Embed Size (px)
Citation preview

Molecular and Cellular Biochemistry 274: 125–131, 2005. c�Springer 2005
A role for CK2α/β in Xenopus early embryonicdevelopment
Isabel Dominguez, Junko Mizuno, Hao Wu, Gregory A. Imbrie,Karen Symes and David C. SeldinHematology-Oncology Section, Department of Medicine, Boston University Medical School, Boston, MA, USA
Abstract
CK2 is expressed widely in early embryonic development in several animal models, however its developmental role is unclear.One of the substrates of CK2 that is important in embryonic development is β-catenin, the transcriptional co-activator of thecanonical Wnt signaling pathway. This pathway has been implicated in diverse aspects of embryonic development, includingone of the earliest events in embryonic development, the establishment of the dorso-ventral embryonic axis. In Xenopus laevis,dorso-ventral axis formation is dependent upon stabilization of β-catenin in the future dorsal side of the embryo. Since CK2phosphorylation of β-catenin stabilizes it, we hypothesized that CK2 might be critical to upregulation of β-catenin in Xenopusembryos and to the process of axis establishment. Our results demonstrate that CK2 is required for dorsal axis formation andis for normal upregulation of Wnt signaling genes and targets. Thus, CK2 is a regulator of endogenous axis formation invertebrates. (Mol Cell Biochem 274: 125–131, 2005)
Key words: β-catenin, dorsal axis formation, protein kinase CK2, Wnt signaling, Xenopus laevis
Introduction
The establishment of the embryonic axes is an essential pro-cess that occurs after fertilization and dictates where embry-onic tissues, organs and systems will be positioned [1]. Themolecular mechanism of generation of the embryonic axesis best studied in frog embryos. Xenopus eggs have an obvi-ous morphological animal-vegetal polarity: the animal poleis pigmented, while the vegetal pole is not, but otherwisethe egg is radially symmetrical (Fig. 1). This polarity is gen-erated during oogenesis, when cellular components includ-ing pigment, proteins, RNAs, and organelles are asymmet-rically located along the animal-vegetal axis. Among themare the “dorsal determinants”, substances of unknown na-ture that are localized in the vegetal cortex and are essentialfor dorsal axis determination. Fertilization occurs in the an-imal pole and the site of sperm entry, marking the futureventral side of the embryo, is recognizable as a darker pig-
Address for offprints: I. Dominguez, EBRC Building, Room X-420, 650 Albany Street, Boston MA 02118 (E-mail: [email protected])
mented spot. After fertilization, an essential process calledcortical rotation occurs, and the end result is the movementof the vegetally localized dorsal determinants to the prospec-tive dorsal side of the embryo, opposite to the sperm en-try point (reviewed in [2], Fig. 1). This breaks the radialsymmetry inherent to the egg and specifies the dorso-ventralaxis.
Although the nature of the vegetal determinants is still un-known, their mechanism of action seems to involve the activa-tion of canonical Wnt signaling. Indeed, dorsal enrichment ofβ-catenin is the first molecular asymmetry essential for dorsalaxis formation found after cortical rotation in Xenopus em-bryos (reviewed in [2]). However, β-catenin does not appearto be the dorsal determinant or move with cortical rotation;thus, it is reasonable to consider that β-catenin is upregulatedthrough canonical Wnt signaling [2, 3]. Evidence suggeststhat Wnt ligands themselves do not participate in dorsal axisformation, even though their overexpression in the ventral
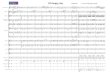
126
Fig. 1. Early dorsal development in Xenopus embryos. Dorsal determinants are localized to the vegetal cortex of the oocyte during oogenesis (left). Afterfertilization of the egg, the dorsal determinants move to the future dorsal side of the embryo through a process that involves cortical rotation and active transport,both mediated by microtubule arrays (middle). The dorsal determinants initiate the dorsal program, putatively by downregulation of GSK3β by GBP and bydsh inhibition of the β-catenin destruction complex. This leads to upregulation of β-catenin in the dorsal cytoplasm as early as the two-cell stage (right).Upregulation of β-catenin in the dorsal side is the earliest detected molecular difference between dorsal and ventral sides of the embryo, and contributes to thedorsal developmental program.
side of the embryo is sufficient to induce an ectopic axis [4].The current model suggests that intracellular mediators in theWnt pathway upregulate β-catenin dorsally (Fig. 2). Indeed,correlating with β-catenin enrichment, shortly after corticalrotation, GSK3β is depleted in the dorsal side of the em-
Fig. 2. Wnt signaling pathway in dorsal axis formation in Xenopus embryos. The canonical Wnt pathway controls intracellular levels of the transcriptionalco-activator β-catenin, but in embryos, this appears to occur independently of ligand-receptor-mediated activation of the pathway (see Wnt/LRP/Frizzledreceptor complex in dashed oval). Dorsal enrichment of β-catenin in Xenopus embryos starts at stage 2 and continues for several hours, leading to dorsalupregulation of β-catenin/TCF target genes such as Xnr5 and Xnr6 at stage 8, and Xsiamois, Xtwin and Xnr3 after the mid-blastula transition (MBT) whenglobal zygotic transcription begins, at stage 8.5 [4, 48].
bryo while dishevelled is found enriched [5, 6]. However themolecular mechanism that leads to β-catenin upregulation inthe early embryo is still uncharacterized. In a search for in-tracellular proteins that could upregulate β-catenin dorsallyafter cortical rotation, we considered protein kinase CK2 as a

127
candidate, based upon its ability to associate with β-cateninand dsh and regulate their stability [7, 8].
CK2 can function as a tetrameric holoenzyme composedof two catalytic subunits (α) and two regulatory subunits (β).The array of subunits and the number of genes that codefor catalytic and regulatory subunits varies greatly betweenspecies [9–11]. Mammals have two alternative catalytic sub-units, α and α′, and a single β regulatory subunit, whileXenopus appears to have two genes encoding α isoforms,no shorter α′ isoform, and one β regulatory subunit [12, 13].There is a role for CK2 during meiosis in mammals and inXenopus, where the CK2β subunit appears to be inhibitingMos activity and the CK2α subunit appears to regulate cdc25activity [14, 15]. Targeting of the single β subunit in mice [16]and inactivation of all subunits in yeast are lethal [17, 18],suggesting that the CK2 holoenzyme is essential for cell sur-vival. Targeting a single catalytic subunit in mice has a moresubtle phenotype; male CK2α′ null mice are infertile dueto abnormal spermatogenesis [19]. However little is knownabout how CK2 regulates early development. The data pre-sented here identify a role for CK2 in dorso-ventral axis de-termination and Wnt signaling.
Materials and methods
In vitro mRNA synthesis, embryo injection andmicromanipulation
Cloning of CK2α and CK2β by RT-PCR using primers tothe published Xenopus sequences [12], and generating thekinase inactive mutant (KI-CK2α) have been described [20].Plasmids used in these studies were linearized and mRNAwas transcribed in vitro as described [20].
Embryos were fertilized in vitro, dejellied in 2% cysteine(pH 7.8) and maintained in 0.1× MMR [21]. Staging wasaccording to Nieuwkoop and Faber [22]. Normal embryoswere micro-dissected with an eyelash knife at various stagesfor protein analysis and microinjected embryos, as detailed[20], were stopped at the stages indicated in the text as de-scribed below. Embryo development was visualized under aLeica MZ6 dissecting microscope and photographed at neu-rula stages. Axis duplication and axis defects were analyzedat tadpole stages.
RT-PCR
RT-PCR was performed as described [23]. Briefly, RNA wasisolated with Trizol according to the manufacturer’s instruc-tions (Invitrogen). Reactions with and without RT were car-ried out. The PCR reactions included 32P-γ -dATP (0.15 µCi)and the products were separated in 5% acrylamide gels and
visualized and quantitated in a Molecular Imager System GS-525 (BioRad). Primers, cycles and annealing temperaturesused for PCR have been reported [20].
Protein analysis in embryos
For endogenous and exogenous protein expression the equiv-alent of 0.15 embryos was loaded directly on gels. ForPKCζ immunoblotting cleared lysates were used since itsmolecular weight is close to yolk. Electroblotting and im-munoblotting are described in detail in [20]. For quantitativeanalysis of each band, integrated pixel density minus back-ground density was determined using Fluor-S MultiImager(BioRad).
Results and discussion
Expression pattern of CK2α and CK2β in early Xenopusembryos: Localization to the upper half of embryos
Our first goal was to determine when and where CK2 subunitsare found during early dorsal development. XCK2α mRNAhas been reported to be present in Xenopus oocytes, blas-tula stage embryos, tadpoles, and several adult tissues, andXCK2β RNA is present at least in the oocyte [13, 24]. To de-termine the spatial distribution of CK2α and β during earlyembryonic development, we utilized in situ hybridization andimmunoblotting.
The expression of CK2α and CK2β proteins was examinedby immunoblotting with specific antibodies against CK2α
and CK2β. Xenopus embryos contain CK2α and CK2β
bands of appropriate 45 kDa and 25 kDa size respectively,while control rat brain extract also has a 41 kDa α′ band(Fig. 3A). Indeed, no gene for Xenopus CK2α′ has beenfound to date [13]. Interestingly, the ratio between CK2α
and CK2β was different in the Xenopus extract comparedwith the rat brain control, suggesting that the CK2β subunitcould be more limiting in early Xenopus embryos than inother tissues.
At early embryonic stages, CK2α and CK2β transcriptswere detected in the upper part of the embryo [20]. To corre-late the mRNA distribution with the distribution of the CK2subunits within the embryo, embryos at the 32-cell stagewere cut into animal (A) and vegetal (V) halves. Immunoblotanalysis (Fig. 3B) showed that both CK2α and β were pref-erentially distributed in the animal half (approximate ratioA/V = 4) while this difference was not that large for otherproteins studied, like tubulin and PKCζ (approximate ratioA/V = 2). This pattern did not change through maternal de-velopment (not shown). The ratio A/V of CK2α and β wasstudied with direct lysis in Laemmli buffer or cleared lysates.
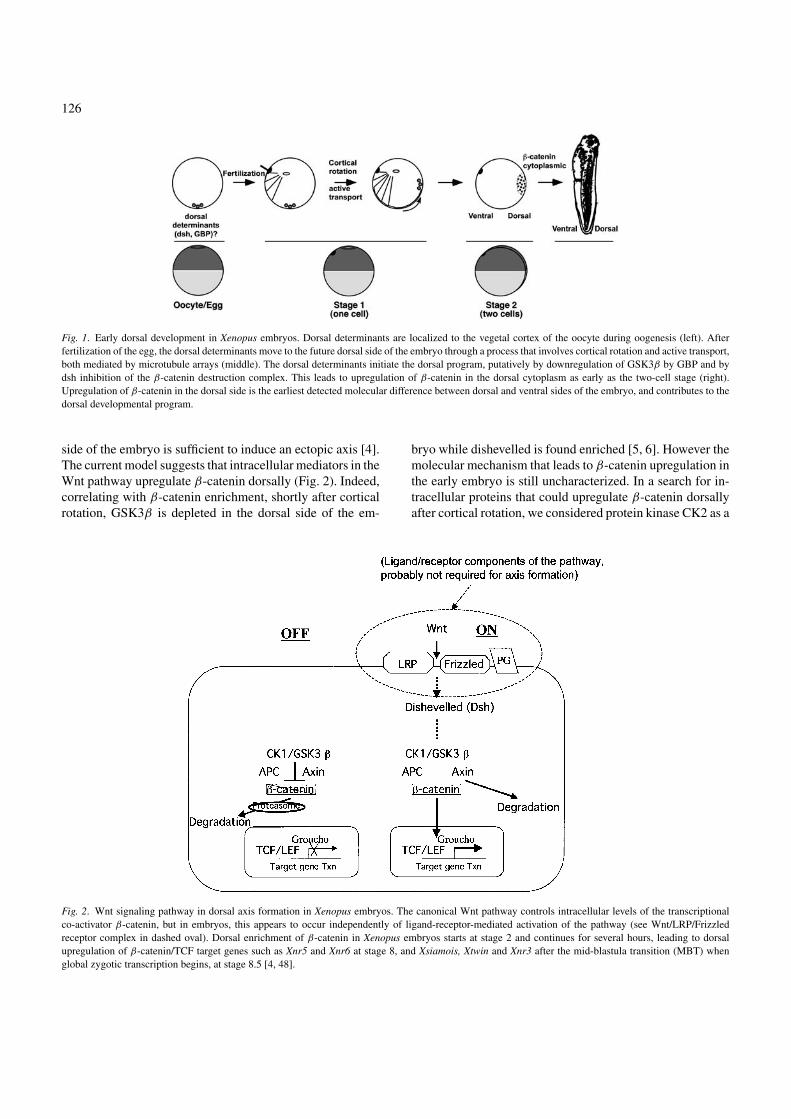
128
Fig. 3. CK2α and β protein expression in early Xenopus development. (A)CK2α and CK2β but not CK2α′ are detected in early Xenopus embryos. Con-trol rat brain lysate (3 µl) supplied as a positive control for the CK2β antibody(BD) and 0.05, 0.1 and 0.15 Xenopus embryo equivalents were loaded. Anti-human CK2α recognizes a 45 kDa CK2α band in stage 6 Xenopus embryolysates, but both CK2α at 45 kDa and CK2α′ at 41 kDa in a control rat brainlysate (upper panel). Anti-human CK2β (lower panel) recognizes a 25 kDaband in both stage 6 embryo and rat brain lysates. The CK2α antibody rec-ognizes an uncharacterized extra band at 38 kDa in the rat brain lysates (•)and a non-specific band (�) in Xenopus extracts. (B) CK2α and CK2β pro-teins are more abundant in the upper part of the embryo. Immunoblottingwith CK2α, CK2β, PKCζ and tubulin antibodies of extracts from embryosat 32-cell stage divided into halves as depicted in the scheme below: A (an-imal) and V (vegetal). CK2α and CK2β are markedly depleted from thevegetal half compared to other enzymes such as PKCζ . Tubulin is shown asa loading control. One representative immunoblot out of eight is shown.
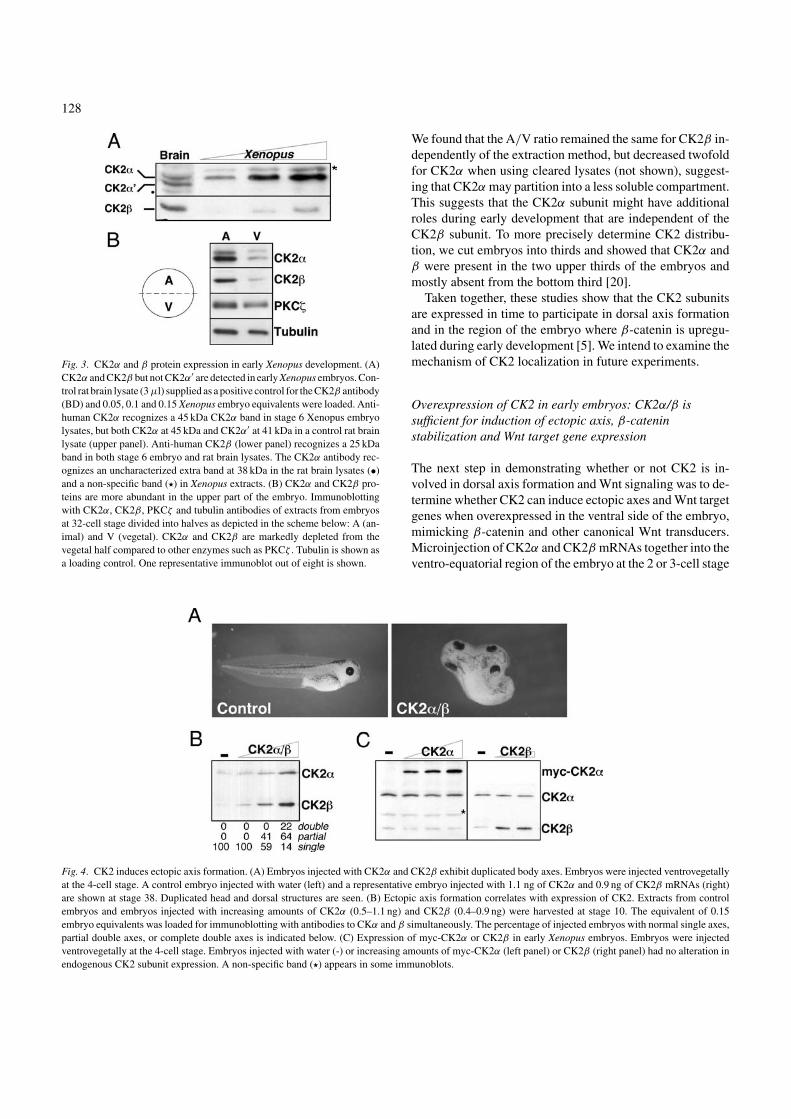
Fig. 4. CK2 induces ectopic axis formation. (A) Embryos injected with CK2α and CK2β exhibit duplicated body axes. Embryos were injected ventrovegetallyat the 4-cell stage. A control embryo injected with water (left) and a representative embryo injected with 1.1 ng of CK2α and 0.9 ng of CK2β mRNAs (right)are shown at stage 38. Duplicated head and dorsal structures are seen. (B) Ectopic axis formation correlates with expression of CK2. Extracts from controlembryos and embryos injected with increasing amounts of CK2α (0.5–1.1 ng) and CK2β (0.4–0.9 ng) were harvested at stage 10. The equivalent of 0.15embryo equivalents was loaded for immunoblotting with antibodies to CKα and β simultaneously. The percentage of injected embryos with normal single axes,partial double axes, or complete double axes is indicated below. (C) Expression of myc-CK2α or CK2β in early Xenopus embryos. Embryos were injectedventrovegetally at the 4-cell stage. Embryos injected with water (-) or increasing amounts of myc-CK2α (left panel) or CK2β (right panel) had no alteration inendogenous CK2 subunit expression. A non-specific band (�) appears in some immunoblots.
We found that the A/V ratio remained the same for CK2β in-dependently of the extraction method, but decreased twofoldfor CK2α when using cleared lysates (not shown), suggest-ing that CK2α may partition into a less soluble compartment.This suggests that the CK2α subunit might have additionalroles during early development that are independent of theCK2β subunit. To more precisely determine CK2 distribu-tion, we cut embryos into thirds and showed that CK2α andβ were present in the two upper thirds of the embryos andmostly absent from the bottom third [20].
Taken together, these studies show that the CK2 subunitsare expressed in time to participate in dorsal axis formationand in the region of the embryo where β-catenin is upregu-lated during early development [5]. We intend to examine themechanism of CK2 localization in future experiments.
Overexpression of CK2 in early embryos: CK2α/β issufficient for induction of ectopic axis, β-cateninstabilization and Wnt target gene expression
The next step in demonstrating whether or not CK2 is in-volved in dorsal axis formation and Wnt signaling was to de-termine whether CK2 can induce ectopic axes and Wnt targetgenes when overexpressed in the ventral side of the embryo,mimicking β-catenin and other canonical Wnt transducers.Microinjection of CK2α and CK2β mRNAs together into theventro-equatorial region of the embryo at the 2 or 3-cell stage

129
induced ectopic axis formation in a dose-response manner(Fig. 4A and B). This effect is comparable to those obtainedwith ectopic expression of transducers of canonical Wnt sig-naling including Xwnt8 [25, 26], dsh [27], KI-GSK3β [28–30] and β-catenin [31]. Morphologic ectopic axis inductionwas accompanied by induction of the embryonic Wnt targetgenes Xsiamois, Xnr3 and Xtwin (see data in [20]). Interest-ingly, microinjection of either CK2 subunit alone failed toproduce a secondary axis [20], suggesting that the CK2 tar-get(s) are phosphorylated only by the pool of CK2 that isin the tetrameric holoenzyme form. Interestingly, althoughthe β subunit has been proposed to stabilize the α subunit,when we overexpress β in the embryo we see no change inα protein levels, and vice versa (Fig. 4C). Thus, the subunitsdo not appear to regulate each other during early Xenopusdevelopment.
These experiments showed that CK2 is sufficient to pro-duce ectopic axes and to induce Wnt target genes (Xsiamois,Xnr3 and Xtwin). Both these outcomes are only observedwhen canonical Wnt signaling is activated, suggesting thatCK2 is a positive regulator of canonical Wnt signaling [4, 32,33]. Interestingly, many molecules that activate Wnt signal-ing can play a role in human tumors [34–37], and this is alsotrue for CK2 [38, 39]. We hypothesized that CK2 might beacting during early Xenopus development through β-cateninor dsh (engrailed is involved much later in development). Infact, our data has demonstrated that CK2 is able to stabilizeβ-catenin in vivo [20], and both β-catenin and dishevelled inmammalian cells [7, 40]. There is also a possibility that CK2could be acting through an as yet uncharacterized substrate.
Antagonism of CK2 in early embryos: CK2 is necessary fornormal Wnt signaling and endogenous axis formation
To conclusively establish that CK2 is a bona fide Wnt trans-ducer and a mediator of early dorsal axis formation by demon-strating that CK2 is required for both these tasks. For this wegenerated a kinase inactive (KI) mutant of the catalytic sub-unit, CK2α, by mutating the ATP-binding lysine (Lys68Arg)critical for catalytic activity and that has been previouslyshown to behave as a dominant negative mutant [41]. Thismutant lacked kinase activity in vitro [20].
To determine whether the kinase-inactive mutant couldblock the ability of injected Wnt to induce an ectopic axis,its mRNA was co-injected together with that of Xwnt8.While Xwnt8 mRNA was able to induce double axes in ap-proximately 50% of the embryos (Fig. 5A), co-injection ofKI-CK2α mRNA and Xwnt8 mRNA led to a marked re-duction in the number of embryos with a duplicated axis(Fig. 5A). No phenotype was observed when mRNA forKI-CK2α mRNA was introduced into the ventral side of theembryo or when KI-CK2α and CK2β mRNA were injected
Fig. 5. KI-CK2α inhibits ectopic dorsal axis formation promoted by Xwnt8and normal dorsal development. (A) KI-CK2α is able to suppress ectopic axisformation by Xwnt8. Two ventrovegetal blastomeres at the two or three cellstage were injected with lacZ mRNA, 0.34 pg of Xwnt8 mRNA, or 0.34 pgof Xwnt8 mRNA together with 1.6 ng of KI-CK2α mRNA. Representativemorphology of the injected embryos is presented at stage 19. At this stage,embryos that are developing ectopic body axes display duplicated neuraltubes (arrowheads), while normal embryos display only one neural tube. (B)Inhibition of the endogenous dorsal axis by KI-CK2α. Two ventrovegetalblastomeres at stage 2 to 3 were injected with 2 ng lacZ mRNA (upperembryo) or 2 ng of KI-CK2α mRNA (other embryos). Morphology of theinjected embryos is presented at stage 38. Inhibition of the endogenous dorsalaxis by KI-CK2α mRNA expression is demonstrated by the absence of themost anterior structures (eyes and cement gland) and marked reduction inthe trunk. This panel was reprinted from Developmental Biology 274(1), I.Dominguez, J. Mizuno, H. Wu, D. Song, K. Symes and D.C.Seldin, Proteinkinase CK2 is required for dorsal axis formation in Xenopus embryos, pp:110–124 (2004), with permission from Elsevier.
ventrally together, demonstrating that the kinase activity ofthe enzyme is required for generation of an ectopic dorsalaxis (not shown). Immunoblotting to verify the expressionof KI-CK2α was performed in all experiments. These resultsdemonstrate that catalytically active CK2 is not only capa-ble of inducing an ectopic axis, but its activity is required forcanonical Wnt signaling. This mutant had no effect on ectopicaxis formation promoted by T393A β-catenin (a β-cateninmutant in the major CK2 phosphorylation site) [20]. This sug-gests that CK2 modulates Wnt signaling through β-cateninphosphorylation or an upstream event in Xenopus embryos.
We next determined the effect of overexpression of the KI-CK2 mutant on endogenous axis formation by injecting thismutant dorsally very early in development. These embryos

130
Fig. 6. Model of CK2 mechanism of action during dorsoventral axis formation. During oogenesis, the dorsal determinants are localized to the vegetal pole of theegg. After fertilization (left), cortical rotation will bring the dorsal determinants to the future dorsal side of the embryo (middle). Here, the dorsal determinantsare placed in a region of the embryo where CK2 is highly expressed and constitutively active. There, downregulation of GSK3β by GBP, inhibition of thedestruction complex by dsh, and phosphorylation of the dorsal determinants or their protein targets in the Wnt pathway by CK2 initiate the dorsal developmentalprogram (right). These targets might include β-catenin, dsh, GBP, or other signaling components. Shading represents the region of the embryo where CK2 islocated.
showed shortened axes, frequently lacking all of the mostanterior structures (Fig. 5B), while dorsal injection of wa-ter or mRNA encoding lacZ mRNA had no effect on en-dogenous axis formation. Immunoblotting to confirm the ex-pression of KI-CK2α was performed in all experiments (notshown). In vitro kinase experiments correlated ventraliza-tion with decreased total CK2 activity in immunoprecipitates[20]. The effects seen in our experiments are similar to thoseof downregulation of GBP or β-catenin function, and muchstronger than the phenotypes obtained by dorsal inhibition ofsome other Wnt transducers [28, 42–46]. This suggests thatthe KI-CK2α mutant had a dominant negative effect overendogenous CK2. Moreover, CK2β co-immunoprecipitateswith both endogenous CK2α and KI-CK2α indicating thatthe KI mutant is able to form tetrameric complexes [20]. Asthe tetrameric form of CK2 seems to be the one involved inWnt signaling, this suggests that sequestration of the CK2β
subunit by the KI-CK2α into tetrameric inactive complexesis the mechanism of action of this mutant. These experimentssuggest that CK2 activity is necessary for endogenous dor-sal axis formation in Xenopus embryos. Epistatic-like ex-periments will address whether β-catenin is the sole targetof CK2 during early embryonic development or if there areother targets like dsh through which CK2 is also acting.
In conclusion, we have shown that CK2 plays an essen-tial role in Wnt signaling and in dorsal axis development. Asshown in Fig. 1, the current model of dorsal axis formationproposes that when the dorsal determinants move during cor-tical rotation, they signal for upregulation of β-catenin. Ourfindings suggest that CK2 plays a role in this process (Fig. 6).In this proposed model, CK2 is absent from the vegetal part ofthe embryo. When the dorsal determinants move to the medialpart of the embryo, they enter the zone of highest expressionof CK2α and β subunits, and there they can be phosphory-lated by CK2. This model is consistent with embryologicalstudies that showed that suggested dorsal determination oc-curs when the dorsal determinants are in contact with the
cytoplasm of the equatorial (medial) part of the embryo [47].Future experiments will continue to address how CK2 is in-volved in dorsal axis formation in Xenopus, through a moredetailed analysis of dorso-ventral as well as animal-vegetalasymmetry in CK2 and by identification of CK2-containingcomplexes and substrates in different regions of the embryoat different stages of development.
Acknowledgments
We would like to thank the members of Dr. K. Symes labora-tory for their help with the Xenopus experiments. We thankmembers of the Seldin laboratory for helpful discussions andcritical reading of the manuscript. We thank Dr. Sergei Sokolfor providing the Xwnt8 and pJ3M plasmids used in thisstudy. We would like to thank the organizers of the Inter-national CK2 meeting for selecting this work to be presentedin a truly exciting meeting. Supported by ACS grant IRG-72-001-28 (I.D.), a pilot research grant from the Departmentof Medicine at Boston University School of Medicine (I.D.),NIH R01 CA87375 (K.S.), NIH R01 CA71796 (D.S.), andP01 ES11624 (D.S.). D.S. is a Scholar of the Leukemia andLymphoma Society.
References
1. Prodon F, Pruliere G, Chenevert J, Sardet C: Establishment and expres-sion of embryonic axes: comparisons between different model organ-isms. Med Sci (Paris) 20: 526–538, 2004
2. Weaver C, Kimelman D: Move it or lose it: axis specification in Xenopus.Development 131: 3491–3499, 2004
3. Rowning BA, Wells J, Wu M, Gerhart JC, Moon RT, Larabell CA:Microtubule-mediated transport of organelles and localization of beta-catenin to the future dorsal side of Xenopus eggs. Proc Natl Acad SciUSA 94: 1224–1229, 1997
4. Sokol SY: Wnt signaling and dorso-ventral axis specification in verte-brates. Curr Opin Genet Dev 9: 405–410, 1999

131
5. Larabell CA, Torres M, Rowning BA, Yost C, Miller JR, Wu M, Kimel-man D, Moon RT: Establishment of the dorso-ventral axis in Xenopusembryos is presaged by early asymmetries in beta-catenin that are mod-ulated by the Wnt signaling pathway. J Cell Biol 136: 1123–1136, 1997
6. Dominguez I, Green JBA: Dorsal downregulation of GSK3(beta) by anon-Wnt-like mechanism is an early molecular consequence of corticalrotation in early Xenopus embryos. Development 127: 861–868, 2000
7. Song DH, Sussman DJ, Seldin DC: Endogenous protein kinase CK2participates in Wnt Signaling in mammary epithelial cells. J Biol Chem275: 23790–23797, 2000
8. Willert K, Brink M, Wodarz A, Varmus H, Nusse R: Casein kinase2 associates with and phosphorylates dishevelled. EMBO J 16: 3089–3096, 1997
9. Faust M, Montenarh M: Subcellular localization of protein kinase CK2.A key to its function? Cell Tissue Res 301: 329–340, 2000
10. Schneider HR, Reichert GH, Issinger OG: Enhanced casein kinase IIactivity during mouse embryogenesis. Identification of a 110-kDa phos-phoprotein as the major phosphorylation product in mouse embryosand Krebs II mouse ascites tumor cells. Eur J Biochem 161: 733–738,1986
11. Ahmed K, Davis AT, Wang H, Faust RA, Yu S, Tawfic S: Significanceof protein kinase CK2 nuclear signaling in neoplasia. J Cell Biochem79: 130–135, 2000
12. Jedlicki A, Hinrichs MV, Allende CC, Allende JE: The cDNAs codingfor the alpha- and beta-subunits of Xenopus laevis casein kinase II.FEBS Lett 297: 280–284, 1992
13. Wilhelm V, Neckelman G, Allende JE, Allende CC: The genomic struc-ture of two protein kinase CK2alpha genes of Xenopus laevis and fea-tures of the putative promoter region. Mol Cell Biochem 227: 175–183,2001
14. Lieberman SL, Ruderman JV: CK2 beta, which inhibits Mos function,binds to a discrete domain in the N-terminus of Mos. Dev Biol 268:271–279, 2004
15. Theis-Febvre N, Filhol O, Froment C, Cazales M, Cochet C, MonsarratB, Ducommun B, Baldin V: Protein kinase CK2 regulates CDC25Bphosphatase activity. Oncogene 22: 220–232, 2003
16. Buchou T, Vernet M, Blond O, Jensen HH, Pointu H, Olsen BB, CochetC, Issinger OG, Boldyreff B: Disruption of the regulatory beta subunitof protein kinase CK2 in mice leads to a cell-autonomous defect andearly embryonic lethality. Mol Cell Biol 23: 908–915, 2003
17. Chen-Wu JL, Padmanabha R, Glover CV: Isolation, sequencing, anddisruption of the CKA1 gene encoding the alpha subunit of yeast caseinkinase II. Mol Cell Biol 8: 4981–4990, 1988
18. Padmanabha R, Chen-Wu JL, Hanna DE, Glover CV: Isolation, se-quencing, and disruption of the yeast CKA2 gene: casein kinase II isessential for viability in Saccharomyces cerevisiae. Mol Cell Biol 10:4089–4099, 1990
19. Xu X, Toselli PA, Russell LD, Seldin DC: Globozoospermia in micelacking the casein kinase II alpha’ catalytic subunit. Nat Genet 23: 118–121, 1999
20. Dominguez I, Mizuno J, Wu H, Song DH, Symes K, Seldin DC: Proteinkinase CK2 is required for dorsal axis formation in Xenopus embryos.Dev Biol 274: 110–124, 2004
21. Newport J, Kirschner M: A major developmental transition in earlyXenopus embryos: I. characterization and timing of cellular changes atthe midblastula stage. Cell 30: 675–686, 1982
22. Nieuwkoop PD, Faber J: Normal Table of Xenopus Laevis. North-Holland Publishing Company, Amsterdam, 1967
23. Schneider VA, Mercola M: Spatially distinct head and heart inducerswithin the Xenopus organizer region. Curr Biol 9: 800–809, 1999
24. Wilhelm V, Rojas P, Gatica M, Allende CC, Allende JE: Expression ofthe subunits of protein kinase CK2 during oogenesis in Xenopus laevis.Eur J Biochem 232: 671–676, 1995
25. Sokol S, Christian JL, Moon RT, Melton DA: Injected Wnt RNA inducesa complete body axis in Xenopus embryos. Cell 67: 741–752, 1991
26. Smith WC, Harland RM: Injected Xwnt-8 RNA acts early in Xenopusembryos to promote formation of a vegetal dorsalizing center. Cell 67:753–765, 1991
27. Sokol SY, Klingensmith J, Perrimon N, Itoh K: Dorsalizing and neural-izing properties of Xdsh, a maternally expressed Xenopus homolog ofdishevelled. Development 121: 3487, 1995
28. Dominguez I, Itoh K, Sokol SY: Role of glycogen synthase kinase 3beta as a negative regulator of dorsoventral axis formation in Xenopusembryos. Proc Natl Acad Sci USA 92: 8498–8502, 1995
29. Pierce SB, Kimelman D: Regulation of Spemann organizer formationby the intracellular kinase Xgsk-3. Development 121: 755–765, 1995
30. He X, Saint-Jeannet JP, Woodgett JR, Varmus HE, Dawid IB: Glyco-gen synthase kinase-3 and dorsoventral patterning in Xenopus embryos.Nature 374: 617–622, 1995
31. Guger KA, Gumbiner BM: Beta-catenin has Wnt-like activity and mim-ics the Nieuwkoop signaling center in Xenopus dorsal-ventral pattern-ing. Dev Biol 172: 115–125, 1995
32. Cadigan KM, Nusse R: Wnt signaling: a common theme in animaldevelopment. Genes Dev 11: 3286–3305, 1997
33. Fagotto F, Guger K, Gumbiner BM: Induction of the primary dorsalizingcenter in Xenopus by the Wnt/GSK/beta-catenin signaling pathway, butnot by Vg1, Activin or Noggin. Development 124: 453–460, 1997
34. Smalley MJ, Dale TC: Wnt signaling and mammary tumorigenesis. JMammary Gland Biol Neoplasia 6: 37–52, 2001
35. Polakis P: Wnt signaling and cancer. Genes Dev 14: 1837–1851, 200036. Brown AM: Wnt signaling in breast cancer: have we come full circle?
Breast Cancer Res 3: 351–355, 200137. Giles RH, van Es JH, Clevers H: Caught up in a Wnt storm: Wnt sig-
naling in cancer. Biochim Biophys Acta 1653: 1–24, 200338. Seldin DC, Leder P: Casein kinase II alpha transgene-induced murine
lymphoma: relation to theileriosis in cattle. Science 267: 894–897, 199539. Landesman-Bollag E, Romieu-Mourez R, Song DH, Sonenshein GE,
Cardiff RD, Seldin DC: Protein kinase CK2 in mammary gland tumori-genesis. Oncogene 20: 3247–3257, 2001
40. Song DH, Dominguez I, Mizuno J, Kaut M, Mohr SC, Seldin DC:CK2 phosphorylation of the armadillo repeat region of beta-cateninpotentiates Wnt signaling. J Biol Chem 278: 24018–24025, 2003
41. Vilk G, Saulnier RB, St Pierre R, Litchfield DW: Inducible expressionof protein kinase CK2 in mammalian cells. Evidence for functionalspecialization of CK2 isoforms. J Biol Chem 274: 14406–14414, 1999
42. Heasman J, Crawford A, Goldstone K, Garner-Hamrick P, Gumbiner B,McCrea P, Kintner C, Noro CY, Wylie C: Overexpression of cadherinsand underexpression of beta-catenin inhibit dorsal mesoderm inductionin early Xenopus embryos. Cell 79: 791–803, 1994
43. Heasman J, Kofron M, Wylie C: Beta-catenin signaling activity dis-sected in the early Xenopus embryo: a novel antisense approach. DevBiol 222: 124–134, 2000
44. Hoppler S, Brown JD, Moon RT: Expression of a dominant-negativeWnt blocks induction of MyoD in Xenopus embryos. Genes Dev 10:2805–2817, 1996
45. Sokol SY: Analysis of dishevelled signalling pathways during Xenopusdevelopment. Curr Biol 6: 1456–1467, 1996
46. Yost C, Farr GH, 3rd, Pierce SB, Ferkey DM, Chen MM, Kimelman D:GBP, an inhibitor of GSK-3, is implicated in Xenopus development andoncogenesis. Cell 93: 1031–1041, 1998
47. Kageura H: Activation of dorsal development by contact between thecortical dorsal determinant and the equatorial core cytoplasm in eggs ofXenopus laevis. Development 124: 1543–1551, 1997
48. Yang J, Tan C, Darken RS, Wilson PA, Klein PS: Beta-catenin/Tcf-regulated transcription prior to the midblastula transition. Development129: 5743–5752, 2002