Embed Size (px)
Citation preview

Gut 1992;33: 1134-1145
PROGRESS REPORT
Acid-base transport systems in gastrointestinalepithelia
D Gleeson
Cell plasma membranes contain a variety oftransport systems that carry acid or base into orout of the cell. Research over the past decade hasled to remarkable advances in our understandingof these acid-base transport systems and hasestablished that they have several major physio-logical roles. They are the primary means bywhich cells regulate their internal pH and theyalso contribute importantly to regulation of cellvolume and possibly cell proliferation. Further-more, in the gastrointestinal tract they mediateabsorption and secretion, not only of acids andbases but of several other electrolytes andnutrients.The major acid-base transport systems
on mammalian cell plasma membranes areillustrated in Figures 1-3; their specific locationson different cell types are discussed later.Hydrogen, potassium (H+,K+) adenosine tri-phosphatase (ATPase) extrude H+ in exchangefor K' using energy derived directly from ATPhydrolysis. They are present on the apical mem-branes of gastric parietal cells' (Fig 1) and, insome species, colon epithelial cells,2 where theymediate acid secretion. However, they are notwidely distributed in gastrointestinal epithelia.
Other acid base transport systems derive theirenergy, not directly from ATP hydrolysis, butfrom coupling the movement of one ion topassive movement of another ion along itselectrochemical gradient. In the case of sodium/hydrogen (Na+/H+) exchange (Figs 1-3), H'extrusion from the cell is coupled, in a 1: 1 ratio,to Na+ entry down its chemical gradient.3-' Thisgradient depends on the sodium pump, Na+,K+ATPase, which is present on the basolateralmembrane of all gastrointestinal epithelial cellsand extrudes 3 Na+ ions in exchange for 2 K+ions, thus maintaining a low intracellular [Na+],high intracellular [K'], and negative intra-cellular potential. Na+/H+ exchange is almostubiquitous in mammalian cells and can beinhibited by the diuretic amiloride.There are several mechanisms for bicarbonate
(HCO3 ) transport across plasma membranes.67In most tissues, the enzyme carbonic anhydrasemediates rapid equilibration of H' and HCO3with C02, which diffuses freely across all cellmembranes. Therefore, HCO3 transport intothe cell is equivalent to H` transport out of thecell and vice versa.
In the case of chloride/bicarbonate (Cl-/HC03 ) exchange (Figs 1-3), HCO3 extrusionfrom the cell is coupled, in a 1:1 ratio, to Clentry along its chemical gradient. This gradirnt
H+ HCO3- 3Na+
Figure 1: Acid-base transport systems on an activated gastricparietal cell. The apical membrane contains H+-K+ A TPaseand the basolateral membrane contains Na+lH+ exchange,Cl-IHCO3 exchange, and Na+-HCO3 co-transport. Thestochiometry of the Na+-HCO3 co-transporter is not known(n= 1, 2, or 3). Note also basolateral membrane Na+-K'ATPase and apical membrane Cl- and K+ channels. Fordetails, see text (section 2). As in Figures 2 and 3, the transportsystems which derive their energy directlyfrom ATPhydrolysis, are shown in closed circles.
is a result of the negative intracellular potential,which maintains a low intracellular [Cl -]. In thecase of Na+-HCO3 co-transport (Figs 1, 3),HCO3- movement is coupled with that ofNa+ inthe same direction. In most, although not all,cells this sytem operates in the direction of Na+and HCO3- entry into the cell, the driving force,as for Na+/H+ exchange, being the out to in Na+gradient. Typically, Cl-/HCO3- exchange andNa+-HCO3- co-transport are not affected byamiloride but can be inhibited by the disulphonicstilbene DIDS.6 7 Finally, HCO3- can exit fromsome cells' uncoupled to other ions, via plasmamembrane HCO3- channels (Fig 3). The drivingforce here is the negative intracellular potential.There have been two major approaches to
characterising acid-base transport systems.Firstly, by studying radiolabelled ion uptake
Gastroenterology Unit,Royal HallamshireHospital, SheffieldD GleesonCorrespondence to:Dr D Gleeson,Gastroenterology Unit, FloorJ, Royal Hallamshire Hospital,Glossop Road, SheffieldS10 2JFAccepted for publication4 November 1991
134
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from
Acid-base transport systems in gastrointestinal epithelia
H+
Na+
Na+
HCO3--
4~
Ci-
2K+
K+ Cl-H+ 3Na+
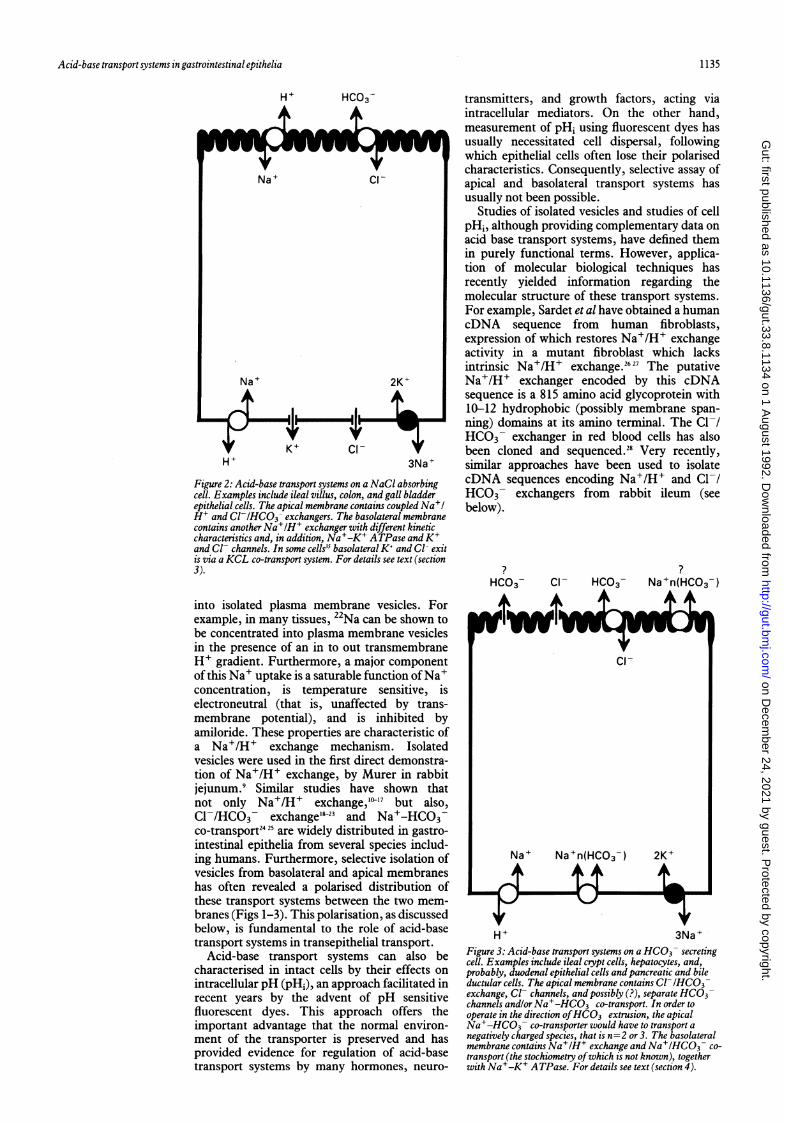
Figure 2: Acid-base transport systems on a NaCI absorbingcell. Examples include ileal villus, colon, and gall bladderepithelial cells. The apical membrane contains coupled Na+lH' and Cl-/HCO3 exchangers. The basolateral membranecontains another Na+lH+ exchanger with different kineticcharacteristics and, in addition, Na+-K' ATPase andK'and Cl- channels. In some cells'5 basolateral K' and Cl- exitis via a KCL co-transport system. For details see text (section3).
into isolated plasma membrane vesicles. Forexample, in many tissues, 22Na can be shown tobe concentrated into plasma membrane vesiclesin the presence of an in to out transmembraneH+ gradient. Furthermore, a major componentof this Na+ uptake is a saturable function ofNa+concentration, is temperature sensitive, iselectroneutral (that is, unaffected by trans-membrane potential), and is inhibited byamiloride. These properties are characteristic ofa Na+/H+ exchange mechanism. Isolatedvesicles were used in the first direct demonstra-tion of Na+/H+ exchange, by Murer in rabbitjejunum.9 Similar studies have shown thatnot only Na+/H+ exchange,"-'7 but also,Cl-HCO3- exchange'8 23 and Na+-HCO3-co-transport2425 are widely distributed in gastro-intestinal epithelia from several species includ-ing humans. Furthermore, selective isolation ofvesicles from basolateral and apical membraneshas often revealed a polarised distribution ofthese transport systems between the two mem-branes (Figs 1-3). This polarisation, as discussedbelow, is fundamental to the role of acid-basetransport systems in transepithelial transport.
Acid-base transport systems can also becharacterised in intact cells by their effects onintracellular pH (pHi), an approach facilitated inrecent years by the advent of pH sensitivefluorescent dyes. This approach offers theimportant advantage that the normal environ-ment of the transporter is preserved and hasprovided evidence for regulation of acid-basetransport systems by many hormones, neuro-
transmitters, and growth factors, acting viaintracellular mediators. On the other hand,measurement of pHi using fluorescent dyes hasusually necessitated cell dispersal, followingwhich epithelial cells often lose their polarisedcharacteristics. Consequently, selective assay ofapical and basolateral transport systems hasusually not been possible.
Studies of isolated vesicles and studies of cellpHi, although providing complementary data onacid base transport systems, have defined themin purely functional terms. However, applica-tion of molecular biological techniques hasrecently yielded information regarding themolecular structure of these transport systems.For example, Sardet et al have obtained a humancDNA sequence from human fibroblasts,expression of which restores Na+/H+ exchangeactivity in a mutant fibroblast which lacksintrinsic Na+/H+ exchange.2627 The putativeNa+/H+ exchanger encoded by this cDNAsequence is a 815 amino acid glycoprotein with10-12 hydrophobic (possibly membrane span-ning) domains at its amino terminal. The Cl-/HCO3- exchanger in red blood cells has alsobeen cloned and sequenced.28 Very recently,similar approaches have been used to isolatecDNA sequences encoding Na+/H+ and Cl-/HCO3 exchangers from rabbit ileum (seebelow).
H+ 3Na+Figure 3: Acid-base transport systems on a HCO3- secretingcell. Examples include ileal crypt cells, hepatocytes, and,probably, duodenal epithelial cells and pancreatic and bileductular cells. The apical membrane contains Cl-IHCO3exchange, Cl- channels, and possibly (?), separate HCO3channels andlor Na+-HCO3 co-transport. In order tooperate in the direction ofHCO3 extrusion, the apicalNa+-HCO3 co-transporter would have to transport anegatively charged species, that is n=2 or 3. The basolateralmembrane contdins Na+IH+ exchange andNa+/HCO3 co-transport (the stochiometry ofwhich is not known), togetherwith Na+-K' ATPase. For details see text (section 4).
1135
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Gleeson
Functional roles of acid base transportsystems
(1) REGULATION OF INTRACELLULAR pH (pHi)In most cells, pHi is actively maintained between7.00 and 7.40, almost one unit above the valueexpected if intracellular and extracellular H+were in electrochemical equilibrium. Further-more, pHi can recover spontaneously to baselinewithin minutes after exposure of cells to acuteacid and alkaline loads.67 Regulation of pHi is ofmajor importance to cell homeostasis becausemany physiological phenomena are pH depen-dent29; diverse examples include the rate ofglycolysis,30 the level of intracellular calcium,3'and plasma membrane K' permeability whichmay regulate intracellular electrical potential.2̀3
In most cells, including several gastro-intestinal cells,3"' the major system regulatingpHi, in the absence of HCO3-, is Na+/H+exchange. Under HCO3- free conditions, block-ing the Na+/H+ exchanger with amiloride orremoval of extracellular Na+ typically causes afall in pHi and inhibits recovery from an intra-cellular acid load (usually achieved by pulseexposure of cells to the weak base ammonia).6 7Rapid recovery of pHi after intracellular acidifi-cation is made possible by a distinctive propertyof the Na+/H+ exchanger, first demonstrated inrenal epithelia by Aronson.42 The exchangercontains a modifier site on its intracellular facewhich is activated by H' ions. Thus, intra-cellular acidosis causes a greater increase in Na+/H' exchange activity than that expected fromthe more favourable in to out H' gradient.3 5738Conversely, the Na+/H+ exchanger is downregulated by intracellular alkalosis and is usuallyinactive above a pHi of 7.20, despite iongradients which favour continued Na+/H+exchange. Hormones and growth factors thatregulate the Na+/H+ exchanger often do soby shifting this inverse relationship betweenactivity and pHi 'up' or 'down.' This shift in pHiresponsiveness may be a result of phosphoryla-tion of the exchanger itself, or of intracellularproteins which regulate its activity.327When cells are maintained under more physio-
logical conditions, where HCO3 and CO2 arepresent, other pHi regulatory systems oftenpredominate over Na+/H+ exchange. Forexample, in hepatocytes,' the intestinal cell lineIEC-6,36 partietal cells,43 and pancreatic acinarcells,4' the major mechanism of pHi recoveryfrom an acid load is not affected by amiloride butis inhibited by DIDS; furthermore, it is depen-dent on the presence ofNa+ and HCO3 but notCl -. These properties are characteristic of aNa+-HCO3 co-transport system. In somecells,363744 pHi recovery from an intracellularalkaline load is mediated by HCO3- extrusionwhich is dependent on the presence of Cl- and isinhibited by DIDS, properties characteristic ofthe Cl -/HCO3- exchanger. This exchanger is insome respects a 'mirror image' of Na+/H+exchange in that it possesses an intracellularmodifier site which is sensitive to hydroxyl(OH -) ions2245 1 and thus, although relativelyinactive at baseline pHi, is activated by intra-cellular alkalosis.
In some cells, Cl-/HCO3- exchange is
coupled to the Na+ gradient, which drives it'uphill,' that is Cl- out of and HCO3- into thecell. Na+ dependent Cl-/HCO3- exchange is animportant pHi regulatory mechanism in inverte-brate cells and in mammalian mesenchymalcells6 7 but has not yet been described in gastro-intestinal epithelia.
(2) GASTRIC ACID SECRETION (FIG 1)Gastric acid secretion is mediated by a H+,K+ATPase on the apical membrane of the parietalcell which extrudes H' in exchange for K+.4The enzyme has recently been cloned andsequenced.3' Its molecular weight is about 95 000Daltons and it consists of two subunits: a largeralpha subunit with several membrane spanningdomains and a smaller beta subunit. The struc-tural, enzymatic, and ion transporting propertiesof gastric H+,K+ ATPase have recently beenreviewed. '
In the resting state, H+,K+ ATPase is local-ised, not on the apical membrane of the parietalcell, but in tubulo-vesicular structures in thecytoplasm which are impermeable to K+ andCl-. Thus, its ability to transport H+ into thesevesicles in exchange for K+ is normally limitedby the availability of intravesicular K+. H+transport into vesicles from resting cells can beobserved however, if these vesicles are preloadedwith K+.`' H+ secretion can also be induced byexposure of resting parietal cells to the K+ionophore valinomycin.52
Stimulation of the parietal cell by secreta-gogues such as histamine, gastrin, and carbacholresults in insertion of these H,K+, ATPasecontaining tubulo-vesicles into the apicalmembrane, the area of which increases severalfold.53 55 The permeability of the apical mem-brane to K+ and Cl- also shows a markedincrease. This is partly due to the appearance ofCl- conductive pathways in the apical mem-brane.5158 These pathways are probably Cl-selective ion channels, as demonstrated recentlyby patch-clamp techniques.59 Whether the apicalmembrane K+ permeability increases via open-ing of separate K+ channels,58 6061 as illustrated inFigure 1, or via activation of a KCI co-transportsystem51 7 iS controversial. It is also unclearwhether the increased apical membrane K+ andCl- permeabilities result from activation oflatent transport systems or, alternatively, frominsertion of a separate class of K+ and Cl-permeable cytoplasmic vesicles into the apicalmembrane.62 Whatever the mechanism, theresult is that: (a) the K+ accumulated by H+ ,K+,ATPase can be recycled into the lumen, so thatH+/K+ exchange is no longer limited by avail-ability of K+; and (b) secretion of Cl- ions canaccompany secretion of H+, thereby preservingelectroneutrality.
Omeprazole,63-5 one of the group of substi-tuted benzimidazoles, is a weak base with a pH of4.0. In its uncharged (unprotonated) form, itdiffuses into acid compartments such as theparietal cell lumen, where it becomes protonatedand positively charged. It is thus trapped in theacid compartment, where it accumulates. Theprotonated omeprazole is then converted to anactive metabolite, the sulphenamide, which
136
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Acid-base transport systems in gastrointestinal epithelia
reacts covalently with cysteine residues on theluminal face of the H+,K+ ATPase alpha unit,resulting in irreversible inhibition of H+,K+ATPase and a parallel inhibition of acidsecretion.'By itself, apical HCl secretion would pro-
gressively alkalinise the parietal cell and depleteit of Cl-. However, parietal cell homeostasisseems to be maintained by modulation of threeother acid base transport systems, all localised onthe basolateral membrane: Cl-IHCO3exchange,346768 which lowers pHi by HCO3extrusion and Na+/H+ exchange,3467 68 and Na+-HCO3 co-transport,4369 both of which act toalkalinise the cell. After stimulation of HCIsecretion by histamine, ClW/HCO3- exchangeacross the basolateral membrane increasesseveral fold70'7; the HCO3- efflux represents theblood 'alkaline tide.' This increase in Cl-/HCO3- exchange can be partly ascribed to morefavourable ion gradients, such as the fall inintracellular [Cl-] resulting from apical HCIexit, but it also reflects a true activation of theexchanger. Any rise in pHi resulting from HCIsecretion should activate Cl-/HCO3- exchangevia its OH- sensitive modifier site72; theexchanger may also be directly activated byintracellular mediators, such as cyclic adenosinemonophosphate (AMP) and calcium, whichactivate acid secretion.7' Simultaneously, there isdownregulation of the basolateral transportsystems which act to alkalinise the cell: Na+-HCO3 co-transport73 and, possibly, Na+/H+exchange7' (although this has been disputed70).The net effect of these various modifications is
to oppose the alkalinising effect of apical HCIsecretion, with the result that patietal cell pHiincreases by less than 010 pH units afterstimulation ofHCI secretion.70`7 The parietal cellthus provides a striking example of how, byselective modulation of apical and basolateralacid-base transport systems, it is possible toreconcile the conflicting demands of varyingtransepithelial transport and maintaining cellhomeostasis.The mechanisms by which secretagogues
exert these multiple effects on the parietal cell areincompletely understood. Carbachol, gastrin,and histamine all bind to specific receptors onparietal cell plasma membranes.50 The effects ofcarbachol and gastrin on acid secretion seem tobe mediated via an increase in intracellularcalcium,7475 achieved partly, as in many celltypes, by inducing inositol phosphate turn-over.75"- Histamine also increases intracellularcalcium,7" by a mechanism independent ofinositol phosphate turnover,75` however, itseffect on acid secretion appears to be mediatedmainly via a rise in intracellular cyclic AMP.79"0Calcium, cyclic AMP, and perhaps other media-tors induce phosphorylation of several intracel-lular proteins,"08' which may result in activationof plasma membrane transport systems.
(3) NaCl ABSORPTION (FIG 2)Electroneutral NaCl absorption is a majordriving force for intestinal fluid absorption.02 07Studies in several species, including humans,have shown that electroneutral Na+ and Cl-
absorption in the ileum,"-0 colon,9' 96 and gallbladder059790 are largely interdependent pro-cesses. Furthermore, they are usually inhibitedby amiloride93-%98 " and the carbonic anhydraseinhibitor acetazolamide,0093 94 9 "I' and also(albeit less consistently) inhibited by HCO3--removal959099101 and by DIDS.94 102 These resultslend support to the model originally proposed byTurnberg on the basis of studies in humans,0whereby Na+ and Cl- are transported across theapical membrane via coupled Na+/H+ and Cl-/HCO3 exchange. More recent studies havedirectly confirmed the presence of both Na+/H+and Cl-/IHCO3- exchangers on the apical mem-branes of gall bladder05 and colonic`5 1621epithelial cells and ileal villus cells'4 '9 (Fig 2).The two exchangers seem to be coupledindirectly via changes in pHi'9; a rise in pHiresulting from stimulation of apical Na+/H+exchange activates apical Cl-IHCO3 exchangevia its OH- sensitive modifier site, as recentlyshown in apical membrane vesicles from ileum-6and colon.22 The Na+ accumulated in the cell isthen extruded in exchange for K' by the baso-lateral Na+,K+ ATPase,'03 and the accumulatedCl- leaves the cell via basolateral anion channelsor in some cells05 via a KCI co-transport system.
Intestinal NaCl absorption is regulated byseveral hormones and neurotransmitters.83 84 It isinhibited by agents that increase intracellularcyclic AMP (for example, cholera toxin, prosta-glandin E2, vasoactive intestinal polypeptide(VIP)), intracellular calcium (serotonin, acetyl-choline, substance P), or intracellular cyclicguanosine monophosphate (GMP) (Escherichiacoli toxin).'03-105 This regulation of NaCl absorp-tion may be achieved largely by modulation ofacid-base transport systems. Cyclic AMP, cyclicGMP, and calcium all inhibit Na+/H+ exchangein isolated enterocytes or in ileal apical mem-brane vesicles.10609 Calcium and cylic AMP alsoinhibit both NaCl absorption and apical mem-brane Na+/H1+ exchange in the gall bladder0 ""as do aldosterone"' 112 and activators of proteinkinase C"13'"4 in the colon. In contrast, gluco-corticoids"5 and alpha adrenergic agonists"6"7increase intestinal NaCl absorption; althoughthe effects of these agents on intestinal Na+/H+exchange are not known, both stimulate renalNa+/H+ exchange."8 "9 Intestinal NaClabsorption is also stimulated by respiratoryacidosis99 01 120 and by exposure to volatile fattyacids,'2' interventions which lower pHi andmight thus activate apical membrane Na+/H+exchange via an intracellular H+ sensitivemodifier site. Indeed, volatile fatty acids haverecently been shown to activate apical membraneNa+/H+ exchange in a human colon cancer cellline.'22 Taken together, these studies providestrong evidence that apical membrane Na+/H+exchange is a major regulated step in NaClabsorption. Much less information is availableregarding hormonal regulation of apical Cl-/HCO3- exchange, however the calcium releas-ing agent serotonin inhibits the exchanger in ilealvillus cells'23 as does cyclic AMP in the gallbladder. "0
Intestinal epithelial cells must also maintainhomeostasis in the face of varying rates oftransepithelial NaCl transport. An increase in
1137
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Gleeson
NaCI absorption initiated by stimulation ofapical Na+/H+ exchange and consequent activa-tion of apical Cl-HCO3- exchange via its OH-sensitive modifier site would result not only insome increase in pHi, but also rises in intra-cellular Na+ and Cl- concentrations, cellvolume, and cell membrane potential. Suchdisturbances in cell homeostasis may, as in theparietal cell, be minimised by simultaneousregulation of apical and basolateral membranetransport systems. In gall bladder'24 andcolonic,'25 epithelia, intracellular [Na+] is main-tained virtually constant despite varying rates ofNaCl absorption, suggesting that there is parellelregulation of basolateral Na+/K+ ATPase. Thebasolateral channels which mediate exit of Cl-and exit of the K' accumulated by Na+/K+ATPase are also regulated in parallel with apicalmembrane transport systems in several absorp-tive epithelia. 126
Intestinal acid-base transport systems may besubject to similar coordinated regulation. Recentobservations in isolated vesicles from jejunal," 127ileal,'28-'32 and colonic'5 133 epithelial cells suggestthat the basolateral membrane also possessesa Na+/H+ exchanger. Furthermore, theapical and basolateral exchangers have differentkinetic characteristics; for example, injejunum'27 and ileum'3 129 131 the apical Na+/H+exchanger is relatively resistant to amiloride. Thisdifference between basolateral and apical Na+/H' exchangers was originally shown on a renalepithelial cell line'34 and may be a general propertyof epithelial cells. 134A It is thus possible that thetwo exchangers have distinct functional roles andare selectively regulated. The apical Na+/H+exchanger might be concerned primarily withtransepithelial transport and respond selectivelyto agents which regulate intestinal NaCl absorp-tion. On the other hand, the basolateral Na+/H+1exchanger might be concerned mainly with cellhomeostasis, contributing to regulation of pHiand, possibly also, cell volume and cell prolifera-tion (see below). The apical and basolateralHCO3 transport systems might also havedistinct characteristics and functional roles.Further evaluation of these possibilities awaitsselective study of apical and basolateral acid-basetransport systems and their regulation in intactNaCl absorbing epithelia.
Malabsorption of NaCl as a result of dysfunc-tion of acid-base transport systems may beimportant in the pathogenesis of diarrhoealdisease. The syndrome of diarrhoea with chlori-dorrhoea and alkalosis, seems to result from anabsence of intestinal Cl /HCO3- exchange.'35 136Other cases of congenital diarrhoea have beenascribed to absence of intestinal apical mem-brane Na+/H+ exchange.'37 In several diseases,including cholera, E coli and amoebic infections,bile acid induced diarrhoea, the carcinoid andWerner Morrison syndromes, and inflammatorybowel disease diarrhoea results from disorderedintestinal electrolyte transport caused by avariety of hormones, toxins, and neurotrans-mitters.83 84 103 138~140 These agents may act viaincreases in enterocyte cyclic AMP (for example,cholera toxin, prostaglandin E2, VIP), cyclicGMP (E coli toxin), or calcium (serotonin,substance P, bile acids). The diarrhoea is partly
mediated by intestinal secretion of Cl- andHCO3- (see below), however, malabsorption ofNaCl, via inhibition of apical membrane Na+/H+exchange and possibly Cl-/HCO3- exchange,also plays a role.
(4) HCO3 SECRETION (FIG 3)HCO3- secretion by the stomach, duodenum,and pancreatic and bile ducts is an importantmeans of neutralising gastric acid and of limitingits damaging effects on the mucosa. BiliaryHCO3 secretion may be an important drivingforce for bile acid independent canalicular bileflow,'4' although whether this secretion originatesfrom hepatocytes or from small bile ductules isunclear. HCO3 secretion by the ileum andcolon is an important cause of watery diar-rhoea.86 103 142
In the stomach and duodenum,143ileum, c9144-146 colon,9192'47-149 and pancreas,'50HCO3- secretion is dependent on the presenceof serosal Na+ and is partly dependent on thepresence of Cl- in the lumen. These findings areconsistent with a model involving Na+ depen-dent HCO3 accumulation by the cell across thebasolateral membrane and HCO3 exit acrossthe apical membrane via Cl -/HCO3 exchange,the accumulated Cl- recycling via apical mem-brane Cl- channels.
Such an asymmetrical distribution of trans-port systems has been shown (using isolatedvesicles) in cells from ileal crypts, which seem tobe the site of HCO3- secretion.'5' The apicalmembrane contains Cl-/HCO3 exchange butnot Na+/H+ exchange. This is in contrast to theapical membrane of the absorptive ileal villuscells, which contains both exchangers'52 (com-pare Figs 2 and 3). The ileal crypt cell basolateralmembrane contains two Na+ dependent mecha-nisms for HCO3 accumulation: Na+-HCO3co-transport25 and Na+/H+ exchange,'52 thelatter accumulating HCO3- indirectly via itsalkalinising effect on the cell. Amiloride onlymodestly inhibits ileal HCO3 secretion,'53suggesting that Na+-HCO3 co-transport maybe the more important of the two mechanisms.
Hepatocytes have a similar distribution of acidbase transport systems: basolateral Na+/H+exchange and Na+-HCO3 co-transport,canalicular membrane Cl-/HCO3- exchangeand Cl- channels.'723 '54 ClV/HCO3- exchangeand Cl- channels are also present on the apicalmembranes of duodenal epithelial cells.'55Finally, both Na+/H+ and Cl/HCO33exchange are present in intact pancreatic'56 andbile'57 ductular cells, although in these cells theirlocalisation remains to be established.
In the duodenum'43 and ileum,8645 amajor component of HCO3- secretion isindependent of luminal Cl- and therefore maynot be mediated by apical Cl-/HCO3- exchange.Possible alternative mechanisms for HCO3 exitacross the apical membrane include HCO3permeable channels8 and Na+-HCO3-co-transport. Na+-HCO3- transport has beendiscussed above as a mechanism for cellularaccumulation ofHCO3-, energised by the out toin Na+ gradient. However, in some epithelia,such as the proximal renal tubule,'59 the Na+-
1138
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Acid-base transport systems in gastrointestinal epithelia
HCO3- co-transporter has a stochiometry ofthree HCO3- ions for each Na+ ion. It thuscarries a net negative charge and normallyoperates in the direction of HCO3- and Na+extrusion, the driving force being the negativeintracellular potential.HCO3 secretion, like NaCl absorption, is
under elaborate neurohormonal control. It isusually stimulated by agents which increaseintracellular cyclic AMP, for example, secretinin the bile and pancreatic ducts and prosta-glandin E2 in the duodenum.'43 Cholera toxininduces severe diarrhoea by stimulating HCO3-and Cl- secretion throughout the intestine, viaincreases in intracellular cyclic AMP.'03 142 160 161
HCO3- secretion predominates in vivo and Cl-secretion in vitro; the reason for this discrepancyis not known. Leukotriones and prostaglandins,often produced in association with intestinalinflammation, also stimulate HCO3 secre-tion. 162 In contrast, HCO3- secretion in theduodenum and ileum is inhibited by alphaadrenergic agonists. 146 158 163
Mediators such as cyclic AMP might induceHCO3 secretion via several mechanisms. Theseinclude activation of apical membrane Cl-recycling by opening of Cl- channels. CyclicAMP causes opening of apical Cl- channels inseveral secretory epithelia. 1' 165 This effect isabsent in patients with cystic fibrosis,'66-'68 whohave impaired pancreatic HCO3- secretion'69and may possibly have a generalised defect ingastrointestinal HCO3 secretion. Cyclic AMPand/or other mediators might also directlyactivate apical HCO3 exit: (a) via Cl-IHCO3exchange, as demonstrated in duodenal apicalmembrane vesicles"10; (b) via HCO3- channels,as in salivary acinar cells8; or (c) via Na'-HCO3-co-transport.
Alternatively, HCO3 secretion may beinduced via activation of basolateral Na+/H+exchange orNa+-HCO3- co-transport, resultingin cellular HCO3- accumulation and therebyincreasing the driving force for HCO3- exitacross the apical membrane. The accompanyingcell alkalinisation should also activate apical Cl -/HCO3- exchange via its OH- sensitive modifiersite.2246 As discussed in the section on NaClabsorption, cyclic AMP and calcium usuallyinhibit Na+/H' exchange in absorptive epithelia.However, recent studies suggest that serotonin,'23which increases intracellular calcium, andforskolin,'7' which increases intracellular cyclicAMP by direct stimulation of adenylate cyclase,both stimulate the basolateral Na+/H' exchangerin ileal crypt cells. The HCO3 secretagoguecarbachol also activates Na+/H+ exchange insalivary acinar cells.172 Yet another mechanismhas been suggested for secretin induction ofHCO3 uptake into bile ductular cells: insertionof cytoplasmic vesicles containing H+ ATPaseinto the basolateral membrane.'73
Evaluation of these possible mechanismsawaits more detailed characterisation of acid-base transport systems in intact HCO3- secret-ing epithelia. However, by analogy with theparietal cell (see section 2), it seems likely thatHCO3- secretion involves parallel regulation ofboth basolateral and apical HCO3- transportsystems, thereby allowing maintenance of a near
constant pHi through varying rates of trans-epithelial HCO3- transport.
Finally, HCO3- secretion may result not fromdirect activation of acid-base transport systemsbut from movement of weak acids in theirprotonated form in the opposite direction, that isfrom lumen to cell. The net result would beconsumption of luminal H' and therefore (in thepresence of carbonic anhydrase), generation ofHCO3 . Ursodeoxycholic acid'74 and certainother bile acids'75 176 induce a 'hyper' choleresis inseveral species, including man,'77 which resultsmainly from active secretion of HCO3 . ThisHCO3- rich choleresis may partly explain theapparently beneficial effects of ursodeoxycholicacid in cholestatic liver diseases,'76 and also itstendency to induce surface gall stone calcifica-tion, one of the factors limiting its efficacy as acholesterol gall stone dissolving agent.'79
Recent studies involving measurement of bothintracellular and canalicular pH in isolated rathepatocyte couplets suggest that ursodeoxycho-lic acid does not directly activate acid basetransport systems in hepatocytes'80 (althoughan effect on bile ductular cells has yet to beexcluded). An alternative hypothesis has beenproposed, based on the observation that these'hyper' choleretic bile acids are excreted by theliver partly in an unconjugated form.5 176 181According to this 'chole-hepatic shunt' hypothe-sis, the unconjugated bile acid is transported inits ionised form across the hepatocyte canalicularmembrane. Unconjugated bile acids have highpKs, and will thus tend to combine with H'in the bile canaliculus, thereby generating aHCO3 ion. The protonated uncharged (andtherefore lipid soluble) bile acid then recyclesacross -the apical membrane of the bile ductularcell, is returned to the liver, redissociates, andthe bile acid anion is re-excreted. In theory, onebile acid molecule could recycle several times,thereby generating several HCO3- ions in thecanaliculus, and this would explain the 'hyper'choleresis. However, each cycle will result inhepatocyte accumulation of one H' ion which,to maintain the generation of biliary HCO3-,must be extruded across the basolateral mem-brane. The HCO3- rich 'hyper' choleresisinduced by ursodeoxycholic acid is partlyinhibited by amiloride and by Na+ removal,'82suggesting that the hepatocyte basolateralNa+/H+ exchanger'7 contributes to this H'extrusion.
(5) ABSORPTION OF OTHER ELECTROLYTES ANDNUTRIENTS; THE ACID MICROENVIRONMENTThe luminal pH near the intestinal apical mem-brane is maintained at a more acid level than bulkluminal pH. The presence of this acid micro-environment was first suggested by studies in the1950s on intestinal absorption of weak acids,183and has been subsequently confirmed bymicroelectrodes in several species, includingman. 184-188Na+/H+ exchange is present on the apical
membranes of jejunal epithelial cells.9'-" Here, itmay function independently of Cl-/HCO3exchange, the presence of which has not beenconvincingly demonstrated in the jejunum.'` A
1139
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Gleeson
role for apical Na+/H+ exchange in maintainingthe acid microenvironment is suggested byobservations that the microenvironment pH isincreased by luminal perfusion with amiloride orwith Na+ free media,'86 189 although other studieshave failed to confirm these findings.'87 Micro-environment pH is also increased by cyclicAMP, cyclic GMP, and activation of proteinkinase C,'90"'' interventions which, as discussedin section (3), inhibit intestinal Na+/H+exchange. In the ileum and colon, microenviron-ment pH is higher than in the jejunum,'84possibly because the apical Na+/H' exchanger islargely neutralised by parallel Cl-IHCO3-exchange (see section 3 and Fig 2).The acid microenvironment contributes to
intestinal absorption of weak acids. It favoursconversion of these weak acids to their proto-nated uncharged form which can then diffuseacross the apical membrane into the cell.'84 Anexample of this process is HCO3- absorption bythe jejunum and gall bladder. In the humanjejunum, Na+ and HCO3- absorption are inter-dependent, are associated with an increase inintraluminal pCO2, and are inhibited by aceta-zolamide.'92 '3 These observations suggest thatHCO3- is not absorbed directly but rather as aresult of H' secretion via apical Na+/H+exchange. The pCO2 of human gall bladderbile exceeds that of hepatic bile and blood,suggesting that the gall bladder may also absorbHCO3 via H' secretion.'9"'96 Gall bladder H'secretion is impaired in patients with calcified(calcium carbonate rich) gall stones; theresult is an abnormally alkaline gall bladderbile that is supersaturated with calciumcarbonate. 11
Volatile fatty acids (VFAs), are produced bybacterial degradation of non-absorbed dietarycarbohydrate and are rapidly absorbed by thecolon, where they constitute a significant energysource. 84 197 198 In several species, includinghumans,'99-202 colonic VFA absorption in vivo isassociated with HCO3- secretion and a lowerluminal pCO2 than in the absence of VFAs.These findings suggest that VFAs combine withluminal H' (thereby depleting CO2 and generat-ing HCO3 ) and are then absorbed passively intheir protonated (uncharged) form. Otherobservations suggest that the acid microenviron-ment, maintained by apical Na+/H+ exchange,also contributes H' to this process and therebyassists in VFA absorption. For example, VFAabsorption is, like the microenvironment pH,'84relatively independent of bulk luminal pH.'97Furthermore, VFA absorption is associated withenhanced colonic Na+ absorption121 99 200 202 andcan be partly inhibited by amiloride202 and byluminal Na+ removal'99 2"3 A third possiblemechanism for VFA absorption is via a VFA-HCO3 exchange mechanism, recently demon-strated on apical membrane vesicles from humanileum2" and rat colon.205The acid microenvironment also plays a role in
intestinal absorption of folic acid20` and oligopep-tides.207 Patients with coeliac disease or Crohn'sdisease have an abnormally alkaline jejunalmicroenvironment,"'820 which may explain thefrequently observed malabsorption of folic acidand other nutrients in these patients.
(6) REGULATION OF CELL VOLUMEWhen exposed to sudden changes in mediumosmolality, cells behave initially as osmometers,swelling in hypo-osmotic media and shrinking inhyperosmotic media. However, cells then tendto return their volume towards normal, despitecontinued exposure to anisotonic media.209Regulatory volume decrease (RVD) andregulatory volume increase (RVI) are typicallyachieved by activation of plasma membrane iontransport systems, which result ultimately in lossor gain of cell electrolytes and water respectively.There are two major mechanisms for RVI in
mammalian cells. The first mechanism is activa-tion of Na+/H+ exchange with coincidental orsecondary (via a rise in pHi) activation of Cl-/HCO3- exchange, resulting in cell accumulationof NaCl. The accumulated Na+ is extruded byNa+/K+ ATPase in exchange for K' and thusthe net effect is cellular accumulation of KCI.This mechanism seems to be the major meansofRIV in gall bladder210 and some renal2' epithe-lial cells, hepatocytes,22 and lymphocytes.23 Theevidence includes observations that RVI in thesecells is dependent on the presence of Na+, Cl,and HCO3- and furthermore, can be inhibitedby amiloride and by DIDS. In addition, activa-tion of Na+/H+ exchange can be demonstratedby measurement of the effects of hyperosmoticstress on pHi.3209213214 The mechanism(s) ofactivation remain obscure but may involve phos-phorylation of the Na+/H+ exchanger.3The other major mechanism of RVI uses the
out to in Na+ gradient to drive K' and Cl- intothe cell.219 This electroneutral Na+-K-2Cl-co-transport system is present on the basolateralmembranes of intestinal epithelial cells`4 and canbe inhibited by the diuretic bumetanide. It is thissystem, rather than Na+/H+ and Cl-/HCO3-exchange, which seems to mediate RVI in jejunalvillus enterocytes.2' The mechanisms of RVI inother intestinal epithelial cells are not known.
Regulatory volume decrease (RVD) after cellswelling in hypo-osmotic media is mediated byKCI loss, for which there are two main mecha-nisms. The first involves opening of K' and Cl-channels and mediates RVD in hepatocytes2"and jejunal villus enterocytes.26 Other cells maylose KCI via activation of a bumetanide inhibit-ible KCI co-transport system.29The above cell volume regulatory mechanisms
have usually been demonstrated by suddenlyexposing cells to large changes in mediumosmolality. It is not clear to what extent thesemechanisms are operative in vivo, where extra-cellular fluid osmolality is usually regulatedwithin a narrow range. However, for epithelialcells to maintain their volume through varyingrates of transepithelial transport, net ion fluxesacross the apical membrane must balance thoseacross the basolateral membrane. Therefore, cellvolume regulatory mechanisms might also play amajor role in balancing ion fluxes across the twomembranes. Indeed, in some epithelia, theopening of basolateral membrane K' and Cl-channels after an increase in Na+ absorptionacross the apical membrane is mediated partlyvia the resulting increase in cell volume. 16Conversely, carbachol induced stimulation ofHCO3- secretion by salivary acinar cells results
1140
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Acid-base transport systems in gastrointestinal epithelia
in activation of basolateral Na+/H+ exchange,which is mediated partly via a decrease in cellvolume,'72 presumably a consequence of apicalHCO3 and water exit.
(7) REGULATION OF CELL PROLIFERATIONOne of the most intriguing and controversialaspects of acid-base transport systems is theirpossible role in the regulation of cellular pro-liferation. In white blood cells, fibroblasts andin various invertebrate cells, several agentswhich induce cell proliferation activate Na/H+exchange5 217; the exchanger is also spontane-ously activated in some tumour cell lines.Furthermore, exposure of cells to amiloride orremoval of Na+ often inhibits their prolifera-tion.5 Based on these observations, it was sug-gested that a rise in pHi resulting from activationof Na+/H+ exchange was a trigger for initiationof cell division.
However, most studies on the effects of mito-gens on pHi have been performed in HCO3 freemedia. Under more physiological conditions,when HCO3 is present, mitogens activate notonly Na+/H+ exchange but also other transportsystems such as Cl-/HCO3 exchange, which insome cells,2829 minimises the changes in pHi.Nevertheless, it is likely that activation of Na+/H' exchange and also the HCO3- transportsystems play a permissive role in cell prolifera-tion by maintaining pHi at a level which willpermit DNA and protein synthesis; these pro-cesses often have pH optima above basal pHi andare inhibited by intracellular acidosis. Indeed, insome cells, simply raising pHi induces prolifera-tion, as was demonstrated recently in fibroblastsby inserting and inducing expression of the genefor H' ATPase.22' It is also possible that acidbase transport systems regulate cell proliferationnot via changes in pHi but via secondary effectson intracellular Na+ or Cl- concentrations or cellvolume.
Gastrointestinal epithelial cell proliferation isinfluenced by many hormonal, paracrine, andintraluminal agents.22' It is still unclear whetheracid-base transport systems are involved inregulating proliferation. However, Na+/H+exchange activity is enhanced in several modelsof increased gastrointestinal cell proliferation,including the neonatal liver,222 the pancreas afterexposure to trophic hormones,223 and theintestine after partial resection224 or exposure tothe carcinogen 1,2 dimethylhydrazine.225 In con-trast, differentiation and cessation of prolifera-tion in some human colon cancer cell lines isassociated with down-regulation of Na+/H+exchange.226 227 Finally, blocking Na+/H+exchange with amiloride or its analogues inhibitsproliferation of a pancreatic cell line and of theliver and jejunum after partial intestinal resec-tion.228229 Further studies in this area will be ofgreat interest.
ConclusionGastrointestinal epithelial cells possess a varietyof acid-base transport systems. The location ofthese transport systems is often highly polarisedbetween the apical and basolateral plasma mem-
branes. They are subject to regulation by manyneural, hormonal, and paracrine factors, actingvia intracellular mediators. There is someevidence that different transport systems, andeven similar transport systems on the apicaland basolateral membranes, are selectivelyregulated. These distinctive properties maypartly explain how acid-base transport systemscan play several major physiological roles,including regulation of pHi and cell volume,transepithelial transport, and, perhaps, cell pro-liferation - roles which might at first seemmutually incompatible.Our understanding of these systems is, how-
ever, still incomplete. For example, the role ofcoordinated regulation of apical and basolateralacid base transport systems in mediatingintestinal NaCl absorption and HCO3 secretionneeds to be systematically evaluated. Thisevaluation will require selective assay of acidbase transport systems on the apical and baso-lateral membranes of polarised gastrointestinalepithelial cells. Such studies have proveddifficult because most epithelia are heterogenousand therefore, measurement of pHi using fluor-escent dyes in one cell type has usually requiredselective cell isolation, following which the cell'spolarised characteristics are often lost.
Recent advances in methodology offer severalpossible solutions to this problem. Firstly,culture of some epithelial cell lines to formmonolayers on semipermeable membranes hasresulted in retention of cell polarity and hasenabled selective study of the apical and baso-lateral membranes of renal'34230 and gastro-intestinal381 22 cell monolayers. Secondly, assay ofacid base transport systems in one cell type in anintact heterogeneous epithelium can sometimesbe achieved by selective loading of pH sensitivedye into that cell type,23' or alternatively by use ofsingle cell fluorescence microscopy.
Yet another advance has been the applicationofmolecular biological techniques to the study ofgastrointestinal acid-base transport systems. Tseet al'3''32 have recently obtained two related butdistinct cDNA sequences from rabbit ileum,which induce expression of Na+/H+ activity ina fibroblast cell line that lacks intrinsic Na+/H+exchange. The first of these putative Na+/H+exchangers has 95% homology with the pre-viously cloned human fibroblast Na+/H+exchanger,262" with which it also shares severalfunctional characteristics, including extremesensitivity to amiloride. Furthermore, immuno-cytochemical studies showed that polyclonalantibodies to a fusion protein incorporating thissequence localise to basolateral but not apicalmembranes from rabbit ileum. The Na+/H+encoded by the second cDNA sequence has beenless extensively characterised but has someproperties common to those of the ileal apicalmembrane Na+/H+ exchanger, including arelative resistance to amiloride. 31 32 Detailedstudies on the neurohormonal regulation of thesetwo putative Na+/H+ exchangers should now bepossible and are awaited with interest. Progresshas also been made in cloning the ileal Cl -/HCO3 exchanger.232Approaches such as these should lead to
further major advances in our understanding of
1 141
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

1142 Gleeson
these fascinating and versatile transport systemsover the next decade.I am grateful to the Wellcome Trust for financial support and toDr Martin Steward, Department of Physiological Sciences,University of Manchester for critical comment on an earlier draftof the manuscript.
1 Rabon EC, Reuben MA. rhe mechanism and structure of the gastricH,-K ATPase. Ann Rev Phvsiol 1990; 52: 321-44.
2 Kaunitz JD, Sachs G. Identification of a vanadate sensitive K+dependent proton pump from rabbit colon. 7 Biol Chem 1986; 261:14005-10.
3 Grinstein S, Rothstein A. Mechanisms of regulation of the Na/Hexchanger. J7 Membr Biol 1986; 90: 1-12.
4 Aronson P'S. Kinetic properties of the plasma membrane Na+/H+exchanger. Ann Rev Plhysiol 1985; 47: 545-60.
5 Grinstein S, Rotin D, Mason MJ. Na+/H+ exchange and growth factorinduced cytosolic pH changes. Role in cellular proliferation. BiochimBiophvs Acta 1989; 988: 73-97.
6 Roos A, Boron WF. Intracellular pH. Physiol Rev 1981; 61: 296-432.7 Madshus IH. Regulation of intracellular pH in eukaryotic cells.
Biochem,J 1988; 250: 1-8.8 Lau KR, Elliott AC, Brown PD. Acetylcholine-induced intracellular
acidosis in rabbit salivary gland acinar cells. Am j Phvsiol 1989; 256:C288-95.
9 Murer H, Hopfer U, Kinne R. Sodium/proton antiport in brush bordermembrane vesicles isolated from rabbit small intestine and kidney.Biochemj 1976; 154: 597-604.
10 Cassano G, Steiger B, Murer H. Na/H and Cl/HCO3 exchange in ratjejunal and rat proximal tubular brush border membrane vesicles.Studies with acridine orange. Pflugers Arch 1984; 400: 309-17.
11 Kleinman JG, Harig JM, Barry JA, Ramaswamy K. Na+ and H+transport in human jejunal brush border membrane vesicles.AmJ7 Phvsiol 1988; 255: G206-1 1.
12 Kichuchi K, Alumrad NN, Ghishan FK. Na+/H+ exchange by brushborder membrane sesicles of human ileum. Gastroenterology 1988; 95:388-93.
13 Ramaswamy K, Harig JM, Kleinman 1G, Harris MS, Barrv JA.Sodium-proton exchange in human ileal brush-border membranevesicles. Biochim Biophys Acta 1989; 981: 193-9.
14 Knickelbein R, Aronson PS, Atherton W, Dobbins JW. Sodium andchloride transport across rabbit ileal brush border. 1. Evidence forNa+/H + exchange. Am3' Physiol 1983; 245: G504-10.
15 Foster ES, Dudeja PK, Brasitus TA. Na+/H+ exchange in rat colonicbrush border apical membrane vesicles. Am J Physiol 1986; 250:G781-7.
16 Rajendran VM, Binder HJ. Characterisation of Na-H exchange in apicalmembrane vesicles of rat colon. J7 Biol Chem 1990; 265: 8408-14.
17 Moseley RH, Meier PJ, Aronson PS, Boyer JL. Na-H exchange in ratliver basolateral but not canalicular plasma membrane vesicles.AmJ Physiol 1986; 250: G35-43.
18 Liedtke CM, Hopfer U. Mechanism of Cl- translocation across smallintestine brush-border membrane. Demonstration of Cl-OH-exchange and Cl- conductance. AmJI Physiol 1982; 242: G272-80.
19 Knickelbein R, Aronson PS, Sehron CM, Seifter J, Dobbins JW.Sodium chloride transport across rabbit ileal brush border. II.Evidence for Cl-HCO3 exchange and mechanism of coupling.Amj Phyvsiol 1985; 249: G236-45.
20 Ramaswamy K, Chung M, Barry IA. Chloride/Bicarbonate exchange inhuman ileal brush border membrane vesicles. Gastroenterology 1988;94: A366.
21 Foster ES, Dudeja PK, Brasitus TA. Cl-OH exchange in rat distalcolon. Amj Physiol 1990; 258: G261-7.
22 Rajendran VM, Osterlin M, Binder HJ. pH-regulated Cl uptake inapical membrane vesicles of rat distal colon. Gastroenterology 1991;100: A700.
23 Meier PJ, Knickelbein RG, Moseley RH, Dobbins JW, Boyer JL.Evidence for carrier mediated chloride/bicarbonate exchange incanalicular rat liver plasma membrane vesicles.J7 Clin Invest 1985; 75:1256-63.
24 Rajendran VM, Oesterlin M, Binder HJ. Sodium uptake across baso-lateral membrane of rat distal colon. Evidence for Na-H exchange andNa anion cotransport.J Clin Invest 1991; 88: 1379-85.
25 Zamir Z, Barry IA, Ramaswamy K. Sodium transport in humanintestinal basolateral membrane vesicles. Gastroenterology 1991; 100:A710.
26 Sardet C, Franchi A, Pouyssegur J. Molecular cloning, primarystructure and expression of the human growth factor activatibleNa+/H+ antiporter. Cell 1989; 56: 271-80.
27 Sardet C, Counillon L, Franchi A, Pouyssegur J. Growth factorsinduce phosphorylation of the Na+/H+ antiporter, a glycoproteinof 110 kilodaltons. Science 1990; 247: 723-6.
28 Kopito RR, Lodish HF. Primary structure and transmembrane orienta-tion of the murine anion exchange protein. Nature 1985; 316: 234-8.
29 Busa WB. Mechanisms and consequences of pH mediated cell regula-tion. Ann Rev Physiol 1986; 48: 389-402.
30 Fidelman ML, Seeholzer SH, Walsh KB, Moore RD. Intracellular pHmediates action of insulin on glycolysis in frog skeletal muscle.AmJ Physiol 1982; 242: C87-93.
31 Siskind MS, McCoy CE, Chobanian A, Schwartz JH. Regulation ofintracellular calcium by cell pH in vascular smooth muscle cells.AmJ Physiol 1989; 256: C234-40.
32 Duffy ME, Dewar DC. IntracellularpH and membrane K+ conductancein rabbit distal colon. AmJ Physiol 1990; 258: C336-43.
33 Henderson RM, Krumpholtz BF, Boyer JL, Graf I- Effect of intra-cellular pH on potassium conductance in liver. Pflugers Arch 1988;412: 334-5.
34 Paradiso AM, Tsien RY, Demarest JR, Machen TE. Na/H andC1/HCO3 exchange in rabbit oxyntic cells using fluorescencemicroscopy. AmJf Physiol 1987; 253: C30-6.
35 Giore I, Hoinard C. Na±+/H±+ exchange in isolated hamster enterocvytes.Its major role in intracellular pH regulation. Gastroenterology 1989; 97:882-7.
36 Wenzl E, Sjaastad MD, Weintraub WH, Machen TE. Intracellular pHregulation in IEC-6 cells, a crvptlike intestinal cell line. Am ] Phs'siol1989; 257: G732-40.
37 Sundaram U, Knickelbein RL, Dobbins JW. Intracellular pH regula-tion in the ileum: Na+/H+ and Cl-/HCO3- exchange in isolatedcrypt and villus cells. Am] Phvsiol 1991; 260: G440-9.
38 Watson AJM, Levine S, Donowitz M, Montrose MH. Kinetics andregulation of a polarized Na+-H+ exchanger from Caco-2 cells, ahuman intestinal cell line. Amj Phvsiol 1991; 261: G229-38.
39 Renner EL, Lake JR, Persico M, Scharschmidt BF. Na+/H+ exchangcactivity in rat hepatocytes; role in regulation of intracellular pH.Am] Phvsiol 1989; 256: G44-52.
40 Gleeson D, Smith ND, Boyer JL. Bicarbonate dependent and indepen-dent intracellular pH regulatory mechanisms in rat hepatocytes.Evidence for Na-HCO3 cotransport. 7 Clin Invest 1989; 84: 312-21.
41 Muallem S, Lalsbeg P. Intracellular pH regulatory mechanisms inpancreatic acinar cells. J. Characteristics of H+ and HCO3-transporters. ] Biol Chem 1990; 265: 12806-12.
42 Aronson PS, Nee J, Suhm MA. Modifier role of internal H + inactivating the Na+-H+ exchanger in renal microvillus membranevesicles. Nature 1982; 229: 161-3.
43 Townslev MC, Machen TE. Na-HCO3 cotransport in rabbit parietalcells. Amj Phvsiol 1989; 257: G350-6.
44 Benedetti A, Strazzabosco M, Boyer JL, Haddad P, Graf J, Boyen J.Cl /HCO3 exchange in rat hepatocytes: role in regulation of intra-cellular pH. AmjPhysiol 1991; 261: G512-21.
45 Mason MJ, Smith JD, Garcia-Soto JdeJ, Grinstein S. Internal pHsensitive site couples Cl-/HCO3- exchange to Na+/H+ antiport inlymphocytes. Amj Physiol 1989; 256: C428-33.
46 Mugharbil A, Knickelbein R, Aronson PS, Dobbins JW. Rabbit ilealbrush border membrane Cl-/HCO3- exchange is modulated by aninternal pH sensitive modifier site. Am J Phvsiol 1990; 259:G666-70.
47 Sachs G, Chang HH, Rabon E, Schachman R, Lewin M, Saccarani G. Anon electrogenic H+ pump in parietal membranes of hog stomach.J]Biol Chem 1976; 255: G313-8.
48 Forte JG, Machen TE, Obrink KJ. Mechanisms of gastric H + and Cl -transport. Ann Rev Phvsiol 1980; 42: 111-26.
49 Sachs G. The parietal cell as a therapeutic target. Scand]7 Gastroenlerol1986; 21 (suppl 1 18): 1-10.
50 Wolfe MM, Soll AH. The physiology of gastric acid secretion. N EngiJMed 1988; 319: 1707-15.
51 Shull GE, Lingrel JB. Molecular cloning of the rat stomach (H +,K+)ATPase..JBiol Chem 1986; 261: 16788-91.
52 Hershey SJ, Steiner L. Stimulation of acid formation in isolated gastricglands by valinomycin. Am] Physiol 1988; 255: G313-8.
53 Ito S. Functional gastric morphology. In: Johnson LR, ed. Phvsiologv ofthe gastrointestinal tract. New York: Raven Press, 1987: 817-51.
54 Forte JG, Forte TM, Black JA, Okamoto C, Wolosin JM. Ultra-structural changes related to functional activity in gastric osxyntic cells.Am] Physiol 1981; 241: G349-58.
55 Smolka A, Helander HF, Sachs G. Monoclonal antibodies against gastricH+,K+-ATPase. AmjPhysiol 1983; 245: G589-96.
56 Perez A, Blissard D, Sachs G, Hershey SJ. Evidence for a chlorideconductance in the secretory membrane of the parietal cell.Am] Physiol 1989; 256: G299-305.
57 Cuppoletti J, Sachs G. Regulation of gastric acid secretion via modula-tion of a chloride conductance. ] Biol Chem 1984; 259: 14952-9.
58 Reenstra WW, Forte JG. Characterisation ofK+ and Cl- conductancesin apical membrane vesicles from stimulated rabbit oxyntic cells.Am] Ptvsiol 1990; 259: G850-8.
59 Saccomani G, Sarnas CGP, Smith PR, Kirk KL, Shoemaker RL.Histamine induced Cl- channels in apical membranes of isolatedrabbit cells. Am]Physiol 1991; 26: C1OOO-l 1.
60 Wolosin JM, Forte JG. Stimulation of oxyntic cell triggers K + and Cl -conductances in the apical H + ,K + ATPase membrane. Am]J Phvsiol1984; 246: C537-45.
61 Gunther RD, Bassilian S, Rabon EC. Cation transport in vesicles fromsecreting rabbit stomach.J Biol Chem 1987; 262: 13966-72.
62 Im WB, Blackman DP, Davis JP. Studies on K+ permeability of ratgastric microsomes. JBiol Chem 1985; 260: 9452-60.
63 Sachs G, Carlsson E, Lindberg P, Wallmark B. Gastric H K ATPase asa therapeutic target. Ann Rev Pharmacol Toxicol 1988; 28: 269-84.
64 Maton PN. Omeprazole. N Engli Med 1991; 324: 965-75.65 Wallmark B. Mechanism of action of Omeprazole. Scand] Gastro 1986;
21 (suppl 118): 11-7.66 Wallmark B, Larsson H, Humble L. The relationship between gastric
acid secretion and gastric H+ ,K + ATPase activity.J Biol Chem 1985;260: 13681-4.
67 Muallem S, Burnham C, Blissard D, Berglindh T, Sachs G. Electrolytetransport across the basolateral membrane of parietal cells.J Biol Chem 1985; 260: 6641-53.
68 Machen TE, Paradiso AM. Regulation of intracellular pH in thestomach. Ann Rev Physiol 1987; 49: 19-33.
69 Curci S, Debellis L, Fromter E. Evidence for rheogenic sodiumbicarbonate cotransport in the basolateral membrane of oxyntic cellsof frog gastric fundus. Pflugers Arch 1987; 408: 497-504.
70 Muallem S, Blissard D, Cragoe EE, SachsG. Activation of Na+/H+ andCl-/HCO3- exchange by stimulation of acid secretion in the parietalcell. ] Biol Chem 1988; 263: 14703-11.
71 Paradiso AM, Townsley MC, Wenzl E, Machen TE. Regulationof intracellular pH in resting and in stimulated parietal cells.Am] Physiol 1988; 257: C554-61.
72 Wenzl E, Machen TE. Intracellular pH dependence of buffer capacityand anion exchange in the parietal cell. Am] IPhvsiol 1989; 257:G741-7.
73 Townsley MC, Machen TE. Na/HCO3 cotransport in oxyrntic cells isinhibited by histamine. FASE:B]E 1988; 2: A752.
74 Negulescu P, Reenstra WW, Machen TE. Intracellular calcium require-ments for stimulus-secretion coupling in parietal cells. Am]7 Phvsiol1989; 256: C241-51.
75 Chew CS, Brown MR. Release of Ca2+ and eles-ation oSf inossitoltriphosphate byr secretagognes in parietal and chief cells isolated fromrabbit gastric mucosa. Biochim Biophvs Acta 1986; 888: 1 16-25.
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Acid-base transport systems in gastrointestinal epithelia 1143
76 Puurtinen J, Schwabe U. Effect of gastric secretagogues on the formationof inositol phosphates in isolated gastric cells of the rat. BrJ7 Pharmtacol1987; 90: 479-90.
77 Chiba r, Fisher SK, Park J, Seguin EB, Agranoff BW, Yamada 'T.Carbamylcholine and gastrin induce inositol lipid turnover in caninegastric parietal cells. AmJ Phvsiol 1988; 255: G99-105.
78 Ljungstrom M, Chew CS. Calcium oscillations and morphologicaltransformations in single cultured gastric parietal cells. AmJI Physiol1991; 260: C67-78.
79 Soll AH, Wollin A. Histamine and cyclic AMP in isolated canine parietalcells. AmJ Phvsiol 1979; 237: E444-52.
80 Brown MR, Chew CS. Carbachol-induced phosphorylation in parietalcells: regulation by [Ca2 + J. AmJ7 Phvsiol 1989; 257: G99-1 10.
81 Urushidani T, Hanzel DK, Forte JG. Protein phosphorylation associ-ated with stimulation of gastric glands. Biochim BRiophvs Acta 1987;930: 209-19.
82 Hopfer U, Liedtke CM. Proton and bicarbonate transport mechanismsin the intestine. Ann Rev Phvsiol 1987; 49: 51-67.
83 Binder HJ. Absorption and secretion of water and electrolytes bysmall and large intestine. In: Sleisenger MH, Fordtran JS, eds.Gastrointestinal disease: pathop/vsiology, diagnosis, management.Philadelphia: W B Saunders, 1988: 1022-44.
84 Field M, Rao MC, Chang EB. Intestinal electrolyte transport anddiarrhoeal disease. N Engl3 Med 1989; 321: 800-6, 879-83.
85 Reuss L. Ion transport across gallbladder epithelium. Phlvsiol Rev 1989;69: 503-45.
86 Sullivan SK, Field M. Ion transport across mammalian small intestine.In: Schultz SG, Field M, Frizzell RA, eds. Handbook of p/vsiologv.Section 6. The gastrointestinal system. Vol IV. Intestinal absorptionand secretion. New York: Oxford University Press, 1991: 287-301.
87 Halm DR, Frizzell RA. Ion transport across the large intestine. In:Schultz SG, Field M, Frizzell RA, eds. Handbook of Physiology.Section 6. The gastrointestinal system. Vol IV. Intestinal absorptionand secretion. New York: Oxford University Press, 1991: 257-73.
88 Turnberg LA, Bieberdorf FA, Morawski SG, Fordtran JS. Inter-relationships of chloride, bicarbonate, sodium and hydrogen transportin the human ileum. J Clin Invest 1970; 49: 557-67.
89 Podesta RB, Mettrick DF. HCO3- and H+ secretion in rat ileum invivo. Aml Phyvsiol 1977; 232: E574-9.
90 Nellans HN, Frizzell RA, Schultz SG. Brush border processes andtransepithelial Na and Cl transport by rabbit ileum. Am J Phvsiol1974;226: 1131-41.
91 Davis GR, Morawski SG, Santa Anna CA, Fordtran JS. Evaluation ofchloride/bicarbonate exchange in the human colon in vivo.J Clin Invest 1983; 71: 201-7.
92 Deverode GJ, Phillips SF. Conservation of sodium, chloride and waterby the human colon. Gastroenterology 1969; 56: 101-9.
93 Binder HJ, Foster ES, Budinger ME, Hayslett JP. Mechanism ofelectroneutral sodium-chloride absorption in the distal colon of therat. Gastroenterology 1987; 93: 449-56.
94 Lubke R, Haag K, Berger E, Knauf H, Gerok W. Ion transport in ratproximal colon in vivo. AmJ Phvsiol 1986; 256: G132-9.
95 Foster ES, Budinger ME, Hayslett JP, Binder HJ. Ion transport in theproximal colon of the rat. Sodium depletion stimulates neutral sodiumchloride absorption.JI Clin Invest 1986; 77: 228-35.
96 Sellin JH, DeSoignie R. Rabbit proximal colon: a distinct transportepithelium. AmJ Phvsiol 1984; 246: G603-10.
97 Frizzell RA, Dugas MC, Schultz SG. Sodium chloride transport byrabbit gallbladder. Direct evidence for a coupled NaCl influx process.J Gen Physiol 1975; 65: 769-95.
98 O'Grady SM, Wolters PJ. Sodium and chloride transport across isolatedporcine gallbladder. Am3' Phvsiol 1989; 257: C45-5 1.
99 Goldfarb DS, Egnor RW, Charney AN. Effect of acid base variables onion transport in rat colon. J Clin Invest 1988; 81: 1903-10.
100 Nellans HN, Frizzell RA, Schultz SG. Effect of acetazolamide onsodium and chloride transport by in vitro rabbit ileum. AmJI Phvsiol1975; 228: 1808-14.
101 DeSoigne R, Sellin JH. Acid base regulation of ion transport in rabbitileum in vitro. Gastroenterology 1990; 99: 132-41.
102 Hatch M, Freel RW, Goldner AM, Earnest DL. Oxalate and chlorideabsorption by the rabbit colon; sensitivity to metabolic and aniontransport inhibitors. Gut 1984; 25: 232-7.
103 Field M. Secretion of water and electrolytes by mammalian smallintestine. In: Johnson LR, ed. Physiology of the gastrointestinal tract.New York: Raven Press, 1981: 963-82.
104 Donowitz M, Welsh MJ. Ca+ + and cyclic AMP in regulation ofintestinal Na, K and Cl transport. Ann Rev Phvsiol 1986; 48: 135-50.
105 Guandalini S, Rao MC, Smith PL, Field M. cGMP modulation ofileal ion transport: in vitro effects of Escherichia coli heat stableenterotoxin. Am J Physiol 1982; 243: G36-41.
106 Semrad CE, Chang EB. Calcium mediated cAMP inhibition ofNa +/H +exchange in isolated chick enterocytes. Am J7 Physiol 1987; 252:C315-22.
107 Hirose R, Chang EB. Effects of serotonin on Na+/H+ exchange andintracellular calcium in isolated chick ettterocvtes. Am7 Phvsiol 1988;254: G891-7.
108 Semrad CE, Cragoe EJ Jr, Chang EB. Inhibition of Na/H exchange inavian intestine by atrial natiuretic factor. J Clin Invest 1990; 86:585-9 1.
109 Emmer E, Rood RP, Wesolek JH, ei al. Role of calcium and calmodulinin the regulation of the rabbit ileal brush border membrane Na + /H +antiporter. J7 Membr Biol 1989; 108: 207-15.
110 Reuss L, Segal Y, Altenberg G. Regulation of ion transport acrossgallbladder epithelium. Ann Rev Phvsiol 1991; 53: 361-73.
Ill Foster ES, Zimmerman TW, Havslett JP, Binder HJ. Corticostertsidalteration of active electrolyte transport in rat distal ctolsn.AmJ7 Physiol 1983; 245: G668-75.
112 Rajendran VM, Kashgarian M, Binder HI. Aldosterone indtlction olfelectrogenic stsdium transport in apical membrane sresicles tsf rat distalcolon. J Bio/ (,hem 1989; 264: 18638-44.
113 Dtnowitz M, Cheng HY, Sharp GW. Effects tof phorbol esters tsnsodium and chloride transport in rat cslon. A^m .7 Pstysiol 1986; 251:(i509- 1 7.
114 Ahn I, Chang EC, Field M\. I'hosrbol ester inhibition of Na/H exchangein rabbit proximal colon. .Am]7 tsPsiss/ 1985; 249: C527-31).
115 Turmanian SG, Binder HJ. Aldosteronc and glucocorticoid rccptor-specific agonists regulatc ioni transport in rat proximal colon.Am] Phvsiol 1990; 258: G492-8.
116 Chang EB, Field M, Miller RJ. Alpha2 adrenergic receptor regulation ofion transport in rabbit ileum. Am' Phtsvisol 1982; 242: G237-42.
117 Sellin JH, DeSoigne R. Regulation of Na-Cl absorption in rabbitproximal colon in vitro. Am]' Phvsiol 1987; 252: G45-5 1.
118 Kinsella JL, Freiberg JM, Sacktor B. (ilucocorticoid activation oftNa t/H+ exchange in renal bruish border sesicles; kinetic effects.Am]' Phvsiol 1985; 248: F233-9.
119 Nord EP, Howard MIJ, Hafezi A, Moradehagi P, Zavjstub S, Insel P'A.Alpha2 adrenergic agonists stimulate Na/H antiport activity in therabbit renal proximal tubule. ] Clin Invest 1987; 80: 1755-62.
120 Vacarezza SC, Charnev AN. Acid base effects on ileal sodiumchloride absorption in vitro. Am]7 Phtsiol 1988; 254: G329-33.
121 Binder HJ, Mehta P. Short chain fatty acids stimulate active soditim andchloride absorption in vitro in the rat distal colon. Gastroenterologs1989; 96: 989-96.
122 Montrose MH. Transport of short chain fatty acids in cultured huimancolon epithelial cells. Gastroenterologv 1990; 98: A549.
123 Sundaram U, Knickelbein RG, Dobbins JW. Mechanism of intestinalsecretion. Effect of serotonin on rabbit ileal crypt and sillus ccils.J Clin Invest 1991; 87: 743-6.
124 Moran WM, Hudson RL, Schultz SG. TIranscellular Na absorption andintracellular Na activities in rabbit gallbladder. Am 7 Phtvsiol 1986;251: G155-9.
125 Turnheim K, Hudson RL, Schultz S(i. Cell Na+ actisities andtranscellular Na+ absorption by descending colon from normal andNa+ deprived rabbits. Pflugers Arch 1987; 410: 279-83.
126 Dawson DC, Richards NW. Basolateral K conductance: rolc in regula-tion of NaCI absorption and secretion. Am ] PhYtsiol 19911; 259:C181-95.
127 Acra SA, Dykes W, Nylander W, Ghishan FK. Characterisation of adistinct Na+/H + exchanger in the basolateral membranes of humanjejunum. Gasiroenterologv 1991; 100: A678.
128 Barros F, Dominguez P, Velasco G, Lazo PS. Na'/H' exchange ispresent in the basolateral membranes from rabbit small intestine.Biochem Biophvs Res Commun 1986; 134: 827-34.
129 Knickelbein RG, Aronson PS, Dobbins JW. Characterisatiotl ofNa+/H t exchangers on villus cells in rabbit ileum. Am 7 P/htsiol1990; 259: G802-6.
130 Ma Al, Tse CM, Watson AJM, et al. Expression and extracellularregulation of the rabbit ilea! villus basolateral membrane Na + /H +exchanger in a fibroblast. Gastroenterologv 1991; 100: A695.
131 lIse CM, Watson AJM, Ma Al, Pouyssegur J, Donowitz M. Cloning andfunctional expression of a second novel rabbit ileal villus epithelial cellNa+/H+ exchanger. Gastroenterologs% 1991;100: A258.
132 Tse CM, Ma Al, Yang VW, et al. Molecular cloning and expression ofa cDNA encoding the rabbit ileal villus basolateral membrane Na/Hexchanger. EMBO] 1991; 10: 1957-67.
133 Dude'ja PK, Foster ES, Brasitus TA. Na + -H + antiporter of rat colonicbasolateral membrane sesicles. Am] Phtvsiol 1989; 257: G624-32.
134 Haggerty JG, Agarwal N, Reilly RF, Adelberg A, Slayman C\X'.Pharmacologicallv different Na/H antiporters on the apical andbasolateral surfaces of cultured porcine kidney cells (LLC-IPKProc Natl Acad Sci USA. 1988; 85: 6797-801.
134A Clark DJ, Limbird LE. Na-H' exchange subtypes: a predictivereview. Am] Physiol 1991; 261: C945-53.
135 Turnberg LA. Abnormalities in intestinal electrolyte transport incongenital chloridorrhoea. Gut 1971; 12: 5-44-50.
136 Bieberdorf FA, Gorden P, Fordtran JS. Pathogenesis of conlgenitalalkalosis with diarrhoea. Implications for the physiology of normalileal electrolyte absorption and secretion. ] Clin Invest 1972; 51:1958-68.
137 Booth IW, Strange G, Murer H, Fenton FR, Milla P. Defective jeptinalbrush border Na+/H+ exchange: a cause of congenital secretorydiarrhoea. Lanceg 1985; i: 1066-9.
138 Fondacaro J. Intestinal ion transport and diarrhoeal disease.Am ] Physiol 1986; 250: G1-8.
139 McGowan K, Piver G, Stoff JS, Donowitz M. Role of prostaglandins andcalcium in the effects of Entamoeba Histolvtica on colonic electrolytctransport. Gastroenterologv 1990; 98: 873-80.
140 Dharmsathaphorn K, Huott PA, Vongkovit P, Beuerlein G, Pandol SI,Ammon HV. Cl - secretion by bile salts. A study of the mechanism ottaction based on a cultured colonic epithelial cell line. 7 C/int Invest1989; 84: 945-53.
141 Hardison WGM, Wood CA. Importance of bicarbonate in the bile acidindependent fraction of bile flow. Am]7 Phvsiol 1978; 285: E158-64.
142 Moore WL Jr, Bieberdorf FA, Morawski SG, Finkelstein RA, FordtranJS. Ion transport during cholera induced ileal secretion on the dtog.7 Clin Invest 1971; 50: 312-8.
143 Flemstrom G, Garner A. Gastroduodenal HCO3 transport: character-istics and proposed role in acidity regulation and mucosal protection.Am ] Phvsiol 1982; 242: G183-93.
144 Hubel KA. Effect of luminal chloride concentration on bicarbonatcsecretion in rat ileum. Am] Phvsiol 1969; 217: 40-5.
145 Smith PL, Cascairo MA, Sullivan SK. Sodium dependence of luminalalkalinisation by rabbit ileal mucosa. Am ] Phvsiol 1985; 249:G358-68.
146 Sellin JH, DeSoignie R. Regulation of bicarbonate transport in rabbitileum: pH stat studies. Am] P'hvsiol 1989; 257: G607-15.
147 Sullivan SK, Smith PL. Bicarbonate secretion by rabbit proximal colon.Am] Ph/vsiol 1986; 251: G436-45.
148 Frizzell RA, Koch MI-, Schultz SG. Itn transport by rabbit colon. 1.Actis-e and passis-ecomponents.]7,elmbr ioI 1976; 27: 297-316.
149 Feldman GM, Stephenson SL.. H and HCO3- flux across aptcalsurface otf rat diistal colon. Am]7 Phtssiol 1990); 259: C35-40.
150 Kuijpers G.AJ, 1)e Pont JHIM1. Role otf proton andi btcarbonatc tratsportin pancreatic cell function. Ann evP/2'tvsiol 1987; 49: 87-103.
151 Welsh MI1, Smith 1'1., Fromm M, Frizzell RA. C rspts are thc sttc ofintestinal fluidi andei lectrolyte secretionl. Scienlej 1982; 218: 1219-21.
152 Knickelbein R(,, Aronson l'S, 1)obbins 1W'. Mecmbrant dtistritbutton ofsodiium:hvdrogen and chloride:bicarbonate exchangcrs to crspt andtvillus cell membranes f~rom rabbit leinu. .7 (,i/tn Inveo 1988; 82:2 158-63.
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

1144 Gleeson
153 AMinhas B, Sullivan SK, Field M. Ileal HCO3- secretion in vitro: effectsof Na and Cl. Gasiroenterology 1990; 98: A548.
154 Renner EL, Lake JR, Scharschmidt BF, Zimmereli B, Meier PJ. Rathepatocytes exhibit basolateral Na- HCO3 cotransport. J Clin Invest1989; 83: 125-35.
155 Brown CDA, Dunk CR, Turnberg LA. Cl- HCO3 exchange and anionconductance in rat duodenal apical membrane vesicles. AmJI Phvsiol1989; 257: G661-7.
156 Stimenhill El, Machen TE, Williams JA. pH regulatory mechanisms inrat pancreatic duct cells. AmJ Physiol 1988; 254: G925-30.
157 Strazzabosco M, Mennone A, Boyer JL. Intracellular pH regulation inisolated rat bile duct epithelial cells. J Clin Invest 1991; 87: 1503-12.
158 Sheerin HE, Field M. Ileal HCO3 secretion: relationship to Na and Cltransport and effect of theophylline. Amj Physiol 1975; 228: 1065-74.
159 Boron WF, Boulpaep EL. Intracellular pH regulation in the renalproximal tubule of the salamander. J Gen Phvsiol 1983; 81: 53-94.
160 Banwell JG, Pierce NF, Mitra RK, Brigham KL, et al. Intestinal fluidand electrolyte transport in human cholera. J Clin Invest 1970; 49:183-95.
161 D)onowitz M, Binder HJ. Effect of enterotoxins of V. cholera, E. coli,and S. dysenteriae type 1 on fluid and electrolyte transport in thecolon. J Infect Dis 1976; 134: 135-43.
162 Chang EB, Musch MW, Mayer L. Interleukins 1 and 3 stimulate anionsecretion in chicken intestine. Gastroenterologv 1990; 98: 1518-24.
163 Nvlander 0, Flemstrom G. Effects of alpha adrenergic agonists andantagonists on duodenal surface epithelial HCO3 secretion in the rat invivo. Acta Physiol Scand 1986; 26: 433-41.
164 Gray MA, Greenwell JR, Argent BE. Secretin regulated chloridechannel on the apical plasma membrane of pancreatic duct cells.J MembrBiol 1988; 105: 131-42.
165 Welsh MJ. Electrolyte transport by airway epithelia. Phvsiol Rev 1987;67: 1143-84.
166 Welsh MJ, Liedtke CM. Chloride and potassium channels in cysticfibrosis airway epithelia. Nature 1986; 322: 467-70.
167 Shoumacher RA, Shoemaker RL, Halm DR, Tallant EA, Wallace RW,Frizzell RA. Phosphorylation fails to activate chloride channels fromcystic fibrosis airway cells. Nature 1987; 330: 752-4.
168 Taylor CJ, Baxter PS, Hardcastle J, Hardcastle PT. Failure to inducesecretion in jejunal biopsies from children with cystic fibrosis. Gut1988; 29: 957-62.
169 Gaskin KJ, Durie PR, Carey M, Wei P, Forster GG. Evidence fora primarv defect of pancreatic HCO3 secretion in cystic fibrosis.Pediatr Res 1982; 16: 554-7.
170 Dunk CR, Brown CDA. Turnberg LA. Regulation of Cl/HCO3exchange in rat duodenal brush border vesicles by cAMP. PflugersArch 1989; 414: 701-5.
171 Sundaram U, Knickelbein R, Dobbins JW. Mechanisms of cAMPmediated ileal secretion. Gastroenterology 1991; 100: A705.
172 Manganel M, Turner RJ. Rapid secretagogue induced activation ofNa+/H + exchange in rat parotid acinar cells. J Biol Chem 1991; 266:10182-8.
173 Buanes T, Grotmol T, Landsverh T, Raeder MG. Secretin empties bileduct cell cytoplasm of vesicles when it initiated ductular HCO3-secretion in the pig. Gastroenterology 1988; 95: 417-24.
174 Dumont M, Erlinger S, Uchman S. Hypercholeresis induced byursodeoxycholic acid and 7-ketolithocholic acid in the rat: possiblerole of bicarbonate transport. Gastroenterology 1980; 79: 82-9.
175 Yoon YB, Hagey LR, Hofmann AF, Guranz D, Michelotti EL,Steinbach JH. Effect of side chain shortening on the physiologicalproperties of bile acids: hepatic transport and effect on biliarysecretion pf 23-nor-ttrsodeoxycholate in rodents. Gastroenterology1986; 90: 837-52.
176 Palmer KR, Gurantz D, Hofmann AF, Clayton LM, Hagey LR,CeLcchetti S. Hypercholeresis induced by nor-chenodeoxycholate inthe biliary fistula rodent. Amj Physiol 1987; 252: G219-28.
177 Knyrim K, Vakil N, Pfab R, Classen M. The effects of intraduodenalbile acid administration on biliary secretion of ionized calcium andcarbonate in man. Hepatology 1989; 10: 134-42.
178 de Caestecker JS, Jazrawi RP, Petroni ML, Northfield TC.Ursodeoxycholic acid in chronic liver disease. Gut 1991; 32: 1061-5.
179 Gleeson D, Ruppin DC, Murphy GM, Dowling RH. Final outcome ofursodeoxycholic acid treatment in 126 patients with radiolucentgallstones. QJ Med 1990; 229: 711-29.
180 StrazzaboscoM, Sakisaka S, HayakawaT, Boyer JL. Effect ofUDCA onintracellular and biliary pH in isolated rat hepatocyte couplets andperfused liver. Amj Physiol 1991; 260: G58-69.
181 Guranz D, Schteingartt CD, Hagey LR, Steinbach JH, Grotmol T,Hofmann AF. Hypercholeresis induced by unconjugated bile acidinfusion correlated with recovery in bile of conjugated bile acids.Hepatology 1991; 13: 540-50.
182 LakeJR, Van DykeRW, Scharschmidt BF. EffectsofNa+ replacementand amiloride on ursodeoxycholic acid stimulated choleresis andbiliary bicarbonate secretion. Amj Physiol 1987; 252: G163-9.
183 Hogben CAM, Tocco DJ, Brodie BB, Schanker LS. On the mechanismof intestinal absorption of drugs. J Pharmacol Exp Ther 1959; 125:275-82.
184 Rechkemmer G. Transport of weak electrolytes. In: Schultz SG, FieldM, Frizzell RA, eds. Handbook of physiology. Section 6. Thegastrointestinal system. Vol IV. Intestinal absorption and secretion.New York: Oxford University Press, 1991: 371-88.
185 Lucas ML, Blair JA. The relationship of the acid microclimate inproximal jejunum and its relationship to luminal acidification.Proc R Soc Lond 1975; 200: 27-41.
186 Lucas ML, Kei FH, Blair1IA. The influence of buffer pH, glucos andsodium ion concentratiott on the acid microclimate of rat proximaljejunum in vitro. Pflugers Arch 1980; 385: 137-42.
187 Shiau YF, Fernandez P, Jackson MI, McMonagle S. Mechanismsmaintaining a low-pH microclimate in the intestine. Am J Phyvsiol1985; 248: G6()8-17.
188 Lucas ML, Cooper BT, Lei FH, Jo)hnson IT, Holmes GKT, Blair IA,et al. Acid microclimate in coeliac and Crohns disease: a model forfolate malabsorption. Gut 1978; 19: 735-42.
189 Shimada T. Factors affecting the microclimate pH in rat jejunum.7JPhysiol 1987; 392: 113-27.
190 MclEwan GiTA, Daniel H, Fett C, Burgess MN, Lucas ML. I'he effect
of Esterichia coli STa enterotoxin and other secretagogues on mucosalsurface pH of rat small intestine in viso. Proc R Soc Lond 1988; 234:219-37.
191 Shimada T, Hoshi 'F. Na+ dependent elevation of the acidic cell surfacepH (microclimate pH) of rat jejunal villus cells induced by cyclicnucleotides and phorbol ester: possible mediators of the regulation ofthe Na +/H + antiporter. Biochim Biophvs Acta 1988; 937: 328-34.
192 Sladen GE, Dawson AM. Effect of bicarbonate on sodium absorption bythe human jejunum. Nature 1968; 218: 267-8.
193 Turnberg LA, Fordtran JS, Carter NW, Rector FC Jr. Mechanism ofbicarbonate absorption and its relationship to sodium transport in thehuman jejunum.J7 Clin Invest 1970; 49: 548-56.
194 Shiffman ML, Sugerman HJ, Moore EW. Human gallbladder mucosalfunction. Effect of concentration and acidification of bile oncholesterol and calcium solubilitv. Gastroenterologv 1990; 99: 1452-9.
195 Martheau C, Sastre B, Iconiomidis N, Portugal H, Pauli A-M, GerolamiA. pH regulation in human gallbladder bile: study in patients with andwithout gallstones. Hepatology 1990; 11: 997-1002.
196 Gleeson D, Hood KA, Murphy GM, Dowling RH. Calcium andcarbonate ion concentrations in gallbladder and hepatic bile. Resultsin patients with radiolucent and calcified stones. Gastroenterology1992; 102: 1707-16.
197 Cummings JH. Short chain fatty acids in the human colon. Gui 1981; 22:763-79.
198 Fleming SE, Arce DS. Volatile fatty acids: their production, absorption,utilization and roles in human health. Clin Gastroenterol 1986; 15:787-814.
199 Rubsamen K, von Engelhardt W. Absorption ot Na, H ions and shortchain fatty acids from the sheep colon. Pflugers Arch 1981; 391:141-6.
200 Ruppin H, Bar-Meir S, Soergel KH, Wood CM, Schmitt MG Jr.Absorption of short chain fatty acids by the human colon.Gastroenierology 1980; 78: 1500-7.
201 Argenzio RA, Whipp SC. Inter-relationship of sodium, chloride,bicarbonate and acetate transport by the colon of the pig. J7 Phvsiol1979; 295: 365-81.
202 Sellin JH, DeSoigne R. Short chain fatty acid absorption in rabbit colonin vitro. Gastroenterology 1990; 99: 676-83.
203 Umesaki Y, Yajima T, Yokokura r, Mutal M. Effect of organic acidabsorption on bicarbonate transport in the rat colon. I'flugers Arch1979; 379: 43-7.
204 Harig JM, Soergel KH, Barry JA, Ramaswamv K. Transport ofpropionate by human ileal brush border membrane vesicles.AmJPhvsiot 1991; 260: G776-82.
205 Mascolo N, Rajendran VM, Binder HJ. Mechanism of short-chainfatty acid uptake by apical membrane vesicles of rat distal colon.Gastroenterology 1991; 101: 331-8.
206 Sellub J, Rosenberg IH. Folate transport in isolated brush bordervesicles from rat intestine.J Biol Chem 1985; 256: 4489-93.
207 Ganapathy V, Leibech L. Role of pH gradient and membrane potentialin dipeptide transport in intestinal and renal brush border membranevesicles from the rabbit..7 Biol Chem 1986; 258: 14189-92.
208 Kitis G, Lucas ML, Bishop H, et al. Altered jejunal surface pH in coeliacdisease: its effect on propranalol and folic acid absorption. Clin Sci1982; 63: 373-80.
209 Hoffmann EK, Simonsen LO. Membrane mechanisnis in volume and.pH regulation in vertebrate cells. Phvsiol Rev 1989; 69: 315-82.
210 Erikson AE, Spring KR. Volume regulation by Nectarus gallbladder:apical Na+/H+ and Cl-/HCO3- exchange. Am3r Phvsiol 1982; 243:C146-50.
211 Herbert SC. Hypertonic cell volume regulation in mouse thick limbs. II.Na+/H+ and Cl-/HCO3- exchange in basolateral membranes.AmJ Physiol 1986; 250: C920-3 1.
212 Corasanti 1G, Gleeson D, Boyer JL. Effects of osmotic stresses onisolated rat hepatocytes I. Ionic mechanisms of cell volume regulation.AmJ' Phvsiol 1990; 258: G290-8.
213 Grinstein S, Rothstein A, Sarkadi B, Gelfand EW. Responses oflymphocytes to anisotonic media: volume regulating behaviour.AmJPhvsiotl 1984; 246: C204-15.
214 Gleeson D, Corasanti JG, Boyer JL. Effects of osmotic stresses onisolated rat hepatocytes II. Modulation of intracellular pH.Am3' Phvsiotl 1990; 258: G299-307.
215 McLeod RI, Hamilton JR. Regulatory solume increase in mammalianjejunal villus cells is due to bumetanide sensitive NaKC12cotransport. AmJ7 Phvsiol 1990; 258: G665-74.
216 McLeod RI, Hamilton JR. Volume regulation initiated by Na+-nutrient cotransport in isolated mammalian villus enterocytes.AmJ7 Physiol 1991; 260: G26-33.
217 Moolenaar WH. Effect of growth factors on intracellular pH regulation.Ann Rev Ptvsiol 1986; 48: 363-75.
218 Bierman AJ, Cragoe EJ Jr, deLaat SW, Moolenaar WH. Bicarbonatedetermines cytoplasmic pH and suppresses mitogen inducedalkalinisation in fibroblastic cells.J7 Biol Chem 1988; 263: 15253-6.
219 Ganz MB, Boyarski G, Sterzel RB, Boron WF. Argenine vasopressinenhancess pHi regulation in the presence of HCO3- by stimulatingthree acid-base transport systems. Naiure 1989; 337: 648-51.
220 Perona R, Serrano R. Increased pH and tumourgenicity of fibroblastsexpressing a yeast proton pump. Naiure 1988; 334: 438-40.
221 Johnson LR. Regulation of gastrointestinal mucosal growth. Phvsiol Rev1988; 68: 456-502.
222 Goodrich AL, Suchy FJ. Na+/H+ exchange in basolateral plasmamembrane vesicles from neonatal rat liver. Am J Phvsiol 1990; 259:G334-9.
223 Bastie MJ, Delvaux M, Dufresne M, Saunier-Blache IS, Vavsse N,Ribet A. Distinct activation oif Na/H exchange by gastrin and CCKpeptide in acini from guinea pig. AmJ7 Phvsiot 1988; 254: G25-32.
224 Sacks Al, Acra 5, Dykes W, Polk B, Barnard J, Ghishan FK. The: ilealNa/H exchanger is upregulated by small intestine resection in theweanling rat. Gasiroenierotogy 1991; 100: A701.
225 Brasitus TA, Dudeja PK, Foster ES. 1,2 dimethylhydrazine inducedalteratiomns in Na+/H+ exchange in rat colonic brush bordfer mem-brane vesicles. Biochim Biophvs Acita 1988; 938: 483-8.
226 Corasanti JG, Deinhart IA. Na+/H + exchange actisity in Hr1-29 coloncarcinoma cells saries as a function osf differentiation. Gtasiro¢enieroto>gy1990; 98: A539.
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from

Acid-base transport systems in gastrointestinal epithelia 1145
227 Corasanti JG, Deinhart JA, Grinciro A. Sodium butyrate down regulatesNa+/H+ exchange activity in T-84 colon carcinoma cells.Gastroenierology 1991; 100: A683.
228 Ulrich-Baker MG, Wang P, Fitzpatrick L, Johnson LR. Amilorideinhibits rat mucosal ornithine decarboxvlase activity and DNAsynthesis. AmJ7 Phvsiol 1988; 254: G408-15.
229 Delvaux M, Bastie MJ, Chentoufi J, Cragoe EJ Jr, Vaysse N, Ribet A.Amiloride and its analogues inhibit Na/H exchange and cell prolifera-
tion in AR42J pancreatic cell line. Am]7 PhYsiol 1990; 259: (842-9.230 Montrose IH, Murer H. Polarity and kinetics of Na t /H t exchangc in
cultured opossum kidney cells. ,m]7 /hv-vsi'ol 1990; 258: Cl 21-33.231 Carter KJ, Saario 1, Seidler U, Silen W. Effect of pCO2 on intracellullar
pH in in sitro frog gastric mucosa. Amjt Pht'siol 1999; 256: G2()6- 13.232 Chow A, Dobbins JWX, Aronson PS, Igarashi 1. Cloning of a rabbit ileal
band 3 related protein, a member of the anion exchangcr gene tamily.Gastroenterologv 1991; 100: A682.
on Decem
ber 24, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.33.8.1134 on 1 August 1992. D
ownloaded from





























