Embed Size (px)
Citation preview

ISSN 0032�9452, Journal of Ichthyology, 2009, Vol. 49, No. 11, pp. 1073–1080. © Pleiades Publishing, Ltd., 2009.
1073
INTRODUCTION
It is known that the larger and longer the river sys�tem is where the Atlantic Salmon Salmo salar repro�duces the more diverse are conditions of its spawningand subsequent life of its juveniles. These conditionsare formed both in the mainstream and in numeroustributaries (Kazakov et al., 1992; Zubchenko et al.,2007; Martynov, 2007). Riffles and rapids of river sys�tems are subdivided into groups of spawning–rearinggrounds at various distances from the river mouth,sometimes by hundreds kilometers as, e.g., in largerivers of the North�West of Russia: the Pechora, theNorthern Dvina, the Mezen, the Ponoy, the Varzuga,and the Tuloma. Such a situation is reflected in theintraspecies diversity of populations of Atlanticsalmon, including the genetic level (Primmer et al.,2006). The number of spawning groups (“run groups”)of salmon increases, the reaching of the smoltificationage of juveniles of one generation is stretched to one–three years, and the period of feeding of fish in the seadiffers by the same interval.
Previously, we described conditions of formation ofstable phenotypic groups of zero class of salmon in dif�ferent types of rearing grounds in the Varzuga (Pavlovet al., 2007). Differences in accumulation of particularfractions of lipids and fatty acids were revealed,
depending on the habitat. Finally, to the end of sum�mer, these differences were reflected in size and weightparameters of phenotypic groups of zero class (Pavlovet al., 2008). It may be assumed that later these differ�ences become more expressed or modified and influ�ence the dates of smoltification.
Thus, it is interesting to follow the age dynamics oflipid status (the state of complex of parameters of lipidmetabolism) in juveniles of salmon of different agegroups in biotopes of the Varzuga and the Arenga, itstributary. These biotopes differ in hydrological condi�tions and food resources. The lipoid status reflects thenatural differentiation of fish by the level of lipoid con�tent, the level of readiness for smoltification, andmigration to the sea. It may be considered as one ofmechanisms on intrapopulational diversity of Atlanticsalmon in long and geomorphologically complicatedrivers.
The present study is aimed at the comparativeinvestigation of the lipid status in salmon juveniles ofdifferent age (parrs 0+, 1+, 2+; smolts 3+).
MATERIAL AND METHODS
The juveniles of Atlantic salmon, parr of age 0+,1+, 2+, and smolts 3+ were caught in the coastal part
Age Dynamics of Lipid Status of Juveniles of Atlantic Salmon (Salmo salar L.) from the Varzuga River
D. S. Pavlova, Z. A. Nefedovab, A. E. Veselovb, N. N. Nemovab, T. R. Ruokolainenb, O. B. Vasil’evab, and P. O. Ripattib
a Institute of Ecology and Evolution, Russian Academy of Sciences, Leninskii pr. 33, Moscow, 119071 Russiab Institute of Biology, Karelian Research Center, Russian Academy of Sciences,
ul. Pushkinskaya 11, Petrozavodsk, 185910 Russiaa e�mail: [email protected]
Received June 17, 2009
Abstract—Heterogeneity of zero class (0+) of Atlantic Salmon Salmo salar from the compared biotopes ofthe Varzuga R. (the Kola Peninsula) is revealed by the content of TAG, PL, 18:2n�6,18:3n�3, the sum ofmonoenoic FA, and parameters of CS/PL and TAG/PL retained in elder age groups of parr (1+ and 2+). Itdepends on the quality of food resources and hydrological conditions. The increase in the level of PS, one ofthe principal regulators of activity of Na+/K+�ATPase—a key enzyme of osmoregulation—in parrs 1+ and,to a greater extent, in smolts is supposed to be the formation of one of the biochemical mechanisms of pre�adaptation of fish to a marine environment. Salmon juveniles 3+ differ from the fish of junior age groups in ahigher content of PUFA (20:4n�6, 20:5n�3, 22:6n�3), a considerable decrease in TAG, in parameterTAG/PL, and the sum of monoenoic FA related to smoltification and preparation for life in a marine envi�ronment. Multidirectional variations of concentrations of minor phospholipids (PI, LPC, SPM) in salmonjuveniles depending on age and habitat may indicate the change in viscosity of biomembranes and, accord�ingly, of activity of some enzymes bound with membranes.
DOI: 10.1134/S003294520911006X
Key words: Atlantic salmon, juveniles, lipid status, phenotypic groups, individual variability.

1074
JOURNAL OF ICHTHYOLOGY Vol. 49 No. 11 2009
PAVLOV et al.
of the rapids of the Varzuga mainstream and in themouth of the Arenga, its tributary (the Kola Peninsula,66°32′5 N, 36°15′5 E). The fish were captured by anelectrofishing apparatus (Fa�2) (Norway) on June 20–25, 2005 and 2006, when, after their distribution fromspawning nests and wintering grounds, the stablegroups of parr were formed and migrating smolts wereaccessible. It should be noted that the investigatedsmolts were not connected with the aforementionedbiotopes and belonged to a random sample of down�stream migrants from the upstream cataracts of themainstream and tributaries of the Varzuga R. Unfortu�nately, as migration was short it was not possible to col�lect the material on smolts (3+) from the Arenga.
The number of zero class in each sample was 30;parrs 1+, 2+, and smolts 3+ from 8 to 30. After catch�ing, to remove the effect of electromagnetic field, theparrs were kept for a day in channel cages. Then the
samples were fixed with 97% alcohol and minced thor�oughly for better extraction of lipids.
The lipid status was estimated by the content oftotal lipids (TL), triacylglycerols (TAG), cholesterol(CS), cholesterol esters (CSE), total phospholipids(PL) and their particular fractions: phosphatidylinos�itol (PI), phosphatidylserin (PS), phosphatidyletha�nolamine (PEA), phosphatidylcholin (PC), lysophos�phatidylcholin (LPC), sphingomyelin (SPM), andfatty acids (FA). Methods of determination of lipidsand fatty acids are given by Pavlov et al. (2008). Inaddition, we may note that lipid fractions CS and CSEwere separated on ten same chromatographic plates inthe same mixture of solvents (petroleum ether�sulphu�ric ether�acetic acid, 90 : 10 : 1) as total Pl and TAG.The quantity of CSE, as well as the total content of PLand TAG, were determined by the hydroxamatemethod (Pavlov et al., 2008), CS was determined by
Table 1. The content of lipids (% of dry weight of sample) in juveniles of Atlantic salmon Salmo salar in relation to age and feed�ing ground, June 2005 and 2006
ParametersAge groups of fish
0+ 1+ 2+ 3+ (smolts)
Number of fish
Length of fish, cm
Weight of fish, g
TL
PL
TAG
CS
CSE
CS/PL
TAG/PL
Note: Here and in Tables 2 and 3: above the line is the Arenga biotope and under the line is the Varzuga. Differences are significant at p <0.05: * between the fish of the same age from the compared biotopes, a between ages 0+ and 1+, b between ages 1+ and 2+, andc between age 2+ and 3+ (smolts).
3030���� 25
25���� 8
12���� –
15����
3.05 0.02±
3.04 0.04±���������������������� 5.14 0.09±
5.12 0.08±��������������������� 7.89 0.30±
7.58 0.20±����������������������
–
11.3 0.4c
±
���������������������
0.18 0.01±
0.18 0.01±��������������������� 1.35 0.06±
1.24 0.06±��������������������� 4.49 0.40±
3.70 0.30±����������������������
–
11.9 0.7c
±
���������������������
9.94 0.26±
12.54 0.46*±��������������������������� 9.24 0.30
a±
10.0 0.40a
±
�����������������������8.40 0.50±
7.33 0.20*b
±
�������������������������� –
6.8 0.6c
±
�������������������
4.59 0.20±
4.37 0.24±���������������������� 5.24 0.20
a±
4.88 0.20±�����������������������
5.67 0.30±
4.12 0.20*b
±
�������������������������� –4.4 0.7±�����������������
1.69 0.13±
2.75 0.27*±������������������������ 1.13 0.10
a±
2.34 0.20*±������������������������
1.33 0.40±
1.49 0.10b
±
����������������������� –
0.5 0.2c
±
�������������������
3.21 0.20±
4.37 0.25*±������������������������
2.82 0.20±
2.56 0.20a
±
����������������������� 1.39 0.10b
±
1.69 0.10b
±
����������������������� –1.8 0.6±�����������������
0.46 0.05±
0.54 0.07±���������������������� 0.05 0.02
a±
0.23 0.10*a
±
�������������������������� 0.01 0.01b
±
0.02 0.01b
±
����������������������� –0.1 0.1±�����������������
0.77 0.07±
1.03 0.07*±������������������������
0.59 0.08±
0.55 0.05a
±
����������������������� 0.25 0.02b
±
0.43 0.05*±������������������������
–0.43 0.07±����������������������
0.37 0.04±
0.63 0.07*±������������������������ 0.22 0.04
a±
0.48 0.04a*±
�������������������������� 0.24 0.07±
0.36 0.05±����������������������
–
0.11 0.02c
±
�����������������������

JOURNAL OF ICHTHYOLOGY Vol. 49 No. 11 2009
AGE DYNAMICS OF LIPID STATUS OF JUVENILES OF ATLANTIC SALMON 1075
the reaction with a staining reagent (Engelbrechtet al., 1974) and expressed in percentages of dry weightof the sample.
RESULTS
On selected parts of the rapids at the coast of theVarzuga R. (the Varzuga biotope) and the mouth of theArenga R. tributary (the Arenga biotope) as was shownpreviously (Pavlov et al., 2007, 2008) the fry fromspawning nests are distributed and elder age groups ofsalmon parr also arrive here after wintering. Salmonfry are distributed from the central part of the Varzuga R.rapids. Differences of life conditions in these biotopesin juveniles of different age groups predetermine theformation of stable phenotypic groups which do notalways differ in their size and weight characteristics(Table 1).
Comparing the content of total lipids in salmonjuveniles of various age (0+, 1+, 2+, 3+) frombiotopes of Varzuga and Arenga in June, it may be con�cluded that their significant (p ≤ 0.01–0.0001)decrease with age occurs due to TAG, CS, and CSE. Aconsiderable decrease in the content of TAG in smoltsis found (3.2–5.9 times) in comparison with that injuveniles of earlier age. Decrease of the part of CS andCSE is noted up to the age of parrs 2+. Due to suchchanges, parameters of CS/PL and TAG/PL in juve�niles in both biotopes also decreased.
On the whole, the content of total PL in juvenilesfrom the Varzuga R. did not change with age. However,by particular groups of PL, multidirectional variations
are recorded (Table 2). There is a noticeable growth ofthe level of PS and SPM, especially in smolts, anddecrease in PI and LPC, at significant differences (p <0.02). In specimens from the Arenga, with age theconcentration of total PL steadily increased due toPEA, PS, and SPM, at significant differences (p <0.03).
Differences of the lipid status in salmon juveniles(0+, 1+, 2+) from two biotopes (Tables 1, 2) arefound: zero class from the Varzuga R. differed fromthose from the Arenga tributary in a higher level oftotal lipids, TAG, CS, PI, PS, PEA, PC, SPM, inparameters CS/PL and TAG/PL; parr 1+ from theVarzuga R. had higher values of TAG (twice), CSE,LPC, parameter CS/PL and lower PI and SPM; inparr 2+ from the Varzuga a decreased content of totallipids, total PL, PEA, and a high value of parameterCS/PL are noted. These values are significantly differ�ent (p < 0.05).
Thus, in juveniles from the Varzuga R., in compar�ison with specimens from the Arenga tributary, thecontent of TAG and SCE prevailed and there was ahigher parameter TAG/PL while the level of total PLwas lower. With age, in juveniles from both rivers therewas a decrease of total lipids, TAG, CSE, CS, param�eters CS/PL and TAG/PL, while the part of PSincreased. At the same time, in the fish from theArenga tributary, the concentration of total PLincreased, mainly due to PEA, PS, and SPM. In com�parison with other age groups, the smolts differed inthe lowest level of total lipids, TAG, SPM, LPC,
Table 2. The content of particular fractions of phospholipids (% of dry weight of sample) in juveniles of Atlantic salmon Salmosalar in relation to age and feeding ground, June 2005 and 2006
PhospholipidsAge groups of fish
0+ 1+ 2+ 3+ (smolts)
PC
LPC
PEA
PS
PI
SPM
Note: The number of fish in age groups is indicated in Table 1.
2.28 0.12±
2.84 0.23*±������������������������ 2.50 0.20±
2.49 0.20±���������������������� 2.60 0.40±
2.20 0.10±���������������������� –
2.42 0.39±����������������������
1.26 0.10±
0.83 0.08*±������������������������ 0.53 0.10
a±
0.92 0.10*±������������������������
0.93 0.30±
0.50 0.10±���������������������� –
0.29 0.10±���������������������
0.44 0.04±
0.59 0.06*±������������������������ 0.69 0.10
a±
0.80 0.10±�����������������������
0.92 0.10b
±
0.48 0.05*b
±
�������������������������� –0.57 0.08±����������������������
0.06 0.01±
0.10 0.02*±������������������������ 0.26 0.03
a±
0.27 0.10a
±
����������������������� 0.22 0.04±
0.22 0.04±����������������������
–
0.43 0.07c
±
�����������������������
0.29 0.02±
0.42 0.05*±������������������������ 0.11 0.02
a±
0.05 0.01*a
±
�������������������������� 0.26 0.10b
±
0.13 0.04±�����������������������
–0.15 0.14±���������������������
0.010 0.001±
0.012 0.002±�������������������������� 0.21 0.03
a±
0.14 0.02*a
±
�������������������������� 0.14 0.03b
±
0.15 0.04±�����������������������
–
0.07 0.02c
±
�����������������������
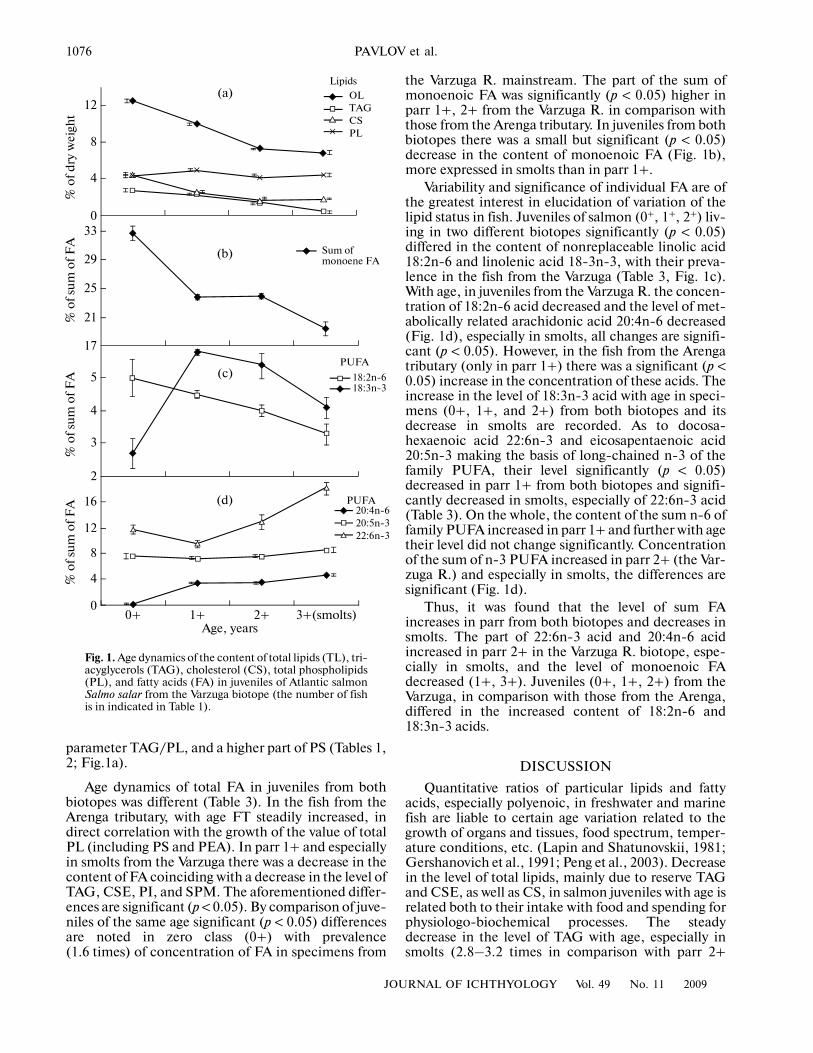
1076
JOURNAL OF ICHTHYOLOGY Vol. 49 No. 11 2009
PAVLOV et al.
parameter TAG/PL, and a higher part of PS (Tables 1,2; Fig.1a).
Age dynamics of total FA in juveniles from bothbiotopes was different (Table 3). In the fish from theArenga tributary, with age FT steadily increased, indirect correlation with the growth of the value of totalPL (including PS and PEA). In parr 1+ and especiallyin smolts from the Varzuga there was a decrease in thecontent of FA coinciding with a decrease in the level ofTAG, CSE, PI, and SPM. The aforementioned differ�ences are significant (p < 0.05). By comparison of juve�niles of the same age significant (p < 0.05) differencesare noted in zero class (0+) with prevalence(1.6 times) of concentration of FA in specimens from
the Varzuga R. mainstream. The part of the sum ofmonoenoic FA was significantly (p < 0.05) higher inparr 1+, 2+ from the Varzuga R. in comparison withthose from the Arenga tributary. In juveniles from bothbiotopes there was a small but significant (p < 0.05)decrease in the content of monoenoic FA (Fig. 1b),more expressed in smolts than in parr 1+.
Variability and significance of individual FA are ofthe greatest interest in elucidation of variation of thelipid status in fish. Juveniles of salmon (0+, 1+, 2+) liv�ing in two different biotopes significantly (p < 0.05)differed in the content of nonreplaceable linolic acid18:2n�6 and linolenic acid 18�3n�3, with their preva�lence in the fish from the Varzuga (Table 3, Fig. 1c).With age, in juveniles from the Varzuga R. the concen�tration of 18:2n�6 acid decreased and the level of met�abolically related arachidonic acid 20:4n�6 decreased(Fig. 1d), especially in smolts, all changes are signifi�cant (p < 0.05). However, in the fish from the Arengatributary (only in parr 1+) there was a significant (p <0.05) increase in the concentration of these acids. Theincrease in the level of 18:3n�3 acid with age in speci�mens (0+, 1+, and 2+) from both biotopes and itsdecrease in smolts are recorded. As to docosa�hexaenoic acid 22:6n�3 and eicosapentaenoic acid20:5n�3 making the basis of long�chained n�3 of thefamily PUFA, their level significantly (p < 0.05)decreased in parr 1+ from both biotopes and signifi�cantly decreased in smolts, especially of 22:6n�3 acid(Table 3). On the whole, the content of the sum n�6 offamily PUFA increased in parr 1+ and further with agetheir level did not change significantly. Concentrationof the sum of n�3 PUFA increased in parr 2+ (the Var�zuga R.) and especially in smolts, the differences aresignificant (Fig. 1d).
Thus, it was found that the level of sum FAincreases in parr from both biotopes and decreases insmolts. The part of 22:6n�3 acid and 20:4n�6 acidincreased in parr 2+ in the Varzuga R. biotope, espe�cially in smolts, and the level of monoenoic FAdecreased (1+, 3+). Juveniles (0+, 1+, 2+) from theVarzuga, in comparison with those from the Arenga,differed in the increased content of 18:2n�6 and18:3n�3 acids.
DISCUSSION
Quantitative ratios of particular lipids and fattyacids, especially polyenoic, in freshwater and marinefish are liable to certain age variation related to thegrowth of organs and tissues, food spectrum, temper�ature conditions, etc. (Lapin and Shatunovskii, 1981;Gershanovich et al., 1991; Peng et al., 2003). Decreasein the level of total lipids, mainly due to reserve TAGand CSE, as well as CS, in salmon juveniles with age isrelated both to their intake with food and spending forphysiologo�biochemical processes. The steadydecrease in the level of TAG with age, especially insmolts (2.8–3.2 times in comparison with parr 2+
4
00+
% o
f su
m o
f F
A
Age, years
12
1+ 2+ 3+(smolts)
8
16
2
3
4
5
17
21
25
29
330
4
8
12(a)
(b)
(c)
(d)
% o
f su
m o
f F
A%
of
sum
of
FA
% o
f d
ry w
eigh
t Lipids
OLTAGCSPL
Sum of
PUFA
monoene FA
18:2n�618:3n�3
PUFA20:4n�620:5n�322:6n�3
Fig. 1. Age dynamics of the content of total lipids (TL), tri�acyglycerols (TAG), cholesterol (CS), total phospholipids(PL), and fatty acids (FA) in juveniles of Atlantic salmonSalmo salar from the Varzuga biotope (the number of fishis in indicated in Table 1).

JOURNAL OF ICHTHYOLOGY Vol. 49 No. 11 2009
AGE DYNAMICS OF LIPID STATUS OF JUVENILES OF ATLANTIC SALMON 1077
from both biotopes) indicates to greater intensity ofoxidative processes and the decrease in the lipid syn�thesis rate in them, related to smoltification (Sheri�dan, 1989). Many experts note that these changesoccur at a considerable expenditure of energy, first ofall of energy�rich lipids (Soivio et al., 1988; Virtanenet al., 1992). With reference to coho salmon Onco�rhynchus kisutch and steelhead trout Salmo gairdneri,Sheridan (1986) demonstrated the decrease of con�centration of lipids in blood serum and in tissues, prin�cipally due to TAG. In earlier investigations, it wasfound that up to the beginning of silvering the salmonslose from 30 to 40% of fat in comparison with parr(Malikova, 1957). The noted decrease in the level ofCS and CSE in parr 1+ and 2+ in comparison withzero class (0+) may be caused by their active use as astructural component of biomembranes, steroid hor�mones, and sources of FA (CSE). In cells, there is abalanced system regulating the content of free choles�terol, a part of which is esterified yielding a depot ofcellular cholesterol and fatty acid as cholesterin esters(Kreps, 1981). The level of total PL in juveniles fromthe Varzuga with age remained permanent, the part ofparticular phospholipids fractions varied: the concen�tration of minor PS and SPM grows and PI and LPCdecrease. In specimens from the Arenga tributary, withage the concentration of total Pl increased due toPEA, PS, and SPM. The increase in the level of PS in
parr 1+ from both biotopes and especially in smoltsmay be related to greater activity of Na+/K+—ATPase, a key enzyme of osmoregulation, and indi�cates to the formation of a biochemical mechanisms ofpreadaptation in the preparation of fish for migrationfrom rivers to the marine environment (Gennis, 1997;Boldyrev et al., 2006). The adaptive role of phospho�lipids in exothermal organisms is very diverse both bythe function which they regulate and by the actionmechanism (Kreps, 1981). In particular, the revealedmultidirectional variations of concentrations of par�ticular phospholipid fractions (PEA, PI, PS, LPC,and SPM) in salmon juveniles depending on age andhabitat may indicate to the change in viscosity ofbiomembranes and, accordingly, be specific regulatorsof activity of some membrane�bound enzymesaccording to conditions of life activities of the organ�ism (Dyatlovitskaya and Bezuglov, 1998; Prokazovaet al., 1998; Boldyrev et al., 2006).
The principal distinction of life conditions of parrfrom the compared biotopes is the different flow rate atmicrostations: it is higher in the Arenga tributary—0.98 (0.82–1.28) m/s—than in the Varzuga main�stream—0.74 (0.70–0.95) m/s. It is known that athigher flow rates the quantity of food organisms drift�ing in water increases and, accordingly, the frequencyof foraging darts made by salmon juveniles increases
Table 3. The content of sum fatty acids (% of dry weight of sample) and of particular fatty acids (% of FA sum) in total lipids injuveniles of Atlantic salmon Salmo salar in relation to age and feeding ground, June 2005 and 2006
Fatty acidsAge group
0+ 1+ 2+ 3+ (smolts)
Sum FA
Sum of monoene FA 19.4 ± 3.1
Linolic 18:2n�6 3.3 ± 0.7c
Linolenic 18:3n�3 4.1 ± 0.7c
Arachidonic 20:4n�6 4.7 ± 0.7c
Eicosapentaenoic 20:5n�3 8.5 ± 0.9c
Docosahexaenoic 22:6n�3 18.3 ± 2.0c
Note: The family of linolenic acid is n�3 and the family of linolic acid is n�6. The number of fish in age groups is indicated in Table 1.
2.47 0.46±
3.94 0.97*±������������������������ 3.53 1.50
a±
3.66 1.96a
±
����������������������� 4.56 0.75b
±
4.21 0.59±�����������������������
–
1.8 0.9c
±
�������������������
31.9 2.6±
32.8 3.0±������������������� 21.3 2.6
a±
23.9 1.6*a
±
������������������������ 21.1 2.7±
24.0 1.6*±����������������������
3.6 0.6±
5.0 1.3*±������������������� 3.8 0.6
a±
4.5 0.3*a
±
���������������������3.5 0.3±
4.0 0.4*b
±
���������������������
2.2 0.5±
2.7 0.9*±������������������� 4.1 0.7
a±
5.8 0.2*a
±
��������������������� 4.0 0.6±
5.4 0.8*±�������������������
0.2 0.1±
0.2 0.1±����������������� 3.8 0.5
a±
3.4 0.4*a
±
���������������������3.9 0.5±
3.8 0.6b
±
�������������������
7.3 0.8±
7.7 1.0±����������������� 7.1 0.6
a±
7.3 0.6a
±
������������������� 6.2 0.3±
7.6 0.7*±�������������������
13.3 1.9±
11.9 1.6*±���������������������� 11.2 1.3
a±
9.7 1.4*a
±
���������������������11.6 2.0±
13.0 2.2b
±
���������������������

1078
JOURNAL OF ICHTHYOLOGY Vol. 49 No. 11 2009
PAVLOV et al.
(Shustov, 1983; Veselov and Kalyuzhin, 2001). Conse�quently, an increased level of the sum of phospholip�ids, low values of TASG and of parameter TAG/PL injuveniles from the Arenga tributary in comparison withparr from the Varzuga may be related to the intensiveexpenditure of lipids at a high locomotion activity offish in the accelerated water flow with greater turbu�lence and flow rate.
The increased content of total lipids, includingreserve TAG and CSE, in all age groups of fish fromthe Varzuga mainstream in comparison with thosefrom the Arenga tributary in June is related to differ�ence in food spectra and foraging mode in salmonjuveniles (Shustov, 1987; Pavlov et al., 2008). Inverte�brates of drift in the coastal zone of the Varzuga main�stream are estimated as the most accessible and attrac�tive feed for salmon juveniles. This corresponds to theprevailing level in juveniles of all ages from the Varzugaof nonreplaceable 18:2n�6 and 18:3n�3 acids whichare not synthesized in the fish organism but are takenwith food (Pravdina, 1975; Sargent et al., 1995; Bellet al., 1997).
Comparison of concentrations of 20:4n�6 acid(0.2% of the sum of FA) and of its precursor 18:2n�6acid (3.6–5.0%) in zero class from both biotopes indi�cated to a low activity (if any) of linoleoyl�CoA desat�urase playing a key role in the transformation of 18:2n�6 to 20:4n�6 acid (Bell et al., 2002). The stabledecrease with age of the level of 18:2n�6 acid in juve�niles from the Varzuga R. (from 5.0 to 3.3% of the sumFA) and the increase in the content of 20:4n�6 acid(from 0.2 to 4.7%) indicate to the activation of enzymesystems of desaturation and elongation of FA. It maybe related to hormonal restructuring in fish, especiallyin smolts (Sheridan, 1989). Among other ecologicalfactors, the qualitative composition of food is a leadingfactor controlling the content of PUFA in tissues ofmost fish (Sushchik, 2008). In freshwater fish, in com�parison with marine fish, food 18:2n�6 and 18:3n�3acids are more transformed into long�chain 20:4n�6,20:5n�3, 22:6n�3 acids (Sargent et al., 1955; Penget al., 2003). Some species of frewshwater fish differ inthe ability of conversion of these acids, which may bedifferent at a particular stage of ontogenesis and belimited by presence of an increased level of food20:5n�3 and 22:6n�3 acids (Sargent et al., 1995; Penget al., 2003). The increase of the level of 20:5n�3 and22:6n�3 acids in parr 2+ in the Varzuga R. biotope andof 20:4n�6 acid in juveniles in both biotopes and espe�cially in smolts may be related to modification of thefatty�acid composition of the freshwater type into themarine type. Usually this happens during preparationfor downstream migration to the sea (Peng et al.,2003). It is known that smolts have an increased levelof long�chain PUFA typical for marine fish (Sheridanet al., 1985; Ogata and Murai, 1989). It is also shownthat 22:6n�3 and 20:4n�6 acids play a significant rolein the regulation of activity of nerve cells and in theformation of the optic system in fish, while their defi�
ciency leads to anomalies in behavioral reactions (Bellet al., 1995; Chyb et al., 1999; Hwang, 2000; Gamohet al., 2001). Such changes in the spectrum of n�3 andn�6 families of PUFA in elder juveniles should be con�sidered the mobilization of adaptive reactions of theorganism (Rabinovich and Ripatti, 1994). This con�clusion is based on the results of imitation modeling ofproperties of fatty�acid chains of various levels ofunsaturation (Rabinovich et al., 2004; Rabinovich,2008). In particular, it was shown that PUFA of thetype 22:6n�3 possess a sharply decreased sensitivity oftheir characteristics (both general and local) tochanges in environmental temperature in comparisonwith the saturated acids. Such trait of theirs is one ofreasons for the increased content of PUFA in bound�ary lipid layers embedded in the membrane ofenzymes.
CONCLUSIONS
(1) The age dynamics of lipid status of juveniles ofAtlantic salmon is characterized by an increased levelof PUFA—20:4n�6, 20:5n�3, and 22:6n�3—and bythe decreased content of high�energy TAG, as well asCS, CSE, and the sum of monoenoic FA indicating tothe intensification of metabolism related to smoltifi�cation and to preparation for life in the marine envi�ronment.
(2) The increase of the level of PS (the most effi�cient regulator of activity of Na+/K+–ATPase, a keyenzyme of osmoregulation) in parr 1+ and especiallyin smolts may be related to the formation of a bio�chemical mechanism of preadaptation of fish tomarine environment.
(3) The found diversity of zero class (0+) of Atlan�tic salmon from biotopes of the Varzuga mainstreamand the Arenga tributary by the content of TAG, totalPL, nonreplaceable 18:2n�6 and 18:3n�3, and the sumof monoenoic FA is retained in elder age groups of parr(1+, 2+). These differences depend on the quality offood resources (the species composition and lipidspectrum of food items) and on hydrological condi�tions of the compared biotopes.
(4) The determined differences in the lipid status ofsalmon juveniles may predetermine stretching (overone–three years) of the oncoming of smoltification inparr of the same generation and, accordingly, early orlate migration to the sea which generally influencesthe formation of the complex age structure of the pop�ulation.
ACKNOWLEDGMENTS
The study is supported by the Russian Foundationfor Basic Research (08�04�00927a, 08�04�91771�AF),the Biodiversity: Inventory, Functioning, Conserva�tion Program of Fundamental Research of the Presid�ium of the Russian Academy of Sciences, and the

JOURNAL OF ICHTHYOLOGY Vol. 49 No. 11 2009
AGE DYNAMICS OF LIPID STATUS OF JUVENILES OF ATLANTIC SALMON 1079
Leading Scientific Schools Program of the Presidentof the Russian Federation (NSh�2104.2008.4).
REFERENCES
1. M. V. Bell, R. S. Batty, J. R. Dick, et al., “Dietary Defi�ciency of Docosahexaenoic Acid Impairs at Low LightIntensities in Juvenile Herring (Clupea harengus L.),”Lipids 30 (5), 443–449 (1995).
2. J. D. Bell, D. R. Tocher, B. M. Farndale, et al., “TheEffect of Dietary Lipid on Polyunsaturated Fatty AcidMetabolism in Atlantic Salmon (Salmo salar) Under�going Parr�Smolt Transformation,” Lipids, No. 32,515–525 (1997).
3. J. D. Bell, R. J. Henderson, D. R. Tocher, et al., “Sub�stituting Fish Oil with Crude Palm Oil in the Diet ofAtlantic Salmon (Salmo salar) Affects Muscle FattyAcid Composition and Hepatic Fatty Acid Metabo�lism,” J. Nutr., No. 132, 222–230 (2002).
4. A. A. Boldyrev, E. I. Kyaivyaryainen, and V. A. Ilyukha,Membranology (KarNTs RAN, Petrozavodsk, 2006) [inRussian].
5. S. Chyb, P. Raghu, and R. C. Hardle, “PolyunsaturatedFatty Acids Activate the Drosophyla Light�SensitiveChannels TRP and TRPL,” Nature 397, 255–259(1999).
6. E. V. Dyatlovitskaya and V. V. Bezuglov, “Lipids as Bio�effectors, Introduction,” Biokhimiya 63 (1), 3–5(1998).
7. F. M. Engelbrecht, F. Mari, and J. T. Anderson, “Cho�lesterol Determination in Serum. A Rapid DirectionMethod,” S.A. Med. J. 48 (7), 250–356 (1974).
8. S. Gamoh, M. Hashimoto, S. Hossain, andS. Masumura, “Chronic Administration of Docosa�hexanoic Acid Improves the Performance of RadialArm Maze Task in Aged Rats,” Clin. Exp. Pharmacol.Physiol. 28 (4), 266–270 (2001).
9. R. B. Gennis, Biomembranes: Molecular Structure andFunction (Springer, New York, 1989).
10. A. D. Gershanovich, V. I. Lapin, and M. I. Sha�tunovskii, “Specific Features of Lipid Metabolism inFish,” Zh. Usp. Sovrem. Biol. 111 (2), 207–219(1991).
11. D. Hwang, “Fatty Acids and Immune Responses—ANew Perspective in Searching for Clues to Mecha�nisms,” Annu. Rev. Nutr. 20, 431–456 (2000).
12. R. V. Kazakov, O. G. Kuz’min, Yu. A. Shustov, andI. L. Shchurov, Atlantic Salmon from the Varzuga River(Gidrometioizdat, St. Petersburg, 1992) [in Russian].
13. E. M. Kreps, Lipids of Cellular Membranes. Brain LipidEvolution. Adaptation Function of Lipids (Nauka, Lenin�grad, 1981) [in Russian].
14. V. I. Lapin and M. I. Shatunovskii, “Specific Featuresof Composition and the Physiological and EcologicalImportance of Lipids in Fish,” Zh. Usp. Sovrem. Biol.92 (3(6)), 380–394 (1981).
15. E. M. Malikova, “Biochemical Assessment of JuvenileSalmon at a State Close to a Downstream�MigratingOne and at a Delay of Silvering in Fresh Water,” Tr.Latv. Otd. Vses. Nauchno�Issled. Inst. Rybn. Khoz.Okeanogr., No. 3, 257–281 (1957).
16. V. G. Martynov, Atlantic Salmon (Salmo salar L.) in theNorth of Russia (Ural. Otd. Ross. Akad. Nauk, Yekate�rinburg, 2007) [in Russian].
17. H. Ogata and T. Murai, “Effects of Dietary Fatty AcidComposition on Growth and Smolting of Underyear�ling Masu Salmon, Oncorhynchus masu,” Aquaculture82 (1–4), 181–190 (1989).
18. D. S. Pavlov, O. V. Meshcheryakova, A. E. Veselov,et al., “Parameters of Energy Metabolism in Juvenilesof Atlantic Salmon Salmo salar Living in the Main�stream and in the Tributary of the Varzuga River (theKola Peninsula),” Vopr. Ikhtiol., No. 6, 819–826(2007) [J. Ichthyol., No. 9, 774–781 (2007)].
19. D. S. Pavlov, Z. A. Nefedova, A. E. Veselov, et al.,“Lipid Status of Fingerlings of the Atlantic SalmonSalmo salar L. from Different Microbiotopes of theVarzuga River,” Vopr. Ikhtiol., No. 5, 48 (2008) [J. Ich�thyol., No. 8, 648–654 (2008)].
20. J. Peng, Y. Larondelle, D. Pham, et al., “Polyunsatu�rated Fatty Acid Profiles of Whole Body Phospholipidsand Triacylglicerols in Anadromous and LandlockedAtlantic Salmon (Salmo salar) Fry,” Comp. Biochem.Physiol. B 134, 335–348 (2003).
21. N. I. Pravdina, “Importance of Fatty Acid Radicals inthe Structural Heterogeneity and Metabolism of Phos�pholipids,” Usp. Sovr. Biol. 79 (2), 205–223 (1975).
22. C. R. Primmer, A. E. Veselov, A. Zubchenko, et al.,“Isolation by Distance within a River System: GeneticPopulation Structuring of Atlantic Salmon, Salmosalar, in Tributaries of the Varzuga River in NorthwestRussia,” J. Mol. Ecol., No. 15, 653–666 (2006).
23. N. V. Prokazova, N. D. Zvezdina, and A. A. Korotaeva,“The Effect of Lysophosphatidylcholine on the Trans�fer of the Transmembrane Signal into the Cell,”Biokhimiya 63 (1), 38–46 (1998).
24. A. L. Rabinovich, “Temperature Dependence of Con�formation Properties of Oligomer Chains of NaturalLipids: Computer Modeling,” Biofizika 53 (3), 426–433 (2008).
25. A. L. Rabinovich and P. O. Ripatti, “PolysaturatedHydrocarbon Chains of Lipids: Structure, Properties,and Functions,” Usp. Sovr. Biol. 114 (5), 581–591(1994).
26. A. L. Rabinovich, P. O. Ripatti, and N. K. Balabaev,“Computer Modeling of Biological Membranes,” inWorks of the Karelian Research Center, Russian Acad�emy of Sciences (Petrozavodsk, 2004), Issue 6, pp. 99–137.
27. X. Rollin, J. Peng, D. Pham, et al., “The Effects ofDietary Lipid and Strain Difference on Polyunsatu�rated Fatty Acid Composition and Conversion inAnadromous and Landlocked Salmon (Salmo salar(L.)) Parr,” Comp. Biochem. Physiol. B 134, 349–366(2003).
28. J. R. Sargent, J. G. Bell, M. V. Bell, et al., “Dietary Ori�gins and Functions of Long�Chain (n�3) Polyunsatu�rated Fatty Acids in Marine Fish,” J. Mar. Biotechnol.,No. 3, 26–28 (1995).
29. M. A. Sheridan, “Effects of Thyroxin, Cortisol,Growth Hormone, and Prolactin on Lipid Metabolismof Coho Salmon, Oncorynchus kisutsh, during Smoltifi�

1080
JOURNAL OF ICHTHYOLOGY Vol. 49 No. 11 2009
PAVLOV et al.
cation,” Gen. Comp. Endocrinol. 64 (2), 220–238(1986).
30. M. A. Sheridan, “Alterations in Lipid MetabolismAccompanying Smoltification and Seawater Adapta�tion of Salmonid Fish,” Aquaculture 82 (1–4), 191–204 (1989).
31. M. A. Sheridan, W. V. Allen, and T. H. Kersteller,“Changes in the Fatty Acid Composition of SteelheadTrout Salmo gairdneri Richardson Assoiciated with ParrSmolt Transformation,” Comp. Biochem. Physiol. B,No. 80, 671–676 (1985).
32. Yu. A. Shustov, Ecology of Juveniles of Atlantic Salmon(Izd�vo Kareliya, Petrozavodsk, 1983) [in Russian].
33. Yu. A. Shustov, Ecological Specific Features and Behav�ior of Juveniles of Atlantic Salmon in Rivers of Kareliaand Kola Peninsula (KarNTs RAN, Petrozavodsk,1987), p. 37 [in Russian].
34. A. Soivio, E. Virtanen, M. Backstrom, et al., “Lohiis�tukkaiden Kunnon ja Vaellusvalminden Seuranta,”Suomen Kalatalous, No. 53, 134–152 (1988).
35. N. N. Sushchik, “The Role of Indispensable FattyAcids in Trophometabolic Interactions in FreshaterEcosystems (Review),” Zh. Obshch. Biol. 69 (4), 299–316 (2008).
36. A. E. Veselov and S. M. Kalyuzhin, Ecology, Behavior,and Distribution of Juvenile Atlantic Salmon (Izd�voKareliya, Petrozavodsk, 2001) [in Russian].
37. E. Virtanen, K. L. Forsman, K. A. Soivio, and L. Sed�erholm�Tana, “Smoltification of Salmon Salmo salarL. of Neva Origin and Criteria of the PhysiologicalQuality of Hatchery�Reared Smolts,” Sb. Nauch. Tr.Nauchno�Issled. Inst. Ozern. Rechn. Rybn. Khoz.,No. 297, 28–43 (1992).
38. K. A. Zubchenko, S. M. Kalyuzhin, A. E. Veselov, et al.,Specific Features of Reproduction of Atlantic Salmon(Salmo salar L.) in the Umba Rive (Kola Peninsula)(Skandinaviya, Petrozavodsk, 2007) [in Russian].





























