Embed Size (px)
Citation preview

The University of Southern MississippiThe Aquila Digital Community
Honors Theses Honors College
Spring 5-11-2012
A Phylogeny of Samydaceae Based on NuclearGBSSI and EMB2765 DNA SequencesChelsa WilliamsThe University of Southern Mississippi
Follow this and additional works at: http://aquila.usm.edu/honors_theses
Part of the Life Sciences Commons
This Honors College Thesis is brought to you for free and open access by the Honors College at The Aquila Digital Community. It has been acceptedfor inclusion in Honors Theses by an authorized administrator of The Aquila Digital Community. For more information, please [email protected].
Recommended CitationWilliams, Chelsa, "A Phylogeny of Samydaceae Based on Nuclear GBSSI and EMB2765 DNA Sequences" (2012). Honors Theses.Paper 70.

The University of Southern Mississippi
A Phylogeny of Samydaceae Based on Nuclear GBSSI and EMB2765 DNA Sequences
by
Chelsa N. Williams
A Thesis
Submitted to the Honors College of The University of Southern Mississippi
in Partial Fulfillment of the Requirements for the Degree of
Bachelor of Science in the Department of Biological Sciences
May 2012


Approved by
_________________________________
Mac H. Alford, Associate Professor
Department of Biological Sciences
_________________________________
Glen Shearer, Jr., Chair
Department of Biological Sciences
_________________________________
David R. Davies
Honors College, Dean
iii

ACKNOWLEDGMENTS
I would like to take this opportunity to express my gratitude to some key people
without whom this thesis would have never been possible. Thanks for the opportunities,
your time, and your patience, Mrs. Paula Mathis and Dean Dave Davies of the
University’s Honors College. Thanks to Mrs. Liliana Hernández for being there to answer
any questions I may have had along the way. A big thank you and mention of recognition
is due for Mrs. Tharanga Samarakoon for being a mentor and huge help to me throughout
this project, meeting me when it was not convenient and answering all of my questions.
The biggest thank you goes to Dr. Mac H. Alford for a multitude of reasons: for giving
me the opportunity for this thesis, for the meetings, for answering all of my questions, for
the patience shown, for the countless returns of e-mail during e-mail correspondence, and
for being a mentor.
iv

ABSTRACT
Samydaceae are a tropical family of woody flowering plants, poorly known
because they were once included in the ‘dustbin’ family Flacourtiaceae and because there
are few economically important species. The family has recently been studied using DNA
sequence data, but most of the data were from plastid DNA and some relationships had
low statistical support. For this study, two nuclear regions, EMB2765 and GBSSI, were
sequenced in order to affirm or refute the relationships inferred from plastid DNA and in
order to find stronger statistical support for relationships. The data collected mostly
affirm previously collected data from plastid DNA.
v

TABLE OF CONTENTS
Title Page...............................................................................................................................i
Approval Page .......................................................................................................................iii
Acknowledgments .................................................................................................................iv
Abstract..................................................................................................................................v
Table of Contents...................................................................................................................vi
Chapter 1: Introduction..............................................................................................1
Chapter 2: Materials and Methods.............................................................................5
Chapter 3: Results......................................................................................................11
Chapter 4: Discussion................................................................................................17
References .............................................................................................................................19
vi

1
Chapter 1: INTRODUCTION
Plants, especially the great majority of tracheophytes considered to be flowering
plants, are extremely important to humans in the areas of nutrition, medicine, food,
shelter, tools, dyes, and overall welfare (Judd et al., 2007). They have impacts on us
ecologically and economically. Plant systematics is the science of plant diversity in
which the fundamental aim is to study the biological diversity of plants on Earth today
and their evolutionary history. A secondary aim is to convey this knowledge about
diversity and relationships in the form of phylogenies and classifications (Judd et al.,
2007). According to Judd et al. (2007), this science works by “putting forward
hypotheses about the existence of branches of the tree of life and testing them with
evidence.” While some think that systematics is separate from other areas of biology,
this area plays an important role in other disciplines such as linguistics, philosophy,
ecology, and molecular biology (Judd et al., 2007). Judd et al. (2007) also maintain that
“systematics is essential to our understanding of and communication about the natural
world.”
While there are many different ways in which to construct a phylogeny, we
attempt to construct one based on evolutionary relationships and root it by using a
relative of the group known as an outgroup. The members of the ingroup, the group being
studied, are more closely related to each other than they are to the outgroup(s) being used
(Judd et al., 2007). In this study, the outgroups used are Lacistema aggregatum
(Lacistemataceae), Hasseltia floribunda, and Prockia crucis (Salicaceae). The ingroup
members originally came from a very encompassing, pantropical family known as

2
Flacourtiaceae (Chase et al., 2002). Warburg (1894), Gilg (1925), Hutchinson (1967),
Lemke (1988), and Takhtajan (1997) are all well-known botanists who worked on
Flacourtiaceae, and all of them recognized that relationships within the family and
relationships of the family to other families was difficult to ascertain. The family has
always been difficult to define and characterize because of various reasons, including
predominant character states shared by many other families and flowers too small to be
easily examined by hand, even with a hand lens (Chase et al., 2002). Over the years,
Flacourtiaceae have earned a reputation as a “catchall” or “dustbin” group. Alwyn Gentry
(2003) expressed, “If you don’t have any idea what family it is, try Flacourtiaceae or
Euphorbiaceae,” and Williams (1965) proclaimed, “When in doubt, put it in the
Flacourtiaceae.”
With the coming of age of molecular biology, relationships of Flacourtiaceae
have begun to be clarified because DNA provided more characters. Molecular studies
have used DNA from three different cellular sources: mitochondrial DNA, plastid DNA,
nuclear DNA, or some combination thereof. Mitochondrial DNA, in general, is not
variable enough to be useful in plants, and copies of nuclear genes are far less abundant
and more complicated than plastid genes. Thus, most studies of plants have, until
recently, utilized plastid DNA as the major data set for studies of relationships.
Based on plastid DNA data, Chase et al. (2002) demonstrated a split into two
major lineages within Flacourtiaceae, and Alford (2003, 2005, in prep.) later reported the
best division of one of those lineages was into the two major families of Samydaceae and
Salicaceae sensu medio.

3
According to Alford (2003, 2005, in prep.), Samydaceae include about 256
species of the former Flacourtiaceae. Many plants within this “new” family have been
reported to have medicinal, insecticidal, fungicidal, and cytotoxic components (Anaya et
al., 2004; Beutler et al., 2000; Itokawa et al., 1990; Jullian et al., 2005), to accumulate
metals at some of the highest levels ever recorded (Jaffré et al., 1979), and to be common
elements of tropical forests worldwide (Phillips & Miller, 2002; Guilherme et al., 1998;
Silvra Jr. et al., 1998). Unfortunately, the family has received very little attention
seemingly due to its lack of economically important members. Chase et al. (2002) only
sampled one species from this newly resurrected family, not giving much insight into this
family. However, some species of Casearia have been shown to have inhibitory activity
against snake venom phosphordiesterase I (Chai et al., 2010) and to have anti-ulcer, anti-
inflammatory, anti-ophidian, and antitumor potentialities (Ferreira et al., 2011). Some
species within Samydaceae are also sporadically used for lumber (Chudnoff, 1984;
Richter & Dallwitz, 2000). The potential for economical uses is out there.
Currently, the ongoing research goal is to infer the phylogeny of and revise the
classification of the Samydaceae. Previously, plastid genes were used in an attempt to
disentangle existing relationships, but these genes posed a multitude of problems. Due to
hybridization, plants can contain plastids from other species, thus potentially resulting in
misleading phylogenies. Plastid DNA also evolves moderately slowly, resulting in data
that may not be variable enough for inferring relationships between closely related genera
and species. This study aims to use nuclear regions to provide more insight and resolution
to the existing relationships within Samydaceae, with the hope that the nuclear DNA has
more variation than the plastid DNA. Each cell contains only one nucleus with two

4
genomic sets, one from “mom” and one from “dad.” The genes chosen for this study
presumably have only one copy per set, though the potential for two or more alleles does
exist. Alford et al. (unpubl. results) have found Casearia to be polyphyletic based on
previous plastid DNA, and those data have shown that there are several other genera
nested within the large genus. Because nuclear DNA regions in other plants have shown
great variability and exhibit a different mode of inheritance than plastids, the hypotheses
for this study are:
1. Nuclear DNA regions in Samydaceae are expected to shower greater variation
than plastid regions, providing more data for inference of phylogenetic history,
and
2. Analyses of the nuclear DNA data will reveal a pattern of relationships identical
to or more resolved than (i.e., not conflicting with) those from analyses of plastid
DNA.

5
Chapter 2: MATERIALS AND METHODS
Species of Samydaceae and three outlier species from Lacistemataceae and
Salicaceae were previously collected, had DNA extracted using a Qiagen DNeasy® Plant
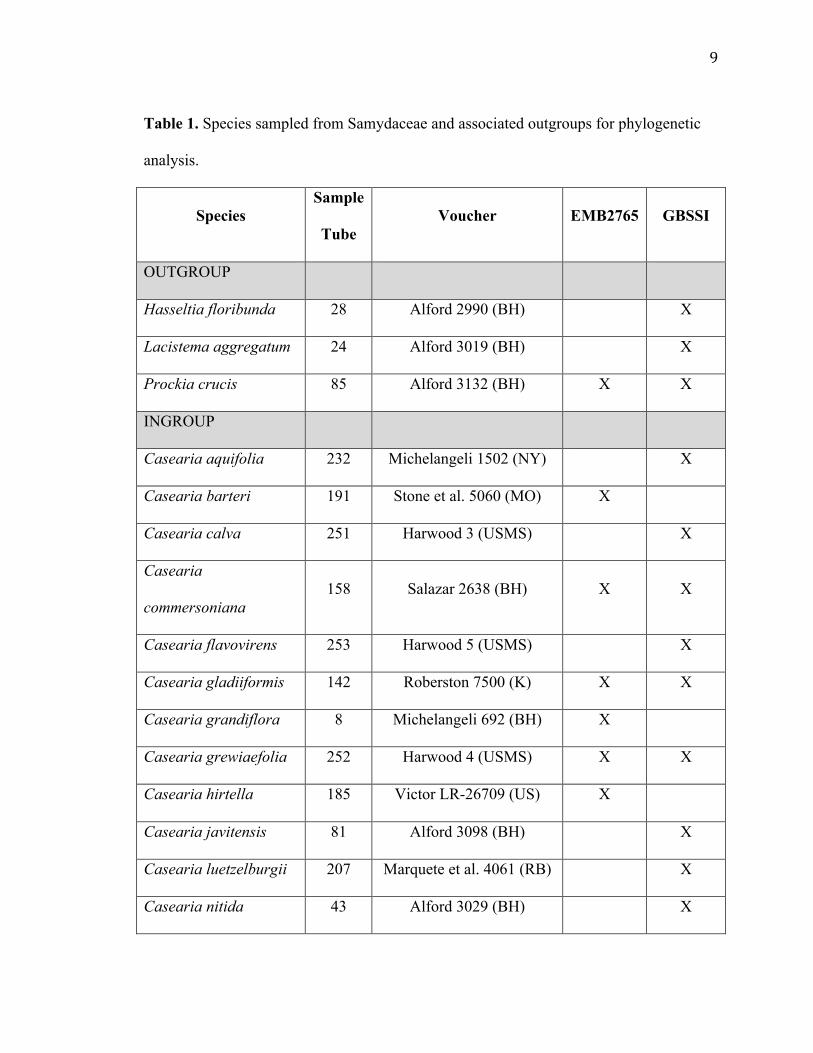
Mini Kit (QIAGEN Sciences, Maryland), and were frozen at -20°C. A list of species
used, including vouchers, can be found in Table 1. Primers chosen for this experiment
included GBSSI forward and reverse primers and EMB2765 forward [9F2] and reverse
[9R] primers. GBSSI is a 714 bp part of the nuclear gene granule bound starch synthase
(called waxy in grasses) which was developed in lab using the genomes of Populus,
Manihot, and Ricinus (Goodstein et al. 2012). The GBSSI forward primer is 5’-
ACTGTRAGCCCTTACTATGC-3’, and the GBSSI reverse primer is 5’-
GTTCCATATCGCATAGCATGC-3’. EMB2765 is a low-copy nuclear gene where
exon 9 was chosen because it was easy to amplify and proved to be most successful for
amplification across a range of Malpighiales and distant outgroups (Wurdack and Davis,
2009). The forward primer, EMB2765ex9F2, was 5’-
TATCCAAATGAGCAGATTATGTGGGA-3’ and the reverse primer, EMB2765ex9R,
was 5’-TTGGTCCAYTGTGCWGCAGAAGGRT-3’, following Wurdack and Davis
(2009).
Thawed samples of DNA were mixed in a 0.2 mL Fisherbrand PCR tube with
attached cap according to the following recipe: 25 µL of Takara ExTaq Premix (Otsu,
Shiga, Japan), 10 µL of PAR enhancer, 8 µL of sterile, deionized distilled water, 2.5 µL
of forward primer, 2.5 µL of reverse primer and 2.0 µL of sample DNA. Once combined,
the 50 µL samples underwent amplification with a PCR regime consisting of 94°C for 3

6
minutes, then 30 cycles of 94°C for 30 seconds, 55°C for 30 seconds, and 72°C for 1
min, then 72°C for 2 minutes, and finally stored at 4°C on a Thermo Electron
Corporation PCR Sprint Thermal Cycler (Milford, Massachusetts). The PAR enhancer
used here is a polymerase chain reaction additive reagent developed in a study by
Samarakoon et al. (unpubl. results). This enhancer is intended to overcome the effects of
PCR inhibitors, and it is composed of trehalose, bovine serum albumin, and polysorbate-
20 (Tween-20) (Samarakoon et al., unpubl. results).
After amplification, DNA samples went through electrophoresis. The gel used
was about 30 mL of 1.0% agarose. 4 µL of the DNA sample was mixed with 1 µL of
loading dye and loaded into the wells. 1.5 µL of Promega BenchTop PCR markers
(Madison, Wisconsin) was mixed with 1 µL of loading dye and loaded into the first well
to create a lane of standard fragments for determining the size of amplified fragments.
The gel was run at 100 V for approximately 20 minutes and then soaked in a Pyrex dish
of distilled water containing 3 µL of ethidium bromide 1% solution for at least 30
minutes. This allowed for staining of any present nucleic acid.
Once the ethidium bromide was allowed to stain the nucleic acids, the gel was
fluoresced using a Fisher Scientific ultraviolet light box (Dubuque, Iowa). DNA samples
fluorescing orange were noted, double-checked for appropriate size, and put aside for
purification and DNA sequencing. These samples were purified using a Qiagen
QIAquick® PCR Purification Kit (QIAGEN Sciences, Maryland) and then packaged for
sequencing by Eurofins MWG Operon in Huntsville, Alabama.
Sequences were provided in .abi format from Eurofins and were loaded in
Sequencher 4.7 – Build 2946 (Ann Arbor, Michigan), where they were proofread.

7
Unusable sequence was discarded, and computer-labeled bases were double-checked;
those mislabeled or not labeled were corrected, and contigs were formed by automatically
assembling the data of the forward and reverse strands. Some samples were not able to
form contigs because of unusable forward or reverse sequences, but the usable forward or
reverse strands were still used.
With the program ClustalX (Thompson, 1997), sequences were aligned using
“complete alignment.” Using these alignments, bases were again checked against the
other DNA samples at particular bases and against the chromatograms in Sequencher.
These sequences were then imported into WinClada (Nixon, 2002) and analyzed using
parsimony to infer the phylogenetic tree and using jackknife analysis for statistical
support of the phylogenetic relationships. Uninformative characters, which are those that
were the same for all samples or only different in one sample, were selected and
deactivated before beginning analyses. Heuristic analyses were run on data from
EMB2765, GBSSI, and combined EMB2765 and GBSSI, in each analysis saving 5000
total trees and initiating 500 replications, with 5 trees saved per replication. The most
parsimonious tree(s) was(were) found and saved. If there was more than one
parsimonious tree, a strict consensus tree was calculated and saved. A jackknife analysis
was then run using 500 replications, each replication consisting of 5 individual
replications, saving 2 trees per replication. The jackknife values were then mapped onto
the most parsimonious tree or strict consensus tree (Figures 2, 3, and 4). Jackknife
analysis involves resampling portions of the available data at random without
replacement and seeing how frequently or infrequently the same results are obtained; the

8
more frequently a result is obtained, the more data there are to support that relationship.
This is the number reported above branches in the phylogenetic tree (Farris et al., 1996).
9
Table 1. Species sampled from Samydaceae and associated outgroups for phylogenetic
analysis.
Species Sample
Tube Voucher EMB2765 GBSSI
OUTGROUP
Hasseltia floribunda 28 Alford 2990 (BH) X
Lacistema aggregatum 24 Alford 3019 (BH) X
Prockia crucis 85 Alford 3132 (BH) X X
INGROUP
Casearia aquifolia 232 Michelangeli 1502 (NY) X
Casearia barteri 191 Stone et al. 5060 (MO) X
Casearia calva 251 Harwood 3 (USMS) X
Casearia
commersoniana 158 Salazar 2638 (BH) X X
Casearia flavovirens 253 Harwood 5 (USMS) X
Casearia gladiiformis 142 Roberston 7500 (K) X X
Casearia grandiflora 8 Michelangeli 692 (BH) X
Casearia grewiaefolia 252 Harwood 4 (USMS) X X
Casearia hirtella 185 Victor LR-26709 (US) X
Casearia javitensis 81 Alford 3098 (BH) X
Casearia luetzelburgii 207 Marquete et al. 4061 (RB) X
Casearia nitida 43 Alford 3029 (BH) X
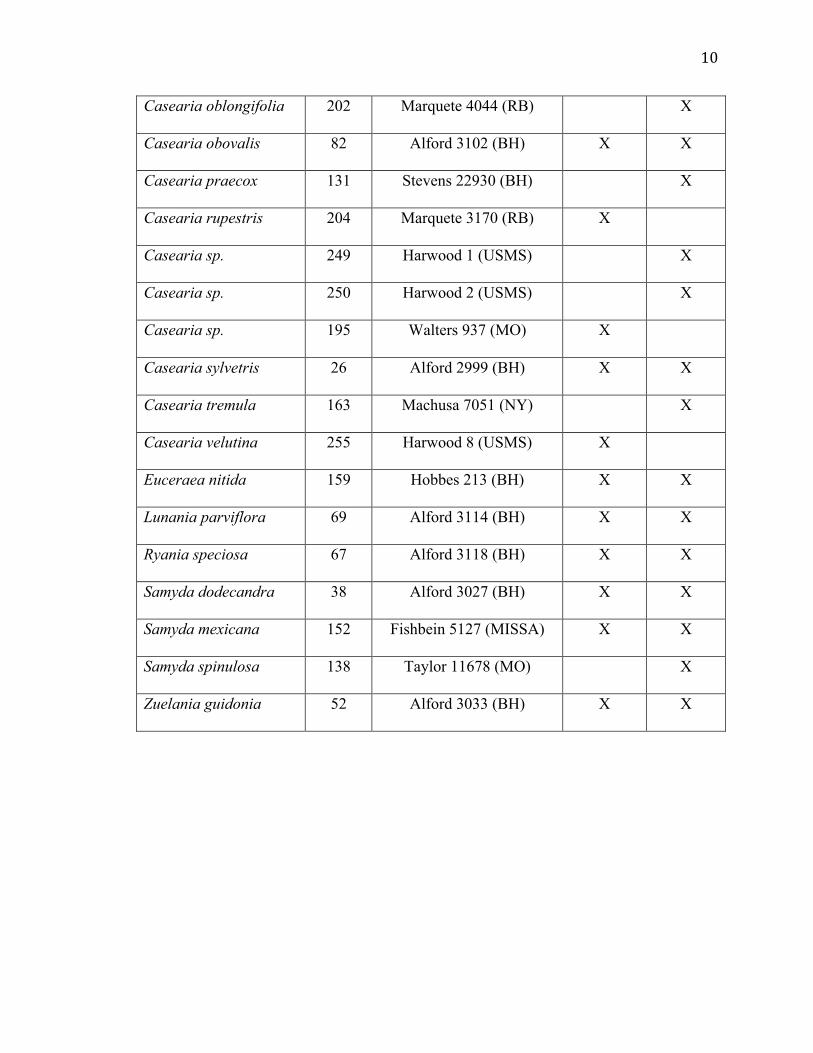
10
Casearia oblongifolia 202 Marquete 4044 (RB) X
Casearia obovalis 82 Alford 3102 (BH) X X
Casearia praecox 131 Stevens 22930 (BH) X
Casearia rupestris 204 Marquete 3170 (RB) X
Casearia sp. 249 Harwood 1 (USMS) X
Casearia sp. 250 Harwood 2 (USMS) X
Casearia sp. 195 Walters 937 (MO) X
Casearia sylvetris 26 Alford 2999 (BH) X X
Casearia tremula 163 Machusa 7051 (NY) X
Casearia velutina 255 Harwood 8 (USMS) X
Euceraea nitida 159 Hobbes 213 (BH) X X
Lunania parviflora 69 Alford 3114 (BH) X X
Ryania speciosa 67 Alford 3118 (BH) X X
Samyda dodecandra 38 Alford 3027 (BH) X X
Samyda mexicana 152 Fishbein 5127 (MISSA) X X
Samyda spinulosa 138 Taylor 11678 (MO) X
Zuelania guidonia 52 Alford 3033 (BH) X X

11
Chapter 3: RESULTS
A total number of 32 species had positive results. Fourteen of these species
produced data for only GBSSI. Six of these species produced data for only EMB2765.
Twelve of these sequences produced results for both GBSSI and EMB2765. Forward and
reverse copies were obtained for each species for every positive result. Not all results
were good sequences, and some characters were determined by forming contigs or
through alignment in ClustalX.
After mopping uninformative characters, 112 characters were potentially
phylogenetically useful (variable) in EMB2765 data, 227 were useful in GBSSI data, and
339 were useful in the merged matrix of both sets of data. See Table 2 for a comparison
of variation with other sampled DNA regions. From the table it is clear that these nuclear
regions exhibit almost as much or more variability than the previously sampled regions.
For EMB2765 data, parsimony analysis resulted in four most parsimonious trees,
with length 217, consistency index 0.62, and retention index 0.68. For the GBSSI data,
there was only one most parsimonious tree with a length of 496, a consistency index of
0.62, and a retention index of 0.71. For the merged matrix of both sets of data, there were
1,082 most parsimonious trees, the length was 715, the consistency index was 0.62, and
the retention index was 0.70.
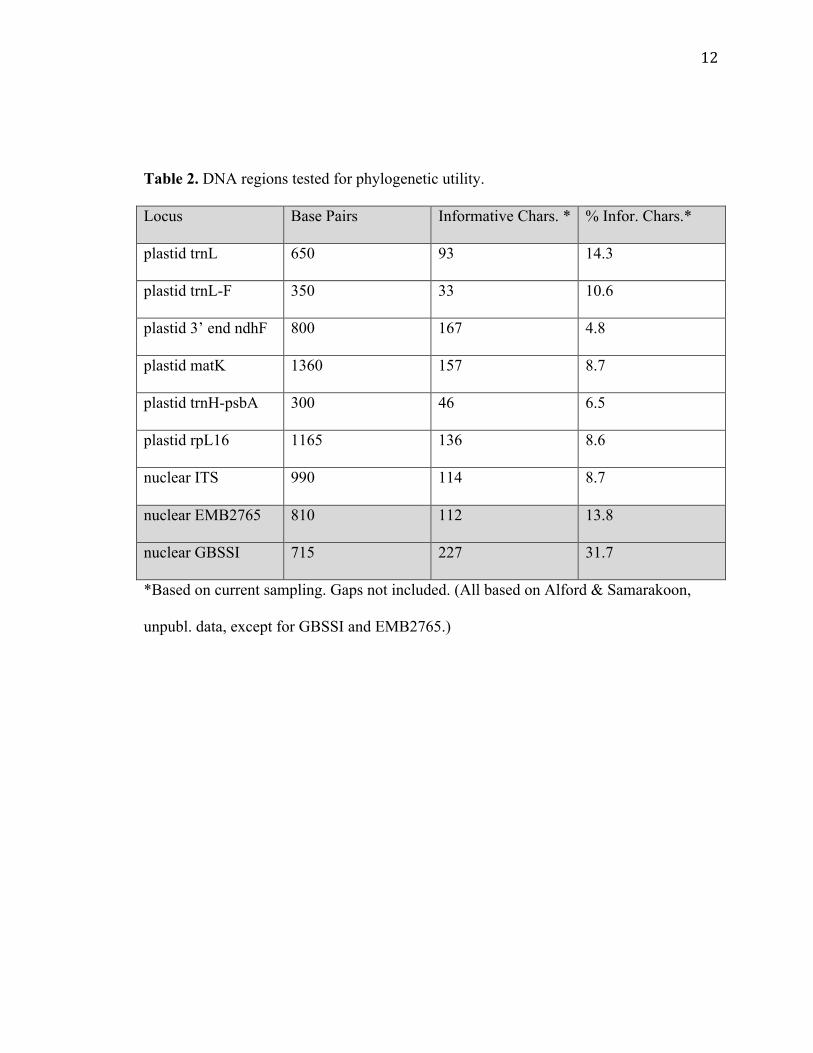
12
Table 2. DNA regions tested for phylogenetic utility.
Locus Base Pairs Informative Chars. * % Infor. Chars.*
plastid trnL 650 93 14.3
plastid trnL-F 350 33 10.6
plastid 3’ end ndhF 800 167 4.8
plastid matK 1360 157 8.7
plastid trnH-psbA 300 46 6.5
plastid rpL16 1165 136 8.6
nuclear ITS 990 114 8.7
nuclear EMB2765 810 112 13.8
nuclear GBSSI 715 227 31.7
*Based on current sampling. Gaps not included. (All based on Alford & Samarakoon,
unpubl. data, except for GBSSI and EMB2765.)

13
Figure 1. Strict consensus tree of Samydaceae based on morphology, 6 plastid, 1 nuclear,
and 1 mitochondrial region (6 MPTs, L=1537, RI=0.79). All taxa except Laetia coriacea
are represented by at least morphology and two DNA regions. Jackknife values ≥60% are
above branches. Based on Alford & Samarakoon, unpubl. data.
100
100 96
98
90
100
100
70
81 94
82
100
75
60
100
100 60
79
73
100
63

14
Figure 2. Phylogeny of Samydaceae based on parsimony analysis of nuclear EMB2765
DNA data. Numbers above the branches are jackknife statistical values.

15
Figure 3. Phylogeny of Samydaceae based on parsimony analysis of nuclear GBSSI
DNA data. Numbers above the branches are jackknife statistical values.

16
Figure 4. Phylogeny of Samydaceae based on parsimony analysis of nuclear GBSSI and
EMB2765 DNA data. Numbers above the branches are jackknife statistical values.

17
Chapter 4: DISCUSSION
While there are some similarities between the trees produced with these nuclear
data (Figures 2-4) and the previous plastid data (Figure 1), there are also differences. Not
every tree contains the same exact species, so it can be a bit difficult to compare simply
by looking at the branches. Compared to the plastid DNA data tree, the trees based on
nuclear data are much better supported statistically. As Figure 2 shows, every branch in
the EMB2765 tree has a jackknife value >58%. Many branches in the GBSSI tree in
Figure 3 also have high values, but four branches are supported by less than 50%. In the
combined EMB2765 and GBSSI nuclear DNA data (Figure 4), the phylogeny is not as
well resolved, due to missing data, but all branches except one in the strict consensus tree
have values >70%.
By looking at the number of variable sites (Table 2) and jackknife values (Figures
2-4), the nuclear DNA data surpass that of the plastid data. More branches in the plastid
DNA data have low jackknife values. The tree based on plastid DNA includes data from
more sampled species. However, the trees agree with each other on some broad points.
The one major difference is the occurrence of Casearia velutina at the base of the
EMB2765 and combined trees. Given that Casearia velutina has all of the features of a
‘standard’ Casearia, this result is likely incorrect or based on a mis-identified sample.
Further study of the voucher specimen or a re-amplification of the DNA should resolve
this issue.
Consistent with the data from Figures 2-4, there are still some genera nested
within Casearia, including Samyda, Euceraea, and Zuelania. Some Casearia (C.

18
commersoniana and C. javitensis) are more closely related to Ryania, as predicted from
morphological data because they have staminodes that occur within the stamens and have
a branched style, just like Ryania. Overall, the data yielded by the EMB2765 and GBSSI
nuclear genes affirms the previous findings using plastid DNA, but with higher statistical
support. Thus, these genes appear to be appropriate regions for the continued study of
relationships within Samydaceae.

19
REFERENCES
Alford, M. H. 2003. Claves para los géneros de Flacourtiaceae de Perú y del Nuevo
Mundo. Arnaldoa 10: 19-38.
Alford, M. H. 2005 Systematic studies in Flacourtiaceae. Ph.D. dissertation, Cornell
University, Ithaca, New York. 290 pp.
Alford M. H. 2012 in prep. Phylogeny, classification, and character evolution of
Salicaceae and the non-cyanogenic Flacourtiaceae. American Journal of Botany.
Anaya, A. L., M. Jiménez, R. Cruz-Ortega, M. L. Macías-Rubalclava, and B. E.
Hernández-Bautista. 2004. Fungicide effects of allelochemicals produced by the leaves of
the tropical tree Zuelania guidonia (Flacourtiaceae). Abstract 18. Second European
Allelopathy Symposium: “Allelopathy—from understanding to application.”
Beutler, J. A., K. L. McCall, K. Herbert, T. Johnson, R. H. Shoemaker, and M. R. Boyd.
2000. Cytotoxic clerodane diterpene esters from Laetia corymbulosa. Phytochemistry 55:
233-236.
Chai, Xing-Yun, Fei-Fei Li, Chang-Cai Bai, Zheng-Ren Xu, Hai-Ming Shi, and Peng-Fei
Tu. 2010. Three New Acylated Glycosides from the Stems of Casearia velutina and their

20
Protective Effect Against H2O2-Induced Impairment in PC12 Cells. Planta Med 76(1):
91-93.
Chase, M. W., S. Zmarzty, M. D. Lledó, K. J. Wurdack, S.M. Swensen, and M. F. Fay.
2002. When in doubt, put it in Flacourtiaceae: a molecular phylogenetic analysis based
on plastid rbcL DNA sequences. Kew Bulletin 57: 141-148.
Chudnoff, M. 1984. Tropical Timbers of the World. USDA Forest Service Agricultural
Handbook No. 607.
Farris, James S., Victor A. Albert, Mari Källersjö, Diana Lipscomb, and Arnold G.
Kluge. 1996. Parsimony Jackknifing Outperforms Neighbor-joining. Cladistics 12:99-
124.
Ferreira, Paulo Michael P., Leticia V. Costa-Lotufo, Manoel O. Moraes, Francisco W. A.
Barros, Aline M. A. Martins, Alberto J. Cavalheiro, Vanderlan S. Bolzani, Andre G.
Santos, and Claudia Pesso. 2011. Folks uses and pharmacological properties of Casearia
sylvestris: a medicinal review. Anais da Academia Brasileira de Ciências 83(4):
Gentry, A. H. 1993. A Field Guide to the Families and Genera of Woody Plants of
Northwest South America (Columbia, Ecuador, Peru). University of Chicago Press,
Chicago.

21
Gilg, E. 1925. Flacourtiaceae, Pages 377-457 in Vol. 21, Die natürlichen
Pflanzenfamilien, 2nd ed. (A. Engler, and K. Prantl. Eds.), Berlin.
Goodstein, D. M., S. Shu, R. Howson, R. Neupane, R. D. Hayes, J. Fazo, T. Mitros, W.
Dirks, U. Hellsten, N. Putnam, and D. S. Rokhsar. 2012. Phytozome: a comparative
platform for green plant genomics, Nucleic Acids Res. 40(D1): D1178-D1186.
www.phytozome.net.
Guilherme, F. G., J. N. Nakajima, C. A. Prado-Lima, and A. Vanini. 1998. As
fitofisionomias e a flora lenhosa nativas do Parque do Sabiá, Uberlândia-MG. Daphne
8(2): 17-30.
Hutchinson, R. 1967. The Genera of Flowering Plants, Clarendon Press, Oxford.
Itokawa, H., N Totsuka, H. Morita, K Takeya, Y. Iitaka, E. P. Schenkel, and M.
Motidome. 1990. New antitumor principles, casearins A-F, for Casearia sylvestris Sw.
(Flacourtiaceae). Chemical and Pharmaceutical Bulletin 38: 3384-3388.
Jaffré, T., W. Kersten, R. R. Brooks, and R. D. Reeves, 1979. Nickel uptake by
Flacourtiaceae of New Caledonia. Proceedings of the Royal Society of London, Series B,
Biological Sciences 205: 385-394.

22
Jullian, V., C. Bonduelle, A. Valentin, L. Aceby, A.-G. Duigou, M.-F. Prevost, and M.
Sauvain. 2005. New clerodane diterpenoids from Laetia procera (Poepp.) Eichler
(Flacourtiaceae), with antiplasmodial and antileishmanial activities. Bioorganic and
Medicinal Chemistry Letters 15: 5065-5070.
Judd, W. S., C. S. Campbell, E. A. Kellogg, P. F. Stevens, and M. J. Donoghue. 2007.
Plant Systematics: A Phylogenetic Approach. Sinauer Press, Sunderland, Massachusetts.
Lemke, D. E. 1988. A synopsis of Flacourtiaceae. Aliso 12: 29-43.
Nixon, K.C. 2002 WinClada, vers. 1.00.08. Ithaca, New York. Available from author at
www.cladistics.com .
Phillips, O. and J. S. Miller. 2002. Global Patterns of Plant Diversity: Alwyn H. Gentry’s
Forest Transect Data Set. St. Louis: Missouri Botanical Garden Press.
Richter, H. G. and M. J. Dallwitz, 2000 onwards. Commerical timbers: descriptions,
illustrations, identification, and information retrieval. Version: 4 May 2000.
http://biodiversity.uno.edu/delta/ .
Silva, Jr., M. C., J. M. Felfili, P. E. Nogueira, and A. V. Rezende. 1998. Análise florística
das matas de galleria do Distrito Federal. Pp. 53-84 IN: Cerrado: Matas de Galeria, J. F.
Ribeiro, ed. Planaltina: EMBRAPA.

23
Takhtajan, A. 1985. Three new families of flowering plants. Bot. Zhurn. (Moscow and
Leningrad) 70: 1691 - 1693.
Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G. Higgins. 1997.
The ClustalX windows interface: flexible strategies for multiple sequences
alignment aided by quality analysis tools. Nucleic Acids Research 25: 4876-4882.
Warburg, O. 1894. Flacourtiaceae. Pages 1-56 in Vol 3(6a), Die natürlichen
Pflanzenfamilien, 1st ed. (A. Engler, and K. Prantl. Eds.). Wilhelm Englemann, Leipzig.
Williams, L. 0. 1965. The story of two sterile specimens. Annals of the Missouri
Botanical Garden 52: 485-486.
Wurdack, K. J. and C. C. Davis. 2009. Malpighiales phylogenetics: gaining ground on
one of the most recalcitrant clades in the Angiosperm tree of life. American Journal of
Botany 96(8): 1551-1570.




![Classical Taxonomy & Molecular Phylogeny [17.1-17.3] SPI 5 Apply evidence from the fossil record, comparative anatomy, amino acid sequences, and DNA structure](https://img.pdfslide.net/doc/110x75/56649ec65503460f94bd1738/classical-taxonomy-molecular-phylogeny-171-173-spi-5-apply-evidence-from.jpg)










![Molecular Phylogeny Inferred from Sequences of Small Title ...repository.kulib.kyoto-u.ac.jp/dspace/bitstream/2433/...and Cantor [16]. The phylogenetic tree was con structed from an](https://img.pdfslide.net/doc/110x75/60ea7e51d31cb125f52ec7c3/molecular-phylogeny-inferred-from-sequences-of-small-title-and-cantor-16.jpg)













