Embed Size (px)
Citation preview

J Mol Cell Cardiol 29, 175–182 (1997)
Adenosine Preconditioning of HumanMyocardium is Dependent upon theATP-sensitive K+ ChannelJoseph C. Cleveland, Jr, Daniel R. Meldrum, Robert T. Rowland,Anirban Banerjee and Alden H. HarkenUniversity of Colorado Health Sciences Center, Department of Surgery, Denver, Colorado, USA
(Received 24 May 1996, accepted in revised form 8 July 1996)
J. C. C, J, D. R. M, R. T. R, A. B A. H. H. Adenosine Preconditioningof Human Myocardium is Dependent upon the ATP-sensitive K+ Channel. Journal of Molecular and CellularCardiology (1997) 29, 175–182. Evidence supports the involvement of adenosine receptor stimulation andactivation of KATP channels in ischemic preconditioning of human myocardium. It is unknown, however, whetherprotection mediated by adenosine receptors is dependent upon the KATP channel in the human heart. The purposeof this study was to determine whether adenosine-mediated protection against a simulated ischemia–reperfusioninjury in human myocardium is dependent upon KATP channels. Isolated human right atrial trabeculae wereplaced in tissue baths at 37°C, oxygenated with a modified Tyrode solution, and field stimulated at 1 Hz.Trabeculae were subjected to 45 min of normothermic simulated ischemia (hypoxic, substrate-free buffer withpacing at 3 Hz.) and 60 min of reperfusion (I/R trabeculae). Trabeculae were preconditioned with simulatedischemia (IPC trabeculae) or adenosine receptor stimulation (adenosine, 125 lmol/l) for 5 min (ADO trabeculae)prior to simulated ischemic–reperfusion injury. Inhibition of the KATP channel with glibenclamide (10 lmol/l) wascombined with adenosine pretreatment (ADO+GLI trabeculae) or alone (GLI trabeculae) prior to simulatedischemic–reperfusion injury. Developed force (DF) at end reperfusion (mean±..) was compared to baselinedeveloped force, and tissue creatine kinase (CK) activity at end reperfusion was measured. I/R trabeculae showed27±2% of baseline DF, whereas IPC trabeculae or ADO trabeculae showed 50±4% and 43±3% of baseline DF,respectively. ADO+GLI trabeculae showed 25±2% and GLI trabeculae showed 23±4% of baseline DF. TissueCK activity was enhanced in the IPC and ADO trabeculae (433±63 U/g wet myocardium, and 415±28 U/gwet myocardium, respectively). I/R trabeculae had 196±26 U/g wet myocardium and ADO+GLI trabeculae had277±38 U/g wet myocardium at end reperfusion. The results suggest that ischemic preconditioning and adenosinereceptor stimulation confer functional protection against simulated ischemic-reperfusion, and adenosine mediatedprotection is eliminated by KATP channel inhibition in human myocardium.
1997 Academic Press Limited
K W: Adenosine receptor stimulation; KATP channels; Ischemic preconditioning; Human myocardium.
preconditioning (Murry et al., 1986) and has nowbeen described in rats (Banerjee et al., 1993), rabbits(Liu et al., 1991), and recently in humans (DeutschIntroductionet al., 1990; Yellon et al., 1993; Walker et al., 1995;Cleveland et al., 1996). The mechanisms underlyingTolerance to a myocardial ischemia–reperfusion in-
jury can be induced by a preceding, transient isch- ischemic preconditioning are not completely elu-cidated, but in animal models evidence suggestsemic stress. This phenomenon is known as ischemic
stress. This phenomenon is known as ischemic that ischemic stress causes the elaboration of endo-
Please address all correspondence to: Joseph C. Cleveland, Jr, University of Colorado Health Sciences Center, Department of Surgery,Box C-305, 4200 E. Ninth Ave, Denver, CO 80262, USA.
0022–2828/97/010175+08 $25.00/0 mc960262 1997 Academic Press Limited

J. C. Cleveland et al.176
genous agonists – including norepinephrine (Baner- bottom clip was fixed while the top clip was attachedto a force transducer. Each organ bath containedjee et al., 1993), adenosine (Liu et al., 1991) and
bradykinin (Brew et al., 1995; Goto et al., 1995). 30 ml of modified Tyrode solution which wasbubbled (40 ml/min) with a 92.5% O2/7.5% CO2 gasThese agents bind to their respective receptors and
trigger a cascade of intracellular signals which mixture during normoxia. This mixture providedfor a P2>360 mmHg, a P2 of 38–42 mmHg,ultimately prime the myocyte for protection against
the subsequent ischemic insult. and buffer pH of 7.35–7.45, which were checkedroutinely with an automated blood gas analyzerWhile mechanistic studies of preconditioning in
human myocardium are still emerging, there is (ABL Instruments). Temperature in the organ bathswas maintained at 37.5°C. During the simulatedevidence that adenosine receptors confer protection
in isolated human atrial (Walker et al., 1995) and ischemic period, the gas mixture was switchedto 92.5% N2/7.5% CO2, which produced a P2<ventricular trabeculae (Cleveland et al., 1996)
against simulated ischemic or hypoxic injuries. The 50 mmHg, and the organ baths were covered toprevent atmospheric gas exchange. Except duringATP-sensitive K+ channel (KATP) is also proposed
as mediating ischemic preconditioning in animal the period of simulated ischemia, the Tyrode bufferwas replaced at 20-min intervals throughout ex-models (Gross and Auchampach, 1992), and is
suggested as an effector of ischemic preconditioning perimentation.Thirty minutes were allowed after suspension ofin human myocardium, as well (Speechly-Dick et
al., 1995). Further, in animals Auchampach and each trabeculae for recovery. After this time, thetrabeculae were gradually stretched to a restingGross (1993) have established a link between ad-
enosine receptors, KATP channels, and ischemic pre- force of 1 g, which was determined to be the optimallength–tension for human atrial trabeculae in ourconditioning. It is unknown, however, whether the
KATP channel is necessary for adenosine to confer laboratory and field stimulated. Field stimulationoccurred with platinum electrodes (Radnoti Glass,functional protection against ischemia–reperfusion
injury in human myocardium. Thus, we hypo- Inc. Monrovia, CA, USA) at a frequency of 1 Hz.The platinum electrodes were positioned on eitherthesized that exogenous adenosine administration
could replicate the functional protection of ischemic side of each trabecula, and were driven with astimulator (Grass SD9 stimulator, Warwick, RI,preconditioning, and that the KATP channel was
necessary for adenosine mediated cardioprotection USA) with 5-ms pulses at a voltage of 10% abovethreshold. Isometric contractile responses were de-in human myocardium.tected by force-displacement transducers (GrassFT03, Warwick, RI, USA), and recorded with acomputerized preamplifier/digitizer (MacLab 8, ADMaterials and MethodsInstruments, Milford, MA, USA) and a Macintoshcomputer (Apple Computer, Cupertino, CA, USA).Isolated atrial trabeculaeContractile function was assessed by recording de-veloped force (DF, g). Trabeculae which failed toRight atrial appendages were obtained from patients
undergoing coronary artery surgery during can- generate at least 0.5 g of developed force wereexcluded from study.nulation of the right atrium for cardiopulmonary
bypass. All trabeculae were from patients withstable coronary artery disease. Patients were ex-cluded from study if they had hemodynamic in- Materialsstability (mean arterial pressure <80 mmHg) within48 h of cardiopulmonary bypass, atrial dys- The modified Tyrode solution was prepared daily
with de-ionized distilled water (ddH20) and consistedrhythmias, or right atrial pressures greater than10 mmHg. Also patients currently taking oral hypo- of (mmol/l): -glucose 5.0; CaCl2 2.0; NaCl 118.0;
KCl 4.0; MgSO4⋅7H20 1.2; NaHCO3 25.0; andglycemic agents were excluded. Informed consentwas obtained from all subjects, and the study was NaH2PO4 1.2. All reagents were from Sigma Chem-
ical Co. (St Louis, MO, USA). In the substrate-freeapproved by University of Colorado Health SciencesCenter. Tyrode solution, choline chloride (7 mmol/l) was
added to maintain constant osmolarity. AdenosineEach appendage was placed in oxygenated, modi-fied Tyrode solution at 4°C. Two to three trabeculae (Sigma) was dissolved in de-ionized distilled water
and was added directly to the organ baths with a(diameter <1.0 mm, and length 4–7 mm) were ob-tained from each appendage, and suspended ver- final concentration of 125 lmol/l. Glibenclamide,
a selective inhibitor of the KATP channel, (Sigma)tically in an organ bath between two clips. The

Adenosine and KATP Channels in Human Heart 177
to I/R. Group 4 (n=6), adenosine+KATP channelblockade (ADO+GLI) were incubated with ad-enosine (125 lmol/l) in combination with gli-benclamide (10 lmol/l) for 5 min followed by10 min of normoxic Tyrode solution with pacing at1 Hz prior to I/R. Group 5 (n=4) glibencalmidecontrol (GLI) were incubated with the KATP channelinhibitor glibenclamide for 5 min followed by 10min of normoxic Tyrode solution with pacing at1 Hz prior to I/R. In addition, control trabeculae(n=3) were equilibrated for 90 min and perfusedwith normoxic Tyrode solution with pacing at 1 Hzfor 120 min to ensure model stability. At end re-perfusion, all trabeculae were removed from theorgan baths, weighed, measured, and rapidly frozenin liquid nitrogen for tissue creatine kinase (CK)activity. Samples were stored at −70°C and CKassay was performed within 2 weeks.
5min
10min
45min
60min
GLI
Perfusion Simulatedischemia
ReperfusionGLI
(n=4)
ADO
Perfusion Simulatedischemia
ReperfusionADO+
GLI(n=6)
ADO
Perfusion Simulatedischemia
ReperfusionADO(n=6)
Perfusion Simulatedischemia
ReperfusionIPC
(n=5)
Perfusion Simulatedischemia
ReperfusionI/R
(n=11)
GLI
= Simulated ischemia = Normoxic perfusion
Figure 1 Experimental groups. All experiments wereproceeded by a 90-min equilibration period which al-lowed for stabilization of DF. Simulated ischemia refersto incubation of trabeculae in substrate-free, hypoxic
Trabecular tissue creatine kinase activityTyrodes’ with pacing at 3 Hz. Normoxic perfusion andreperfusion refer to incubation of trabeculae in oxy-genated Tyrodes’ with substrate and pacing at 1 Hz. ADO End reperfusion tissue CK activity was determinedrefers to adenosine, GLI to glibenclamide. as previously described (Banerjee et al., 1991). In
brief, trabeculae were added to 10 volumes of coldwas dissolved in dimethyl sulfoxide (DMSO) and
isotonic extraction buffer consisting of (mmol/l):then dissolved in Tyrode to yield a concentration
imidazole acetate 50; Mg2+ acetate 10; KH2PO4of 10 lmol/l in the organ bath. The concentration4; EDTA 2; N-acetylcysteine 0.05; sulfur in 0.8%
of DMSO in the bath was below 0.1%.ethanol 0.012; pH 7.6. Samples were homogenizedwith a vertishear tissue homogenizer (parallelblades 0.5 cm apart) at half maximal speed for 20 sExperimental design(10 equally-spaced bursts) followed by cent-rifugation at 2000 g for 5 min and 20 000 g forTrabeculae were subjected to a 90-min equilibration
period to allow for stabilization of developed force, 10 min. The final supernatant was diluted to lessthan 0.25 absorbance units (Au/min). The assayand subsequently all experiments were concluded
for 120 min. Individual trabeculae were randomly was performed with Sigma diagnostic kit No. 47-UVon an automated spectrophotometer (Centrifichemassigned to one of five experimental groups. All
experimental groups were challenged with a 45- 500 discrete auto-analyzer, Union Carbide) in cu-vettes maintained at 30°C. Samples and reagentsmin period of simulated ischemia, which consisted
of hypoxic, substrate-free Tyrode solution with pa- were maintained at 4°C prior to assay. Results arepresented as units of CK activity per gram wetcing at 3 Hz, followed by 60 min of reperfusion with
normoxic, Tyrode solution with pacing at 1 Hz (I/R). weight of tissue.The experimental protocols are depicted in Figure 1.Group 1 (n=11), ischemia–reperfusion (I/R) under-went 15 min of normoxic perfusion prior to I/R.Group 2 (n=5), simulated ischemic preconditioned(IPC) were subjected to 5 min of hypoxic, substrate- Statistical analysisfree Tyrode solution with pacing at 3 Hz. followedby 10 minutes normoxic Tyrode’s with pacing at All data are presented as mean±standard error
(±..). All values were compared using analysis1 Hz prior to I/R. Group 3 (n=6), adenosine pre-conditioned (ADO) were incubated with adenosine of variace (ANOVA) with application of a post hoc
Bonferroni/Dunn test. P<0.05 was accepted to rep-(125 lmol/l) for 5 min followed by 10 min of nor-moxic Tyrode solution with pacing at 1 Hz prior resent a significant difference between groups.
J. C. Cleveland et al.178
Table 1 Baseline Developed Force and Atrial Trabecular Dimensions
Group Developed Length (mm) Diameter (mm) Mass (mg)force (g)
I/R 1.02±0.14 5.1±0.3 0.79±0.08 7.1±0.5IPC 1.03±0.13 5.3±0.8 0.89±0.05 6.8±1.0
ADO 0.99±0.19 5.7±0.8 0.73±0.06 5.7±0.6ADO+GLI 1.06±0.15 5.8±0.4 0.82±0.09 7.3±0.7
GLI 1.22±0.15 6.5±0.6 0.77±0.03 7.2±0.4
Values are mean±..
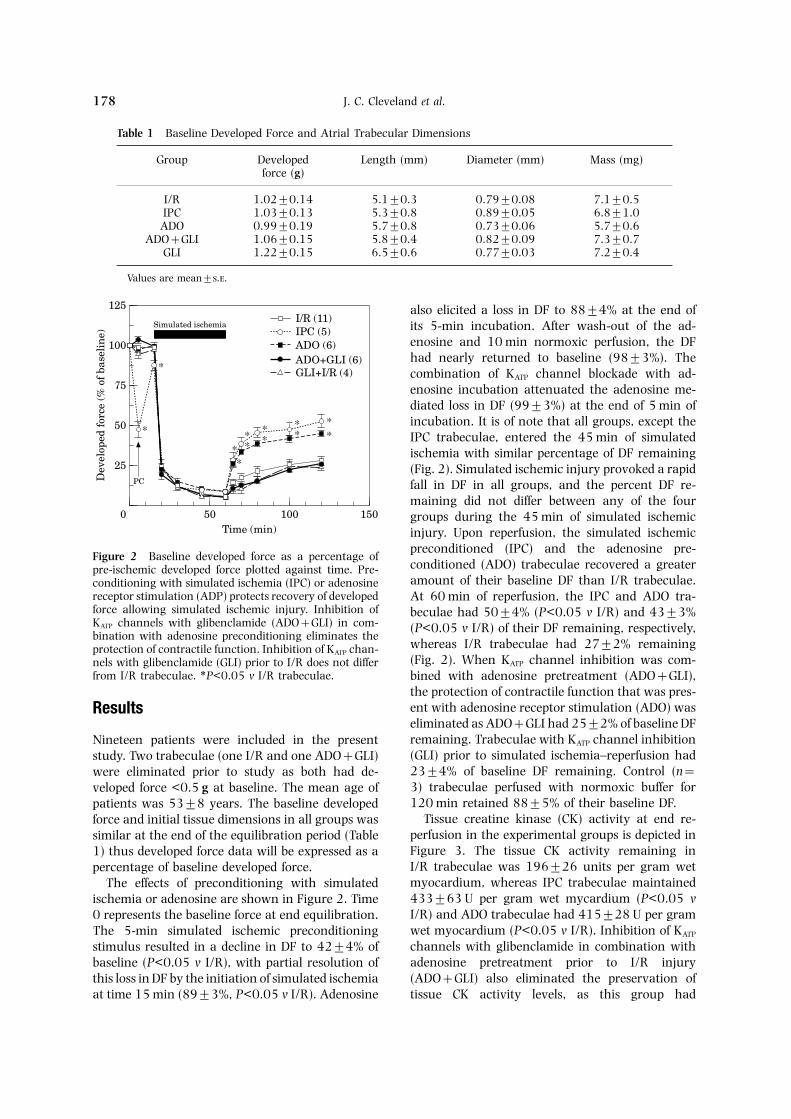
also elicited a loss in DF to 88±4% at the end ofits 5-min incubation. After wash-out of the ad-enosine and 10 min normoxic perfusion, the DFhad nearly returned to baseline (98±3%). Thecombination of KATP channel blockade with ad-enosine incubation attenuated the adenosine me-diated loss in DF (99±3%) at the end of 5 min ofincubation. It is of note that all groups, except theIPC trabeculae, entered the 45 min of simulatedischemia with similar percentage of DF remaining(Fig. 2). Simulated ischemic injury provoked a rapidfall in DF in all groups, and the percent DF re-maining did not differ between any of the fourgroups during the 45 min of simulated ischemicinjury. Upon reperfusion, the simulated ischemic
150
125
Time (min)
Dev
elop
ed f
orce
(%
of
base
lin
e)
50
100
75
50
25
1000
Simulated ischemia
PC
I/R (11)IPC (5)ADO (6)ADO+GLI (6)GLI+I/R (4)*
***
***
****
*
preconditioned (IPC) and the adenosine pre-Figure 2 Baseline developed force as a percentage of
conditioned (ADO) trabeculae recovered a greaterpre-ischemic developed force plotted against time. Pre-amount of their baseline DF than I/R trabeculae.conditioning with simulated ischemia (IPC) or adenosine
receptor stimulation (ADP) protects recovery of developed At 60 min of reperfusion, the IPC and ADO tra-force allowing simulated ischemic injury. Inhibition of beculae had 50±4% (P<0.05 v I/R) and 43±3%KATP channels with glibenclamide (ADO+GLI) in com- (P<0.05 v I/R) of their DF remaining, respectively,bination with adenosine preconditioning eliminates the
whereas I/R trabeculae had 27±2% remainingprotection of contractile function. Inhibition of KATP chan-(Fig. 2). When KATP channel inhibition was com-nels with glibenclamide (GLI) prior to I/R does not differ
from I/R trabeculae. ∗P<0.05 v I/R trabeculae. bined with adenosine pretreatment (ADO+GLI),the protection of contractile function that was pres-ent with adenosine receptor stimulation (ADO) wasResultseliminated as ADO+GLI had 25±2% of baseline DFremaining. Trabeculae with KATP channel inhibitionNineteen patients were included in the present(GLI) prior to simulated ischemia–reperfusion hadstudy. Two trabeculae (one I/R and one ADO+GLI)23±4% of baseline DF remaining. Control (n=were eliminated prior to study as both had de-3) trabeculae perfused with normoxic buffer forveloped force <0.5 g at baseline. The mean age of120 min retained 88±5% of their baseline DF.patients was 53±8 years. The baseline developed
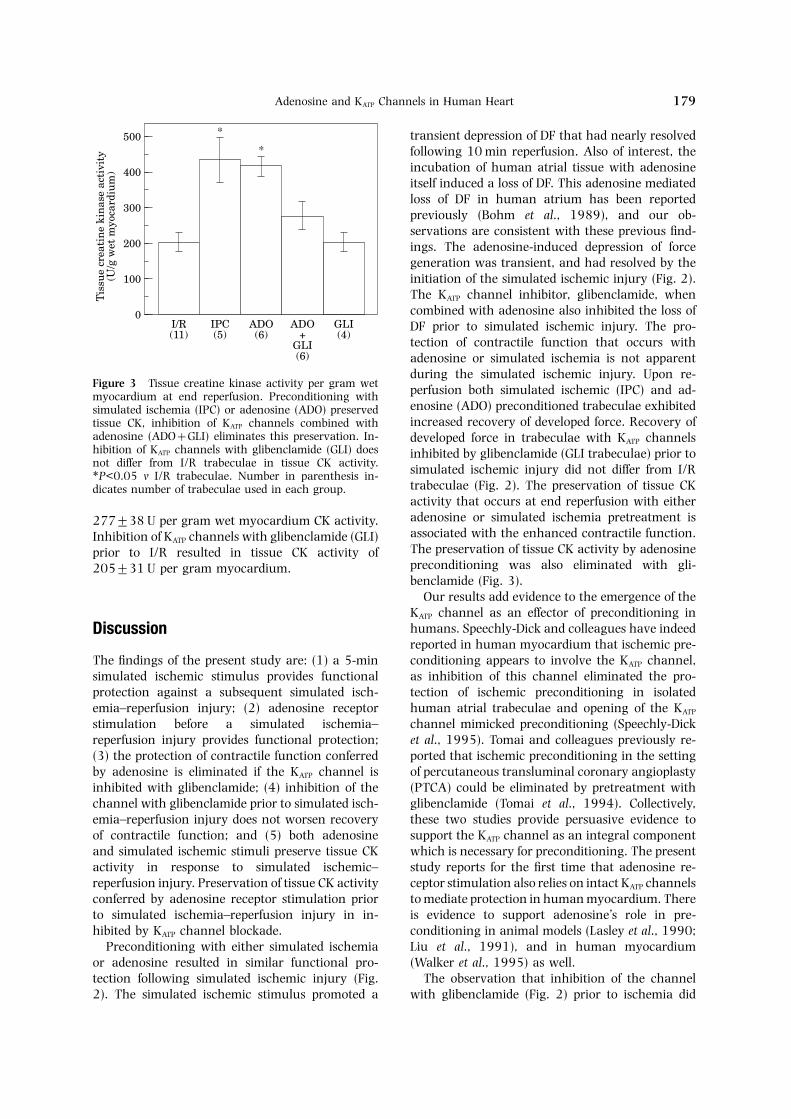
Tissue creatine kinase (CK) activity at end re-force and initial tissue dimensions in all groups wasperfusion in the experimental groups is depicted insimilar at the end of the equilibration period (TableFigure 3. The tissue CK activity remaining in1) thus developed force data will be expressed as aI/R trabeculae was 196±26 units per gram wetpercentage of baseline developed force.myocardium, whereas IPC trabeculae maintainedThe effects of preconditioning with simulated433±63 U per gram wet mycardium (P<0.05 vischemia or adenosine are shown in Figure 2. TimeI/R) and ADO trabeculae had 415±28 U per gram0 represents the baseline force at end equilibration.wet myocardium (P<0.05 v I/R). Inhibition of KATPThe 5-min simulated ischemic preconditioningchannels with glibenclamide in combination withstimulus resulted in a decline in DF to 42±4% ofadenosine pretreatment prior to I/R injurybaseline (P<0.05 v I/R), with partial resolution of(ADO+GLI) also eliminated the preservation ofthis loss in DF by the initiation of simulated ischemia
at time 15 min (89±3%, P<0.05 v I/R). Adenosine tissue CK activity levels, as this group had
Adenosine and KATP Channels in Human Heart 179
transient depression of DF that had nearly resolvedfollowing 10 min reperfusion. Also of interest, theincubation of human atrial tissue with adenosineitself induced a loss of DF. This adenosine mediatedloss of DF in human atrium has been reportedpreviously (Bohm et al., 1989), and our ob-servations are consistent with these previous find-ings. The adenosine-induced depression of forcegeneration was transient, and had resolved by theinitiation of the simulated ischemic injury (Fig. 2).The KATP channel inhibitor, glibenclamide, whencombined with adenosine also inhibited the loss ofDF prior to simulated ischemic injury. The pro-tection of contractile function that occurs withadenosine or simulated ischemia is not apparent
GLI(4)
500
0
Tis
sue
crea
tin
e ki
nas
e ac
tivi
ty(U
/g w
et m
yoca
rdiu
m)
I/R(11)
400
300
200
100
IPC(5)
ADO(6)
ADO+
GLI(6)
*
*
during the simulated ischemic injury. Upon re-Figure 3 Tissue creatine kinase activity per gram wet perfusion both simulated ischemic (IPC) and ad-myocardium at end reperfusion. Preconditioning with
enosine (ADO) preconditioned trabeculae exhibitedsimulated ischemia (IPC) or adenosine (ADO) preservedincreased recovery of developed force. Recovery oftissue CK, inhibition of KATP channels combined with
adenosine (ADO+GLI) eliminates this preservation. In- developed force in trabeculae with KATP channelshibition of KATP channels with glibenclamide (GLI) does inhibited by glibenclamide (GLI trabeculae) prior tonot differ from I/R trabeculae in tissue CK activity. simulated ischemic injury did not differ from I/R∗P<0.05 v I/R trabeculae. Number in parenthesis in-
trabeculae (Fig. 2). The preservation of tissue CKdicates number of trabeculae used in each group.activity that occurs at end reperfusion with eitheradenosine or simulated ischemia pretreatment is277±38 U per gram wet myocardium CK activity.associated with the enhanced contractile function.Inhibition of KATP channels with glibenclamide (GLI)The preservation of tissue CK activity by adenosineprior to I/R resulted in tissue CK activity ofpreconditioning was also eliminated with gli-205±31 U per gram myocardium.benclamide (Fig. 3).
Our results add evidence to the emergence of theKATP channel as an effector of preconditioning inhumans. Speechly-Dick and colleagues have indeedDiscussionreported in human myocardium that ischemic pre-conditioning appears to involve the KATP channel,The findings of the present study are: (1) a 5-min
simulated ischemic stimulus provides functional as inhibition of this channel eliminated the pro-tection of ischemic preconditioning in isolatedprotection against a subsequent simulated isch-
emia–reperfusion injury; (2) adenosine receptor human atrial trabeculae and opening of the KATP
channel mimicked preconditioning (Speechly-Dickstimulation before a simulated ischemia–reperfusion injury provides functional protection; et al., 1995). Tomai and colleagues previously re-
ported that ischemic preconditioning in the setting(3) the protection of contractile function conferredby adenosine is eliminated if the KATP channel is of percutaneous transluminal coronary angioplasty
(PTCA) could be eliminated by pretreatment withinhibited with glibenclamide; (4) inhibition of thechannel with glibenclamide prior to simulated isch- glibenclamide (Tomai et al., 1994). Collectively,
these two studies provide persuasive evidence toemia–reperfusion injury does not worsen recoveryof contractile function; and (5) both adenosine support the KATP channel as an integral component
which is necessary for preconditioning. The presentand simulated ischemic stimuli preserve tissue CKactivity in response to simulated ischemic– study reports for the first time that adenosine re-
ceptor stimulation also relies on intact KATP channelsreperfusion injury. Preservation of tissue CK activityconferred by adenosine receptor stimulation prior to mediate protection in human myocardium. There
is evidence to support adenosine’s role in pre-to simulated ischemia–reperfusion injury in in-hibited by KATP channel blockade. conditioning in animal models (Lasley et al., 1990;
Liu et al., 1991), and in human myocardiumPreconditioning with either simulated ischemiaor adenosine resulted in similar functional pro- (Walker et al., 1995) as well.
The observation that inhibition of the channeltection following simulated ischemic injury (Fig.2). The simulated ischemic stimulus promoted a with glibenclamide (Fig. 2) prior to ischemia did

J. C. Cleveland et al.180
not exacerbate simulated ischemia–reperfusion-in- enosine mediated cardioprotection is dependentupon the KATP channel in human myocardium.duced contractile dysfunction requires comment.
Cole et al. (1991) observed that inhibition of the There is much evidence to support a car-dioprotective role for openings of KATP channels.KATP channel with glibenclamide prior to ischemia
worsened myocardial ischemic injury, whereas Grover et al. (1990) established a protective rolefor both cromakalin and pinancidil upon recoveryopening of the channel with pinancidil protected
recovery of mechanical function in guinea-pig myo- of contractile function and limitation of infarct sizein dogs and rats. Although the exact nature ofcardium. Similarly, Thornton et al. (1993) also
observed that inhibition of the channel with gli- this protection is unknown, it is hypothesized thatopening of the KATP channel shortens action po-benclamide produced larger infarcts compared to
hearts subjected to ischemia alone. Finally, Gross tential duration, thus allowing less Ca+2 entry dur-ing ischemia. In support of this hypothesis, Behlingand Auchampach (1992) administered an opener
of KATP channels which markedly reduced infarct and Mallone (1995) found a reduction in cytosoliccalcium in rat hearts given a KATP channel openersize in dogs subjected to myocardial regional isch-
emia–reperfusion. Collectively, these previous re- prior to ischemia. However, Yao and Gross (1994)showed that the channel opener bimakalim exertedports suggest that KATP channel may have an
endogenously protective role in the ischemic myo- infarct size reduction following ischemia but didnot accelerate the shortening of action potentialcardium. The mechanism of this endogenous pro-
tection against ischemic injury remains unknown duration. Thus, opening of KATP channels is likelyto involve cellular mechanisms of protection otherand was not addressed in the present study.
The initial coupling of adenosine A1 receptors to than electrophysiological alterations.The present study employed isolated human rightthe KATP channel was proposed by Kirsch et al. in
neonatal rat ventricular myocytes (1990). The link atrial trabeculae contracting isometrically to ex-plore preconditioning in human myocardium.between adenosine receptor stimulation, intact KATP
channels, and ischemic preconditioning was ini- There are important metabolic (Davies et al., 1947)and physiological (Schwinger et al., 1993; Urthalertially reported by Auchampach and Gross and fur-
ther suggested that opening of the KATP channel was et al., 1975) differences in atrial and ventriculartissues. Differences in the intracellular signalingan important mediator of ischemic preconditioning.
This study established that preconditioning with initiated by adenosine receptor stimulation havealso been described between human atrial andadenosine and dipyridamole resulted in a marked
infarct-size reduction in dogs subjected to regional ventricular tissue (Bohm et al., 1989). It is possiblethat our observations are specific to human atrialmyocardial ischemia–reperfusion. When KATP chan-
nels were inhibited with glibenclamide, the pro- tissue, and that the adenosine mediated dependenceupon KATP channels is not operative in humantective effects of adenosine and dipyridamole
preconditioning were eliminated (Auchampach and ventricle. However, previous work from our laborat-ory indicates that ischemic preconditioning or ad-Gross, 1993).
Toombs et al. (1993) also reported that ad- enosine receptor stimulation prior to a hypoxicinjury in diseased, explanted human ventricularenosine’s protection of rabbit myocardium was de-
pendent upon KATP channel activation. Of interest, trabeculae yielded qualitatively similar protectionto that seen in the present study (Cleveland et al.,this group not only reported that adenosine’s in-
farct-size reduction was reversed with gli- 1996). Human right atrium is an attractive modelfor studying mechanisms of preconditioning inbenclamide, but higher doses of glibenclamide were
associated with larger infarcts. Collectively, both human myocardium because it is readily available,relatively disease-free, and represents a stable pre-animal and human studies suggest that endogenous
adenosine released during the transient stimulus of paration for study of contractile function. Theseadvantages have been described by othersischemic preconditioning may mediate endogenous
cardioprotective properties through activation of (Speechly-Dick et al., 1995) who have employed asimilar model for the study of human pre-KATP channels. Kitakaze et al. (1996), however, have
proposed an alternative hypothesis coupling the conditioning.The original description of myocardial pre-adenosine receptor to the KATP channel. Their results
implicate opening of the KATP channel which then conditioning by Murry et al. (1986) reported areduction in myocellular necrosis with a precedingactivates ectosolic 5′-nucleotidase, with subsequent
adenosine release. Further work is required to fully ischemic stress. In the particular model of this study,it is difficult to measure a reduction in myocellularcharacterize the link between adenosine and KATP
channels, but the present study suggests that ad- necrosis, but we interpret the recovery of contractile

Adenosine and KATP Channels in Human Heart 181
B EC, M MB, R TF, G-Rfunction (DF) as an important parameter of pre-F, MI J RC, H AH, B A, 1995.conditioning mediated protection. Others (JenkinsThe role of bradykinin in cardiac functional protection
et al., 1995) have reported that the improved re- after global ischemia-reperfusion in the rat heart. Amcovery of function following global ischemia is pro- J Physiol 269: H1370–H1378.
C JC , W MM, M DR, R-portional to a reduction in infarction size. Our tissue RT, R TF, S BC, H AH,CK data indirectly implies that preconditioning withB A, 1966. Ischemic preconditioning in humansimulated ischemia or adenosine receptor stimu-and rat ventricle. Am J Physiol (in press).
lation improves myocellular viability. The tissue CK C WC, MP CD, S D, 1991. ATP-reg-data, however, does not directly address whether ulated K+ channels protect the myocardium against
ischemia/reperfusion damage. Circ Res 69: 571–581.either of the two preconditioning strategies pro-D F, F E, S H, 1947. The distributionmoted lessening of myocellular necrosis. Thus, we
of nucleotide, phosphocreatine, and glycogen in thecan only speculate whether the enhanced tissueheart. J Physiol 106: 154–166.
CK activity which was present in the adenosine- D E, B M, K W, H J, H-or ischemic-preconditioned trabeculae represents a H, L W, 1990. Adaptation to ischemia
during percutaneous transluminal coronary an-metabolic or viability adaptation conferred by eithergioplasty: clinical, hemodynamic, and metabolic fea-preconditioning strategy.tures. Circulation 82: 2044–2051.The present study reports that preconditioning
G M, L Y, Y X, A J, C MV, Dwith simulated ischemia or with adenosine receptor JM, 1995. Role of bradykinin in protection of ischemicstimulation prior to a prolonged simulated ischemic preconditioning in rabbit hearts. Circ Res 77: 611–621.
G GJ, A J, 1992. Blockade of ATP-sensitiveinjury in human myocardium promotes recoverypotassium channels prevents myocardial pre-of contractile function and enhanced tissue CKconditioning in dogs. Circ Res 70: 223–233.activity at end reperfusion. The protection of con-
G GJ. S PG, D S, 1990. The protectivetractile function or augmented tissue CK activity effects of cromakalim and pinancidil on reperfusionwhich was present in the adenosine-pretreated tra- function and infarct size in isolated perfused rat hearts
and anesthetized dogs. Cardiovasc Drugs Ther 4: 465–beculae was eliminated with KATP channel in-474.hibition. The link between adenosine and the KATP
J DP, P WB, Y DM, 1995. Ischaemicchannel in human myocardium may add importantpreconditioning in a model of global ischemia; infarct
insights into the mechanism(s) of ischemic pre- size reduction, but no reduction in stunning. J Mol Cellconditioning in human myocardium. Finally, as Cardiol 27: 1623–1632.
K GE, C J, B L, B AM, 1990.both adenosine receptor stimulation and the KATPCoupling of ATP-sensitive K+ channels to A1 receptorschannel are accessible pharmacologically, they mayby G proteins in rat ventricular myocytes. Am J Physiolultimately be employed to provide protection259: H820–H826.
against elective clinical myocardial ischemia– K M, M T, N K, K K,reperfusion injury. S Y, C M, M H, I M, H M,
K T, 1996. Role of activation of ectosolic 5′-nucleotidase in the cardioprotection mediated by open-ing of K+ channels. Am J Physiol 270: H1744–H1756.
References L RD, R J, VW DGL, M RM ,1990. Adenosine A1 receptor mediated protection of
A JA, G GJ, 1993. Adenosine A1 receptors, the globally ischemic isolated rat heart. J Mol CellCardiol 22: 39–47.KATP channels, and ischemic preconditioning in dogs.
Am J Physiol 264: H1327–H1336. L G, T JD, W DVM, S AHJ, OR, D JM, 1991. Protection against infarctionB A, G MA, B JM, R KB, W-
GJR, 1991. Oxygen metabolite effects on creatine afforded by preconditioning in mediated by A1 ad-enosine receptors in rabbit heart. Circulation 84: 350–kinase and cardiac energetics after reperfusion. Am J
Physiol 262: H590–H597. 356.M CE, J RB, R KA, 1986. Pre-B A, L-W C, R K, M M,
B E, C C, B DD, K AH, 1993. conditioning with ischemia: a delay of lethal cell injuryin ischemic myocardium. Circulation 74: 1124–1136.Preconditioning against myocardial dysfunction after
ischemia and reperfusion by an a1-adrenergic mech- S R, B M, K A, U R, UP, K E, R B, E E, 1993. Force-anism. Circ Res 73: 656–670.
B RW, M HJ, 1995. KATP channel openers frequency-relation in human and atrial and ventricularmyocardium. Mol Cell Biochem 119: 73–78.protect against increased cytosolic calcium during isch-
emia and reperfusion. J Mol Cell Cardiol 27: 1809– S-D ME, G GJ, Y DM, 1995. Doesischemic preconditioning in the human involve protein1817.
B M, P B, U M, E E, 1989. kinase C and the ATP-dependent K+ channel? CircRes 77: 1030–1035.Characterization of A1 adenosine receptors in atrial
and ventricular myocardium from diseased human T JD, T C, S D, D JM,1993. Blockade of ATP-sensitive potassium channelshearts. Circ Res 65: 1201–1211.

J. C. Cleveland et al.182
increases infarct size but does not prevent pre- U F, W A, H L, J T, 1975. Com-parison of contractile performance of canine atrial andconditioning in rabbit hearts. Circ Res 72: 44–49.
T F, C F, G A, V F, DP R, ventricular muscles. Circ Res 37: 762–771.W DM, W JM, P WB, P CW,P P A, C L, G PA,
1994. Ischemic preconditioning during coronary an- Y DM, 1995. Preconditioning in isolated super-fused human muscle. J Mol Cell Cardiol 27: 1349–1357.gioplasty is prevented by glibenclamide, a selective
ATP-sensitive K+ channel blocker. Circulation 90: 700– Y Z, G GJ, 1994. Effects of the KATP channel openerbimakalim on coronary blood flow, monophasic action705.
T C, MG D, J W, V-J J, potential duration, and infarct size in dogs. Circulation89: 1769–1775.1993. Protection of ischaemic–reperfusion injury with
adenosine pretreatment is reversed by inhibition of Y DM, A A, P WB, 1993. Pre-conditioning the human myocardium. Lancet 342:ATP sensitive potassium channels. Cardiovasc Res 27:
623–629. 276–277.