Embed Size (px)
Citation preview

International Journal of Food Microbiology, 8 (1989) 225-232 225 Elsevier
JFM 90006
A new colorimetric nucleic acid hybridization assay for Listeria in foods
W. King, S. Raposa, J. Warshaw, A. Johnson, D. Halbert and J.D. Klinger
GENE- TRAK Systems, Framingham, MA, U.S.A.
Non-isotopic colorimetric detection has been applied in a rapid nucleic acid dipstick hybridization assay for detection of Listeria spp. in food and environmental samples. The assay takes approximately 2.5-3 h following a two day broth and plate enrichment. Hybridization occurs between fluorescein labeled detector probes, poly(deoxyadenosine)-tailed capture probes, and Listeria-specific regions of 16 S ribosomal RNA. These target:probe complexes are captured on poly(deoxythymidine) coated plastic dipsticks. Detection is based on binding of horseradish peroxidase conjugated anti-fluorescein antibody to the hybridization complex, and enzyme-mediated color development. The colorimetric endpoint is read on a photometer at 450 nm. In these initial studies, 306 inoculated dairy, meat and seafood samples, and 200 environmental samples were tested. When compared with total culture results the hybridization assay had unconfirmed positive and false-negative rates of approximately 1.4-2.9% and 0.8-4.7%, respectively.
Key words: Listeria; Nucleic acid hybridization; Bacterial identification; Rapid methods
Introduction
Con tamina t ed foods have now been firmly established as agents for t ransmiss ion of h u m a n listeriosis (Archer and Young, 1988; Hird, 1987; L i n n a n et al., 1988; Seeliger, 1988). This realization has been accompanied by intensive studies to develop more sensitive and reliable methods for detect ion of Listeria in food
products . Recent efforts in this regard include improved enr ichment media and cul ture methods; procedures which enhance recovery of Listeria from part icular food types (milk, meats, etc.); rapid methods such as flow-cytometry, ELISA, and nucleic acid hybridizat ion; selective and differential media for direct enumera t ion of Listeria; and, simplified culture methods for screening and ident if icat ion of suspect Lister ia isolates. Unfor tuna te ly , there have been few studies compar ing the efficacy of various methods on identical samples, bu t it appears that even the improved cul tural methods still suffer f rom lack of sensitivity a n d / o r precision (Klinger,
Correspondence address." J.D. Klinger, GENE-TRAK Systems, 31 New York Avenue, Framingham, MA 01701, U.S.A.
0168-1605/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

226
1988; Doyle and Schoeni, 1987; Truscott and McNab, 1988). We previously reported development of a nucleic acid hybridization assay for
detection of Listeria spp. in foods and environmental samples (Klinger et al., 1988; Klinger and Johnson, 1988). The assay was based on hybridization of 32p-labeled synthetic DNA oligomer probes with genus-specific regions of 16 S rRNA in treated samples after two days of culture enrichment. Following cell lysis, Listeria rRNA was trapped onto a nylon membrane which served as the hybridization matrix. Detection of the hybridized labeled probe was by counting beta radiation on the filters and comparison to positive and negative controls. This assay was initially optimized for dairy foods and environmental samples. Results for indicated sample types compared very favorably with reference (e.g., FDA) culture methods. In this paper, we report our preliminary experience with a new, non-isotopic colorimetric dipstick hybridization assay for Listeria spp. in foods and environmental samples. The new procedure requires about 2.5 h after a two day enrichment period with the endpoint being read on a simple photometer or spectrophotometer.
Materials and Methods
Probes
Like the isotopic hybridization version, the basis of the colorimetric assay is detection of hybrids consisting of unique regions of Listeria 16 S rRNA and synthetic, labelled DNA oligomer probes. In the colorimetric assay, two types of probes are employed, i.e., fluorescein labeled 'detector' probes and an unlabeled 'capture probe'. Details of probe development have been previously described (Klinger et al., 1988). Listeria 16 S rRNA sequences were obtained from the type strain of Listeria mono~ytogenes (ATCC 15313) using the reverse transcriptase terminating method (Lane et al., 1985). Prediction of unique regions likely to be useful as probes was done by comparison of Listeria 16 S sequences with those of other related genera. Probes were prepared on BioSearch 8600 or Applied Biosys- terns 380A synthesizers. Detector probes were terminated with cytidine nucleoside modified at the C-4 position with a diaminopropyl spacer. These oligomer probes were purified by reverse-phase HPLC prior to reaction with fluorescein iso- thyocyanate to yield sequences of approximately 30-50 bases containing two fluorescein moieties. Probes were additionally purified using Sephadex G-25 chro- matography and by HPLC. Capture probes were prepared in similar fashion without modified nucleotides. Prior to final purification, the probes were elongated at the 3' terminus by reaction with dATP and terminal transferase to yield 'tails' of approximately 100-200 deoxyadenosine monophosphate residues. The final probe cocktail contained detector probes and capture probes (approx. 600 ng /ml and 200 ng/ml , respectively) in 5 M GuSCN, 80 mM EDTA, 0.4 M TRIS, pH 8.0, stored at room temperature. This probe mixture was tested for specificity in the format as described below.

227
Assay method
To characterize assay specificity, strains of Listeria and non-Listeria organisms were grown overnight in modified Listeria enrichment broth ( 'MEB') , tryptic soy broth, or brain heart infusion broth (Difco) at 35 ° C, or other modified appropriate temperatures to assure dense growth (approx. 108-109/ml). Cultures were diluted 1 : 100 in broth prior to assay. The majority of strains were food and clinical isolates obtained from reference culture collections (Klinger et al., 1988). Others (especially non-Listeria competitor organisms) were obtained from the ATCC, or isolated from retail food samples and identified in our laboratories using appropriate systems (e.g., API R Rapid STREP) and supplementary reactions (motility, CAMP reactions, etc.).
The assay was performed on enrichment cultures of various food and environ- mental samples. 25 g samples of food were mixed in 225 ml of modified Lovett 's Listeria enrichment broth ( 'MEB') , buffered by the addition of MOPS buffer (3-[N-morpholino] propanesulfonic acid; see Klinger et al., 1988), and incubated 20-24 h at 35 o C. For inoculated food studies, samples were obtained from retail outlets in Massachusetts. 0.1 ml volumes of dilutions of overnight broth cultures of challenge organisms ( L. monocytogenes, L. innocua and L. ivanovii ) were mixed with 25 g of each dairy food sample in 225 ml MEB, or 1 g of each meat or seafood sample in 100 ml MEB, to achieve inoculum levels of approx. 0 (control), 10, or 100 cells per sample. Swabs from commercial food manufacturing environmental sources were added directly to smaller volumes (10-25 ml) of either MEB or Listeria enrichment broth (as described by McClain and Lee, 1988), and incubated at 35 °C for 20-24 h. Multiple swabs were obtained from each site in an at tempt to provide duplicate samples for assay by hybridization (MEB enrichment) and by the USDA culture method. Following primary enrichment of either foods or environmental samples, volumes of approx. 0.125 ml of the selective broths were spread with sterile cotton or dacron-tipped applicators over the entire surfaces of 100 mm plates containing LPM agar (Listeria plating medium; McClain and Lee, 1988). Plates were incubated an additional 20-24 h at 35°C prior to the hybridization assay. Following this second incubation, material (presumed growth) was removed from the surface of each LPM plate using a sterile applicator and suspended by vigorous mixing for at least 5 s in 1 ml PBS. Partial cell lysis of organisms in test samples (0.3 ml of the PBS suspension in 12 x 75 mm glass tubes) was obtained in a two-step process, i.e., initially by the addition of 0.1 ml of enzyme solution containing lysozyme 3-7.5 m g / m l (Sigma), mutanolysin 150-750 un i t s /ml (Sigma), 10 mM EDTA, 100 mM TRIS, and incubation at 37°C for 15 min. This was followed by a second 15 min 37°C incubation after the addition of 0.1 ml lysis solution contain- ing proteinase K, 5 m g / m l , (Boerhinger Mannheim) and Sarkosyl 50 m g / m l (Sigma). Probe solution containing 5 M GuSCN was preheated at 65°C and immediately added (0.5 ml) to the sample tubes in a 37 °C waterbath. A polystyrene 'dipstick', coated in a proprietary fashion with deoxythymidine homopolymer [poly(dT)] was added to each sample tube and the incubation at 37°C was continued for 60 rain. If Listeria is present in the sample, hybridization of both

228
detector and capture probes to specific 16 S rRNA targets occurs. Additionally, during this incubation these probe: target complexes will bind to the solid dipstick surfaces via poly(dA):poly(dT) interactions. Dipsticks were removed from the sample tubes and washed extensively in 0.15 M NaC1, 0.05 M TRIS, 0.002 M EDTA, Tween 20, 1 mg/ml , pH 7.5 at room temperature and then briefly at 65 ° C. Dipsticks were then added to 12 × 75 mm tubes containing 0.75 ml of appropriately diluted rabbit anti-fluorescein antiserum conjugated to horseradish peroxidase, and incubated for 20 min at room temperature. After incubation with enzyme conjugate, dipsticks were washed a second time, blotted, and added to tubes containing 0.75 ml of substrate-chromogen solution (H20 2 and tetramethyl benzidine) and incubated for 30 min at room temperature. Color development was arrested by the addition of 0.25 ml of 4 N HzSO 4 following the removal of the dipsticks. Reactions of test samples were read at 450 nm using a spectrophotometer or GENE-TRAK R pho- tometer and compared to those of positive and negative controls. Controls consisted of 1 : 100 dilutions of a frozen or formalin-killed suspension of L. monocytogenes (positive) and an undiluted formalin-killed overnight culture of Streptococcus faecium. Absorbance values of 0.1 or greater were considered positive for the presence of Listeria.
Culture methods
Samples were assayed by both the colorimetric hybridization assay and by at least one standard reference culture procedure in a blind fashion. Results from the two methods were then compared to determine relative assay performance, Dairy foods (25 g samples) were cultured as recommended in the FDA procedure (Lovett, 1988). One gram samples of meats or seafoods were processed according to the USDA culture method. In addition, samples from both the MEB primary enrich- ments and the material suspended in PBS from the secondary LPM plates (i.e., the actual samples used in the hybridization assay) were streaked to LPM for recovery of Listeria. Data from this latter procedure are referred to in the Results section as ' total culture' results. In our experience, isolation rates using this approach are often somewhat higher than those of the current standard reference methods.
Results
A total of 291 Listeria strains were tested (1 : 100) in the colorimetric hybridiza- tion assay (Table I); all were positive. The majority (195) were L. rnonocytogenes, though all other Listeria spp. were included, and all major serotypes were repre- sented. Test results (A450) for 284 of the 291 strains (97.6%) were above 0.5; 7 strains gave A450 values between 0.25 and 0.50. To mimic 'worst-case' situations, overnight broth cultures of non-Listeria organisms were used undiluted to determine exclusivity. A total of 65 strains representing 59 species were tested. Gram-positive genera included, Bacillus, Brochothrix, Clostridium, Corynebacterium, Enterococcus, Kurthia, Lactobacillus, Micrococcus, Rhodococcus, Staphylococcus, and Streptococcus.
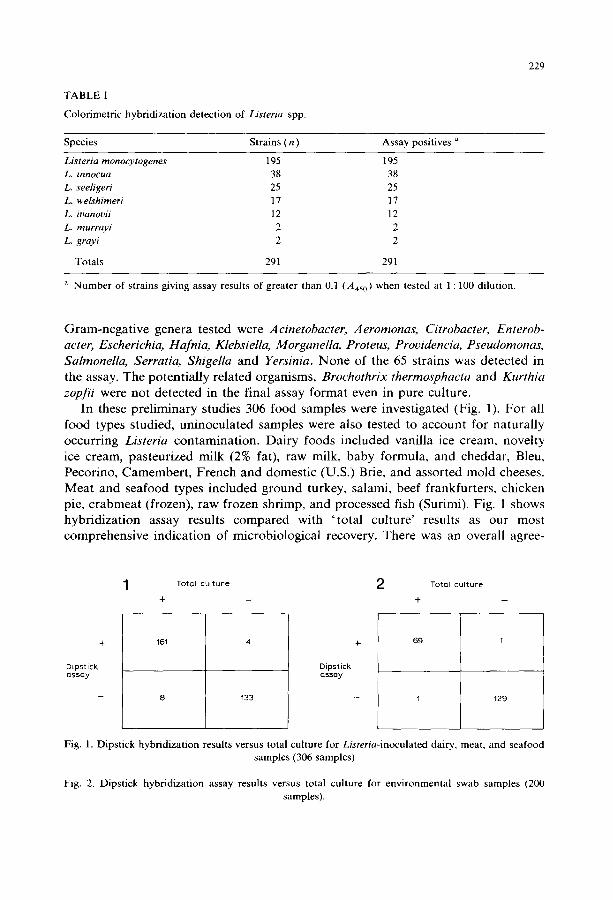
TABLE I
Colorimetric hybridization detection of Listeria spp.
229
Species Strains ( n ) Assay positives a
Listeria monocytogenes 195 195 L. innocua 38 38 L. seeligeri 25 25 L. welshimeri 17 17 L. ioanovii 12 12 L. murrayi 2 2 L. grayi 2 2
Totals 291 291
a Number of strains giving assay results of greater than 0.1 (A450) when tested at 1 : 100 dilution.
G r a m - n e g a t i v e genera tested were Acinetobacter, Aeromonas, Citrobacter, Enterob- acter, Eseherichia, Hafnia, Klebsiella, Morganella, Proteus, Prouidencia, Pseudomonas, Salmonella, Serratia, Shigella and Yersinia. N o n e of the 65 s trains was de tec ted in the assay. The po ten t i a l ly re la ted organisms, Brochothrix thermosphacta and Kurthia zopfii were not de tec ted in the final assay fo rmat even in pure culture.
In these p re l imina ry studies 306 food samples were inves t iga ted (Fig. 1). F o r all food types s tudied, un inocu la ted samples were also tested to account for na tura l ly occur r ing Listeria con tamina t ion . Da i ry foods inc luded vani l la ice cream, novel ty ice cream, pas teur ized mi lk (2% fat), raw milk, baby formula , and cheddar , Bleu, Pecorino, Camember t , F r ench and domes t ic (U.S.) Brie, and assor ted mold cheeses. Mea t and seafood types inc luded g round turkey, salami, beef f rankfur ters , chicken pie, c r a b m e a t (frozen), raw frozen shr imp, and processed fish (Surimi). Fig. 1 shows hybr id i za t ion assay results c o m p a r e d with ' t o t a l cul ture ' results as our most comprehens ive ind ica t ion of microbio log ica l recovery. There was an overal l agree-
T o t a l c u l t u r e 2 To ta l c u l t u r e
+ - - + - -
+
D i p s t i c k a s s a y
161 4
8 133
+
D i p s t i c k a s s a y
6 9 1
1 129
Fig. 1. Dipstick hybridization results versus total culture for Listeria-inoculated dairy, meat, and seafood samples (306 samples)
Fig. 2. Dipstick hybridization assay results versus total culture for environmental swab samples (200 samples).

230
ment of 96.1% (294/306) between the colorimetric assay and culture. Positive hybridization results were unconfirmed for four samples yielding an apparent false-positive rate of 2.9%. Results for 8 culture-positive samples were below the A45 o cut-off value of 0.1 and were scored as negative, for a false-negative rate of 4.7%. In these experiments, dairy products were also assayed using the FDA culture procedure (Lovett, 1988), and meats and seafoods were cultured using the method suggested by the USDA (McClain and Lee, 1988). Eighty-four meat or seafood samples were tested by hybridization and culture. Eight of 53 (15.1%) were negative by the USDA culture method; all 53 were detected by hybridization. Likewise, among the 222 dairy product samples, 108 of 112 hybridization positive samples were confirmed by at least one culture method. Only 47 of the 222 samples were culture positive by the FDA procedure, resulting in a false-negative rate for that method of 57.6%. Twenty sites (200 swabs) in food manufacturing environments were sampled with replicate swabs and these samples were assayed by both hybridization and the USDA culture method. Among these samples there was nearly complete agreement (198/200) between colorimetric assay results and total culture (Fig. 2). Not shown in Fig. 2 is the direct comparison between the USDA method and hybridization method results on 'matched ' swabs taken from identical environmental sites. Somewhat more sites were scored as Listeria positive by the USDA method, yielding an apparent hybridization false-negative rate of approxi- mately 10%. It should be noted, however, that the comparison assays were not done on identical samples, i.e., one swab was placed in MEB for the hybridization assay, and a theoretically 'matched ' swab was processed in Listeria enrichment broth for USDA culture. The choice of MEB as the primary enrichment for the hybridization method was based on our observation of superior growth of Listeria isolated from food and environmental sources. In five of the seven instances in which Listeria were recovered by the USDA method and the matched sample was hybridization negative, no Listeria were isolated on LPM subculture from either the primary MEB or from material suspended from the secondary LPM plate. After accounting for such sampling errors, we found that the assay gave a false-positive rate of 0.8% and a false-negative rate of 2.7%. In other experiments using such ' matched environmen- tal samples', the situation was nearly exactly reversed with the reference culture method showing a significant false-negative rate when compared to hybridization (data not shown). We believe that this suggests that the bulk of the disparity between culture and hybridization results for these environmental samples is due to sampling error in which the swab for one method actually contacted and contained Listeria cells, whereas the 'matched ' swab did not.
Discussion
The problem of detecting Listeria in contaminated foods illustrates a number of points characteristic of new trends in food microbiology (Archer and Young, 1988). Listeria as it relates to foods has been considered a 'new and emerging' organism

231
and more work is needed to establish the precise significance of its presence, and its natural history and ecology in the production, storage, and processing of a large variety of food types. Given these uncertainties and the high potential mortality rates associated with listeriosis, it is unlikely that regulatory agencies will be able to rationally assign acceptable, safe, threshold levels for the presence of Listeria in foods. This is further substantiated by the very remote possibilities for obtaining relevant information on human oral infectious doses from animal experiments. The major portion of all the literature on Listeria concerns it use as a tool in immunological studies and underscores species, host strain, virulence and challenge route variability in resultant outcomes in natural and experimental challenges. Additionally, until about two years ago relatively little emphasis had been placed on diagnostic analytical methods particularly optimized for detection of Listeria in foods.
With these points in mind we previously developed a hybridization assay for all Listeria spp. in foods and environmental samples using isotopic detection. The method was rapid and gave results which compared very favorably with the FDA culture method. The latter procedure was chosen as the reference culture method for two reasons: (1) at the time we began our studies it was the only 'official ' regulatory method available in the U.S., and (2) we had designed our assay for dairy products and these are conventionally assayed using FDA-sanctioned methods. We chose to use probes to detect all Listeria spp. (i.e., not just L. monocytogenes) so that the assay could be used as a broad screen to rapidly eliminate negatives, and to detect species of Listeria other than L. monocytogenes which may be present as indicator organisms. The assay has performed well and has been well-received; however, the need for the isotopic probe and the restricted product claims (dairy and environ- mental samples) have somewhat limited its utility.
The colorimetric assay format described here is very simple to use. Our initial studies indicate that it is at least as sensitive and specific as standard reference culture methods, and in some instances, more so. Also, as the need has become apparent for testing meats and seafoods, we have begun to build a data base on the methods ' performance with these product types. Another appealing aspect of the dipstick format is its potential application to assays for other foodborne pathogens. We have recently reported its use as a model system for detection of Yersinia (Chan et al., 1988a), and as a fully developed assay for Salmonella (Chan et al., 1988b).
The present study has introduced an enrichment approach consisting of 24 h in MEB followed by a period of growth on the surface of LPM agar plates. Material consisting of the original inoculum and apparent growth is removed from the plate using a sterile swab and suspended for the hybridization assay. We have found this procedure to be superior to methods (as we originally proposed) employing sequen- tial broth enrichments. We feel that enhanced recovery from the surface of a selective plate probably relates to reduction of one or more anti-Listeria inhibitory activities of competitor organisms which is more apparent in a broth milieu. In more recent experiments we have found that transfer of up to 0.5 ml of enrichment broth to the selective plate is not only feasible and convenient, but that the overall assay performance (and documented Listeria recovery) is significantly increased. Further

232
investigation of the potential basis for the increased recovery is underway and will be reported fully elsewhere.
Finally, we wish to emphasize that we have presented initial results for this new colorimetric hybridization assay. A configuration of the assay more closely ap- proximating a commercially available product has very recently been described (King et al., 1989).
References
Archer, D.L. and Young, F.E. (1988) Contemporary issues: diseases with a food vector. Clin. Microbiol. Rev. 1, 377-398.
Chan, S., Pitman, T., Shah, J., King, W. and Lawrie, J. (1988a) Non-radioactive DNA assay for detection and identification of food-borne Yersinia. Abstr. Annu. Meet. Am. Soc. Microbiol., p. 280.
Chan, S.W., Wilson, S., Hsu, H-Y, King, W., Halbert, D.N. and Klinger, J.D. (1988b) Model non-iso- topic hybridization systems for detection of foodborne bacteria: preliminary results and future prospects. In: S.-d. Kung (Ed.), Proc. Int. Symp. Biotechnology and Food Quality, in press.
Dormelly, C.W. (1988) Historical perspectives on methodology to detect Listeria monocytogenes. J. Assoc. Off. Anal. Chem. 71, 644-646.
Doyle, M.P. and Schoeni, J.L. (1987) Comparison of procedures for isolating Listeria mono~ytogenes in soft, surface-ripened cheese. J. Food Protect. 50, 4-6.
Hird, D.W. (1987) Review of evidence for zoonotic listeriosis. J. Food Protect. 50, 429-433. King, W., Raposa, S., Warshaw, J., Johnson, A., Halbert, D. and Klinger, J.D. (1989) Detection of
Listeria in foods using nucleic acid probes. In: A.J. Miller and J.L. Smith (Eds.), Proc. Comprehen- sive Conference on Listeria monocytogenes, Soc. Indust. Microbiol., in press.
Klinger, J.D. (1988) Isolation of Listeria: a review of procedures and future prospects. Infection 16, Suppl. 2, $98-S105.
Klinger, J.D., Johnson, A., Croan, D., Flynn, P., Whippie, K., Kimball, M., Lawrie, J. and Curiale, M. (1988) Comparative studies of a nucleic acid hybridization assay for Listeria in foods. J. Assoc. Off. Anal. Chem. 71,669-673.
Klinger, J.D. and Johnson, A.R. (1988) A rapid nucleic acid hybridization assay for Listeria in foods. Food Technol. 42, 66-70.
Lane, D.J., Pace, B., Olse, G.J., Stahl, D.A., Sogin, M.L. and Pace, N.R. (1985) Rapid determination of 16 S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. U.S.A. 82, 6955-6959.
Linnan, M.J., Mascola, L., Lou, X.D., Goulet, V., May, S., Salminen, C., Hird, D.W., Yonekura, L., Hayes, P., Weaver, R., Audurier, A., Plikaytis, B.D., Fannin, S.L., Kieks, A. and Broome, C.V. (1988) Epidemic listeriosis associated with Mexican-style cheese. N. Engl. J. Med. 319, 823-828.
Lovett, J. (1988) Isolation and identification of Listeria monocytogenes in dairy products. J. Assoc. Off. Anal. Chem. 71,658-660.
Mclain, D. and Lee, W.H. (1988) Development of USDA-FSIS method for isolation of Listeria rnonocytogenes from raw meat and poultry. J. Assoc. Off. Anal. Chem. 71,660-664.
Seeliger, H.P.R. (1988) Listeriosis - history and actual developments. Infection 16, Suppl., $80-$84. Truscott, R.B. and McNab, W.B. (1988) Comparison of media and procedures for isolation of Listeria
rnonocytogenes from ground beef. J. Food Protect. 51,626-628.





























