE ROWE1'2, NICHOLAS S. C. EAGER1 and PAUL M. BRICKELL1*1The Medical Molecular Biology Unit, Department of Biochemistry and 2Department of Anatomy and Developmental Biology, UniversityCollege and Middlesex School of Medicine, The Windeyer Building, Cleveland Street, London, W1P 6DB, UK
* Author for correspondence
Summary
Retinoic acid (RA) affects differentiation and morpho-genesis in various developmental systems and is believedto act through nuclear RA receptors that belong to thesteroid/thyroid hormone family of ligand-binding tran-scription factors. Three closely related receptors, RAR-a, -/Jand -y, with distinct expression patterns, have beenidentified and a fourth receptor, hRXR-a, whichresponds to RA but which has low homology to RAR-a,-P and -y, was recently discovered. Here we report theisolation of a cDNA clone encoding a chicken homologueof hRXR-a (cRXR) and show that a cRXR transcript of2.5 kb is expressed in a range of embryonic chick tissues.By in situ hybridization to sections from stage 24 andstage 27 chick embryos, we show that cRXR transcriptsare expressed at high levels in the liver and in elements of
the developing peripheral nervous system derived fromthe neural crest, including dorsal root ganglia, cranialganglia, enteric ganglia and peripheral nerve tracts. Atstage 16, in the posterior trunk region, cRXR transcriptsare expressed by cells in the neural crest and in neuralcrest cells migrating into the sclerotome, indicating thatneural crest cells express cRXR transcripts before overtdifferentiation into peripheral nervous tissue. Thisdistribution suggests a novel role for RA in thedeveloping peripheral nervous system, mediated bycRXR. In addition, it identifies cRXR as a marker for aspecific population of neural-crest-derived cells.
Key words: RXR, retinoic acid receptor, peripheralnervous system, neural crest.
Introduction
The retinoids are a group of related compounds,including vitamin A (retinol) and its metabolite retinoicacid (RA), that profoundly affect differentiation andmorphogenesis in various developmental systems.Local application of RA to developing chick limb budsinduces duplication of the digits (Tickle et al. 1982) andthere is evidence to suggest that RA may be a naturalsignalling substance in the chick limb bud (Thaller andEichele, 1987; Brickell and Tickle, 1989; Brockes,1989). Local application of RA also induces specificmalformations of the chick face (Wedden, 1987). InXenopus laevis embryos, RA acts on the developingcentral nervous system to transform anterior neuraltissue to a posterior neural specification (Durston et al.1989). In mammals, retinoids are potent teratogens,producing malformations of craniofacial structures, thelimb, the heart, the thymus and the central nervoussystem (Morriss and Thorogood, 1978; Lammer et al.1985; Satre and Kochhar, 1989). In addition to theseeffects, retinoids markedly affect the differentiationand maintenance of epithelial cells both in vivo and invitro (Asselineau et al. 1989; Kopan and Fuchs, 1989)
and are used in the treatment of a number of chronicdermatoses (Lammer et al. 1985).
RA acts by binding to nuclear RA receptors, whichare members of the steroid/thyroid hormone receptorsuperfamily. The resultant ligand-receptor complexesare capable of regulating the transcription of targetgenes (Green and Chambon, 1988; La Rosa and Gudas,1988; de Th6 et al. 1990). Three closely related RAreceptors, encoded by distinct genes, have beenidentified in humans and mice. These are RAR-a(Petkovich et al. 1987; Giguere et al. 1987), RAR-/3(Brand et al. 1988: Benbrook et al. 1988) and RAR-y(Zelent et al. 1989; Krust et al. 1989). A number ofisoforms of RAR-y, generated by alternative splicing ofthe primary transcript, have also been identified(Kastner et al. 1990; Giguere et al. 1990). Theexpression patterns of the three RAR genes are quitedistinct (Dolle" et al. 1989; Ruberte et al. 1990; Zelent etal. 1989). Recently, a fourth species of nuclear receptorthat responds to RA (hRXR-a) was identified inhumans (Mangelsdorf et al. 1990). Whilst hRXR-or isstill a member of the steroid/thyroid hormone receptorsuperfamily, it has no significant homology to RAR-o-,RAR-0 and RAR-y in the RA-binding domain and

772 A. Rowe, N. S. C. Eager and P. M. Brickell
therefore appears to represent an evolutionarily distinctRA response pathway. Transcriptional transactivationby hRXR-tfis considerably less sensitive to RA than istransactivation by hRAR-a-, an approximately five-foldhigher concentration of RA being required for a 50 %maximal response with hRXR-a (Mangelsdorf et al.1990). This suggests that a closely related metabolite orstructural analogue of RA, rather than RA itself, maybe the natural ligand of hRXR-tf. High level expressionof hRXR-a' was detected in rat liver, by northernblotting analysis (Mangelsdorf et al. 1990).
Here we report the isolation of a cDNA cloneencoding a chicken homologue of hRXR-a (cRXR) andshow by in situ hybridisation that, in addition to highlevel expression in liver, cRXR transcripts are unex-pectedly expressed in elements of the developingperipheral nervous system derived from the neuralcrest.
Materials and methods
Isolation of cDNA clonesApproximately 2X105 recombinant bacteriophage from a 10day (stage 36) chick embryo cDNA library constructed inAgtll (Clontech) were screened with a 380bp Kpnl-Pstlfragment of the human RAR-orcDNA clone p63 (Petkovich etal. 1987), which had been labelled with [<*-32P]dCTP (NewEngland Nuclear) by random priming (Feinberg and Vogel-stein, 1984). This probe contained sequences encoding the A,B and most of the C (DNA-binding) domain of human RAR-a. Plaque lifts on to Hybond-N (Amersham International)were performed according to the manufacturer's instructions.Hybridisation to duplicate filters was performed for 16 h at65°C in a solution containing 6xSSC, 1% (w/v) SDS,5xDenhardt's solution, O.lmgml"1 yeast total RNA and106ctsmin~' ml"1 of radiolabelled probe. The most stringentpost-hybridisation wash was for 30min at 55 °C in lxSSC,0.1% (w/v) SDS. The filters were autoradiographed andpositive clones were purified by rescreening under the sameconditions. The EcoRl insert of clone AR2 was subcloned intothe plasmid vector pBluescript SK+ (Stratagene) for sub-sequent analysis, yielding plasmid pR2.
Nucleotide sequencingDideoxy sequencing was performed using Sequenase (USBiochemicals) according to the manufacturer's instructions,either from double-stranded pR2 DNA or following transferof selected fragments into M13 mpl8 or mpl9. Oligonucleo-tide primers were synthesised in our laboratory. Nucleotidesequences were analysed using the Microgenie programme(Beckmann).
RNA isolation and northern blottingFertilised chicken eggs were obtained from Poyndon Farm,Waltham Cross, Herts, and were incubated at 38±1°C. Theembryos were staged according to Hamburger and Hamilton(1951), dissected into fresh PBS and snap-frozen in liquidnitrogen. Total RNA was isolated from snap-frozen tissue bythe acid guanidinium thiocyanate-phenol-chloroformmethod (Chomczynski and Sacchi, 1987) and was thenfractionated on a 1 % agarose MOPS-formaldehyde gel(10 ng per track) and blotted on to Genescreen Plusmembrane (Dupont). Sample quality and quantity werechecked by ethidium bromide staining of non-denaturing
agarose gels and by hybridisation of filters with probes fortype I or type II collagen transcripts, as described previously(Devlin etal. 1988). Filters were hybridised with a 32P-labelledRNA probe synthesised with T7 RNA polymerase (BCL), aspreviously described (Devlin et al. 1988), using the BamHI-£co RI fragment of AR2 (Fig. 1; nucleotides 786-1713) as atemplate, after subcloning into pBluescript SK+ (Stratagene)and linearisation with BamYil. Hybridisation was performedfor 16 h at 65 °C in a solution containing 60% (v/v)formamide, 6xSSC, 20mM sodium phosphate, 1% (w/v)SDS, 5xDenhardt's solution, 7% (w/v) dextran sulphate,O.lmgml"1 denatured herring testis DNA, O.lmgml yeasttotal RNA, 0.01 mgml"1 poly(A) and 10*ctsmin"'ml"1 ofradiolabelled probe. The most stringent post-hybridisationwash was for 30min at 80°C in O.lxSSC, 0.1 % (w/v) SDS.
In situ hybridisationChick embryos, staged according to Hamburger and Hamil-ton (1951) were embedded in wax as described by Davidson etal. (1988). 7 fan sections were then cut, collected on slidescoated with 3-aminopropyltriethoxysilane (TESPA, Sigma)and baked for 6-16h at 60°C. In situ hybridisation wasperformed essentially as described by Hogan et al. (1986),using a 35S-labelled antisense RNA probe synthesised fromthe same cRXR template as for the northern hybridisationprobe. Hybridisation was performed at 55°C. As a negativecontrol, adjacent sections were hybridised with a 35S-labelledsense strand RNA probe synthesised from the same template.After post-hybridisation washes and treatment with40/igmr1 RNAase A (Wilkinson etal. 1987), the slides wereautoradiographed (Hogan et al. 1986) and stained with 0.05 %(w/v) malachite green.
Results
Isolation of a cDNA clone for a chicken homologueof hRXR-aIn order to isolate chicken homologues of the humanand murine RARs, a stage 36 (10 day) chick embryocDNA library was screened at low stringency with aradiolabelled probe derived from the human RAR-acDNA clone p63 (Petkovich et al. 1987). Amongst theclones isolated was AR2. The restriction map andnucleotide sequence of the insert of AR2 and thepredicted amino acid sequence of the protein that itwould encode are shown in Fig. 1. The predicted aminoacid sequence is strongly homologous to that of thehuman nuclear retinoic acid receptor hRXR-a- (Man-gelsdorf et al. 1990), not only in the putative DNA-binding domain, but also in the putative ligand-bindingdomain (Fig. 2). It is also strongly homologous in thesedomains to the predicted protein product of the cDNAclone H-2RIIBP (Hamada et al. 1989), which Mangels-dorf et al. (1990) have suggested might represent amurine RXR-/J species (Fig. 2). In contrast, thepredicted amino acid sequence of AR2 is less homolo-gous to those of human RAR-a, RAR-/3 and RAR-y inthe DNA-binding domain, and shows no significanthomology to the ligand-binding domains of theseproteins (Fig. 2) or to those of any other knownmembers of the steroid/thyroid hormone receptorsuperfamily (data not shown). The cDNA clone AR2

A)
B)
probe
E xs Pv N H B Sa
gaattcCGGTGAGAGGAGACGGTCGTGTTTTGAAAGGCGAGAGATGAACTGAAGATTAAACATGTATGGGAATTATCCTCACTTCATTAAGTTTCCTGCGGGCTTCGGCAATTCCCCTGTTM Y G N Y P H F I K F P A G F G N S P VCATGCCAGCTCCACGTCTGTGAGCCCATCATCCAGCCTGTCCGTGGGGAGCACGGTGGAC 181H A S S T S V S P S S S L S V G S T V D 40
GGACACCACAACTACCTCGAGGCCCCCACAAACGCCTCCCGGGCGCTGCCATCCCCCATG 241G H H N Y L E A P T N A S R A L P S P M 60
AACACCATTGGGTCTCCAGTGAACGCGTTGGGCTCACCCTACAGGGTCATCGCATCCTCC 301H T I G S P V N A L G S P Y K V I A S S 80
ATTGGCTCGCATCCCGTTGCTCTGTCGTCCTCAGCCCCGGGCATGAATTTTGTGACCCAC 361I G S H P V A L S S S A P G M N F V T H 100
AGCCCACAGCCCAATGTGCTCAACAATGTCAGCAGCTCGGAGGACATCAAGCCCTTGCCA 421S P Q P N V L N N V S S S E D I K P L P 120
GGTCTACCAGGGATTGGGAACATGAATTATCCATCCACAAGCCCAGGTTCTTTAGCCAAA 481G L P G I G N M N Y P S T S P G S L A K 140
CACATCTGTGCCATCTGTGGGGACAGGTCTTCAGGGAAGCACTATGGGGTGTACAGCTGC 541H I C A I C G D R S S G K H Y G V Y S C 160
GAGGGCTGCAAGGGCTTCTTTAAGAGGACCATCCGGAAAGACCTGATCTACACCTGCCGT 601E G C K G F F K R T I R K D L I Y T C R 180
GACAACAAGGACTGCCTCATCGACAAGCGCCAGCGCAACCGCTGCCAGTACTGCCGCTAT 661D N K D C L I D K R O R N R C O Y C R Y 200
CAGAAGTGCCTCGCCATGGGGATGAAGAGGGAAGCCGTGCAGGAGGAGAGGCAGGGGAGC 721O K C L A M G M K R E A V Q E E R Q G S 220
AGGGAGCGCAGTGAGAACGAAGCTGAATCCACGAGCGGCGGCAGCGAGGACATGCCTGTG 781R E R S E N E A E S T S G G S E D M P V 240
GAGAGGATCCTGGAAGCCGAGCTGGCAGTCGAACCCAAGACGGAGGCATACAGCGACGTG 841E R I L E A E L A V E P K T E A Y S D V 260
AACACGGAGAGCTCAACCAATGACCCCGTCACCAACATCTGCCACGCTGCTGACAAGCAG 901N T E S S T N D P V T N I C H A A D K Q 280CTCTTCACCCTCGTCGAGTGGGCCAAGCGCATCCCCCACTTCTCCGACCTGACCCTGGAG 961L F T L V E W A K R I P H F S D L T L E 300
GACCAAGTCATCCTCCTGCGGGCAGGCTGGAACGAGCTGCTCATCGCATCCTTCTCCCAT 1021D Q V I L L R A G W N E L L I A S F S H 320
CGCTCCGTGTCGGTGCAGGATGGCATCCTGCTGGCCACAGGCTTGCATGTGCACCGCAGC 1081R S V S V Q D G I L L A T G L H V H R S 340
AGTGCTCACAGTGCAGGCGTGGGCTCCATCTTTGACAGAGTTTTGACAGAGCTGGTGTCC 1141S A H S A G V G S I F D R V L T E L V S 360
AAAATGAAGGACATGCAGATGGATAAGTCGGAGCTGGGGTGCCTGCGAGCCATTGTCCTC 1201K M K D M Q M D K S E L G C L R A I V L 380
TTCAACCCAGACGCCAAGGGCCTGTCCAGCCCCTCCGAAGTGGAGTCGCTGCGGGAAAAG 1261F N P D A K G L S S P S E V E S L R E K 400
GTCTACGCCACGCTGGAAGCCTACACGAAGCAGAAGTACCCCGAGCAGCCGGGGCGGTTT 1321V Y A T L E A Y T K Q K Y P E Q P G R F 420
GCCAAACTCCTCCTGCGCCTGCCAGCACTACGGTCCATCGGGCTGAAGTGCCTGGAGCAC 1381A K L L L R L P A L R S I G L K C L E H 440
CTCTTCTTCTTCAAGCTGATCGGGGACACCCCCATCGACACCTTCCTTATGGAGATGCTG 1441L F F F K L I G D T P I D T F L M E M L 460
GAGACACCCCTGCAGGTCACTTGAGGGTCCGCTCCCCCCCGCCATATCCCCACACAGCTC 1501E T P L Q V T * 467
CCACCCCTCCTCTCCGGGCTGCAAGAGCCGCCCTCCACCCTCTGCTCCTCTTTGGTGGGA 1561TTGTTGTGTCTTTGTACAGCTGTAAATCGTCCCCTCTAACCCTCCCGGTGGGTTGCGGAC 1621CCTCTGCCCAGCTAACCACACACCTCCCTGTACAGATAATTGCTTTAAATTATTTTTTC A 1681CTCTCAATAAAAGCCAACAACAACCGgaattc 1713
Fig. 1. (A) Restriction map of the insert of cDNA cloneAR2. 5' and 3' untranslated regions are shown as solidlines. The coding region is boxed. The shaded regionrepresents the putative DNA-binding domain. Sequencespresent in the probe used in northern blotting and in situhybridisation analyses are indicated. B, BamHl; Ba, Ball;E, EcoRl; H, Hinfi; N, Ncol; P, Pstl; Pv, PvwII; S, Smal;Sa, Sad; X, Xhol. (B) Nucleotide sequence and predictedamino acid sequence derived from cDNA clone AR2.EcoRi linker sequences are shown in lower case. Theputative DNA-binding domain is underlined.
therefore appears to derive from a mRNA encoding thechick homologue of hRXR-ar, or a very closely relatedprotein. The protein product of AR2 will be referred toas cRXR. The homology between cRXR and hRXR-aris significant, but less marked, in the amino-terminalregion (Fig. 2). This might simply indicate that theamino-terminal region has been relatively poorlyconserved in evolution. Alternatively, cRXR might bederived from a second member of the RXR family, orcould represent an alternatively spliced transcript of thechicken RXR-a-gene, as has been observed for RAR-y(Kastner etal. 1990; Giguere etal. 1990). The resolution
RXR in the peripheral nervous system 773
1 142/3 208/9 233/4 468
WcRXR
hRXRo*
l
37
134/5 200/1 225/6 462
hRARo-
hRARjS
hRARy
H-2RIIBP
Fig. 2. Comparison of the predicted structure of cRXRwith those of human RXR-o- (Mangelsdorf et al. 1990),RAR-o- (Petkovich et al. 1987), RAR-£ (Brand et al. 1988)and RAR-y (Krust et al. 1989), and with the proteinproduct of the cDNA clone H-2RIIBP (Hamada et al.1989) which is thought to be murine RXR-/3 (Mangelsdorfet al. 1990). The N-terminal amino acid sequence of theH-2RIIBP product is not known and is indicated by dashedlines. The positions of the putative DNA-binding andligand-binding domains of cRXR, derived by analogy withhRXR-a-are indicated. The amino acid residues at theboundary of each domain are numbered. The structures ofthe other nuclear receptors are similarly depicted. In eachdomain, the percentage amino acid identity with cRXR isindicated.
of these issues must await the cloning of other membersof the RXR gene family from human and chicken.
Northern blotting analysis of cRXR gene expressionA BamUl-EcoRI fragment of AR2 (Fig. 1A,B, nucleo-tides 786-1713) was used as a template for the synthesisof a radiolabelled antisense RNA probe for cRXRtranscripts. This probe was complementary to se-quences encoding the putative ligand-binding domainof cRXR, which are not highly conserved amongstmembers of the steroid/thyroid hormone receptorfamily. The probe did not contain sequences comp-lementary to those encoding the putative DNA-bindingdomain, which are conserved amongst members of thisfamily. This minimised the risk of cross-hybridisation ofthe probe to transcripts of other genes within thefamily. Northern blotting analysis (Fig. 3) identified asingle cRXR transcript of approximately 2.5 kb in chicklimb buds, heads and bodies at stages 22 and 25. Thistranscript was also found at low levels in a range ofadult chicken tissues, and at high levels in adult liver(data not shown). The distribution in the adult chickenis similar to that found by northern blotting in the adultrat, using the human RXR-a-probe (Mangelsdorf et al.1990).
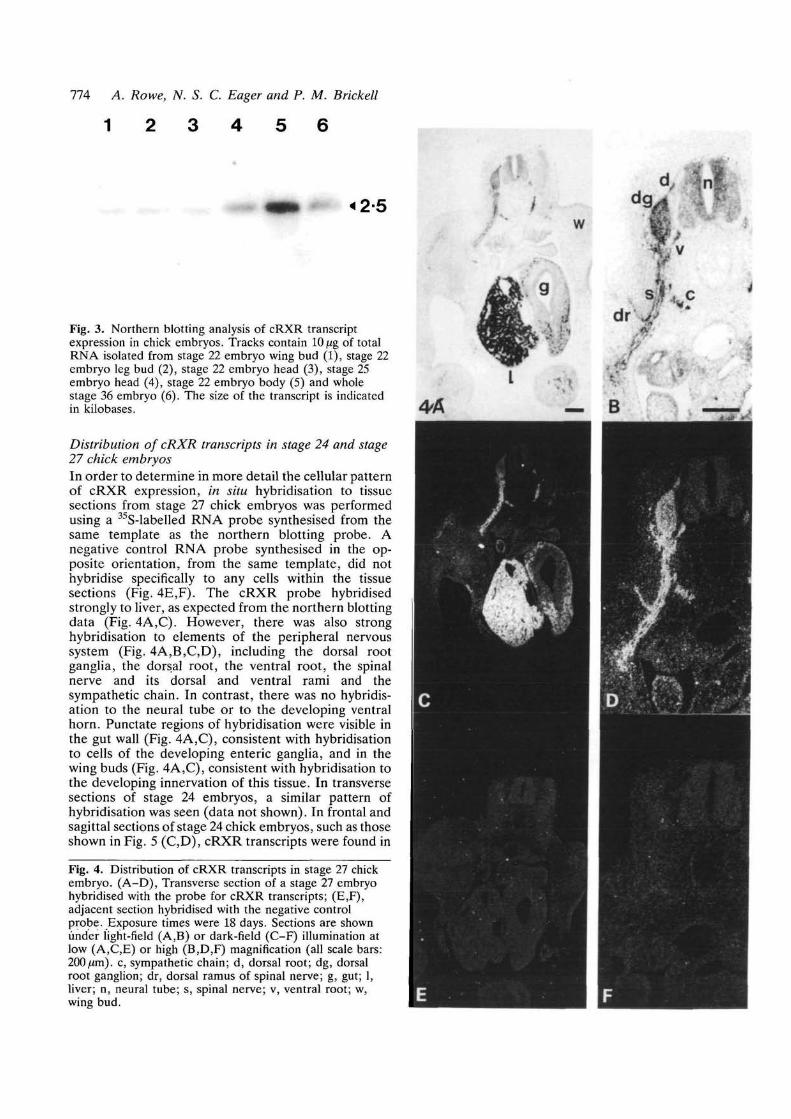
774 A. Rowe, N. S. C. Eager and P. M. Brickell
1 2 3 4 5 6
«2-5
Fig. 3. Northern blotting analysis of cRXR transcriptexpression in chick embryos. Tracks contain 10pt% of totalRNA isolated from stage 22 embryo wing bud (1), stage 22embryo leg bud (2), stage 22 embryo head (3), stage 25embryo head (4), stage 22 embryo body (5) and wholestage 36 embryo (6). The size of the transcript is indicatedin kilobases.
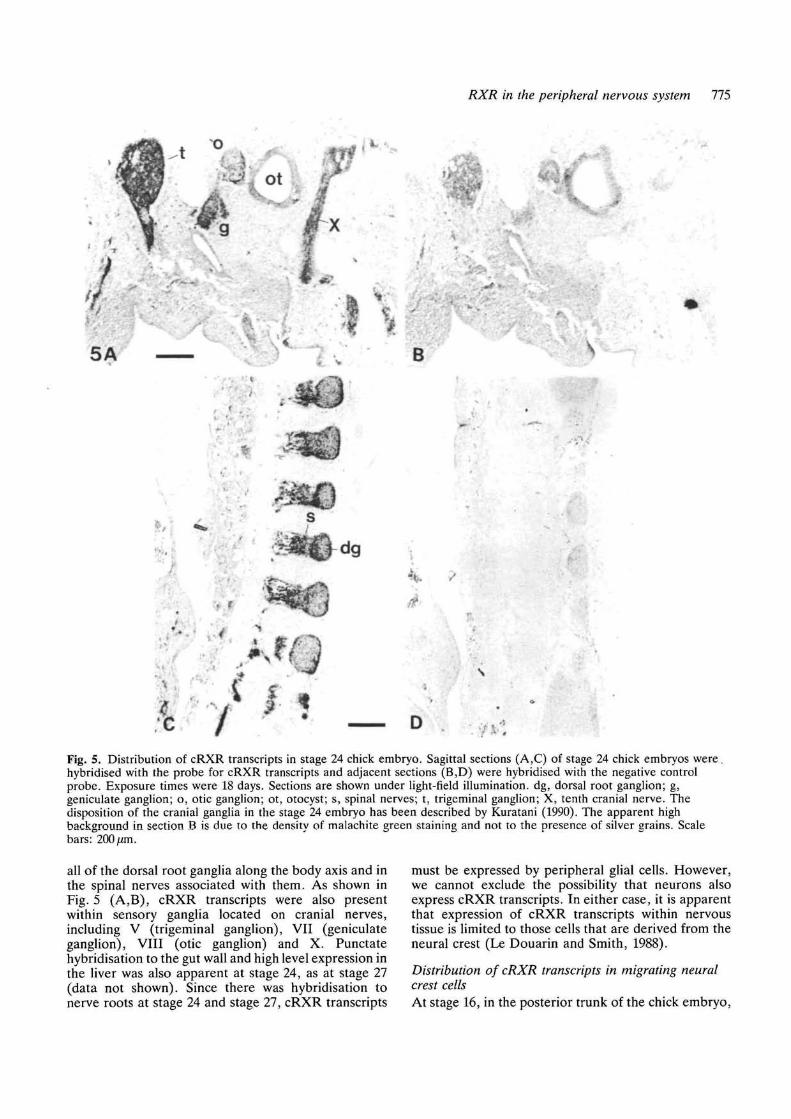
Distribution of cRXR transcripts in stage 24 and stage27 chick embryosIn order to determine in more detail the cellular patternof cRXR expression, in situ hybridisation to tissuesections from stage 27 chick embryos was performedusing a 35S-labelled RNA probe synthesised from thesame template as the northern blotting probe. Anegative control RNA probe synthesised in the op-posite orientation, from the same template, did nothybridise specifically to any cells within the tissuesections (Fig. 4E,F). The cRXR probe hybridisedstrongly to liver, as expected from the northern blottingdata (Fig. 4A,C). However, there was also stronghybridisation to elements of the peripheral nervoussystem (Fig. 4A,B,C,D), including the dorsal rootganglia, the dorsal root, the ventral root, the spinalnerve and its dorsal and ventral rami and thesympathetic chain. In contrast, there was no hybridis-ation to the neural tube or to the developing ventralhorn. Punctate regions of hybridisation were visible inthe gut wall (Fig. 4A,C), consistent with hybridisationto cells of the developing enteric ganglia, and in thewing buds (Fig. 4A,C), consistent with hybridisation tothe developing innervation of this tissue. In transversesections of stage 24 embryos, a similar pattern ofhybridisation was seen (data not shown). In frontal andsagittal sections of stage 24 chick embryos, such as thoseshown in Fig. 5 (C,D), cRXR transcripts were found in
Fig. 4. Distribution of cRXR transcripts in stage 27 chickembryo. (A-D), Transverse section of a stage 27 embryohybridised with the probe for cRXR transcripts; (E,F),adjacent section hybridised with the negative controlprobe. Exposure times were 18 days. Sections are shownunder light-field (A,B) or dark-field (C-F) illumination atlow (A,C,E) or high (B,D,F) magnification (all scale bars:200fim). c, sympathetic chain; d, dorsal root; dg, dorsalroot ganglion; dr, dorsal ramus of spinal nerve; g, gut; 1,liver; n, neural tube; s, spinal nerve; v, ventral root; w,wing bud.
RXR in the peripheral nervous system 775
• \
• ' • ' u ^ ', • • ' ' . »
Fig. 5. Distribution of cRXR transcripts in stage 24 chick embryo. Sagittal sections (A,C) of stage 24 chick embryos were,hybridised with the probe for cRXR transcripts and adjacent sections (B,D) were hybridised with the negative controlprobe. Exposure times were 18 days. Sections are shown under light-field illumination, dg, dorsal root ganglion; g,geniculate ganglion; o, otic ganglion; ot, otocyst; s, spinal nerves; t, trigeminal ganglion; X, tenth cranial nerve. Thedisposition of the cranial ganglia in the stage 24 embryo has been described by Kuratani (1990). The apparent highbackground in section B is due to the density of malachite green staining and not to the presence of silver grains. Scalebars: 200 ̂ m.
all of the dorsal root ganglia along the body axis and inthe spinal nerves associated with them. As shown inFig. 5 (A,B)> cRXR transcripts were also presentwithin sensory ganglia located on cranial nerves,including V (trigeminal ganglion), VII (geniculateganglion), VIII (otic ganglion) and X. Punctatehybridisation to the gut wall and high level expression inthe liver was also apparent at stage 24, as at stage 27(data not shown). Since there was hybridisation tonerve roots at stage 24 and stage 27, cRXR transcripts
must be expressed by peripheral glial cells. However,we cannot exclude the possibility that neurons alsoexpress cRXR transcripts. In either case, it is apparentthat expression of cRXR transcripts within nervoustissue is limited to those cells that are derived from theneural crest (Le Douarin and Smith, 1988).
Distribution of cRXR transcripts in migrating neuralcrest cellsAt stage 16, in the posterior trunk of the chick embryo,

776 A. Rowe, N. S. C. Eager and P. M. Brickell
6A
ts n
B
Fig. 6. Expression of cRXR transcripts in stage 16 chickembryo. Adjacent transverse sections of the posterior trunkregion of a stage 16 chick embryo were hybridised with theprobe for cRXR transcripts (A) or the negative controlprobe (B) and are shown under light-field illumination.Panel C is a higher power photograph of a portion of panelA. Exposure times were 18 days, c, neural crest; d,dermatome; n, notochord; s, sclerotome; t, neural tube.Scale bars: 200 /an.
neural crest cells continue to migrate into the sclero-tome, where some will differentiate into cells of thedorsal root ganglia and other elements of peripheralnervous tissue (Le Douarin and Smith, 1988). As shownin Fig. 6, cells in the neural crest and cells migratingfrom the neural crest into the sclerotome at stage 16expressed cRXR transcripts. In more anterior regionsof the stage 16 embryo, the development of theperipheral nervous system is more advanced and cRXRtranscripts were found, for example, in the trigeminalganglion (data not shown).
Discussion
We have described the isolation and characterisation ofa cDNA clone encoding a chicken homologue (cRXR)
of the recently discovered human nuclear receptorhRXR-a, and have shown that cRXR transcripts areexpressed in the liver and in elements of the peripheralnervous system that are derived from the neural crest.We have also shown that cRXR transcripts are presentin migrating neural crest cells, prior to the overtdevelopment of peripheral nervous tissue.
It is probable that there will prove to be a family ofclosely related RXR-like receptors, analogous to theRAR-like family (Mangelsdorf et al. 1990). Whilst theother members of this putative family have not yet beencharacterised, Mangelsdorf et al. (1990) have suggestedthat the cDNA clone H-2RIIBP, which was isolated byHamada et al. (1989), might represent a murine RXR-/Sspecies. Whilst cRXR and the H-2RIIBP product areclearly related, the homology between them is lessstrong than that between cRXR and hRXR-a- (Fig. 2).
Mangelsdorf et al. (1990) showed that hRXR-a canrespond to RA by activating transcription from apromoter containing a palindromic thyroid hormoneresponse element, although transactivation by hRXR-ais less sensitive to RA than is transactivation by humanRAR-cr. Our preliminary data show that cRXR can alsoactivate transcription from this promoter in response toRA.
The distribution of cRXR transcripts, which we havedescribed, is quite different to that of RAR-a-, RAR-/Sand RAR-y transcripts. This indicates that the biologi-cal roles of the RXR-iike and RAR-like receptors aredifferent. In the mouse, RAR-y transcripts are presentin various tissues whose development is sensitive toretinoid treatment, including primary mesenchyme,cartilage and differentiating squamous keratinizingepithelia (Zelent et al. 1989; Ruberte et al. 1990). Cellsexpressing RAR-y do not have a common embryologi-cal origin and include derivatives of the endoderm,mesoderm, ectoderm and cranial neural crest. Thedistribution of RAR-a and RAR-/J transcripts has notbeen described in such detail, but RAR-ar transcriptsappear to have a rather general distribution, whilstRAR-/J transcripts are restricted to particular regions ofmesenchyme that tend not to express RAR-y tran-scripts (Dolle" et al. 1989; Ruberte et al. 1990). RAR-or,RAR-/3 and RAR-y transcripts have not been found inthe peripheral nervous system at any stage of develop-ment (Dolle" et al. 1989; Ruberte et al. 1990). Thedistribution of cRXR transcripts does, however, showsome similarity to that of cellular retinoic acid-bindingprotein (CRABP) in stage 24 chick embryos (Maden etal. 1989) and in mouse embryos at similar stages(Maden et al. 1990). For example, CRABP is expressedin dorsal root ganglia, sensory nerves, sympatheticganglia and enteric ganglia. It is not clear whether theroles of cRXR and CRABP in these tissues areintegrated in any way. However, CRABP is notexpressed in the ventral root, which does express cRXRtranscripts and, conversely, high levels of CRABP arepresent in the neural tube, which does not expresscRXR transcripts. The distribution of cRXR transcriptstherefore correlates more closely with neural crestorigin than does CRABP expression, and cRXR clearly

RXR in the peripheral nervous system 111
provides a valuable molecular marker for a specificpopulation of neural-crest-derived cells.
Mangelsdorf et al. (1990) speculated that the highlevel expression of hRXR-a'in the liver may indicate arole for the hRXR- a family of receptors as regulators ofvitamin A storage, metabolism or mobilisation by thisorgan. Our data support this idea, but also indicate animportant role in the neural-crest-derived cells of theperipheral nervous system. The nature of this role isunclear. RA can induce nerve growth factor receptorexpression in immature chick sympathetic neurons (H.Rohrer, personal communication), but it is not knownwhether this is a transcriptional effect or whether thesecells express cRXR. Retinoids are known to haveteratogenic effects on the central nervous system(Durston et al. 1989; Lammer et al. 1985) and on tissuesderived from cranial neural crest cells, includingcraniofacial structures, heart and thymus (Lammer etal. 1985). However, retinoid-induced malformations ofthe trunk neural crest-derived structures of the periph-eral nervous system have not been described (Lammeret al. 1985; Smith-Thomas et al. 1987). Retinoids doinduce decreased cell-substratum adhesion in bothcranial and trunk neural crest cells in vitro (Smith-Thomas et al. 1987) and it has been suggested thisinhibits neural crest cell migration in vivo (Pratt et al.1987). We have shown that cells migrating from theneural crest of the posterior trunk at stage 16 expresscRXR transcripts, raising the possibility that cRXRmediates these effects of retinoids. However, it is likelythat the effects of retinoids on neural crest cells in vitroare mediated by the disruption of cell membranestructure rather than by the induction of changes ingene expression (Smith-Thomas et al. 1987). Similarly,experimental vitamin A deficiency in animals results inperipheral nerve degeneration, but this is secondary toa selective cessation of bone growth, rather thanrepresenting a direct effect on peripheral nervous tissue(Duchen and Jacobs, 1984). It may be that the effects ofRA upon the development and function of theperipheral nervous system are subtle, and a search forsuch effects might be informative. Alternatively, it ispossible that a closely related metabolite or structuralanalogue of RA, rather than RA itself, is the naturalligand of cRXR (Mangelsdorf et al. 1990). However, aswith the RAR family of RA receptors, a fullerunderstanding of function will require the identificationof target genes regulated in vivo.
We thank Pierre Chambon for the gift of the human RAR-orcDNA clone p63 and are grateful to Lewis Wolpert, CheryllTickle and Patrick Anderson for their comments. This work issupported by the Medical Research Council of Great Britainand by The Wellcome Trust.
References
ASSEUNEAU, D., BERNARD, B. A., BAILLY, C. AND DARMON, M.(1989). Retinoic acid improves epidermal morphogenesis. DeviBiol. 133, 322-335.
BENBROOK, D., LERNHARDT, E. AND PFAHL, M. (1988). A new
retinoic acid receptor identified from a hepatocellularcarcinoma. Nature 333, 669-672.
BRAND, N. J., PETKOVICH, M., KRUST, A., CHAMBON, P., DE THE,
H., MARCHIO, A., TIOLLAIS, P. AND DEJEAN, A. (1988).
Identification of a second human retinoic acid receptor. Nature332, 850-853.
BRICKELL, P. M. AND TICKLE, C. (1989). Morphogens in chick limbdevelopment. BioEssays 11, 145-149.
BROCKES, J. P. (1989). Retinoids, homeobox genes and limbmorphogenesis. Neuron 2, 1285-1294.
CHOMCYNSKI, P. AND SACCHI, N. (1987). Single-step method ofRNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162, 156-159.
DAVIDSON, D., GRAHAM, E., SIME, C. AND HILL, R. (1988). A
gene with sequence similarity to Drosophila engrailed isexpressed during the development of the neural tube andvertebrae in the mouse. Development 104, 305-316.
DE THE, H., DEL MAR VIVANCO-RUIZ, M., TIOLLAIS, P.,
STUNNENBERG, H. AND DEJEAN, A. (1990). Identification of aretinoic acid responsive element in the retinoic acid receptor Bgene. Nature 343, 177-180.
DEVLIN, C. J., BRICKELL, P. M., TAYLOR, E. R., HORNBRUCH, A.,
CRAIG, R. K. AND WOLPERT, L. (1988). In situ hybridisationreveals differential spatial localization of mRNAs for type I andtype II collagen in the chick limb bud. Development 103,111-118.
DOLLE, P., RUBERTE, E., KASTNER, P., PETKOVICH, M., STONER, C.
M., GUDAS, L. AND CHAMBON, P. (1989). Differential expressionof genes encoding a, B and y retinoic acid receptors andCRABP in the developing limbs of the mouse. Nature 342,702-705.
DUCHEN, L. W. AND JACOBS, J. M. (1984). Nutritional deficienciesand metabolic disorders. In Neuropathology (ed. J. H. Adams,J. A. N. Corsellis and L. W. Duchen), pp. 573-626. London:Edward Arnold.
DURSTON, A. J., TMMERMANS, J. P. M., HAGE, W. J., HENDRIKS,
H. F. J., D E VRIES, N. J., HEIDEVELD, M. AND NIEUWKOOP, P.
D. (1989). Retinoic acid causes an anteroposteriortransformation in the developing central nervous system. Nature340, 140-144.
FEINBERG, A. P. AND VOGELSTEIN, B. (1984). A technique forradiolabelling DNA restriction endonuclease fragments to a highspecific activity. Anal. Biochem. 137, 266-267.
GIGUERE, V., ONG, E. S., SEGUI, P. AND EVANS, R. M. (1987).
Identification of a receptor for the morphogen retinoic acid.Nature 330, 624-629.
GIGUERE, V., SHAGO, M., ZIRNGIBL, R., TATE, P., ROSSANT, J.
AND VARMUZA, S. (1990). Identification of a novel isoform ofthe retinoic acid receptor y expressed in the mouse embryo.Molec. cell. Biol. 10, 2335-2340.
GREEN, S. AND CHAMBON, P. (1988). Nuclear receptors enhanceour understanding of transcription regulation. Trends Genet. 4,309-314.
HAMADA, K., GLEASON, S. L., LEVI, B-Z., HIRSCHFELD, S.,
APPELLA, E. AND OZATO, K. (1989). H-2RIIBP, a member ofthe nuclear hormone receptor superfamily that binds to both theregulatory element of major histocompatibility class I genes andthe oestrogen response element. Proc. natn. Acad. Sci. U.S.A.86, 8289-8293.
HAMBURGER, V. AND HAMILTON, H. L. (1951). A series of normalstages in the development of the chick embryo. J. Morph. 88,49-52.
HOGAN, B., COSTANINI, F. AND LACY, E. (1986). Manipulating the
Mouse Embryo, a Laboratory Manual. New York: Cold SpringHarbor Press.
KASTNER, P H . , KRUST, A., MENDELSOHN, C , GARNIER, J. M.,
ZELENT, A., LEROY, P., STAUB, A. AND CHAMBON, P. (1990).Murine isoforms of retinoic acid receptor y with specific patternsof expression. Proc. natn. Acad. Sci. U.S.A. 87, 2700-2704.
KOPAN, R. AND FUCHS, E. (1989). The use of retinoic acid toprobe the relation between hyperproliferation-associatedkeratins and cell proliferation in normal and malignantepidermal cells. J. Cell Biol. 109, 295-307.
KRUSX, A., KASTNER, P., PETKOVICH, M., ZELENT, A. AND

778 A. Rowe, N. S. C. Eager and P. M. Brickell
CHAMBON, P. (1989). A third human retinoic acid receptor,hRAR-y. Proc. natn. Acad. Sci. U.S.A. 86, 5310-5314.
KURATANI, S. (1990). Development of glossopharyngeal nervebranches in the early chick embryo with special reference tomorphology of the Jacobson's anastomosis. Anat. Embryol. 181,253-269.
LAMMER, E. J., CHEN, D. T., HOAR, R. M., AGNISH, N. D.,BENKE, P. J., BRAUN, J. T., CURRY, C. J., FERNHOFF, P. M.,GRIX, A. W., LOTT, I. T., RICHARD, J. M. AND SUN, S. C.(1985). Retinoic acid embryopathy. New Engl. J. Med. 313,837-841.
LA ROSA, G. J. AND GUDAS, L. J. (1988). Early retinoic acid-induced F9 teratocarcinoma stem cell gene ERA-1: Alternatesplicing creates transcripts for a homeobox-containing proteinand one lacking the homeobox. Molec. cell. Biol. 8, 3906-3917.
LE DOUARIN, N. M. AND SMITH, J. (1988). Development of theperipheral nervous system from the neural crest. A. Rev. CellBiol. 4, 375-404.
MADEN, M., ONO, D. E. AND CHYTIL, F. (1990). Retinoid-bindingprotein distribution in the developing mammalian nervoussystem. Development 109, 75-80.
MATJEN, M., ONG, D. E., SUMMERBELL, D., CHYTIL, F. AND HIRST,E. A. (1989). Cellular retinoic acid-binding protein and the roleof retinoic acid in the development of the chick embryo. DeviBiol. 135, 124-132.
MANGELSDORF, D. J., ONG, E. S., DYCK, J. A. AND EVANS, R. M.(1990). Nuclear receptor that identifies a novel retinoic acidresponse pathway. Nature 345, 224-229.
MORRISS, G. M. AND THOROGOOD, P. V. (1978). An approach tocranial neural crest cell migration and differentiation inmammalian embryos. In Development in Mammals vol. 3 (ed.M. H. Johnson), Amsterdam: Elsevier North Holland, pp.363^11.
PETKOVICH, M., BRAND, N. J., KRUST, A. AND CHAMBON, P.
(1987). A human retinoic acid receptor which belongs to thefamily of nuclear receptors. Nature 330, 444-450.
PRATT, R. M., GOULDING, E. H. AND ABBOTT, B. D. (1987).Retinoic acid inhibits migration of cranial neural crest cells invitro. J. Craniofac. genet, dev. Biol 7, 205-217.
RUBERTE, E., DOLLE, P., KRUST, A., ZELENT, A., MORRIS-KAY, G.AND CHAMBON, P. (1990). Specific spatial and temporaldistribution of retinoic acid receptor gamma transcripts duringmouse embryogenesis. Development 108, 213-222.
SATRE, M. A. AND KOCHHAR, D. M. (1989). Elevations in theendogenous levels of the putative morphogen retinoic acid inembryonic mouse limb-buds associated with limbdysmorphogenesis. Devi Biol. 133, 529-536.
SMITH-THOMAS, L., LOTT, I. AND BRONNER-FRASER, M. (1987).Effects of isotretinoin on the behaviour of neural crest cells invitro. Devi Biol. 123, 276-281.
THALLER, C. AND EICHELE, G. (1987). Identification and spatialdistribution of retinoids in the developing chick limb bud.Nature 327, 625-628.
TICKLE, C , ALBERTS, B., WOLPERT, L. AND LEE, J. (1982). Localapplication of retinoic acid to the limb bud mimics the action ofthe polarising region. Nature 296, 564-566.
WEDDEN, S. E. (1987). Epithelial-mesenchyme interactions in thedevelopment of chick facial primordia and the target of retinoidaction. Development 99, 341-351.
WILKINSON, D. G., BAILES, J. A. AND MCMAHON, A. P. (1987).Expression of the proto-oncogene int-1 is restricted to specificneural cells in the developing mouse embryo. Cell 50, 79-88.
ZELENT, A., KRUST, A., PETKOVICH, M., KASTNER, P. ANDCHAMBON, P. (1989). Cloning of murine a and /S retinoic acidreceptors and a novel receptor predominantly expressed in skin.Nature 339, 714-717.
(Accepted 7 December 1990)














![In Vitro Assays with Rock Shell RXR · In Vitro Assays with Rock Shell RXR 0510 [3H] - 9-cis RA (nM)Specific binding (dpm x 10-3) 0 0.2 0.4 0.6 0.8 1 bound (nM) Bound/free Kd = 15.2](https://img.pdfslide.net/doc/110x75/5f7a328515a1ad100532adca/in-vitro-assays-with-rock-shell-in-vitro-assays-with-rock-shell-rxr-0510-3h-.jpg)