Embed Size (px)
Citation preview

Plant Molecular Biology41: 45–55, 1999.© 1999Kluwer Academic Publishers. Printed in the Netherlands.
45
A tobacco cryptic constitutive promoter, tCUP, revealed by T-DNA tagging
Elizabeth Foster1, Jiro Hattori1, Helene Labbe1, Therese Ouellet1, Pierre R. Fobert2,3, Lisa E.James1, V.N. Iyer2 and Brian L. Miki1,∗1Eastern Cereal and Oilseed Research Centre, Agriculture and Agri-Food Canada, Ottawa, Ontario, CanadaK1A 0C6 (∗author for correspondence: Room 2091 K.W. Neatby Building);2Department of Biology, CarletonUniversity, Ottawa, Ontario, Canada K1S 5B6;3present address: National Research Council, Plant BiotechnologyInstitute, 100 Gymnasium Place, Saskatoon, Saskatchewan, Canada S7N 0W9∗∗
Received 28 October 1998; accepted in revised form 23 June 1999
Key words:cryptic, constitutive, promoter, T-DNA tagging, tobacco
Abstract
We have isolated a constitutive promoter sequence,tCUP, from tobacco by T-DNA tagging using a promoterlessGUS-nos3′ reporter gene construct. The T-DNA integration event produced a translational fusion with the GUSgene that is expressed widely in organs, at both the mRNA and enzyme activity levels. In tobacco transformedwith a tCUP-GUS-nos3′ gene, GUS specific activity in leaves was within a range of values similar to those ofplants transformed with the widely used constitutive promoter gene fusion, CaMV 35S promoter-GUS-nos3′.Characteristics of thetCUPpromoter sequence differ from those of other plant constitutive promoters; for instance,the tCUP sequence lacks a TATA box. Transcription initiates at a single site within the tCUP sequence which issimilar to a transcriptional start site consensus sequence determined for plant genes. ThetCUPpromoter is crypticas RNA accumulation at the transcriptional start site is not detected in untransformed tobacco. Thus,tCUP is thefirst example of a cryptic, constitutive promoter isolated from plants. ThetCUP-GUS-nos3′ gene fusion producedGUS activity in tissues of all species tested suggesting thattCUPmay utilize fundamental transcription mechanismsfound in plants.
Introduction
Only a few constitutive promoters have been clonedand characterized from plants, including promotersfrom the actin genes of rice (Zhanget al., 1991) andArabidopsis(An et al., 1996), the translation initiationfactor 4A gene of tobacco (Mandelet al., 1995), theGOS2 gene of rice (de Pateret al., 1992) and ubiq-uitin genes inArabidopsis(Callis et al., 1990) andmaize (Christensenet al., 1992). Therefore, in exper-iments with transgenic plants only a limited numberof plant promoters have been used when constitutive
The nucleotide sequence data reported will appear in theEMBL, GenBank and DDBJ Nucleotide Sequence Databases underthe accession number AF133844 (tCUP).∗∗author(s): E Foster, J Hattori, H Labbe, T Ouellet, LE James,BL Miki. For the department of Agriculture and Agri-Food, Gov-ernment of Canada. © Minister of Public Works and GovernmentServices Canada 1998.
gene expression is desired. Indeed, the most widelyused constitutive promoter in transgenic plants is the35S promoter isolated from cauliflower mosaic virusand its derivatives (Odellet al., 1985; Datlaet al.,1993; Holtorfet al., 1995; Wilminket al., 1995).
Most plant promoters have traditionally been de-rived from genes with desirable expression patterns.T-DNA tagging (Feldmann, 1991; Waldenet al., 1991)has proven to be a useful method for identifyingsuch genes, as T-DNAs may insert preferentially intotranscribed regions of the genome. Alternatively, pro-moter and enhancer trapping in transgenic plants, byscreening for reporter gene expression, can also beeffective for the isolation of regulatory sequences ofexpressed genes (Weiet al., 1997). A high-frequencyAgrobacteriumtransformation system is being devel-oped to facilitate saturation mutagenesis with T-DNAsfor large-scale functional analysis of genes (Mathur
46
et al., 1998) and this may eventually augment the di-versity of gene regulatory elements that are currentlyavailable to study and exploit.
Sequences which normally do not function as generegulatory elements are referred to as cryptic (Irnigeret al., 1992; Fobertet al., 1994; Palmeret al., 1994;Troyanovsky and Leube, 1994; Kapoun and Kaufman,1995; Teeet al., 1995). Such cryptic regulatory ele-ments are inactive, but become activated as promotersor regulatory elements when placed in new contexts.Cryptic elements have predominantly been foundwithin or adjacent to genes, but have also been dis-covered in untranscribed regions of the genome. Re-cent examples of cryptic elements include enhancerswithin an intron of theDrosophila proboscipediagene(Kapoun and Kaufman, 1995), promoters within in-trons of yeast actin (Irnigeret al., 1992) and humancomplement component C4 genes (Teeet al., 1995),and elements within promoters of a human cytoker-atin pseudogene (Troyanovsky and Leube, 1994) andthe mouse cytochrome CYP4A6 gene (Palmeret al.,1994). We have previously discovered a novel cryp-tic promoter fromNicotiana tabacumwhich directsseed-coat-specific expression and originated within anoncoding region of the tobacco genome (Fobertet al.,1994).
Here, we report the activation by T-DNA taggingof a promoter in the tobacco genome and the iso-lation of the first example of a cryptic, constitutivepromoter. Thetobaccoconstitutive promoter, referredto astCUP, is expressed widely in organs and exhibitsGUS specific activity in tobacco leaves at levels simi-lar to the 35S promoter. Notable features of thetCUPsequence include its cryptic nature and its lack ofpromoter consensus sequences, distinguishing it fromother plant constitutive promoters.
Materials and methods
T-DNA tagging of tobacco
Tobacco transgenic line T1275 was obtained byAgrobacterium-mediated transformation with a binaryvector containing a promoterlessGUScoding region(Fobertet al., 1991). Segregation analysis of the trans-genic plants was performed as described (Fobertet al.,1991).
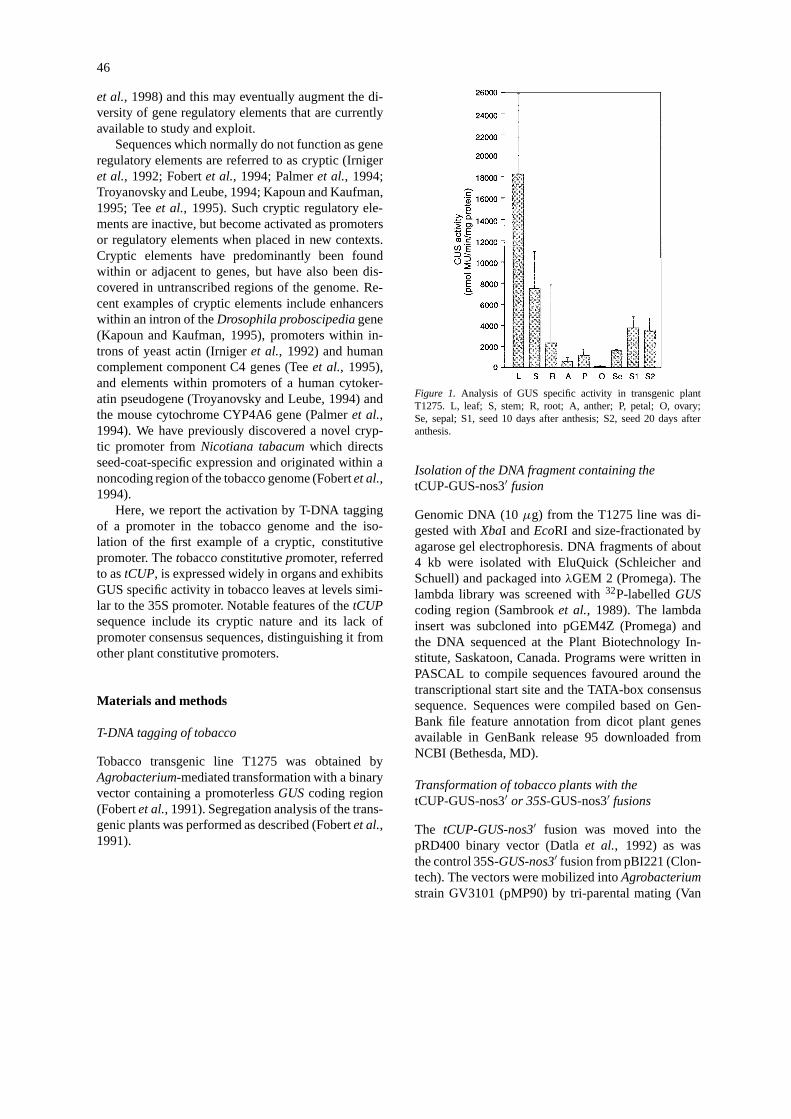
Figure 1. Analysis of GUS specific activity in transgenic plantT1275. L, leaf; S, stem; R, root; A, anther; P, petal; O, ovary;Se, sepal; S1, seed 10 days after anthesis; S2, seed 20 days afteranthesis.
Isolation of the DNA fragment containing thetCUP-GUS-nos3′ fusion
Genomic DNA (10µg) from the T1275 line was di-gested withXbaI andEcoRI and size-fractionated byagarose gel electrophoresis. DNA fragments of about4 kb were isolated with EluQuick (Schleicher andSchuell) and packaged intoλGEM 2 (Promega). Thelambda library was screened with32P-labelledGUScoding region (Sambrooket al., 1989). The lambdainsert was subcloned into pGEM4Z (Promega) andthe DNA sequenced at the Plant Biotechnology In-stitute, Saskatoon, Canada. Programs were written inPASCAL to compile sequences favoured around thetranscriptional start site and the TATA-box consensussequence. Sequences were compiled based on Gen-Bank file feature annotation from dicot plant genesavailable in GenBank release 95 downloaded fromNCBI (Bethesda, MD).
Transformation of tobacco plants with thetCUP-GUS-nos3′ or 35S-GUS-nos3′ fusions
The tCUP-GUS-nos3′ fusion was moved into thepRD400 binary vector (Datlaet al., 1992) as wasthe control 35S-GUS-nos3′ fusion from pBI221 (Clon-tech). The vectors were mobilized intoAgrobacteriumstrain GV3101 (pMP90) by tri-parental mating (Van

47
Haute et al., 1983). Agrobacteriumtransformationof tobacco cultivar SR1 was as described (Fobertet al., 1991). GUS fluorogenic assays in triplicateand histochemical staining were performed as de-scribed (Jefferson, 1987). Western blot analysis wasperformed after SDS-PAGE and blotting using theChemiluminescence Western Blotting Kit (BoehringerMannheim) according to the manufacturer’s instruc-tions, using rabbit anti-β-glucuronidase IgG (Molecu-lar Probes) primary antibody, sheep anti-rabbit IgG-POD (Boehringer Mannheim) secondary antibody,and the same protein extracts as subjected to fluo-rogenic assay. Within an experiment, plant materialswere harvested at similar stages of development fromplants grown under similar conditions.
RNase protection assays
Total RNA was isolated from seedling, leaf, stem,root, flower bud, petal and ovary of untransformedtobacco and tobacco transformed withtCUP-GUS-nos3′, and seedlings from transformed line T1275using the TriPure Isolation Reagent (BoehringerMannheim) according to the manufacturer’s instruc-tions. Total RNA from seed 14 days after anthesiswas isolated as previously described (Fobertet al.,1994). RNase protection assays were performed us-ing the RPA II kit (Ambion). Probes were preparedusing the Riboprobe Gemini System II (Promega).Probes prepared to detect the transcriptional start sitein transgenic line T1275 and in tobacco transformedwith tCUP-GUS-nos3′ were situated about−446 to+596 relative to the detected transcriptional start site.Antisense and sense probes were prepared to detecttranscripts on either strand on either side of the tran-scriptional start site from untransformed tobacco fromabout−446 to +169. For quantification, autoradi-ograms were scanned and analysed using the Gel Print2000i digital camera system and GPTools softwarefor determination of relative grey scale of exposure(BioPhotonics Corporation, Michigan).
Results
Constitutive expression of GUS in a T-DNA taggedline
About 1000 tobacco transgenic lines were created byT-DNA tagging with a vector containing a promoter-lessGUS coding region, encoding the bacterial en-zymeβ-glucuronidase, andnopaline synthasetermi-
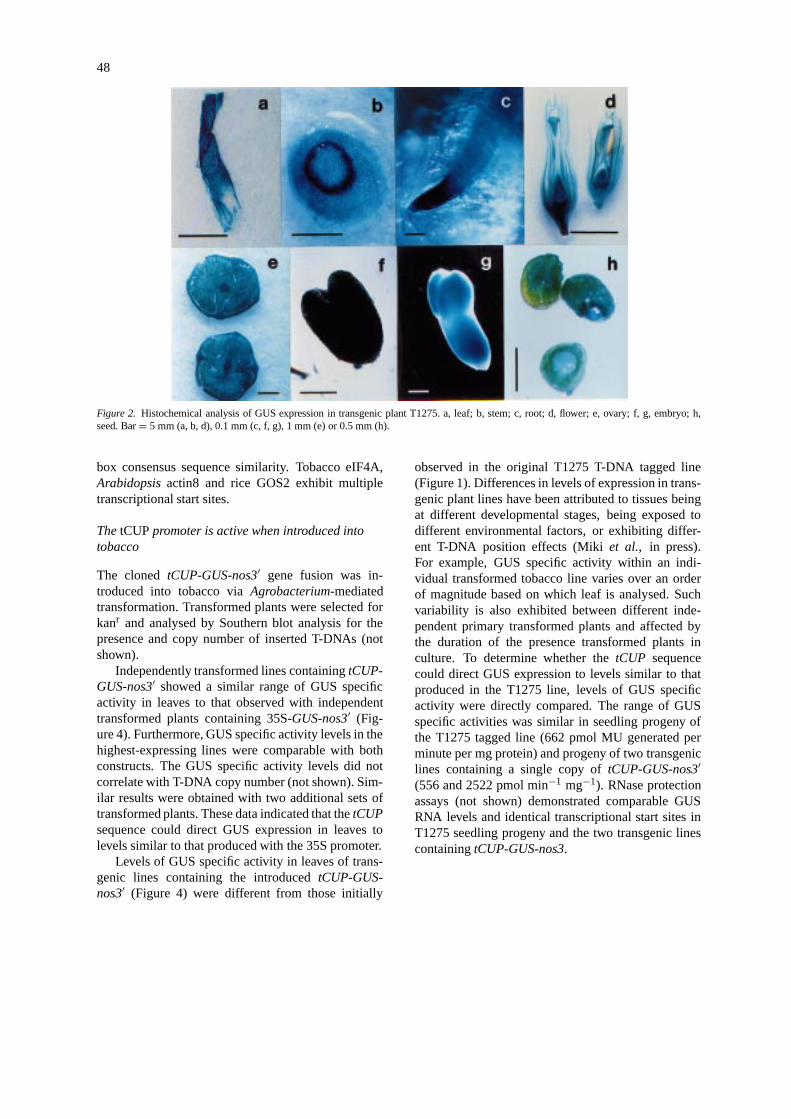
nator (nos3′) at the T-DNA right border, along with akanamycin resistance marker gene,nptII (Fobertet al.,1991). In one of the transgenic lines, T1275, GUSspecific activity was detected in leaf, at a high level,and in other organs of the plant examined (Figure 1).GUS expression was also detected histochemically intissues of all organs examined (Figure 2).
In the T1275 line, a 3:1 segregation ratio of the T-DNA kanr marker (262 kanr : 88 kans) was observed.A Southern blot of T1275 genomic DNA digested withEcoRI which cuts the T-DNA once, between the loca-tions of theGUSandnptII probes, showed a single,hybridizing band with either probe (Fobert, 1992).These data are consistent with insertion of a singleT-DNA copy at a single locus. Furthermore, the ac-cumulated data suggested that the tobacco genomicsequence adjacent to the integrated promoterlessGUScoding region supplied thecis-regulatory elementsneeded for constitutive expression of GUS.
Cloning of the constitutive promoter,tCUP
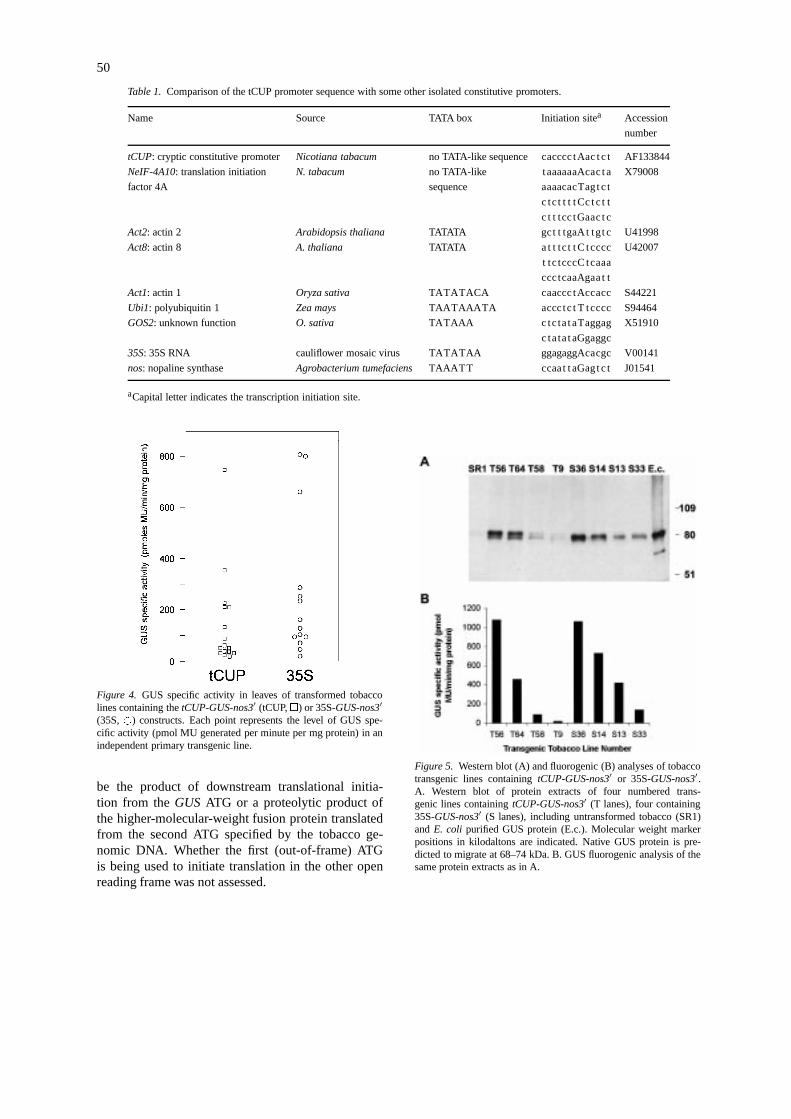
A 4.4 kb XbaI-EcoRI DNA fragment detected bySouthern blot analysis, which contains a 2.2 kb to-bacco genomic sequence upstream of the T-DNAinsertion point (referred to as thetCUP sequence)fused with theGUS coding region andnos3′ termi-nator, was cloned and sequenced. A restriction mapof the 4.4 kb cloned fragment and thetCUP sequencearound the transcriptional and translational start sitesis presented in Figure 3. Sequence comparison withthe original T-DNA vector sequence indicated thepoint of T-DNA integration within the T-DNA rightborder. The transcriptional start site was delimitedby RNase protection assay (not shown) and was lo-cated at a single position about 220 bp upstream ofthe translational initiation codon of theGUS codingregion in the T-DNA. The sequence around the tran-scriptional start site exhibits similarity with sequencesfavoured at the transcriptional start site that we com-piled from available dicot plant genes in GenBank(T/A T/C A+1, A C/A C/A A/C/T A A A/T). Se-quence similarity is not detected about 30 bp upstreamof the transcriptional start site with the TATA-box con-sensus that we compiled from available dicot plantgenes (C T A T A A/T A T/A A). We also com-pared the tCUP sequence in these regions with thoseof some of the other isolated plant, viral or bacterialpromoters that exhibit constitutive expression whenintroduced into transgenic plants (Table 1). Unlike thetCUP sequence, all but tobacco eIF4A exhibit TATA-
48
Figure 2. Histochemical analysis of GUS expression in transgenic plant T1275. a, leaf; b, stem; c, root; d, flower; e, ovary; f, g, embryo; h,seed. Bar= 5 mm (a, b, d), 0.1 mm (c, f, g), 1 mm (e) or 0.5 mm (h).
box consensus sequence similarity. Tobacco eIF4A,Arabidopsisactin8 and rice GOS2 exhibit multipletranscriptional start sites.
ThetCUPpromoter is active when introduced intotobacco
The clonedtCUP-GUS-nos3′ gene fusion was in-troduced into tobacco viaAgrobacterium-mediatedtransformation. Transformed plants were selected forkanr and analysed by Southern blot analysis for thepresence and copy number of inserted T-DNAs (notshown).
Independently transformed lines containingtCUP-GUS-nos3′ showed a similar range of GUS specificactivity in leaves to that observed with independenttransformed plants containing 35S-GUS-nos3′ (Fig-ure 4). Furthermore, GUS specific activity levels in thehighest-expressing lines were comparable with bothconstructs. The GUS specific activity levels did notcorrelate with T-DNA copy number (not shown). Sim-ilar results were obtained with two additional sets oftransformed plants. These data indicated that thetCUPsequence could direct GUS expression in leaves tolevels similar to that produced with the 35S promoter.
Levels of GUS specific activity in leaves of trans-genic lines containing the introducedtCUP-GUS-nos3′ (Figure 4) were different from those initially
observed in the original T1275 T-DNA tagged line(Figure 1). Differences in levels of expression in trans-genic plant lines have been attributed to tissues beingat different developmental stages, being exposed todifferent environmental factors, or exhibiting differ-ent T-DNA position effects (Mikiet al., in press).For example, GUS specific activity within an indi-vidual transformed tobacco line varies over an orderof magnitude based on which leaf is analysed. Suchvariability is also exhibited between different inde-pendent primary transformed plants and affected bythe duration of the presence transformed plants inculture. To determine whether thetCUP sequencecould direct GUS expression to levels similar to thatproduced in the T1275 line, levels of GUS specificactivity were directly compared. The range of GUSspecific activities was similar in seedling progeny ofthe T1275 tagged line (662 pmol MU generated perminute per mg protein) and progeny of two transgeniclines containing a single copy oftCUP-GUS-nos3′(556 and 2522 pmol min−1 mg−1). RNase protectionassays (not shown) demonstrated comparable GUSRNA levels and identical transcriptional start sites inT1275 seedling progeny and the two transgenic linescontainingtCUP-GUS-nos3.

49
Figure 3. Restriction map and partial nucleotide sequence of the 4.4 kbXbaI–EcoRI cloned DNA fragment constituting thetCUP-GUS-nos3′reporter gene fusion. The restriction map indicates the restriction enzyme sites by name and the transcriptional start site with the arrow. Arrowssituated above the restriction map represent the antisense and sense RNase protection assay probes used to detect the transcriptional start site(antisense) and test for RNA accumulated in untransformed tobacco corresponding to the region about−446 and+169 (sense and antisense).Map scale is indicated in kb. Beneath the restriction map in capitals is the partial nucleotide sequence of the clonedtCUP-GUS-nos3′ genefusion from about−71 of the transcriptional start site to the methionine initiator codon of theGUScoding region from the T-DNA. Included atthe 3′ end of the sequence are 48 nucleotides at the right border of the T-DNA sequence in bold italics. The transcript initiates at a single sitewithin the region indicated in bold. Methionine amino acids specified by the tobacco genomic DNA are indicated in bold, putative translationproducts are shown and stop codons are indicated (∗∗∗). Sequence is numbered according to the transcriptional start site consensus within theregion of the single transcriptional start site.
tCUP is a GUS translational fusion
During sequence analysis, two ATG codons withinthe tobacco genomic DNA were found between thetranscriptional start site and theGUS coding regionof tCUP-GUS-nos3′. The first ATG is in a differ-ent reading frame but the second one is in the samereading frame as theGUScoding region, which maylead to production of an N-terminal extension on theGUS protein (Figure 3). Western blot analysis of pro-tein extracts from tobacco plants transformed withtCUP-GUS-nos3′ consistently demonstrated the pro-duction of immunodetectable GUS protein having ahigher molecular weight than that in tobacco trans-formed with35S-GUS-nos3′, which was not predictedto produce a fusion protein (Figure 5A). The higher-molecular-weight band was detected in alltCUP-GUS-nos3′ transformed plants tested. Furthermore,
translationin vitro in wheat germ extract of GUS pro-tein with the predicted N-terminal fusion (equivalentto thetCUP-GUS-nos3′ translation product) produceda higher-molecular-weight band by SDS-PAGE thanthat of GUS without an N-terminal fusion (equiva-lent to the 35S-GUS-nos3′ translation product) (notshown). This suggested that aGUS translational fu-sion was initiated at the second ATG which is inframe with theGUScoding region and derived fromthe tobacco genomic DNA. Furthermore, GUS ac-tivity levels were not affected by the translationalfusion present intCUP-GUS-nos3′, as relative levelsof immunodetectable GUS protein reflected the rel-ative levels of GUS specific activity over the rangeof expression observed in bothtCUP-GUS-nos3′ and35S-GUS-nos3′ containing plants (Figure 5B). Thelower-molecular-weight band seen on the western blotof tCUP-GUS-nos3′ transgenic lines (Figure 5A) may
50
Table 1. Comparison of the tCUP promoter sequence with some other isolated constitutive promoters.
Name Source TATA box Initiation sitea Accession
number
tCUP: cryptic constitutive promoter Nicotiana tabacum no TATA-like sequence cacccc tAac t c t AF133844
NeIF-4A10: translation initiation N. tabacum no TATA-like taaaaaaAcac t a X79008
factor 4A sequence aaaacacTagt c t
c tc t t t t Cc t c t t
c t t tcc tGaac t c
Act2: actin 2 Arabidopsis thaliana TATATA gct t tgaAt tgt c U41998
Act8: actin 8 A. thaliana TATATA a t t tc t t C tcccc U42007
t tc tcccC tcaaa
ccctcaaAgaa t t
Act1: actin 1 Oryza sativa TATATACA caaccc tAccacc S44221
Ubi1: polyubiquitin 1 Zea mays TAATAAATA accctc t T tcccc S94464
GOS2: unknown function O. sativa TATAAA c tctat aTaggag X51910
c tatat aGgaggc
35S: 35S RNA cauliflower mosaic virus TATATAA ggagaggAcacgc V00141
nos: nopaline synthase Agrobacterium tumefaciens TAAATT ccaat t aGagt c t J01541
aCapital letter indicates the transcription initiation site.
Figure 4. GUS specific activity in leaves of transformed tobaccolines containing thetCUP-GUS-nos3′ (tCUP,�) or 35S-GUS-nos3′(35S,#) constructs. Each point represents the level of GUS spe-cific activity (pmol MU generated per minute per mg protein) in anindependent primary transgenic line.
be the product of downstream translational initia-tion from theGUS ATG or a proteolytic product ofthe higher-molecular-weight fusion protein translatedfrom the second ATG specified by the tobacco ge-nomic DNA. Whether the first (out-of-frame) ATGis being used to initiate translation in the other openreading frame was not assessed.
Figure 5. Western blot (A) and fluorogenic (B) analyses of tobaccotransgenic lines containingtCUP-GUS-nos3′ or 35S-GUS-nos3′ .A. Western blot of protein extracts of four numbered trans-genic lines containingtCUP-GUS-nos3′ (T lanes), four containing35S-GUS-nos3′ (S lanes), including untransformed tobacco (SR1)andE. coli purified GUS protein (E.c.). Molecular weight markerpositions in kilodaltons are indicated. Native GUS protein is pre-dicted to migrate at 68–74 kDa. B. GUS fluorogenic analysis of thesame protein extracts as in A.

51
Figure 6. RNase protection assay of RNA isolated from leaf (L),stem (St), root (R), flower bud (F) and developing seed (Se) tissuesof progeny of a transformed tobacco line containing one copy oftCUP-GUS-nos3′ (10µg RNA) and untransformed tobacco (30µgRNA). Undigested probe (P), yeast RNA negative control (−) lanesand size markers are indicated. RNase protection assays shown useda probe to detect sense transcripts between about−446 and+596of tCUP-GUS-nos3′ or between about−446 and+169 of untrans-formed tobacco. The protected fragment in transformed plants isabout 596 bp (upper arrowhead) and, if present, accumulated tran-scripts initiated at this site in untransformed plants are predicted toprotect a fragment of about 169 bp (lower arrowhead). Upper bandin RNA-containing lanes was added to samples to indicate loss ofsample during assay.
Constitutive expression oftCUP in tobacco
GUSexpression levels were examined in progeny oftransgenic tobacco lines containingtCUP-GUS-nos3′.RNA was detected in all organs of tobacco examined(leaf, stem, root, flower bud, and developing seed,Figure 6; leaf, root, flower bud, petal, ovary anddeveloping seed, Table 2).GUSRNA levels were mea-sured in the different organs tested from an individualprogeny plant from each of two transgenic lines (Ta-ble 2). The transcriptional start sites were mapped byRNase protection assay to the same position in eachorgan tested (leaf, stem, root, flower bud, and devel-oping seed, Figure 6; petal and ovary, not shown).This confirmed that elements which confer transcrip-tion widely in organs are contained in the isolatedtCUP sequence.
Table 2. RelativeGUSRNA levels (grey scaleunits) detected by RNase protection in organsof a single progeny of each of two transformedlines (T51 and T64) containing a single copyof tCUP-GUS-nos3′.
Organ RelativeGUSRNA level
(grey scale units)
T51 T64
Leaf 852 185
Root 270 96
Flower bud 173 28
Petal 121 48
Ovary 322 nda
Seed (14 dpa) 324 811
anot determined.
GUS enzyme specific activity was measured in themajor organs of kanr progeny from three indepen-dently transformed lines containingtCUP-GUS-nos3′(Table 3). The variation in the relative levels of GUSspecific activity in the different organs (Table 3) wasgreater than that observed for the relativeGUSRNAlevels (Table 2). Transformed plants containing35S-GUS-nos3′ exhibited a similar pattern of GUS activityin organs. Histochemical analysis of developing seeds,seedlings and flowers of transgenic tobacco contain-ing tCUP-GUS-nos3′ showed GUS expression com-parable to transgenic tobacco containing the controlconstitutive gene fusion 35S-GUS-nos3′ (Figure 7).
Activation of a cryptic promoter
RNase protection assays were performed using RNAfrom organs of untransformed tobacco and sense andantisense probes. Both probes spanned thetCUP se-quence from about−446 to+169 on both sides ofthe transcriptional start site (Figure 3). In all tissuestested with the antisense probe (leaf, stem, root, flowerbud, and developing seed, Figure 6; petal and ovary,not shown) protected fragments were not detected, in-dicating that native transcripts were not perceptiblein untransformed tobacco that correspond to the tran-scriptional start site in the T1275 line (not shown)and tobacco transformed withtCUP-GUS-nos3′ (Fig-ure 6). Protected fragments, which may correspondto transcripts from the opposite strand, were not de-tected with the sense probe in any organ tested (leaf,stem, root, flower bud, developing seed, petal andovary with sense probe, not shown). The RNase pro-tection conditions can detect RNAs representing an

52
Table 3. Average GUS specific activity (pmol MU generated per minuteper mg total protein) in organs of 24 progeny of transformed tobaccolines containing tCUP-GUS-nos3′ compared with those containing35S-GUS-nos3′ or untransformed tobacco. Eight kanr progeny of eachof three independent lines were tested. Two of the three lines trans-formed with tCUP-GUS-nos3′ and two of three with 35S-GUS-nos3′contained a single T-DNA insert, while the thirdtCUP-GUS-nos3′line contained four T-DNA copies and the third 35S-GUS-nos3′ linecontained three inserts.
Organ GUS specific activity (pmol MU min−1 mg−1) ± SEM
in transformed tobacco
tCUP-GUS-nos3′ 35S-GUS-nos3′ Untransformed
Leaf 1172± 233 2119± 432 3.0
Stem 672± 132 1168± 228 7.6
Root 2938± 449 3905± 507 22.2
Seed 393± 45.8 1027± 380 10.0
Petals 42.5± 7.4 174± 25.8 1.3
Anthers 19.6± 3.0 35.9± 9.8 0.4
Pistils 14.8± 2.2 31.8± 4.6 1.8
Sepals 111± 22.4 465± 82.0 2.5
Ovary 5.5± 0.5 21.9± 3.5 2.7
estimated 0.001% to 0.01% of total RNA levels (Ouel-let et al., 1992). These data suggest that the regionof the tCUP sequence in which transcript initiationwas detected intCUP-GUS-nos3′ does not direct thecomparable accumulation of transcribed RNA at thenative location in organs of untransformed tobacco,and is probably inactive. This is consistent with theactivation of a cryptic promoter by insertion of theT-DNA. Other explanations for the lack of detectabletranscripts in untransformed tobacco could include ac-cumulation below detectable levels, or transcription ina restricted pattern in untested tissues.
Discussion
The data presented here support the isolation of acryptic, constitutive promoter,tCUP, from tobacco.The tCUP sequence activatesGUS reporter gene ex-pression in all organs of transgenic tobacco tested.The tCUP sequence also directs expression of GUSin tobacco leaves to levels comparable to those ob-tained with a widely used constitutive promoter, thecauliflower mosaic virus 35S promoter. To our knowl-edge,tCUP is the first example of a cryptic, constitu-tive promoter and furthermore provides an example ofa cryptic promoter of plant origin which may prove
to be as useful as the 35S promoter for directingconstitutive transgene expression.
In preliminary experiments, thetCUP sequencewas found to direct transgene expression in a widerange of plant species. Leaf tissue of canola, tobacco,soybean, alfalfa, pea andArabidopsis, and cell sus-pensions of oat, maize, wheat and barley exhibitedGUS-positive blue spots after transient bombardment-mediated assays and histochemical GUS activity stain-ing (not shown). The number of GUS-positive bluespots was higher in dicot leaf tissues than in monocotcell suspensions. Nonetheless, the expression of thetCUP sequence in both dicots and monocots suggeststhat it may be useful for directing gene expression in awide range of plant species.
ThetCUPsequence does not contain sequence fea-tures generally associated with plant promoters andimplicated in their function, such as TATA-box se-quences. This lack of promoter consensus sequencesdistinguishes thetCUP sequence from most of theother promoters commonly studied in plants, includ-ing constitutive promoters. For instance, the ubiquitinUbi1 gene from maize contains a TATA-box sequence,other elements and sequence motifs bearing sequencesimilarity to other promoters and initiates transcriptionpredominantly at three adjacent nucleotides (Chris-tensenet al., 1992). Constitutively expressed actingenes fromArabidopsisalso exhibit putative TATA-box sequences; the ACT2 gene has a unique start site,while the ACT8 gene has multiple starts (Anet al.,1996). Although a translation initiation factor 4A genefrom tobacco also lacks a TATA box it initiates tran-scription at multiple sites, unliketCUP (Mandelet al.,1995).
A notable sequence characteristic oftCUP is thesimilarity to the consensus sequence for plant genessurrounding the transcriptional start site. Other func-tional elements which have been located at the tran-scriptional start site include a rice initiatorciselement(TCCA+1AG) which directs the siting of transcrip-tion initiation (Zhu et al., 1995) and the transcrip-tional Initiator element (Inr) consensus (Py Py A+1,N T/A Py Py) that is capable of independently direct-ing transcription and is conserved among vertebratesand invertebrates (Lo and Smale, 1996). The sequencesurrounding the transcriptional start site of tCUP hassequence similarity with the functionally defined riceciselement and the Inr element. Likewise, the consen-sus sequence at the transcriptional start site of plantgenes that we compiled shares sequence similaritywith the above functionally defined elements. In the

53
Figure 7. Histochemical analyses of GUS expression in seeds (a, d), flowers (b, e) and seedlings (c, f) of transgenic plants containingtCUP-GUS-nos3′ (a–c) in comparison to those containing the constitutive control 35S-GUS-nos3′ gene fusion (d–f).
absence of a TATA box, the transcriptional start sitesequence may play an important functional role inexpression from thetCUPpromoter.
Sequences which normally do not function as reg-ulatory elements but become activated as promotersor regulatory elements when placed in new con-texts are referred to as cryptic (Irnigeret al., 1992;Palmeret al., 1994; Fobertet al., 1994; Troyanovskyand Leube, 1994; Kapoun and Kaufman, 1995; Teeet al., 1995). Cryptic elements have predominantlybeen found within or adjacent to genes; howeverour previous discovery in tobacco of a cryptic seed-coat-specific promoter, T218 (Fobertet al., 1994),demonstrated that a cryptic element could also be lo-cated within an untranscribed region of the genome.The tCUP promoter is cryptic at its native locationin untransformed tobacco as transcript accumulationresulting from promoter activity was not detected oneither strand. GenBank searches revealed no signifi-
cant sequence similarity with thetCUPsequence, withone exception. Two short regions of amino acid iden-tity (of ca. 66%) were detected between two openreading frames on the antisense strand of thetCUPgenomic sequence and an open reading frame of anArabidopsisexpressed sequence and its genomic clone(GenBank accession number W43439 and AB006703,respectively). This region of amino acid identity is sit-uated upstream of the sequences which confertCUPpromoter activity (Fosteret al., manuscript in prepara-tion).
Evidence gathered to date on the sequence down-stream of the T-DNA integration point in transgenicline T1275 did not reveal significant similarity withthe GenBank database or rearrangements around thepoint of T-DNA integration (Fosteret al., manuscriptin preparation). Such rearrangements might alter thelocation of existing promoters. Through separate stud-ies on the site of T-DNA integration in the T1275

54
line we hope to provide knowledge about the originsand evolution of cryptic regulatory elements within thegenome.
The available evidence leads us to now suggestthat inactive elements may be more abundant in plantsthan previously believed. It is generally assumed basedon previous studies (Konczet al., 1989; Hermanet al., 1990) that T-DNAs integrate preferentially intotranscribed regions of the genome. However, to ourknowledge we are the first researchers to explorethe hypotheses that cryptic elements capable of acti-vating plant gene expression are abundant and mayalso be preferential targets for T-DNA integration.Because many higher-plant genomes consist mostlyof non-coding or intergenic sequences which do notencode expressed genes, and because traditional ap-proaches for the isolation of gene regulatory elementsconcentrated on the isolation and characterization ofexpressed genes and their associated sequences, manysuch useful cryptic regulatory sequences would beoverlooked. Thus, T-DNA tagging is one tool bywhich a greater variety and range of useful generegulatory elements can be isolated.
Acknowledgements
We thank Dr Ravi Chibbar, Nick Leung, KarenCaswell (Plant Biotechnology Institute, Saskatoon)for assisting with monocot transient expression analy-ses, Dr Sylvia McHugh (Iogen Corporation, Ottawa)for providing SR1 root RNA, and Dr Laurian Robert(Eastern Cereal and Oilseed Research Centre, Agri-culture and Agri-Food Canada) and Dr Mark Gijzen(Southern Crop Protection and Food Research Cen-tre, Agriculture and Agri-Food Canada). ECORCcontribution number 991381.
References
An, Y.-Q., McDowell, J.M., Huang,S., McKinney, E.C., Chambliss,S. and Meagher, R.B. 1996. Strong, constitutive expression oftheArabidopsis ACT2/ACT8actin subclass in vegetative tissues.Plant J. 10: 107–121.
Callis, J., Raasch, J.A. and Vierstra, R.D. 1990. Ubiquitin extensionproteins ofArabidopsis thaliana. J. Biol. Chem. 265: 12486–12493.
Christensen, A.H., Sharrock, R.A. and Quail, P.H. 1992. Maizepolyubiquitin genes; structure, thermal perturbation of expres-sion and transcript splicing, and promoter activity followingtransfer to protoplasts by electroporation. Plant Mol. Biol. 18:675–689.
Datla, R.S., Hamerlindl, J.K., Panchuk, B., Pelcher, L.E. and Keller,W. 1992. Modified binary plant transformation vectors with thewild-type gene encoding NPTII. Gene 211: 383–384.
Datla, R.S., Bekkaoui, F., Hammerlindl, J.K., Pilate, G., Dun-stan, D.I. and Crosby, W.L. 1993. Improved high-level consti-tutive foreign gene expression in plants using an AMV RNA4untranslated leader sequence. Plant Sci. 94: 139–149.
de Pater, B., van der Mark, F., Rueb, S., Katagiri, F., Chua, N.-H.,Schilperoort, R.A. and Hensgens, L.A.M. 1992. The promoter ofrice geneGOS2is active in various different monocot tissues andbinds rice nuclear factor ASF-1. Plant J. 2: 837–844.
Feldmann, K.A. 1991. T-DNA insertion mutagenesis in Arabidop-sis: mutational spectrum. Plant J. 1: 71–82.
Fobert, P.R. 1992. Characterization of chromosomal sites of T-DNA integration by activation of a promoterlessβ-glucuronidase(GUS) gene linked to the T-DNA right border repeat. Ph. D.thesis, Carleton University.
Fobert, P.R., Miki, B.L., Iyer V.N. 1991. Detection of gene regula-tory signals in plants revealed by T-DNA-mediated fusions. PlantMol. Biol. 17: 837–851.
Fobert, P.R., Labbé, H., Cosmopolous, J., Gottlob-McHugh, S.,Ouellet, T., Hattori, J., Sunohara, G., Iyer, V.N. and Miki, B.L.1994. T-DNA tagging of a seed coat-specific cryptic promoter intobacco. Plant J. 6: 567–577.
Herman, L., Jacobs, A., Van Montagu, M. and Depicker, A. 1990.Plant chromosome/marker gene fusion assay for study of normaland truncated T-DNA integration events. Mol. Gen. Genet. 224:248–256.
Holtorf, S., Apel, K. and Bohlmann, H. 1995. Comparison ofdifferent constitutive and inducible promoters for the overexpres-sion of transgenes inArabidopsis thaliana. Plant Mol. Biol. 29:637–646.
Irniger, S., Egli, C.M., Kuenzler, M. and Braus, G.H. 1992.The yeast actin intron contains a cryptic promoter that can beswitched on by preventing transcriptional interference. Nucl.Acids Res. 20: 4733–4739.
Jefferson, R.A. 1987. Assaying chimeric genes in plants: the GUSgene fusion system. Plant Mol. Biol. Rep. 5: 387–405.
Kapoun, A.M. and Kaufman, T.C. 1995. A functional analysisof 5′, intronic and promoter regions of the homeotic geneproboscipediain Drosophila melanogater. Development 121:2127–2141.
Koncz, C., Martini, N., Mayerhofer, R., Koncz-Kalman, Z., Korber,H., Redei, G.P. and Schell, J. 1989. High-frequency T-DNA-mediated gene tagging in plants. Proc. Natl. Acad. Sci. USA 86:8467–8471.
Lo, K. and Smale, S.T. 1996. Generality of a functional initiatorconsensus sequence. Gene 182: 13–22.
Mandel, T., Fleming, A.J. Krahenbuhl, R. and Kuhlemeier, C. 1995.Definition of constitutive gene expression in plants: the transla-tion initiation factor 4A gene as a model. Plant Mol. Biol. 29:995–1004.
Mathur, J., Szabados, Z, Schaefer, S., Grunenberg, B., Lassow, A.,Jonas-Straube, E., Schell, J., Koncz, C. and Koncz-Kalman, Z.1998. Gene identification with sequenced T-DNA tags generatedby transformation ofArabidopsiscell suspension. Plant J. 13:707–716.
Miki, B.L.A., McHugh, S.G., Labbé, H., Ouellet, T., Tolman,J.H. and Brandle, J.E. In press. Genetic transformation of to-bacco: gene expression and applications. In: Y.P.S. Bajaj (Ed.),Biotechnology in Agriculture and Forestry, Springer-Verlag,Berlin.

55
Odell, J.T., Nagy, F. and Chua, N.-H. 1985. Identification of DNAsequences required for activity of the cauliflower mosaic virus35S promoter. Nature 313: 810–812.
Ouellet, T., Rutledge, R.G. and Mik, B.L. 1992. Members of theBrassicaacetohydroxyacid synthase multigene family ofBras-sica napushave divergent patterns of expression. Plant J. 2:321–330.
Palmer, C.N., Hsu, M.-H., Muerhoff, A.S., Griffin, K.J. andJohnson, E.F. 1994. Interaction of the peroxisome proliferator-activated receptor alpha with the retinoid X receptor alpha un-masks a cryptic peroxisome proliferator response element thatoverlaps an ARP-1-binding site in the CYP4A6 promoter. J. Biol.Chem. 269: 18083–18089.
Sambrook, J., Fritsch, E.F. and Maniatis, T. 1989. MolecularCloning: A Laboratory Manual, 2nd ed., Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
Sanford, J., Smith, F. and Russell, J. 1993. Optimizing the biolis-tic process for different biological applications. Meth. Enzymol.217: 483–509.
Tee, M.K., Babalola, G.O., Aza-Blanc, P., Speek, M., Gitelman,S.E. and Miller, W.L. 1995. A promoter within intron 35 of thehumanC4Agene initiates abundant adrenal-specific transcriptionof a 1 kb RNA: location of a cryptic CYP21 promoter element?Hum. Mol. Genet. 4: 2109–2116.
Troyanovsky, S. and Leube, R. 1994. Activation of the silent humancytokeratin 17 pseudogene-promoter region by cryptic enhancerelements of the cytokeratin 17 gene. Eur. J. Biochem. 225: 61–69.
Van Haute, E., Joos, H., Maes, M., Warren, G., Van Montagu, M.and Schell, J. 1983. Intergenic transfer and exchange recombina-tion of restriction fragments cloned in pBR322: a novel strategyfor the reversed genetics of the Ti plasmids ofAgrobacteriumtumefaciens. EMBO J. 2: 411–417.
Walden, R., Hayashi, H. and Schell, J. 1991. T-DNA as a gene tag.Plant J. 1: 281–288.
Wei, W., Twel, D. and Lindsey, K. 1997. A novel nucleic acid heli-case gene identified by promoter trapping inArabidopsis. PlantJ. 11: 1307–1314.
Wilmink, A., van de Ven, B. and Dons, J. 1995. Activity of constitu-tive promoters in various species from the Liliaceae. Plant Mol.Biol. 28: 949–955.
Zhang, W., McElroy, D. and Wu, R. 1991. Analysis of riceAct15′region activity in transgenic rice plants. Plant Cell 3: 1155–1165.
Zhu, Q., Dabi, T. and Lamb, C. 1995. TATA box and initiator func-tions in the accurate transcription of a plant minimal promoter invivo. Plant Cell 7: 1681–1689.





























